Introduction
Neutrophils are the most abundant type of leukocyte
in the human bloodstream and are recruited from the circulation to
sites of infection, inflammation or damage (1). As key components of the innate immune
system, neutrophils have various functions, such as releasing
granule proteins, generating reactive oxygen species (ROS) and
releasing neutrophil extracellular traps (NETs) (1,2).
NETs are web-like structures made of chromatin and
are decorated with histones, myeloperoxidase (MPO), neutrophil
elastase and cathepsin G. NETs are known for their ability to
defend against pathogens, including Staphylococcus aureus
(3), pneumococci (4) and streptococci (5). Previous studies (3,5,6) have
highlighted the ‘double-edged sword’ concept, as NETs have been
identified to have a role in the host immune defense, while also
being implicated in the pathophysiology of various diseases,
including thrombosis. A further understanding of how neutrophils
cast their NETs is critical for exploring the pathogenesis of
diseases associated with NETs.
Numerous pathogens may induce neutrophils to form
NETs via different signals, such as Toll-like receptors, Fc
receptors, chemokine receptors and cytokine receptors (1). Brinkmann et al (7) first described the NETosis pathway in
2004; this cell death process depends on the generation of ROS by
NADPH oxidase (8) and the
generation of citrullinated histone H3 (citH3) by peptidylarginine
deiminase type 4 (PADI4) (9).
Neutrophils are also able to release NETs in a cell
death-independent manner called vital NETosis. In this process,
NETs are released rapidly and NADPH oxidase function is not
necessary (10,11). Recently, Chen et al (12) reported that cytosolic
lipopolysaccharide or cytosolic gram-negative bacteria are able to
activate the noncanonical (caspase-4/11) inflammasome and trigger
gasdermin D-dependent neutrophil cell death recently called
noncanonical NETosis, which is still debatable. The involvement of
a number of pathways implies that the progress of NET formation is
complex; however, the short life of neutrophils makes it difficult
to explore the molecular and cellular mechanisms of NET
formation.
HL-60 cells may be differentiated into
granulocyte-like cells using all-trans retinoic acid (ATRA) or
dimethyl sulfoxide (DMSO). A model based on a stable cell line has
been used to reduce dissonant results due to different experimental
settings or conditions (13-15)
and differentiated HL-60 (dHL-60) cells have been used to study
neutrophil functions, including NETosis (16-18).
However, the inefficiency of dHL-60 cells in generating NETs makes
it challenging to completely replace neutrophils in the analysis of
NET formation. In the present study, the cell culture and
differentiation conditions that lead to the most effective release
of NETs from dHL-60 were optimized.
Methods
Reagents
Fetal bovine serum (FBS), goat sera, RPMI-1640
medium and PBS were purchased from Biological Industries Israel
Beit Haemek. X-VIVO™ 15 medium was purchased from Lonza (cat. no.
04-418Q). FITC Annexin V Apoptosis Detection kits were purchased
from BD Pharmingen (cat. no. 556547). Anti-CD11b-PE, anti-CD16-APC
and anti-CD66b-FITC were purchased from Biolegend (cat. no. 101207,
302011 and 305104, respectively). ATRA was purchased from
MedChemExpress (cat. no. 302-79-4). DMSO was purchased from
Sigma-Aldrich (Merck KGaA; cat. no. D8418-100). Neutrophil
isolation kits were purchased from TBD (cat. no. LZS11131). citH3
antibody (histone H3 citrulline R2 + R8 + R7), anti-MPO antibody
and Ca2+ ionophore (CI) were purchased from Abcam (cat.
nos. ab5103, ab9535 and ab120287, respectively). Goat anti-mouse
Alexa Fluor 488, ProLong Gold Antifade reagent and histone H3 mouse
antibody were purchased from Cell Signaling Technology, Inc. (cat.
nos. 4408s, 9071S and 14269s, respectively). Goat anti-rabbit Alexa
Fluor 568, Total ROS Assay kits and Quant-iT™ PicoGreen™
double-strand (ds)DNA Assay kits were purchased from Thermo Fisher
Scientific, Inc. (cat. no. A-11011, 88-5930-74 and P7589,
respectively). R-PE-antibody conjugation kits were purchased from
Fcmacs (cat. no. FMS-ABPE0001).
Cell culture and differentiation
The HL-60 cell line was obtained from the American
Type Culture Collection. Cells were cultured in RPMI-1640 medium
supplemented with 10% FBS or X-VIVO™ 15 medium in a humidified
atmosphere containing 5% CO2 at 37˚C. Cells were
passaged every three days and only cells passaged no more than 15
times were used for the experiments. ATRA (1 µmol/l) or DMSO
(1.25%) were used to induce HL-60 cells to differentiate. CD11b,
CD16 and CD66b expression was detected to evaluate the rate of
HL-60 differentiation using a BD FACSCanto II flow cytometer (BD
Biosciences). FITC Annexin V Apoptosis Detection kits were used to
determine the viability of differentiated cells using a BD
FACSCanto II flow cytometer (BD Biosciences).
Quantification of ROS production using
flow cytometry
Total ROS Assay kits were used to quantify ROS in
cells. The 500 X ROS Assay Stain stock solution was diluted to 1X
using the ROS Assay Buffer. The cells were centrifuged for 5 min at
350 x g and washed with PBS. Washed cells (1x106
cells/ml) were incubated with 1X ROS assay stain for 60 min in a
37˚C incubator with 5% CO2, followed by treatment with
phorbol 12-myristate 13-acetate (PMA; 50 nmol/l) or CI (4 µmol/l)
for 1.5 h to induce the production of ROS. The analysis was
performed with a BD FACSCanto II flow cytometer using the 488 nm
(blue) laser in the FITC channel.
NETs visualization by fluorescence
microscopy
In order to avoid abnormal NET generation caused by
serum, the dHL-60 cells were centrifuged for 5 min at 350 x g at
4˚C and washed with PBS. Washed cells (5x105 cells/ml)
were incubated for 1 h in RPMI-1640 in confocal dishes and the
cells were then stimulated with 50 nmol/l of PMA or 4 µmol/l of CI
for 4 h in a humidified atmosphere containing 5% CO2 at
37˚C. Cells were fixed with 4% paraformaldehyde for 20 min at room
temperature and permeabilized with 0.3% Triton X-100 for 10 min at
room temperature. Samples were blocked with 10% goat serum for 1 h
at 37˚C and then incubated with antibodies directed against MPO
(1:100 dilution) or H3 (1:100 dilution) overnight at 4˚C. After
washing with PBS, the cells were incubated with goat anti-rabbit
Alexa Fluor 568 (1:1,000 dilution) and goat anti-mouse Alexa Fluor
488 (1:500 dilution) for 1 h at 37˚C in the dark. Cells were
incubated with DAPI for 10 min at room temperature and exposed to
ProLong Gold Antifade reagent according to the manufacturer's
protocols. Cells were visualized using a confocal TCS SP8
microscope (Leica Microsystems).
Comparison of NETs composition
Cells were visualized using a confocal microscope
and the composition of NETs was calculated via four random
fluorescence scanning planes using ImageJ software (version 1.46r;
National Institute of Health) (6).
The average size of individual NETs was calculated by measuring the
area of DNA stained by DAPI, the average size of NET-associated
proteins was calculated from the area of MPO or histone H3. The
composition of NETs was determined by calculating the average size
of NET-associated proteins divided by the average size of the area
occupied by DNA.
Quantification of NETs
PicoGreen was used to detect dsDNA following the
manufacturer's protocol. Washed dHL-60 cells were transferred to
96-well plates (1x106 cells/ml in RPMI-1640) and the
cells were stimulated with 50 nmol/l PMA or 4 µmol/l CI for 4 h.
The supernatant was gently removed and replenished with 100 µl PBS
and an equivalent amount of 1X Quant-iT PicoGreen reagent was added
to the 96-well plates, followed by incubation for 5 min at room
temperature. Fluorescence was detected at an excitation wavelength
of 502 nm with an emission wavelength of 523 nm using a
spectrofluorometer (SpectraMax M2; Molecular Devices).
CitH3 quantification by flow
cytometry
CitH3 quantification was performed as previously
described (19), with certain
modifications. Anti-citH3 antibody was pre-conjugated with
PE-antibody using antibody conjugation kits. Cells were cultured in
RPMI-1640 and stimulated with PMA (50 nmol/l) or CI (4 µmol/l) for
2 h at 37˚C in 5% CO2, fixed in 2% paraformaldehyde for
20 min, washed with PBS and centrifuged for 20 min at 16,000 x g at
4˚C. Cells were incubated with citH3-PE for 30 min at room
temperature in the dark, washed with PBS, centrifuged for 20 min at
16,000 x g at 4˚C and analyzed using a BD FACSCanto II flow
cytometer.
Statistical analysis
SPSS Statistics 23.0 for Win10 (IBM Corp.) was used
for data processing. Normally distributed data were expressed as
the mean ± standard deviation. One-way ANOVA was used to analyze
differences among groups. As a multiple-comparisons test, Dunnett's
test was selected if one column represented control data and
Tukey's test was chosen when comparing all pairs of columns.
Two-way ANOVA with Bonferroni's test for multiple comparisons was
used when there were two independent variables. P<0.05 was
considered to indicate a statistically significant difference.
Results
Differentiation and apoptosis of HL-60
cells under ATRA and DMSO
Since the HL-60 differentiation time was different
in previous studies (13,15,20-22),
the present study aimed to determine the differentiation protocol
that led to the highest expression of the neutrophil surface
marker, while maintaining high cell viability. The differentiation
was observed for three to seven days using the most commonly used
differentiation agents, ATRA and DMSO. High concentrations of serum
have been reported to inhibit differentiation (23). X-VIVO™ 15 is a serum-free medium,
which has been reported to improve cell differentiation (24). Hence, the effects of serum
(RPMI-1640 supplemented with 10% FBS) and serum-free medium
(X-VIVO™ 15 medium) on the differentiation of HL-60 cells were also
compared.
To investigate the differentiation efficiency,
CD11b, CD16 and CD66b were used as the neutrophil surface markers
(20,25-27)
and Apoptosis Detection kits were used for the evaluation of cell
viability. Both ATRA and DMSO need five days to produce efficient
expression of CD11b, CD16 and CD66b in dHL-60 cells incubated in
normal medium with serum or X-VIVO™ 15 medium (Fig. 1A-N). With increasing differentiation
time, the expression of CD11b, CD16 and CD66b decreased in certain
groups (Fig. 1M-O). The association
between the differentiation time and the survival rate of cells was
determined and the results indicated that the cell viability was
similar from the third to the fifth day; however, cell death
increased significantly from the sixth day (Fig. 1P).
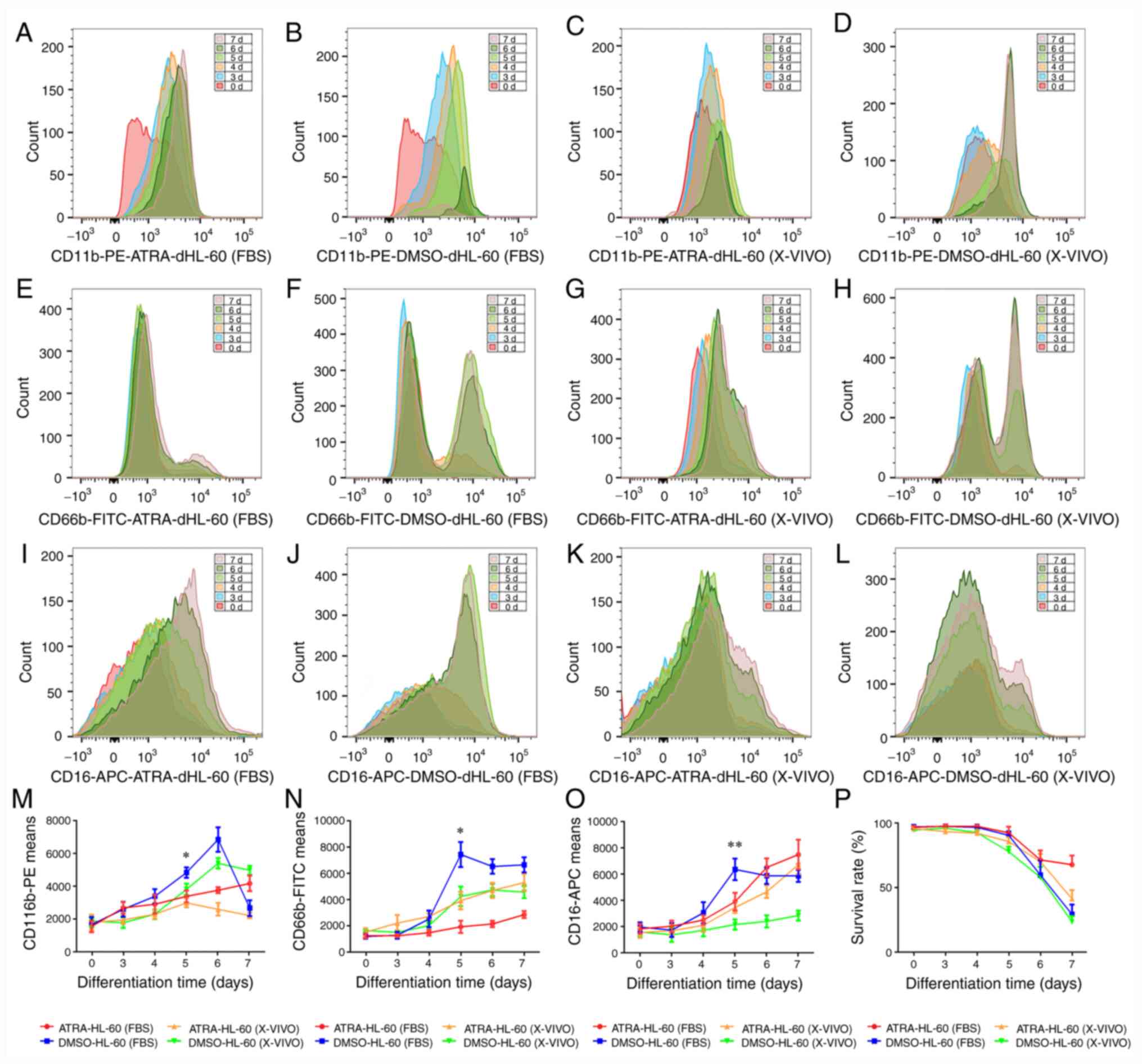 | Figure 1Differentiation and apoptosis of
HL-60 cells in the presence of ATRA and DMSO. HL-60 cells were
differentiated into a neutrophil-like state by stimulation with
ATRA (1 µmol/l) or DMSO (1.25%) for three to seven days in
serum-containing medium (RPMI-1640 supplemented with 10% FBS) or
serum replacement medium (X-VIVO™ 15 Medium). (A-D) CD11b, (E-H)
CD16 and (I-L) CD66b expression was used as a neutrophil surface
marker to measure the differentiation state in (A, E, I)
ATRA-dHL-60 (FBS), (B, F and J) DMSO-dHL-60 (FBS), (C, G and K)
ATRA-dHL-60 (X-VIVO) and (D, H and L) DMSO-dHL-60 (X-VIVO). (M-O)
CD11b, CD66b and CD16 expression was compared between the four
protocols during three to seven days of differentiation; one-way
ANOVA with Dunnett's test for multiple comparisons was used for
comparing the expression of surface markers on the fifth
differentiation day. (M) CD11b expression in was significantly
higher in the DMSO-dHL-60 (FBS) group than in the ATRA-dHL-60
(X-VIVO) group, *P<0.05. (N) The expression of CD66b
was also significantly higher in the DMSO-dHL-60 (FBS) group
compared with that in the ATRA-dHL-60 (FBS) group,
*P<0.05. (O) The DMSO-dHL-60 (FBS) group had the
highest expression of CD16 among the groups;
**P<0.01. (P) Cell viability was similar from the
third day to the fifth day and decreased significantly from the
sixth day. Samples were measured using flow cytometry and data were
analyzed using FlowJo software. The experiments were performed
three times and a representative experiment is presented. ATRA,
all-trans retinoic acid; DMSO, dimethyl sulfoxide; dHL-60,
differentiated HL-60 cells; d, days. |
Since induction for five days produced the best
results in terms of differentiation markers and cell viability, the
expression of CD11b, CD16, and CD66b was compared in different
groups. The group with the best differentiation potential
[DMSO-dHL-60 (FBS)] was used as a representative control group. It
was observed that CD11b expression was significantly higher in the
DMSO-dHL-60 (FBS) group than in the ATRA-dHL-60 (X-VIVO) group
(Fig. 1M). The expression of CD66b
was also significantly higher in the DMSO-dHL-60 (FBS)group as
compared with that in the ATRA-dHL-60 (FBS) group (Fig. 1N). After induction for 5 days, the
DMSO-dHL-60 (FBS) group had the highest expression of CD16 among
the groups (Fig. 1O).
In summary, induction for five days provided the
best results in terms of differentiation markers and cell
viability, while among the four methods studied, the
differentiation protocol involving the induction of differentiation
using DMSO in serum-containing medium produced the best
differentiation potential.
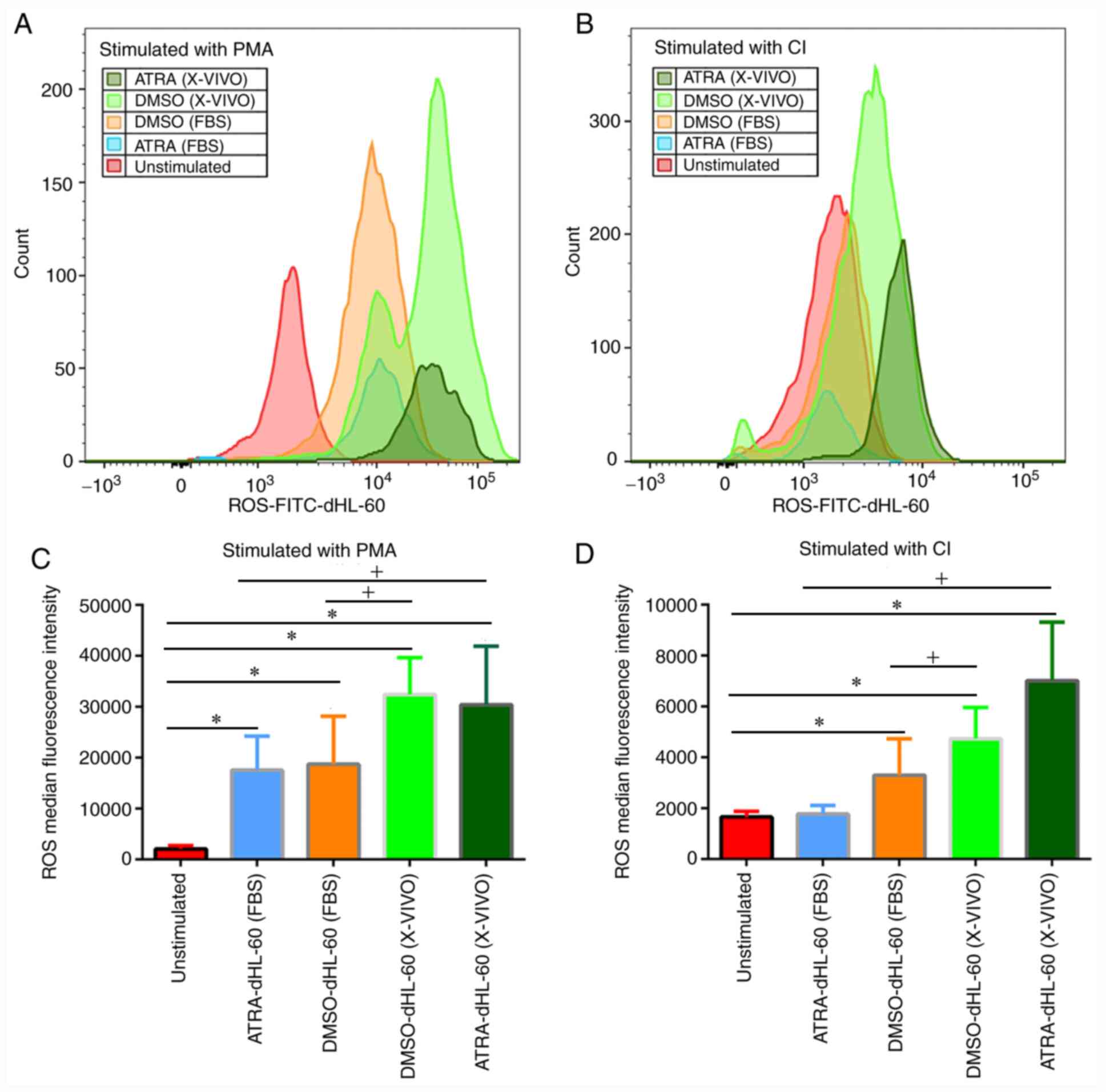
ROS formation in dHL-60 cells after
stimulation with PMA or CI
In the above experiments, it was determined that
induction for five days produced the best results in terms of
differentiation markers and cell viability. In subsequent
experiments, the ability of dHL-60 cells to produce ROS after five
days of differentiation was tested. ROS formation was analyzed
using Total Reactive Oxygen Species Assay kits, which contain the
necessary reagents and buffers for identifying ROS in cells by flow
cytometry in the FITC channel. It was determined that all four
different protocols may induce HL-60 cells to produce ROS upon
stimulation with PMA (Fig. 2A and
C). Both ATRA-dHL-60 and
DMSO-dHL-60 cells cultured in serum-free medium released more ROS
than the cells in serum-containing medium. DMSO-dHL-60 (X-VIVO) was
the most efficient of the four groups (Fig. 2C). Under stimulation with CI, the
ATRA-dHL-60 (FBS) group was not able to form ROS. ROS formation was
more effective in serum-free medium than in serum-containing medium
with the same agents and ATRA-dHL-60 (X-VIVO) was the most
efficient of all four groups (Fig.
2B and D).
Stimulation of differentiated HL-60
cells in serum-free medium results in highly efficient NETs
production
In the above experiments, it was determined that ROS
formation was more effective in serum-free medium than in
serum-containing medium. As the release of NETs occurs downstream
of ROS generation, the ability of dHL-60 cells to produce NETs
after stimulation was tested in subsequent experiments. By
fluorescent microscopy, it was observed that all four
differentiation protocols are able to release NETs upon stimulation
by both PMA and CI (Fig. 3A).
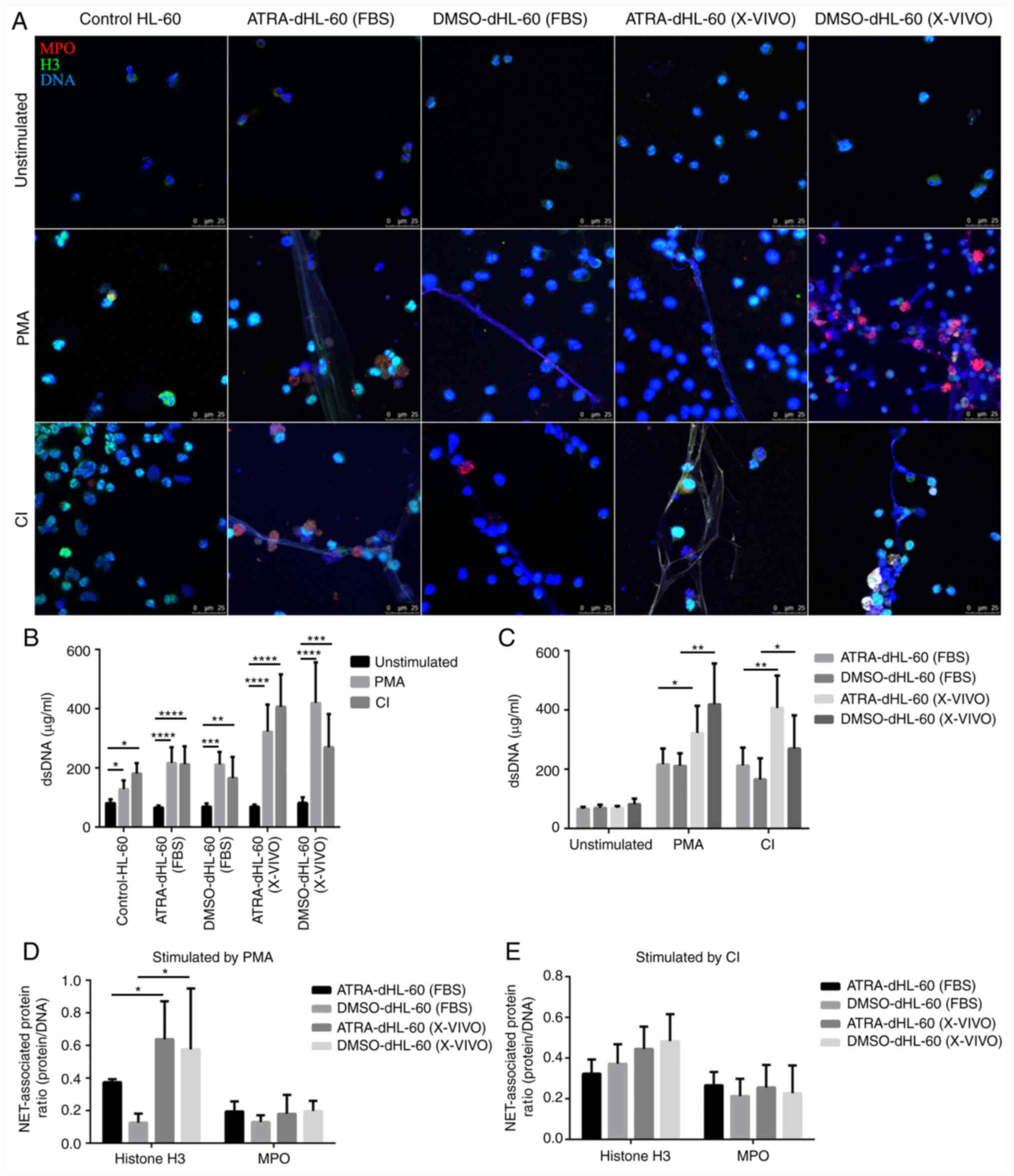 | Figure 3NETs released from dHL-60 post-PMA
and CI stimulation. (A) NETs formation was detected by fluorescent
microscopy. Samples were stained with antibodies against MPO (red)
and histone H3 (green). DNA was counterstained with DAPI (original
magnification, x20; scale bar, 25 µm). (B and C) DNA release was
detected by PicoGreen. (B) One-way ANOVA with Dunnett's test for
multiple comparisons was used for comparing the DNA release between
the stimulated and unstimulated groups. All four different
protocols can induce HL-60 cells to produce NETs upon stimulation
with PMA and CI. *P<0.05, **P<0.01,
***P<0.001 and ****P<0.0001. (C)
One-way ANOVA with Tukey's test for multiple comparisons was used
for comparing the DNA release between the serum-containing medium
group and the serum-free medium group with the group with the same
differentiation agents, *P<0.05,
**P<0.01. In B and C, values are expressed as the
mean ± standard error of the mean from six independent experiments.
(D and E) The composition of NETs was detected by calculating the
areas of NET-associated proteins (MPO and histone H3) divided by
DNA area. (D) Stimulation with PMA and compared between the
serum-containing medium group and serum-free medium group with the
same differentiation agents, *P<0.05. (E) Stimulation
with CI and compared between the serum-containing medium group and
serum-free medium group with the same differentiation agents. Data
were measured using ImageJ software. NETs, neutrophil extracellular
traps; MPO, myeloperoxidase; dHL-60, differentiated HL-60 cells;
ATRA, all-trans retinoic acid; DMSO, dimethyl sulfoxide; PMA,
phorbol 12-myristate 13-acetate; CI, Ca2+ ionophore;
dsDNA, double-strand DNA. |
Quantification of NETs using PicoGreen indicated
that these cells release DNA when stimulated with PMA or CI;
however, differentiated HL-60 cells in serum-free medium appeared
to produce more NETs than those in serum-containing medium
(Fig. 3B). Therefore, the numerical
value of NETs was compared according to the medium in which the
cells were incubated. The results indicated that both ATRA and DMSO
dHL-60 cells in serum-free medium formed more NETs than those in
serum-containing medium upon stimulation with PMA or CI. The
DMSO-dHL-60 (X-VIVO) group was the most efficient in releasing NETs
post-PMA stimulation, while the ATRA-dHL-60 (X-VIVO) group was the
most efficient in releasing NETs post-CI stimulation (Fig. 3C).
Next, the composition of NETs was compared by
calculating the areas of NET-associated proteins (MPO and histone
H3) divided by DNA areas determined using fluorescent microscopy
(6). When dHL-60 cells were
stimulated with PMA, both ATRA and DMSO dHL-60 groups in serum-free
medium released a greater ratio of histone H3 to DNA than those in
serum-containing medium (Fig. 3D);
however, the ratio of MPO expression exhibited no difference. There
was no statistically significant difference in the composition of
NET-associated proteins between the groups upon CI stimulation
(Fig. 3E). Furthermore, the
composition of NET-associated proteins differed for different
stimulation agents (data not shown).
Histone citrullination in dHL-60 cells
after stimulation with PMA or CI
The conversion of arginine into citrulline by PADI4
may contribute to chromatin relaxation, which is frequently
required for NET extrusion via suicidal and vital NETosis (28). To evaluate the ability for histone
citrullination in different protocols after stimulation, the
formation of citH3 was detected using a flow cytometer. The results
indicated that none of the dHL-60 cells contained citH3 upon PMA
treatment (Fig. 4A and C). After CI treatment, the proportion of
citH3-positive cells among dHL-60 cells was 10.4% in the
ATRA-dHL-60 (FBS), 6.58% in the DMSO-dHL-60 (FBS), 32.8% in the
ATRA-dHL-60 (X-VIVO) and 18.4% in the DMSO-dHL-60 (X-VIVO) group;
however, ATRA-dHL-60 (X-VIVO) was the most effective group
regarding citH3 formation after CI stimulation (Fig. 4B and C).
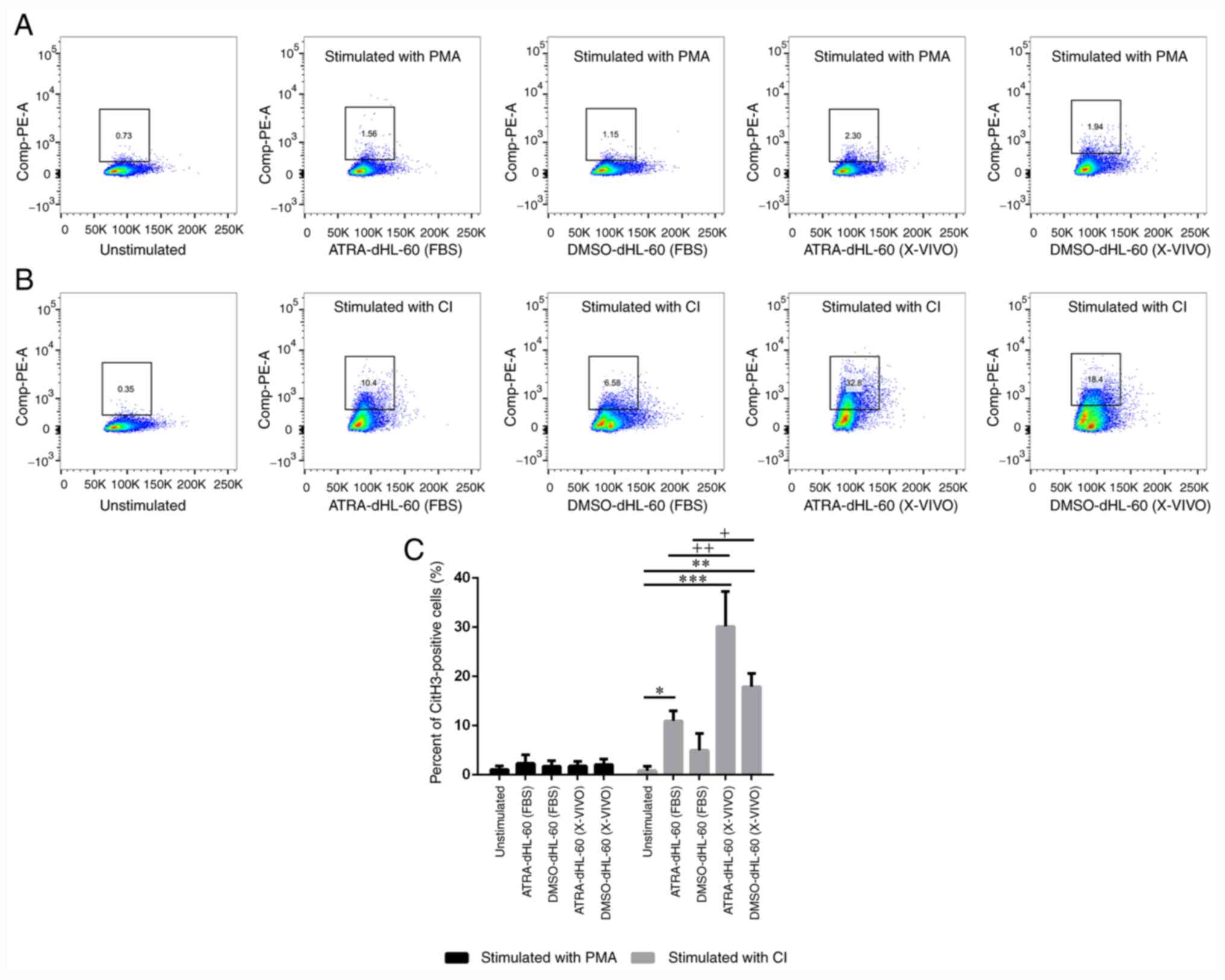 | Figure 4Histone citrullination in dHL-60
cells after stimulation. (A and B) Representative flow cytometry
images for the identification of citH3-positive cells defined as
NETs. (A) Stimulation with PMA and (B) stimulation with CI. (C)
Two-way ANOVA with Bonferroni's test for multiple comparisons was
used for comparing the percentage of citH3-positive cells among the
groups. After stimulation with CI, the percentage of citH3-positive
cells was significantly higher in ATRA-dHL-60 (FBS), ATRA-dHL-60
(X-VIVO) and DMSO-dHL-60 (X-VIVO) groups as compared with that in
unstimulated group, *P<0.05 **P<0.01
and ***P<0.001. The percentage of citH3-positive
cells was significantly higher in serum-free group as compared with
that in serum-containing group [DMSO-dHL-60 (X-VIVO) vs.
DMSO-dHL-60 (FBS); +P<0.05 and ATRA-dHL-60 (X-VIVO)
vs. ATRA-dHL-60 (FBS), ++P<0.01] when stimulation
with CI. NETs, neutrophil extracellular traps; dHL-60,
differentiated HL-60 cells; ATRA, all-trans retinoic acid; DMSO,
dimethyl sulfoxide; PMA, phorbol 12-myristate 13-acetate; CI,
Ca2+ ionophore; citH3, citrullinated histone H3. |
Discussion
In the present study, the ability of dHL-60 cells to
form NETs under different differentiation conditions and with
different stimulation agents was assessed. The experimental
evidence suggested that the optimal experimental protocol involved
differentiation of HL-60 cells for five days and culture in
serum-free medium. With respect to the ability of dHL-60 cells to
release NETs, DMSO-dHL-60 (X-VIVO) cells were most efficient at
producing NETs and ROS upon PMA stimulation, while ATRA-dHL-60
(X-VIVO) cells were most efficient at producing NETs and citH3 upon
CI stimulation.
The biology of neutrophils remains to be fully
elucidated and the short life span of neutrophils makes it
difficult to study their functions. HL-60 cells are a human myeloid
leukemia cell line, which may undergo granulocytic differentiation
after stimulation using agents such as ATRA and DMSO (13,29-33).
There are various differentiation conditions. For stimulation using
DMSO, Song et al (32) and
Wang et al (34) treated
HL-60 cells for three days, Gupta et al (22) suggested incubation for three to four
days, Manda-Handzlik et al (13) treated the cells for five days and
Razvina et al (16) treated
the cells for seven days. The treatment time with ATRA was also
different in previous studies (13,20,22,32).
Since neutrophilic differentiation is frequently coordinated with
cell death, exploration of HL-60 differentiation conditions is
conducive to effective differentiation. The present results
indicated that incubation with DMSO or ATRA for five days was the
most suitable period for inducing neutrophil marker expression and
cell viability.
NETs have critical roles in the host defense
mechanism mediated by neutrophils; therefore, numerous studies have
used dHL-60 to explore the molecular and cellular mechanisms of
NETs formation (14,16,17,29).
However, the capacity of dHL-60 to release NETs was weak compared
with that of neutrophils (13,30). A
maximum of 28% of NETs release was reached after 4 h of
co-incubation of dHL-60 with PMA, while blood-derived neutrophils
produced almost 100% NETs after 4 h of stimulation (30). It is important to increase the
ability of dHL-60 cells to release NETs. High concentrations of
serum have been reported to inhibit differentiation (23). To suppress the adverse effects of
serum, a serum-free medium (X-VIVO™ 15 medium), which has been
reported to improve cell differentiation (24), was used in the present study. The
present results confirmed that the ability of dHL-60 cells cultured
in serum-free media to release NETs upon PMA or CI stimulation was
greater than that of cells cultured in serum-containing media.
PMA and CI are commonly used for the study of
NETosis. PMA is able to activate numerous pathways in neutrophils
and mediate suicidal NETosis. Upon treatment with PMA, NADPH
oxidase is activated via protein kinase C and the Raf-MAPK
kinase-ERK signaling pathway, leading to ROS generation and MPO
activation and resulting in chromatin decondensation in the nucleus
(7,35). In this cell death process, the
generation of ROS by NADPH oxidase is critical. Consistent with the
weak ability of dHL-60 cells to release NETs, they do not release
ROS as vigorously as neutrophils (29,30,36).
Imaizumi and Breitman (33)
reviewed several studies and concluded that the capacity for ROS
production by DMSO-dHL-60 cells was stronger than that of
ATRA-dHL-60 cells upon PMA stimulation. Sham et al (31) reported the opposite result. In the
present study, it was observed that the capacity for ROS production
was similar between DMSO-dHL-60 (FBS) cells and ATRA-dHL-60 (FBS)
cells; however, ROS formation was more effective in serum-free
medium than in serum-containing medium with the same agents and
DMSO-dHL-60 (X-VIVO) was the most effective.
Neutrophils may form NETs independent of oxidant
production. CI-induced NET formation is dependent on AKT signaling
and calcium-induced mitochondrial ROS, but not on the NADPH oxidase
pathway (18,37). The present study also determined
that the ROS release stimulated by CI was much lower than with PMA.
Serum may influence the generation of ROS and NET formation
(38) and the presence of DNase I
in serum also affects the degradation of NETs (39). To avoid any interference of serum
with ROS and NET generation, all dHL-60 cells cultured in media
were washed and cultured with RPMI-1640 during stimulation.
The formation of NETs is a process dependent on both
ROS formation and histone citrullination. It was indicated that
dHL-60 cells only exhibited histone citrullination upon CI
stimulation but not upon PMA stimulation. This result was
consistent with the results of previous neutrophil stimulation
experiments using CI and PMA (18).
The present study indicated that citH3 formation after CI
stimulation was highest in the ATRA-dHL-60 (X-VIVO) group. Song
et al (32) reported that
ATRA stimulation led to significant upregulation of PADI4
expression at both the mRNA and protein levels in HL-60 cells in a
time-dependent manner. It was speculated that the effective histone
citrullination ability of ATRA-dHL-60 cells may be due to the
upregulation of PADI4.
In conclusion, the optimal experimental protocol for
HL-60 cells was differentiation for five days and culture in
serum-free medium. The DMSO-dHL-60 (X-VIVO) group was most
efficient in terms of NETs and ROS production upon PMA stimulation,
while the ATRA-dHL-60 (X-VIVO) group was most efficient in terms of
NETs and citH3 generation upon CI stimulation. Based on the
different pathways of PMA and CI, DMSO-dHL-60 (X-VIVO) may be
proposed as a model for the study of ROS-high NETosis, while
ATRA-dHL-60 (X-VIVO) may be suitable for ROS-low NETosis.
Acknowledgements
The authors would like to thank Mrs Ying Yin
(Department of Central Laboratory, Wuxi People's Hospital
Affiliated to Nanjing Medical University, Wuxi, China) for
performing the flow cytometry detection and Miss Jianxin Tan
(Department of Central Laboratory, Wuxi People's Hospital
Affiliated to Nanjing Medical University, Wuxi, China) for
acquiring laser confocal microscopy images.
Funding
Funding: This study was funded by the Medical Innovation Team of
Jiangsu Province (grant no. CXTDB 2017016), the General Program of
Nanjing Medical University (grant no. 2017NJMUZD119), Wuxi
Municipal Bureau on Science and Technology (grant no. NZ2019026),
the Young Project of Wuxi Health and Family Planning Commission
(grant no. Q202054) and the Major Program of Wuxi Health and Family
Planning Commission (grant no. Z202016).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LL and DZ designed, supervised the present study and
can authenticate the raw data in the study. YG and FG performed the
experiments and were involved in writing the manuscript. QW
performed quality control and adjustment on the flow cytometry
experiment. KW conducted cell culture and differentiation. SP
analyzed the flow cytometry results. ZP completed the statistical
analyzes of all results. SX analyzed the fluorescence microscopy
data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Soehnlein O, Steffens S, Hidalgo A and
Weber C: Neutrophils as protagonists and targets in chronic
inflammation. Nat Rev Immunol. 17:248–261. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Castanheira FVS and Kubes P: Neutrophils
and NETS in modulating acute and chronic inflammation. Blood.
133:2178–2185. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Lefrancais E, Mallavia B, Zhuo H, Calfee
CS and Looney MR: Maladaptive role of neutrophil extracellular
traps in pathogen-induced lung injury. JCI Insight.
3(e98178)2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Mohanty T, Fisher J, Bakochi A, Neumann A,
Cardoso JFP, Karlsson CAQ, Pavan C, Lundgaard I, Nilson B,
Reinstrup P, et al: Neutrophil extracellular traps in the central
nervous system hinder bacterial clearance during pneumococcal
meningitis. Nat Commun. 10(1667)2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Tanaka M, Kinoshita-Daitoku R, Kiga K,
Sanada T, Zhu B, Okano T, Aikawa C, Iida T, Ogura Y, Hayashi T, et
al: Group A streptococcus establishes pharynx infection by
degrading the deoxyribonucleic acid of neutrophil extracellular
traps. Sci Rep. 10(3251)2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
van Dam LS, Kraaij T, Kamerling SWA,
Bakker JA, Scherer UH, Rabelink TJ, van Kooten C and Teng YKO:
Intrinsically distinct role of neutrophil extracellular trap
formation in antineutrophil cytoplasmic antibody-associated
vasculitis compared to systemic lupus erythematosus. Arthritis
Rheumatol. 71:2047–2058. 2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Brinkmann V, Reichard U, Goosmann C,
Fauler B, Uhlemann Y, Weiss DS, Weinrauch Y and Zychlinsky A:
Neutrophil extracellular traps kill bacteria. Science.
303:1532–1535. 2004.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Fuchs TA, Abed U, Goosmann C, Hurwitz R,
Schulze I, Wahn V, Weinrauch Y, Brinkmann V and Zychlinsky A: Novel
cell death program leads to neutrophil extracellular traps. J Cell
Biol. 176:231–241. 2007.PubMed/NCBI View Article : Google Scholar
|
|
9
|
D'Cruz AA, Speir M, Bliss-Moreau M,
Dietrich S, Wang S, Chen AA, Gavillet M, Al-Obeidi A, Lawlor KE and
Vince JE: The pseudokinase MLKL activates PAD4-dependent NET
formation in necroptotic neutrophils. Sci Signal.
11(eaao1716)2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Yousefi S, Mihalache C, Kozlowski E,
Schmid I and Simon HU: Viable neutrophils release mitochondrial DNA
to form neutrophil extracellular traps. Cell Death Differ.
16:1438–1444. 2009.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Pilsczek FH, Salina D, Poon KK, Fahey C,
Yipp BG, Sibley CD, Robbins SM, Green FH, Surette MG, Sugai M, et
al: A novel mechanism of rapid nuclear neutrophil extracellular
trap formation in response to Staphylococcus aureus. J Immunol.
185:7413–7425. 2010.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Chen KW, Monteleone M, Boucher D,
Sollberger G, Ramnath D, Condon ND, von Pein JB, Broz P, Sweet MJ
and Schroder K: Noncanonical inflammasome signaling elicits
gasdermin D-dependent neutrophil extracellular traps. Sci Immunol.
3(eaar6676)2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Manda-Handzlik A, Bystrzycka W, Wachowska
M, Sieczkowska S, Stelmaszczyk-Emmel A, Demkow U and Ciepiela O:
The influence of agents differentiating HL-60 cells toward
granulocyte-like cells on their ability to release neutrophil
extracellular traps. Immunol Cell Biol. 96:413–425. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Manda-Handzlik A, Bystrzycka W, Cieloch A,
Glodkowska-Mrowka E, Jankowska-Steifer E, Heropolitanska-Pliszka E,
Skrobot A, Muchowicz A, Ciepiela O, Wachowska M and Demkow U:
Nitric oxide and peroxynitrite trigger and enhance release of
neutrophil extracellular traps. Cell Mol Life Sci. 77:3059–3075.
2020.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Obama T, Ohinata H, Takaki T, Iwamoto S,
Sawada N, Aiuchi T, Kato R and Itabe H: Cooperative action of
oxidized low-density lipoproteins and neutrophils on endothelial
inflammatory responses through neutrophil extracellular trap
formation. Front Immunol. 10(1899)2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Razvina O, Jiang S, Matsubara K, Ohashi R,
Hasegawa G, Aoyama T, Daigo K, Kodama T, Hamakubo T and Naito M:
Differential expression of pentraxin 3 in neutrophils. Exp Mol
Pathol. 98:33–40. 2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Alyami HM, Finoti LS, Teixeira HS, Aljefri
A, Kinane DF and Benakanakere MR: Role of NOD1/NOD2 receptors in
Fusobacterium nucleatum mediated NETosis. Microb Pathog. 131:53–64.
2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Douda DN, Khan MA, Grasemann H and
Palaniyar N: SK3 channel and mitochondrial ROS mediate NADPH
oxidase-independent NETosis induced by calcium influx. Proc Natl
Acad Sci USA. 112:2817–2822. 2015.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Gavillet M, Martinod K, Renella R, Harris
C, Shapiro NI, Wagner DD and Williams DA: Flow cytometric assay for
direct quantification of neutrophil extracellular traps in blood
samples. Am J Hematol. 90:1155–1158. 2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Rincon E, Rocha-Gregg BL and Collins SR: A
map of gene expression in neutrophil-like cell lines. BMC Genomics.
19(573)2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Liu CL, Tangsombatvisit S, Rosenberg JM,
Mandelbaum G, Gillespie EC, Gozani OP, Alizadeh AA and Utz PJ:
Specific post-translational histone modifications of neutrophil
extracellular traps as immunogens and potential targets of lupus
autoantibodies. Arthritis Res Ther. 14(R25)2012.PubMed/NCBI View
Article : Google Scholar
|
|
22
|
Gupta D, Shah HP, Malu K, Berliner N and
Gaines P: Differentiation and characterization of myeloid cells.
Curr Protoc Immunol. 104:22F.5.1–22F.5.28. 2014.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Pedruzzi E, Fay M, Elbim C and
Gougerot-Pocidalo MA: Differentiation of PLB-985 myeloid cells into
mature neutrophils, shown by degranulation of terminally
differentiated compartments in response to N-formyl peptide and
priming of superoxide anion production by granulocyte-macrophage
colony-stimulating factor. Br J Haematol. 117:719–726.
2002.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Chen L, Xie XY, Nie JQ, Chen DL, Huang AP,
Fang F, Qu MY, Nan X, He LJ, Fan Z, et al: Mononuclear cells of
umbilical cord blood differentiation to granulocyte cell in vitro.
Zhonghua Xue Ye Xue Za Zhi. 38:532–536. 2017.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
25
|
Perdomo J, Leung HH, Ahmadi Z, Yan F,
Chong JJ, Passam FH and Chong BH: Neutrophil activation and NETosis
are the major drivers of thrombosis in heparin-induced
thrombocytopenia. Nat Commun. 10(1322)2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Lelliott PM, Momota M, Shibahara T, Lee
MS, Smith NI, Ishii KJ and Coban C: Heparin induces neutrophil
elastase dependent vital and lytic NET formation. Int Immunol.
32:359–368. 2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Lood C, Blanco LP, Purmalek MM,
Carmona-Rivera C, De Ravin SS, Smith CK, Malech HL, Ledbetter JA,
Elkon KB and Kaplan MJ: Neutrophil extracellular traps enriched in
oxidized mitochondrial DNA are interferogenic and contribute to
lupus-like disease. Nat Med. 22:146–153. 2016.PubMed/NCBI View
Article : Google Scholar
|
|
28
|
Burgener SS and Schroder K: Neutrophil
extracellular traps in host defense. Cold Spring Harb Perspect
Biol. 12(a037028)2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Wang Y, Li M, Stadler S, Correll S, Li P,
Wang D, Hayama R, Leonelli L, Han H, Grigoryev SA, et al: Histone
hypercitrullination mediates chromatin decondensation and
neutrophil extracellular trap formation. J Cell Biol. 184:205–213.
2009.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Yaseen R, Blodkamp S, Lüthje P, Reuner F,
Völlger L, Naim HY and von Köckritz-Blickwede M: Antimicrobial
activity of HL-60 cells compared to primary blood-derived
neutrophils against Staphylococcus aureus. J Negat Results Biomed.
16(2)2017.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Sham RL, Phatak PD, Belanger KA and
Packman CH: Functional properties of HL60 cells matured with
all-trans-retinoic acid and DMSO: Differences in response to
interleukin-8 and fMLP. Leuk Res. 19:1–6. 1995.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Song G, Shi L, Guo Y, Yu L, Wang L, Zhang
X, Li L, Han Y, Ren X, Guo Q, et al: A novel PAD4/SOX4/PU.1
signaling pathway is involved in the committed differentiation of
acute promyelocytic leukemia cells into granulocytic cells.
Oncotarget. 7:3144–3157. 2016.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Imaizumi M and Breitman TR: Retinoic
acid-induced differentiation of the human promyelocytic leukemia
cell line, HL-60, and fresh human leukemia cells in primary
culture: A model for differentiation inducing therapy of leukemia.
Eur J Haematol. 38:289–302. 1987.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Wang Y, Wysocka J, Sayegh J, Lee YH,
Perlin JR, Leonelli L, Sonbuchner LS, McDonald CH, Cook RG, Dou Y,
et al: Human PAD4 regulates histone arginine methylation levels via
demethylimination. Science. 306:279–283. 2004.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Steinberg SF: Structural basis of protein
kinase C isoform function. Physiol Rev. 88:1341–1378.
2008.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Remijsen Q, Vanden Berghe T, Wirawan E,
Asselbergh B, Parthoens E, De Rycke R, Noppen S, Delforge M,
Willems J and Vandenabeele P: Neutrophil extracellular trap cell
death requires both autophagy and superoxide generation. Cell Res.
21:290–304. 2011.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Parker H, Dragunow M, Hampton MB, Kettle
AJ and Winterbourn CC: Requirements for NADPH oxidase and
myeloperoxidase in neutrophil extracellular trap formation differ
depending on the stimulus. J Leukoc Biol. 92:841–849.
2012.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Kamoshida G, Kikuchi-Ueda T, Nishida S,
Tansho-Nagakawa S, Kikuchi H, Ubagai T and Ono Y: Spontaneous
formation of neutrophil extracellular traps in serum-free culture
conditions. FEBS Open Bio. 7:877–886. 2017.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Hakkim A, Furnrohr BG, Amann K, Laube B,
Abed UA, Brinkmann V, Herrmann M, Voll RE and Zychlinsky A:
Impairment of neutrophil extracellular trap degradation is
associated with lupus nephritis. Proc Natl Acad Sci USA.
107:9813–9818. 2010.PubMed/NCBI View Article : Google Scholar
|