Introduction
The current coronavirus disease 2019 (COVID-19)
pandemic emerged in late 2019 in Wuhan, China and has spread
rapidly throughout the world. It is estimated that 20% of
individuals infected with the causative viral agent, severe acute
respiratory syndrome coronavirus 2 (SARS-CoV-2), develop COVID-19
disease, which is causing the collapse and saturation of health
systems in several countries (1).
The approximate fatality rate of the disease ranges from 4 to 10%
and the associated symptoms include high fever, dry cough,
tiredness, sore throat, diarrhea, conjunctivitis, smell or taste
loss and, in more serious conditions, difficulty breathing and the
inability to speak and move (2).
Severe disease is related to the presence of comorbidities, such as
hypertension, diabetes, cardiovascular diseases and respiratory
diseases (3). Mexico, in
particular, has a high rate of hypertension, obesity and diabetes,
placing its population in an unfavorable condition to deal with the
pandemic (4).
SARS-CoV-2 is a single-stranded RNA virus ~65-125 nm
in diameter, which can be transmitted by air or by contact with
contaminated surfaces. The main routes of entry into the body are
the nasal, oral or ocular routes (5). Once the virus enters the body, it will
infect mucosa, in particular the nasal and laryngeal mucosa
(6). As the infection progresses,
it travels to the lungs through the respiratory tract (7). From the lungs, the virus can enter the
peripheral blood, leading to viremia and the attack of other organs
(8). The host receptor protein used
by the virus to infect cells is angiotensin-converting enzyme 2
(ACE2) (9,10) and expression of this receptor has
been observed in various tissues, such as the heart, liver,
testicles, kidneys and intestines. Typically, the ACE2 protein is
responsible for the regulation of heart and kidney function, as
well as the control of blood pressure (11).
Among the 14 open reading frames (ORFs) identified
in the SARS-CoV-2 genome, ORF3a codes for 274 of the amino acid
proteins proposed to be responsible for its high virulence in
humans (12). It has been described
to be similar to ion channel proteins that can help to create a
more permeable membrane in infected cells (13). ORF3a presents a plethora of
activities, among which are the reduction of type I interferon
(IFN) production, fibrinogen induction, production and release of
cytokines, activation of the NF-κB pathway, production of
chemokines, Golgi reticulum fragmentation, endoplasmic reticulum
stress, accumulation of intracellular vesicles and cell death
(14). All of these characteristics
identify ORF3a as an ideal therapeutic target in the fight against
COVID-19.
Vita Deyun® is a solution based on 1 mg
silymarin, 1 mg glutathione (GSH), 1 mg vitamin C and 0.050 mg
selenium (https://vitadeyun.com.mx). The
individual components of this solution have been extensively
investigated for their effects on different biological aspects of
cells and organisms, and they have demonstrated benefits as
anti-inflammatory agents, antioxidants and enhancers of the immune
response, as described in detail below.
Silymarin is a natural compound derived from the
plant milk thistle (Silybum marianum) and it has been
traditionally used to treat hepatological conditions (15). Evidence of its beneficial uses range
from antioxidant and anti-inflammatory to immunomodulatory
activities (16-19).
In addition, it has been shown that silymarin has a low toxicity of
≤40 µg/kg weight and ≤425 µg/ml in cell culture (20). Furthermore, a protective effect
during lung injury has been demonstrated through decreasing the
infiltration of inflammatory cells, suppressing the activity of
myeloperoxidase, decreasing the production of nitric oxide and the
activity of nitric oxide synthase, and reducing the protein levels
of pro-inflammatory mediators, superoxide dismutase, catalase and
GSH peroxidase (21).
GSH is an antioxidant comprising three amino acids,
glutamine, glycine and cysteine, making it a small molecule with
low molecular weight. GSH is normally produced in hepatocytes and
exported into circulation to be transported for use in other cells;
moreover, GSH in plasma is maintained at a very low concentration
because cell uptake by numerous other cells is rapid (22,23).
This process requires the action of gamma-glutamyl transpeptidase,
which is commonly found on the surface of cells (24). There is evidence that GSH deficiency
leads to increased levels of oxidative stress. Indeed, a protective
role of GSH has been demonstrated against inflammatory lung
diseases (25-27).
GSH is essential for numerous innate and adaptive immune system
functions, including proliferation of T lymphocytes (28,29),
phagocytic activity of neutrophils (30), dendritic cell functions (31) and presentation of antigens by
antigen-presenting cells (32). One
of the first steps in antigen degradation and processing is the
reduction of disulfide bonds, which requires GSH (33). Although, GSH inhibits the production
of certain inflammatory cytokines, it is necessary that dendritic
cells maintain an adequate level of IFN-γ (34), an essential condition for host
defense against intracellular pathogens. There are >650 studies
registered in ClinicalTrials.gov (www.clinicaltrials.gov, accessed: 30 December 2020)
claiming to demonstrate the various benefits of using GSH.
Regarding ascorbic acid, there is ample evidence
from in vitro studies, animal experiments and clinical
trials that vitamin C might exert an antiviral effect (35,36).
Based on this finding and since, to the best of our knowledge, no
specific antiviral drug has yet been proven to be truly effective
against COVID-19, the supplementation of micronutrients, such as
vitamin C has become relevant in the management of COVID-19, as it
might improve the immunological response against SARS-CoV-2.
Viral infections are characterized by deficient
micronutrient stores, particularly vitamins and trace elements,
such as zinc, iron, selenium, magnesium and copper. Lifestyle and
appropriate nutrition may offer further antiviral approaches for
public health (37). Selenium, one
of the compounds of Vita Deyun, is a natural trace element
essential to human health and has a key and complex role in the
immune system (38). Selenium has a
history of reducing the incidence and severity of various viral
infections; it is well-documented that selenium deficiency is
associated with higher susceptibility to RNA viral infections and
more severe disease outcomes (39).
As described above, the individual components of
Vita Deyun have been proven to have beneficial effects in viral and
respiratory diseases, which supports the proposal for evaluation of
the effect of Vita Deyun on SARS-CoV-2 ORF3a. Therefore, the aim of
the present study was to evaluate the in vitro effect of
Vita Deyun solution on the expression of ACE2 in diverse
cell lines, particularly in the presence of SARS-CoV-2 ORF3a
expression.
Materials and methods
Cell culture and treatment
HaCaT, DOK, A549, H1299 and Lenti-X 293T cells were
cultured in DMEM medium supplemented with 10% fetal bovine serum,
100 U/ml penicillin, and 1 mg/ml streptomycin. The cultures were
maintained at 37˚C in a 5% CO2 atmosphere. The
aforementioned products were acquired from Gibco®
(Thermo Fisher Scientific, Inc.). HaCaT cells are human
keratinocytes spontaneously transformed from histologically normal
skin and were obtained from Dr Petra Boukamp (German Cancer
Research Center) (40). The DOK
cells are human dysplastic oral keratinocytes (from the tongue) and
were provided by the cell bank of the Center for Scientific
Instrumentation of the University of Granada (41). The A549 and H1299 cell lines are
derived from lung cancer tissues and were acquired from the ATCC.
Lenti-X 293T cells were obtained from Clontech Laboratories, Inc.
Certain cell lines underwent additional authentication using the
Multiplex human Cell Authentication test performed by Multiplexion
GmbH (www.multiplexion.de).
Vita Deyun solution (Envasadora D´Aqua, S.A de C.V.)
contains 1 mg silymarin, 1 mg GSH, 0.05 mg Selenium, 1 mg ascorbic
acid and 0.1 mg sodium benzoate in 100 ml water. For the treatment,
3x106 cells were seeded on p100 petri dishes in 9.5 ml
complete DMEM and left for 24 h. They were then treated with 500 µl
Vita Deyun solution for 4 h and used for RNA isolation. Cells were
cultivated at 37˚C in a 5% CO2 atmosphere, unless
otherwise stated. Cells in culture were continually monitored
microscopically when seeding them or before and after a treatment
by using an inverted microscope Primovert with a x400 magnification
(ZEISS company).
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
For RNA isolation (total RNA), the GeneJet RNA
Purification Kit (Thermo Fisher Scientific, Inc.) was used
according to the manufacturer's protocol. Thereafter, to obtain
cDNA, the Transcriptor First Strand cDNA Synthesis Kit (Roche
Applied Science) was utilized. Briefly, 5 µg total RNA was primed
with oligo(dT) and reverse transcribed as recommended by the
manufacturer's instructions. Specific primers for all analyzed
genes were engineered by utilizing Oligo v.6.0 software (Molecular
Biology Insights, Inc.) and sequences were obtained from the
National Center of Biotechnology Information (NCBI). Ribosomal
protein L32 (RPL32) and ribosomal protein S18 (RPS18) served as
reference genes in the present study. The primer sequences are
presented in Table I. For qPCR
reactions, the Light Cycler 2.0 system and the LightCycler
FastStart DNA Master PLUS SYBR-Green I Kit (Roche Applied Science)
were used. The thermocycling conditions were as follows: A first
denaturalization step of 10 min at 95˚C, followed by 40 cycling
steps of 10 sec at 95˚C, 10 sec using the temperature shown for
each primer pair in Table I, and 12
sec at 72˚C. Expression levels were normalized using the expression
of the reference genes as internal controls. The expression of the
control groups was set as the reference value and the fold-change
variations in gene expression were calculated using the Pfaffl
mathematical model (42).
 | Table IPrimers used for reverse
transcription-quantitative PCR amplification. |
Table I
Primers used for reverse
transcription-quantitative PCR amplification.
| | Primers
(5'-3') | |
|---|
| Gene | Forward | Reverse | Product length
(bp) | Temperature
(˚C) |
|---|
| ACE2 |
TTCTTTGTCACTGCACCTAA |
AATGCCAACCACTATCACTC | 225 | 58 |
| IL-6 |
TACAAAAGTCCTGATCCAGTTC |
AAGAAGGAATGCCCATTAAC | 245 | 60 |
| IL-6R |
GGCACGCCTTGGACAGAATCC |
CCGCAGCTTCCACGTCTTCTT | 252 | 62 |
| IL-18 |
CCCCGGACCATATTTATTATA |
CATGTCCTGGGACACTTCT | 202 | 58 |
| TNF-α |
CCTGTAGCCCATGTTGTAGCA |
GCCTTGGCCCTTGAAGAG | 171 | 62 |
| RPL32 |
GCATTGACAACAGGGTTCGTAG |
ATTTAAACAGAAAACGTGCACA | 320 | 60 |
| RPS18 |
CGATGGGCGGCGGAAAA |
CAGTCGCTCCAGGTCTTCACGG | 283 | 60 |
Lentivirus transfection, transduction
and cell line establishment
Viral particles were produced using Lenti-X 293T
cells. Briefly, 3x106 Lenti-X-293T cells were seeded one
day before transfection. For the transfection assays,
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) was used according to the manufacturer's protocol
and utilizing a mix of Lenti-X HT packaging systems (4 µg) and 1 µg
of one of the following lentiviral vectors, pLVX-Puro (Takara Bio,
Inc.) or the pLVX-EF1alpha-SARS-CoV-2-ORF3a-2xStrep-IRES-Puro
vector (cat no. 141383; Addgene, Inc.) (43). After 48 h, to eliminate detached
cells and cell detritus, the medium containing the viral particles
was filtered through a 0.45-µm polyethersulfone membrane (Merck
KGaA). The titer of viral particles was tested using Lenti-X GoStix
(Takara Bio, Inc.). The medium containing the viral particles was
aliquoted and stored at -80˚C until use. For the transduction
assays, 1x106 target cells were seeded one day before
transduction. The target cells were transduced with 200 µl medium
containing lentiviral particles (~1x106
Inclusion-Forming Units). To obtain stable cell lines, after 72 h
of transduction, the cells were incubated with 0.5 µg/ml puromycin
for 3 weeks. The medium containing puromycin was replaced every
three days; stable cells were cultured under normal conditions
without puromycin. Cells were always cultivated or treated at 37˚C
in a 5% CO2 atmosphere.
Statistical analysis
Data were obtained from three independent
experiments with paired observations (each experiment was performed
simultaneously and in duplicate). The data are presented as the
mean ± standard deviation. The one-way ANOVA test for multiple
comparisons was employed to compare data from the different
treatments. The Bonferroni test was used as post hoc test and
P<0.05 was considered to indicate a statistically significant
difference. Statistical analysis was performed using the GraphPad
prism v.5 software (GraphPad Software, Inc.).
Results
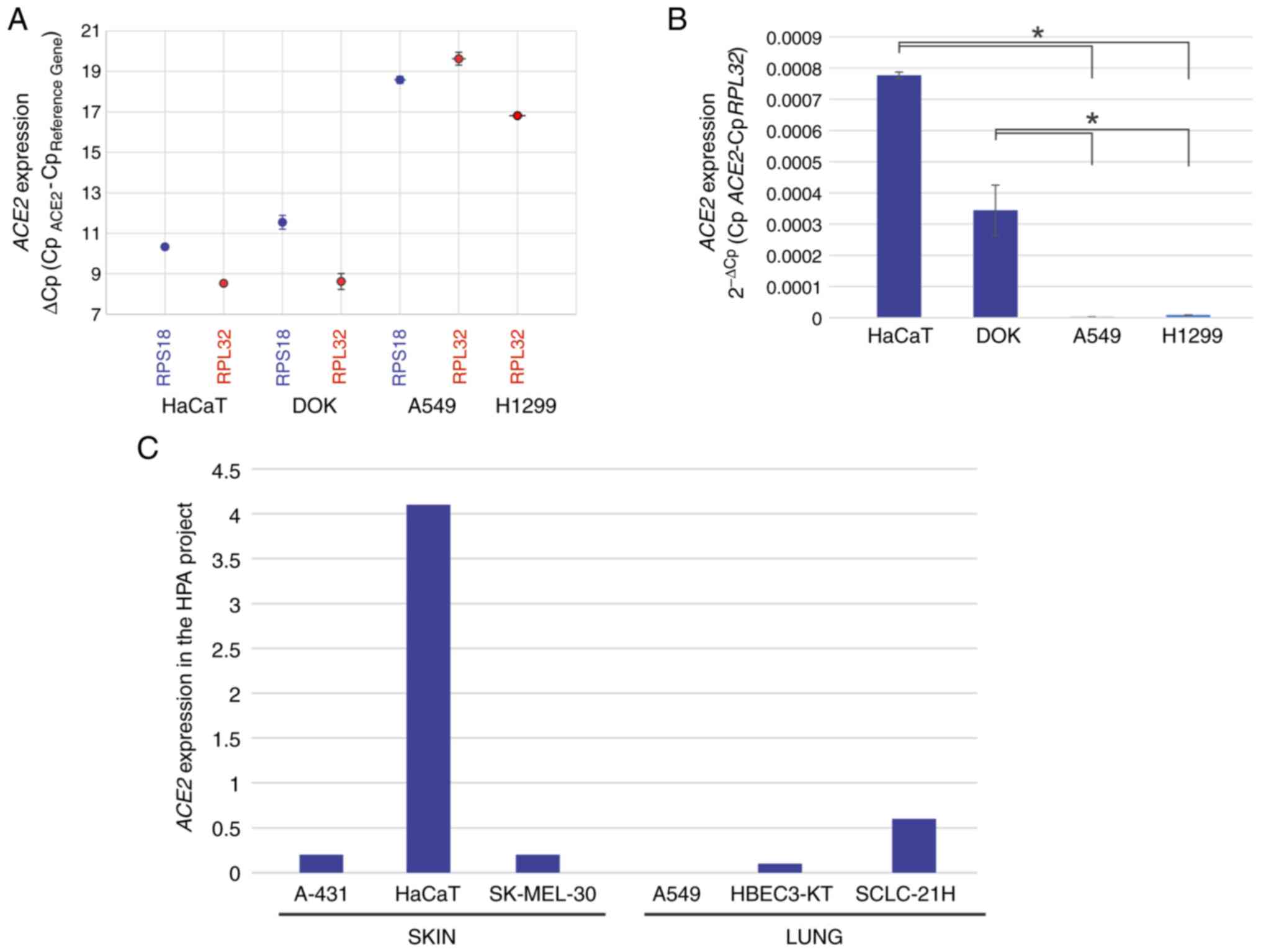
ACE2 is highly expressed in oral and
skin epithelial cells and has low expression in lung cells
With the aim of achieving a suitable study model
that would allow for evaluation of the effects of Vita Deyun
solution on the expression of ACE2 and pro-inflammatory
cytokines, the lung-derived A549 and H1299 cell lines, the oral
epithelial DOK cell line and the skin-derived HaCaT cell line were
analyzed for the expression of ACE2 using RT-qPCR. As
presented in Fig. 1A, the
ΔCp values for ACE2 were 8-12 in HaCaT and DOK
cells, whereas in A549 and H1229 cells, the ΔCp values
were 17-20. As ΔCp values represent an inverse
correlation to expression, a marked decrease in ACE2 levels
was determined in the lung-derived cell lines. For improved
interpretation of these results, the data are presented as
2-ΔCp values (mRNA expression density), as shown in
Fig. 1B. In addition, to
corroborate these results, data regarding the reported expression
of ACE2 was obtained from the Human Protein Atlas (HPA)
project version 20.1 (https://www.proteinatlas.org/ENSG00000130234-ACE2/summary/rna),
where information regarding RNAseq gene expression in different
cell lines is available (RNA summary-Normalized-HPA cell lines). As
presented in Fig. 1C, the
expression of ACE2 was high in HaCaT cells, but not in the
skin-derived A-431 and SK-MEL-30 cell lines included in the HPA
project. By contrast, the lung-derived HBEC3-KT and SCLC-21H cells
exhibited generally low expression of ACE2 and very low
expression was also observed in the A549 cells.
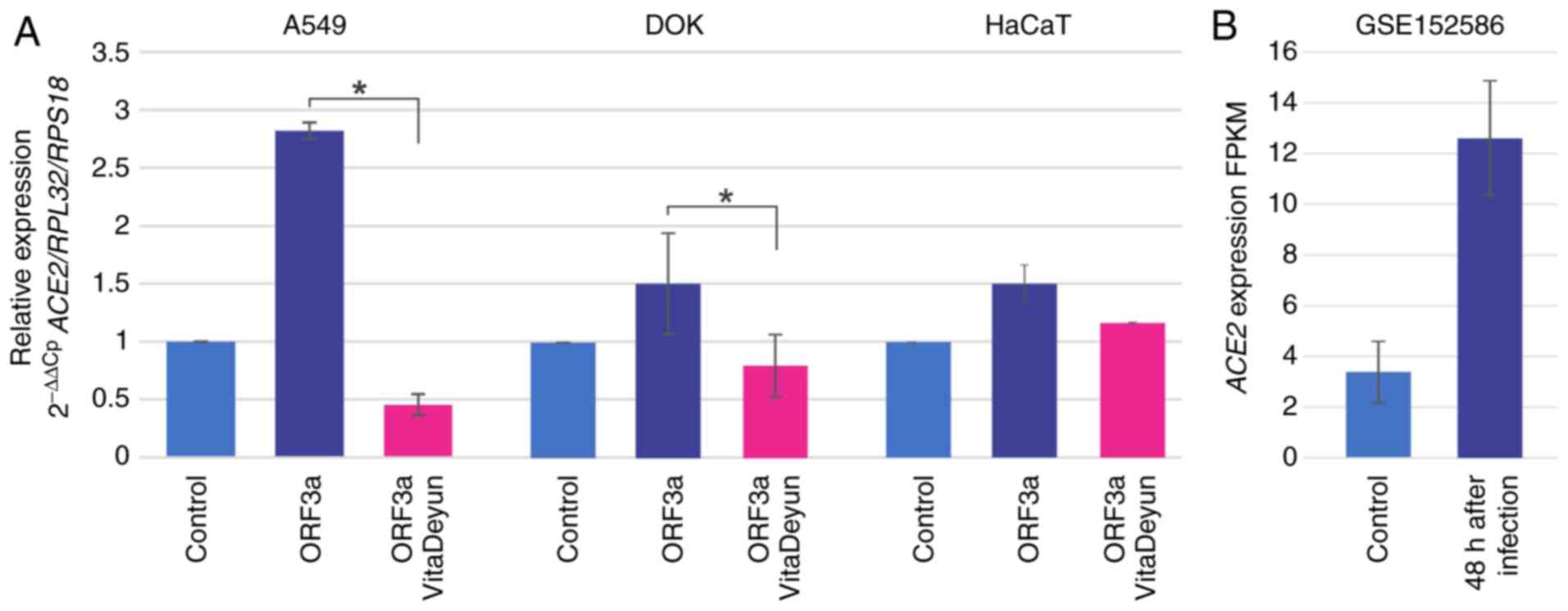
SARS-CoV-2 ORF3a modulates ACE2
expression in oral- and lung-derived epithelial cells and this
effect was hampered by Vita Deyun treatment
To analyze the influence of ORF3a expression in
oral- and lung-derived epithelial cells, DOK and A549 cells were
transduced with lentivirus carrying ORF3a of SARS-CoV-2. HaCaT
cells were included in the present study due to their high levels
of ACE2 mRNA expression. ORF3a induced an increment in
ACE2 mRNA levels in all of the analyzed cells, most notably
in the lung-derived A549 cells, where the expression was increased
by almost 3-fold (Fig. 2A). The
cells were treated for 24 h with the food supplement Vita Deyun to
evaluate whether it modulated the effect of ORF3a on ACE2,
and the expression levels of ACE2 were determined by
RT-qPCR. Notably, it was observed that Vita Deyun treatment
markedly inhibited the expression of ACE2 induced by ORF3a
and, in certain cases, the expression levels of ACE2 were
even lower than the basal levels of the control (untreated) cells
(Fig. 2A). Morphologically, no
change or adverse effect on the growth of the cells was observed in
the presence of Vita Deyun when compared to the untreated cells
(data not shown). The data from the GSE152586 study of the Gene
Expression Omnibus from NCBI (https://www.ncbi.nlm.nih.gov/geo/) was analyzed to
further support the present findings. In the GSE152586 study,
human-derived alveolar organoids were infected with SARS-CoV-2 and,
following 48 h of infection, gene expression was evaluated using
RNAseq (44). The data showed an
evident increase in ACE2 expression following infection
(Fig. 2B).
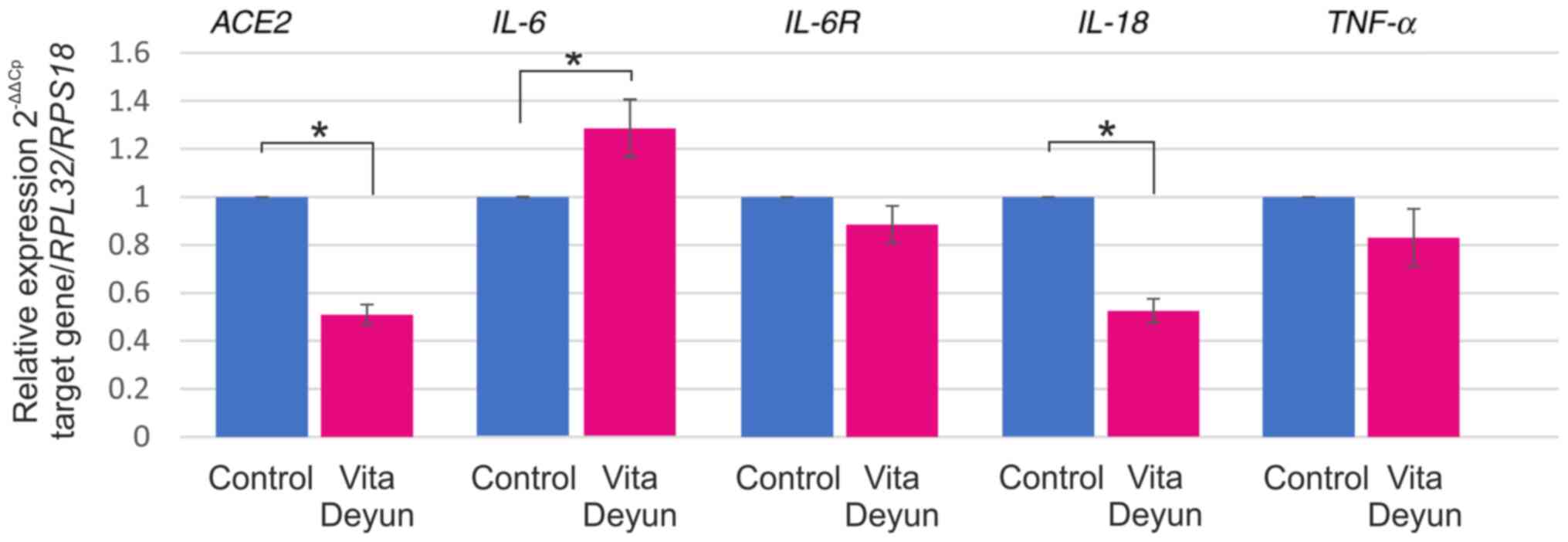
Differential responses in cytokine
expression following Vita Deyun treatment of epithelial cells
constitutively expressing ORF3a
In addition to its effect on ACE2 expression,
the effect of Vita Deyun on the expression of interleukin (IL)-6,
IL-6 receptor (IL-6R), IL-18 and tumor necrosis factor (TNF)-α was
evaluated. A model of A549 cells that constitutively expresses
ORF3a from SARS-CoV-2 was established. Lentiviral particles
carrying ORF3a and a puromycin resistance sequence were generated
and used to transduce A549 cells. The generation of this model was
particularly difficult, since ORF3a expression was toxic for the
cells and a high percentage of dead cells was microscopically
observed (data not shown). Following selection with puromycin, only
a few clones were obtained, and the ORF3a-positive cells were
observed to have a lower proliferation rate (data not shown).
Furthermore, DOK and HaCaT cells were transduced with ORF3a, but
the cells did not survive. The A549 ORF3a-positive cells were
treated with Vita Deyun and the expression of ACE2 and the
aforementioned cytokines was evaluated. As shown in Fig. 3, a significant decrease in the
ACE2 mRNA levels was observed. Whereas a slight increase in
IL-6 mRNA expression was observed. Slight decreases were
observed in IL-6R and TNF-α and a considerable
downregulation of IL-18 was observed.
Discussion
Sufficient evidence exists to indicate that ACE2 is
the main host cell receptor of SARS-CoV-2 for entry into cells to
cause infection. ACE2 expression in oral-, skin- and
lung-derived epithelial cells was observed in the present study.
DOK and HaCaT cells were found to express higher levels of
ACE2 than the analyzed lung-derived cells. Xu et al
(45) also reported high levels of
ACE2 receptor on the epithelial cells of oral mucosa.
A notable observation that was determined in the
present study is that the SARS-CoV-2 ORF3a protein induces the
expression of ACE2. This observation was demonstrated using
two different methodological strategies, as shown in Figs. 2 and 3. To the best of our knowledge, the
present study is the first to demonstrate this effect. The lack of
studies could be partly due to the fact that ORF3a is harmful to
cells; as mentioned in the results section of the present study,
only stably transformed A549 cells constitutively expressing ORF3a
were obtained and similar attempts were not successful using HaCaT
or DOK cell lines due to cell lethality. Recently, Ren et al
(46) demonstrated that ORF3a of
SARS-CoV-2 induces apoptosis in cells.
Moreover, the present study focused on determining
whether the food supplement Vita Deyun is capable of regulating the
expression of ACE2, the receptor reported to be recognized
by SARS-CoV-2(47). As
hypothesized, Vita Deyun reduced the expression of ACE2 in
various cell models (Fig. 2).
One of the components of the dietary supplement Vita
Deyun is silymarin, for which there is evidence of antiviral
properties. Silymarin has been observed to hijack the replication
system of the chikungunya virus, reducing viral replication and
decreasing the production of viral proteins (16-19).
Silymarin also inhibits the entry of hepatitis C virus into its
target cells and, in infected cells, it inhibits the expression of
viral RNA and the production of viral proteins (48). To the best of our knowledge, there
are currently no clinical reports that involve the use of silymarin
against COVID-19; however, there are studies that suggest its
potential use to combat this disease, as mentioned below. Taking
into account the biophysical and structural evidence that spike
proteins possess high binding affinity toward host ACE2 and the
viral hemagglutinin esterase (HE) glycoprotein receptor, Patel
et al (49) used a
combination of various computational approaches to identify
potential inhibitors of HE by employing the naturally occurring
plant-based anticancer compound-activity-target database
(http://crdd.osdd.net/raghava/npact/).
Subsequently, the best-scoring molecules were validated using
molecular dynamics simulations and the authors identified silymarin
(among other compounds) as a potential HE inhibitor with improved
binding energy. Furthermore, Kumar et al (50) demonstrated that the binding of
phytochemicals, such as sarsasapogenin, ursonic acid, curcumin,
ajmalicine, novobiocin and silymarin to the SARS-CoV-2
nonstructural protein 15 protein might play a key role in
inhibiting SARS-CoV-2 replication. In addition, Gorla et al
(51) screened essential flavonoids
against possible protein targets, such as the SARS-CoV-2 spike
glycoprotein receptor-binding domain of the S protein (RBD-S) and
the host ACE2 protease domain (PD-ACE-2) using in silico
molecular docking studies and determined that biochanin A and
silymarin bind significantly at the active sites of RBD-S and
PD-ACE-2.
Another component of Vita Deyun is GSH. Kim et
al (52) applied advanced
bioinformatics computational approaches to identify which existing
Food and Drug Administration-approved drugs could block
coronaviruses from entering cells by binding to ACE2 or
transmembrane protease, serine 2, or attenuate the expression of
genes induced by coronaviruses; the authors identified various ACE
inhibitors, a β-lactam antibiotic, two antiviral agents
(Fosamprenavir and Emricasan) and GSH. A common denominator in
COVID-19 patients appears to be impaired redox homeostasis, which
is responsible for the accumulation of reactive oxygen species. GSH
levels could be critical in extinguishing the exacerbated
inflammation observed in COVID-19 cases. Therefore, it was proposed
by Silvagno et al (53) that
restoring GSH levels in patients with severe symptoms of COVID-19
may be a therapeutic alternative to prevent and control the
disease. In agreement with this, Guloyan et al (54) suggested that the use of liposomal
GSH could be beneficial in COVID-19 patients.
Regarding selenium, Taylor and Radding (55) mentioned that the resulting
collateral damage due to increased oxidative stress and
inflammation would be exacerbated by dietary deficiency of selenium
and GSH precursors. In the review by Hiffler and Rakotoambinina
(39), it was observed that
low-selenium status is common in individuals considered at risk of
developing severe COVID-19, particularly in the elderly. In a study
conducted by Moghaddam et al (56) in Germany, it was shown that serum
selenium levels were highly correlated with COVID-19 outcome in
hospitalized patients and the authors determined that selenium
deficiency is associated with mortality risk from COVID-19.
Vitamin C has been reported to have multiple
pharmacological characteristics, including antiviral, antioxidant,
anti-inflammatory and immunomodulatory effects, which establish it
as a potential therapeutic option in the management of COVID-19
(57,58). Indeed, previous studies have shown
promising results when intravenously applying high doses of vitamin
C for the management of COVID-19 (59,60).
In accordance with these studies, Feyaerts and Luyten (61) proposed that using a relatively low
dose of vitamin C as a prophylaxis, or an intravenous-based
high-dose regimen in cases of severe COVID-19, may be beneficial.
Similarly, Carr and Rowe (62)
asserted that, due to its excellent safety profile, low cost and
potential for rapid upscaling of production, the administration of
vitamin C to patients with hypovitaminosis C and severe respiratory
infections, such as COVID-19, appears warranted.
Regarding the downregulation of IL-18 and
TNF-α induced by Vita Deyun (as presented in Fig. 3), it has been recently reported that
use of curcumin and silymarin alone or in combination for treatment
of gamma ray-induced nephrotoxicity in rats resulted in a marked
decline in the serum levels of the two cytokines (63).
Currently, there are various drugs and vaccines
being evaluated for their safety, efficacy and effect against
COVID-19, although a significant period of time will be required
for their validation. Therefore, it is also important to consider
the use of natural components that have no side effects in humans
as alternative tools to combat COVID-19. Numerous nutraceuticals
have a proven ability to boost the immune system in addition to
having antiviral, antioxidant and anti-inflammatory effects
(64). However, ongoing clinical
trials are expected to provide more definitive evidence.
Vita Deyun solution is a combination of various
phytonutrients in the form of a food supplement that, based on the
current in vitro results and the abovementioned evidence,
may help to boost the immune system and prevent virus spread,
thereby potentially reducing the symptoms of COVID-19. However, it
is clear that clinical trials are required to fully determine its
efficacy. Currently, clinical protocols that have already been
authorized by ethics committees are being conducted in various
states of Mexico to determine the efficacy of Vita Deyun against
the diverse symptoms associated with COVID-19.
Acknowledgements
The authors are grateful for the support received
from our technicians in the laboratory: Ms Sarah Naomi
Guevara-Hernández, Ms Holanda Vanesa Ramos-Sánchez and Mr Carlos
Eliu Moreno-Rios (Division of Immunology, Western Biomedical
Research Center, Mexican Social Security Institute).
Funding
Envasadora D'Aqua, S.A. de C.V. and Summa Company, S.A. de C.V.
financially supported the present study and covered the publication
costs (IMSS agreement reference 09-B5-61-2800/2020/00).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
AAL and LFJS designed the study, conducted the
transient and stable transfections, performed the RT-qPCR assays
and wrote the manuscript. LFJS analyzed the genomic data from the
published databases. ALU and JSC were involved in the analysis and
interpretation of the results. AAL and LFJS confirmed the
authenticity of all the raw data. All authors read and approved the
final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the National
Scientific Research Committee of the Mexican Social Security
Institute (registration no. R-2020-785-155).
Patient consent for publication
Not applicable.
Competing interests
Note that Envasadora D´Aqua, S.A de C.V. and Summa
Company, S.A de C.V., who produce and distribute the supplement
Vita Deyun and also funded the present study, had no role in the
design, execution, interpretation or writing of the manuscript.
References
|
1
|
Acter T, Uddin N, Das J, Akhter A,
Choudhury TR and Kim S: Evolution of severe acute respiratory
syndrome coronavirus 2 (SARS-CoV-2) as coronavirus disease 2019
(COVID-19) pandemic: A global health emergency. Sci Total Environ.
730(138996)2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Wang C, Horby PW, Hayden FG and Gao GF: A
novel coronavirus outbreak of global health concern. Lancet.
395:470–473. 2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Yang J, Zheng Y, Gou X, Pu K, Chen Z, Guo
Q, Ji R, Wang H, Wang Y and Zhou Y: Prevalence of comorbidities and
its effects in patients infected with SARS-CoV-2: A systematic
review and meta-analysis. Int J Infect Dis. 94:91–95.
2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Burki T: COVID-19 in Latin America. Lancet
Infect Dis. 20:547–548. 2020.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Shereen MA, Khan S, Kazmi A, Bashir N and
Siddique R: COVID-19 infection: Origin, transmission, and
characteristics of human coronaviruses. J Adv Res. 24:91–98.
2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zhang X, Chen X, Chen L, Deng C, Zou X,
Liu W, Yu H, Chen B and Sun X: The evidence of SARS-CoV-2 infection
on ocular surface. Ocul Surf. 18:360–362. 2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Guan WJ, Ni ZY, Hu Y, Liang WH, Ou CQ, He
JX, Liu L, Shan H, Lei CL, Hui DS, et al: Clinical characteristics
of coronavirus disease 2019 in China. N Engl J Med. 382:1708–1720.
2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Letko M, Marzi A and Munster V: Functional
assessment of cell entry and receptor usage for SARS-CoV-2 and
other lineage B betacoronaviruses. Nat Microbiol. 5:562–569.
2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Song W, Gui M, Wang X and Xiang Y: Cryo-EM
structure of the SARS coronavirus spike glycoprotein in complex
with its host cell receptor ACE2. PLoS Pathog.
14(e1007236)2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Li F, Li W, Farzan M and Harrison SC:
Structure of SARS coronavirus spike receptor-binding domain
complexed with receptor. Science. 309:1864–1868. 2005.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Anguiano L, Riera M, Pascual J and Soler
MJ: Circulating ACE2 in cardiovascular and kidney diseases. Curr
Med Chem. 24:3231–3241. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Pacheco JM, Henry TM, O'Donnell VK,
Gregory JB and Mason PW: Role of nonstructural proteins 3A and 3B
in host range and pathogenicity of foot-and-mouth disease virus. J
Virol. 77:13017–13027. 2003.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Lu W, Zheng BJ, Xu K, Schwarz W, Du L,
Wong CK, Chen J, Duan S, Deubel V and Sun B: Severe acute
respiratory syndrome-associated coronavirus 3a protein forms an ion
channel and modulates virus release. Proc Natl Acad Sci USA.
103:12540–12545. 2006.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Chan CM, Tsoi H, Chan WM, Zhai S, Wong CO,
Yao X, Chan WY, Tsui SK and Chan HY: The ion channel activity of
the SARS-coronavirus 3a protein is linked to its pro-apoptotic
function. Int J Biochem Cell Biol. 41:2232–2239. 2009.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Wang X, Zhang Z and Wu SC: Health benefits
of Silybum marianum: Phytochemistry, pharmacology, and
applications. J Agric Food Chem. 68:11644–11664. 2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Saller R, Meier R and Brignoli R: The use
of silymarin in the treatment of liver diseases. Drugs.
61:2035–2063. 2001.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Bosisio E, Benelli C and Pirola O: Effect
of the flavanolignans of Silybum marianum L. on lipid peroxidation
in rat liver microsomes and freshly isolated hepatocytes. Pharmacol
Res. 25:147–154. 1992.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Carini R, Comoglio A, Albano E and Poli G:
Lipid peroxidation and irreversible damage in the rat hepatocyte
model. Protection by the silybin-phospholipid complex IdB 1016.
Biochem Pharmacol. 43:2111–2115. 1992.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Mira L, Silva M and Manso CF: Scavenging
of reactive oxygen species by silibinin dihemisuccinate. Biochem
Pharmacol. 48:753–759. 1994.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Lani R, Hassandarvish P, Chiam CW,
Moghaddam E, Chu JJ, Rausalu K, Merits A, Higgs S, Vanlandingham D,
Abu Bakar S and Zandi K: Antiviral activity of silymarin against
chikungunya virus. Sci Rep. 5(11421)2015.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zhao F, Shi D, Li T, Li L and Zhao M:
Silymarin attenuates paraquat-induced lung injury via Nrf2-mediated
pathway in vivo and in vitro. Clin Exp Pharmacol Physiol.
42:988–998. 2015.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Anderson ME and Meister A: Dynamic state
of glutathione in blood plasma. J Biol Chem. 255:9530–9533.
1980.PubMed/NCBI
|
|
23
|
Sies H and Graf P: Hepatic thiol and
glutathione efflux under the influence of vasopressin,
phenylephrine and adrenaline. Biochem J. 226:545–549.
1985.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Kozak EM and Tate SS:
Glutathione-degrading enzymes of microvillus membranes. J Biol
Chem. 257:6322–6327. 1982.PubMed/NCBI
|
|
25
|
Kloek J, Van Ark I, De Clerck F, Bloksma
N, Nijkamp FP and Folkerts G: Modulation of airway
hyperresponsiveness by thiols in a murine in vivo model of allergic
asthma. Inflamm Res. 52:126–131. 2003.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Lee YC, Lee KS, Park SJ, Park HS, Lim JS,
Park KH, Im MJ, Choi IW, Lee HK and Kim UH: Blockade of airway
hyperresponsiveness and inflammation in a murine model of asthma by
a prodrug of cysteine, L-2-oxothiazolidine-4-carboxylic acid. FASEB
J. 18:1917–1919. 2004.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Hodge S, Matthews G, Mukaro V, Ahern J,
Shivam A, Hodge G, Holmes M, Jersmann H and Reynolds PN: Cigarette
smoke-induced changes to alveolar macrophage phenotype and function
are improved by treatment with procysteine. Am J Respir Cell Mol
Biol. 44:673–681. 2011.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Sido B, Braunstein J, Breitkreutz R,
Herfarth C and Meuer SC: Thiol-mediated redox regulation of
intestinal lamina propria T lymphocytes. J Exp Med. 192:907–912.
2000.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Hadzic T, Li L, Cheng N, Walsh SA, Spitz
DR and Knudson CM: The role of low molecular weight thiols in T
lymphocyte proliferation and IL-2 secretion. J Immunol.
175:7965–7972. 2005.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Oliver JM, Albertini DF and Berlin RD:
Effects of glutathione-oxidizing agents on microtubule assembly and
microtubule-dependent surface properties of human neutrophils. J
Cell Biol. 71:921–932. 1976.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Kuppner MC, Scharner A, Milani V, Von
Hesler C, Tschop KE, Heinz O and Issels RD: Ifosfamide impairs the
allostimulatory capacity of human dendritic cells by intracellular
glutathione depletion. Blood. 102:3668–3674. 2003.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Peterson JD, Herzenberg LA, Vasquez K and
Waltenbaugh C: Glutathione levels in antigen-presenting cells
modulate Th1 versus Th2 response patterns. Proc Natl Acad Sci USA.
95:3071–3076. 1998.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Short S, Merkel BJ, Caffrey R and McCoy
KL: Defective antigen processing correlates with a low level of
intracellular glutathione. Eur J Immunol. 26:3015–3020.
1996.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Murata Y, Ohteki T, Koyasu S and Hamuro J:
IFN-gamma and pro-inflammatory cytokine production by
antigen-presenting cells is dictated by intracellular thiol redox
status regulated by oxygen tension. Eur J Immunol. 32:2866–2873.
2002.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Furuya A, Uozaki M, Yamasaki H, Arakawa T,
Arita M and Koyama AH: Antiviral effects of ascorbic and
dehydroascorbic acids in vitro. Int J Mol Med. 22:541–545.
2008.PubMed/NCBI
|
|
36
|
White LA, Freeman CY, Forrester BD and
Chappell WA: In vitro effect of ascorbic acid on infectivity of
herpesviruses and paramyxoviruses. J Clin Microbiol. 24:527–531.
1986.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Alkhatib A: Antiviral functional foods and
exercise lifestyle prevention of coronavirus. Nutrients.
12(2633)2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Zhang J, Saad R, Taylor EW and Rayman MP:
Selenium and selenoproteins in viral infection with potential
relevance to COVID-19. Redox Biol. 37(101715)2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Hiffler L and Rakotoambinina B: Selenium
and RNA virus interactions: Potential implications for SARS-CoV-2
infection (COVID-19). Front Nutr. 7(164)2020.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Boukamp P, Petrussevska RT, Breitkreutz D,
Hornung J, Markham A and Fusenig NE: Normal keratinization in a
spontaneously immortalized aneuploid human keratinocyte cell line.
J Cell Biol. 106:761–771. 1988.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Chang SE, Foster S, Betts D and Marnock
WE: DOK, a cell line established from human dysplastic oral mucosa,
shows a partially transformed non-malignant phenotype. Int J
Cancer. 52:896–902. 1992.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Pfaffl MW: A new mathematical model for
relative quantification in real-time RT-PCR. Nucleic Acids Res.
29(e45)2001.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Gordon DE, Jang GM, Bouhaddou M, Xu J,
Obernier K, White KM, O'Meara MJ, Rezelj VV, Guo JZ, Swaney DL, et
al: A SARS-CoV-2 protein interaction map reveals targets for drug
repurposing. Nature. 583:459–468. 2020.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Katsura H, Sontake V, Tata A, Kobayashi Y,
Edwards CE, Heaton BE, Konkimalla A, Asakura T, Mikami Y, Fritch
EJ, et al: Human lung stem cell-based alveolospheres provide
insights into SARS-CoV-2-mediated interferon responses and
pneumocyte dysfunction. Cell Stem Cell. 27:890–904.e8.
2020.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Xu H, Zhong L, Deng J, Peng J, Dan H, Zeng
X, Li T and Chen Q: High expression of ACE2 receptor of 2019-nCoV
on the epithelial cells of oral mucosa. Int J Oral Sci.
12(8)2020.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Ren Y, Shu T, Wu D, Mu J, Wang C, Huang M,
Han Y, Zhang XY, Zhou W, Qiu Y and Zhou X: The ORF3a protein of
SARS-CoV-2 induces apoptosis in cells. Cell Mol Immunol.
17:881–883. 2020.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Wan Y, Shang J, Graham R, Baric RS and Li
F: Receptor recognition by the novel coronavirus from Wuhan: An
analysis based on decade-long structural studies of SARS
Coronavirus. J Virol. 94:e00127–20. 2020.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Wagoner J, Negash A, Kane OJ, Martinez LE,
Nahmias Y, Bourne N, Owen DM, Grove J, Brimacombe C, McKeating JA,
et al: Multiple effects of silymarin on the hepatitis C virus
lifecycle. Hepatology. 51:1912–1921. 2010.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Patel CN, Kumar SP, Pandya HA and Rawal
RM: Identification of potential inhibitors of coronavirus
hemagglutinin-esterase using molecular docking, molecular dynamics
simulation and binding free energy calculation. Mol Divers.
25:421–433. 2021.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Kumar S, Kashyap P, Chowdhury S, Kumar S,
Panwar A and Kumar A: Identification of phytochemicals as potential
therapeutic agents that binds to Nsp15 protein target of
coronavirus (SARS-CoV-2) that are capable of inhibiting virus
replication. Phytomedicine. (153317)2020.PubMed/NCBI View Article : Google Scholar : (Epub ahead of
print).
|
|
51
|
Gorla US, Rao GK, Kulandaivelu US, Alavala
RR and Panda SP: Lead finding from selected flavonoids with
antiviral (SARS-CoV-2) potentials against COVID-19: An in-silico
evaluation. Comb Chem High Throughput Screen: Aug 18, 2020 (Epub
ahead of print).
|
|
52
|
Kim J, Zhang J, Cha Y, Kolitz S, Funt J,
Escalante Chong R, Barrett S, Kusko R, Zeskind B and Kaufman H:
Advanced bioinformatics rapidly identifies existing therapeutics
for patients with coronavirus disease-2019 (COVID-19). J Transl
Med. 18(257)2020.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Silvagno F, Vernone A and Pescarmona GP:
The role of glutathione in protecting against the severe
inflammatory response triggered by COVID-19. Antioxidants (Basel).
9(624)2020.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Guloyan V, Oganesian B, Baghdasaryan N,
Yeh C, Singh M, Guilford F, Ting YS and Venketaraman V: Glutathione
supplementation as an adjunctive therapy in COVID-19. Antioxidants
(Basel). 9(914)2020.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Taylor EW and Radding W: Understanding
selenium and glutathione as antiviral factors in COVID-19: Does the
Viral Mpro Protease target host selenoproteins and
glutathione synthesis? Front Nutr. 7(143)2020.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Moghaddam A, Heller RA, Sun Q, Seelig J,
Cherkezov A, Seibert L, Hackler J, Seemann P, Diegmann J, Pilz M,
et al: Selenium deficiency is associated with mortality risk from
COVID-19. Nutrients. 12(2098)2020.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Abobaker A, Alzwi A and Alraied AHA:
Overview of the possible role of vitamin C in management of
COVID-19. Pharmacol Rep. 72:1517–1528. 2020.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Mousavi S, Bereswill S and Heimesaat MM:
Immunomodulatory and antimicrobial effects of vitamin C. Eur J
Microbiol Immunol (Bp). 9:73–79. 2019.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Waqas Khan HM, Parikh N, Megala SM and
Predeteanu GS: Unusual early recovery of a critical COVID-19
patient after administration of intravenous vitamin C. Am J Case
Rep. 21(e925521)2020.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Liu F, Zhu Y, Zhang J, Li Y and Peng Z:
Intravenous high-dose vitamin C for the treatment of severe
COVID-19: Study protocol for a multicentre randomised controlled
trial. BMJ Open. 10(e039519)2020.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Feyaerts AF and Luyten W: Vitamin C as
prophylaxis and adjunctive medical treatment for COVID-19?
Nutrition. 79-80(110948)2020.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Carr AC and Rowe S: The emerging role of
vitamin C in the prevention and treatment of COVID-19. Nutrients.
12(3286)2020.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Abdel-Magied N and Elkady AA: Possible
curative role of curcumin and silymarin against nephrotoxicity
induced by gamma-rays in rats. Exp Mol Pathol.
111(104299)2019.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Mrityunjaya M, Pavithra V, Neelam R,
Janhavi P, Halami PM and Ravindra PV: Immune-boosting, antioxidant
and anti-inflammatory food supplements targeting pathogenesis of
COVID-19. Front Immunol. 11(570122)2020.PubMed/NCBI View Article : Google Scholar
|