Introduction
Coronary heart disease (CHD) is a significant
disease that threatens human health worldwide, causing ~7.4 million
deaths annually (1). The mortality
rate of CHD has declined over time due to advances in prevention,
diagnosis and treatment of this disease (2); nevertheless, it varies from country
to country (2). A total of ~75% of
the U.S. counties have achieved a reduction in CHD mortality by 20%
in 10 years (103.4/100,000 individuals). However, the regional
differences in the magnitude of the reduction in CHD mortality is
noteworthy (3). The narrow focus
on the estimation of the decreased mortality may result in missed
opportunities to prevent and treat cardiovascular diseases and to
explore additional mechanisms regarding the factors that lead to
successful mortality reductions. It is interesting to note that the
death caused by ischemic cardiovascular disease accounts for 93.17%
of the total deaths from cardiovascular disease. Advanced age,
smoking, diabetes (4) and
psychosocial stress (5) are all
risk factors of cardiovascular disease. To date, the most effective
modalities for vascular reperfusion in ischemic cardiovascular
disease include thrombolysis, percutaneous coronary intervention
(6), and coronary artery bypass
grafting (7). Nurse-led
patient-centered care can significantly improve smoking habits,
alcohol consumption, adherence to physical activity and total
cholesterol levels (8). However,
despite the beneficial use of reperfusion of the ischemic
myocardium, the subsequent myocardial ischemia-reperfusion injury
(MIRI) can cause further damage to the patient, resulting in a poor
prognosis (9). Therefore, there is
an urgent requirement to explore novel strategies to alter the
status of prevention and treatment of MIRI.
Inflammation is an important part of MIRI.
Ischemia-reperfusion injury activates the innate immune response,
resulting in an excessive inflammatory response (10). Excessive inflammatory responses
lead to the secretion of reactive oxygen species (ROS), the
infiltration of immune cells and the damage of the vascular
endothelium, which in turn damages the myocardial tissue (11). Interleukin (IL)-32 is a relatively
newly discovered pro-inflammatory cytokine present in natural
killer cells (NK) (12). A recent
study showed that IL-32 can promote cell differentiation,
participate in apoptosis, induce the production of other
pro-inflammatory cytokines (13)
and chemokines through multiple signaling transduction pathways,
and play an important role in inflammatory responses and autoimmune
diseases (14,15). For example, IL-32 can directly
affect specific immunity by inducing monocytes to differentiate
into macrophages (16). IL-32
induces the expression of TNF-α, IL-1β and macrophage inflammatory
protein-2 in mouse macrophages and stimulates human mononuclear
cells to secrete prostaglandin E2, which is an important
inflammatory mediator of cartilage and bone destruction in
rheumatoid arthritis (17). The
aforementioned studies indicated that IL-32 is an important
pro-inflammatory cytokine, and an important chemokine and
pro-inflammatory cytokine inducer. Given that IL-32 has been
implicated in Crohn's disease (18) and rheumatoid arthritis, both of
which are associated with inflammation, the present study
hypothesized that this cytokine may play a primary role in the
development of MIRI.
Human cardiomyocytes were used to establish an in
vitro model to mimic MIRI by hypoxia and reoxygenation (H/R)
(19). The present study
investigated the effects of IL-32 on inflammation, oxidative stress
and apoptosis in H/R-induced cells. The mechanism of action of
IL-32 was also explored. By investigating the potential role of
IL-32 in MIRI, the present study aimed to explore the development
of preventive or therapeutic approaches for treating CHD.
Materials and methods
Cell culture and treatment
Human cardiomyoblasts (Immortalized; BFN60808678,
BLUEFBIO, www.bluefcell.com) were cultured in
DMEM (Gibco; Thermo Fisher Scientific, Inc.) containing 10% FBS
(Beijing Solarbio Science & Technology Co., Ltd.) and 1%
penicillin-streptomycin (Beijing Solarbio Science & Technology
Co., Ltd.). The cells in the control group were maintained in an
incubator with 5% CO2 at 37˚C for 48 h. The cells
(1.5x105 cells/well) in the H/R treatment group were
placed in an incubator containing 94% N2, 5%
CO2, and 1% O2 at 37˚C to induce hypoxia for
3 h; subsequently, they were placed in an incubator with 5%
CO2 at 37˚C for an additional 4, 8, and 16 h for
reoxygenation.
Cell transfection
The cells (5x105 cells/well) in the
six-well plate were transfected with small interfering RNA (siRNA)
to knockdown IL-32 expression and the non-targeted siRNA was used
as the negative control (NC). The cells were transfected with
pcDNA3.1 vector to overexpress nucleotide-binding oligomerization
domain 2 (NOD2) for 48 h at 37˚C and the empty vector (GenePharm,
Inc.) was used as the NC. The method was performed according to the
instructions provided by the manufacturer (FuGENE HD transfection
reagent; Roche Diagnostics). The cells were collected for
subsequent experiments 48 h after transfection. The expression
levels of IL-32 or NOD2 were assessed following 36 h of cell
culture. IL-32 siRNA-1 target (5'-3'): GAGCTGGAGGACGACTTCAAA; IL-32
siRNA-2 target: GAAGGTCCTCTCTGATGACAT; siRNA-NC:
GGCGTGCAGCAGGAAATACTA.
Cell counting kit (CCK)-8 assay
The cells (5x103/well) in the 96-well
plates were treated as aforementioned. The cell viability was
evaluated following 2 h of culture with the addition of the CCK-8
solution (10 µl; Beyotime Institute of Biotechnology) at 37˚C. The
optical density (OD) was measured using a microplate reader (450
nm; Hiwell-Diatek).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from cells using
TRIzol® reagent (Thermo Fisher Scientific, Inc.) and
cDNA was reverse transcribed using a Reverse Transcriptase kit
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. The reverse transcription product was diluted and RT-qPCR
was performed using a QuantiTect® SYBR-Green PCR kit
(Qiagen, Inc.). The thermocycling conditions were as follows: 95˚C
for 30 sec, 40 cycles of 95˚C for 30 sec, 60˚C for 30 sec, and 72˚C
for 30 sec. The 2-ΔΔCq method (20) was used to analyze the data. β-actin
was used for normalization. The primer sequences are shown in
Table I.
 | Table IPrimer sequences used for reverse
transcription-quantitative PCR. |
Table I
Primer sequences used for reverse
transcription-quantitative PCR.
| Primer name | Primer sequence
(5'-3') |
|---|
| IL-32 | F:
CTCTCTCGGCTGAGTATTTGTG |
| | R:
GCTCGACATCACCTGTCCAC |
| TNF-α | F:
TGGGATCATTGCCCTGTGAG |
| | R:
GGTGTCTGAAGGAGGGGGTA |
| IL-6 | F:
GTCCAGTTGCCTTCTCCCTGG |
| | R:
CCCATGCTACATTTGCCGAAG |
| IL-1β | F:
TGAGCTCGCCAGTGAAATGAT |
| | R:
TCCATGGCCACAACAACTGA |
| NOD2 | F:
CTTCTGGAGAAGTCCCGCAC |
| | R:
TCTGTGCCTGAAAAGCCTCC |
| β-actin | F:
CTTCGCGGGCGACGAT |
| | R:
CCACATAGGAATCCTTCTGACC |
Western blotting
Protein extraction was performed and a Nano 300
spectrophotometer (YPH-Bio) was used for protein quantification.
SDS-PAGE using 10% gels was performed to separate the protein
samples (25 µg/lane) and the separated proteins were subsequently
transferred to a polyvinylidene membrane, followed by blocking in
non-fat milk for 1 h at room temperature. Following incubation of
the blots with primary antibodies [IL-32 (cat. no. 11079-1-AP;
1:1,000; ProteinTech Group, Inc.), phosphorylated (p-)p65 (cat. no.
GTX133899; 1:1,000; GeneTex, Inc.), cyclooxygenase-2 (COX-2; cat.
no. 12375-1-AP; 1:2,000; ProteinTech Group, Inc.), p65 (cat. no.
GTX102090; 1:2,000; GeneTex, Inc.), Bax (cat. no. 50599-2-lg;
1:5,000; ProteinTech Group, Inc.), cleaved caspase 3 (cat. no.
GTX03281; 1:1,000; GeneTex, Inc.), caspase 3 (cat. no. GTX110543;
1:1,000; GeneTex, Inc.), Bcl2 (cat. no. 26593-1-AP; 1:2,000;
ProteinTech Group, Inc.), NOD2 (cat. no. GTX30694; 1:1,000;
GeneTex, Inc.), NADPH oxidase 2 (NOX2; cat. no. 19013-1-AP;
1:1,000; ProTeintech Group, Inc.), p-ERK (cat. no. 28733-1-AP;
1:5,000; ProteinTech Group, Inc.), ERK (cat. no. 11257-1-AP;
1:2,000; ProteinTech Group, Inc.), β-actin (cat. no. 20536-1-AP;
1:5,000; ProteinTech Group, Inc.)] at 4˚C overnight, the strips
were incubated at room temperature with an HRP-conjugated secondary
antibody (cat. no. SA00001-2; 1:5,000; ProteinTech Group, Inc.) for
2 h. An ECL kit (GK10008; GlpBio) was used for visualization.
β-actin was used for normalization and ImageJ software (v1.8.0;
National Institutes of Health) was used for densitometry.
Determination of lactate dehydrogenase
(LDH) activity and oxidative stress indices
LDH (ab102526; Abcam), malondialdehyde (MDA; cat.
no. A003-4-1; Nanjing Jiancheng Bioengineering Institute), and
superoxide dismutase (SOD; cat. no. A001-1; Nanjing Jiancheng
Bioengineering Institute) assay kits were used to assess the
activity of the corresponding biochemical function indices.
Briefly, the cells were collected and lysed and the samples were
obtained according to the different requirements of the kits. The
OD values were measured using a microplate reader. The
determination of ROS was performed using a 2'-7'dichlorofluorescin
diacetate (DCFH-DA) assay kit (cat. no. E004-1-1; Nanjing Jiancheng
Bioengineering Institute). Diluted DCFH-DA was added to the wells
and the cells were incubated for 30 min at 37˚C. Subsequently, the
cells were collected and washed with PBS twice. Following
centrifugation at 300 x g for 5 min at 4˚C, the supernatant was
removed, and the cells were suspended in PBS for detection. Images
of the results were captured using a fluorescence microscope
(magnification, x100; Olympus Corporation) and quantified with
ImageJ software.
TUNEL assay
The cells (5x105/well) were seeded in a
24-well plate and cultured until they reached 80% confluence.
Following H/R treatment, the cell smears were immersed in 4%
paraformaldehyde and fixed for 30 min at room temperature, followed
by washing with PBS. The working solution in the TUNEL assay kit
(E-CK-A334; Elabscience Biotechnology, Inc.) was added for 1 h at
37˚C in the dark according to the manufacturer's protocols. The
nuclei were counterstained with 1 mg/ml DAPI for 5 min at room
temperature in the dark and the slides were then mounted with
anti-fade mounting medium. The coverslip was removed, sealed and
the apoptotic cells in six randomly selected fields were observed
under a fluorescence microscope (magnification, x200).
Statistical analysis
GraphPad Prism 8.0 (GraphPad Software, Inc.) was
used for statistical analysis. All experiments were performed 3
times and the data are presented as the mean ± SD. Difference
between multiple groups were compared using a one-way ANOVA with
Tukey's post-hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Downregulation of IL-32 expression
attenuates H/R-induced viability reduction, LDH release, and
induction of oxidative stress
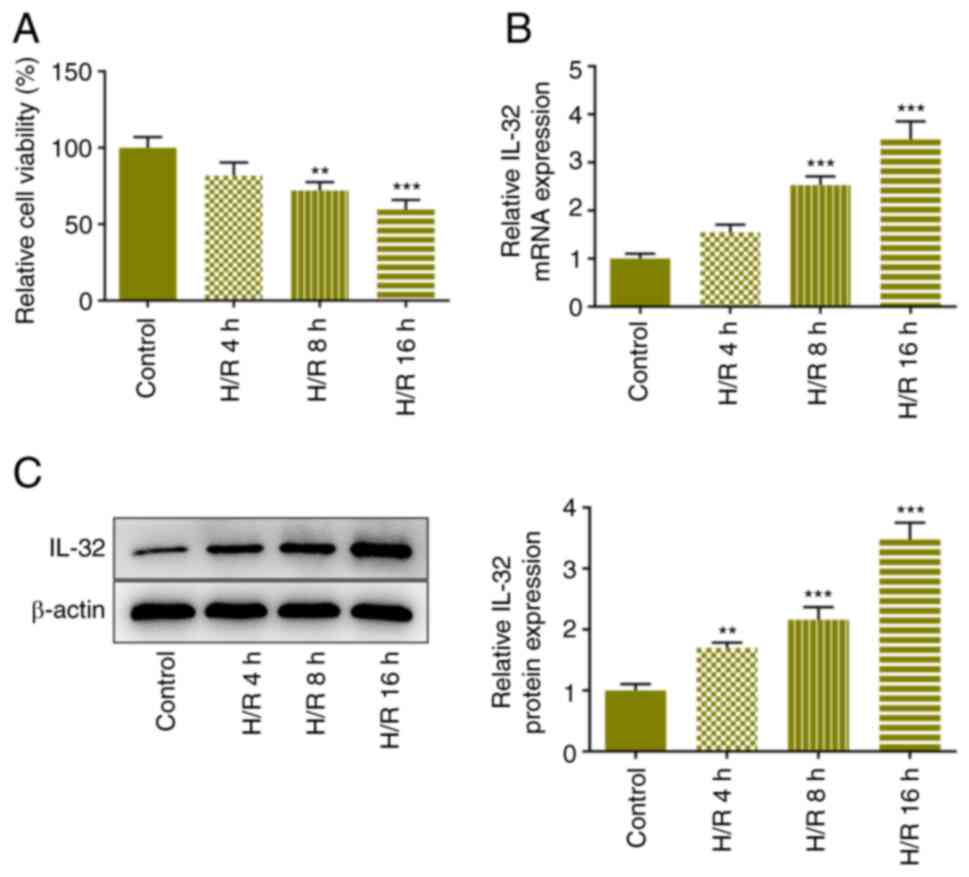
Following induction of reoxygenation in the cells
for 4, 8, and 16 h, the viability in each group was determined
using the CCK-8 assay. The viability in the treated cells was
significantly reduced compared with that of the control group,
notably in the H/R 16 h group (Fig.
1A). IL-32 expression levels in these groups of cells were
assessed using RT-qPCR and western blot analyses. The expression
levels of IL-32 were increased in a dose-dependent manner following
the increase in the reoxygenation time (Fig. 1B and C). To highlight the role of IL-32, the
cells were cultured for 16 h under reoxygenation conditions in the
subsequent experiments. IL-32 siRNA was constructed and the cells
were transfected with siRNA to downregulate IL-32 expression
levels. The efficacy was verified using RT-qPCR and western
blotting assays in the normal (Fig.
2A and B) and H/R-treated
transfected cells (Fig. 2C and
D) and the siRNA-IL-32-1 group was
selected for the subsequent assays. The effects of the
downregulation of IL-32 expression on cell viability and LDH levels
were evaluated using the CCK-8 and LDH assay kits, respectively.
Downregulation of IL-32 expression significantly elevated cell
viability (Fig. 2E) and reduced
the LDH release (Fig. 2F), partly
reversing the impact of H/R treatment. The levels of MDA (Fig. 2G), SOD (Fig. 2H), and ROS (Fig. 2I) in each group were also assessed
using the specific assay kits. H/R treatment significantly elevated
MDA and ROS levels, whereas it decreased the SOD level in the
cells. Moreover, downregulation of IL-32 expression facilitated the
decline in MDA and ROS levels, accompanied by an increase in SOD
levels.
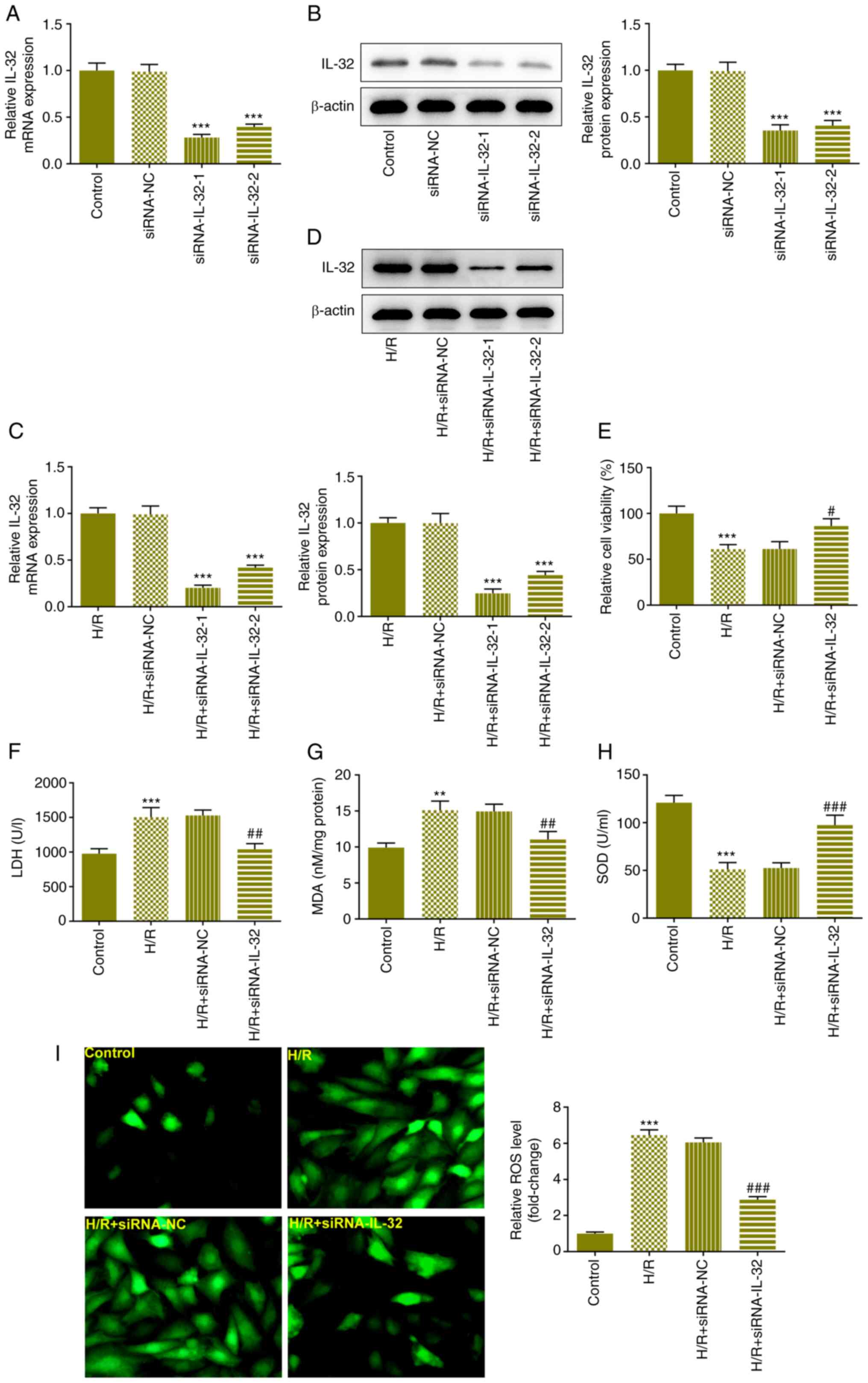 | Figure 2Downregulation of IL-32 expression
attenuates H/R-induced viability reduction, LDH release and
oxidative stress. (A) IL-32 siRNA was constructed and the cells
were transfected with siRNA to downregulate IL-32 levels. The
expression levels of IL-32 in the cells were verified with RT-qPCR
and (B) western blot assays. (C) The expression levels of IL-32 in
the H/R-treated cells were verified with RT-qPCR and (D) western
blot assays. (E) The effects of the downregulation of IL-32
expression on cell viability were evaluated using the Cell Counting
Kit-8 assay. (F) The effects of the downregulation of IL-32
expression on LDH levels were evaluated using the LDH assay kit.
The levels of (G) MDA, (H) SOD and (I) ROS in each group were
assessed using specific assay kits. **P<0.01 and
***P<0.001 vs. control, siRNA-NC or H/R + siRNA-NC;
#P<0.05, ##P<0.01 and
###P<0.001 vs. H/R + siRNA-NC. IL, interleukin; H/R,
hypoxia and reoxygenation; LDH, lactate dehydrogenase; siRNA, small
interfering RNA; RT-qPCR, reverse transcription-quantitative PCR;
MDA, malondialdehyde; SOD, superoxide dismutase; ROS, reactive
oxygen species. |
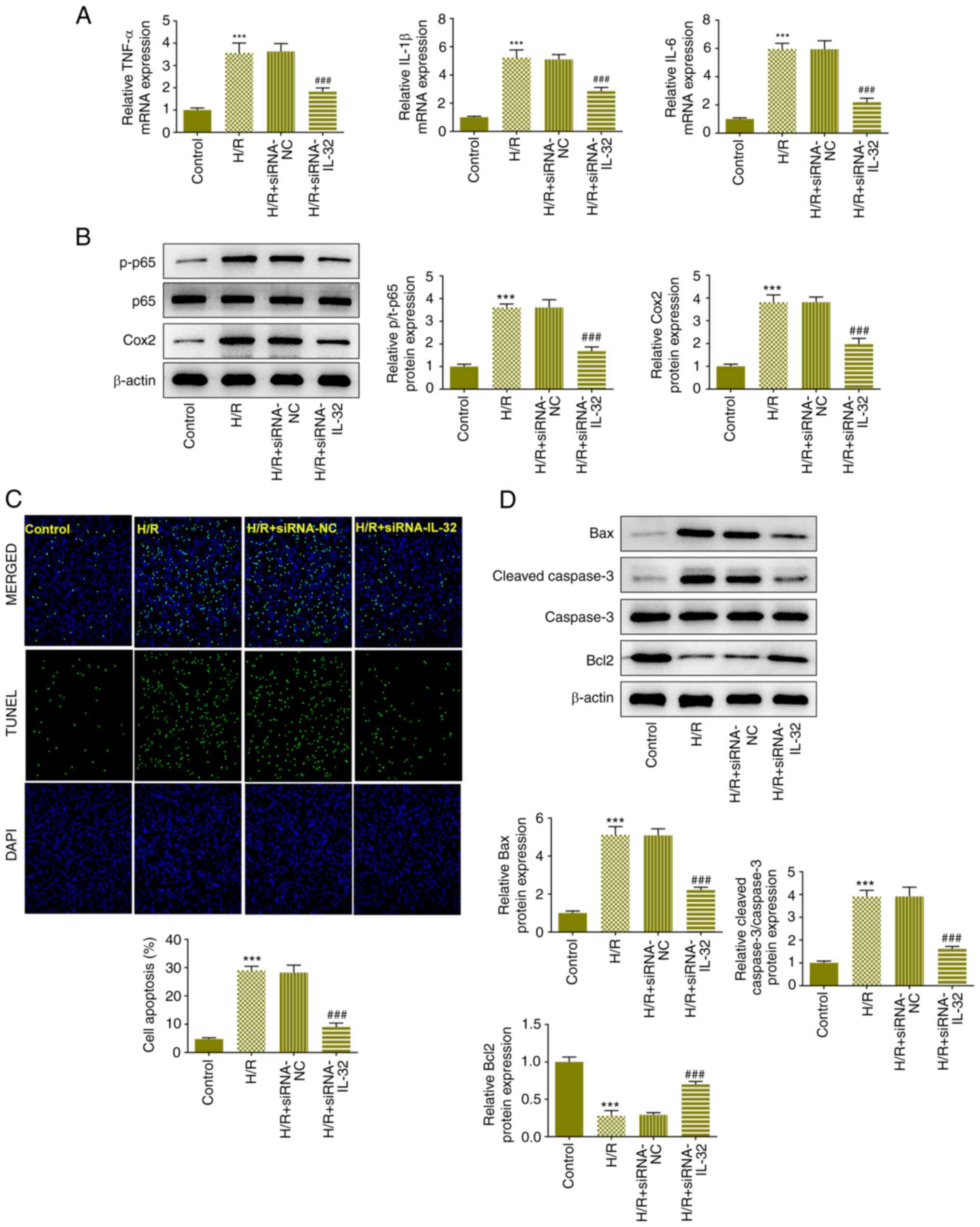
Downregulation of IL-32 expression
attenuates H/R-induced inflammation and apoptosis
The expression levels of TNF-α, IL-1β and IL-6 were
all increased in the H/R group and decreased to a certain degree
due to downregulation of IL-32 expression, according to the results
of RT-qPCR (Fig. 3A). Moreover,
the expression levels of p-p65, p65 and COX-2 were assessed with
western blot analysis. H/R treatment significantly increased the
levels of p-p65 and COX-2, whereas IL-32 reduced their expression
levels (Fig. 3B). In addition, the
induction of apoptosis in each group was determined with TUNEL
(Fig. 3C) and western blot
analyses (Fig. 3D). The
fluorescence activity of the H/R group was significantly increased
compared with that of the control group. IL-32 downregulation
effectively alleviated the enhancement of the fluorescence
activity. H/R treatment increased Bax and cleaved caspase 3 levels,
along with the reduction in Bcl-2 levels, whereas IL-32
downregulation partially reversed these changes.
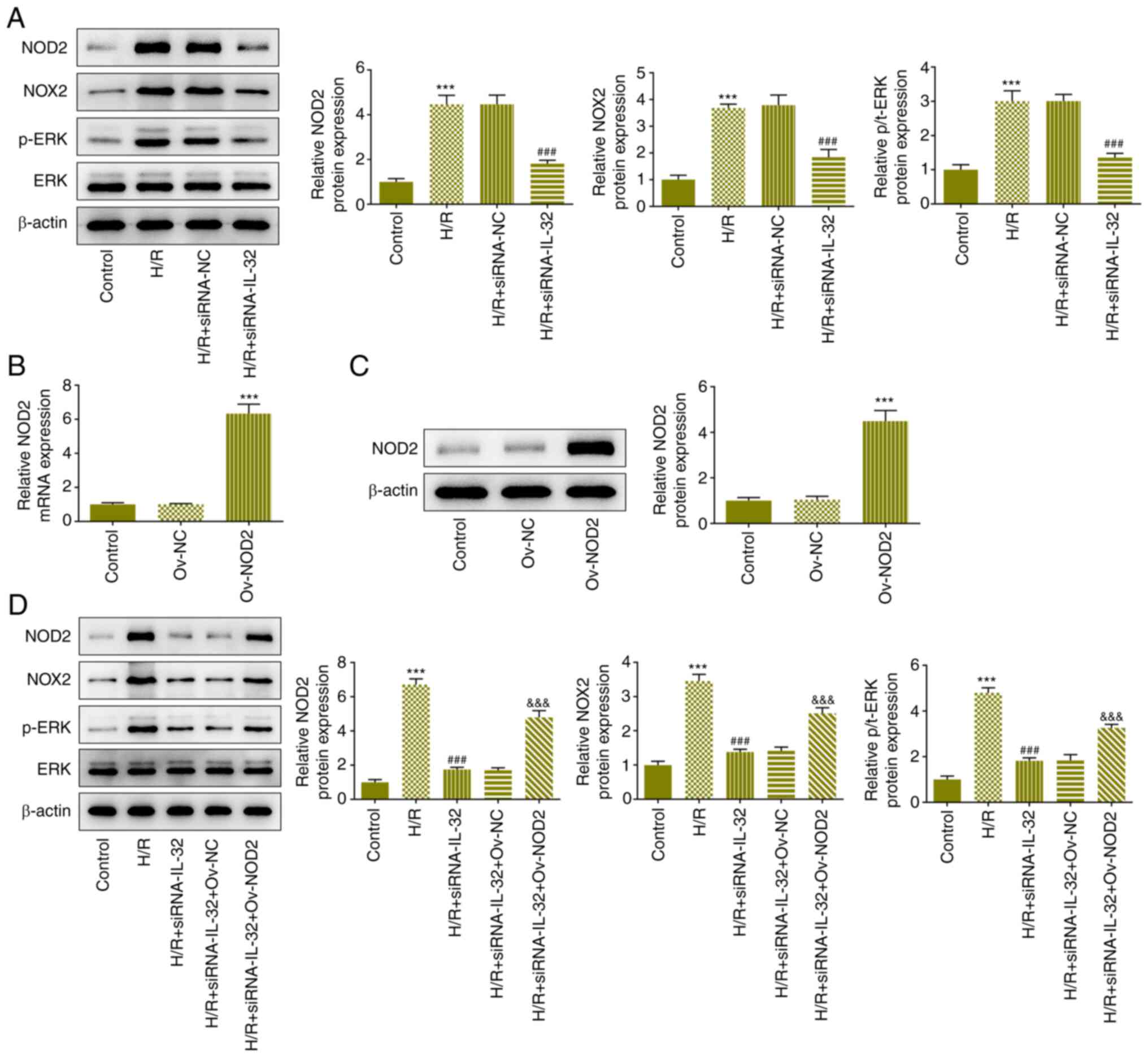
IL-32 regulates the NOD2/NOX2/MAPK
signaling pathway
To determine the regulatory axis of IL-32, the
expression levels of the proteins related to the NOD2/NOX2/MAPK
signaling pathway were determined using western blotting. The
expression levels of NOD2, NOX2 and p-ERK were elevated in the H/R
treatment group, and partly declined following downregulation of
IL-32 expression (Fig. 4A). To
verify the role of NOD2 signaling in the regulatory mechanism of
IL-32, NOD was overexpressed. Following the verification of the
NOD2 overexpressing models (Fig.
4B and C), the cells were
co-transfected with IL-32 siRNA and NOD2 plasmids to explore the
effects of this regulatory axis. NOD2 overexpression activated the
NOD2, NOX2 and p-ERK signaling pathways, reversing the effects of
the downregulation of IL-32 expression on the expression levels of
these proteins (Fig. 4D).
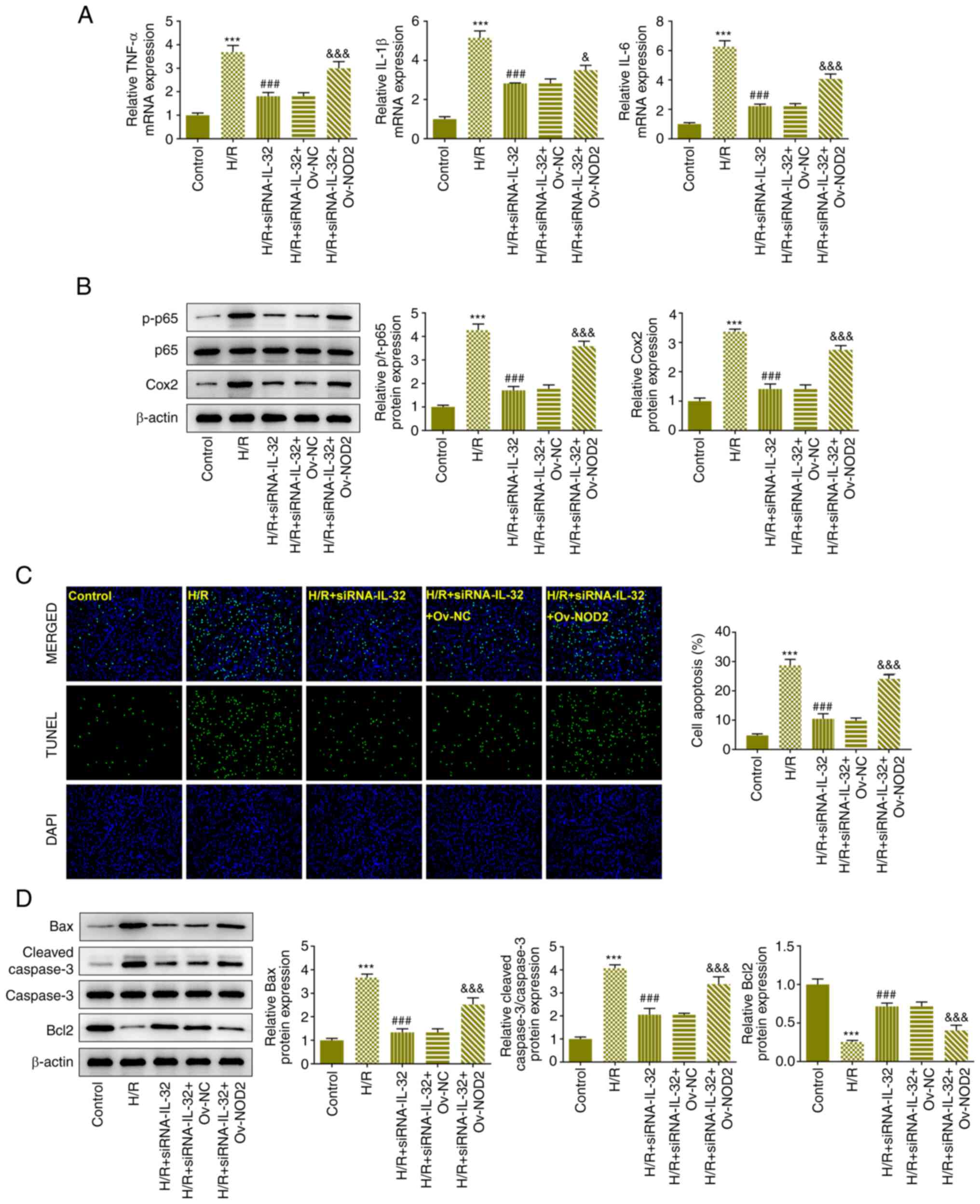
IL-32 functions via the NOD2/NOX2/MAPK
signaling pathway
NOD2 overexpression reduced cell viability (Fig. 5A) and elevated LDH levels (Fig. 5B), indicating that IL-32 regulation
of the viability and LDH release was influenced by NOD2 expression.
Subsequently, the effects of NOD2 overexpression on the induction
of oxidative stress were evaluated. Overexpression of NOD2
increased the levels of MDA and reduced SOD levels (Fig. 5C and D). Overexpression of NOD2 increased the
levels of ROS (Fig. 5E),
indicating that it could disturb the effect of the downregulation
of IL-32 expression on oxidative stress in the cells. Moreover,
NOD2 overexpression significantly increased the expression levels
of TNF-α, IL-1β and IL-6 (Fig.
6A), as well as the expression levels of p-p65 and COX-2
(Fig. 6B). These results suggested
that NOD2 signaling mediated the regulation of IL-32 on cell
inflammation. Furthermore, the effects of the downregulation of
IL-32 expression on the apoptotic rate were also reversed by NOD2
overexpression as demonstrated by increased fluorescence levels
(Fig. 6C), elevated Bax and
cleaved caspase 3, as well as reduced Bcl2 protein expression
levels (Fig. 6D).
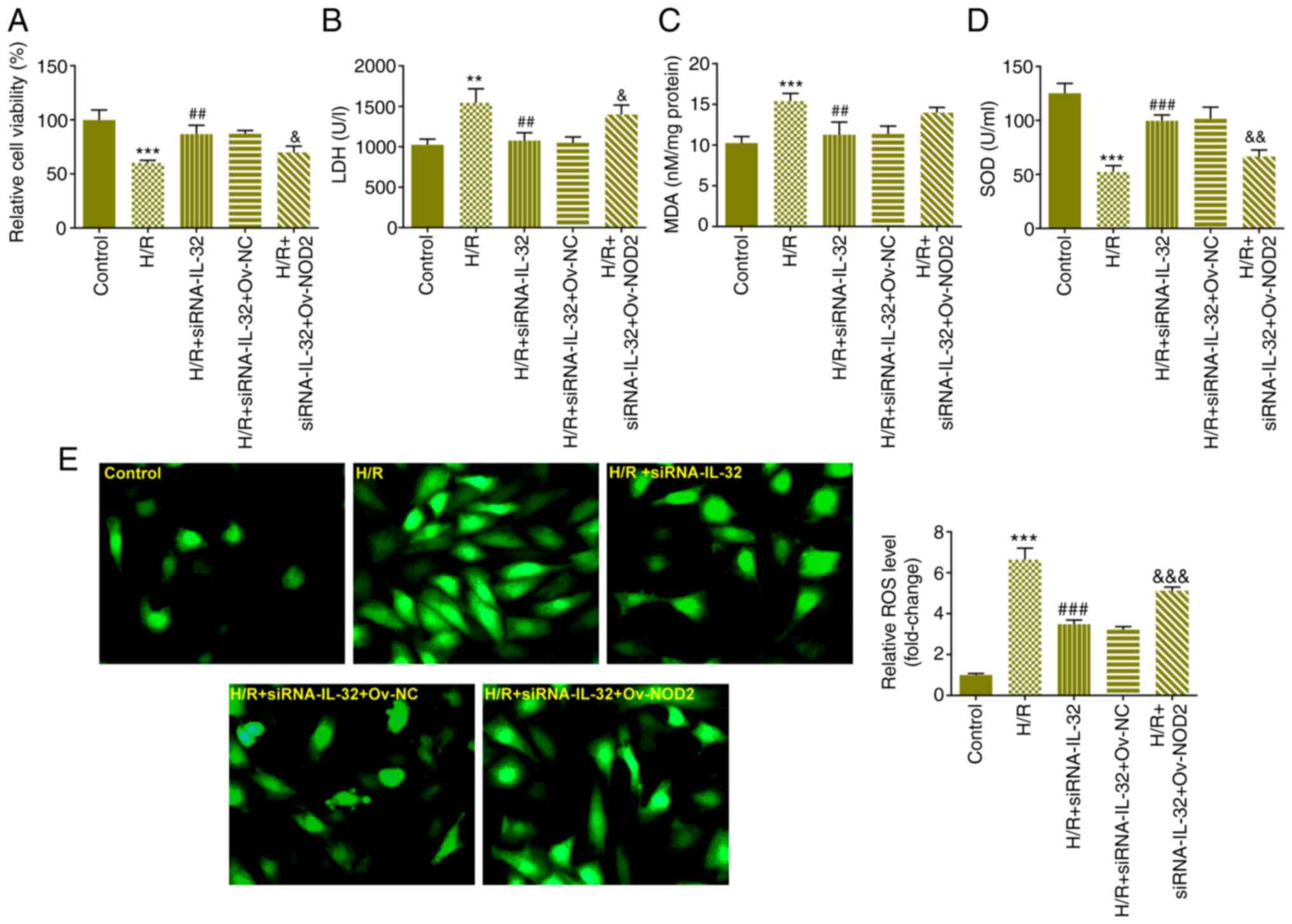 | Figure 5IL-32 induces oxidative stress via
the NOD2/NOX2/MAPK signaling pathway. (A) The effects of NOD2
overexpression on cell viability were evaluated using the Cell
Counting Kit-8 assay. (B) The effects of NOD2 overexpression on LDH
levels were evaluated using the LDH assay kit. The effects of NOD2
overexpression on the levels of (C) MDA, (D) SOD and (E) ROS were
assessed using specific assay kits. **P<0.01 and
***P<0.001 vs. control; ##P<0.01 and
###P<0.001 vs. H/R; &P<0.05,
&&P<0.01 and
&&&P<0.001 vs. H/R + siRNA-IL-32 + Ov-NC.
IL, interleukin; NOD, nucleotide-binding oligomerization domain;
NOX, NADPH oxidase; LDH, lactate dehydrogenase; MDA,
malondialdehyde; SOD, superoxide dismutase; ROS, reactive oxygen
species; H/R, hypoxia and reoxygenation; siRNA, small interfering
RNA; NC, negative control. |
Discussion
Despite the decrease noted globally in the burden of
cardiovascular disease, this condition is rapidly increasing in
developing countries (21).
Reperfusion is a commonly used clinical treatment for
cardiovascular disease (22).
Addressing the reperfusion arrhythmia following the restoration of
blood supply is a promising strategy for treating cardiovascular
disease complications. Although IL-32 is a key cytokine its
function in MIRI has not been previously revealed. It is considered
that the investigation of the disease mechanism of MIRI can provide
a novel concept for the development of therapeutic strategies for
the treatment of this condition. Myocardial membrane permeability
is increased in patients with MIRI, resulting in a rapid rise in
LDH levels in the body (23). The
results of the present study demonstrated that IL-32 levels were
increased following H/R, whereas increased IL-32 levels promoted a
cascade reaction in cardiomyocytes, which was accompanied by
increased LDH release, inflammatory response, oxidative stress and
apoptosis. This suggests that IL-32 plays a primary role in
MIRI-induced myocardial injury. In a previous study, IL-32
expression levels were increased in the tissues of patients with
aortic valve calcification and in primary human aortic valve
interstitial cells, whereas following treatment with recombinant
IL-32, the IL and TNF-α expression levels were increased (24). These findings suggested that
inhibition of IL-32 levels could target a variety of cardiovascular
diseases.
Furthermore, the present study demonstrated that
IL-32 may disrupt normal cardiomyocyte functions by activating the
NOD2/NOX2 signaling pathway. NOD2 is one of the earliest discovered
members of the Nod-like receptor family, which induces
self-oligomerization by recognizing pathogen-associated molecular
patterns (25); subsequently, it
activates NF-κB, promotes the transcription of pro-inflammatory
cytokines and initiates the immune-inflammatory response (26). In addition, increased NOX activity
causes a series of pathological conditions, such as the development
of cardiovascular disease (27).
The function of NOX in cardiomyocytes is divided into the two
following types: Immature type and mature type. The mature type is
mainly NOX2, which has shown increased expression during tissue
ischemia. NOX or ROS inhibitors have been revealed to significantly
reduce cell apoptosis (28,29).
Moreover, elevated NOX2 levels were detected in the coronary
arteries of patients with coronary atherosclerosis and in active
and necrotic cardiomyocytes of patients with acute myocardial
infarction (30,31). This phenomenon suggests that the
production of NOX2 by myocardial ischemia triggers myocardial
apoptosis, while the application of NOX2 inhibitors can protect the
myocardium and avoid ischemic injury. In the present study, IL-32
was shown to mediate the regulation of NOD2 and NOX2 expression in
cardiomyocytes. The application of the monoclonal antibody against
IL-32 or other inhibitors of IL-32 may be more effective in
alleviating cardiomyocyte apoptosis.
The MAPK signaling pathway is present in the
majority of cells and can transfer extracellular signals to cells
and their nuclei, which is intimately relevant to cell
proliferation and apoptosis (32).
The ERK signaling pathway is the earliest discovered canonical
Ras-Raf-MAPK signaling pathway (33). Cytokine receptors can activate the
ERK pathway (34). Activated ERK
can transfer signals into the nucleus and phosphorylate nuclear
transcription factors involved in the regulation of cell
proliferation and differentiation (35). Therefore, the present study
examined the expression levels of the ERK protein and the results
demonstrated that IL-32 promoted ERK phosphorylation by inducing
the activation of the NOD2/NOX2 signaling pathway. Since IL-32 is a
secreted protein, it is not clear whether its site of action is
intracellular or extracellular (36). In addition, our findings are based
on cellular experiments. The investigation of the function of IL-32
requires additional in vitro and in vivo studies.
In summary, the present study indicated that IL-32
participated in cardiomyocyte oxidative stress, inflammation, and
apoptosis during H/R treatment via the NOD2/NOX2/MAPK signaling
pathway. The current study demonstrated the potential role of IL-32
in MIRI and provides a theoretical basis for the development of
relevant therapeutic methods for this disease.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL and ZW contributed to design, experiments and
analysis. YL contributed to the draft and ZW revised the manuscript
for important intellectual content. YL and ZW have read and
approved the final manuscript, and also confirm the authenticity of
the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sethi NJ, Safi S, Korang SK, Hróbjartsson
A, Skoog M, Gluud C and Jakobsen JC: Antibiotics for secondary
prevention of coronary heart disease. Cochrane Database Syst Rev.
2(CD003610)2021.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Stolpe S, Kowall B and Stang A: Decline of
coronary heart disease mortality is strongly effected by changing
patterns of underlying causes of death: An analysis of mortality
data from 27 countries of the WHO European region 2000 and 2013.
Eur J Epidemiol. 36:57–68. 2021.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Woodruff RC, Casper M, Loustalot F and
Vaughan AS: Unequal local progress towards healthy people 2020
objectives for stroke and coronary heart disease mortality. Stroke.
52:e229–e232. 2021.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Aday AW and Matsushita K: Epidemiology of
peripheral artery disease and polyvascular disease. Circ Res.
128:1818–1832. 2021.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Hagström E, Norlund F, Stebbins A,
Armstrong PW, Chiswell K, Granger CB, López-Sendón J, Pella D,
Soffer J, Sy R, et al: Psychosocial stress and major cardiovascular
events in patients with stable coronary heart disease. J Intern
Med. 283:83–92. 2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Saito Y and Kobayashi Y: Update on
antithrombotic therapy after percutaneous coronary intervention.
Intern Med. 59:311–321. 2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Bakaeen FG, Gaudino M, Whitman G, Doenst
T, Ruel M, Taggart DP, Stulak JM, Benedetto U, Anyanwu A, Chikwe J,
et al: 2021: The American association for thoracic surgery expert
consensus document: Coronary artery bypass grafting in patients
with ischemic cardiomyopathy and heart failure. J Thorac Cardiovasc
Surg. 162:829–850.e1. 2021.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Chiang CY, Choi KC, Ho KM and Yu SF:
Effectiveness of nurse-led patient-centered care behavioral risk
modification on secondary prevention of coronary heart disease: A
systematic review. Int J Nurs Stud. 84:28–39. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Zheng J, Chen P, Zhong J, Cheng Y, Chen H,
He Y and Chen C: HIF-1α in myocardial ischemia-reperfusion injury
(review). Mol Med Rep. 23(352)2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Shen Y, Liu X, Shi J and Wu X: Involvement
of Nrf2 in myocardial ischemia and reperfusion injury. Int J Biol
Macromol. 125:496–502. 2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Xiang M, Lu Y, Xin L, Gao J, Shang C,
Jiang Z, Lin H, Fang X, Qu Y, Wang Y, et al: Role of Oxidative
stress in reperfusion following myocardial ischemia and its
treatments. Oxid Med Cell Longev. 2021(6614009)2021.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Han S and Yang Y: Interleukin-32: Frenemy
in cancer? BMB Rep. 52:165–174. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Kang JY and Kim KE: Prognostic value of
interleukin-32 expression and its correlation with the infiltration
of natural killer cells in cutaneous melanoma. J Clin Med.
10(4691)2021.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Yao Q, Wang B, Jia X, Li Q, Yao W and
Zhang JA: Increased human interleukin-32 expression is related to
disease activity of graves' disease. Front Endocrinol (Lausanne).
10(613)2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
de Albuquerque R, Komsi E, Starskaia I,
Ullah U and Lahesmaa R: The role of interleukin-32 in autoimmunity.
Scand J Immunol. 93(e13012)2021.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Netea MG, Lewis EC, Azam T, Joosten LA,
Jaekal J, Bae SY, Dinarello CA and Kim SH: Interleukin-32 induces
the differentiation of monocytes into macrophage-like cells. Proc
Natl Acad Sci USA. 105:3515–3520. 2008.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Joosten LA, Netea MG, Kim SH, Yoon DY,
Oppers-Walgreen B, Radstake TR, Barrera P, van de Loo FA, Dinarello
CA and van den Berg WB: IL-32, a proinflammatory cytokine in
rheumatoid arthritis. Proc Natl Acad Sci USA. 103:3298–3303.
2006.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Dinarello CA and Kim SH: IL-32, a novel
cytokine with a possible role in disease. Ann Rheum Dis. 65 (Suppl
3):iii61–iii64. 2006.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Cheng Y, Cheng L, Gao X, Chen S, Wu P,
Wang C and Liu Z: Covalent modification of Keap1 at Cys77 and
Cys434 by pubescenoside a suppresses oxidative stress-induced NLRP3
inflammasome activation in myocardial ischemia-reperfusion injury.
Theranostics. 11:861–877. 2021.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Teo KK and Rafiq T: Cardiovascular risk
factors and prevention: A perspective from developing countries.
Can J Cardiol. 37:733–743. 2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Gagno G, Ferro F, Fluca AL, Janjusevic M,
Rossi M, Sinagra G, Beltrami AP, Moretti R and Aleksova A: From
brain to heart: Possible role of amyloid-β in ischemic heart
disease and ischemia-reperfusion injury. Int J Mol Sci.
21(9655)2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Qin Z, Shen S, Qu K, Nie Y and Zhang H:
Mild hypothermia in rat with acute myocardial ischaemia-reperfusion
injury complicating severe sepsis. J Cell Mol Med. 25:6448–6454.
2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Tsai CL, Chiu YM, Lee YJ, Hsieh CT, Shieh
DC, Tsay GJ, Bau DT and Wu YY: Interleukin-32 plays an essential
role in human calcified aortic valve cells. Eur Cytokine Netw.
29:36–47. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Mann JK, Shen J and Park S: Enhancement of
muramyl dipeptide-dependent NOD2 activity by a self-derived
peptide. J Cell Biochem. 118:1227–1238. 2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Okanishi H, Hayashi K, Sakamoto Y, Sano T,
Maruyama H, Kagawa Y and Watari T: NOD2 mRNA expression and
NFkappaB activation in dogs with lymphocytic plasmacytic colitis. J
Vet Intern Med. 27:439–444. 2013.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Zhang Y, Murugesan P, Huang K and Cai H:
NADPH oxidases and oxidase crosstalk in cardiovascular diseases:
Novel therapeutic targets. Nat Rev Cardiol. 17:170–194.
2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Cadenas S: ROS and redox signaling in
myocardial ischemia-reperfusion injury and cardioprotection. Free
Radic Biol Med. 117:76–89. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Huang W, Xiong Y, Chen Y, Cheng Y and Wang
R: NOX2 is involved in CB2-mediated protection against lung
ischemia-reperfusion injury in mice. Int J Clin Exp Pathol.
13:277–285. 2020.PubMed/NCBI
|
|
30
|
Pejenaute Á, Cortés A, Marqués J, Montero
L, Beloqui Ó, Fortuño A, Martí A, Orbe J and Zalba G: NADPH oxidase
overactivity underlies telomere shortening in human
atherosclerosis. Int J Mol Sci. 21(1434)2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Sirker A, Murdoch CE, Protti A, Sawyer GJ,
Santos CX, Martin D, Zhang X, Brewer AC, Zhang M and Shah AM:
Cell-specific effects of Nox2 on the acute and chronic response to
myocardial infarction. J Mol Cell Cardiol. 98:11–17.
2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Sun Y, Liu WZ, Liu T, Feng X, Yang N and
Zhou HF: Signaling pathway of MAPK/ERK in cell proliferation,
differentiation, migration, senescence and apoptosis. J Recept
Signal Transduct Res. 35:600–604. 2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Guo YJ, Pan WW, Liu SB, Shen ZF, Xu Y and
Hu LL: ERK/MAPK signalling pathway and tumorigenesis. Exp Ther Med.
19:1997–2007. 2020.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Niogret C, Birchmeier W and Guarda G:
SHP-2 in lymphocytes' cytokine and inhibitory receptor signaling.
Front Immunol. 10(2468)2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Lavoie H, Gagnon J and Therrien M: ERK
signalling: A master regulator of cell behaviour, life and fate.
Nat Rev Mol Cell Biol. 21:607–632. 2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Hong JT, Son DJ, Lee CK, Yoon DY, Lee DH
and Park MH: Interleukin 32, inflammation and cancer. Pharmacol
Ther. 174:127–137. 2017.PubMed/NCBI View Article : Google Scholar
|