Introduction
Hemangioma is a benign tumor that is characterized
by endothelial hyperplasia in the blood vessels, which primarily
occurs in infants and female patients (1,2).
Patients with hemangioma may suffer from disease-related syndromes
such as PHACE and LUMBAR syndromes, or exhibit skin lesions during
the disease growth phase (2,3). To
reduce the risk of complications experienced by patients with
hemangioma, early identification by physical examination,
ultrasound or MRI allows for immediate intervention with
appropriate measures (3,4). Propranolol treatment, laser therapy
and surgical resection are among the potential treatment options
available for patients with hemangioma. However, certain patients
with hemangioma do not respond well to these treatment strategies
(2,5,6).
Therefore, to optimize the management protocols of patients with
hemangioma, it is necessary to explore the pathogenic mechanism of
this disease to identify novel treatment targets.
MicroRNAs (miRNAs/miRs) are a class of endogenous,
non-coding RNAs ~22 nucleotides in length, which mainly mediate the
post-transcriptional regulation of mRNA (7). Among miRNAs, miR-203 has been shown
to serve a key role in cell proliferation, migration and/or
invasion in several malignancies (8-10).
By decreasing the expression of regulator of G-protein signaling 17
(RGS17), miR-203 has been shown to suppress cell proliferation and
invasion in non-small cell lung cancer (8). In addition, miR-203 has been reported
to inhibit the proliferation and invasion of osteosarcoma cells,
but promote their apoptosis, by targeting runt-related
transcription factor 2 (RUNX2) expression (9). miR-203 has also been reported to
inhibit epithelial-mesenchymal transition and invasion of renal
cell carcinoma cells by inactivating the PI3K/AKT signaling pathway
(10). However, whether
miR-203a-3p exhibits similar effects on hemangioma remains unclear
and the molecular mechanism requires further investigation.
Vascular epithelial growth factor A (VEGFA) is a
critical regulator of a number of cellular processes, including
proliferation, apoptosis and invasion (11-13).
Notably, it has been demonstrated that VEGFA can serve a role in
the pathogenesis of hemangioma in several ways, including through
the promotion of angiogenesis (14). In addition, GG genotype of +405 G/C
VEGFA polymorphism is associated with a lower risk for infantile
hemangioma (15). The PI3K/AKT
pathway has been shown to regulate various biological processes,
including cell survival, mobility and malignant transformation
(16,17). In particular, this signaling
pathway has been reported to be associated with the pathogenesis
and progression of hemangioma (18).
Our previous study revealed that miR-203a-3p was
involved in the regulation of hemangioma progression mediated by
long non-coding RNA MEG8(19).
Therefore, the present study aimed to explore the effects of
miR-203a-3p on endothelial cell proliferation, apoptosis and
invasion in hemangioma, with specific focus on its downstream
targets and associated signaling pathways.
Materials and methods
Cell lines
Human primary hemangioma endothelial cells (HemECs)
and the 293T cell line were purchased from the BeNa Culture
Collection. HemECs were maintained in complete endothelial cell
medium (Gibco; Thermo Fisher Scientific, Inc.), whereas the 293T
cells were maintained in DMEM (Gibco; Thermo Fisher Scientific,
Inc.) containing 10% FBS (Gibco; Thermo Fisher Scientific, Inc.).
All cells were cultured in a humidified incubator with 5%
CO2 at 37˚C. Ethics approval for the use of HemECs was
waived by the Ethics Committee of the Affiliated Hospital of Hebei
University of Engineering (Handan, China).
Cell transfection
miR-203a-3p mimics, the miR-203a-3p inhibitor and
their corresponding negative controls (NCs) were purchased from
Changchun Changsheng Gene Biotechnology Co., Ltd. The VEGFA
overexpression plasmid (pEX2-VEGFA) and the empty NC plasmid
(pEX2-NC) were purchased from Shenzhen Morecell Biomedical Co.,
Ltd. HemECs (1x106 cells) were transfected with 50 nM
miR-203a-3p mimics, 50 nM miR-203a-3p inhibitors, 50 nM NC mimics,
50 nM NC inhibitors, 0.8 µg pEX2-VEGF or 0.8 µg pEX2-NC by
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol at 37˚C
for 6 h. The sequences for miR-203a-3p or NC mimics and inhibitors
used were as follows: miR-203a-3p mimics,
5'-GUGAAAUGUUUAGGACCACUAG-3'; miR-203a-3p inhibitors,
5'-CUAGUGGUCCUAAACAUUUCAC-3'; NC mimics,
5'-UUGUACUACACAAAAGUACUG-3' and NC inhibitors,
5'-CAGUACUUUUGUGUAGUACAA-3'.
miR-203a-3p manipulation
The cultured HemECs were divided into the following
five groups: i) Control group (without any treatment); ii) NC mimic
group (transfected with NC mimics); iii) miR mimic group
(transfected with miR-203a-3p mimics); iv) NC inhibitor group
(transfected with the NC inhibitor); and v) miR inhibitor group
(transfected with the miR-203a-3p inhibitor). Reverse
transcription-quantitative PCR (RT-qPCR), cell proliferation,
apoptosis, invasion and western blotting assays were performed
following transfection.
Luciferase reporter assay
The binding sites between miR-203a-3p and VEGF were
predicted using RNAhybrid v22 (https://bibiserv.cebitec.uni-bielefeld.de/rnahybrid).
The 3'untranslated regions of VEGFA containing miR-203a-3p-binding
wild type (WT) and mutant type (MT) sites were inserted into the
pGLuc vector, which was synthesized by Beyotime Institute of
Biotechnology. The 0.8 µg VEGFA-WT or 0.8 µg VEGFA-MT plasmids were
then co-transfected with the 50 nM miR-203a-3p mimics or 50 nM NC
mimics into 1x106 293T cells using Lipofectamine 2000.
Following cell culture for 48 h, the cells were harvested and
assessed using a luciferase assay kit (Beyotime Institute of
Biotechnology) according to the kit's protocol. The luciferase
enzyme activity was normalized to Renilla luciferase enzyme
activity.
VEGFA regulation experiment
HemECs were cultured and divided into the following
five groups: i) Control group (without any treatment), ii) NC mimic
+ pEX2-NC group (co-transfected with NC mimics and the pEX2-NC
plasmid), iii) miR mimic + pEX2-NC group (co-transfected with
miR-203a-3p mimics and the pEX2-NC plasmid), iv) NC mimic +
pEX2-VEGFA group (co-transfected with NC mimics and the pEX2-VEGFA
plasmid) and v) miR mimic + pEX2-VEGFA group (co-transfected with
miR-203a-3p mimics and the pEX2-VEGFA plasmid). Further
experiments, namely RT-qPCR, western blotting, cell proliferation,
apoptosis and invasion assays, were performed following
transfection.
RT-qPCR
Total RNA was extracted from the HemECs using a
RNeasy Mini kit (Qiagen China Co. Ltd.) 48 h after transfection.
The RNA was reverse transcribed into cDNA using a High Capacity
cDNA Reverse Transcription Kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.) according to the kit's protocol. qPCR was
performed using a SYBR®-Green PCR Master Mix (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The qPCR was carried
out with following procedure: 95˚C hold for 10 min; 40 cycles of
denaturation at 95˚C for 15 sec and annealing/extension at 61˚C for
30 sec. The expression levels of miR-203a-3p and VEGFA were
evaluated using the 2-ΔΔCq method
(20) with U6 or β-actin used as
endogenous controls, respectively. The primer sequences are listed
in Table SI.
Cell proliferation assay
Cell Counting Kit-8 (CCK-8; MilliporeSigma) was
utilized to assess the proliferation of HemECs. Briefly, HemECs
were plated in 96-well plates (2x103 cells/well). At 0,
24, 48 and 72 h following transfection, 10 µl CCK-8 reagent was
added before the mixture was incubated for an additional 2 h at
37˚C. The optical density values at 450 nm were subsequently
evaluated using an automated enzyme immunoassay analyzer (BioTek
China).
Cell apoptosis assay
A TUNEL apoptosis kit (Beyotime Institute of
Biotechnology) was used for detecting the apoptosis of HemECs
following transfection. Briefly, transfected HemECs were plated
into 24-well plates (1x104 cells/well) and cultured for
48 h. The HemECs were then fixed with 4% paraformaldehyde (Beyotime
Institute of Biotechnology) for 30 min at 37˚C and incubated with
0.1% Triton X-100 (Beyotime Institute of Biotechnology) for 5 min
at 4˚C. Subsequently, 50 µl TUNEL solution was added and the cells
were incubated for 1 h at 37˚C, followed by DAPI (Sangon Biotech
Co., Ltd.) staining for 15 min at room temperature. After adding
Antifade Mounting Medium (Beyotime Institute of Biotechnology), the
cells were sealed and observed under an inverted fluorescence
microscope (Motic China Group Co., Ltd.) with 3 random fields being
chosen.
Cell invasion assay
The invasive ability of HemECs following
transfection was assessed using Transwell chambers. Briefly, 48 h
following transfection, HemECs, which were suspended in 1% FBS
containing complete endothelial cell medium, were seeded into the
upper Matrigel-coated chambers (Corning, Inc.) at a density of
5x104 cells/well, whilst complete endothelial cell
medium containing 10% FBS was added into the lower chamber. The
upper chamber was coated by 70 µl Matrigel, which was diluted with
complete endothelial cell medium at a ratio of 1:8, at 37˚C for 1
h. The cells were cultured for 24 h before their invasive ability
was assessed by staining with 0.5% crystal violet (MilliporeSigma)
for 15 min at room temperature after being fixed with 4%
paraformaldehyde (MilliporeSigma). The images were taken under an
inverted light microscope (Motic China Group Co., Ltd.).
Western blot analysis
Total proteins from HemECs at 48 h following
transfection were lysed using RIPA buffer (MilliporeSigma) and
quantified using bicinchoninic acid assay (Thermo Fisher
Scientific, Inc.). Of the extracted proteins, 20 µg per lane were
separated by 4-20% SDS-PAGE and transferred onto nitrocellulose
membranes (Pall Life Sciences). After blocking with 5% BSA
(MilliporeSigma) at 37˚C for 1 h, the membranes were incubated at
4˚C overnight with primary antibodies (all obtained from
Invitrogen; Thermo Fisher Scientific, Inc.) against Bcl-2 (1:1,000
dilution; cat. no. PA5-11379), cleaved (C)-caspase 3 (1:1,000; cat.
no. PA5-96077), VEGFA (1:1,000; cat. no. PA5-16754), phosphorylated
(p)-PI3K (1:1,000; cat. no. PA5-118549), PI3K (1:1,000; cat. no.
PA5-99518), p-AKT (1:1,000; cat. no. MA5-38243), AKT (1:1,000; cat.
no. 44-609G) and β-actin (1:2,000; cat. no. PA5-85291).
Subsequently, the membranes were incubated with a HRP-conjugated
goat anti-rabbit secondary antibody (1:10,000; cat. no. 65-6120;
Invitrogen; Thermo Fisher Scientific, Inc.) at 37˚C for 1 h. The
bands were visualized using the ECL-PLUS reagent (Thermo Fisher
Scientific, Inc.). Densitometric quantification was completed by
ImageJ 1.8 (National Institutes of Health).
Statistical analysis
GraphPad Prism 7.0 (GraphPad Software, Inc.) was
used to analyze the data, which were triplicates and exhibited as
mean ± standard deviation. One-way ANOVA followed by Tukey's
post-hoc test was used for comparisons among multiple groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of miR-203a-3p on the
proliferation, apoptosis and invasion of HemECs
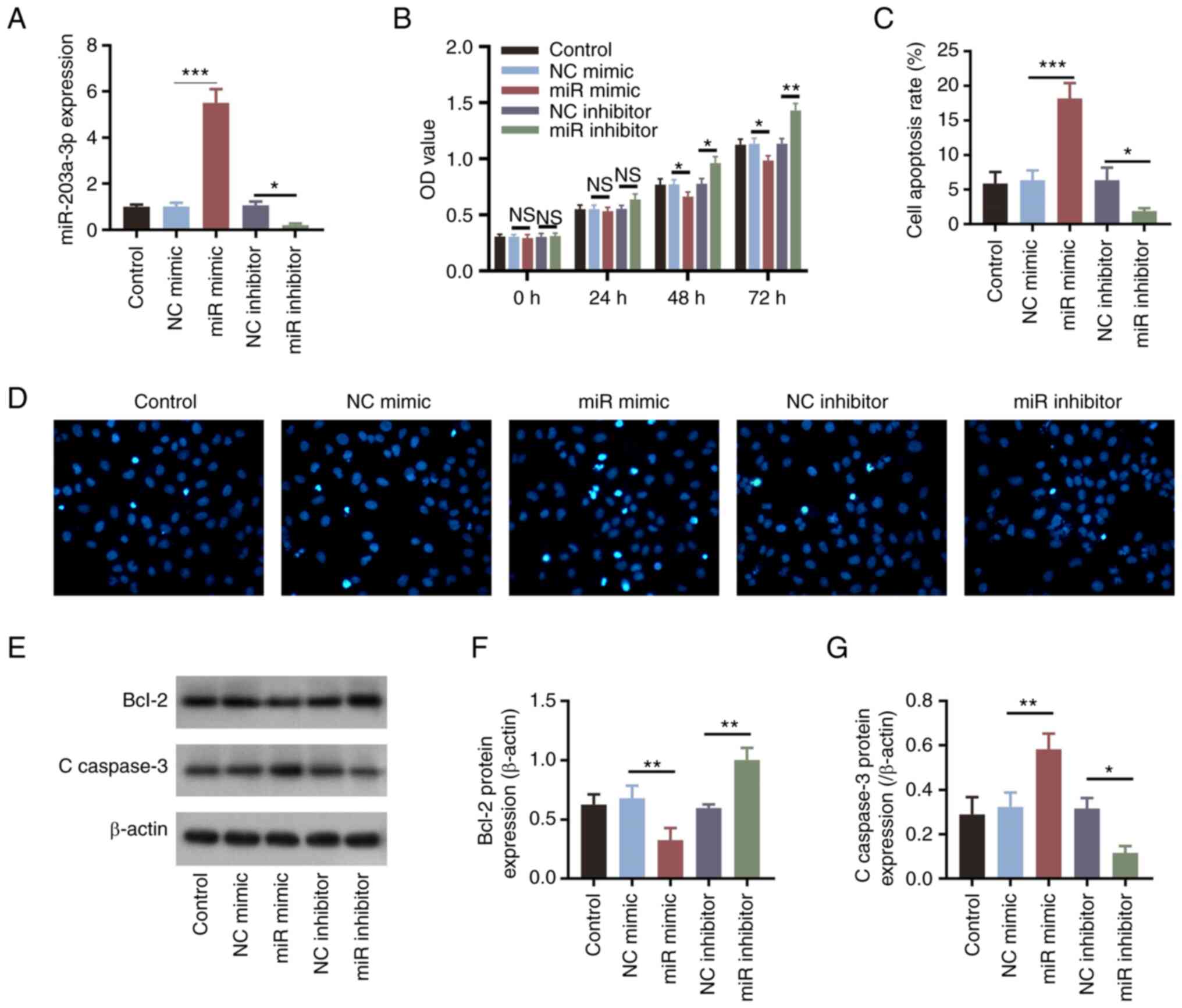
The expression levels of miR-203a-3p were
significantly increased following transfection of the cells with
miR-203a-3p mimics compared with those in the NC mimics group
(P<0.001; Fig. 1A). By
contrast, miR-203a-3p expression was significantly decreased
following the transfection of HemECs with the miR-203a-3p inhibitor
compared with that in the NC inhibitor group (P<0.05; Fig. 1A). These observations suggested
that transfection was successful. In addition, compared with those
in their corresponding NC groups, miR-203a-3p mimics and
miR-203a-3p inhibitor transfection induced significant decreases
and increases in cell proliferation at 48 and 72 h, respectively
(all P<0.05; Fig. 1B).
Subsequently, it was revealed that apoptosis was significantly
increased by transfection with the miR-203a-3p mimics (P<0.001)
but significantly alleviated by the miR-203a-3p inhibitor
(P<0.05; Fig. 1C and D), compared with that in their
corresponding NC groups. Furthermore, the expression levels of
apoptotic markers Bcl-2 and C-caspase 3 were measured; miR-203a-3p
mimics transfection significantly suppressed Bcl-2 expression
whilst increasing that of C-caspase 3, whereas transfection with
the miR-203a-3p inhibitor resulted in the opposite effects compared
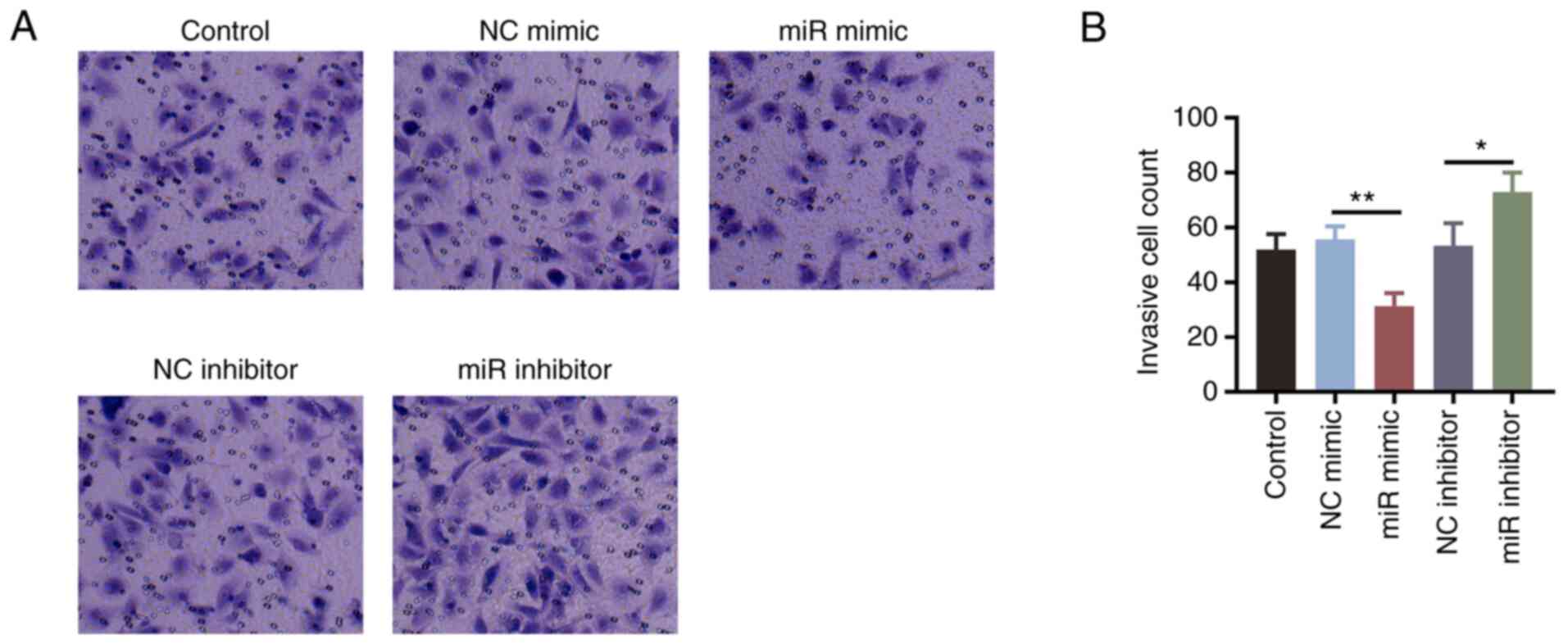
with miR-203a mimics (all P<0.05; Fig. 1E-G). The number of invasive cells
was significantly decreased in cells transfected with miR-203a-3p
mimics (P<0.01) but significantly elevated in cells transfected
with the miR-203a-3p inhibitor (P<0.05; Fig. 2A and B), compared with that in their
corresponding NC groups.
miR-203a-3p regulates VEGFA expression
and the PI3K/AKT signaling pathway in HemECs
A binding site on the 3'untranslated region of VEGFA
mRNA for miR-203a-3p was predicted (Fig. 3A). Therefore, VEGFA-MT and VEGFA-WT
sequences were designed based on this binding site. The relative
luciferase activity was significantly decreased (P<0.001) in
cells co-transfected with VEGFA-WT and miR-203a-3p mimics compared
with that in cells transfected with the NC mimics, suggesting
direct binding between VEGFA mRNA and miR-203a-3p; however, the
relative luciferase activity did not differ between cells
co-transfected with VEGFA-MT and miR-203a-3p mimics compared with
that in cells transfected with the NC mimics (Fig. 3B). In addition, miR-203a-3p mimics
transfection significantly decreased the expression levels of
VEGFA, whereas transfection with the miR-203a-3p inhibitor exerted
opposite effects (all P<0.05; Fig.
3C-E), compared with those in their corresponding NC groups.
These results suggested that miR-203a-3p may negatively regulate
VEGFA expression. In addition, compared with those in their
corresponding NC groups, miR-203a-3p overexpression significantly
decreased the phosphorylation levels of PI3K and AKT, whereas
miR-203a-3p knockdown resulted in the opposite effects (all
P<0.05; Fig. 3F and G).
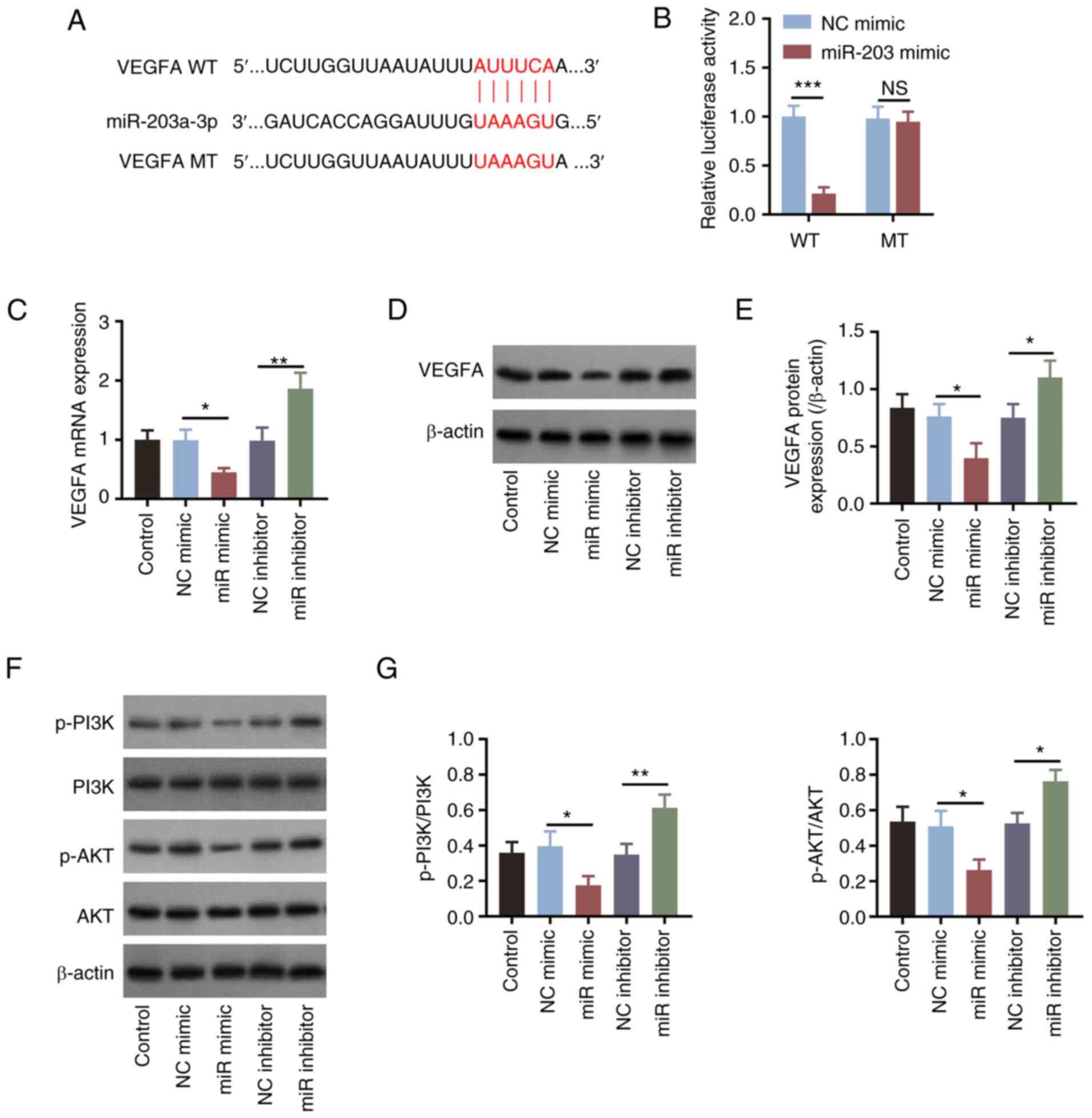 | Figure 3miR-203a-3p downregulates VEGFA
expression and PI3K/AKT signaling in HemECs. (A) Design of the WT
and MT sequences of VEGFA for luciferase reporter gene assays. (B)
Comparison of relative luciferase activity among the WT and MT
groups after miR-203 mimics co-transfection. (C) Comparison of
VEGFA mRNA expression levels among the groups after the
manipulation of miR-203 expression. (D) Detection of the VEGFA
protein expression by western blotting after the manipulation of
miR-203 expression, (E) which was semi-quantified. (F) Detection of
PI3K and AKT phosphorylation by western blot analysis. (G)
Semi-quantification of the p-PI3K/PI3K and p-AKT/AKT ratios among
the groups in HemECs following the manipulation of miR-203
expression. *P<0.05, **P<0.01 and
***P<0.001. miR-203a-3p, microRNA-203a-3p; VEGF,
vascular endothelial growth factor; WT, wild type; MT, mutant type;
p-, phosphorylated; HemECs, human hemangioma endothelial cells; NS,
non-significant; NC, negative control. |
miR-203a-3p regulates proliferation,
apoptosis and invasion by inhibiting VEGFA in HemECs
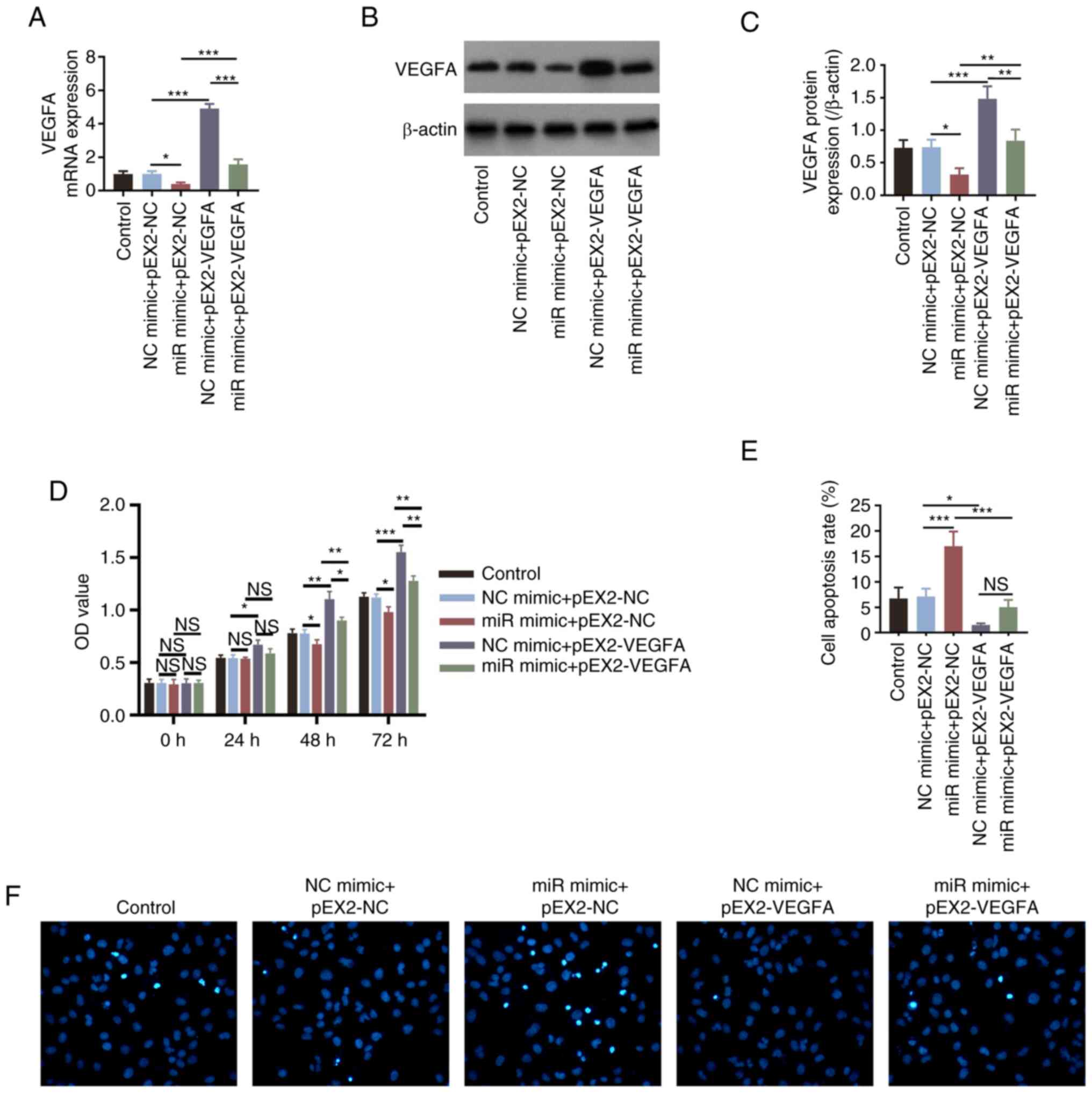
Transfection of HemECs with the VEGFA overexpression
plasmids significantly increased the expression levels of VEGFA
(all P<0.001; Fig. S1A-C) and
attenuated miR-203a-3p mimics-induced reduction of VEGFA (all
P<0.001; Fig. 4A-C), suggesting
successful transfection. Subsequently, overexpression of VEGFA
significantly increased cell proliferation (all P<0.05; Fig. S1D) and also significantly reversed
the miR-203a-3p mimics-induced decreases in cell proliferation
after 48 and 72 h in HemECs (all P<0.01; Fig. 4D). By contrast, VEGFA
overexpression significantly inhibited cell apoptosis (P<0.01;
Fig. S1E and F), in addition to significantly
reversing the miR-203a-3p mimics-induced increases in HemEC
apoptosis (P<0.001; Fig. 4E and
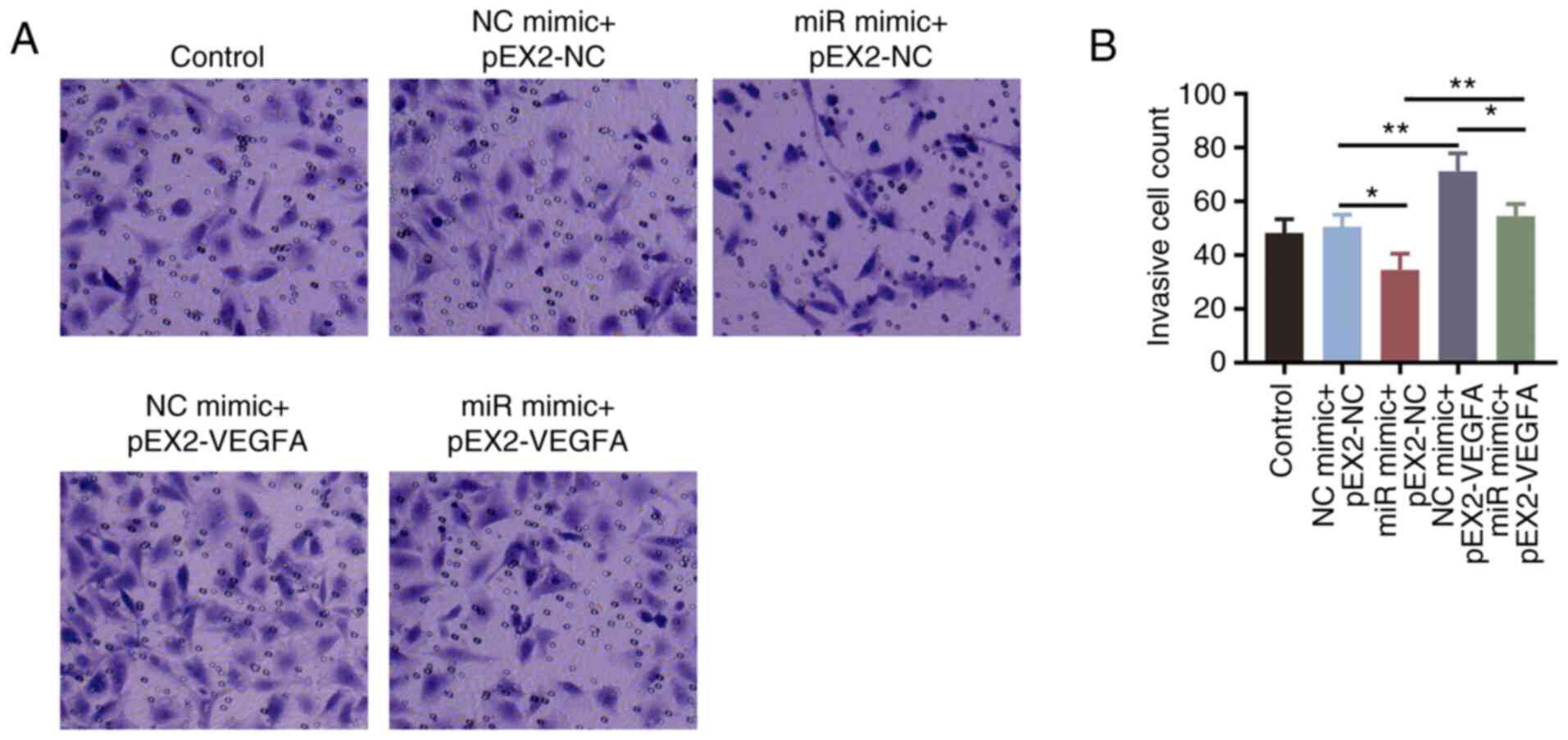
F). The number of invasive cells
was found to be significantly increased by VEGFA overexpression
(P<0.01; Fig. S1G and H), which also significantly reversed the
miR-203a-3p mimics-induced reductions in invasive cell numbers
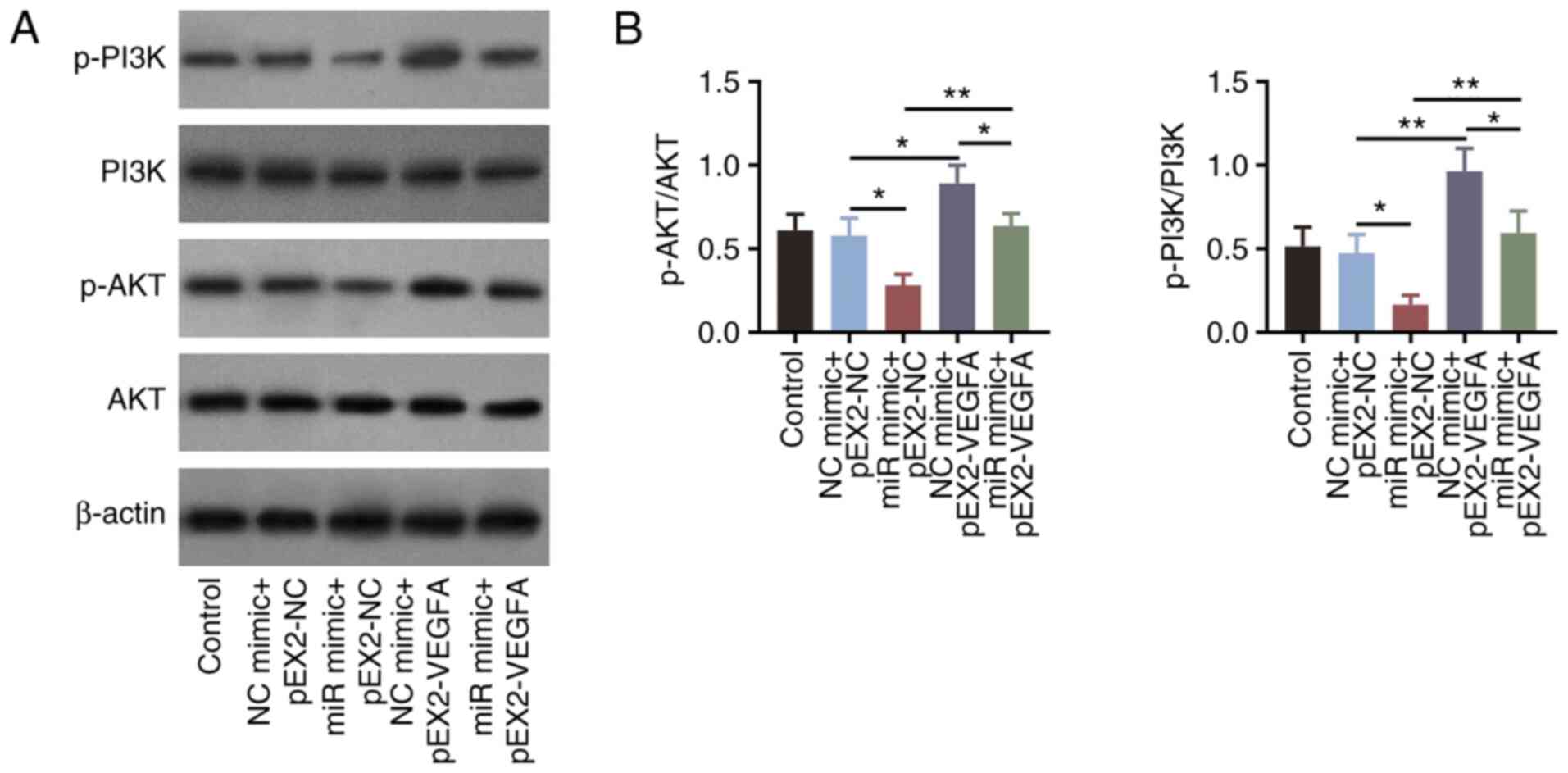
(P<0.01; Fig. 5A and B).
miR-203a-3p inhibits the PI3K/AKT
signaling pathway by targeting VEGFA in HemECs
To further explore if miR-203a-3p could regulate the
PI3K/AKT signaling pathway through VEGFA, additional experiments
were performed. Overexpression of VEGFA significantly increased the
phosphorylation levels of PI3K (P<0.01) and AKT (P<0.05;
Fig. S1I-K). In addition,
overexpression of VEGFA significantly reversed the miR-203a-3p
mimics-induced decreases in PI3K and AKT phosphorylation in HemECs
(all P<0.01; Fig. 6A and
B).
Discussion
A number of miRNAs have been reported to be involved
in the progression of hemangioma. In particular, miR-200c-3p has
been reported to promote the proliferation of vascular endothelial
cells by targeting the Notch signaling pathway, which serves a key
role in the pathogenesis of hemangioma (21). In another previous study,
downregulation of miR-556-3p expression was reported to suppress
hemangioma cell proliferation whilst accelerating apoptosis by
targeting VEGFC expression (22).
Furthermore, miR-139-5p may negatively modulate cell proliferation
and migration in hemangioma stem cells by targeting insulin-like
growth factor 1(23). Other
studies have also reported that some miRNAs can regulate the
progression of hemangioma, including miR-382-5p and miR-125b-5p
(24,25). Although these previous studies have
revealed miRNAs can regulate the proliferation and apoptosis of
hemangioma cells, it remains necessary to explore alternative
biological pathways that are involved in the biological functions
of hemangioma cells. This information could then be applied for
designing potential treatment strategies for hemangioma. Our
preliminary study revealed that miR-203 expression can be regulated
by the lncRNA MEG8 and may participate in the modulation of
hemangioma progression via inhibiting the Notch signaling pathway
(19). However, the molecular
mechanism by which miR-203a-3p can participate in modulation of
hemangioma progression remains unclear. As a miRNA that has been
extensively studied, miR-203 functions as a tumor suppressor in a
variety of malignancies (26).
Overexpression of miR-203 has been found to inhibit cell invasion
and proliferation in hepatocellular carcinoma cells (27), whereas the expression levels of
miR-203 have been revealed to be downregulated during lung cancer
tumorigenesis (8). In lung cancer
cells, miR-203 can suppress the invasion and proliferation of the
A549 and Calu-1 cell lines by targeting RGS17 expression (8). In estrogen-dependent breast cancer
cells, miR-203 has been reported to inhibit their viability and
migration by suppressing the protein expression of estrogen
receptor α (28). In addition,
miR-203 knockdown has been revealed to inhibit cisplatin-induced
cell apoptosis in osteosarcoma cells in vitro by targeting
RUNX2 expression (29). Therefore,
it is reasonable to hypothesize that miR-203 may participate in the
regulation of cell viability, apoptosis and migration in
hemangioma. In the present study, overexpression of miR-203a-3p
decreased the proliferation and invasion of HemECs, whilst
increasing apoptosis of these cells. By contrast, knocking down
miR-203a-3p expression resulted in the opposite effects. Possible
reasons for these findings could be that miR-203a-3p regulated
activation of the VEGFA-mediated PI3K/AKT pathway upstream of cell
proliferation, apoptosis and invasion, or that miR-203a-3p could
have regulated cell invasion and proliferation by targeting RGS17
expression (8).
The VEGF family consists of cytokines known to be
angiogenic factors that participate in blood vessel formation and
lymphangiogenesis (30). VEGFA is
the main member of this family of proteins, which has been revealed
to regulate cell proliferation, apoptosis and invasion (11-13,31,32).
In particular, VEGFC has also been reported to impact the
progression of hemangioma (22). A
previous study suggested that VEGFA serves a key role in the
pathogenesis of hemangioma, specifically through the proliferation
and invasion physiological pathways (24). In addition, the miR-206/VEGFA axis
has been documented to serve an important role in the suppression
of HemEC proliferation after the expression of
metastasis-associated lung adenocarcinoma transcript 1 has been
knocked down (33). It has also
been shown that VEGFA is a master regulator of angiogenesis in
hemangioma (14,34). In the present study, it was
demonstrated that VEGFA could regulate hemangioma proliferation,
apoptosis and invasion. According to previous studies, VEGFA is one
of the targets of miR-203a-3p and has been shown to be a key
regulatory mechanism during the pathogenesis of laryngeal carcinoma
and progression of cervical cancer (35,36).
In the present study, it was found that miR-203a-3p regulated the
cellular functions of hemangioma through VEGFA. The reason could be
attributed to VEGFA being a direct target of miR-203a-3p as
previously reported (35).
AKT is a serine threonine kinase acting downstream
of PI3K, which regulates a high number of critical cellular
processes (37). Inhibition of the
PI3K/AKT/mTOR signaling pathway has been shown to restore the
autophagic process in epidermal keratinocyte cells from psoriatic
mice (38). In addition,
suppression of AKT/mTOR signaling has been observed to promote
chemo-radiotherapy resistance in lung squamous cell carcinoma cells
(39). In another study, AKT was
found to serve a critical role during the life cycle of acute and
persistent murine norovirus strains in macrophages (40). In addition to these cellular
activities, it has been reported that the PI3K/AKT signaling
pathway serves a key role in hemangioma. According to an earlier
study, activation of the PI3K/AKT signaling pathway may affect
tumorigenesis of lung hemangioma through the downstream mTOR
(41). Furthermore, the PI3K/AKT
pathway has been demonstrated to suppress hemangioma proliferation
by downregulating the expression of proliferating cell nuclear
antigen, which may further influence the progression of hemangioma
(42). In the present study, it
was found that the PI3K/AKT signaling pathway was regulated by
miR-203a-3p in hemangioma, which was in line with the data
previously reported in other tumors, such as renal cell carcinoma,
papillary thyroid cancer and glioblastoma (10,43,44).
In addition, it was confirmed that miR-203a-3p can directly
suppress VEGFA expression and the PI3K/AKT pathway, which could
suppress the proliferation of hemangioma, thereby uncovering the
miR-203a-3pVEGFA/PI3K/AKT axis.
In conclusion, in the present study, miR-203a-3p
overexpression was found to inhibit endothelial cell proliferation,
suppress invasion and promote apoptosis by inactivating the
VEGFA-mediated PI3K/AKT pathway in hemangioma. These findings may
assist in identifying suitable targets for the management of
hemangioma.
Supplementary Material
VEGFA can regulate cell proliferation,
apoptosis, invasion and PI3K/AKT signaling in HemECs. (A)
Comparison of VEGFA mRNA expression among the groups after VEGFA
overexpression. (B) Detection of VEGFA protein expression by
western blotting after VEGFA overexpression, (C) which was
semi-quantified. (D) Cell proliferation and (E) apoptosis rates
among the groups after VEGFA overexpression. (F) Representative
images of TUNEL staining in E (magnification, x200). (G) Comparison
of invaded cells among the groups after VEGFA overexpression. (H)
Representative images of invasive cells after crystal violet
staining (magnification, x200). (I) Detection of PI3K and AKT
phosphorylation by western blotting after VEGFA overexpression.
Comparison of (J) p-PI3K/PI3K and (K) p-AKT/AKT ratios,
*P<0.05, **P<0.01 and
***P<0.001. NC, negative control; OD, optical
density; p-, phosphorylated; VEGF, vascular endothelial growth
factor; NS, not significant.
Primers used for reverse transcription
quantitative PCR.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by funds received from
the following grant: Scientific Research Fund Project of Hebei
Provincial Health Commission, Molecular Mechanism of Propranolol
Induced Apoptosis in Hemangioma Endothelial Cells (grant no.
20200582).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZH and LZ designed the present study. Material
preparation and data collection were performed by YL and DD. ZH, LZ
and JG analyzed the data. All authors contributed to the
interpretation of data, methodology and reviewing and editing of
the manuscript. ZH and LZ confirm the authenticity of all the raw
data. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Ethics approval for the use of HemECs was waived for
the present study by the Ethics Committee of the Affiliated
Hospital of Hebei University of Engineering (Handan, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jung HL: Update on infantile hemangioma.
Clin Exp Pediatr. 64:559–572. 2021.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Sebaratnam DF, Rodriguez Bandera AL, Wong
LF and Wargon O: Infantile hemangioma. Part 2: Management. J Am
Acad Dermatol. 85:1395–1404. 2021.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Valdebran M and Wine Lee L:
Hemangioma-related syndromes. Curr Opin Pediatr. 32:498–505.
2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
DeHart A and Richter G: Hemangioma: Recent
advances. F1000Res. 8(F1000)2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ji Y, Chen S, Wang Q, Xiang B, Xu Z, Zhong
L, Yang K, Lu G and Qiu L: Intolerable side effects during
propranolol therapy for infantile hemangioma: frequency, risk
factors and management. Sci Rep. 8(4264)2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Kowalska M, Debek W and Matuszczak E:
Infantile hemangiomas: An update on pathogenesis and treatment. J
Clin Med. 10(4631)2021.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Rolle K, Piwecka M, Belter A, Wawrzyniak
D, Jeleniewicz J, Barciszewska MZ and Barciszewski J: The sequence
and structure determine the function of mature human miRNAs. PLoS
One. 11(e0151246)2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Chi Y, Jin Q, Liu X, Xu L, He X, Shen Y,
Zhou Q, Zhang J and Jin M: miR-203 inhibits cell proliferation,
invasion, and migration of non-small-cell lung cancer by
downregulating RGS17. Cancer Sci. 108:2366–2372. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lin W, Zhu X, Yang S, Chen X, Wang L,
Huang Z, Ding Y, Huang L and Lv C: MicroRNA-203 inhibits
proliferation and invasion, and promotes apoptosis of osteosarcoma
cells by targeting Runt-related transcription factor 2. Biomed
Pharmacother. 91:1075–1084. 2017.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Han N, Li H and Wang H: MicroRNA-203
inhibits epithelial-mesenchymal transition, migration, and invasion
of renal cell carcinoma cells via the inactivation of the PI3K/AKT
signaling pathway by inhibiting CAV1. Cell Adh Migr. 14:227–241.
2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Ma M, Zhang J, Gao X, Yao W, Li Q and Pan
Z: miR-361-5p Mediates SMAD4 to promote porcine granulosa cell
apoptosis through VEGFA. Biomolecules. 10(1281)2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zhang T, Qian Y, Yuan C, Wu Y, Qian H, Lu
H, Hu C and Li W: Propranolol suppresses proliferation and
migration of HUVECs through regulation of the miR-206/VEGFA axis.
Biomed Res Int. 2021(7629176)2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Geng A, Luo L, Ren F, Zhang L, Zhou H and
Gao X: miR-29a-3p inhibits endometrial cancer cell proliferation,
migration and invasion by targeting VEGFA/CD C42/PAK1. BMC Cancer.
21(843)2021.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Makkeyah SM, Elseedawy ME, Abdel-Kader HM,
Mokhtar GM and Ragab IA: Vascular endothelial growth factor
response with propranolol therapy in patients with infantile
hemangioma. Pediatr Hematol Oncol. 39:215–224. 2022.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Oszajca K, Szemraj J, Wyrzykowski D,
Chrzanowska B, Salamon A and Przewratil P: Single-nucleotide
polymorphisms of VEGF-A and VEGFR-2 genes and risk of infantile
hemangioma. Int J Dermatol. 57:1201–1207. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Sanaei MJ, Razi S, Pourbagheri-Sigaroodi A
and Bashash D: The PI3K/Akt/mTOR pathway in lung cancer; oncogenic
alterations, therapeutic opportunities, challenges, and a glance at
the application of nanoparticles. Transl Oncol.
18(101364)2022.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Barzegar Behrooz A, Talaie Z, Jusheghani
F, Los MJ, Klonisch T and Ghavami S: Wnt and PI3K/Akt/mTOR survival
pathways as therapeutic targets in glioblastoma. Int J Mol Sci.
23(1353)2022.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Ji Y, Chen S, Li K, Li L, Xu C and Xiang
B: Signaling pathways in the development of infantile hemangioma. J
Hematol Oncol. 7(13)2014.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Hu Z, Liu X, Guo J, Zhuo L, Chen Y and
Yuan H: Knockdown of lncRNA MEG8 inhibits cell proliferation and
invasion, but promotes cell apoptosis in hemangioma, via
miR-203-induced mediation of the Notch signaling pathway. Mol Med
Rep. 24(872)2021.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Hu X, Bai S, Li L, Tian P, Wang S, Zhang
N, Shen B, Du J and Liu S: MiR-200c-3p increased HDMEC
proliferation through the notch signaling pathway. Exp Biol Med
(Maywood). 246:897–905. 2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Jin W, Chen L, Gao F, Yang M, Liu Y and
Wang B: Down-regulation of miR-556-3p inhibits hemangioma cell
proliferation and promotes apoptosis by targeting VEGFC. Cell Mol
Biol (Noisy-le-grand). 66:204–207. 2020.PubMed/NCBI
|
|
23
|
Wu Y, Li H, Xie J, Wang F, Cao D and Lou
Y: miR1395p affects cell proliferation, migration and adipogenesis
by targeting insulinlike growth factor 1 receptor in hemangioma
stem cells. Int J Mol Med. 45:569–577. 2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Yuan X, Xu Y, Wei Z and Ding Q: CircAP2A2
acts as a ceRNA to participate in infantile hemangiomas progression
by sponging miR-382-5p via regulating the expression of VEGFA. J
Clin Lab Anal. 34(e23258)2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Wang T, Zhang FL, Zhao Y, Guo DD and Yang
R: Effects of miR-125b-5p on the proliferation and apoptosis of
human hemangioma endothelial cells HemES and its mechanism.
Zhongguo Ying Yong Sheng Li Xue Za Zhi. 37:247–253. 2021.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
26
|
Zong M, Feng W, Wan L, Yu X and Yu W:
miR-203 affects esophageal cancer cell proliferation, apoptosis and
invasion by targeting MAP3K1. Oncol Lett. 20:751–757.
2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Chen H, Kong M, Chen Y, Jiang Y, Wen M and
Zhang X: Prognostic significance of miR-203 and ZEB1 expression in
early-stage hepatocellular carcinoma. J Cancer. 12:4810–4818.
2021.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Lin J, Wang L, Gao J and Zhu S: MiR-203
inhibits estrogen-induced viability, migration and invasion of
estrogen receptor alpha-positive breast cancer cells. Exp Ther Med.
14:2702–2708. 2017.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Huang Z, Huang L, Liu L, Wang L, Lin W,
Zhu X, Su W and Lv C: Knockdown of microRNA-203 reduces cisplatin
chemo-sensitivity to osteosarcoma cell lines MG63 and U2OS in vitro
by targeting RUNX2. J Chemother. 33:328–341. 2021.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Shibuya M and Claesson-Welsh L: Signal
transduction by VEGF receptors in regulation of angiogenesis and
lymphangiogenesis. Exp Cell Res. 312:549–560. 2006.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Iyer S and Acharya KR: Tying the knot: The
cystine signature and molecular-recognition processes of the
vascular endothelial growth factor family of angiogenic cytokines.
FEBS J. 278:4304–4322. 2011.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Ye X, Gaucher JF, Vidal M and Broussy S: A
structural overview of vascular endothelial growth factors
pharmacological ligands: From macromolecules to designed
peptidomimetics. Molecules. 26(6759)2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wang S, Ren L, Shen G, Liu M and Luo J:
The knockdown of MALAT1 inhibits the proliferation, invasion and
migration of hemangioma endothelial cells by regulating
MiR-206/VEGFA axis. Mol Cell Probes. 51(101540)2020.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Cai C, Böttcher MC, Werner JA and Mandic
R: Differential expression of VEGF121, VEGF165 and VEGF189 in
angiomas and squamous cell carcinoma cell lines of the head and
neck. Anticancer Res. 30:805–810. 2010.PubMed/NCBI
|
|
35
|
Xu L, Shen B, Chen T and Dong P: miR-203
is involved in the laryngeal carcinoma pathogenesis via targeting
VEGFA and Cox-2. Onco Targets Ther. 9:4629–4637. 2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Zhu X, Er K, Mao C, Yan Q, Xu H, Zhang Y,
Zhu J, Cui F, Zhao W and Shi H: miR-203 suppresses tumor growth and
angiogenesis by targeting VEGFA in cervical cancer. Cell Physiol
Biochem. 32:64–73. 2013.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Dimri M and Satyanarayana A: Molecular
signaling pathways and therapeutic targets in hepatocellular
carcinoma. Cancers (Basel). 12(491)2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Tang ZL, Zhang K, Lv SC, Xu GW, Zhang JF
and Jia HY: LncRNA MEG3 suppresses PI3K/AKT/mTOR signalling pathway
to enhance autophagy and inhibit inflammation in TNF-α-treated
keratinocytes and psoriatic mice. Cytokine.
148(155657)2021.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Shen C, Shyu DL, Xu M, Yang L, Webb A,
Duan W and Williams TM: Deregulation of AKT-mTOR signaling
contributes to chemoradiation resistance in lung squamous cell
carcinoma. Mol Cancer Res. 20:425–433. 2022.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Owusu IA, Passalacqua KD, Mirabelli C, Lu
J, Young VL, Hosmillo M, Quaye O, Goodfellow I, Ward VK and Wobus
CE: Akt plays differential roles during the life cycles of acute
and persistent murine norovirus strains in macrophages. J Virol.
96(e0192321)2022.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Amin RM, Hiroshima K, Miyagi Y, Kokubo T,
Hoshi K, Fujisawa T and Nakatani Y: Role of the PI3K/Akt, mTOR, and
STK11/LKB1 pathways in the tumorigenesis of sclerosing hemangioma
of the lung. Pathol Int. 58:38–44. 2008.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Ou JM, Qui MK, Dai YX, Dong Q, Shen J,
Dong P, Wang XF, Liu YB and Fei ZW: Combined blockade of AKT/mTOR
pathway inhibits growth of human hemangioma via downregulation of
proliferating cell nuclear antigen. Int J Immunopathol Pharmacol.
25:945–953. 2012.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Yang X, Li X, Lin Q and Xu Q:
Up-regulation of microRNA-203 inhibits myocardial fibrosis and
oxidative stress in mice with diabetic cardiomyopathy through the
inhibition of PI3K/Akt signaling pathway via PIK3CA. Gene.
715(143995)2019.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Czechowicz JA, Benjamin T, Bly RA, Ganti
SN, Balkin DM, Perkins JA, Frieden IJ and Rosbe KW: Airway
hemangiomas in PHACE syndrome: A multicenter experience.
Otolaryngol Head Neck Surg. 165:182–186. 2021.PubMed/NCBI View Article : Google Scholar
|