Introduction
Diabetic nephropathy (DN) is a serious complication
of diabetes mellitus (DM) clinically characterized as increased
urinary albumin excretions, reduced glomerular filtration rate and
progressive deteriorations in renal function (1). It is reported that approximately
20-40% of patients with diabetes develop DN in their disease course
(2,3). DN is the leading cause of chronic
kidney disease (CKD) and end-stage renal disease (ESRD), and is
acknowledged as an independent risk factor for cardiovascular
diseases, seriously threatening human health (4,5).
Currently, strict glycemic, blood pressure, and lipid control are
the main managements of DN (1);
however, ~40% of diabetic patients progress to ERSD, eventually
requiring dialysis, which imposes heavy health care costs (6). Given the increased prevalence of the
DN, seeking novel biomarkers for the early detection of DN and
elucidating the pathogenesis of DN are essential in order to
provide appropriate treatments to prevent this disease.
The 5-lipoxygenase gene (ALOX5), a member of the
ALOX family responsible for the oxidative metabolism of
polyunsaturated fatty acids, is involved in arachidonate
metabolism, a metabolic pathway in which cysteinyl leukotrienes are
biosynthetically initiated (7,8).
ALOX5 has been reported to be closely associated with multiple
physiological and pathological processes, such as inflammation,
oxidative stress, and cell differentiation, thus influencing
various diseases, including cancers, myocardial infarction, and
bone diseases (9-11).
Interestingly, ALOX5 was generally reported to be aberrantly
upregulated in patients suffering from different diseases, such as
esophageal adenocarcinoma and gastric cancer (12,13).
Notably, ALOX5 was also reported to be aberrantly upregulated in
type 1 diabetes (T1D) and type 2 diabetes (T2D). Increased
expression of ALOX5 was linked to the tissue inflammation in
diabetes, and the inhibitor of ALOX5 was suggested as a candidate
for DM (14-16).
In addition, ALOX5 appears to be involved in the progression of
diabetic retinal disease, emphasizing the importance of ALOX5 in
DM-related diseases (17). In
addition, ALOX5-deficient mice with diabetic retinopathy exhibited
less leukostasis, superoxide production, and nuclear factor (NF)-κB
expression, suggesting that the inflammation in diabetic
retinopathy was ALOX5-dependent (18). Moreover, ALOX5 products have been
demonstrated to be involved in renal tubulointerstitial injury,
indirectly indicating that ALOX5 is associated with renal tissue
injury (19). Furthermore, a
previous study demonstrated that although ALOX5 is usually located
in leukocytes, dendritic cells, macrophages and neutrophils, it can
also be found in areas of the kidney, including the cortex, outer
medulla, and inner medulla (20).
Collectively, ALOX5 appears to participate into multiple processes
of DM-related diseases and renal diseases, as DN is one of the
complications of DM, it is hypothesized that ALOX5 may be involved
in the progression of DN.
Therefore, the present study aimed to investigate
the role of ALOX5 in the progression of DN, and to explore the
underlying mechanism of action, addressing the therapeutic
potential of ALOX5 in DN. To better carry the exploration, an in
vitro cell model of DN was first constructed. HG-induced SV40
MES-13 has been widely recognized as an in vitro cell model
of DN, and has been widely applied in numerous studies (21-24).
Accordingly, SV40 MES-13 cells were treated with HG to simulate DN
in vitro in the present study.
Materials and methods
Cell culture and treatment
Mouse renal mesangial cells (SV40 MES-13) were
obtained from American Type Culture Collection and cultured in
Dulbecco's modified Eagle's medium (DMEM; Thermo Fisher Scientific,
Inc.) supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc.) and 1% streptomycin/penicillin (HyClone;
Cytiva) in a 5% CO2 incubator under 37˚C. Cells were
classified into normal glucose (NG; 5.6 mM glucose) and high
glucose (HG; 30 mM) groups according to the treatment of various
concentrations of glucose for 24 h.
Cell transfection
siRNAs against ALOX5 (si-ALOX5-1,
5'-GCCGGACUGAUGUACCUGUUUTT-3'; si-ALOX5-2,
5'-CCUGUUCAUCAACCGCUUCAUTT-3') and siRNA negative control
(si-control, 5'-UUCUCCGAACGUGUCACGUTT-3') were obtained from
Shanghai GenePharma Co., Ltd. SV40 MES-13 cells were transfected
with si-control (50 nM) or si-ALOX5 (50 nM) using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37˚C. At 48 h post transfection, the
transfected cells were harvested for the further experiments.
Cell viability assay
Transfected or non-transfected cells were seeded
into 96-well plates (3x103 cells/well) and then
incubated with different doses of glucose. After 24 and 48 h of
incubation, 10 µl Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc.) solution was added to each well for another
incubation at 37˚C for 1 h in an incubator with 5% CO2.
Finally, the OD value of each group was determined at an absorbance
of 450 nm using a microplate reader (Bio-Rad Laboratories,
Inc.).
Immunofluorescence
SV40 MES-13 cells were seeded into 6-well plates
(5x104 cells/well) and cultured for 24 h. Then, cells
were washed with PBS and fixed with 4% paraformaldehyde for 30 min
at room temperature and permeabilized using 0.5% Triton X-100.
Subsequently, the cells were blocked in 2.5% bovine serum albumin
(BSA) at room temperature for 1 h and incubated with anti-Ki67
(1:500; cat. no. orb389335; Biorbyt Ltd.) or anti- proliferating
cell nuclear antigen (PCNA; 1:50; cat. no. orb48485; Biorbyt Ltd.)
antibodies at 4˚C overnight. The following day, the cells were
incubated with the Alexa Fluor 488-flagged goat anti-rabbit
secondary antibody (1:500; cat. no. A0423; Beyotime Institute of
Biotechnology). Subsequently, the cells were incubated with DAPI
for 3 min, and the fluorescence signals were captured using a DMRA2
fluorescence microscope (Leica Microsystems GmbH).
Flow cytometric analysis
Cell cycle and cell apoptosis were evaluated using a
flow cytometer. Briefly, SV40 MES-13 cells were harvested and
washed using ice-cold PBS twice. For the cell cycle assay, cells
were fixed with 70% ice-cold ethanol overnight. The cells were then
incubated with 50 µg/ml RNase at 37˚C for 30 min, followed by
staining with 50 µg/ml propidium iodide (PI) at 4˚C for 30 min in
the dark. For cell apoptosis assay, cells were resuspended with 500
µl binding buffer, followed by staining with 5 µl Annexin V-FITC
and 5 µl PI at room temperature for 15 min in the dark. Finally,
the flow cytometric analysis was performed by a FACScan (Beckman
Coulter, Inc.) and the data were analyzed using FlowJo software
version 10.6.2 (Tree Star, Inc.).
Enzyme-linked immunosorbent assay
(ELISA)
The concentrations of inflammatory cytokines in the
culture medium, including TNF-α, IL-6 and IL-1β, were detected
using their corresponding ELISA kits: Mouse TNF-α ELISA kit (cat.
no. orb565170), mouse IL-6 ELISA kit (cat. no. orb565132) or mouse
IL-8 ELISA kit (cat. no. orb566249) from Biorbyt Ltd., according to
the manufacturer's instructions.
Reverse transcription-quantitative PCR
(RT-qPCR)
The total RNA was extracted from cells using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. The purity of RNA was measured
using NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific,
Inc.). First-strand complementary DNA (cDNA) synthesis was then
performed using SuperScript IV One-Step RT-PCR system (Invitrogen;
Thermo Fisher Scientific, Inc.). RT-qPCR was conducted using the
SYBR-Green PCR Master Mix (Applied Biosystems; Thermo Fisher
Scientific, Inc.) on ABI 7500 Real-time PCR system (Bio-Rad
Laboratories, Inc.). The primer sequences used were listed as
follows: ALOX5 forward, 5'-CCTCAGCCTCATTGGCTCTG-3' and reverse,
5'-TAGGAGTCCACCGCGCC-3'; GAPDH forward, 5'-CAGGAGAGTGTTTCCTCGTCC-3'
and reverse, 5'-TTTGCCGTGAGTGGAGTCAT-3'. The mRNA expression was
normalized to GAPDH and quantified using the comparative
quantification method (2-ΔΔCq) (25).
Western blotting
The cells were harvested using RIPA lysis buffer
(Beyotime Institute of Biotechnology) to extract total protein. A
bicinchoninic acid (BCA) kit (Sigma-Aldrich; Merck KGaA) was
applied to measure the protein concentration according to the
manufacturer's instructions. The same amount of protein (30
µg/lane) was electrophoresed on polyacrylamide gels and transferred
to polyvinylidene fluoride (PVDF) membranes (Millipore; Merck
KGaA). After blocking with 5% skimmed milk for 2 h at room
temperature, the membranes were incubated with primary antibodies
against ALOX5 (1:1,000; cat. no. orb536948), Bcl-2 (1:1,000; cat.
no. orb10173), Bax (1:2,000; cat. no. orb31066), and cleaved
caspase-3 (1:1,000, cat. no. orb126608; all Biorbyt Ltd.),
phosphorylated NF-κB inhibitor α (p-IκBα) (1:1,000; product no.
2859), IκBα (1:1,000; product no. 4812), p-p65 (1:1,000; product
no. 3033), p65 (1:1,000; product no. 8242; all from Cell Signaling
Technology, Inc.), collagen type IV (COL4; 1:1,000; cat. no.
SAB4200500; Sigma-Aldrich; Merck KGaA), fibronectin (FN; 1:1,000;
cat. no. 15613-1-AP; ProteinTech, Inc.), transforming growth factor
(TGF)-β1 (1:1,000; cat. no. 21898-1-AP; ProteinTech, Inc.), and
GAPDH (1:1,000; product no. 2118; Cell Signaling Technology, Inc.)
at 4˚C overnight. The following day, the membranes were incubated
with the horseradish peroxidase-conjugated goat anti-rabbit IgG
secondary antibody (1:2,000; product code ab6721; Abcam) for 1 h at
room temperature. The protein bands were visualized by enhanced
chemiluminescence system (ECL; Thermo Fisher Scientific., Inc.) and
quantified using ImageJ software version 1.46 (National Institutes
of Health). Since GAPDH expression was not changed in HG-induced
SV40 MES-13 cells in previous studies (22,24),
GAPDH was used as the internal control.
Statistical analysis
All data were analyzed using GraphPad Prism version
6 (GraphPad Software, Inc.) and presented as the mean ± standard
deviation (SD) from at least three independent experiments.
Comparisons between different groups were carried out using
Student's unpaired t-test or one-way AVOVA followed by Tukey's post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
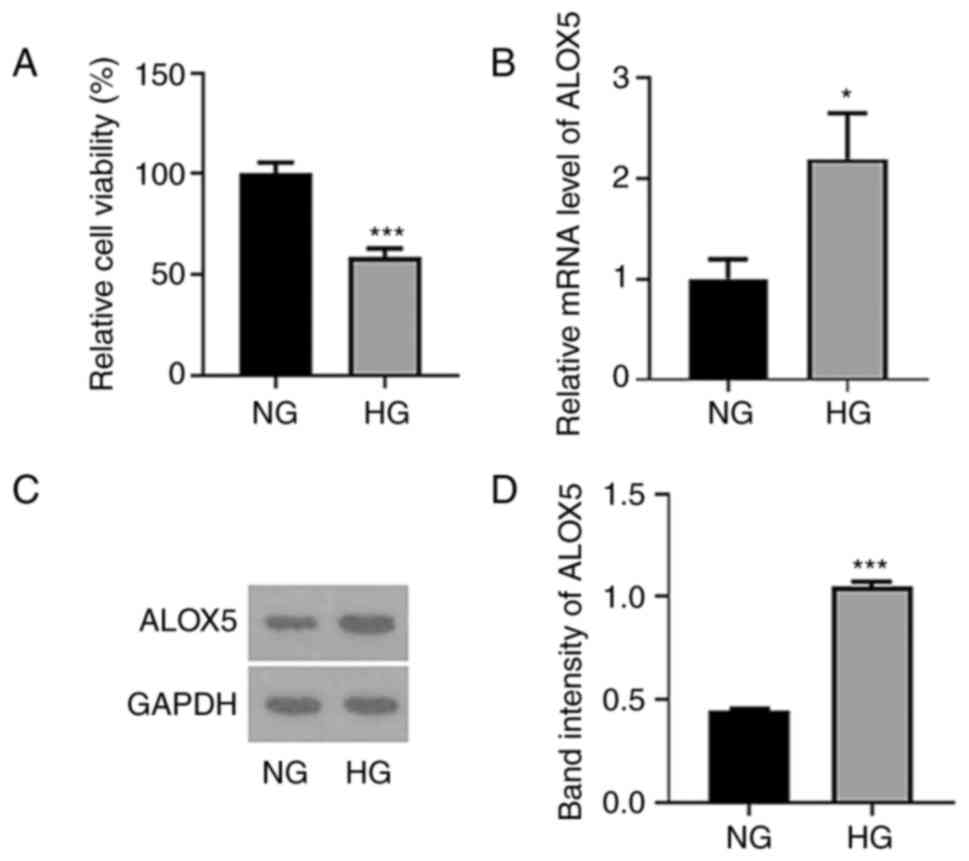
ALOX5 is upregulated in HG-induced
SV40 MES-13 cells
Firstly, SV40 MES-13 cells were treated with HG (30
mM glucose) for 24 h to simulate DN in vitro, and cells
treated with NG (5.6 mM glucose) were regarded as the control. As
revealed in Fig. 1A, the cell
viability was significantly reduced following HG treatment. In
addition, both the mRNA level and protein expression of ALOX5 were
significantly upregulated upon HG treatment (Fig. 1B-D), suggesting that HG induced a
high level of ALOX5 in SV40 MES-13 cells.
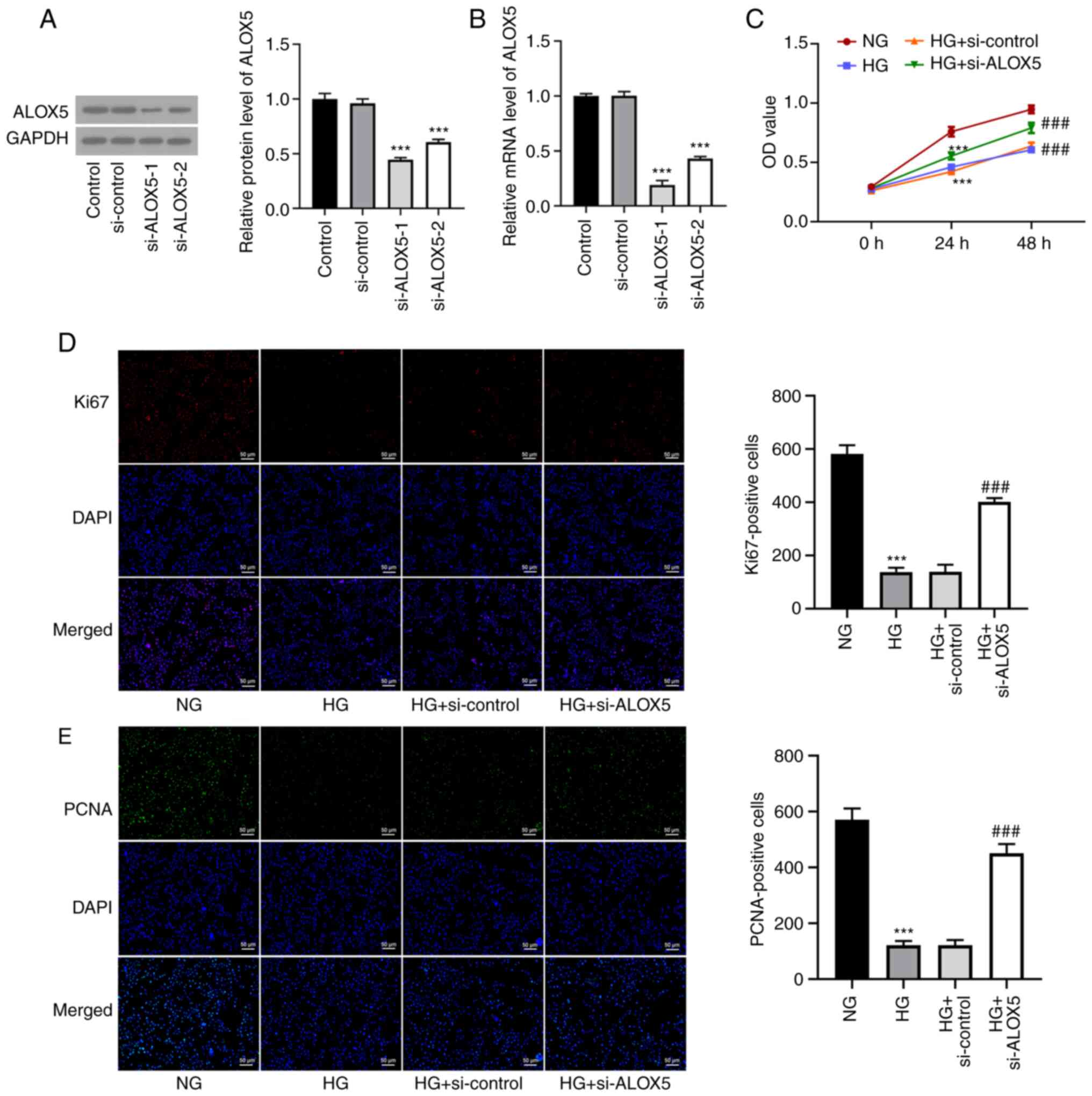
Silencing of ALOX5 promotes cell
proliferation in HG-induced SV40 MES-13 cells
To investigate the role of ALOX5 in DN, SV40 MES-13
cells were transfected with si-ALOX5-1 or si-ALOX5-2 to silence
ALOX5. As revealed in Fig. 2A and
B, compared with si-control, both
si-ALOX5-1 and si-ALOX5-2 significantly reduced the mRNA level and
protein expression of ALOX5. Since the si-ALOX5-1 group was more
efficient than the si-ALOX5-2 group in silencing ALOX5, it was used
for subsequent experiments. A series of cellular experiments was
then performed. As revealed in Fig.
2C, it was observed that at both 24 and 48 h post HG
stimulation, the decreased cell viability under HG stimulation was
significantly improved by ALOX5 silencing. Generally, the induction
time is dependent on the induction efficacy and the research goal.
In this study, upon 24 h of an induction of HG, a reduced cell
viability was not only obtained, compared with NG, but a
significantly upregulated expression level of ALOX5 was also found,
confirming a high level of ALOX5 in HG-induced SV40 MES-13 cells,
which was suitable for the following investigation on the specific
role of ALOX5 upon HG stimulation. Thus, 24 h was the time-point
used for investigation in this study. Subsequently, the expression
levels of Ki67 and PCNA, two classical hallmarks of cell
proliferation (26), were markedly
decreased by HG treatment for 24 h, which were partly abolished by
interference of ALOX5 (Fig. 2D and
E). These results indicated that
interference of ALOX5 improved cell proliferation ability of SV40
MES-13 cells under HG stimulation.
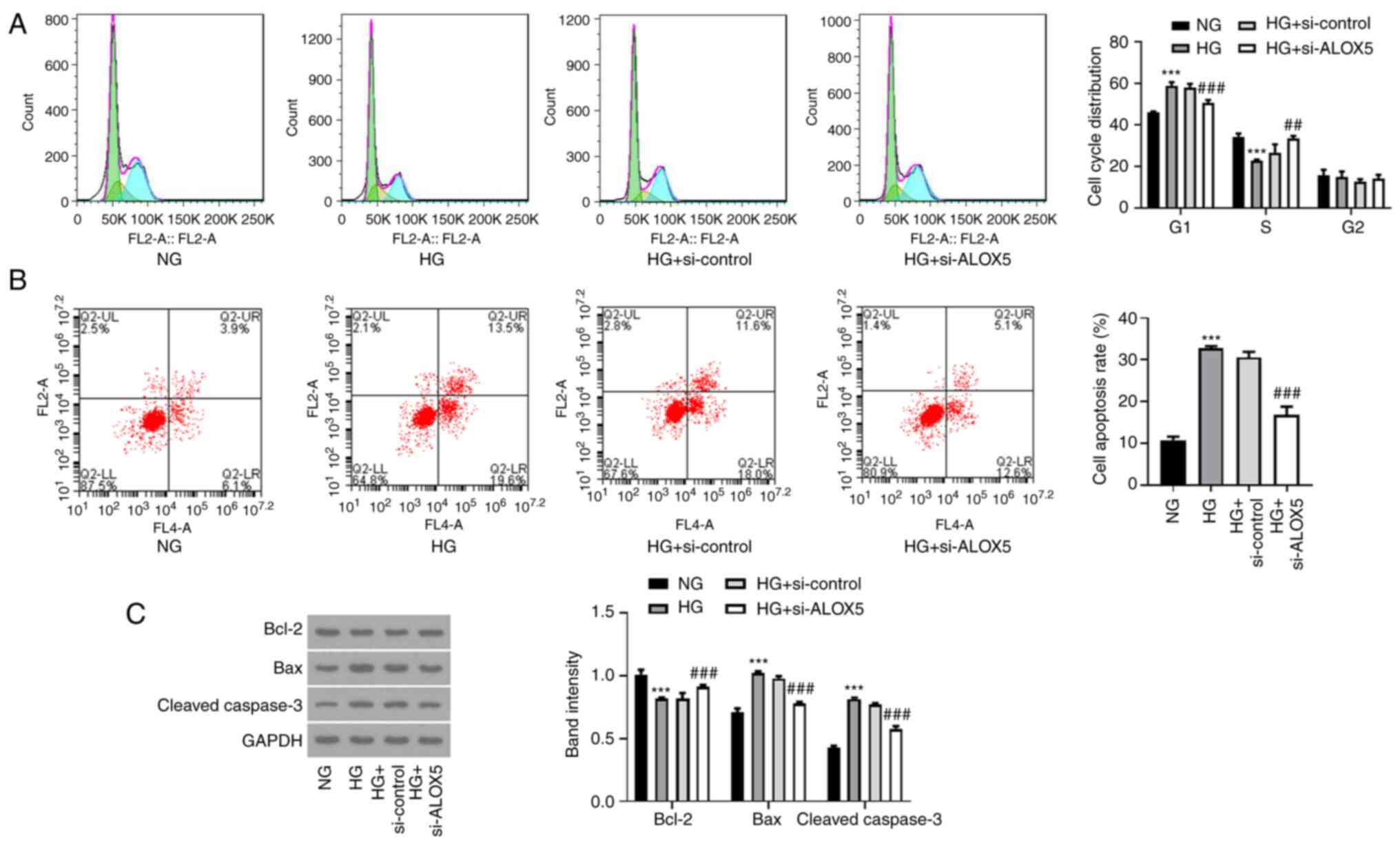
Silencing of ALOX5 promotes cell cycle
progression and suppresses cell apoptosis in HG-induced SV40 MES-13
cells
Subsequently, a flow cytometric assay was conducted
to analyze the effects of ALOX5 on cell cycle progression and cell
apoptosis. As revealed in Fig. 3A,
after treatment with HG, cells were arrested in the G1 phase, and
the cell proportion in the S phase was reduced, reflecting that HG
treatment blocked cell cycle progression. However, interference of
ALOX5 reduced the cell proportion in the G1 phase and increased
that in the S phase, indicating that interference of ALOX5 promoted
the progression of the cell cycle. In addition, HG stimulation
significantly increased the cell apoptosis rate of SV40 MES-13
cells, which was partly abolished by interference of ALOX5
(Fig. 3B). In addition, the
reduced protein expression of Bcl-2 and the elevated protein
expression of Bax and cleaved caspase-3 after HG induction were
also partly hindered by silencing of ALOX5 (Fig. 3C). These results suggested that
HG-induced blockage of cell cycle progression and a high rate of
cell apoptosis could be partly attenuated by interference of
ALOX5.
Silencing of ALOX5 alleviates
inflammatory response and fibrosis in HG-induced SV40 MES-13
cells
Next, a series of inflammatory cytokines and
fibrosis-related proteins were assessed to detect the effects of
ALOX5 on inflammation and fibrosis in a cell DN model in
vitro. As shown in Fig. 4A-C,
the upregulated levels of TNF-α, IL-6 and IL-8 upon HG stimulation
were significantly suppressed when ALOX5 was silenced, indicating
that silencing of ALOX5 markedly inhibited inflammatory response in
HG-induced SV40 MES-13 cells. In addition, the upregulated protein
expression levels of COL4, FN, and TGF-β1 upon HG stimulation were
markedly inhibited by silencing of ALOX5 (Fig. 4D), suggesting that interference of
ALOX5 attenuated HG-induced fibrosis.
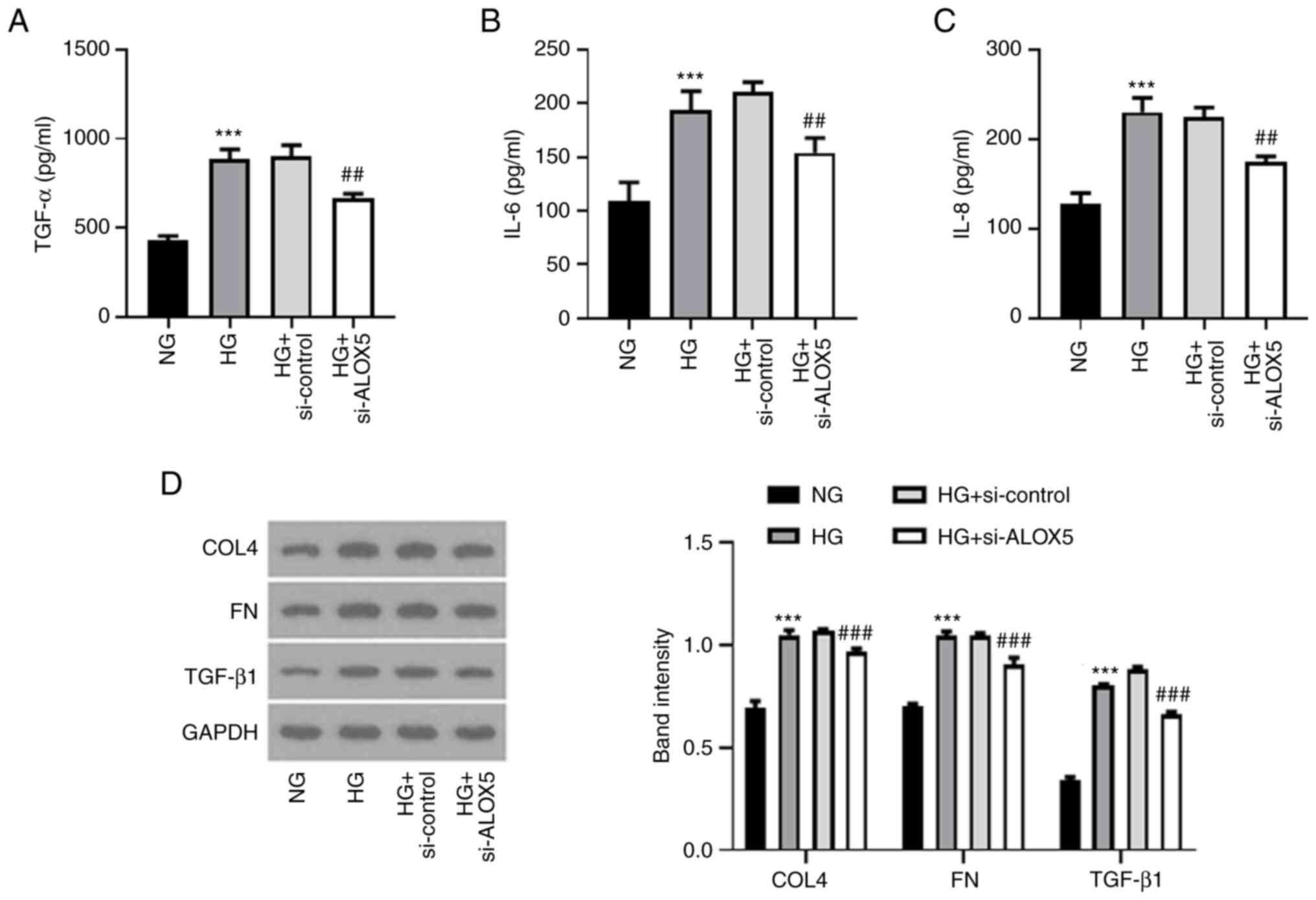 | Figure 4Silencing of ALOX5 alleviates
inflammatory response and fibrosis in HG-induced SV40 MES-13 cells.
(A-C) The HG-induced SV40 MES-13 cells were transfected with
si-ALOX5 or si-control, and the inflammatory cytokines, including
TNF-α, IL-6 and IL-8 were evaluated using their corresponding ELISA
kits. (D) The expression level of fibrosis-related proteins was
assessed using western blotting. ***P<0.001 vs. NG;
##P<0.01 and ###P<0.001 vs. HG +
si-control. ALOX5, 5-lipoxygenase; HG, high glucose; si-, siRNA;
ELISA, enzyme-linked immunosorbent; NG, normal glucose; COL4,
collagen type IV; FN, fibronectin; TGF-β1, transforming growth
factor β1. |
Silencing of ALOX5 weakens NF-κB
signaling pathway in HG-induced SV40 MES-13 cells
Finally, NF-κB, as an important inflammatory
stimulus for DN (27,28), was also assessed in the present
study. As revealed in Fig. 5, HG
stimulation enhanced the expression levels of p-IκBα and p-p65,
indicating that NF-κB signaling was activated upon HG stimulation,
whereas silencing of ALOX5 reduced the expression levels of p-IκBα
and p-p65. These results indicated that ALOX5 regulated NF-κB
activation.
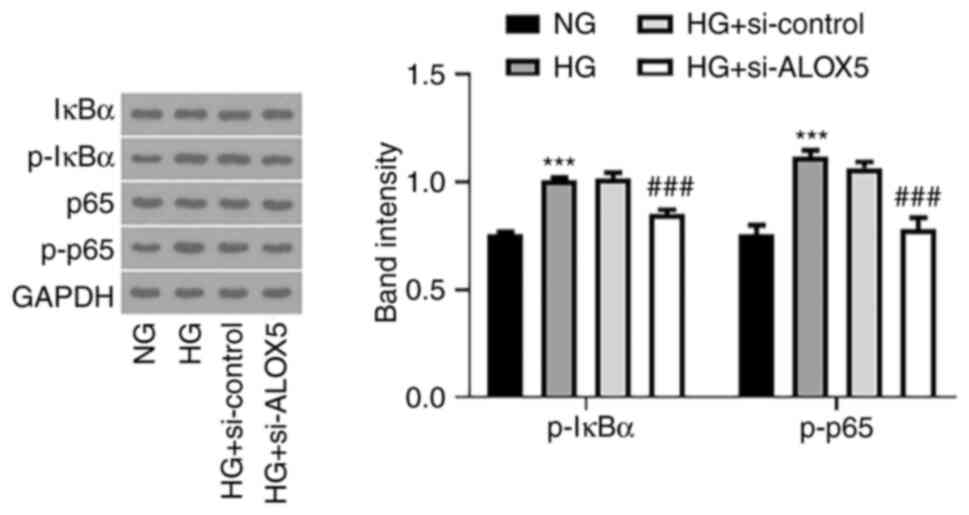 | Figure 5Silencing of ALOX5 weakens the NF-κB
signaling pathway in HG-induced SV40 MES-13 cells. The HG-induced
SV40 MES-13 cells were transfected with si-ALOX5 or si-control, and
the protein expression of p-IκBα, IκBα, p-p65, and p65 was measured
using western blotting. ***P<0.001 vs. NG;
###P<0.001 vs. HG + si-control. ALOX5,
5-lipoxygenase; HG, high glucose; si-, siRNA; p-, phosphorylated;
IκBα, NF-κB inhibitor α; NG, normal glucose. |
Discussion
DN is one of the most challenging kidney diseases.
This disease leads to heavy financial and health-care burdens
globally (29), therefore, it is
evident that further understanding of the pathogenic mechanisms
underlying DN is urgently required to prevent disease progression.
Firstly, to better carry out our investigation, an in vitro
cell model of DN was firstly constructed. HG-induced SV40 MES-13
has been widely recognized as an in vitro cell model of DN,
and has been widely applied in numerous studies (21-24).
In addition, mannitol is usually used as a negative control to
eliminate the effects of osmotic pressure caused by HG; however, in
a previous study mannitol did not produce any change in comparison
with the control, indicating that osmotic pressure is not a
variable factor between NG and HG (24), thus numerous studies only set NG
and HG, without mannitol as a negative control, for the
investigation in HG-induced kidney injury (21-23).
Accordingly, the present study used NG and HG groups for the
investigation. In the present study, it was observed that ALOX5 was
aberrantly upregulated by HG stimulation in SV40 MES-13 cells. A
series of cellular biological experiments revealed that silencing
of ALOX5 could efficiently attenuate HG-induced renal cell injuries
by mitigating cell apoptosis and cell growth restriction,
antagonizing inflammatory response and fibrosis in SV40 MES-13
cells. Mechanistically, interference of ALOX5 inhibited NF-κB
signaling, which may account for the regulatory function of ALOX5
in HG-induced SV40 MES-13 cells. These results suggested that ALOX5
may serve as a promising candidate for novel therapeutic strategies
of DN treatment. Furthermore, silencing of ALOX5 exerted a
protective effect against HG-induced injuries in SV40 MES-13 cells,
raising the possibility that ALOX5 inhibitors or ALOX5-targeting
drugs may be alternative strategies for the clinical treatment of
DN.
DN is a multifactorial disease involving a variety
of pathogenic molecular processes and histopathological structure.
Fibrosis and inflammation are important pathologic characteristics
of DN. The production of fibrotic and pro-inflammatory cytokines
can directly damage the kidney structure and promote the deposition
of extracellular matrix components (EMC), thereby contributing to
the onset and progression of DN (30,31).
In addition, apoptosis is a strictly controlled cell death process,
which is involved in cell growth in multiple diseases. It was
reported by Zheng et al that HG treatment promoted the
apoptosis of mouse β-TC-tet cells, and that cell apoptosis was up
to 30% (32). In addition, another
study revealed that the apoptosis rate of RSC96 Schwann cells was
increased to ~40% following HG treatment for 48 h (33). However, according to different
handling factors such as experiment personnel, there may exist some
differences in the actual data between different studies. In the
present study, promotion of apoptosis (~30%) was demonstrated
following HG treatment for 24 h in SV40 MES-13 cells, indicating
that HG caused severe injury in renal cells. The accumulating
evidence confirms that renal cell apoptosis is also a classical
hallmark of DN, whereby apoptotic cells may be partly attributed to
renal inflammation (34,35). Qi et al determined that
NORAD could aggravate the progression of DN by promoting the
proliferative ability and inhibiting the apoptosis of glomerular
mesangial cells (21). Ma et
al reported that downregulation of lncRNA NEAT1 inhibited cell
proliferation, fibrosis, and inflammation but promoted cell
apoptosis in a DN cellular model, shedding light on the application
of lncRNA NEAT1 downregulation in the treatment of DN (36). It was also demonstrated by Liu
et al that praliciguat inhibited the progression of DN
partly by suppressing inflammation and apoptosis (37). Consistently, in the present study,
it was determined that silencing of ALOX5 not only exerted
anti-inflammatory and anti-fibrotic effects, but also exerted
anti-apoptotic effects, accompanied with the promotive effects on
cell viability, Ki67-positive and PCNA-positive cells, and cell
cycle progression. These results provided ample evidence that
silencing of ALOX5 could retain anti-fibrosis, anti-inflammation
and anti-apoptosis, thereby suppressing the progression of DN,
revealing the potential application of ALOX5 interference for the
treatment of DN.
Furthermore, NF-κB signaling has been demonstrated
to be a key inflammatory pathway in the pathogenesis of DN
inflammation and fibrosis in both clinical and animal studies
(28,38). Under normal conditions, NF-κB binds
to anchor protein IκBα, forming an inactive complex in the
cytoplasm, which suppresses the activation of NF-κB signaling
(39). In a diabetic state, IκBα
can be phosphorylated in its proteasomal degradation by the IKK
complex, leading to the activation and phosphorylation of NF-κB
subunit p65, which is then translocated towards the nucleus and
triggers the overproduction of pro-inflammatory cytokines,
including TNF-α and IL-6. Particularly, the hyperphosphorylation of
IκBα and p65 have been observed in DN in vivo or in
vitro, confirming that DN was dependent on NF-κB activation,
and limiting the activation of NF-κB has been widely demonstrated
to protect the rats against DN (27,40-42).
Consistently, the present study also demonstrated the
hyperphosphorylation of IκBα and p65, as well as a severe
inflammatory response, in HG-induced SV40 MES-13 cells. In
addition, it has been reported that ALOX5 and its metabolite LTB4
are capable of activating NF-κB in cancer cells, indicating a close
association between ALOX5 and NF-κB and a potential regulatory
effect of ALOX5 on NF-κB signaling (43,44).
In the present study, it was further demonstrated that silencing of
ALOX5 downregulated p-p65 and p-IκBα in HG-induced SV40 MES-13
cells, revealing that interference of ALOX5 could regulate the
functional NF-κB subunit to block HG-induced activation of NF-κB
signaling in a DN cell model in vitro.
Although the role of ALOX5 in HG-induced SV40 MES-13
cells was investigated in the present study, thus far, these
findings have not been verified in vivo or clinically. In
addition, whether glomerular mesangial cells express ALOX5 under
normal or disease conditions in vivo should be verified in
future studies. Furthermore, despite the fact that silencing of
ALOX5 was determined to suppress NF-κB signaling in HG-induced SV40
MES-13 cells, whether the effect of ALOX5 is NF-κB-dependent
remains to be investigated. Moreover, all of the findings
concerning the cellular biological activities in the present study
were based on transfection with si-ALOX5-1, therefore, a second
siRNA may be beneficial to verify the absence of off-target
effects. In addition, the involvement of other signaling pathways
underlying the regulatory role of ALOX5 in HG-induced renal injury
cannot be excluded. Further studies are required to reveal the
functions and precise mechanisms of ALOX5 in DN in vivo and
in vitro.
In conclusion, it was demonstrated that ALOX5 was
aberrantly upregulated in HG-induced renal cell injury, and
silencing of ALOX5 ameliorated DN in vitro by attenuating
the inflammatory response, fibrosis and cell apoptosis. Mechanistic
analysis revealed the importance of NF-κB signaling underlying the
regulation of ALOX5 in DN. The present study contributed to an
improved understanding of the mechanism underlying ALOX5 involved
in DN. ALOX5 may be considered as an attractive therapeutic target
to prevent the development of DN.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
All data generated and/or analyzed during the
present study are included in this published article.
Authors' contributions
XC designed the study. XC, HX, YL, QO and SD
conducted the experiments and analyzed the data. HX drafted the
manuscript and XC revised the manuscript. All authors read and
approved the final manuscript.. XC and HX confirm the authenticity
of all the raw data and are agreed to be accountable for all
aspects of the work.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sagoo MK and Gnudi L: Diabetic
nephropathy: An overview. Methods Mol Biol. 2067:3–7.
2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Gheith O, Farouk N, Nampoory N, Halim MA
and Al-Otaibi T: Diabetic kidney disease: World wide difference of
prevalence and risk factors. J Nephropharmacol. 5:49–56.
2016.PubMed/NCBI
|
|
3
|
Narres M, Claessen H, Droste S, Kvitkina
T, Koch M, Kuss O and Icks A: The incidence of end-stage renal
disease in the diabetic (compared to the non-diabetic) population:
A systematic review. PLoS One. 11(e0147329)2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Si X, Li P, Zhang Y, Zhang Y, Lv W and Qi
D: Renoprotective effects of olmesartan medoxomil on diabetic
nephropathy in streptozotocin-induced diabetes in rats. Biomed Rep.
2:24–28. 2014.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Shi Y and Hu FB: The global implications
of diabetes and cancer. Lancet. 383:1947–1948. 2014.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Weir MA and Herzog CA: Beta blockers in
patients with end-stage renal disease-evidence-based
recommendations. Semin Dial. 31:219–225. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Dobrian AD, Lieb DC, Cole BK,
Taylor-Fishwick DA, Chakrabarti SK and Nadler JL: Functional and
pathological roles of the 12- and 15-lipoxygenases. Prog Lipid Res.
50:115–131. 2011.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Chen F, Ghosh A, Lin J, Zhang C, Pan Y,
Thakur A, Singh K, Hong H and Tang S: 5-lipoxygenase pathway and
its downstream cysteinyl leukotrienes as potential therapeutic
targets for Alzheimer's disease. Brain Behav Immun. 88:844–855.
2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Wang Y, Skibbe JR, Hu C, Dong L, Ferchen
K, Su R, Li C, Huang H, Weng H, Huang H, et al: ALOX5 exhibits
anti-tumor and drug-sensitizing effects in MLL-rearranged leukemia.
Sci Rep. 7(1853)2017.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Lisovyy OO, Dosenko VE, Nagibin VS,
Tumanovska LV, Korol MO, Surova OV and Moibenko OO:
Cardioprotective effect of 5-lipoxygenase gene (ALOX5) silencing in
ischemia-reperfusion. Acta Biochim Pol. 56:687–694. 2009.PubMed/NCBI
|
|
11
|
Wu Y, Sun H, Song F, Huang C and Wang J:
Deletion of Alox5 gene decreases osteogenic differentiation but
increases adipogenic differentiation of mouse induced pluripotent
stem cells. Cell Tissue Res. 358:135–147. 2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zhu L, Yang F, Wang L, Dong L, Huang Z,
Wang G, Chen G and Li Q: Identification the ferroptosis-related
gene signature in patients with esophageal adenocarcinoma. Cancer
Cell Int. 21(124)2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Tang J, Zhang C, Lin J, Duan P, Long J and
Zhu H: ALOX5-5-HETE promotes gastric cancer growth and alleviates
chemotherapy toxicity via MEK/ERK activation. Cancer Med.
10:5246–5255. 2021.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Heemskerk MM, Giera M, Bouazzaoui FE, Lips
MA, Pijl H, van Dijk KW and van Harmelen V: Increased PUFA content
and 5-Lipoxygenase pathway expression are associated with
subcutaneous adipose tissue inflammation in obese women with type 2
diabetes. Nutrients. 7:7676–7690. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
ul Ain Q, Greig NH, Nawaz MS, Rashid S and
Kamal MA: Exploring N(1)-p-fluorobenzyl-cymserine as an inhibitor
of 5-lipoxygenase as a candidate for type 2 diabetes and
neurodegenerative disorder treatment. CNS Neurol Disord Drug
Targets. 13:197–202. 2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ramalho T, Filgueiras L, Silva-Jr IA,
Pessoa AFM and Jancar S: Impaired wound healing in type 1 diabetes
is dependent on 5-lipoxygenase products. Sci Rep.
8(14164)2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Schwartzman ML, Iserovich P, Gotlinger K,
Bellner L, Dunn MW, Sartore M, Grazia Pertile M, Leonardi A, Sathe
S, Beaton A, et al: Profile of lipid and protein autacoids in
diabetic vitreous correlates with the progression of diabetic
retinopathy. Diabetes. 59:1780–1788. 2010.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Gubitosi-Klug RA, Talahalli R, Du Y,
Nadler JL and Kern TS: 5-Lipoxygenase, but not 12/15-lipoxygenase,
contributes to degeneration of retinal capillaries in a mouse model
of diabetic retinopathy. Diabetes. 57:1387–1393. 2008.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Landgraf SS, Silva LS, Peruchetti DB,
Sirtoli GM, Moraes-Santos F, Portella VG, Silva-Filho JL, Pinheiro
CS, Abreu TP, Takiya CM, et al: 5-Lypoxygenase products are
involved in renal tubulointerstitial injury induced by albumin
overload in proximal tubules in mice. PLoS One.
9(e107549)2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Reinhold SW, Vitzthum H, Filbeck T, Wolf
K, Lattas C, Riegger GA, Kurtz A and Krämer BK: Gene expression of
5-, 12-, and 15-lipoxygenases and leukotriene receptors along the
rat nephron. Am J Physiol Renal Physiol. 290:F864–F872.
2006.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Qi H, Yao L and Liu Q: NORAD affects the
progression of diabetic nephropathy through targeting miR-520h to
upregulate TLR4. Biochem Biophys Res Commun. 521:190–195.
2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Du Y, Yang YT, Tang G, Jia JS, Zhu N and
Yuan WJ: Butyrate alleviates diabetic kidney disease by mediating
the miR-7a-5p/P311/TGF-beta1 pathway. FASEB J. 34:10462–10475.
2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Yang X, Luo W, Li L, Hu X, Xu M, Wang Y,
Feng J, Qian J, Guan X, Zhao Y and Liang G: CDK9 inhibition
improves diabetic nephropathy by reducing inflammation in the
kidneys. Toxicol Appl Pharmacol. 416(115465)2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Xu J, Xiang P, Liu L, Sun J and Ye S:
Metformin inhibits extracellular matrix accumulation, inflammation
and proliferation of mesangial cells in diabetic nephropathy by
regulating H19/miR-143-3p/TGF-β1 axis. J Pharm Pharmacol.
72:1101–1109. 2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Jurikova M, Danihel L, Polak S and Varga
I: Ki67, PCNA, and MCM proteins: Markers of proliferation in the
diagnosis of breast cancer. Acta Histochem. 118:544–552.
2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Liu T, Zhang L, Joo D and Sun SC: NF-κB
signaling in inflammation. Signal Transduct Target Ther.
2(17023)2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Donate-Correa J, Martín-Núñez E,
Muros-de-Fuentes M, Mora-Fernández C and Navarro-González JF:
Inflammatory cytokines in diabetic nephropathy. J Diabetes Res.
2015(948417)2015.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Samsu N: Diabetic nephropathy: Challenges
in pathogenesis, diagnosis, and treatment. Biomed Res Int.
2021(1497449)2021.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Moreno JA, Gomez-Guerrero C, Mas S, Sanz
AB, Lorenzo O, Ruiz-Ortega M, Opazo L, Mezzano S and Egido J:
Targeting inflammation in diabetic nephropathy: A tale of hope.
Expert Opin Investig Drugs. 27:917–930. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Chow FY, Nikolic-Paterson DJ, Ozols E,
Atkins RC and Tesch GH: Intercellular adhesion molecule-1
deficiency is protective against nephropathy in type 2 diabetic
db/db mice. J Am Soc Nephrol. 16:1711–1722. 2005.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Zheng H, Li X, Yang X, Yan F, Wang C and
Liu J: miR-217/Mafb axis involve in high glucose-induced β-TC-tet
cell damage via regulating NF-κB signaling pathway. Biochem Genet.
58:901–913. 2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Dong J, Li H, Bai Y and Wu C: Muscone
ameliorates diabetic peripheral neuropathy through activating
AKT/mTOR signalling pathway. J Pharm Pharmacol. 71:1706–1713.
2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Rane MJ, Song Y, Jin S, Barati MT, Wu R,
Kausar H, Tan Y, Wang Y, Zhou G, Klein JB, et al: Interplay between
Akt and p38 MAPK pathways in the regulation of renal tubular cell
apoptosis associated with diabetic nephropathy. Am J Physiol Renal
Physiol. 298:F49–F61. 2010.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Yu Q, Zhang M, Qian L, Wen D and Wu G:
Luteolin attenuates high glucose-induced podocyte injury via
suppressing NLRP3 inflammasome pathway. Life Sci. 225:1–7.
2019.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Ma J, Zhao N, Du L and Wang Y:
Downregulation of lncRNA NEAT1 inhibits mouse mesangial cell
proliferation, fibrosis, and inflammation but promotes apoptosis in
diabetic nephropathy. Int J Clin Exp Pathol. 12:1174–1183.
2019.PubMed/NCBI
|
|
37
|
Liu G, Shea CM, Jones JE, Price GM, Warren
W, Lonie E, Yan S, Currie MG, Profy AT, Masferrer JL and Zimmer DP:
Praliciguat inhibits progression of diabetic nephropathy in ZSF1
rats and suppresses inflammation and apoptosis in human renal
proximal tubular cells. Am J Physiol Renal Physiol. 319:F697–F711.
2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Oguiza A, Recio C, Lazaro I, Mallavia B,
Blanco J, Egido J and Gomez-Guerrero C: Peptide-based inhibition of
IκB kinase/nuclear factor-κB pathway protects against
diabetes-associated nephropathy and atherosclerosis in a mouse
model of type 1 diabetes. Diabetologia. 58:1656–1667.
2015.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Schröfelbauer B, Polley S, Behar M, Ghosh
G and Hoffmann A: NEMO ensures signaling specificity of the
pleiotropic IKKβ by directing its kinase activity toward IκBα. Mol
Cell. 47:111–121. 2012.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Manna K, Mishra S, Saha M, Mahapatra S,
Saha C, Yenge G, Gaikwad N, Pal R, Oulkar D, Banerjee K and Das
Saha K: Amelioration of diabetic nephropathy using pomegranate peel
extract-stabilized gold nanoparticles: Assessment of NF-κB and Nrf2
signaling system. Int J Nanomedicine. 14:1753–1777. 2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Zhang Y, Ren S, Ji Y and Liang Y:
Pterostilbene ameliorates nephropathy injury in
streptozotocin-induced diabetic rats. Pharmacology. 104:71–80.
2019.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Kolati SR, Kasala ER, Bodduluru LN,
Mahareddy JR, Uppulapu SK, Gogoi R, Barua CC and Lahkar M: BAY
11-7082 ameliorates diabetic nephropathy by attenuating
hyperglycemia-mediated oxidative stress and renal inflammation via
NF-κB pathway. Environ Toxicol Pharmacol. 39:690–699.
2015.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Zhao Y, Wang W, Wang Q, Zhang X and Ye L:
Lipid metabolism enzyme 5-LOX and its metabolite LTB4 are capable
of activating transcription factor NF-κB in hepatoma cells. Biochem
Biophys Res Commun. 418:647–651. 2012.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Cheng JH, Zhang WJ, Zhu JF, Cui D, Song
KD, Qiang P, Mei CZ, Nie ZC, Ding BS, Han Z, et al: CaMKIIγ
regulates the viability and self-renewal of acute myeloid leukaemia
stem-like cells by the Alox5/NF-κB pathway. Int J Lab Hematol.
43:699–706. 2021.PubMed/NCBI View Article : Google Scholar
|