Introduction
Gestational diabetes mellitus (GDM) is one of the
most common complications associated with pregnancy, which affects
~14% of pregnancies worldwide (1).
It is a risk for mothers and their children because GDM can cause
either short- or long-term adverse health effects (2). Although several clinical symptomatic
supportive treatment options are available, such as lifestyle
intervention (diet and exercise) and insulin therapy, all have
demonstrated limited efficacy (3).
Therefore, it is necessary to identify novel prognostic markers and
molecular therapeutic targets for improving the diagnosis and
treatment of GDM.
GDM is characterized by a transitory form of
diabetes induced by insulin resistance and pancreatic β-cell
dysfunction during pregnancy (4,5). The
former is a common feature of GDM, where alleviating insulin
resistance using pioglitazone clinically has become an effective
approach for treating this condition (6,7).
Pancreatic β-cells is a unique population of cells that store and
produce insulin in response to metabolic demand, and dysfunction in
this cell type is a central component of the pathogenesis of GDM
(8). When the quantity of insulin
secreted by pancreatic β-cells is insufficient to compensate for
the reduced insulin sensitivity of tissues, glucose homeostasis is
disrupted, resulting in GDM (9).
Responsible for impoverished insulin synthesis and secretion,
increased cell apoptosis and proinflammatory cytokine production in
pancreatic β-cells are frequently reported in GDM (10-13),
which attracts extensive attention to decipher the underlying
molecular mechanisms behind the phenotypes. However, the molecular
effectors that regulate pancreatic β-cell function in spite of
multitudinous efforts especially inflammation and apoptosis, remain
elusive.
MicroRNAs (miRNAs or miRs) is a family of short,
non-coding RNAs that are ~22 nucleotides in length and typically
regulate gene expression by binding to the 3'-untranslated region
(3'-UTR) of target gene mRNAs, which causes translational
repression or degradation (14,15).
Accumulating evidence suggests that miRNAs can exert key roles in
the pathogenesis and development of diabetes, including GDM
(16,17). Several miRNAs have been previously
demonstrated to be differently expressed in GDM, including miR-132,
miR-181a and miR-29c, implicating their potential diagnostic value
in GDM (18,19). Furthermore, miRNAs have been
reported to regulate β-cell development and function. miR-320a was
found to target musculoaponeurotic fibrosarcoma oncogene homolog F
to suppress insulin secretion whilst inducing apoptosis in
pancreatic β cells (20). In
addition, the expression of miR-221/222 was significantly
upregulated in β-cells from the high-fat diet (HFD)-fed mice and
db/db mice, and overexpression of miR-221/222 impaired the insulin
production and secretion of β-cells and resulted in glucose
intolerance in vivo (21). In GDM,
miRNA dysregulation can contribute to the pathogenesis of GDM by
regulating the inflammatory response in pancreatic β cells
(22,23). miR-101a has been demonstrated to
impair the glucose-induced inflammatory response by decreasing the
expression of the transcription factor One Cut homeobox2(24). Another previous study showed that
silencing miRNA-222 suppressed the inflammatory response in GDM
mice by promoting C-X-C motif chemokine 4 receptor expression
(25). Therefore, identifying
additional miRNAs as biomarkers for clinical diagnosis and
pathogenesis would be beneficial for exploring novel and
prospective strategies for GDM treatment.
In the present study, dataset from the GSE98043 mRNA
expression profiling microarray were analyzed to identify core
miRNAs and pathways that can contribute to GDM. Subsequently, the
effect and mechanism of any miRNAs found in this screen in the
regulation of pancreatic β cell proliferation, apoptosis and
insulin secretion, in addition to the inflammatory response, were
investigated in MIN6 cells under high glucose (HG) conditions. It
is hoped that results of these experiments can provide a
theoretical basis for the investigation into the pathogenesis and
potential drug targets for GDM.
Materials and methods
MicroRNA expression profile data from
Gene Expression Omnibus (GEO)
MicroRNA array expression profile data (GSE98043)
was downloaded from GEO database (https://www.ncbi.nlm.nih.gov/geo/) (26). GSE98043 was analyzed using
Agilent-070156 Human_miRNA_V21.0_Microarray 046064 (Feature Number
version) and consisted of 4 plasma samples from pregnant women (2
from normal controls and 2 from GDM patients). Differentially
expressed miRNAs (DEmiRNA) were identified through the R ‘limma’
package (Version 4.2), which is a widely used tool that can be used
to analyze data from any GEO series and significance analysis of
microarray (SAM), to determine the differential expression of
microRNAs among groups (27).
miRNAs were considered to be differentially expressed according to
the P<0.05 threshold from the limma analysis and median false
discovery rate <0.05 from SAM. Data were visualized as heat maps
using the online tool Morpheus (https://software.broadinstitute.org/morpheus/).
Blood sample collection
Whole blood samples were collected from 30 women who
underwent pregnancies with GDM and 30 healthy women who underwent
healthy pregnancies (control) at the Department of Gynecology,
Handan Central Hospital (Handan, China) between November, 2019 and
November, 2020. GDM was diagnosed according to the World Health
Organization/International Association of the Diabetes and
Pregnancy Study Groups criteria (28,29).
The GDM patients with pre-gestational diabetes, multiple gestations
accompanied with further complications and those taking medications
were all excluded and pregnancies with GDM who did not fulfill any
of the exclusion criteria were recruited in the current study. The
control participants without GDM, pre-eclampsia, low birthweight,
or preterm delivery were selected in this study. The main
clinicopathological parameters are reported and summarized in
Table I, and data are presented as
means ± SD for both control and GDM groups. The present study was
approved by the Ethics Committee of Handan Central Hospital.
Briefly, the whole blood samples were obtained from all individuals
through venous blood collection prior to breakfast, before they
were put into a heparin sodium-containing anticoagulation tube. All
blood samples were obtained with informed consent. The blood
glucose levels were measured using a glucose assay kit (cat. no.
ab65333; Abcam).
 | Table IClinicopathological characteristics
of individuals with GDM and healthy controls. |
Table I
Clinicopathological characteristics
of individuals with GDM and healthy controls.
|
Characteristics | Control (n=30) | GDM (n=30) | P-value |
|---|
| Age, years | 31.32±2.45 | 32.02±2.18 | 0.529 |
| Pregnancy period,
weeks | 39.84±0.94 | 39.67±0.86 | 0.635 |
| Birth weight,
kg | 3.41±0.27 | 3.99±0.42 | 0.020 |
| BMI,
kg/m2 | 23.22±1.23 | 24.54±1.86 | 0.029 |
Cell culture and culture
conditions
The MIN6 murine pancreatic β-cell line, a
well-established cellular model for studying islet β-cell function,
was purchased from The Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences. They were cultured with DMEM (Gibco;
Thermo Fisher Scientific, Inc.) containing 12% FBS (Gibco; Thermo
Fisher Scientific, Inc.), 50 µM β-mercaptoethanol, penicillin (100
U/ml) and streptomycin (100 U/ml; Sigma-Aldrich; Merck KGaA) at
37˚C in a humidified atmosphere under 5% CO2.
In the present study, normal glucose (3.3 mM
glucose) was set as the control group. MIN6 cells were incubated in
complete medium containing 16.7 mM glucose (final concentration in
the medium) for 24 h at 37˚C, which was set as the HG group
(30). By contrast, cells in the
HG + miR-143-3p mimics or mimic-negative control (NC) group were
transfected with miR-143-3p mimics or mimics NC for 24 h at 37˚C,
before being cultured in 16.7 mM glucose for another 24 h at 37˚C.
In addition, cells in the HG + miR-143-3p mimics +
pcDNA-TAK1/pcDNA-empty group were co-transfected with the
miR-143-3p mimic and the pcDNA-TAK1 plasmid or pcDNA-empty vector
at 37˚C for 6 h, and subsequently the transfection reagent was
removed and the medium containing 16.7 mM glucose was added for 48
h at 37˚C.
Cell transfection
miR-143-3p mimics, mimics negative control (NC),
miR-143-3p inhibitor, inhibitor NC and pcDNA-TAK1 were designed and
synthesized by Shanghai GenePharma Co., Ltd. and the pcDNA 3.1
vector (pcDNA empty) was used as a control for the overexpression
vector pcDNA-TAK1. The miR-143-3p mimics sequence was
5'-UGAGAUGAAGCACUGUAGCUC-3'; whereas the mimics NC sequence was
5'-ACUGAGCUCGUAGUAUGCAAG-3'. The miR-143-3p inhibitor sequence was
5'-GAGCUACAGUGCUUCAUCUCA-3', whilst the inhibitor NC sequence was
5'-AGUCUCGCGCUUACUGAAUCA-3'.
When the MIN6 cell confluency reached 75%,
miR-143-3p mimics (50 nM) or inhibitor (100 nM) and/or
pcDNA-TAK1/pcDNA empty vector (2 µg; accession no. NM_172688.3)
were transfected or co-transfected into the cells using the
Lipofectamine 2000 reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) by following the manufacturer's protocol. After 48 h in
total, the cells were collected for further experiments.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA from whole blood and cultured cells were
extracted using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. For the
reverse transcription of miRNA and mRNA, cDNA was synthesized using
an miScript II RT kit (Qiagen GmbH) at 37˚C for 60 min and
PrimeScript™ RT reagent kit (Takara Bio, Inc.) at 42˚C for 30 min,
respectively. Subsequent qPCR was performed using the
SYBR® Premix Ex Taq™ (Takara Bio, Inc.) on an ABI Prism
7500 Sequence Detection System (Thermo Fisher Scientific, Inc.).
The following thermocycling conditions were used: Pre-denaturation
at 95˚C for 10 sec; followed by 40 cycles of denaturation at 95˚C
for 15 sec and annealing at 55˚C for 15 sec, and extension at 72˚C
for 40 sec; and a final extension of 10 min at 72˚C. The following
primer sequences were used: miR-143-3p forward,
5'-CTGGCGTTGAGATGAAGCAC-3' and reverse, 5'-CAGAGCAGGGTCCGAGGTA-3';
U6 forward, 5'-CTCGCTTCGGCAGCACA-3' and reverse,
5'-AACGCTTCACGAATTTGCGT-3'; TGFβ-activated kinase 1 (TAK1) forward,
5'-AGCTTGATGACACGCTGTTG-3' and reverse, 5'-GAGTTGCTCTGCCCTTCATC-3';
GAPDH forward, 5'-CTCATGACCACAGTCCATGCCATCACTG-3' and reverse,
5'-CATGAGGTCCACCACCCTGTTGCTGTA-3'. miR-143-3p expression was
normalized to that of U6, whereas TAK1 expression was normalized to
that of GAPDH. Data were analyzed by using the 2-ΔΔCq
method (31).
Detection of glucose-stimulated
insulin secretion
Cells were incubated in DMEM containing 3.3 or 16.7
mM glucose at 37˚C for 24 h, respectively. After cell sonication as
below: 3 sec On and 5 sec OFF on ice for 10 min, the supernatant
was collected after centrifugation at 12,000 x g for 10 min at 4˚C.
Glucose-stimulated insulin secretion into supernatant was measured
using an insulin ELISA kit (Cat no. P01325) from RayBiotech, Inc.
according to the manufacturer's protocols.
Cell viability assay
Cells were seeded into a 96-well plate with
1x104 cells per well. When the MIN6 cell confluency
reached 75%, the cells were transfected at 37˚C for 24 h with the
miRNA mimics/inhibitor or plasmids using the Lipofectamine 2000
transfection reagent, before being treated with 16.7 mM glucose.
After 0, 12, 24 and 48 h treatment, 10 µl CCK-8 solution (Dojindo
Molecular Technologies, Inc.) was added to each well, before the
cells were continuously incubated for another 2 h at 37˚C and the
absorbance was read using a microplate reader (BioTek Instruments,
Inc.) at 450 nm according to the manufacturer's protocols.
Apoptosis assay
Cells were seeded into six-well plates at
1x106 cells per well. When the MIN6 cell confluency
reached 75%, the cells were transfected at 37˚C for 24 h with the
miRNA mimics/inhibitor or plasmids using the Lipofectamine 2000
transfection reagent, before they were treated with 16.7 mM
glucose. After 24 h treatment, MIN6 β-cells in each group were
detached by trypsinization and collected by centrifugation at
12,000 x g for 10 min at 4˚C. To measure apoptosis, the Annexin
V-FITC Apoptosis Staining/Detection (Abcam) was used according to
the manufacturer's instructions. The cells (1x105 cells)
were then resuspended in pre-cooled 500 µl binding buffer and
incubated with 5 µl Annexin V-FITC and 1 µl propidium iodide (PI).
After incubation at room temperature in the dark for 15 min, cell
apoptosis was analyzed using a FACScan flow cytometer (BD
Biosciences). The MultiCycle Software version 5.0 (Phoenix Flow
Systems, San Diego, CA, USA) for Windows 7 (Microsoft Corporation,
Redmond, WA, USA) was used to analyze the experimental data.
Caspase 3 activity assay
Caspase-3 activity was measured using a Caspase-3
Activity kit (Cat no. C1115, Beyotime Institute of Biotechnology)
according to the manufacturer's protocol. Briefly, cells were
harvested 24 h after 16.7 mM glucose treatment at 37˚C and lysed in
RIPA buffer (cat no. P0013B, Beyotime Institute of Biotechnology).
Caspase-3 activity assay was performed in 96-well plates by
incubating 10 µl of each cell lysate sample in 80 µl reaction
buffer containing 10 µl of the caspase-3 substrate Ac-DEVD-pNA; (2
mM) at 37˚C for 2 h. The samples were measured with a microplate
reader (Model 680; Bio-Rad Laboratories, Inc., Hercules, CA, USA)
at an absorbance of 405 nm.
ELISA
In the culture supernatants, TNF-α (cat no. MTA00B),
IL-8 (cat no. DY442) and IL-6 (cat no. M6000B) levels were measured
using ELISA kits (R&D Systems, Inc.) according to the
manufacturer's protocols. Absorbance was detected using an
automatic multi-well spectrophotometer (Bio-Rad Laboratories, Inc.)
at 450 nm.
Luciferase reporter assay
TargetScan's bioinformatics prediction (version 7.2;
www.targetscan.org/vert_72) was used to
predict gene targets for miR-143-3p. Gene ontology
enrichment analysis and alternative analyses were carried out using
EnrichR (version: January 23rd). An adjusted P-value cut-off of
below 0.05 was used to filter results. The fragment of the
3'-untranslated region (UTR) of TAK1 [wild-type (wt) or mutant
(mut)] was amplified and cloned into the pGL-control vector
(Promega). Site-directed mutagenesis of the TAK1 3'-UTR at the
putative miR-143-3p binding site was performed using a QuikChange
II Site-Directed Mutagenesis kit (Agilent Technologies, Inc., Santa
Clara, CA, USA). MIN6 cells at a density of 1x105 per
well were seeded into a 24-well plate until they reached ~60%
confluence. In total, 2.5 µg wild-type (wt)-TAK1-UTR-pGL3 or mutant
(mut)-TAK1-UTR-pGL3 plasmids was co-transfected with 100 nM
miR-143-3p mimics or inhibitors using Lipofectamine®
2000 (Thermo Fisher Scientific, Inc.). Following 48 h transfection,
the luciferase activity was measured using a Dual-luciferase
reporter assay system (Promega Corporation). pRL-TK plasmid,
containing the Renilla luciferase activity, was transfected
as an internal control.
Western blotting
Total cellular proteins were extracted using RIPA
buffer (Beyotime Institute of Biotechnology) and protein
concentration was determined using the BCA kit (Pierce; Thermo
Fisher Scientific, Inc.). Protein isolation and immunoblotting were
performed as previously described (32). Briefly, total protein (40 µg/lane)
was separated by 8% SDS-PAGE and electrophoretically transferred
onto a PVDF membrane (MilliporeSigma). Subsequently, membranes were
blocked with 5% skim milk for 1 h at room temperature. The
membranes were incubated with specific primary antibodies against
TAK1 (cat no. 5206, 1:1,000), cleaved caspase 3 (cat. no. 9661,
1:1,000), inhibitor of NF-κB (IκB) kinase subunit β (IKKβ; cat. no.
8943; 1:1,000), phosphorylated (p-)-IKKβ (cat. no. 2697; 1:1,000),
anti-IκBα (cat. no. 4812; 1:1,000), p-IκBα (cat. no. 2859; 1:1,000)
or β-actin (cat. no. 4970; 1:1,000) at 4˚C overnight. Following
incubation with HRP-conjugated anti-rabbit IgG secondary antibody
(cat. no. 7074; 1:2,000) for 1 h at room temperature, the bands
were detected using ECL Prime Western Blotting Detection
Reagent(Cytiva). All antibodies were purchased from Cell Signaling
Technology, Inc. The intensities of the bands of interest were
analyzed using the ImageJ software (version 1.46; National
Institutes of Health).
Statistical analysis
Data are represented as the mean ± SD from ≥ three
replicates and analyzed using the SPSS 22.0 software package (IBM
Corp.). Significance was determined using unpaired Student's t-test
between two groups whereas for multiple comparisons among > two
groups one-way ANOVA followed by Tukey's post hoc test was used.
P<0.05 was considered to indicate a statistically significant
difference. Correlation analysis was performed using Pearson's
correlation coefficient analysis.
Results
miR-143-3p is downregulated in the
peripheral blood of patients with GDM
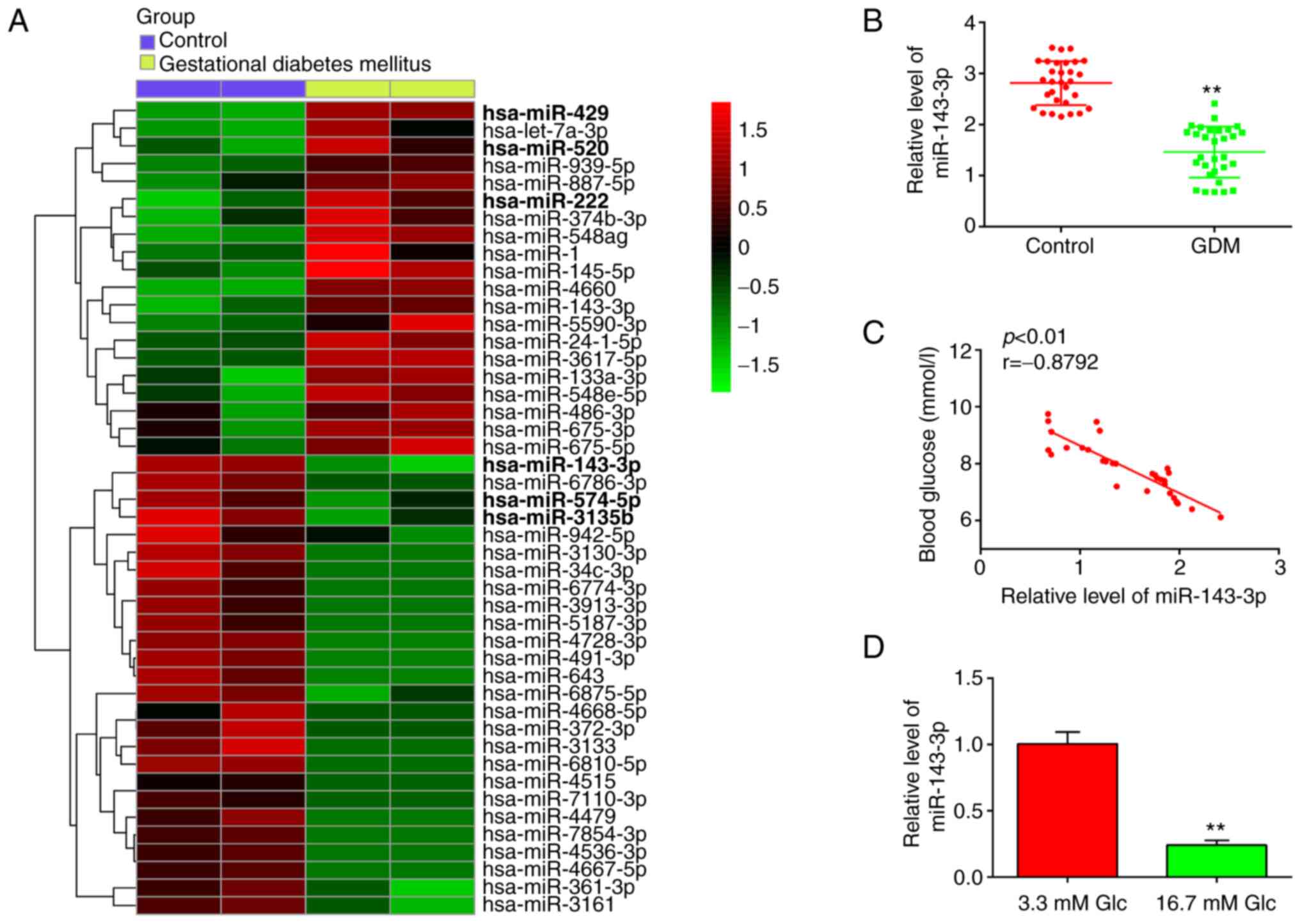
Firstly, 46 miRNAs, including 20 upregulated and 26
downregulated miRNAs, were found from the GEO dataset with the
accession number GSE98043 (Fig.
1A). In this list, 20 miRNAs were upregulated and 26 miRNAs
were downregulated in the blood samples between the GDM and Control
group. Notably, miR-143-3p, miR-574-5p and miR-3135b were found to
be downregulated, whereas miR-429, miR-520 and miR-222 were
particularly upregulated, compared with those in the control group
(Fig. 1A). Association between
miRNAs miR-574-5p, miR-3135b, miR-429, miR-520, miR-222 and GDM has
been previously reported (25,26,33,34),
suggesting the experimental validity of our microarray findings.
Notably, miR-143-3p showed the highest degree of downregulation
among other miRNAs screened in this dataset, consistent with
previous studies (35,36). Furthermore, miR-143-3p has been
previously reported to serve an important role in the inflammatory
response and apoptosis in several diseases, such as mycoplasmal
pneumonia and autoimmune hepatitis (37,38).
However, its role in the development of inflammatory response and
apoptosis in GDM remains unclear. Therefore, miR-143-3p was
selected for further study.
To validate the expression levels of miR-143-3p,
RT-qPCR was performed in blood samples isolated from 30 pregnant
patients with GDM and 30 healthy pregnant individuals. As shown in
Fig. 1B, miR-143-3p levels were
significantly lower in the peripheral blood of patients with GDM
compared with those in healthy individuals. Pearson correlation
analysis revealed an inverse correlation between the blood glucose
concentration and miR-143-3p levels among the patients with GDM
(r=-0.8792; Fig. 1C).
Subsequently, HG-induced MIN6 cell model was established as
previously described (28).
RT-qPCR results demonstrated that miR-143-3p expression was
significantly decreased in the HG group compared with that in the
control group (Fig. 1D). These
data suggest that miR-143-3p may be involved in the pathogenesis of
GDM.
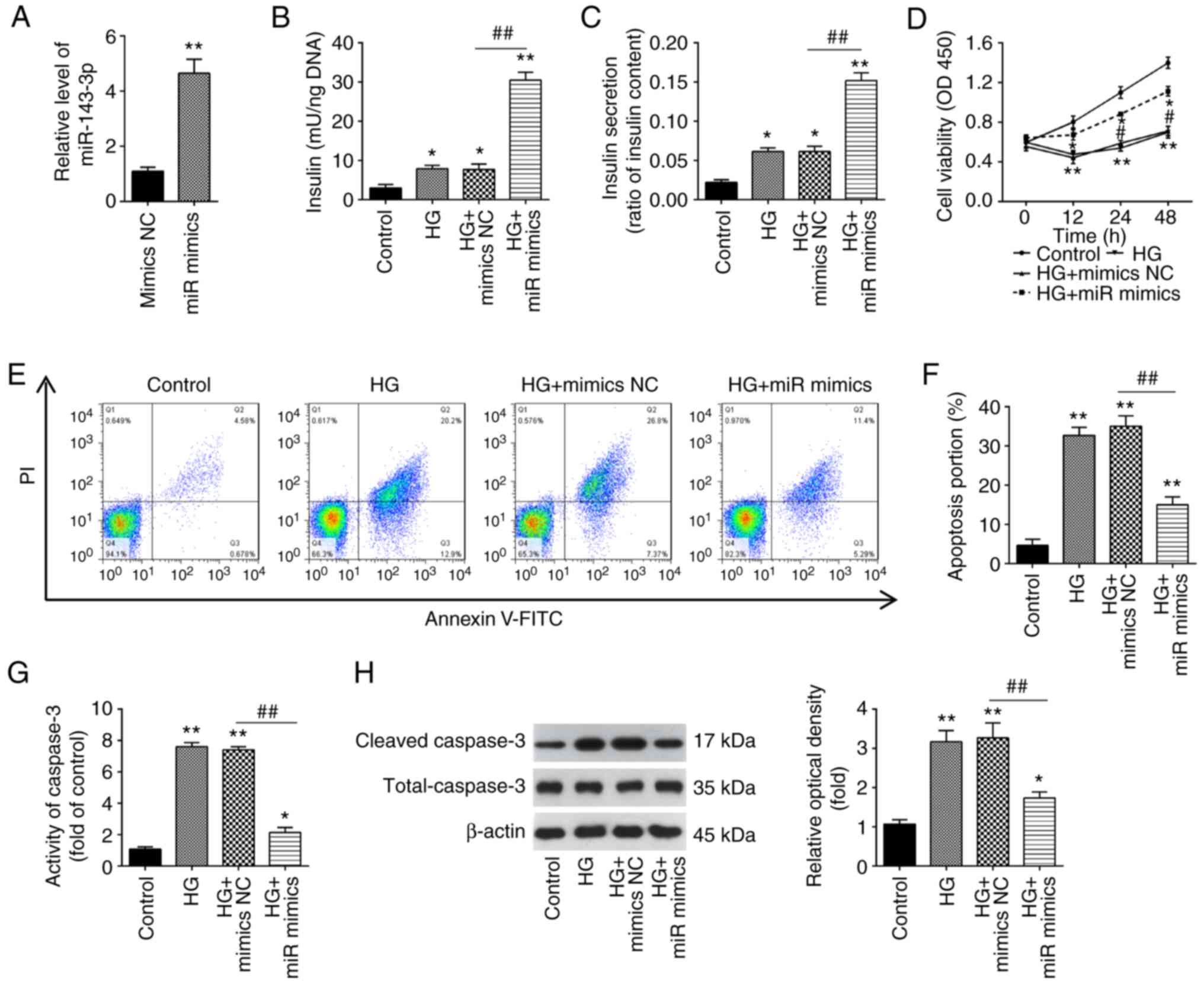
Overexpression of miR-143-3p promotes
insulin secretion and inhibites cell apoptosis in HG-stimulated
MIN6 cells
To investigate the function of miR-143-3p in GDM,
MIN6 cells were transfected with the miR-143-3p mimics or mimics
NC, before being cultured under 16.7 mM glucose (HG). According to
the results of RT-qPCR, miR-143-3p was found to be significantly
increased after miR-143-3p transfection compared with that in cells
transfected with mimics NC, suggesting that the transfection of
miR-143-3p mimics was successful (Fig.
2A). As shown in Fig. 2B and
C, HG resulted in a significant
increase in insulin concentration and secretion in MIN6 cells
compared with those in the control group, which was significantly
prevented by miR-143-3p overexpression beforehand. Furthermore, the
effects of miR-143-3p on MIN6 cell viability and apoptosis was next
assessed. A significant decrease in cell viability in HG-stimulated
MIN6 cells was detected, but this inhibitory effect was reversed
after the overexpression of miR-143-3p (Fig. 2D). In addition, HG-induced cell
apoptosis was significantly reversed by miR-143-3p overexpression
in MIN6 cells (Fig. 2E).
Similarly, caspase 3 activity assay also revealed that caspase 3
activity was significantly increased by HG, which was likewise
significantly reversed by the overexpression of miR-143-3p in MIN6
cells (Fig. 2F and G). The caspase 3 activity assay data were
supported by those from western blotting, which revealed
significantly increased caspase 3 cleavage in response to HG but
was in turn significantly reversed by miR-143-3p overexpression
(Fig. 2H). These data indicate
that miR-143-3p overexpression can promote insulin secretion whilst
inhibiting cell apoptosis in HG-treated MIN6 cells.
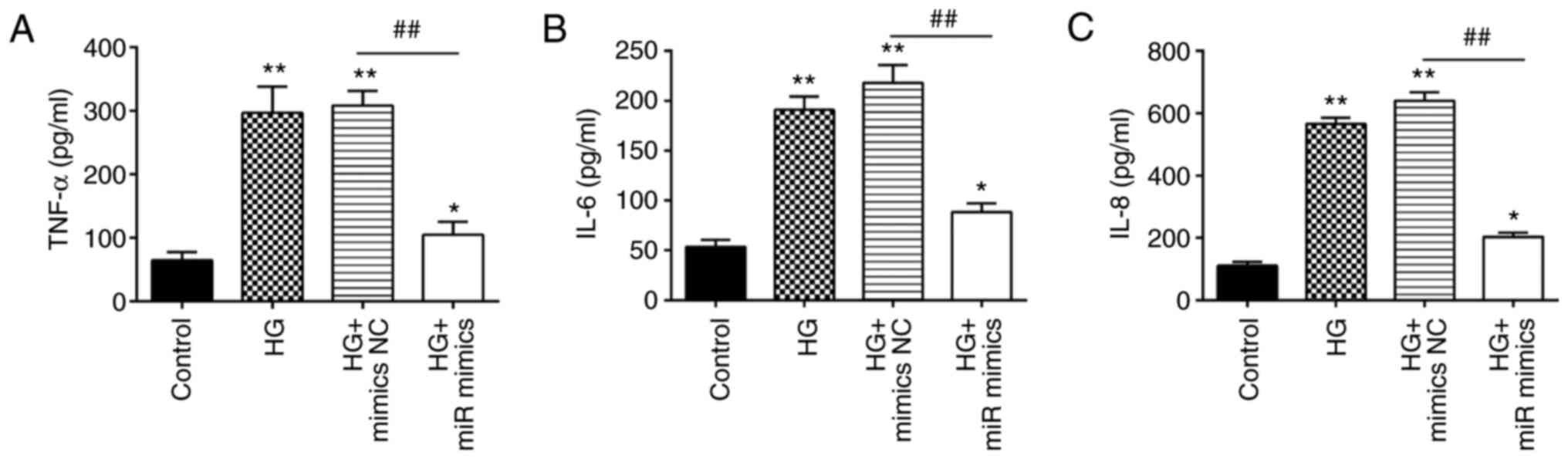
Overexpression of miR-143-3p protects
MIN6 cells against HG-induced inflammation
Apart from insulin resistance, excessive
inflammation is also one of the key factors involved in the
pathogenesis of GDM (39,40). Therefore, the effects of miR-143-3p
on the inflammatory response in MIN6 cells after HG treatment were
next investigated. HG stimulation led to a significant increase in
the levels of proinflammatory factors TNF-α, IL-6, and IL-8
(Fig. 3A-C). However,
overexpression of miR-143-3p significantly decreased the levels of
these proinflammatory factors previously raised by HG in MIN6
cells. These data suggest that the overexpression of miR-143-3p can
suppress the HG-induced inflammatory response in MIN6 cells.
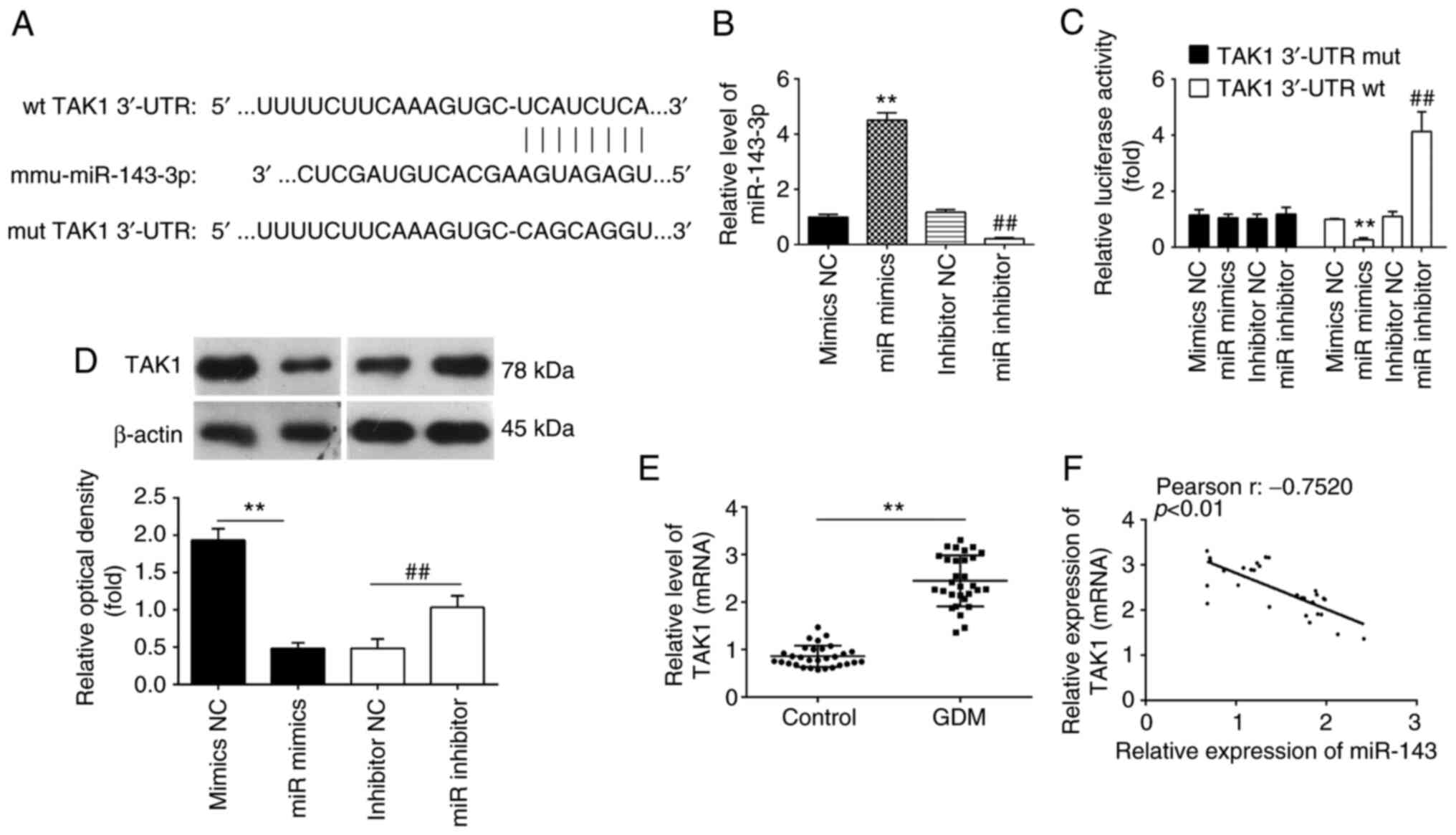
TAK1 is a direct target of miR-143-3p
in MIN6 cells
To investigate the underlying molecular mechanism
involved in the miR-143-3p-mediated suppression of inflammation and
apoptosis, the TargetScan software was next used to analyze the
putative target genes of miR-143-3p. Bioinformatics analyses
predicted that 3'-UTR regions of the TAK1 gene contains a potential
binding site for miR-143-3p (Fig.
4A). Therefore, MIN6 cells were next transfected with either
the miR-143-3p mimics or miR-143-3p inhibitor. The expression
levels of miR-143-3p were significantly increased and decreased
after transfection with the miR-143-3p mimics and miR-143-3p
inhibitor, respectively, compared with those in their corresponding
NC (Fig. 4B). To verify if TAK1 is
indeed a direct target of miR-143-3p, a dual-luciferase reporter
assay was performed. Compared with that in their corresponding NCs,
overexpression of miR-143-3p was found to significantly decrease
the luciferase activities of the 3'-UTR segment of TAK1, whilst
luciferase activities of the same construct was significantly
increased by miR-143-3p knockdown (Fig. 4C). Subsequently, the potential
effects of miR-143-3p on the expression of TAK1 was measured on a
protein level in MIN6 cells by western blot analysis. As shown in
Fig. 4D, compared with that in
their corresponding NCs, the expression of TAK1 proteins was
significantly reduced following the overexpression of miR-143-3p,
but was significantly upregulated following the knockdown of
miR-143-3p, in the MIN6 cells. Since the expression levels of
miR-143-3p were lower in the whole blood samples of patients with
GDM, the expression levels of TAK1 were also detected in the
samples from 30 pregnant individuals with GDM and 30 healthy
pregnant individuals. As shown in Fig.
4E, TAK1 expression was significantly upregulated in whole
blood samples of patients with GDM compared with that of healthy
individuals. In addition, Pearson correlation analysis revealed an
inverse correlation between TAK1 and miR-143-3p expression among
the GDM patients (r=-0.7520; Fig.
4F). These results suggest that TAK1 is a direct functional
target of miR-143-3p in MIN6 cells.
Overexpression of miR-143-3p
alleviates HG-induced pancreatic β-cell dysfunction by targeting
TAK1
To clarify if TAK1 is involved the inhibitory
effects of miR-143-3p on HG-induced inflammation and apoptosis in
MIN6 cells, the TAK1 expression vector pcDNA-TAK1 and miR-143-3p
mimics were co-transfected into MIN6 cells followed by HG
treatment. First, the expression of the TAK1 protein was markedly
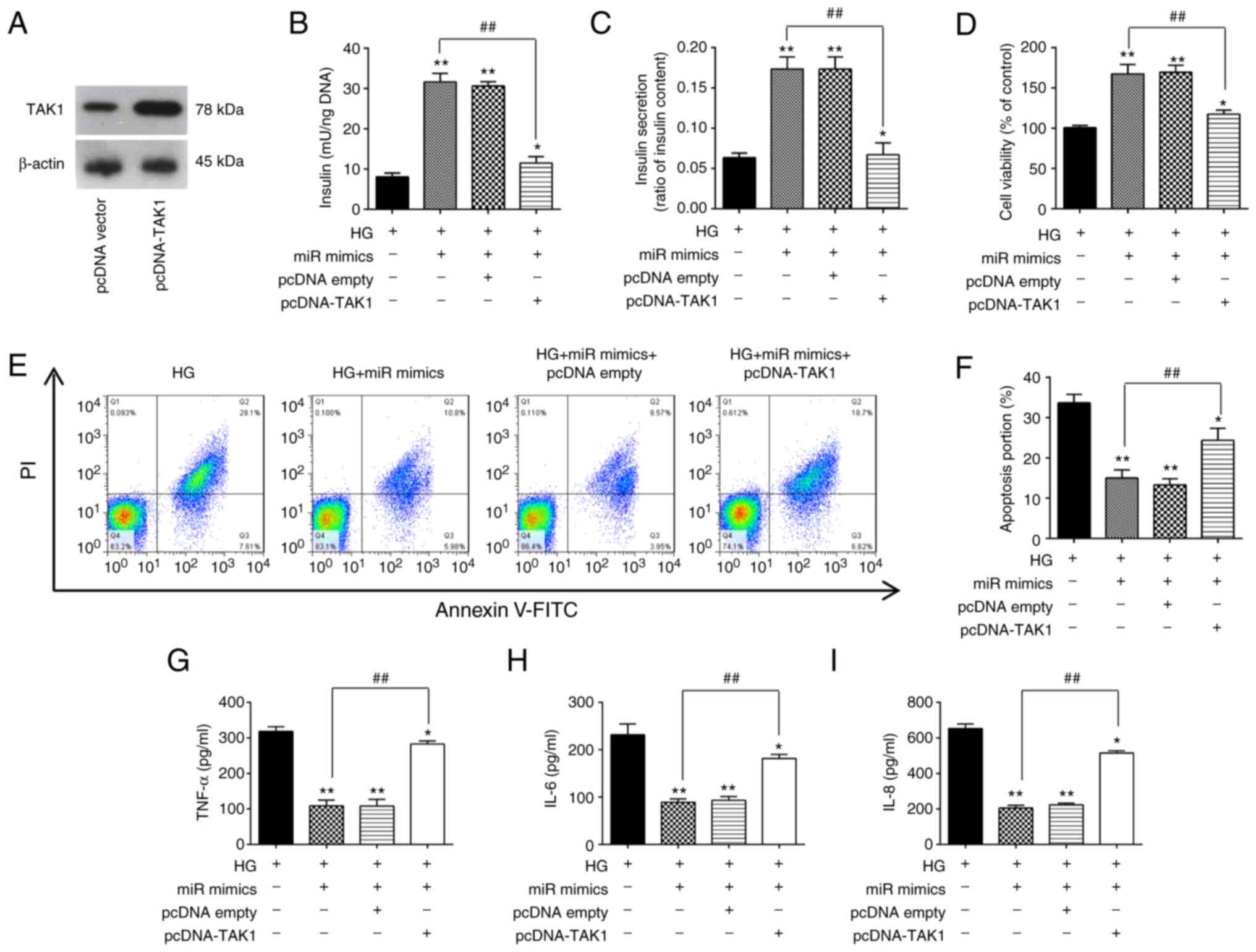
increased in MIN6 cells after transfection with the pcDNA-TAK1
plasmid compared with that in cells transfected with the empty
plasmid (Fig. 5A). Insulin
concentration and insulin secretion were next measured, both of
which were significantly increased by miR-143-3p overexpression in
HG-treated MIN6 cells (Fig. 5B and
C). However, these miR-143-3p
overexpression-mediated increases were in turn significantly
reversed by TAK1 overexpression (Fig.
5B and C). In addition, the
increased cell viability induced by miR-143-3p mimic transfection
was also significantly reversed by TAK1 overexpression in
HG-treated MIN6 cells (Fig. 5D).
Subsequently, apoptosis and inflammatory response in HG-treated
MIN6 cells were measured. As shown in Fig. 5E and F, the inhibitory effects of miR-143-3p
overexpression on the apoptosis in HG-treated MIN6 cells were also
significantly reversed by TAK1 overexpression. Similarly, the
reduction of proinflammatory cytokine secretion caused by
miR-143-3p overexpression were all significantly reversed by TAK1
overexpression (Fig. 5G-I). These
data therefore strongly supports the notion that miR-143-3p
overexpression can protect MIN6 cell function by negatively
targeting TAK1.
Overexpression of miR-143-3p blocks
the TAK1/NF-κB pathway in MIN6 cells
It has been extensively reported that NF-κB
functions as a key regulator in the inflammatory process in a wide
range of diseases, particularly inflammation-related diseases
including GDM (41,42). The present study showed that
miR-143-3p overexpression can negatively regulate the expression of
TAK1 in MIN6 cells, which is an important upstream activator of the
NF-κB pathway. Therefore, the potential effects of miR-143-3p on
the activation of TAK1/NF-κB signaling in vitro were next
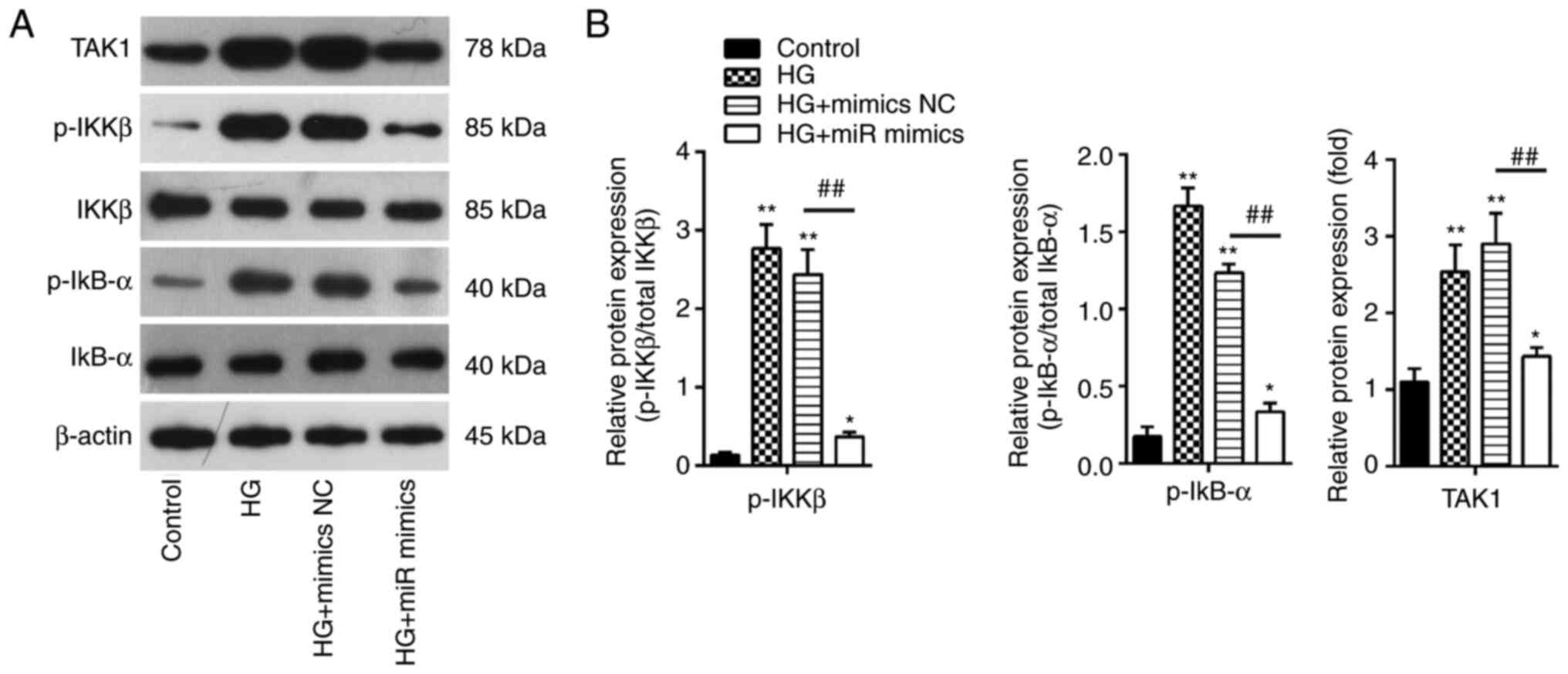
examined. The protein expression levels of TAK1, p-IKKβ and p-IκBα
were detected by western blotting. It was found that HG treatment
led to a significant increase in IKKβ and IκBα phosphorylation in
addition to increasing TAK1 expression in MIN6 cells compared with
those in the control group (Fig.
6A). However, these effects were significantly reversed by
miR-143-3p overexpression in MIN6 cells (Fig. 6B). These results suggest that
miR-143-3p overexpression blocked the TAK1/NF-κB pathway in MIN6
cells under HG conditions.
Discussion
In the present study, miR-143-3p levels were found
to be lower in the whole blood from patients with GDM, which were
in turn inversely correlated with blood glucose concentrations.
Furthermore, overexpression of miR-143-3p promoted cell viability
and insulin secretion, inhibited apoptosis and the inflammatory
response in pancreatic β cells cultured under high-glucose
conditions. It was also found that the TAK1/NF-κB signaling pathway
may be involved. These findings suggest that miR-143-3p can
regulate the pathogenesis of GDM and may serve as a novel
therapeutic target for GDM.
Several studies have reported that GDM can alter the
expression profile of miRNAs, such that they can be exploited as
diagnostic targets for GDM that regulate cell physiology (30,43).
Chen et al (44) previously
demonstrated that high expression levels of miR-351 in liver
tissues can negatively regulate the PI3K/AKT pathway by interacting
with flotillin 2, thereby acting as an inhibitor of GDM. In another
study, Pfeiffer et al (45)
reported miR-330-3p as being significantly upregulated in women
with GDM compared to nondiabetic controls and miR-330-3p to be
significantly related to the percentage of caesarean deliveries.
This strong novel association of circulating miR-330-3p with risk
of primary caesarean delivery as a pregnancy outcome linked with
poor maternal glycaemic control. In addition, Fu et al
(46) reported that miR-875-5p
silencing notably reduced expression levels of pro-inflammatory
markers by targeting thioredoxin reductase 1 cytoplasmic in rats
with GDM. By contrast, Zhang et al (25) revealed that silencing miR-222
expression can suppress the inflammatory response in mice with GDM
by promoting CXCR4 whist inactivating NOD-, LRR- and pyrin
domain-containing protein 3 inflammasomes. In the present study,
after retrieving a miRNA microarray dataset (GSE98043), large
numbers of miRNAs were found to be aberrantly expressed in the
whole blood of patients with GDM, including miR-143-3p, which was
downregulated. In addition, there was a significant inverse
correlation between miR-143-3p levels and the blood glucose
concentrations. These data suggest that miR-143-3p may be involved
in GDM, implying its potential role in GDM.
The role of miR-143-3p in GDM has been studied
previously (35,47). Decreased miR-143-3p placental
expression was reported in macrosomia, which may be associated with
maternal and infant health problems (36). These previous studies suggest a
potential role for miR-143-3p in GDM. Several studies have shown
that miR-143-3p is involved in the initiation of the inflammatory
response and apoptosis in various disease settings (48). miR-143-3p overexpression attenuated
the Derp1-induced inflammatory response and apoptosis of BEAS-2B
cells by downregulating high mobility group box 1 (HMGB1)
expression in Bronchial asthma (49). In addition, miR-143-3p
overexpression has been demonstrated to alleviate the pulmonary
inflammatory response and suppress alveolar epithelial cell
apoptosis in mice with mycoplasmal pneumonia (37). miR-143-3p was also found to at
least partially reverse obesity-induced insulin resistance by
downregulating insulin-like growth factor 2 receptor expression in
differentiated hADSCs (50).
However, the role of miR-143-3p in GDM remains elusive and requires
further investigation. In the present study, miR-143-3p
overexpression was shown to enhance cell viability and insulin
secretion, inhibited the apoptosis and inflammatory response in
pancreatic β cells following HG stimulation. This suggests that
miR-143-3p may be involved in the pathogenesis of GDM by regulating
pancreatic β-cell function.
Although miR-143-3p is emerging as an important
functional component in GDM development, the molecular mechanisms
remain poorly understood. In the present study, TAK1, also known as
MAP3K7, was found to be a target of miR-143-3p that can be targeted
by miR-143-3p, suggesting that the development of GDM may be
associated with the TAK1 signaling pathway. Furthermore, the
seemingly therapeutic effects induced by miR-143-3p overexpression
on the pancreatic β cells treated with HG were reversed by TAK1
overexpression. As a member of the MAPK kinase kinase family, TAK1
can be activated by various inflammatory mediators, such as IL-1β
(51-53).
Once activated, TAK1 phosphorylates and activates the IKK complex,
especially IKKβ, leading to the degradation of IκBα and triggers
the activation of the transcription factor NF-κB (54). This activated NF-κB then promotes
the transcription of genes associated with pathways controlling
inflammatory responses, cell proliferation and apoptosis, impairing
β-cell survival and function (55). After transfection with the
miR-143-3p mimics, the expression levels of p-IKKβ and p-IκBα were
significantly decreased in HG-treated MIN6 cells, suggesting the
inactivation of NF-κB pathway. Combing the results from previous
studies and those from the present study that the inflammatory
responses can be relieved by suppressing TAK1/NF-κB activity, where
miR-143-3p may be a potential therapeutic target for GDM treatment
(56,57).
There remain to be some limitations to the present
study. Although it investigated the cellular function of miR-143-3p
and its underlying mechanism in GDM, in vivo studies are
required to validate these preliminary in vitro results
obtained. In addition, only one cell line, MIN6, was used in the
present study. Therefore, further study with other cell lines, such
as INS-1 cells, is necessary for verification. For the screening of
differentially expressed miRNAs in the present study, only the
GSE98043 dataset was used. For bioinformatics analysis, ≥ two
datasets are required to screen differentially expressed microRNAs
to increase reliability. Therefore, new datasets and a more
in-depth analysis are required in any future studies.
In conclusion, results from the present study
provided novel findings that the overexpression of miR-143-3p can
ameliorate pancreatic β cell dysfunction by inhibiting the
TAK1/NF-κB pathway. These data provide novel insights into the
pathogenesis of GDM, which may facilitate the identification of
novel diagnostic or therapeutic targets for GDM.
Acknowledgements
Not applicable.
Funding
Funding: This study was supported by the Mandatory Planning
Projects of Handan Science and Technology Bureau (grant no.
19422083009-9).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CRL and LPY conceived and designed the experiments.
CRL, HQF and LNZ performed the experiments. CRL, HQF, LNZ, YRG and
JJM analyzed the data. LPY and CRL confirm the authenticity of all
the raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Handan Central Hospital (Approval number: 2019014
Handan, China). All blood samples were obtained with informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Boulton A: Strengthening the international
diabetes federation (IDF). Diabetes Res Clin Pract.
160(108029)2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Zhu WW, Yang HX, Wang C, Su RN, Feng H and
Kapur A: High prevalence of gestational diabetes mellitus in
beijing: Effect of maternal birth weight and other risk factors.
Chin Medical J (Engl). 130:1019–1025. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Bellamy L, Casas JP, Hingorani AD and
Williams D: Type 2 diabetes mellitus after gestational diabetes: A
systematic review and meta-analysis. Lancet. 373:1773–1779.
2009.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Alejandro EU, Mamerto TP, Chung G,
Villavieja A, Gaus NL, Morgan E and Pineda-Cortel MRB: Gestational
diabetes mellitus: A harbinger of the vicious cycle of diabetes.
Int J Mol Sci. 21(5003)2020.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Diagnostic criteria and classification of
hyperglycaemia first detected in pregnancy: A World Health
Organization Guideline. Diabetes Res Clin Pract. 103:341–363.
2014.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Samuel VT and Shulman GI: The pathogenesis
of insulin resistance: Integrating signaling pathways and substrate
flux. J Clin Invest. 126:12–22. 2016.PubMed/NCBI View
Article : Google Scholar
|
|
7
|
Ebner M, Lucic I, Leonard TA and Yudushkin
I: PI(3,4,5)P3 Engagement restricts Akt activity to cellular
membranes. Mol Cell. 65:416–431.e6. 2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Ashcroft FM and Rorsman P: Diabetes
mellitus and the β cell: The last ten years. Cell. 148:1160–1171.
2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Prieto D, Contreras C and Sanchez A:
Endothelial dysfunction, obesity and insulin resistance. Curr Vasc
Pharmacol. 12:412–426. 2014.PubMed/NCBI View Article : Google Scholar
|
|
10
|
McGarry JD and Dobbins RL: Fatty acids,
lipotoxicity and insulin secretion. Diabetologia. 42:128–138.
1999.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Donath MY, Schumann DM, Faulenbach M,
Ellingsgaard H, Perren A and Ehses JA: Islet inflammation in type 2
diabetes: From metabolic stress to therapy. Diabetes Care. 31
(Suppl 2):S161–S164. 2008.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Weir GC, Marselli L, Marchetti P, Katsuta
H, Jung MH and Bonner-Weir S: Towards better understanding of the
contributions of overwork and glucotoxicity to the beta-cell
inadequacy of type 2 diabetes. Diabetes Obes Metab. 11 (Suppl
4):S82–S90. 2009.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Busch AK, Cordery D, Denyer GS and Biden
TJ: Expression profiling of palmitate- and oleate-regulated genes
provides novel insights into the effects of chronic lipid exposure
on pancreatic beta-cell function. Diabetes. 51:977–987.
2002.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Shukla GC, Singh J and Barik S: MicroRNAs:
Processing, maturation, target recognition and regulatory
functions. Mol Cell Pharmacol. 3:83–92. 2011.PubMed/NCBI
|
|
16
|
Ma J, Wang J, Liu Y, Wang C, Duan D, Lu N,
Wang K, Zhang L, Gu K, Chen S, et al: Comparisons of serum miRNA
expression profiles in patients with diabetic retinopathy and type
2 diabetes mellitus. Clinics (Sao Paulo). 72:111–115.
2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Sebastiani G, Guarino E, Grieco GE,
Formichi C, Poggi CD, Ceccarelli E and Dotta F: Circulating
microRNA (miRNA) expression profiling in plasma of patients with
gestational diabetes mellitus reveals upregulation of miRNA
miR-330-3p. Front Endocrinol (Lausanne). 8(345)2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhao C, Dong J, Jiang T, Shi Z, Yu B, Zhu
Y, Chen D, Xu J, Huo R, Dai J, et al: Early second-trimester serum
miRNA profiling predicts gestational diabetes mellitus. PLoS One.
6(e23925)2011.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Collares CV, Evangelista AF, Xavier DJ,
Rassi DM, Arns T, Foss-Freitas MC, Foss MC, Puthier D,
Sakamoto-Hojo ET, Passos GA and Donadi EA: Identifying common and
specific microRNAs expressed in peripheral blood mononuclear cell
of type 1, type 2, and gestational diabetes mellitus patients. BMC
Res Notes. 6(491)2013.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Du H, Yin Z, Zhao Y, Li H, Dai B, Fan J,
He M, Nie X, Wang CY, Wang DW and Chen C: miR-320a induces
pancreatic beta cells dysfunction in diabetes by inhibiting MafF.
Mol Ther Nucleic Acids. 26:444–457. 2021.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Fan L, Shan A, Su Y, Cheng Y, Ji H, Yang
Q, Lei Y, Liu B, Wang W, Ning G, et al: MiR-221/222 inhibit insulin
production of pancreatic beta-cells in mice. Endocrinology.
161(bqz027)2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wang P, Wang H, Li C, Zhang X, Xiu X, Teng
P and Wang Z: Dysregulation of microRNA-657 influences inflammatory
response via targeting interleukin-37 in gestational diabetes
mellitus. J Cell Physiol. 234:7141–7148. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Wang P and Wang Z, Liu G, Jin C, Zhang Q,
Man S and Wang Z: MiR-657 promotes macrophage polarization toward
M1 by targeting FAM46C in gestational diabetes mellitus. Mediators
Inflamma. 2019(4851214)2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Zheng Y, Wang Z, Tu Y, Shen H, Dai Z, Lin
J and Zhou Z: miR-101a and miR-30b contribute to inflammatory
cytokine-mediated beta-cell dysfunction. Lab Invest. 95:1387–1397.
2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Zhang H, Luan S, Xiao X, Lin L, Zhao X and
Liu X: Silenced microRNA-222 suppresses inflammatory response in
gestational diabetes mellitus mice by promoting CXCR4. Life Sci.
266(118850)2021.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Wang F, Li Z, Zhao M, Ye W, Wu H, Liao Q,
Bu S and Zhang Y: Circulating miRNAs miR-574-5p and miR-3135b are
potential metabolic regulators for serum lipids and blood glucose
in gestational diabetes mellitus. Gynecol Endocrinol. 37:665–671.
2021.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Smyth GK: Linear models and empirical
bayes methods for assessing differential expression in microarray
experiments. Stat Appl Genet Mol Biol. 3(Article3)2004.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Weinert LS: International association of
diabetes and pregnancy study groups recommendations on the
diagnosis and classification of hyperglycemia in pregnancy: Comment
to the international association of diabetes and pregnancy study
groups consensus panel. Diabetes Care. 33(e97): author reply e98.
2010.PubMed/NCBI View Article : Google Scholar
|
|
29
|
International Association of Diabetes and
Pregnancy Study Groups Consensus Panel. Metzger BE, Gabbe SG,
Persson B, Buchanan TA, Catalano PA, Damm P, Dyer AR, de Leiva A,
Hod M, et al: International association of diabetes and pregnancy
study groups recommendations on the diagnosis and classification of
hyperglycemia in pregnancy. Diabetes Care. 33:676–682.
2010.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Zhao H and Tao S: MiRNA-221 protects islet
beta cell function in gestational diabetes mellitus by targeting
PAK1. Biochem Biophys Res Commun. 520:218–224. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Wen J, Han S, Cui M and Wang Y:
[Retracted] long noncoding RNA MCM3APAS1 drives ovarian cancer
progression via the microRNA1433p/TAK1 axis. Oncol Rep.
46(178)2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wei L, Cao C, Ma X, Wang X, Wang M and
Zhang P: Elevated serum and urine MiR-429 contributes to the
progression of gestational diabetes mellitus. Clin Lab.
67:10.7754/Clin.Lab.2020.200909. 2021.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Wen J and Bai X: miR-520h Inhibits cell
survival by targeting mTOR in gestational diabetes mellitus. Acta
Biochim Pol. 68:65–70. 2021.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Muralimanoharan S, Maloyan A and Myatt L:
Mitochondrial function and glucose metabolism in the placenta with
gestational diabetes mellitus: Role of miR-143. Clin Sci (Lond).
130:931–941. 2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Zhang JT, Cai QY, Ji SS, Zhang HX, Wang
YH, Yan HT and Yang XJ: Decreased miR-143 and increased miR-21
placental expression levels are associated with macrosomia. Mol Med
Rep. 13:3273–3280. 2016.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Wang Y, Li H, Shi Y, Wang S, Xu Y, Li H
and Liu D: miR-143-3p impacts on pulmonary inflammatory factors and
cell apoptosis in mice with mycoplasmal pneumonia by regulating
TLR4/MyD88/NF-kappaB pathway. Biosci Rep.
40(BSR20193419)2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Tu H, Chen D, Cai C, Du Q, Lin H, Pan T,
Sheng L, Xu Y, Teng T, Tu J, et al: microRNA-143-3p attenuated
development of hepatic fibrosis in autoimmune hepatitis through
regulation of TAK1 phosphorylation. J Cell Mol Med. 24:1256–1267.
2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Hajifaraji M, Jahanjou F, Abbasalizadeh F,
Aghamohammadzadeh N, Abbasi MM and Dolatkhah N: Effect of probiotic
supplements in women with gestational diabetes mellitus on
inflammation and oxidative stress biomarkers: A randomized clinical
trial. Asia Pac J Clin Nutr. 27:581–591. 2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Sha H, Zeng H, Zhao J and Jin H:
Mangiferin ameliorates gestational diabetes mellitus-induced
placental oxidative stress, inflammation and endoplasmic reticulum
stress and improves fetal outcomes in mice. Eur J Pharmacol.
859(172522)2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Zhang Y, Qu L, Ni H, Wang Y, Li L, Yang X,
Wang X and Hou Y: Expression and function of lncRNA MALAT1 in
gestational diabetes mellitus. Adv Clin Exp Med. 29:903–910.
2020.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Kuzmicki M, Telejko B,
Wawrusiewicz-Kurylonek N, Lipinska D, Pliszka J, Wilk J, Zielinska
A, Skibicka J, Szamatowicz J, Kretowski A and Gorska M: The
expression of genes involved in NF-kappaB activation in peripheral
blood mononuclear cells of patients with gestational diabetes. Eur
J Endocrinol. 168:419–427. 2013.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Elliott HR, Sharp GC, Relton CL and Lawlor
DA: Epigenetics and gestational diabetes: A review of epigenetic
epidemiology studies and their use to explore epigenetic mediation
and improve prediction. Diabetologia. 62:2171–2178. 2019.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Chen SH, Liu XN and Peng Y: MicroRNA-351
eases insulin resistance and liver gluconeogenesis via the PI3K/AKT
pathway by inhibiting FLOT2 in mice of gestational diabetes
mellitus. J Cell Mol Med. 23:5895–5906. 2019.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Pfeiffer S, Sanchez-Lechuga B, Donovan P,
Halang L, Prehn JHM, Campos-Caro A, Byrne MM and López-Tinoco C:
Circulating miR-330-3p in late pregnancy is associated with
pregnancy outcomes among lean women with GDM. Sci Rep.
10(908)2020.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Fu S, Fu S, Ma X, Yang X and Ling J:
miR8755p regulates IR and inflammation via targeting TXNRD1 in
gestational diabetes rats. Mol Med Rep. 23(303)2021.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Hromadnikova I, Kotlabova K, Dvorakova L
and Krofta L: Diabetes mellitus and cardiovascular risk assessment
in mothers with a history of gestational diabetes mellitus based on
postpartal expression profile of microRNAs associated with diabetes
mellitus and cardiovascular and cerebrovascular diseases. Int J Mol
Sci. 21(2437)2020.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Yu B, Zhao Y, Zhang H, Xie D, Nie W and
Shi K: Inhibition of microRNA-143-3p attenuates myocardial
hypertrophy by inhibiting inflammatory response. Cell Biol Int.
42:1584–1593. 2018.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Cai XJ, Huang LH, Zhu YK and Huang YJ:
LncRNA OIP5AS1 aggravates house dust miteinduced inflammatory
responses in human bronchial epithelial cells via the
miR1433p/HMGB1 axis. Mol Med Rep. 22:4509–4518. 2020.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Lin X, Du Y, Lu W, Gui W, Sun S, Zhu Y,
Wang G, Eserberg DT, Zheng F, Zhou J, et al: CircRNF111 protects
against insulin resistance and lipid deposition via regulating
miR-143-3p/IGF2R axis in metabolic syndrome. Front Cell Dev Biol.
9(663148)2021.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Ajibade AA, Wang HY and Wang RF: Cell
type-specific function of TAK1 in innate immune signaling. Trends
Immunol. 34:307–316. 2013.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Delaney JR and Mlodzik M: TGF-beta
activated kinase-1: New insights into the diverse roles of TAK1 in
development and immunity. Cell Cycle. 5:2852–2855. 2006.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Fan Y, Yu Y, Mao R, Zhang H and Yang J:
TAK1 Lys-158 but not Lys-209 is required for IL-1beta-induced
Lys63-linked TAK1 polyubiquitination and IKK/NF-κB activation. Cell
Signal. 23:660–665. 2011.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Hayden MS and Ghosh S: Shared principles
in NF-kappaB signaling. Cell. 132:344–362. 2008.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Karin M and Ben-Neriah Y: Phosphorylation
meets ubiquitination: The control of NF-[kappa]B activity. Ann Rev
Immunol. 18:621–663. 2000.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Lee JH, Lee JH and Rane SG: TGF-β
signaling in pancreatic islet β cell development and function.
Endocrinology. 162(bqaa233)2021.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Ardestani A and Maedler K: The hippo
signaling pathway in pancreatic β-cells: Functions and regulations.
Endocr Rev. 39:21–35. 2018.PubMed/NCBI View Article : Google Scholar
|