Introduction
Asthma, also known as bronchial asthma, is a common
chronic airway inflammatory disease. It is characterized by
reversible airflow obstruction, airway inflammation, persistent
airway hyperresponsiveness and airway remodeling and the release of
inflammatory mediators that can cause tissue damage and airway
dysfunction (1-3).
Asthma is a globally significant non-communicable disease with
major public health consequences for both children and adults and
high morbidity and mortality (4).
Pediatric asthma, also known as pediatric bronchial asthma, is the
most common chronic disease noted in children worldwide and its
mortality has reached a high level with a mortality rate ranging
from 0-0.7 per 100,000 individuals in children (5). An increasing trend of this disease
has been noted in recent years (6,7).
Therefore, it is important to explore the molecular mechanisms
involved in pediatric asthma and develop more effective and
feasible strategies for this disease.
The forkhead box (FOX) transcription factor family
is composed of a group of evolutionarily conserved transcriptional
regulators that play an important role in embryonic growth and
development and maintaining cellular homeostasis (8-10).
FOXF1, a member of the FOX family of transcription factors, has
been previously shown to be important for lung development,
homeostasis, and injury responses (11,12).
It has been reported that haploinsufficiency of FOXF1 leads
to severe lung malformations and suppresses the development of
pulmonary capillaries during both embryonic and early postnatal
periods (13). In addition,
heterozygous deletions and point mutations in the FOXF1 gene
locus were observed in >40% of patients with alveolar-capillary
dysplasia and misalignment of pulmonary veins (14). An additional study reported that
endothelial-specific deletion of both FOXF1 alleles causes
notable inflammatory cell infiltration, pulmonary edema and
pulmonary hemorrhage in adult mice (15). Although the key role of FOXF1 in
the development of lung injury has been established, the molecular
mechanisms regulated by FOXF1 during asthma remain unclear.
Therefore, the present study aimed to explore the expression levels
of FOXF1 in asthma and the mechanisms by which FOXF1 influences the
development of this disease.
Materials and methods
Cell culture and treatment
The human bronchial epithelial cell line BEAS-2B was
purchased from the American Type Culture Collection. The cells were
cultured in DMEM containing 10% FBS, 1% penicillin/streptomycin in
an incubator at 37˚C in the presence of 5% CO2 for 24 h.
Subsequently, BEAS-2B cells were stimulated with 10 ng/ml TGF-β1
for 24 h at 37˚C. LiCl (β-catenin activator; 20 mM; Sigma-Aldrich;
Merck KGaA) was used to treat the cells for 3 h at 37˚C. Untreated
cells were regarded as the control group.
Cell transfection
FOXF1-specific pcDNA overexpression vector
(pcDNA-FOXF1; 20 µg), cadherin (CDH) 11-specific pcDNA
overexpression vector (pcDNA-CDH11; 20 µg), and the corresponding
negative control (pcDNA3.1; 20 µg) were synthesized by Suzhou
GenePharma Co., Ltd. These recombinant plasmids were transfected
into BEAS-2B cells using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) for 48 h at 37˚C
according to the manufacturer's protocols. Cells were collected
after 48 h transfection for the following experiments.
Cell Counting Kit-8 (CCK-8) assay
BEAS-2B cell viability was evaluated using the CCK-8
assay. The transfected cells were seeded into 96-well plates at a
density of 5x104 cells/ml and cultured in DMEM with 10%
FBS at 37˚C. A total of 10 µl CCK-8 solution (Sangon Biotech Co.,
Ltd.) was added to each well following 24, 48 and 72 h of culture
and incubated for 2 h. The absorbance was measured at 450 nm with a
microplate reader (Bio-Rad Laboratories, Inc.).
Flow cytometry
Apoptosis was detected by the FITC Annexin
V/propidium iodide (PI) Apoptosis Detection Kit I (Guangzhou
RiboBio Co., Ltd.). Briefly, the cells were collected by
centrifugation (1,000 x g for 5 min) at room temperature, washed
with precooled PBS at 4˚C and re-suspended in binding buffer
(Guangzhou RiboBio Co., Ltd.). The cells were incubated with 5 µl
Annexin V-FITC (20 µg/ml) at room temperature for 15 min and with
10 µl PI (10 mg/ml) in a dark room for 5 min at room temperature.
Apoptotic cells were subsequently analyzed using a BD FACS Calibur
flow cytometer (BD Biosciences) and FlowJo software (v10.4; FlowJo
LLC) was used for apoptosis analysis.
ELISA analysis
The cell supernatants of BEAS-2B cells were
collected. The concentration levels of tumor necrosis factor-α
(TNF-α), interleukin (IL)-6 and IL-1β in each group were measured
using Human TNF-alpha Quantikine ELISA kit (cat. no. DTA00D), Human
IL-6 Quantikine ELISA kit (cat. no. D6050) and Human IL-1
beta/IL-1F2 Quantikine ELISA kit (cat. no. DLB50) (R&D Systems,
Inc.), respectively. The optical density of each well was assayed
at 450 nm using a microplate spectrophotometer.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from BEAS-2B cells using the
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) in accordance with the manufacturer's protocols. The quality
and concentration of RNA were detected using NanoDrop 2000 (Thermo
Fisher Scientific) at 260 and 280 nm. Reverse transcription of
first-strand cDNAs was performed using PrimeScript RT Master Mix
(Perfect Real Time; Takara Bio, Inc.) according to the
manufacturer's instructions. cDNA amplification was performed by
RT-qPCR using the SYBR Premix Ex Taq™ II kit (Takara Bio, Inc.).
The following thermocycling conditions were used for qPCR:
Pre-denaturation at 95˚C for 1 min, followed by 40 cycles of
denaturation at 95˚C for 15 sec, annealing at 60˚C for 40 sec and
extension at 72˚C for 15 sec. The primer sequences for PCR are:
FOXF1, forward 5'-GCCATCCAGAGTTCACCCAC-3', and reverse
5'-GAAGCCGAGCCCGTTCAT-3'; CDH11, forward
5'-GGGTTGCCCAAGCTTAATGG-3', and reverse 5'-TTTGATGTCTTTGCGGGGGA-3';
GAPDH, forward 5'-GGGAAACTGTGGCGTGAT-3', and reverse
5'-GAGTGGGTGTCGCTGTTGA-3'. The results were normalized to the
expression levels of GAPDH and were measured using the
2-ΔΔCq method (16).
Immunofluorescence staining
The collected BEAS-2B cells were fixed in 4%
polyoxymethylene for 1 h at room temperature and permeabilized with
0.5% Triton X-100 for 10 min at room temperature. Following
blocking with 2% BSA for 1 h at room temperature, cells were
incubated with an anti-alpha smooth muscle actin primary antibody
(α-SMA; 1:500; cat. no. ab32575; Abcam) at 4˚C overnight.
Subsequently, the secondary antibody (1:400; cat. no. ab150077;
Abcam) was added and incubated for 1 h at room temperature. The
cells were subsequently counterstained with DAPI (Beyotime
Institute of Biotechnology) for 10 min at 37˚C and examined using a
confocal microscope.
Western blotting
The total protein was extracted from the cells using
RIPA buffer (Hunan Auragene Biotechnology Co., Ltd.). The BCA
Protein Assay kit (Beijing Dingguo Changsheng Biotechnology Co.,
Ltd.) was used to detect the protein concentration according to the
manufacturer's protocols. An equal amount of protein (60 µg/lane)
was loaded on 8% SDS-polyacrylamide gels and subsequently
transferred to a pure nitrocellulose blotting membrane (Pall Life
Sciences). Following blocking with 5% non-fat milk in 0.1%
Tris-buffered saline with 0.1% Tween-20 for 1 h at room
temperature, the membranes were incubated with primary antibodies
at 4˚C overnight for the following proteins: FOXF1 (cat. no.
ab168383), Bcl-2 (cat. no. ab32124), Bax (cat. no. ab32503),
poly(ADP ribose) polymerase (PARP; cat. no. ab191217), cleaved PARP
(cat. no. ab32064), α-SMA (cat. no. ab108531), fibronectin (cat.
no. ab2413), collagen IV (cat. no. ab6586), E-cadherin (cat. no.
ab40772), N-cadherin (cat. no. ab76011), vimentin (cat. no.
ab92547), CDH11 (cat. no. ab151302) (all 1:1,000), β-catenin
(1:5,000; cat. no. ab32572), c-Myc (1:1,000; cat. no. ab32072),
c-jun (1:1,000; cat. no. ab40766), and β-actin (1:1,000; cat. no.
ab8227) (all Abcam). The membranes were washed and incubated with
horseradish peroxidase-labeled secondary antibody (Cell Signaling
Technology, Inc.) for 1 h at room temperature. Finally, bands were
visualized using an enhanced chemiluminescence detection system
(Merck KGaA) and immunoreactivity was detected using Image J
software (version 1.49; National Institutes of Health).
Statistical analysis
Statistical analysis was conducted using SPSS 22.0
(IBM Corp.) and GraphPad Prism 6 (GraphPad Software, Inc.). The
unpaired Student's t-test was used for the comparison between two
groups. The differences among multiple groups were analyzed using
one-way ANOVA with a post hoc Bonferroni multiple comparison test.
The data are presented as mean ± standard deviation of three
independent experiments. P<0.05 was considered to indicate a
statistically significant difference.
Results
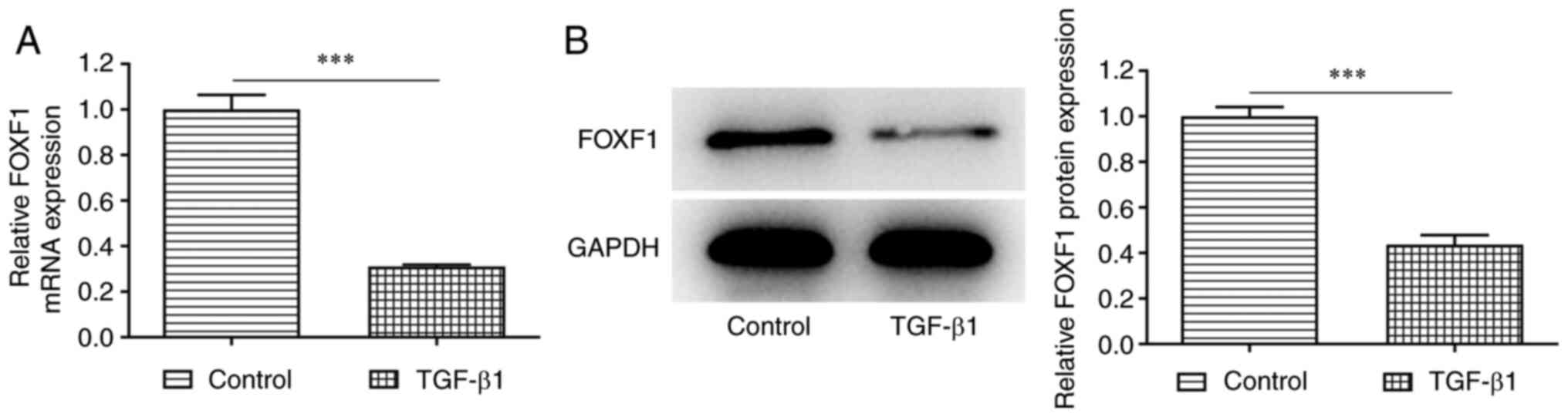
TGF-β1 induces downregulation of FOXF1
expression in BEAS-2B cells
To explore the role of FOXF1 in bronchial epithelial
cells, the expression levels of FOXF1 were initially investigated
in untreated and TGF-β1-treated BEAS-2B cells. RT-qPCR and western
blotting indicated a significant decrease in both mRNA and protein
expression levels of FOXF1 in TGF-β1-treated BEAS-2B cells compared
with the corresponding levels observed in the control cells
(Fig. 1). To conclude, FOXF1
expression was low in TGF-β1-treated BEAS-2B cells.
Upregulation of FOXF1 expression
reduces TGF-β1-induced damage and release of inflammatory factors
in BEAS-2B cells
To investigate the roles of FOXF1 in proliferation,
apoptosis and inflammation in BEAS-2B cells, FOXF1 was
overexpressed in untreated and TGF-β1-treated BEAS-2B cells. The
transfection efficiency was evaluated using RT-qPCR and western
blotting, which revealed that FOXF1 expression was significant
increased following transfection of pcDNA-FOXF1 compared with
transfection control (Fig. 2A and
B). Subsequently, the CCK-8 assay
was used to assess proliferation. The results indicated that TGF-β1
treatment significantly inhibited proliferation in BEAS-2B cells
compared with the control group; whereas overexpression of FOXF1
significantly promoted proliferation in TGF-β1-treated BEAS-2B
cells compared with the TGF-β1 + pcDNA3.1 negative control cells
(Fig. 2C). Furthermore, TGF-β1
treatment significantly increased the apoptotic rate of BEAS-2B
cells compared with the control, while the apoptotic rate of
TGF-β1-treated BEAS-2B cells transfected with pcDNA-FOXF1 was
significantly reduced compared with the transfection control
(Fig. 2D and E). In addition, TGF-β1 treatment
significantly reduced Bcl-2 expression and significantly enhanced
the expression levels of Bax and PARP compared with the control
group. Whereas a significant increase in Bcl-2 and significant
decreases of Bax and cleaved-PARP expression were observed after
transfection with pcDNA-FOXF1 compared with the TGF-β1 + pcDNA3.1
group (Fig. 2F). In addition,
FOXF1 overexpression significantly suppressed the levels of IL-6,
TNF-α and IL-1β in TGF-β1-treated BEAS-2B cells compared with
TGF-β1 + pcDNA3.1- treated cells (Fig.
2G). Overall, FOXF1 alleviated TGF-β1-stimulated viability
injury and apoptosis in BEAS-2B cells.
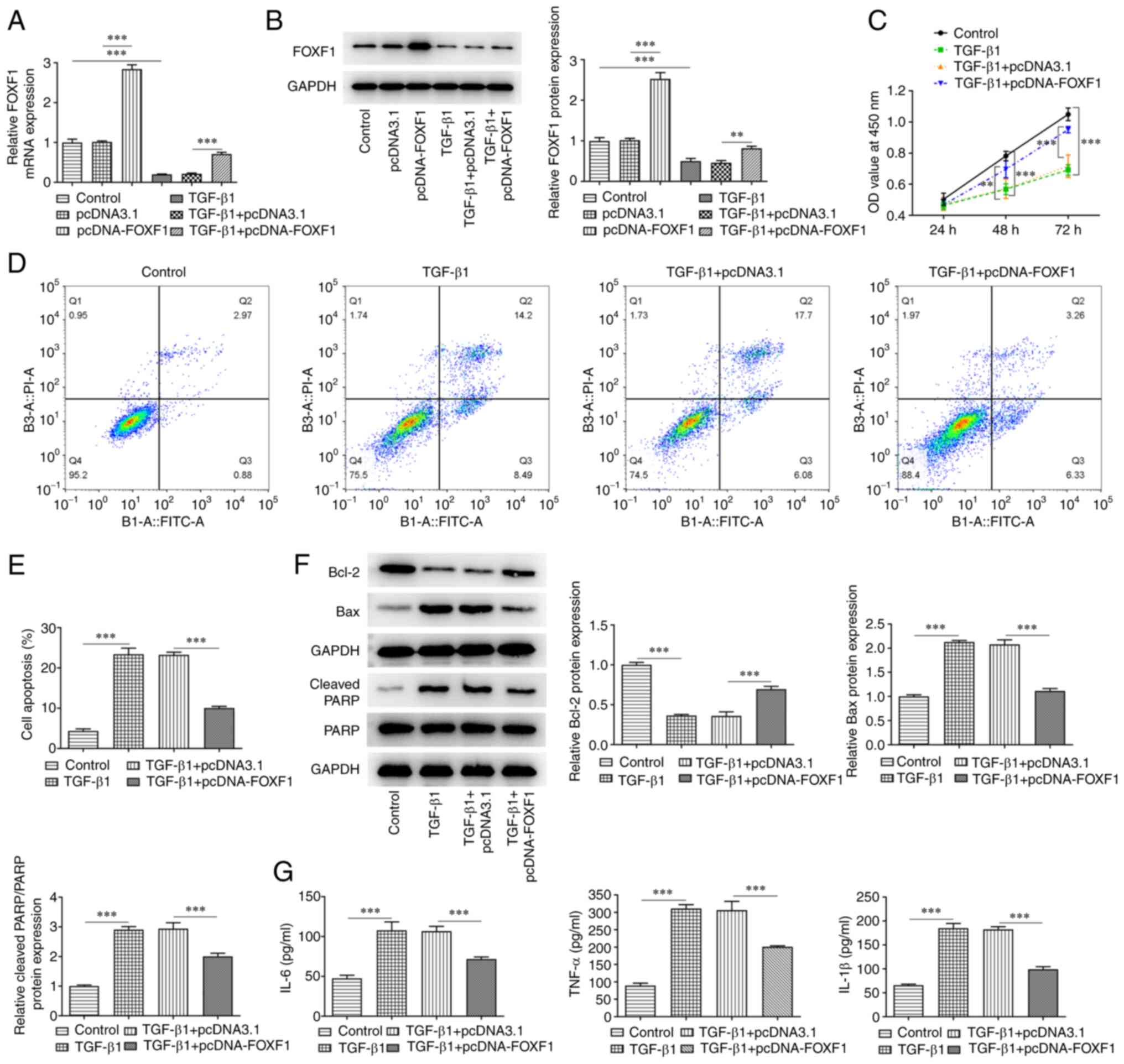 | Figure 2Upregulation of FOXF1 reduces
TGF-β1-induced damage and release of inflammatory factors in
BEAS-2B cells. (A) mRNA and (B) protein expression level of FOXF1
in TGF-β1-treated BEAS-2B cells were detected using reverse
transcription-quantitative PCR and western blotting, respectively.
(C) Cell Counting Kit-8 assay was used to assess proliferation.
Flow cytometry was carried out to (D) identify and (E) quantify
apoptosis in TGF-β1-treated BEAS-2B cells transfected with
pcDNA-FOXF1. (F) Western blotting was performed to detect the
protein expression level of Bcl-2, Bax, PARP and cleaved PARP. (G)
Levels of IL-6, TNF-α and IL-1β were detected using ELISA.
**P<0.01 and ***P<0.001. FOXF1,
forkhead box F1; TGF-β1, transforming growth factor β1; PI,
propidium iodide; FITC-A, FITC-Annexin-V; PARP, poly(ADP ribose)
polymerase; IL, interleukin; TNF-α, tumor necrosis factor-α. |
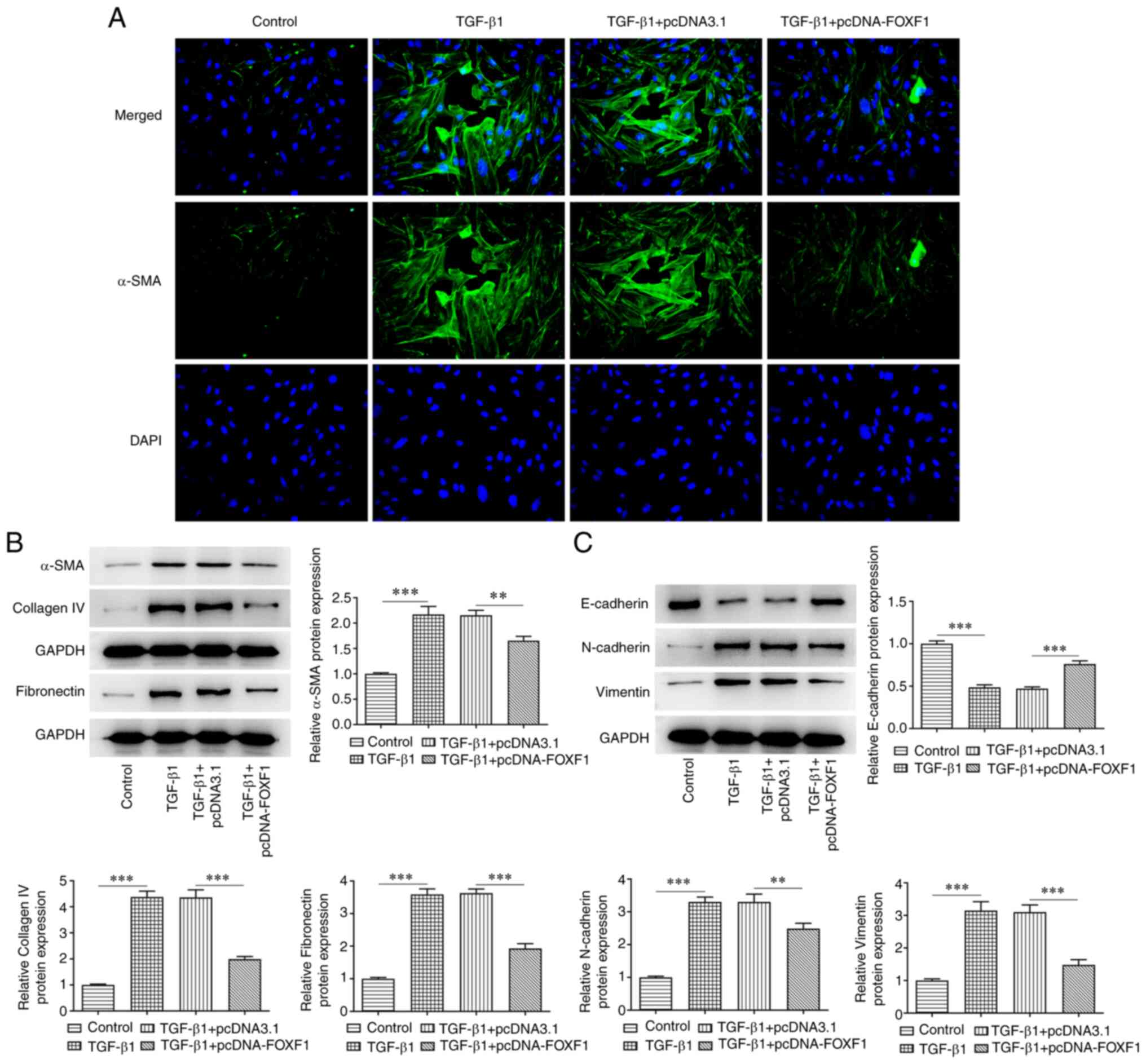
Upregulation of FOXF1 inhibits
fibrosis and epithelial-mesenchymal transition (EMT) in
TGF-β1-treated BEAS-2B cells
The biological role of FOXF1 was investigated with
regard to the induction of BEAS-2B cell fibrosis and EMT. Treatment
of the cells with TGF-β1 markedly enhanced the relative
fluorescence intensity of α-SMA in BEAS-2B cells, which was
subsequently reduced following FOXF1 overexpression (Fig. 3A). Moreover, TGF-β1 treatment
contributed to significantly increased levels of α-SMA, fibronectin
and collagen IV compared with the control group; while FOXF1
overexpression exhibited significant reversal of the expression of
these same markers compared with the TGF-β1 + pcDNA3.1 group
(Fig. 3B). Furthermore, compared
with the control group, significantly decreased levels of
E-cadherin and significantly increased levels of N-cadherin and
vimentin were observed in TGF-β1-treated BEAS-2B cells. FOXF1
overexpression was again able to significantly reverse the effects
of TGF-β1 treatment on BEAS-2B cells compared with the TGF-β1 +
pcDNA3.1 group (Fig. 3C). In
summary, FOXF1 suppressed TGF-β1-elicited fibrosis and EMT in
BEAS-2B cells.
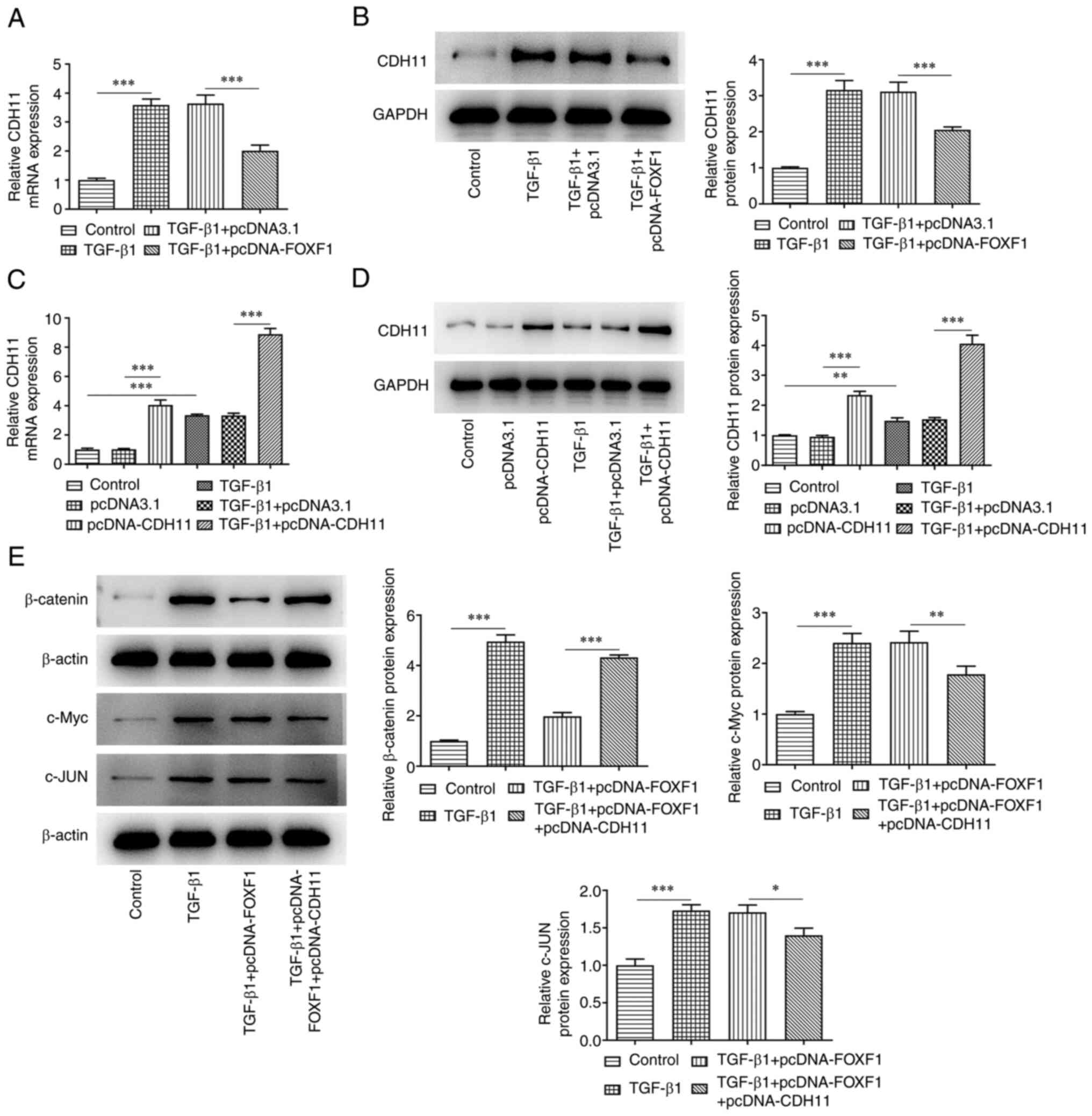
FOXF1 regulates the Wnt/β-catenin
signaling pathway by inhibiting CDH11 expression
The mechanism underlying the regulatory role of
FOXF1 in TGF-β1-treated BEAS-2B cells was investigated. As
presented in Fig. 4A and B, TGF-β1 treatment of the cells
significantly enhanced the levels of CDH11 compared with the
control group; whereas, FOXF1 overexpression led to a significant
reduction of CDH11 levels in TGF-β1-treated BEAS-2B cells compared
with the TGF-β1 + pcDNA3.1 group. Subsequently, CDH11 was
overexpressed in untreated and TGF-β1-treated BEAS-2B cells. The
transfection efficiency was evaluated using RT-qPCR and western
blotting and CDH11 expression was significantly increased in
transfected cells compared with the controls (Fig. 4C and D). Western blotting analysis indicated
that TGF-β1 treatment significantly increased the expression levels
of β-catenin, c-Myc, and c-Jun in BEAS-2B cells compared with the
control group. FOXF1 overexpression reversed the effects of TGF-β1
on β-catenin levels in TGF-β1 + pcDNA-FOXF1 group compared with
TGF-β1 + pcDNA3.1 group, though minimal changes were noted to the
expression levels of c-Myc and c-Jun. However, CDH11 overexpression
significantly reversed the effects of FOXF1 overexpression on the
expression levels of β-catenin, c-Myc and c-jun (Fig. 4E). Collectively, FOXF1
down-regulated CDH11 to inactivate Wnt/β-catenin signaling.
FOXF1 inhibits CDH11-mediated
Wnt/β-catenin signaling in TGF-β1-treated BEAS-2B cells
The CCK-8 assay revealed that CDH11 overexpression
and LiCl treatment significantly reduced the optical density values
of TGF-β1-treated BEAS-2B cells transfected with the FOXF1
overexpression plasmid compared with the TGF-β1 + pcDNA-FOXF1 group
(Fig. 5A). Moreover, a significant
increase was noted in the apoptotic rate of TGF-β1-treated BEAS-2B
cells co-transfected with pcDNA-FOXF1 and pcDNA-CDH11 or treated
with LiCl compared with that in cells transfected with pcDNA-FOXF1
alone (Fig. 5B and C). In addition, western blotting
indicated that pcDNA-CDH11 and LiCl treatment significantly
reversed the effects of FOXF1 overexpression on the protein
expression levels of Bcl-2, Bax, and cleaved PARP/PARP in
TGF-β1-treated BEAS-2B cells compared with cells transfected with
pcDNA-FOXF1 alone (Fig. 5D).
Furthermore, pcDNA-CDH11 and treatment with LiCl markedly enhanced
the relative fluorescence intensity of α-SMA in BEAS-2B cells
following decreased fluorescence after transfection with the FOXF1
overexpression vector (Fig. 5E).
In agreement with these findings, it was observed that the
previously significantly downregulated expression levels of α-SMA,
fibronectin and collagen IV were significantly elevated in
TGF-β1-treated BEAS-2B cells overexpressing FOXF1 following
transfection with pcDNA-CDH11 and treatment with LiCl compared with
cells transfected with pcDNA-FOXF1 alone (Fig. 6A). Furthermore, the significantly
increased levels of E-cadherin and the significantly reduced levels
of N-cadherin and vimentin noted in TGF-β1-treated BEAS-2B cells
overexpressing FOXF1 were significantly reversed following
transfection with pcDNA-CDH11 and treatment with LiCl compared with
cells transfected with pcDNA-FOXF1 alone (Fig. 6B). The latter treatments
significantly increased the levels of IL-6, TNF-α and IL-1β in
TGF-β1-treated BEAS-2B cells transfected with the FOXF1
overexpression plasmid (Fig. 6C).
Overall, CDH11 elevation of activation of Wnt/β-catenin signaling
reversed the impacts of FOXF1 on TGF-β1-treated BEAS-2B cells.
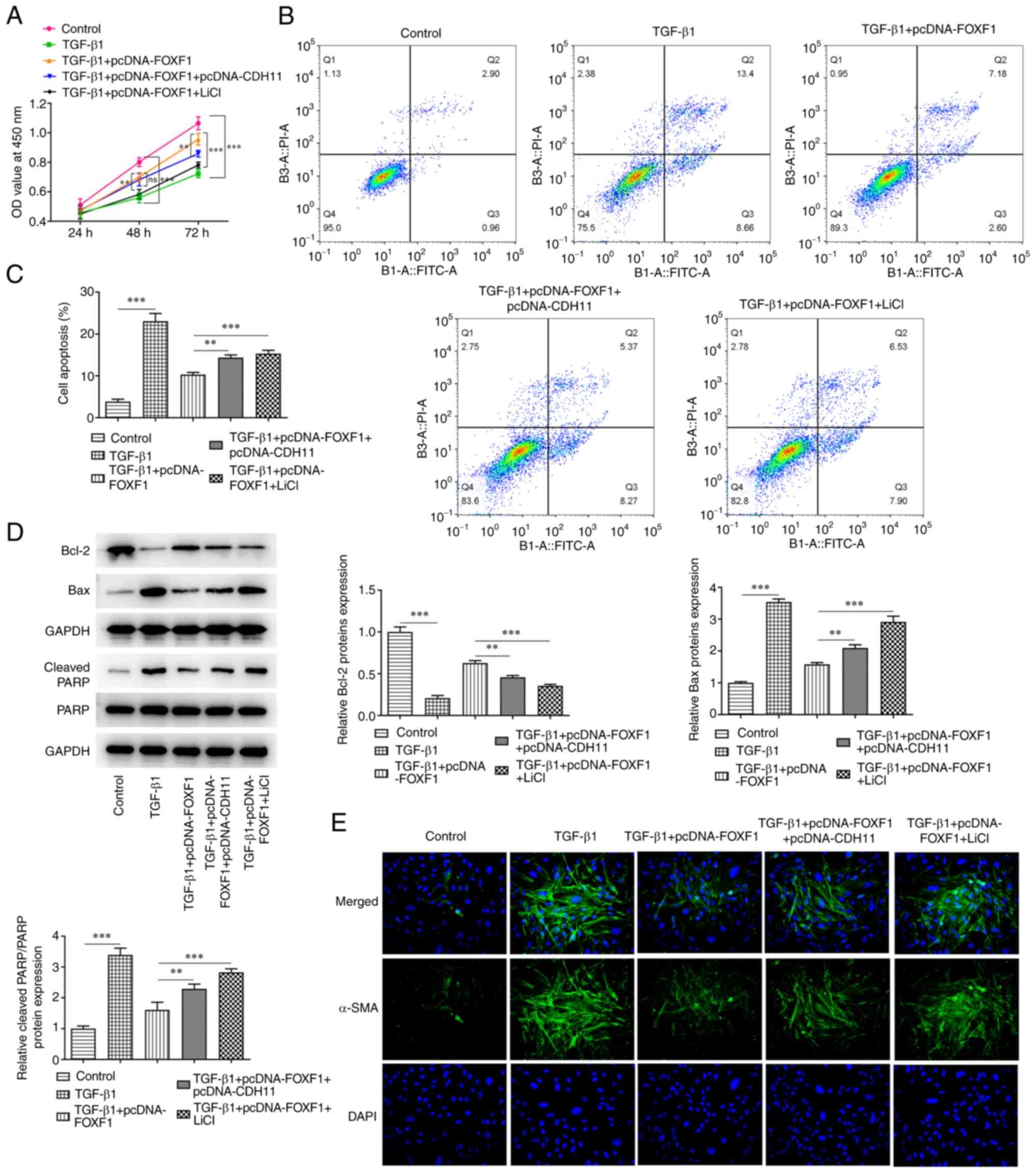 | Figure 5FOXF1 reduces TGF-β1-induced BEAS-2B
cell injury by inhibiting CDH11-mediated Wnt/β-catenin signaling.
(A) Cell Counting Kit-8 assay was used to detect cell
proliferation. Flow cytometry was performed to (B) identify and (C)
quantify apoptosis. (D) Western blotting was performed to detect
the protein expression level of Bcl-2, Bax, PARP and cleaved PARP.
(E) Immunofluorescence was used to detect the expression of α-SMA
(magnification, 200x). **P<0.01,
***P<0.001. FOXF1, forkhead box F1; TGF-β1,
transforming growth factor β1; OD, optical density; PI, propidium
iodide; FITC-A, FITC-Annexin-V; PARP, poly(ADP ribose) polymerase;
α-SMA, smooth muscle α-actin. |
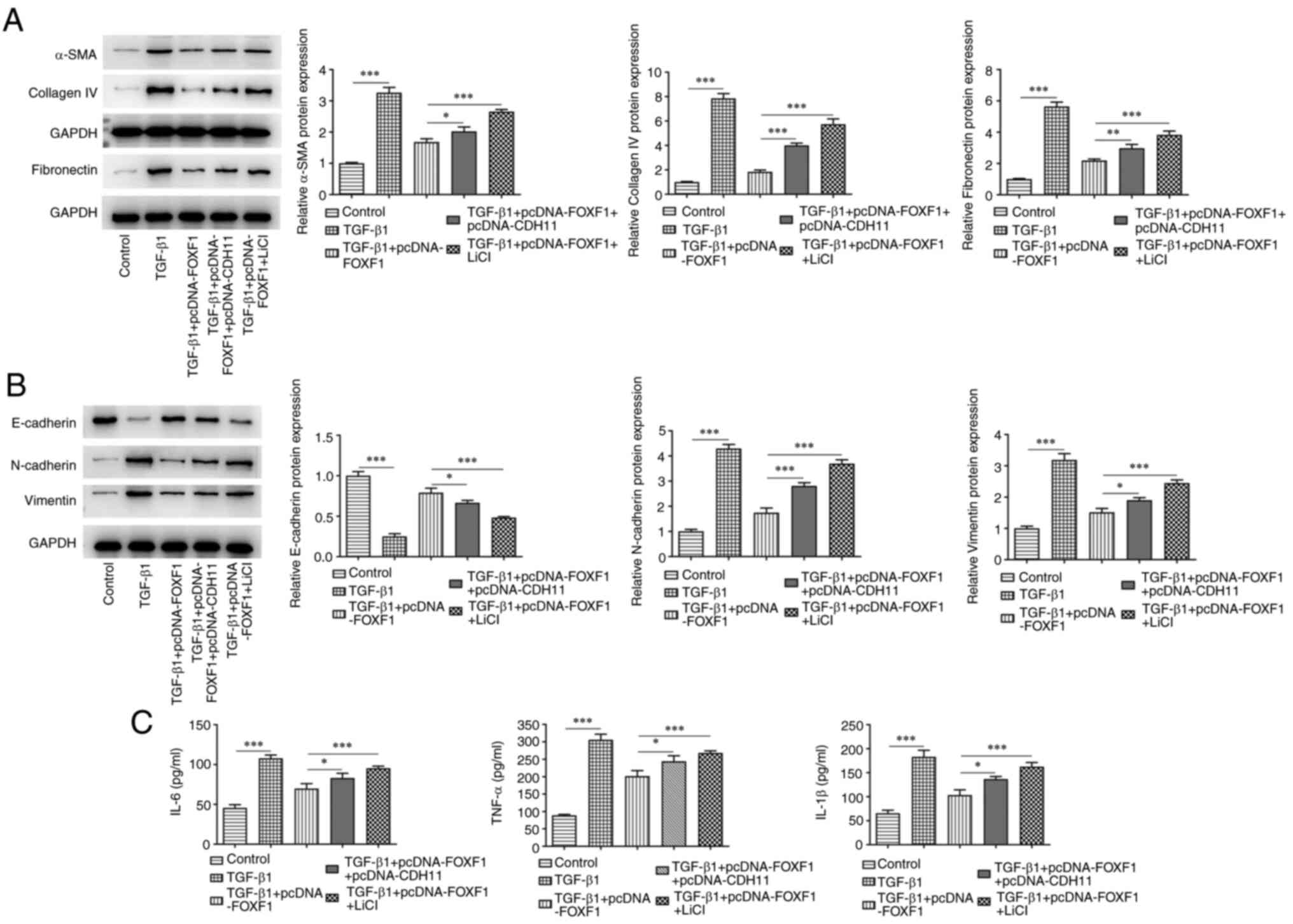 | Figure 6FOXF1 reduces TGF-β1-induced BEAS-2B
cell fibrosis, epithelial-mesenchymal transition and inflammation
by inhibiting CDH11-mediated Wnt/β-catenin signaling. (A) Western
blotting was performed to detect the protein levels of α-SMA,
fibronectin and collagen IV. (B) Protein expression levels of
E-cadherin, N-cadherin and vimentin were assessed using western
blotting. (C) Levels of IL-6, TNF-α and IL-1β were detected using
ELISA. *P<0.05, **P<0.01,
***P<0.001. FOXF1, forkhead box F1; TGF-β1,
transforming growth factor β1; CDH11, cadherin 11; α-SMA, smooth
muscle α actin; IL, interleukin; TNF-α, tumor necrosis
factor-α. |
Discussion
Asthma is a chronic airway inflammatory disease,
with children being mainly susceptible to the disease (17). The release of inflammatory
mediators can cause tissue damage and airway dysfunction (18). Various types of medical treatments
are available for childhood asthma; however, their efficacy remains
unsatisfactory (19-21).
In the present study, a mechanism of action of asthma was examined
that demonstrated FOXF1 may attenuate TGF-β1-induced bronchial
epithelial cell injury by inhibiting CDH11-mediated Wnt/β-catenin
signaling.
FOXF1 plays an important role in regulating lung
development and lung injury (22).
A previous study has shown that FOXF1 stimulates pulmonary
angiogenesis and alveolation during recovery from neonatal
hyperoxia injury (23). In
addition, FOXF1 maintains endothelial barrier function and prevents
edema following lung injury (15).
A previous study has shown that FOXF1 protects against BEAS-2B
apoptosis and oxidative stress induced by paraquat (22). Therefore, it is speculated that
FOXF1 is involved in the regulation of airway epithelial cell
injury induced by asthma. In the present study, TGF-β1 treatment
induced the downregulation of FOXF1 expression in BEAS-2B cells and
FOXF1 overexpression reduced TGF-β1-induced apoptosis by increasing
Bcl-2 expression, reducing the levels of Bax and cleaved PARP and
inhibiting the release of inflammatory factors IL-6, TNF-α and
IL-1β. In addition, FOXF1 overexpression suppressed the production
of α-SMA, fibronectin and collagen IV to reduce TGF-β1-induced
fibrosis; it also limited EMT by enhancing the expression of
E-cadherin while decreasing the levels of N-cadherin and vimentin.
These findings are in line with previous results demonstrating that
the FOXF1 transcription factor can promote lung regeneration
following partial pneumonectomy and can induce fetal lung
mesenchymal cell proliferation and promote lung morphogenesis
(24,25).
CDH11 is a member of the cadherin family of
proteins, with its gene located on chromosome 16q22.1(26). Previous studies have reported that
CDH11 serves key roles in the occurrence and development of several
diseases, including tumors and arthritis (27,28).
Wang et al (29)
demonstrated that microRNA-451a suppresses airway remodeling by
regulating CDH11 in an allergic asthma model in neonatal mice. In
addition, FOXF1 transcription factor has been demonstrated to
inhibit CDH11 expression (30). It
also inhibits pulmonary fibrosis by blocking CDH2-CDH11 cadherin
conversion in myofibroblasts (30). In the current study, it was
revealed that TGF-β1 treatment increased CDH11 expression in
BEAS-2B cells, whereas FOXF1 overexpression downregulated CDH11
expression following TGF-β1 stimulation. Furthermore, FOXF1
ameliorated TGF-β1-triggered viability injury, apoptosis, fibrosis
and EMT in BEAS-2B cells. CDH11 overexpression reversed the effects
of FOXF1 overexpression in TGF-β1-treated BEAS-2B cells. These data
indicated that FOXF1might function in TGF-β1-treated BEAS-2B cells
by regulating CDH11.
CDH11 has been revealed to induce cancer cell
apoptosis, suppress cell motility and invasion and inhibit cancer
progression via the Wnt/β-catenin pathway (31). It has also been shown to be a key
upstream regulator of the Wnt/β-catenin pathway (31). Dong et al (32) reported that upregulation of CDH11
in osteoarthritis activates the Wnt/β-catenin pathway. Huang et
al (33) demonstrated that
vitamin D alleviates airway remodeling in asthma by downregulating
the expression levels of the Wnt/β-catenin pathway proteins Wnt5a
and β-catenin. Furthermore, Yang et al (34) revealed that curcumin reduces lung
inflammation in a mouse model of asthma via the Wnt/β-catenin
signaling pathway. In the present study, TGF-β1 treatment
significantly increased the expression levels of β-catenin, c-Myc
and c-Jun in BEAS-2B cells. Also, the reduced β-catenin, c-Myc, and
c-Jun expression levels imposed by FOXF1 in TGF-β1-treated BEAS-2B
cells were reversed by CDH11. Moreover, FOXF1 ameliorated
TGF-β1-triggered viability injury, apoptosis, fibrosis and EMT in
BEAS-2B cells, which were restored by the activation of
Wnt/β-catenin signaling, which suggested that the Wnt/β-catenin
pathway regulated the biological response of TGF-β1-treated BEAS-2B
cells via the subsequent regulation of FOXF1/CDH11.
In summary, the results of the present study
suggested that FOXF1 overexpression increased BEAS-2B cell
proliferation and repressed apoptosis and inflammation. In
addition, overexpression of FOXF1 was demonstrated to reduce
fibrosis and EMT in TGF-β1-treated BEAS-2B cells. These protective
effects may rely on the regulation of the CDH11-mediated
Wnt/β-catenin pathway, which may provide a novel fundamental
insight into the pathogenesis of asthma and may be useful in
developing therapeutic strategies for the treatment of pediatric
asthma. Limitations of the present study included the use of only
one cell line and the use of an in vitro experiment to
explore the role of FOXF1. It is necessary to investigate these
effects in an in vivo asthma model to support the
conclusions of the current study.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
All data generated and/or analyzed during this study
are included in this published article.
Authors' contributions
QC and QT designed the study, drafted and revised
the manuscript. XL, LL and LW analyzed the data and searched the
literature. All authors performed the experiments. All authors read
and approved the final manuscript. QC and QT confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ntontsi P, Photiades A, Zervas E, Xanthou
G and Samitas K: Genetics and epigenetics in asthma. Int J Mol Sci.
22(2412)2021.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Mims JW: Asthma: Definitions and
pathophysiology. Int Forum Allergy Rhinol. 5 (Suppl 1):S2–S6.
2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Sockrider M and Fussner L: What is asthma?
Am J Respir Crit Care Med. 202:P25–P26. 2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Jones TL, Neville DM and Chauhan AJ:
Diagnosis and treatment of severe asthma: A phenotype-based
approach. Clin Med (Lond). 18:S36–S40. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Serebrisky D and Wiznia AL: Pediatric
asthma: A global epidemic. Ann Glob Health. 85(6)2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Devonshire AL and Kumar R: Pediatric
asthma: Principles and treatment. Allergy Asthma Proc. 40:389–392.
2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Azmeh R, Greydanus DE, Agana MG, Dickson
CA, Patel DR, Ischander MM and Lloyd RD Jr: Update in pediatric
asthma: Selected issues. Dis Mon. 66(100886)2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Jonsson H and Peng SL: Forkhead
transcription factors in immunology. Cell Mol Life Sci. 62:397–409.
2005.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Herman L, Todeschini AL and Veitia RA:
Forkhead transcription factors in health and disease. Trends Genet.
37:460–475. 2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Maiese K: Forkhead transcription factors:
Formulating a FOXO target for cognitive loss. Curr Neurovasc Res.
14:415–420. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Braeuer RR, Walker NM, Misumi K,
Mazzoni-Putman S, Aoki Y, Liao R, Vittal R, Kleer GG, Wheeler DS,
Sexton JZ, et al: Transcription factor FOXF1 identifies
compartmentally distinct mesenchymal cells with a role in lung
allograft fibrogenesis. J Clin Invest. 131(e14734)2021.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Milewski D, Shukla S, Gryder BE, Pradhan
A, Donovan J, Sudha P, Vallabh S, Pyros A, Xu Y, Barski A, et al:
FOXF1 is required for the oncogenic properties of PAX3-FOXO1 in
rhabdomyosarcoma. Oncogene. 40:2182–2199. 2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Mahlapuu M, Ormestad M, Enerbäck S and
Carlsson P: The forkhead transcription factor Foxf1 is required for
differentiation of extra-embryonic and lateral plate mesoderm.
Development. 128:155–166. 2001.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Stankiewicz P, Sen P, Bhatt SS, Storer M,
Xia Z, Bejjani BA, Ou Z, Wiszniewska J, Driscoll DJ, Maisenbacher
MK, et al: Genomic and genic deletions of the FOX gene cluster on
16q24.1 and inactivating mutations of FOXF1 cause alveolar
capillary dysplasia and other malformations. Am J Hum Genet.
84:780–791. 2009.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Cai Y, Bolte C, Le T, Goda C, Xu Y, Kalin
TV and Kalinichenko VV: FOXF1 maintains endothelial barrier
function and prevents edema after lung injury. Sci Signal.
9(ra40)2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Gans MD and Gavrilova T: Understanding the
immunology of asthma: Pathophysiology, biomarkers, and treatments
for asthma endotypes. Paediatr Respir Rev. 36:118–127.
2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Padem N and Saltoun C: Classification of
asthma. Allergy Asthma Proc. 40:385–388. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Abul MH and Phipatanakul W: Severe asthma
in children: Evaluation and management. Allergol Int. 68:150–157.
2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Hoch HE, Houin PR and Stillwell PC: Asthma
in children: A brief review for primary care providers. Pediatr
Ann. 48:e103–e109. 2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Guilbert TW, Bacharier LB and Fitzpatrick
AM: Severe asthma in children. J Allergy Clin Immunol Pract.
2:489–500. 2014.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zheng F, Liu T, Zhu J, Xie Y, Wu L and Lin
Z: FoxF1 protects rats from paraquat-evoked lung injury following
HDAC2 inhibition via the microRNA-342/KLF5/IκB/NF-κB p65 axis. Exp
Cell Res. 395(112208)2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Bolte C, Ustiyan V, Ren X, Dunn AW,
Pradhan A, Wang G, Kolesnichenko OA, Deng Z, Zhang Y, Shi D, et al:
Nanoparticle delivery of proangiogenic transcription factors into
the neonatal circulation inhibits alveolar simplification caused by
hyperoxia. Am J Respir Crit Care Med. 202:100–111. 2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Bolte C, Flood HM, Ren X, Jagannathan S,
Barski A, Kalin TV and Kalinichenko VV: FOXF1 transcription factor
promotes lung regeneration after partial pneumonectomy. Sci Rep.
7(10690)2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Ustiyan V, Bolte C, Zhang Y, Han L, Xu Y,
Yutzey KE, Zorn AM, Kalin TV, Shannon JM and Kalinichenko VV: FOXF1
transcription factor promotes lung morphogenesis by inducing
cellular proliferation in fetal lung mesenchyme. Dev Biol.
443:50–63. 2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Ruan W, Pan R, Shen X, Nie Y and Wu Y:
CDH11 promotes liver fibrosis via activation of hepatic stellate
cells. Biochem Biophys Res Commun. 508:543–549. 2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Yang Z, Yan C, Yu Z, He C, Li J, Li C, Yan
M, Liu B, Wu Y and Zhu Z: Downregulation of CDH11 promotes
metastasis and resistance to paclitaxel in gastric cancer cells. J
Cancer. 12:65–75. 2021.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Assefnia S, Dakshanamurthy S, Auvil JM,
Hampel C, Anastasiadis PZ, Kallakury B, Uren A, Foley DW, Brown ML,
Shapiro L, et al: Cadherin-11 in poor prognosis malignancies and
rheumatoid arthritis: Common target, common therapies. Oncotarget.
5:1458–1474. 2014.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Wang T, Zhou Q and Shang Y: MiRNA-451a
inhibits airway remodeling by targeting Cadherin 11 in an allergic
asthma model of neonatal mice. Int Immunopharmacol.
83(106440)2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Black M, Milewski D, Le T, Ren X, Xu Y,
Kalinichenko VV and Kalin TV: FOXF1 inhibits pulmonary fibrosis by
preventing CDH2-CDH11 cadherin switch in myofibroblasts. Cell Rep.
23:442–458. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Chen X, Xiang H, Yu S, Lu Y and Wu T:
Research progress in the role and mechanism of Cadherin-11 in
different diseases. J Cancer. 12:1190–1199. 2021.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Dong J, Li L, Fang X and Zang M:
Exosome-encapsulated microRNA-127-3p released from bone
marrow-derived mesenchymal stem cells alleviates osteoarthritis
through regulating CDH11-mediated Wnt/β-catenin pathway. J Pain
Res. 14:297–310. 2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Huang Y, Wang L, Jia XX, Lin XX and Zhang
WX: Vitamin D alleviates airway remodeling in asthma by
down-regulating the activity of Wnt/β-catenin signaling pathway.
Int Immunopharmacol. 68:88–94. 2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Yang X, Lv JN, Li H, Jiao B, Zhang QH,
Zhang Y, Zhang J, Liu YQ, Zhang M, Shan H, et al: Curcumin reduces
lung inflammation via Wnt/β-catenin signaling in mouse model of
asthma. J Asthma. 54:335–340. 2017.PubMed/NCBI View Article : Google Scholar
|