Introduction
Granulocyte colony-stimulating factor (G-CSF)
regulates the survival, proliferation, and differentiation of
neutrophils and activates and stimulates the functions of mature
neutrophils (1). A broad consensus
has emerged regarding the clinical utility of G-CSF in neutropenic
conditions resulting from chemotherapy (2). G-CSF is a hematopoietic cytokine that
plays a role in hematopoietic stem and progenitor cell (HSPC)
mobilization from the bone marrow niches to the peripheral blood
stream (3). Both hematopoietic
tissue and bone marrow are located inside the bone, and the
functional interaction between the bone and hematopoietic systems
was recently revealed (3-6).
Bone remodeling is the ‘replacement of old bone with
new bone’ and is based on the coordination of osteoclasts and
osteoblasts (7,8). These cells sequentially perform the
resorption of old damaged bone and the formation of new bone,
respectively (7,8). Osteocytes, which are derived from
osteoblasts, are additional cells involved in bone remodeling. They
are distributed throughout the bone matrix within a network of
lacunae and canaliculi to communicate indirectly with bone-related
cells (7,8). If the network of osteocytes is poorly
connected, bone remodeling may be disturbed. G-CSF affects bone
metabolism by inhibiting osteoblast activity (6) and increasing osteoclast activity
(9,10) in the bone marrow. G-CSF treatment
in patients with severe congenital neutropenias was shown to induce
osteopenia or osteoporosis (11,12).
Soshi et al (13) reported
that the bone mineral density of lumbar vertebrae and femora was
significantly decreased in G-CSF-treated rats. Wu et al
(14) investigated the effects of
G-CSF administration on the skeleton. The test data show that mice
treated with G-CSF have significantly lower stiffness, Young's
modulus, and fracture strength of the bone in their femurs when
compared with the controlled mice treated with saline. We focused
on and investigated G-CSF as one of the regulatory factors of bone
remodeling.
The effects of G-CSF on bone remodeling, including
the jawbone, in vivo remain unclear. We focused on the
effect of G-CSF administration on bone metabolism of the jawbone.
Most dentists and oral and maxillofacial surgeons face the problems
of delayed healing of the extraction socket due to a decrease in
bone turnover, which is associated with serious difficulties in the
maintenance of patient oral health. Moreover, although
osteonecrosis of the jaw associated with medications, including
bisphosphonates and some anticancer drugs (15), is well known, failure of tooth
extraction socket healing occurs frequently. We previously reported
that bisphosphonate therapy induced delayed healing of the
extraction socket as the cumulative administration period was
prolonged (16). The aim of the
present study was to investigate the effect of G-CSF administration
on the bone healing of tooth extraction sockets.
Materials and methods
Animal handling
The present study was approved by the ethics
committee of the Hyogo College of Medicine (Hyogo, Japan) (approval
number 19-032). Five-week-old male C57BL/6J mice were obtained from
SLC Japan. The mice were housed in a light- and
temperature-controlled environment. Food and water were available
ad libitum.
Agents
Granulocyte colony-stimulating factor (G-CSF:
Filgrastim) was purchased from KYOWA KIRIN Co. Ltd..
Experimental methods and design
Following a week of acclimatization, the now
6-week-old male mice weighting 23-27 g were randomly divided into
two groups (n=8 each) and treated with or without G-CSF
(experimental group: G-CSF and control group: saline solution).
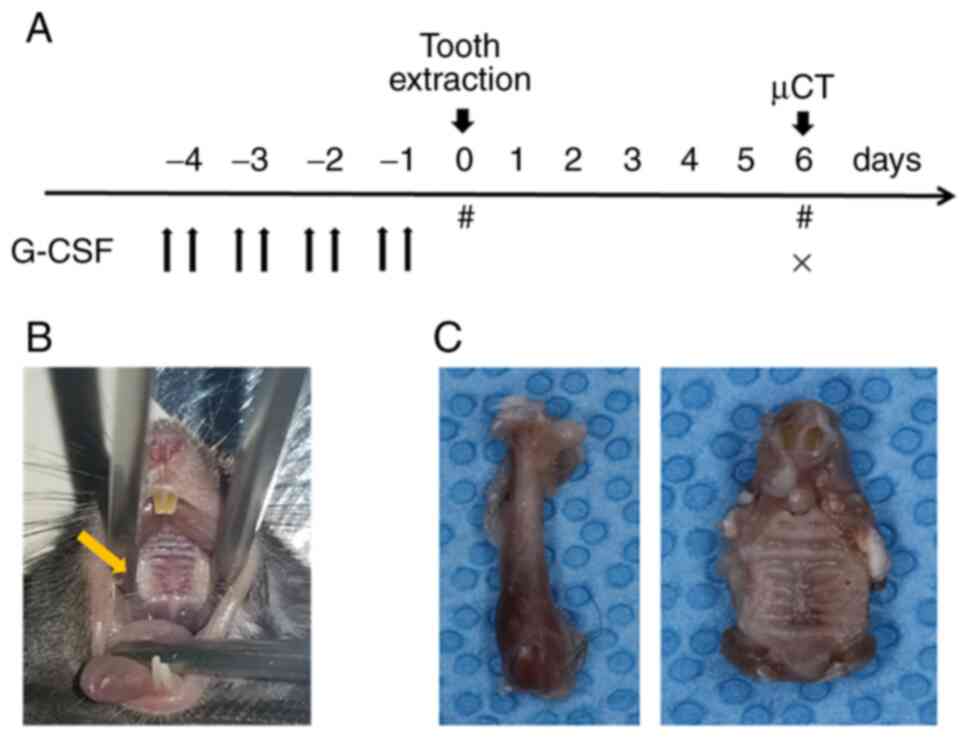
G-CSF (250 µg/kg/day) was injected intraperitoneally every 12 h for
4 days prior to tooth extraction (Fig.
1A). Extraction of the unilateral maxillary first molar was
performed using a spoon excavator under anesthesia with 2%
isoflurane (Pfizer Japan, Inc.) (Fig.
1B). The G-CSF dosage and duration of administration were based
on the protocols described previously by Asada et al
(4). The femurs and maxillae were
harvested at 6 days after the extraction for histological and
histochemical studies (Fig. 1C).
Two or three mice of each group were difficult to analyze due to
tooth root fracture, alveolar bone fracture, or other
complications. Consequently, the minimum number of animals was five
in each group (n≥5 each). At each respective specified time point
of each group, eight mice per group were anesthetized and 300-700
µl blood was harvested via intracardiac puncture for peripheral
blood analysis. For euthanasia, an overdose of sodium pentobarbital
(100 mg/ml) was administered intraperitoneally, and decapitation
was performed using scissors.
Determination of the number of
leukocytes in blood
Blood was harvested via intracardiac puncture under
anesthesia with 2% isoflurane at 1 and 7 days subsequent to 4 days
of G-CSF injection. The number of leukocytes in the blood was
counted as an indicator of HSPC mobilization. Following leukocyte
staining by peroxidase at 70˚C, blood cell components other than
leukocytes were excluded, and their samples were measured by flow
cytometry using the Advia 2120i (Siemens Healthineers).
Microcomputed tomography analysis and
bone morphometry of femurs and maxillae
Microcomputed tomography (µCT) scanning was
performed to measure bone morphological indices of mouse femurs and
maxillae as described previously (14). The femurs and maxillae in each
group were harvested, stored in 70% ethanol at 4˚C, and analyzed
using a micro (µ)-CT scanner (Scan Xmate-L090; Comscan Techno Co.,
Ltd.). Scanning was conducted at 75 kV and 105 mA with a spatial
resolution of approximately 9.073 mm/pixel. For the morphometric
analysis, the bone volume fraction (BV/TV), trabecular thickness
(Tb.Th), trabecular number (Tb.N), and trabecular separation
(Tb.Sp) were determined using TRI/3D-BON software (RATOC System
Engineering Co., Ltd.). The femoral region of interest (ROI) was
the distal femoral metaphysis, and the maxillary ROI was the
alveolar septum of the first molar.
Assays for bone metabolism markers in
serum
Blood was harvested via intracardiac puncture under
anesthesia at 1 and 7 days subsequent to 4 days of G-CSF injection,
and serum was collected after 30 min at room temperature and spun
down at 3,000 rpm for 10 min. The serum samples were used to
measure serum markers of bone resorption (serum band 5 of
tartrate-resistant acid phosphatase; TRACP-5b) and bone formation
(procollagen type 1 N-terminal propeptide; P1NP). TRACP-5b and P1NP
were determined by enzyme-linked immunosorbent assays (Mouse
ACP5/TRAP ELISA Kit, cat. no. LS-F40333-1, LSBio, Inc.; P1NP assay,
cat. no. SEA957Hu, Cloud-Clone Corp., respectively), according to
the manufacturers' protocols. All samples were tested in duplicate
within each assay. The number of mice analyzed in each group was
dependent on the amount of collected blood (n≥5 each).
Histopathology and
immunohistochemistry
The mouse maxillae were immediately placed in 10%
neutral buffered formalin for 24 h and decalcified in 10%
ethylenediaminetetraacetic acid at room temperature for 2 weeks.
Paraffin sections of 4 µm thickness were cut using conventional
methods and stained with hematoxylin and eosin (H&E). Stained
sections were photomicrographed using the Olympus microscope (BX53,
Japan), and were histomorphometrically analyzed using NIH ImageJ
(version 1.47) for quantification of the neoplastic bone area,
except for two cases that were associated with the presence of root
fragments in the extraction socket.
Tartrate-resistant acid phosphatase (TRAP) staining
of the mouse maxillae was performed as described previously
(15). Briefly, samples were
placed in 0.2 M acetate buffer (0.2 M sodium acetate and 50 mM L(+)
tartaric acid in double-distilled water, pH 5.0) for 20 min at room
temperature. The sections were subsequently incubated with 0.5
mg/ml naphthol AS-MX phosphate (Sigma-Aldrich Co.) and 1.1 mg/ml
Fast Red TR Salt (Sigma) in 0.2 M acetate buffer for 1 to 4 h at
37˚C until the osteoclasts appeared bright red. Osteoclasts were
identified as multinucleated TRAP-positive cells. The number of
TRAP-positive cells were counted in the maxillae, and expressed as
the number/mm2.
Silver-stained nucleolar organizer regions (AgNOR)
staining was performed using a silver staining solution prepared by
combining silver nitrate (2 volumes of 50% aqueous solution) (Wako
Pure Chemical Industries, Ltd.) and formic acid (1 volume of 1%
solution containing 2% gelatin) (14). The number of canaliculi per
osteocyte lacuna (N.Ot.Ca/Ot.Lc.) was counted in 10 cells in four
randomly selected non-overlapping defined ROIs at 400x.
The localization of chemokine (C-X-C motif) ligand
12 (stromal cell-derived factor 1) (CXCL12/SDF-1) in femurs and
maxillae was investigated. CXCL12/SDF-1 was detected in
immersion-fixed paraffin-embedded sections of mouse femurs and
maxillae using anti-CXCL12/SDF-1 mouse monoclonal antibody (cat.
no. MAB350, R&D Systems) at 25 µg/ml overnight at 4˚C. Tissue
was stained using the HR-conjugated anti-mouse IgG goat antibody
(cat. no. 414322, Nichirei Corporation) for 10 min and
counterstained with hematoxylin.
Calcein labeling
Nine and 2 days before euthanasia, five mice in each
group were given an intraperitoneal calcein (10 mg/kg) injection
for double labeling. The calcein double labels were analyzed with
an excitation wavelength of 485 nm and an emission wavelength of
510 nm using the Olympus microscope. The mineral apposition rate
(MAR, µm/day) was defined as the distance between the midpoints of
the double label divided by the number of days between the calcein
injections (17-19).
Statistical analysis
All data are expressed as mean ± standard deviation.
Statistical analysis was performed using the Mann-Whitney U test to
identify significant differences for two independent group
comparisons. P<0.05 was considered to indicate a statistically
significant difference.
Results
Body weight
There were no significant differences in body weight
between the control and G-CSF groups during the experimental period
(data not shown).
Determination of the number of
leukocytes in blood
The number of leukocytes in the G-CSF group was
significantly higher than the control group a day after the 4-day
G-CSF administration, whereas no significant difference was
detected at 7 days after the G-CSF administration (Table I). The G-CSF decreased CXCL12
protein expression in the bone marrow, and the disruption of
CXCL12/CXCR4 signaling is a key step in G-CSF-induced HPC
mobilization (4-6).
In the control group, CXCL12/SDF-1-positive cells were found in the
femur and maxillae (Fig. S1A and
C), the number of which was
reduced by G-CSF treatment (Fig.
S1B and D).
 | Table INumber of blood leucocytes. |
Table I
Number of blood leucocytes.
| A, 1 day after
G-CSF 4 days administration |
|---|
| Group | Leucocytes,
x102/µl blood |
|---|
| Control | 9.38±4.00 |
| G-CSF |
27.33±10.63a |
| B, 7 days after
G-CSF 4 days administration |
| Group | Leucocytes,
x102/µl blood |
| Control | 14.13±6.29 |
| G-CSF | 15.25±11.81 |
µCT examination and bone morphometry
of the femur and maxillae
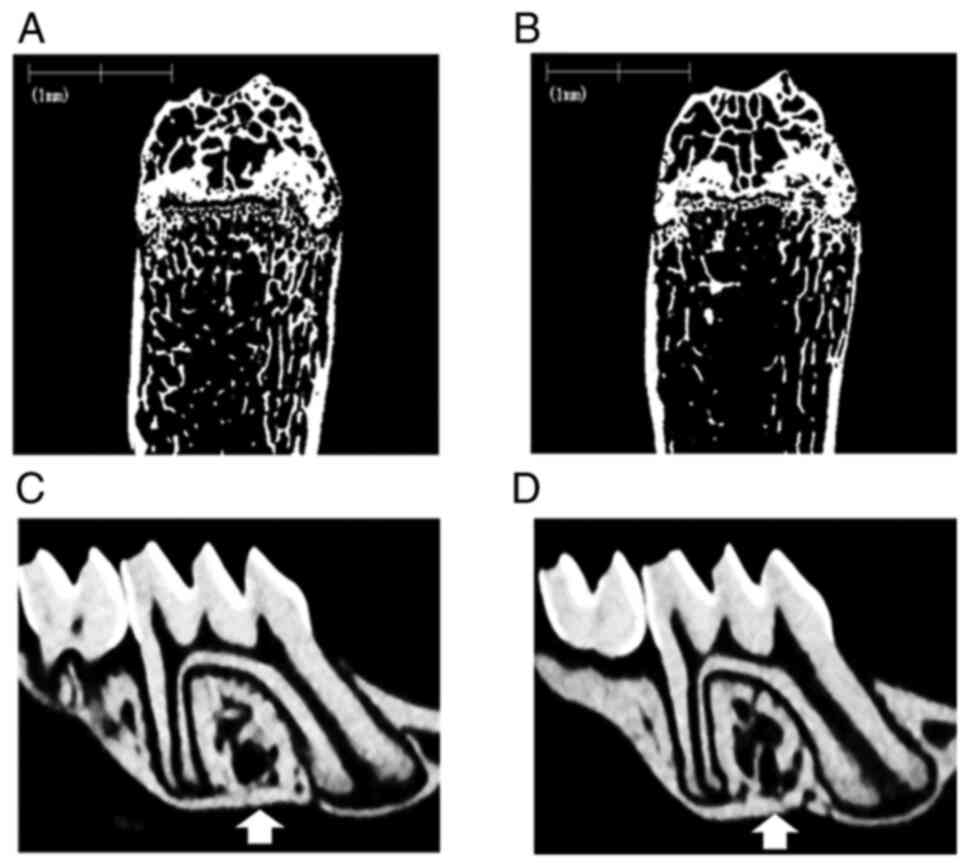
The most typical µ-CT images of the femur and
maxillae for the G-CSF and control groups are shown in Fig. 2A-D. The G-CSF group exhibited a
loss of trabecular bone in the femur (Fig. 2B) and the alveolar septum (Fig. 2D) compared with the control group
(Fig 2A and C). Bone morphometric analysis of the
proximal femurs and the alveolar septa of the first molar in the
maxillae was performed to determine the BV/TV, Tb.Th, Tb.N, and
Tb.Sp (Table II). Significant
differences in the BV/TV, Tb.N and Tb.Sp of the proximal femurs
were observed between the G-CSF and control groups. Significant
differences in the BV/TV, Tb. N and Tb.Sp of the alveolar septa
were observed between the G-CSF and control groups. G-CSF
administration resulted in clear trabecular bone loss in not only
the femur, but also the alveolar bone in the maxilla.
| Table IIBone histomorphometric analysis. |
Table II
Bone histomorphometric analysis.
| A, Distal
femur |
|---|
| | Parameter |
|---|
| Group | BV/TV, % | Tb.Th, µm | Tb.N, 1/mm | Tb.Sp, µm |
|---|
| Control | 17.84±1.64 | 38.46±2.21 | 4.69±0.20 | 175.67±10.34 |
| G-CSF |
11.54±1.37a | 37.64±2.35 |
3.06±0.20a |
290.31±22.09a |
| B, Alveolar septa
of first molar in the maxillae |
| | Parameter |
| Group | BV/TV, % | Tb.Th, µm | Tb.N, 1/mm | Tb.Sp, µm |
| Control | 45.13±8.02 | 62.73±8.57 | 7.25±1.06 | 77.98±19.89 |
| G-CSF |
29.66±5.01a | 53.82±8.58 |
5.60±1.08a |
131.10±36.24a |
Bone metabolism markers in serum
There was no significant difference in serum P1NP
levels, a marker of bone formation, between the control and G-CSF
groups 1 day after the 4-day G-CSF administration. By contrast, 7
days after the 4-day G-CSF administration, serum P1NP levels were
significantly lower in the G-CSF group as compared with the levels
in the control group (control group, 2.16±0.77 ng/ml; G-CSF group
1.33±0.68 ng/ml; Table III).
Serum TRACP-5b levels, a marker of bone resorption, were not
significantly different in the G-CSF group as compared with the
control group (Table IV). These
data are consistent with the morphometry results and suggest that
G-CSF administration progressively decreases the trabecular bone
volume.
| Table IIISerum P1NP. |
Table III
Serum P1NP.
| A, 1 day after
G-CSF 4 days administration |
|---|
| Group | P1NP, ng/ml |
|---|
| Control | 2.19±0.15 |
| G-CSF | 2.62±0.52 |
| B, 7 days after
G-CSF 4 days administration |
| Group | P1NP, ng/ml |
| Control | 2.16±0.77 |
| G-CSF |
1.33±0.68a |
| Table IVSerum TRAP-5b. |
Table IV
Serum TRAP-5b.
| A, 1 day after
G-CSF 4 days administration |
|---|
| Group | TRAP-5b, ng/ml |
| Control | 1.11±0.88 |
| G-CSF | 2.59±2.69 |
| B, 7 days after
G-CSF 4 days administration |
| Group | TRAP-5b, ng/ml |
| Control | 2.02±2.17 |
| G-CSF | 2.63±2.27 |
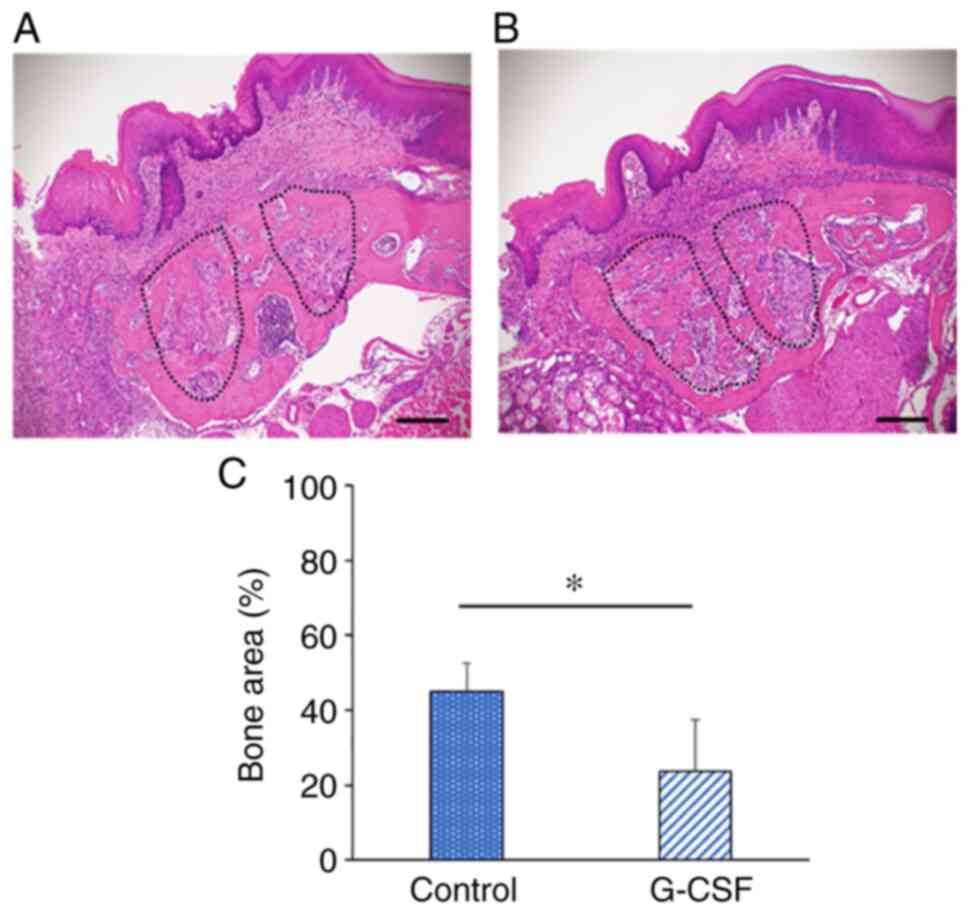
Histological evaluation
Sections of maxilla including the tooth extraction
region were stained with H&E and examined histologically at 6
days after tooth extraction in the control and G-CSF groups. The
tooth extraction socket displayed a tendency to be filled with new
bone in the control group (Fig.
3A). By contrast, fibrous connective tissues and immature bone
were observed in the extraction socket and bone healing was delayed
in the G-CSF group compared with the control group (Fig. 3B). The bone area in the extraction
socket was significantly smaller in the G-CSF group (23.6%) than in
the control group (45.1%) (Fig.
3C). The histological appearance of the G-CSF treated mice bony
regenerate was decreased.
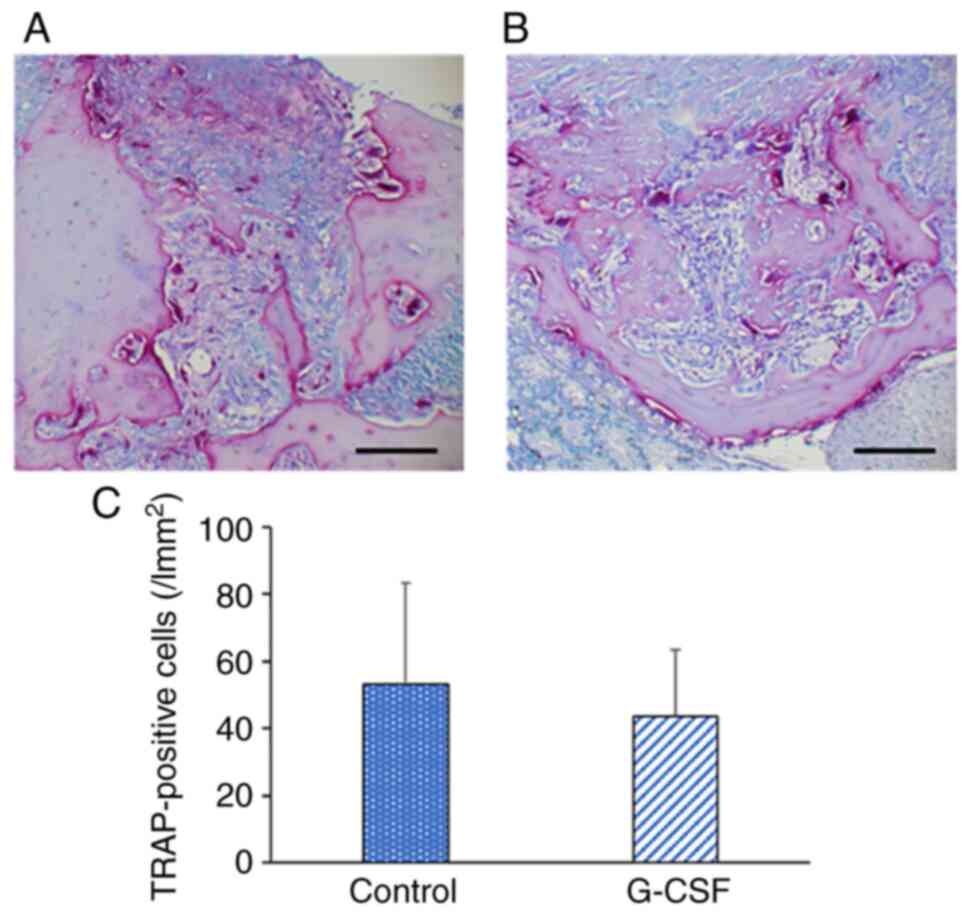
Osteoclast activity
TRAP-positive osteoclasts were present on the bone
surface in the alveolar bone in the control and G-CSF groups at 6
days after the extraction (Fig. 4A
and B). There was no significant
difference in the number of TRAP-positive cells of the alveolar
bone between these two groups (Fig.
4C).
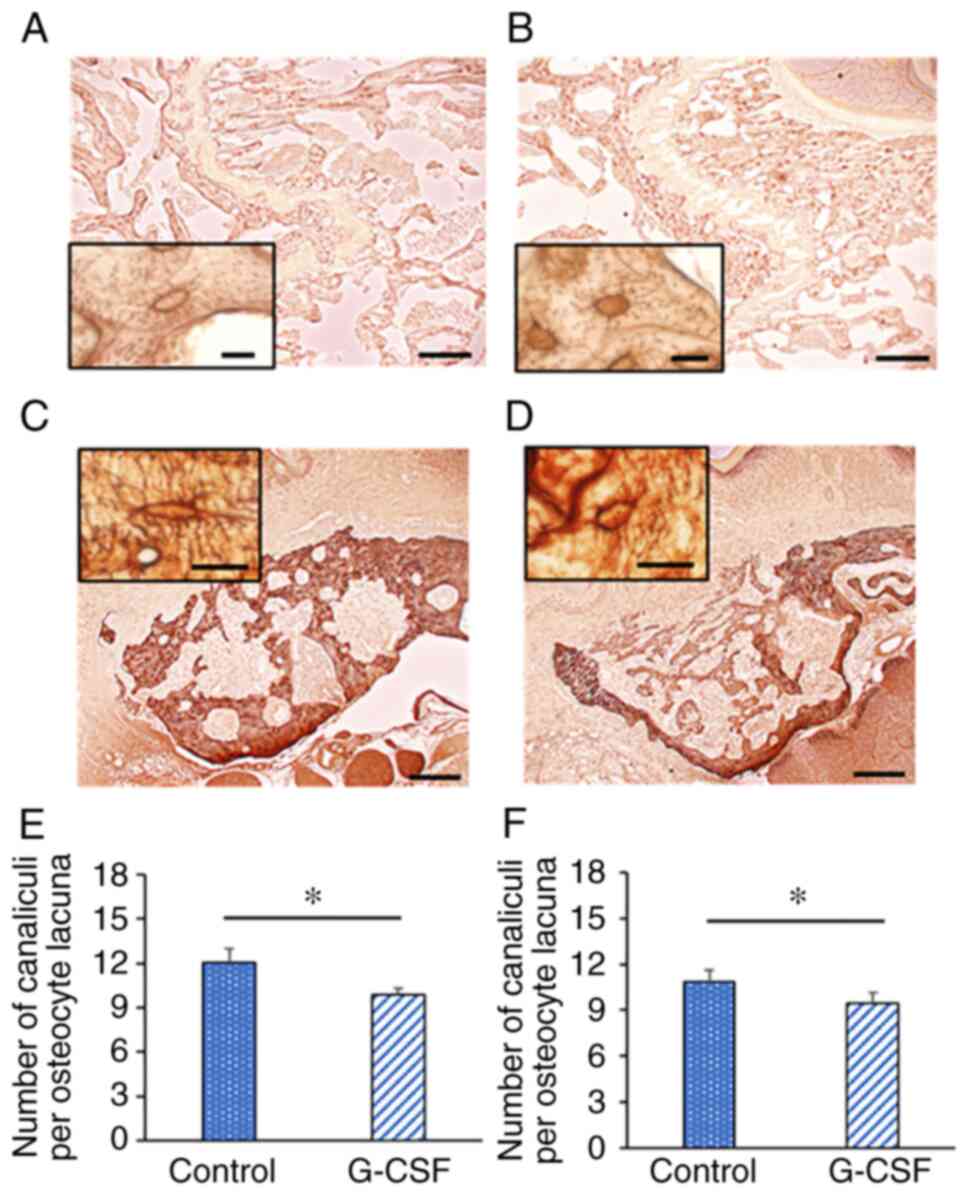
Osteocytic canalicular morphology
To investigate morphological changes in the femur
and maxilla, we performed AgNOR staining (Fig. 5A-E). The number of canaliculi per
osteocyte in the femur and maxilla was significantly decreased by
G-CSF administration (N.Ot.Ca/Ot.Lc. femur: control group,
12.04±0.96; G-CSF group, 9.87±0.45; Fig. 5E, N.Ot.Ca/Ot.Lc. maxilla: control
group, 10.86±0.74; G-CSF group, 9.43±0.69; Fig. 5F).
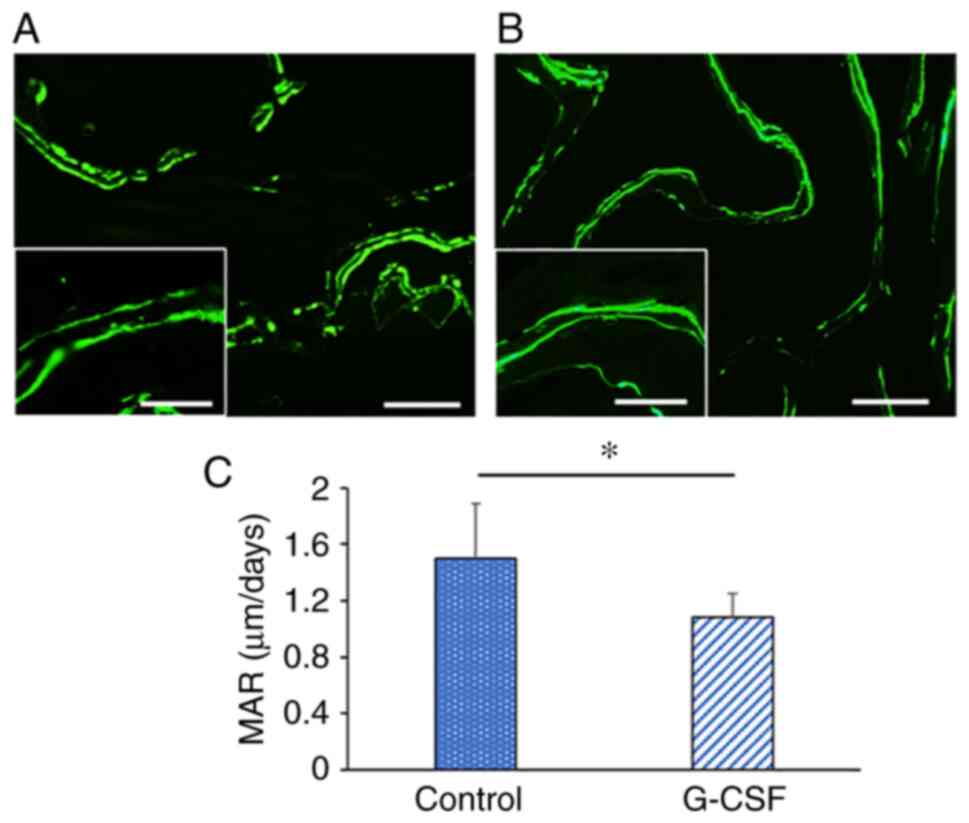
Dynamic parameters
Following calcein injection, two clear fluorescent
lines were observed in newly formed bone of the maxilla in the
control and G-CSF groups (Fig. 6A
and B). Analysis of the bone
formation parameter revealed that the G-CSF group presented a
significantly lower MAR compared with the control group (control
group, 1.50±0.39 µm/day; G-CSF group, 1.09±0.17 µm/day; Fig. 6C).
Discussion
Healthy bones are maintained by bone remodeling, a
physiological process in which old or damaged bone is resorbed by
osteoclasts and new bone is subsequently formed by osteoblasts
(20). In normal bone remodeling,
bone resorption and formation are closely associated, such that
bone mass and quality are unaltered. However, this physiological
process can be disrupted by a variety of factors, including
age-related factors, osteoporosis, drugs, and secondary diseases,
leading to the development of various bone diseases (20). Bone remodeling is systematically
regulated by a variety of systemic [e.g. estrogen (21) and parathyroid hormone (22)] and local factors [e.g. bone
morphogenetic proteins (23) and
transforming growth factor-β (23)] (2,24).
We investigated G-CSF as one of the factors in the regulation of
physiological bone remodeling.
G-CSF administration suppresses osteoblast and
osteocyte function via sympathetic nervous system-mediated
β2-adrenergic signaling, reduces the support system for HSPCs to
remain in the bone marrow microenvironment, and causes HSPCs to
leave the bone marrow niche and be released into the bloodstream
(4,25). Therefore, we hypothesized that
G-CSF, which suppresses osteoblasts and osteocytes, is a potential
candidate for a systemic factor that affects the bone remodeling
microenvironment. In the present study, significant differences in
the BV/TV, Tb. N and Tb.Sp of the femur and alveolar bone were
observed between the G-CSF and control groups. The trabecular bone
of the femur and alveolar bone were reduced in the G-CSF group
compared with the control group. These results suggested that G-CSF
may play important roles in the inhibition of bone turnover in not
only the femur but also the alveolar bone of mice.
The periodontal tissues of the maxillary first
molars of mice were previously surveyed for qualitative
histological tissue changes concomitant with aging, and significant
changes were observed with increasing age, including a reduction
and narrowing of the periodontal ligament space, particularly at
the root apices and interradicular regions, leading to ankylosis
(26). Tooth extraction in elder
mice is much more difficult than in young mice because of root
ankylosis. Moreover, the aging of the bone marrow microenvironment
has been shown to contribute to the decline in HSPC function over
time (27). In the present study,
our interest is boney regenerate. The bone turnover rate in the
maxilla was significantly higher than in the femur (28). Therefore, we used a young mouse
tooth extraction model in this study.
The serum P1NP level, a marker of bone formation,
was lower in the G-CSF group as compared with the control group six
days after G-CSF administration. There was no difference in the
serum TRACP-5b level (bone resorption/osteoclast number marker)
between the G-CSF and control groups. These data are consistent
with the morphometry results and suggest that G-CSF decreases the
trabecular bone volume by reducing bone formation. These results
suggested that GCS-F may play important roles in the inhibition of
bone turnover in the tooth extraction sockets of mice.
In this study, we extracted a unilateral maxillary
first molar of the mice and observed the tooth extraction sockets
histopathologically. Pan et al (29) examined tooth extraction wound
healing in laboratory mice using quantitative µCT imaging, and the
extraction sockets were nearly 50% filled within one week. Their
result corresponded with our result. By contrast, fibrous
connective tissues and immature bone were observed in the
extraction socket and bone healing was delayed in the G-CSF group
compared with the control group. Bone area in the extraction socket
was significantly smaller in the G-CSF group (23.6%) than in
control group (45.1%). We evaluated the number of TRAP-positive
cells in the maxilla, including the tooth extraction socket,
finding in the maxilla that they were non-significantly lower in
the G-CSF group than in the control group. This result suggested
that G-CSF administration did not play an important role for
osteoclasts in the inhibition of bone turnover of the tooth
extraction socket area. Osteocytes represent 90-95% of all bone
cells in the adult skeleton (30).
They produce and secrete sclerostin, receptor activator of nuclear
factor κΒ ligand, and osteoprotegerin to communicate indirectly
with bone-related cells through small tunnels termed canaliculi
(30,31). We showed a reduction in the
N.Ot.Ca/Ot.Lc. in the maxilla on G-CSF administration. We also
evaluated the MAR, which was lower in the G-CSF group than in the
control group. These results suggested that G-CSF plays an
important role in the inhibition of bone turnover in the tooth
extraction sockets of mice. The G-CSF group displayed significantly
delayed bone healing of the extraction socket in comparison with
the control group.
There are some limitations in this study. Firstly,
we only evaluated bone healing of the extraction socket. Future
research will continue to explore other bone healings. One example
is comparing the bone healing between tooth extraction model and
femur fracture model in mice. Secondly, at first, we designed the
study with eight mice per group. However, two or three mice of each
group were difficult to analyze due to tooth root fracture,
alveolar bone fracture or insufficient blood sample volumes.
Consequently, the number of mice was a minimum of five per group.
Moreover, we aim to investigate the effects of G-CSF on HSPC in
vitro in future work.
In conclusion, G-CSF administration reduced the
number of canaliculi per osteocyte and inhibited the connection of
the osteocyte networks. Consequently, osteoblast activation was
inhibited, and the bone remodeling changed to a state of low bone
turnover. For the above reasons, it was considered that the bone
healing of the socket after tooth extraction was delayed.
Supplementary Material
Representative images of
CXCL12/SDF-1-positive cells in bone marrow samples.
Immunoreactivity (black arrows) for CXCL12 in femoral metaphyseal
trabecules of the (A) control and (B) G-CSF-treatment groups and
around the extraction socket in the maxillary bone of the (C)
control and (D) G-CSF-treatment groups is shown as CXCL12/
SDF-1-positive cells located among the hematopoietic cells. The
number of CXCL12/SDF-1-positive cells was decreased by G-CSF
treatment. Scale bar indicates 50 μm. CXCL12, chemokine
(C-X-C motif) ligand 12; SDF-1, stromal cell-derived factor 1.
Acknowledgements
The authors would like to thank Ms. Shinobu Osawa
(Department of Oral and Maxillofacial Surgery, School of Medicine,
Hyogo Medical University, Nishinomiya, Japan) for technical
assistance.
Funding
Funding: This study was supported by JSPS KAKENHI [grant nos.
20H01118 (to MO), 21K10106 (to KT), 21K17126 (to MU) and 21H04285
(to KT)], and by Grant-in-Aid for Researchers, Hyogo Medical
University, 2021 (to MU).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MO, KaT, MU, KN and HK conceived and designed the
present study. MO, KaT, MU, KoT, HH, NY and KY performed the
experiments. MO, KaT, HH, KN and HK analyzed the data and performed
statistical analysis. KN and HK confirm the authenticity of all the
raw data. MO, KaT, KN and HK wrote the manuscript and created the
figures. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments and experimental protocols
were approved by the ethics committee of the Hyogo Medical
University (Hyogo, Japan) (approval number 19-032).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hübel K and Engert A: Clinical
applications of granulocyte colony-stimulating factor: An update
and summary. Ann Hematol. 82:207–213. 2003.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Turhan AB, Binay C, Bor O and Simsek E:
The effects of short-term use of granulocyte colony-stimulating
factor on bone metabolism in child cancer patients. North Clin
Istanb. 5:277–281. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Bendall LJ and Bradstock KF: G-CSF: From
granulopoietic stimulant to bone marrow stem cell mobilizing agent.
Cytokine Growth Factor Rev. 25:355–367. 2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Asada N, Katayama Y, Sato M, Minagawa K,
Wakahashi K, Kawano H, Kawano Y, Sada A, Ikeda K, Matsui T and
Tanimoto M: Matrix-embedded osteocytes regulate mobilization of
hematopoietic stem/progenitor cells. Cell Stem Cell. 12:737–747.
2013.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Lévesque JP, Hendy J, Takamatsu Y, Simmons
PJ and Bendall LJ: Disruption of the CXCR4/CXCL12 chemotactic
interaction during hematopoietic stem cell mobilization induced by
GCSF or cyclophosphamide. J Clin Invest. 111:187–196.
2003.PubMed/NCBI View
Article : Google Scholar
|
|
6
|
Semerad CL, Christopher MJ, Liu F, Short
B, Simmons PJ, Winkler I, Levesque JP, Chappel J, Ross FP and Link
DC: G-CSF potently inhibits osteoblast activity and CXCL12 mRNA
expression in the bone marrow. Blood. 106:3020–3027.
2005.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Katsimbri P: The biology of normal bone
remodelling. Eur J Cancer Care (Engl). 26(e12740)2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Hadjidakis DJ and Androulakis II: Bone
remodeling. Ann N Y Acad Sci. 1092:385–396. 2006.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Takamatsu Y, Simmons PJ, Moore RJ, Morris
HA, To LB and Lévesque JP: Osteoclast-mediated bone resorption is
stimulated during short-term administration of granulocyte
colony-stimulating factor but is not responsible for hematopoietic
progenitor cell mobilization. Blood. 92:3465–3473. 1998.PubMed/NCBI
|
|
10
|
Watanabe T, Suzuya H, Onishi T, Kanai S,
Kaneko M, Watanabe H, Nakagawa R, Kawano Y, Takaue Y, Kuroda Y and
Talmadge JE: Effect of granulocyte colony-stimulating factor on
bone metabolism during peripheral blood stem cell mobilization. Int
J Hematol. 77:75–81. 2003.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Bonilla MA, Dale D, Zeidler C, Last L,
Reiter A, Ruggeiro M, Davis M, Koci B, Hammond W, Gillio A and
Welte K: Long-term safety of treatment with recombinant human
granulocyte colony-stimulating factor (r-metHuG-CSF) in patients
with severe congenital neutropenias. Br J Haematol. 88:723–730.
1994.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Bishop NJ, Williams DM, Compston JC,
Stirling DM and Prentice A: Osteoporosis in severe congenital
neutropenia treated with granulocyte colony-stimulating factor. Br
J Haematol. 89:927–928. 1995.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Soshi S, Takahashi HE, Tanizawa T, Endo N,
Fujimoto R and Murota K: Effect of recombinant human granulocyte
colony-stimulating factor (rh G-CSF) on rat bone: Inhibition of
bone formation at the endosteal surface of vertebra and tibia.
Calcif Tissue Int. 58:337–340. 1996.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Wu YD, Chien CH, Chao YJ, Hamrick MW, Hill
WD, Yu JC and Li X: Granulocyte colony-stimulating factor
administration alters femoral biomechanical properties in C57BL/6
mice. J Biomed Mater Res A. 87:972–979. 2008.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Kishimoto H, Noguchi K and Takaoka K:
Novel insight into the management of bisphosphonate-related
osteonecrosis of the jaw (BRONJ). Jpn Dent Sci Rev. 55:95–102.
2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Shudo A, Kishimoto H, Takaoka K and
Noguchi K: Long-term oral bisphosphonates delay healing after tooth
extraction: A single institutional prospective study. Osteoporos
Int. 29:2315–2321. 2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Tamaoka J, Takaoka K, Hattori H, Ueta M,
Maeda H, Yamamura M, Yamanegi K, Noguchi K and Kishimoto H:
Osteonecrosis of the jaws caused by bisphosphonate treatment and
oxidative stress in mice. Exp Ther Med. 17:1440–1448.
2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Takaoka K, Yamamura M, Nishioka T, Abe T,
Tamaoka J, Segawa E, Shinohara M, Ueda H, Kishimoto H and Urade M:
Establishment of an animal model of bisphosphonate-related
osteonecrosis of the jaws in spontaneously diabetic torii rats.
PLoS One. 14(e0144355)2015.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Dempster DW, Compston JE, Drezner MK,
Glorieux FH, Kanis JA, Malluche H, Meunier PJ, Ott SM, Recker RR
and Parfitt AM: Standardized nomenclature, symbols, and units for
bone histomorphometry: A 2012 update of the report of the ASBMR
histomorphometry nomenclature committee. J Bone Miner Res. 28:2–17.
2013.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Feng X and McDonald JM: Disorders of bone
remodeling. Annu Rev Pathol. 6:121–145. 2011.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Khosla S and Monroe DG: Regulation of bone
metabolism by sex steroids. Cold Spring Harb Perspect Med.
8(a031211)2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wein MN and Kronenberg HM: Regulation of
bone remodeling by parathyroid hormone. Cold Spring Harb Perspect
Med. 8(a031237)2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zou ML, Chen ZH, Teng YY, Liu SY, Jia Y,
Zhang KW, Sun ZL, Wu JJ, Yuan ZD, Feng Y, et al: The smad dependent
TGF-β and BMP signaling pathway in bone remodeling and therapies.
Front Mol Biosci. 8(593310)2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Siddiqui JA and Partridge NC:
Physiological bone remodeling: Systemic regulation and growth
factor involvement. Physiology (Bethesda). 31:233–245.
2016.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Li SD, Chen YB, Qiu LG and Qin MQ: G-CSF
indirectly induces apoptosis of osteoblasts during hematopoietic
stem cell mobilization. Clin Transl Sci. 10:287–291.
2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Tonna EA: Histological age changes
associated with mouse parodontal tissues. J Gerontol. 28:1–12.
1973.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Matteini F, Mulaw MA and Florian MC: Aging
of the hematopoietic stem cell niche: New tools to answer an old
question. Front Immunol. 12(738204)2021.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Wang JY, Huo L, Yu RQ, Rao NJ, Lu WW and
Zheng LW: Skeletal site-specific response of jawbones and long
bones to surgical interventions in rats treated with zoledronic
acid. Biomed Res Int. 2019(5138175)2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Pan J, Pilawski I, Yuan X, Arioka M, Ticha
P, Tian Y and Helms JA: Interspecies comparison of alveolar bone
biology: Tooth extraction socket healing in mini pigs and mice. J
Periodontol. 91:1653–1663. 2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Bonewald LF: Osteocytes as dynamic
multifunctional cells. Ann N Y Acad Sci. 1116:281–290.
2007.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Kitaura H, Marahleh A, Ohori F, Noguchi T,
Shen WR, Qi J, Nara Y, Pramusita A, Kinjo R and Mizoguchi I:
Osteocyte-related cytokines regulate osteoclast formation and bone
resorption. Int J Mol Sci. 21(5169)2020.PubMed/NCBI View Article : Google Scholar
|