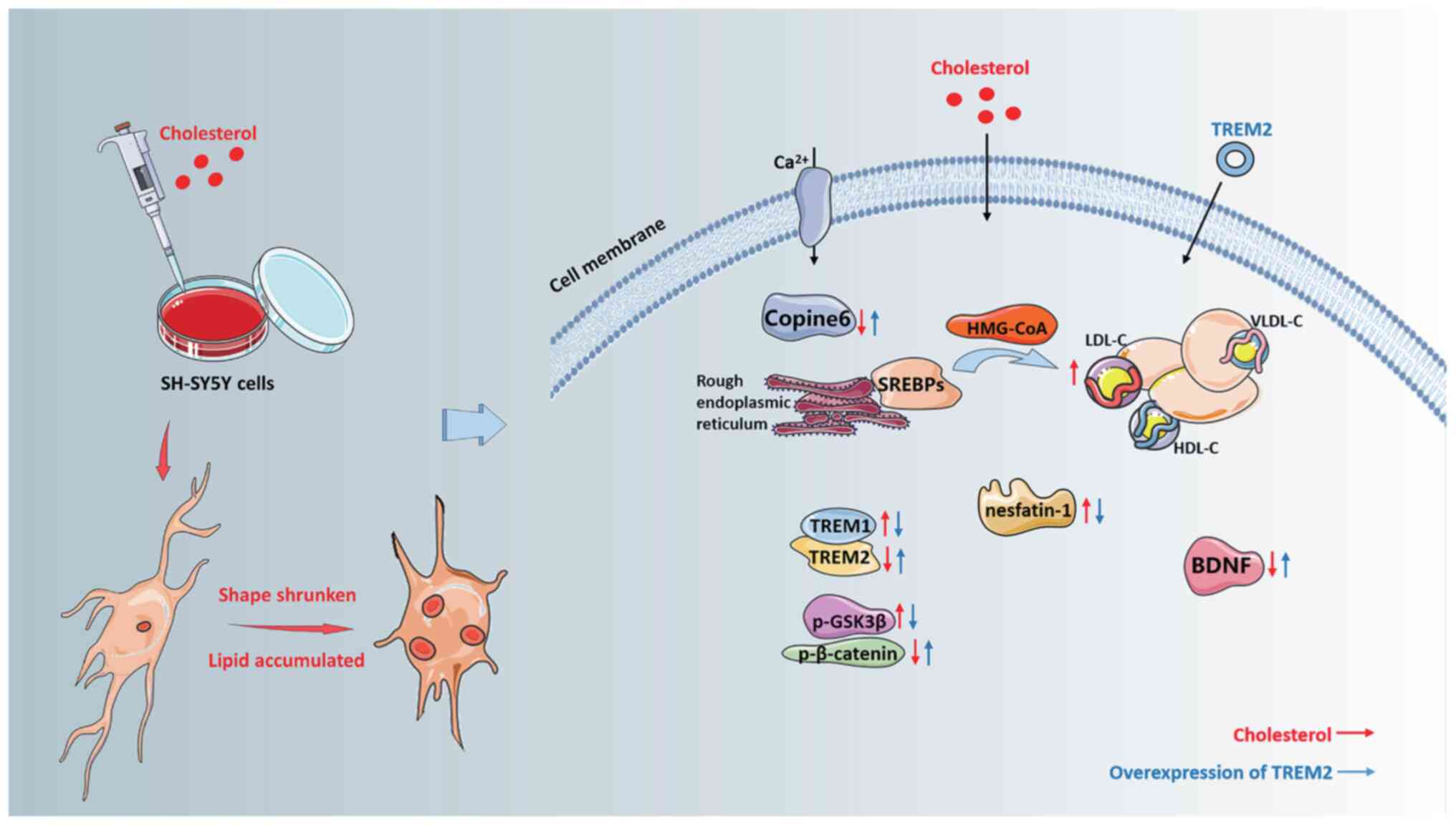
Introduction
Cholesterol is a major component of brain lipids and
plays a crucial role in the maintenance of neuronal cell function,
neurotransmission, and synapse formation; however, an increasing
number of studies have demonstrated the adverse effects of
excessive plasma cholesterol levels (1-3).
In 2018, the new version of the Blood Cholesterol Management
Guidelines was jointly developed and released by the American
College of Cardiology, American Heart Association, and other
departments, and this version clearly notes that high levels of
cholesterol, particularly low-density lipoprotein cholesterol
(LDL-C), can increase the risk of cardiovascular and
cerebrovascular diseases such as heart attack and stroke (4,5).
Moreover, excessive cholesterol is reportedly closely related to
the progression of cognitive impairment and even Alzheimer's
disease (AD) (6-8).
Cholesterol is normally maintained at low levels in neurons, which
inhibits β-amyloid (Aβ) accumulation and enables astrocytes to
regulate Aβ accumulation by cholesterol signaling (9). However, imbalanced plasma cholesterol
levels, particularly elevated LDL-C and decreased high-density
lipoprotein cholesterol levels, are negative factors affecting
cognitive function (7,10,11)
and are positively correlated with abnormal amyloid deposition in
the brain (11). Moreover,
disturbed neuronal cholesterol homeostasis has been observed in AD
(12), and the accumulation of
intracellular cholesterol reportedly reduces the clearance of
defective mitochondria (13) and
thus contributes to the pathogenesis of AD. Furthermore, it has
been reported that the targeted deletion of cholesterol synthesis
in astrocytes decreases the burden of amyloid and tau in an AD
mouse model (9). In our previous
study, high-fat diet-fed rats showed not only hyperglycemia and
hyperlipidemia with increased plasma concentrations of total
cholesterol (TC), total triglycerides, and LDL-C, but also
neuropsychiatric impairments, including decreased learning and
memory abilities, as well as depression-like behavior, accompanied
by reduced expression of brain-derived neurotrophic factor (BDNF)
and related changes in synaptic plasticity in the hippocampus and
prefrontal cortex (PFC) (14).
These findings suggest a strong potency of high cholesterol against
neuropsychiatric injuries. However, there remain other
controversial results suggesting that high levels of cholesterol
are a potential protective factor for cognitive decline (15) and are negatively correlated with
the occurrence of dementia (16).
Therefore, it is important to investigate the exact effects of
cholesterol on neural cells and whether there is a clear connection
and causality between turmoil lipids and cognitive impairment
(17).
Sterol regulatory element binding proteins (SREBPs)
are important regulators of cholesterol homeostasis that regulate
several enzymes needed for the synthesis of cholesterol, fatty
acids, triacylglycerol, and phospholipids (18). Dysregulation of SREBP-2 signaling
has been observed in AD and related models, and this dysregulation
is attributed to increased cholesterol levels and alterations in
the tau protein (12). Nesfatin-1
is a peptide of 82 amino acids produced by hydrolysis of the
N-terminus of nucleobindin2, which is widely distributed in central
and peripheral tissues (19).
Increased plasma concentrations of nesfatin-1 have been detected in
diabetic patients (20) and rats
with metabolic syndrome (21), and
are involved in the regulation of glucose and lipid metabolism
(22). It has been reported that
chronic infusion of nesfatin-1 can prevent hepatic steatosis,
partly by downregulating the expression of SREBP1(23). Furthermore, nesfatin-1 has been
demonstrated to be associated with the regulation of cognition and
mood (24,25). In line with these findings, an
increased plasma concentration of nesfatin-1 has been observed in
rats with hyperlipidemia (26) or
stress (27) in our previous
studies (28,29), and the results showed that the
intraperitoneal injection of nesfatin-1 induced anxiety and
depression-related behaviors in rats accompanied by adaptive
changes in the expression of BDNF and synaptic plasticity in the
hippocampus and PFC. These findings suggest the potential role of
nesfatin-1 in linking metabolic disorders with neuropsychiatric
injuries.
The classic Wnt/β-catenin signaling pathway is a
highly conserved signaling pathway and one of the most conserved
regulators of evolution in embryonic development and adult tissue
homeostasis (30-32).
Dysfunction of the Wnt/β-catenin signaling pathway has been
demonstrated to play an important role in the pathogenesis of AD,
Parkinson's disease, depression, and other neuropsychiatric
diseases (10,33), and is partly involved in
controlling neuronal activity-regulated BDNF secretion (34,35).
The results of our previous study showed that the impaired learning
and memory abilities and decreased expression of BDNF found in the
hippocampus and PFC of high-fat diet-fed rats were associated with
imbalanced expression of key molecules in the Wnt/β-catenin
signaling pathway, including increased expression of CyclinD1 and
decreased expression of phosphorylated β-catenin (26). Taken together with the protective
role of BDNF in regulating cholesterol homeostasis (36), these findings suggest that the
Wnt/β-catenin signaling pathway may be involved in neuropsychiatric
dysfunction and the changes in the BDNF abundance caused by glucose
and lipid metabolism disorders (37).
Triggering receptor expressed on myeloid cells
(TREM), which was first discovered in 2000, is a relatively novel
type of immunoglobulin family inflammatory stimulating receptor
(38). TREM1 and TREM2 are two
subtypes of this receptor, and they have been demonstrated to be
involved in not only inflammatory and immune responses but also the
regulation of neuropsychiatric function, including learning and
memory. In addition to the relationship between AD and the gene
polymorphisms of TREM1 or TREM2(39), it has also been reported that TREM2
can promote the survival of microglia by activating the
Wnt/β-catenin pathway (40),
accelerate the clearance of amyloid protein and reduce oxidative
stress injury in hippocampal neurons (41). Upregulation of the expression of
TREM2 can inhibit hippocampal neuronal apoptosis (12) and improve the spatial cognitive
impairment in a mouse model of AD (42). In addition, inhibiting the
expression of TREM2 can promote the abnormal accumulation of
amyloid protein (43) and
aggravate the impairment of spatial learning ability in P301S
transgenic mice (44). However,
Jiang et al (42) reported
that the overexpression of TREM2 in microglia could effectively
improve AD-like neuropathological changes and spatial learning and
memory impairments in 7-month-old APPswe/PS1dE9 mice, including
amyloid deposition, neuroinflammation, and neuronal and synaptic
loss. However, their subsequent study revealed that the same
intervention could not improve neuropathological damage and memory
impairment in 18-month-old APPswe/PS1dE9 mice (45). These findings suggest that the role
of TREM2 may be altered according to different pathological
statuses. Consistently, in our previous study, differential
expression levels of TREM2 were found in the hippocampus of rats of
different ages and this was correlated with their performance in
the Morris water maze. Moreover, imbalanced expression of TREM2 was
also found in the hippocampus and PFC of rats fed a high-fat diet,
suggesting that TREM2 may be involved in neuropsychiatric injury
caused by abnormal glucose and lipid metabolism (46).
The aim of the present study was to investigate the
effect of high cholesterol stimulation on the morphology and
function of neuronal cells, and to explore the potential mechanism
of action of TREM2 in neuronal cells.
Materials and methods
Cell culture
The neuroblastoma cell line SH-SY5Y was obtained
from Wuhan Sunnbio Technology Co, Ltd. The cells were plated in
DMEM (HyClone, Cytiva), 10% FBS (Biological Industries; Sartorius
AG), and 1% penicillin/streptomycin (Beyotime Institute of
Biotechnology) in a humidified incubator at 37˚C and supplied with
5% CO2.
Cholesterol challenge
16 mg of water-soluble cholesterol (MilliporeSigma)
was dissolved in 10 ml PBS buffer solution to prepare a cholesterol
mixture of 1,000 µM, which was successively diluted 10 times with
DMEM before use. In the MTT assay, the cells were stimulated with
0.1-100 µM cholesterol for 3, 6, 9, 12, 24, or 48 h, and the
effects of different concentrations and times on cell viability
were observed. For all subsequent experiments, cells were treated
with 100 µM for 24 h.
MTT assay
SH-SY5Y cells were plated in a 96-well culture plate
at a density of 1x104 cells per well. After treatment
with cholesterol, 10 µl MTT (Beijing Solarbio Science &
Technology Co., Ltd.) was added to each well and the plate was
incubated at 37˚C for 4 h. The supernatant was discarded, 150 µl
DMSO was added per well, and the plate was incubated at 37˚C for 10
min. Net, the absorbance was measured with a microplate reader at
490 nm.
Oil red O staining
SH-SY5Y cells were plated in 6-well culture plates
at a density of 2x105 cells per well. At the end of the
stimulation period, the cells were washed three times with PBS and
fixed with 4% paraformaldehyde for 10 min at 37˚C. Next, cells were
washed twice, then stained with 0.5% Oil red O for 30 min at 37˚C.
After staining, the cells were washed once with 60% isopropanol,
washed with PBS until a colorless solution was obtained, and
observed under a fluorescent inverted microscope at 50x
magnification.
Cell cycle analysis
Cells (5x106) were collected and fixed in
70% ice-cold ethanol at -20˚C overnight. The cells were then
collected and resuspended in PBS supplemented with 100 ng/ml RNase
A and 50 ng/ml propidium iodide (PI) for 30 min at 37˚C. After
staining, the distribution of cell cycle stage was assessed using a
flow cytometer (CytoFLEX A00-1-1102, Beckman Coulter Biotechnology)
and analysis software (CytExpert 2.4, Beckman Coulter
Biotechnology).
Western blotting
SH-SY5Y cells were lysed using RIPA lysis buffer
(cat. no. P0013B, Beyotime Institute of Biotechnology) supplemented
with containing protease inhibitors (cat. no. ST507-10mL, Beyotime
Institute of Biotechnology) and phosphatase inhibitors (cat. no.
ST019-10mM, Beyotime Institute of Biotechnology). The protein
samples were prepared, resolved using 12.5% SDS gels by SDS-PAGE,
and transferred to PVDF membranes (cat. no. IPVH00010;
MilliporeSigma). The membranes were blocked using 5% skimmed milk
for 2 h at room temperature and incubated with antibodies against
β-catenin, p-β-catenin (1:800; Abcam), TREM1 (1:2,000; Abcam),
TREM2 (1:1,000; Abcam), CyclinD1 (1:1,000; Abcam), nesfatin-1
(1:5,000; Technology), GSK3β, p-GSK3β (Ser9) (1:1,000; Cell
Signaling Technology, Inc.), Copine-6 (1:1,000; Santa Cruz
Biotechnology, Inc.), BDNF (1:1,000; Abcam), or β-actin (1:1,000;
OriGene Technologies, Inc.) overnight at 4˚C and then incubated
with a horseradish peroxidase-conjugated secondary antibody
(1:5,000; OriGene Technologies, Inc.) at 37˚C for 2 h. Densitometry
analysis was performed using ImageJ 1.53 (National Institute of
Health).
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and converted
to cDNA using a RT kit according to the manufacturer's protocol
(Takara Bio, Inc.). qPCR was performed in a thermal cycler with the
following amplification program: 95˚C for 3 min followed by 40
cycles of 95˚C for 10 sec and 55˚C for 30 sec. The sequences of the
primers used were: Human SREBP1 forward, 5'ACACAGCAACCAGAAACTCAA3'
and reverse, 5'AGTGTGTCCTCCACCTCAGTCT3'; human SREBP2 forward,
5'AGCAGAGTTCCTTCTGCCAT3' and reverse, 5'GACAGTAGCAGGTCACAGGT3'; and
human β-actin forward, 5'TTGCCGACAGGATGCAGAA3' and reverse,
5'GCCGATCCACACGGAGTACTT3'. Expression was calculated using the
2-∆∆Cq method and normalized to the respective loading
control.
Overexpression of TREM2
During the logarithmic growth phase, SH-SY5Y cells
were counted and plated into 6-well culture plates with 2 ml cell
suspension (2x105 cells/well). After the cells had
adhered, the medium was discarded, and the cells were washed with
PBS twice. A total of 1,600 µl Opti-MEM (Thermo Fisher Scientific,
Inc.) was gently added to each well and the cells were then
incubated at 37˚C for 12 h. Subsequently, 0.5 µg empty vector
(pCMV-T7-3xFLAG-MCS-Neo, Wuhan MiaoLing Plasmid Company) or 0.5 µg
TREM2 vector (pECMV-3xFLAG-TREM2, Wuhan MiaoLing Plasmid Company)
were dissolved in 20 µl of sterile water, and 5 µl of the dissolved
plasmid was added to 5 µl Lipofectamine® 2000 (Thermo
Fisher Scientific, Inc.) and 200 µl Opti-MEM, and this solution was
incubated at room temperature for 20 min. After incubation, the
mixture was added to the corresponding wells in the 6-well plate,
and the cells were incubated at 37˚C for a further 8 h. Next, the
culture medium was removed, the cells were washed with PBS twice,
and treated with either control (normal medium) or
cholesterol-containing medium, and further incubated at 37˚C for 24
h. The experiment consisted of six groups: Control, cholesterol,
control + empty vector, control + TREM2 vector, cholesterol empty
vector, and cholesterol + TREM2 vector.
Statistical analysis
All data were analyzed using SPSS (version 17.0,
SPSS Inc.). Data are presented as the means ± SEM of at least three
repeats. Data were compared using Student's t-test or a one-way
ANOVA followed by a Tukey's post hoc test. GraphPad Prism 8
(GraphPad Software, Inc.) was used for Pearson correlation
analysis. P<0.05 was considered to indicate a statistically
significant difference.
Results
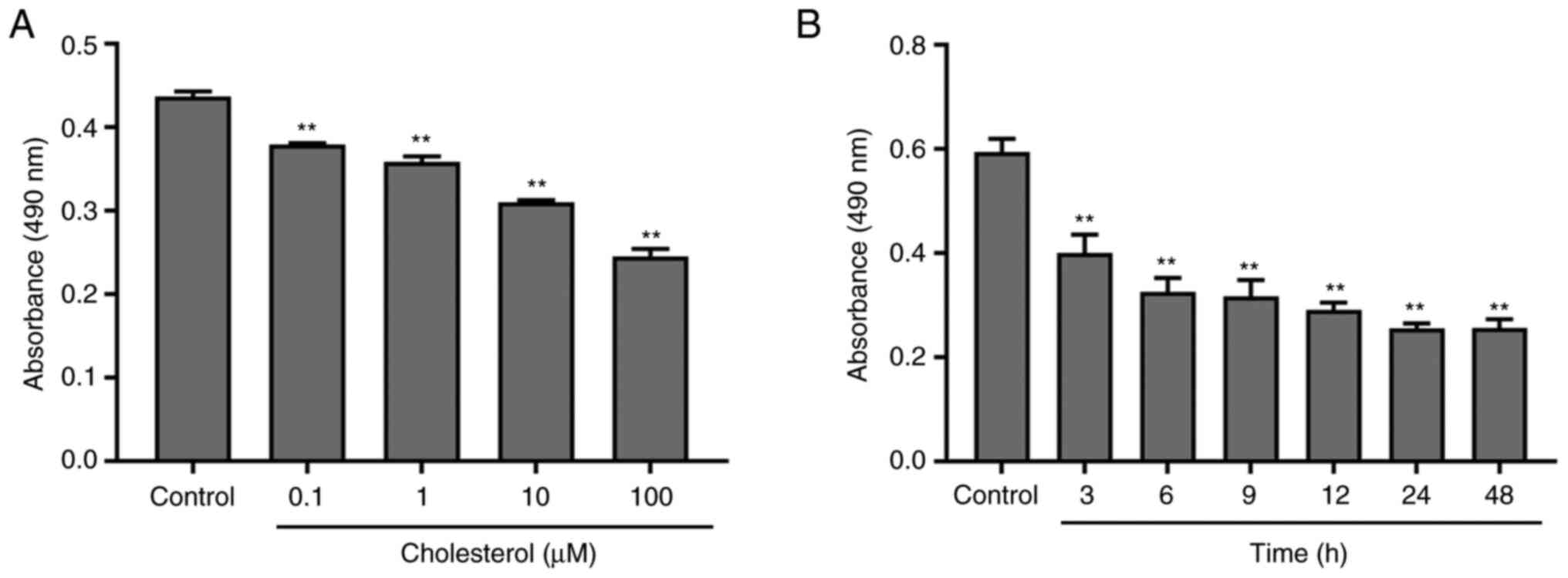
Dose- and time-dependent effects of cholesterol
treatment on the viability of SH-SY5Y cells. SH-SY5Y cells were
cultured and treated with cholesterol (0.1, 1, 10, or 100 µM) for
24 h. As shown in Fig. 1A,
compared with that of the control group, the viability of SH-SY5Y
cells gradually decreased in a dose-dependent manner, and the
viability of cells treated with 100 µM cholesterol was ~50%
(F0.05, 4=70.344, P≤0.0001). Fig. 1B shows the time-dependent effect of
cholesterol on cell viability; a plateau in the effect of time was
observed at 24 h (F0.05, 6=24.095, P≤0.0001). Therefore,
for subsequent experiments, cells were treated with 100 µM
cholesterol for 24 h.
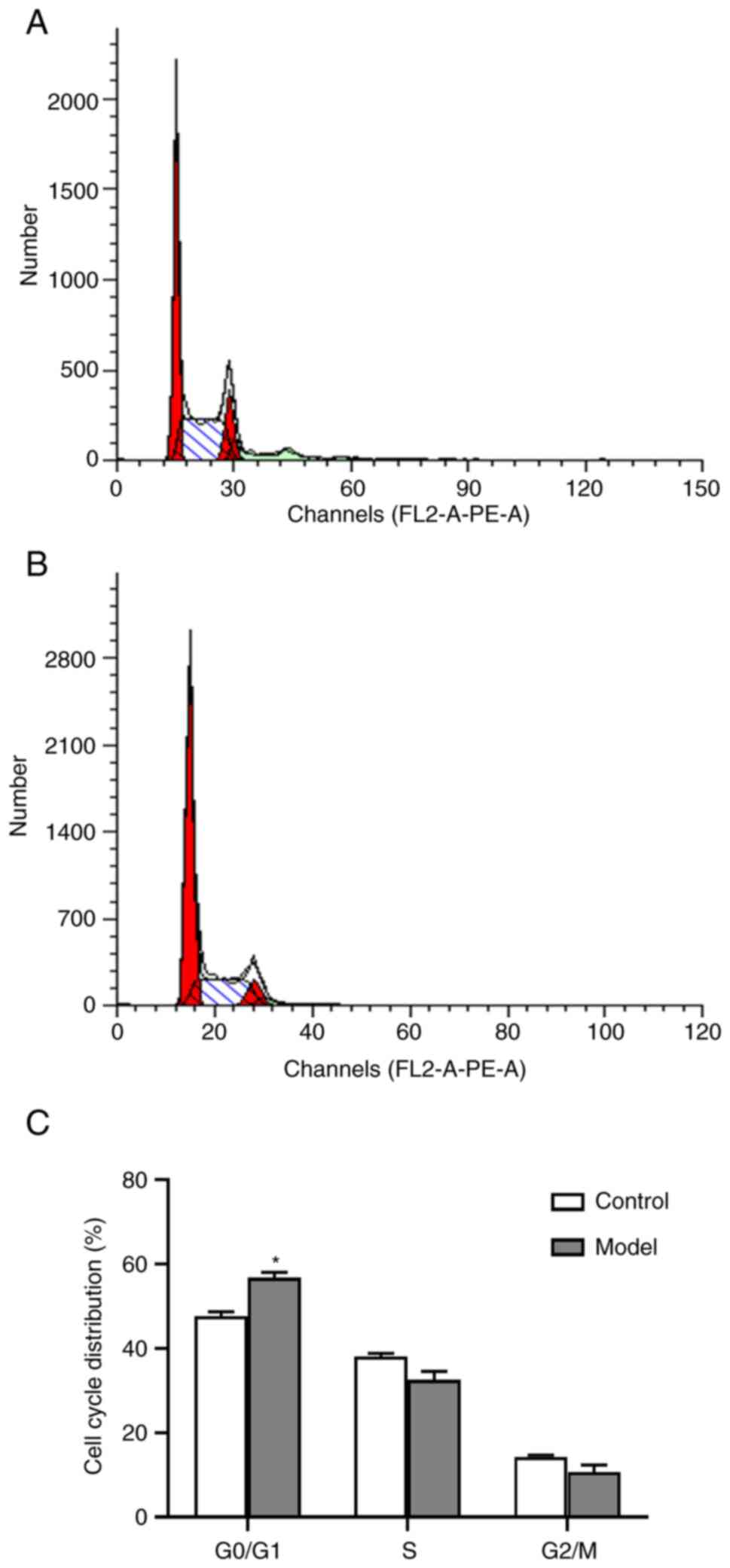
Effect of high cholesterol treatment
on the cell cycle distribution of SH-SY5Y cells
As shown in Fig. 2,
the number of SH-SY5Y cells in the G0/G1 after 24 h of stimulation
with 100 µM cholesterol was significantly higher than that of the
control group (t=4.354, P=0.022). However, cholesterol treatment
had no significant effect on the proportion of cells in the S and
G2/M phases.
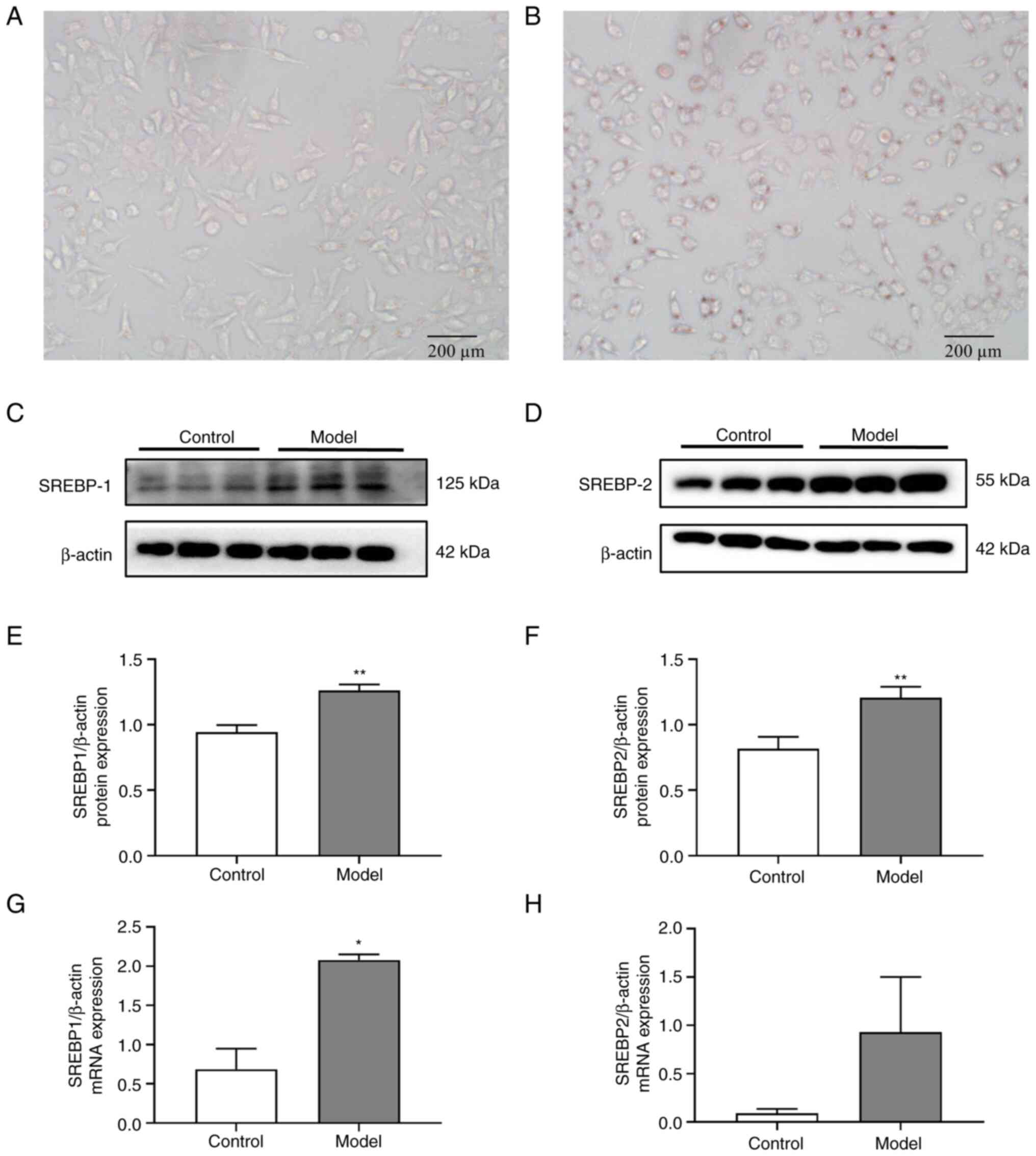
Effects of high cholesterol treatment
on lipid deposition and SREBP expression in SH-SY5Y cells
Fig. 3A and
B show a typical graph of the Oil
red O staining of each group. Compared with the results observed in
the control group, high cholesterol treatment altered the
morphology of SH-SY5Y cells from a spindle-like to an oval-like
shape and increased the number of Oil red O-stained cells, with
stained fat granules observed both inside and outside the cell. The
protein (Fig. 3C-F) and mRNA
(Fig. 3G and H) expression levels of SREBP1 (t=7.655,
P≤0.0001 and t=6.994, P=0.020; respectively) and SREBP2 (t=5.643,
P≤0.0001, t=1.582, P=0.254) in the cholesterol-stimulated SH-SY5Y
cells were markedly higher than that in the control group.
Effect of high cholesterol treatment
on the expression of BDNF, Copine-6, TREM1, and TREM2 in SH-SY5Y
cells
Fig. 4A and
B show the protein expression
levels of BDNF, Copine-6, TREM1, and TREM2 in SH-SY5Y cells.
Treatment with 100 µM cholesterol for 24 h markedly decreased the
protein expression levels of BDNF (t=2.732, P=0.020), Copine-6
(t=6.144, P≤0.0001), and TREM2 (t=8.189, P≤0.0001) and markedly
increased the protein expression levels of TREM1 (t=2.871, P=0.015)
compared with that in the control group. Pearson correlation
analysis showed that the protein expression levels of BDNF in
SH-SY5Y cells were positively correlated with the expression of
Copine-6 (r=0.888, P=0.003, Fig.
4C) and TREM2 (r=0.901, P=0.002, Fig. 4D) but negatively correlated with
the expression of TREM1 (r=-0.715, P=0.046, Fig. 4E).
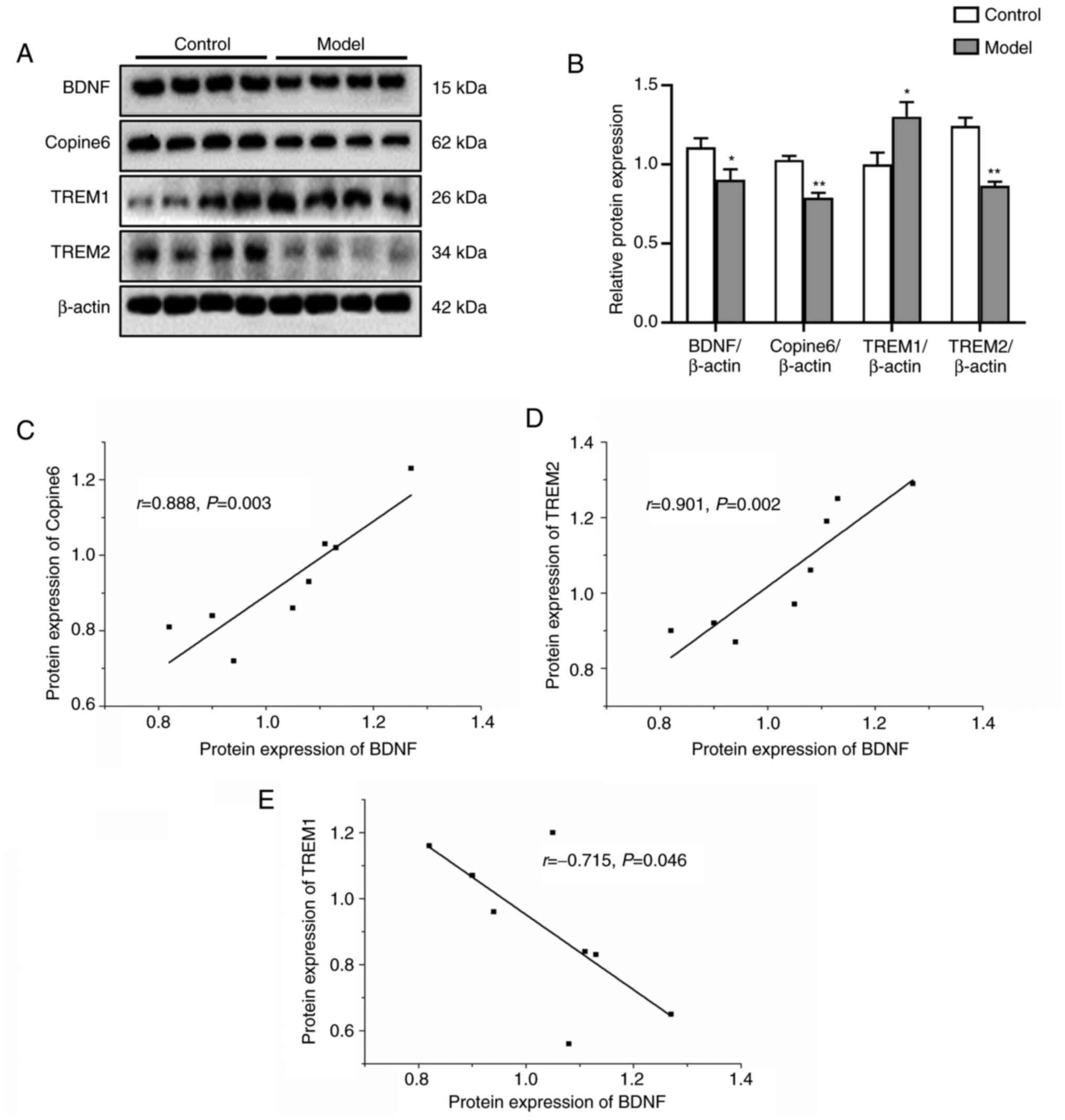 | Figure 4Effect of high cholesterol on the
protein expression levels of BDNF, Copine-6, TREM1, and TREM2 in
SH-SY5Y cells. Data are presented as the mean ± SEM of three
repeats each from two separate passages (n=6). (A) Representative
blots of BDNF, Copine-6, TREM1, and TREM2 expression in SH-SY5Y
cells. (B) Densitometry analysis of the western blotting. (C-E)
Pearson correlation analysis of BDNF with Copine-6, TREM1 and
TREM2. *P<0.05, **P<0.01 vs. control.
Model, SH-SY5Y cells treated with 100 µM for 24 h; TREM, triggering
receptor expressed on myeloid cells. |
Effects of high cholesterol treatment
on the protein expression levels of nesfatin-1 and key molecules in
the Wnt/β-catenin signaling pathway in SH-SY5Y cells
As shown in Fig. 5A
and B, the cholesterol-stimulated
SH-SY5Y cells exhibited a lower p-β-catenin/β-catenin protein
expression ratio and higher protein expression levels of nesfatin-1
(t=4.299, P=0.001) and CyclinD1 (t=5.619, P≤0.0001) and an increase
in the p-GSK3β/GSK3β ratio (t=2.883, P=0.015) compared with the
control cells. No significant differences in CyclinD1 expression
levels were observed between the two groups. Pearson correlation
analysis showed that the protein expression levels of nesfatin-1 in
SH-SY5Y cells was positively correlated with the expression of
p-GSK3β (r=0.821, P=0.013, Fig.
5C), CyclinD1 (r=0.764, P=0.027, Fig. 5D) and TREM1 (r=0.706, P=0.050,
Fig. 5E) but negatively correlated
with the expression of TREM2 (r=-0.719, P=0.044, Fig. 5F), p-β-catenin (r=-0.869, P=0.005,
Fig. 5G) and Copine-6 (r=-0.760,
P=0.029, Fig. 5H).
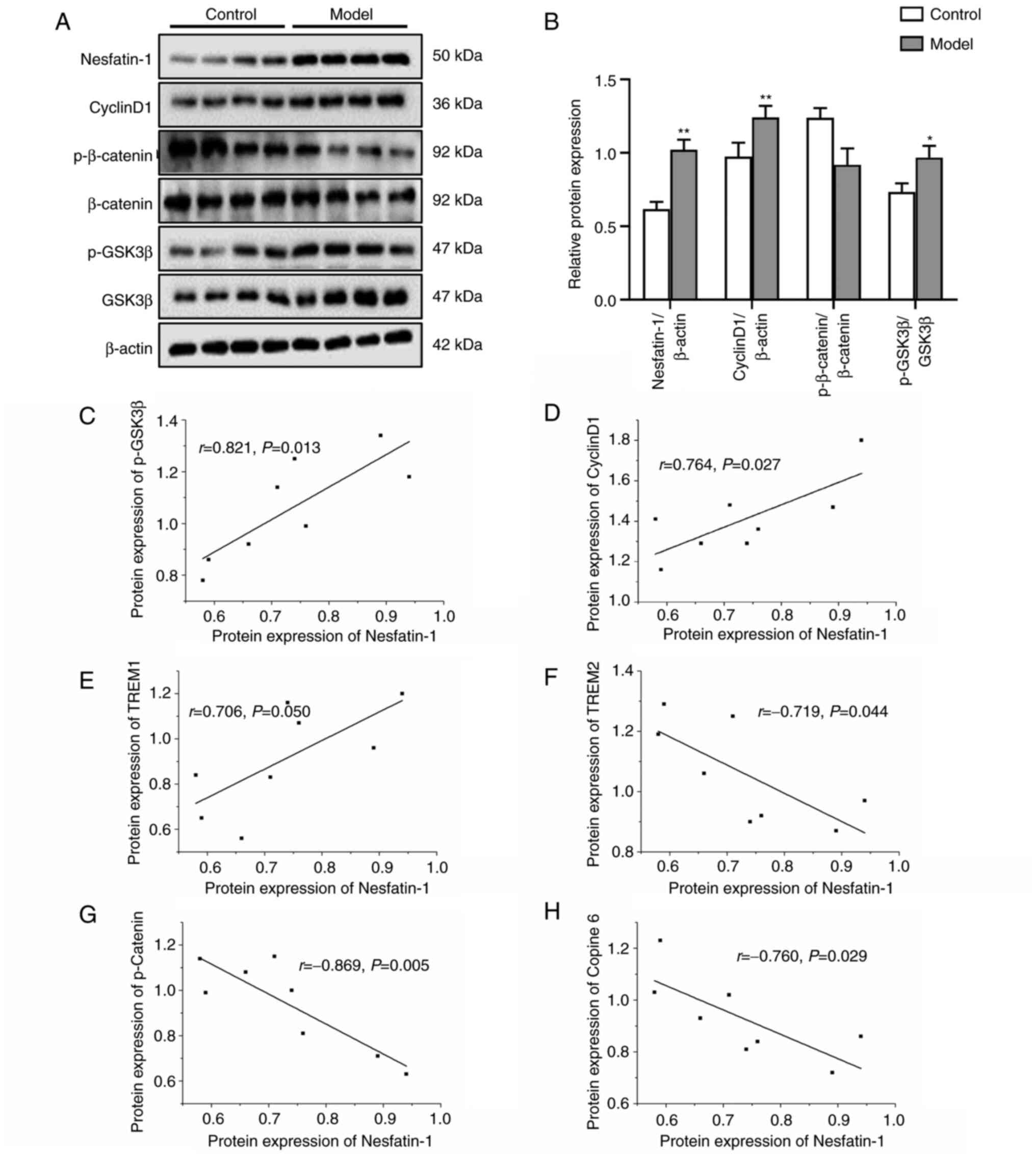 | Figure 5Effects of high cholesterol on the
protein expression levels of nesfatin-1 and key molecules in the
Wnt/β-catenin signaling pathway in SH-SY5Y cells. Data are
presented as the mean ± SEM of three repeats each from two separate
passages (n=6). (A) Representative blots of nesfatin-1, CyclinD1,
p-β-catenin, β-catenin, p-GSK3β, GSK3β protein expression in
SH-SY5Y cells. (B) Densitometry analysis of the western blotting
results. (C-H) Pearson correlation analysis of nesfatin-1 with
p-GSK3β, CyclinD1, TREM1, TREM2, p-β-catenin and Copine 6.
*P<0.05, **P<0.01 vs. control. Model,
SH-SY5Y cells treated with 100 µM for 24 h. |
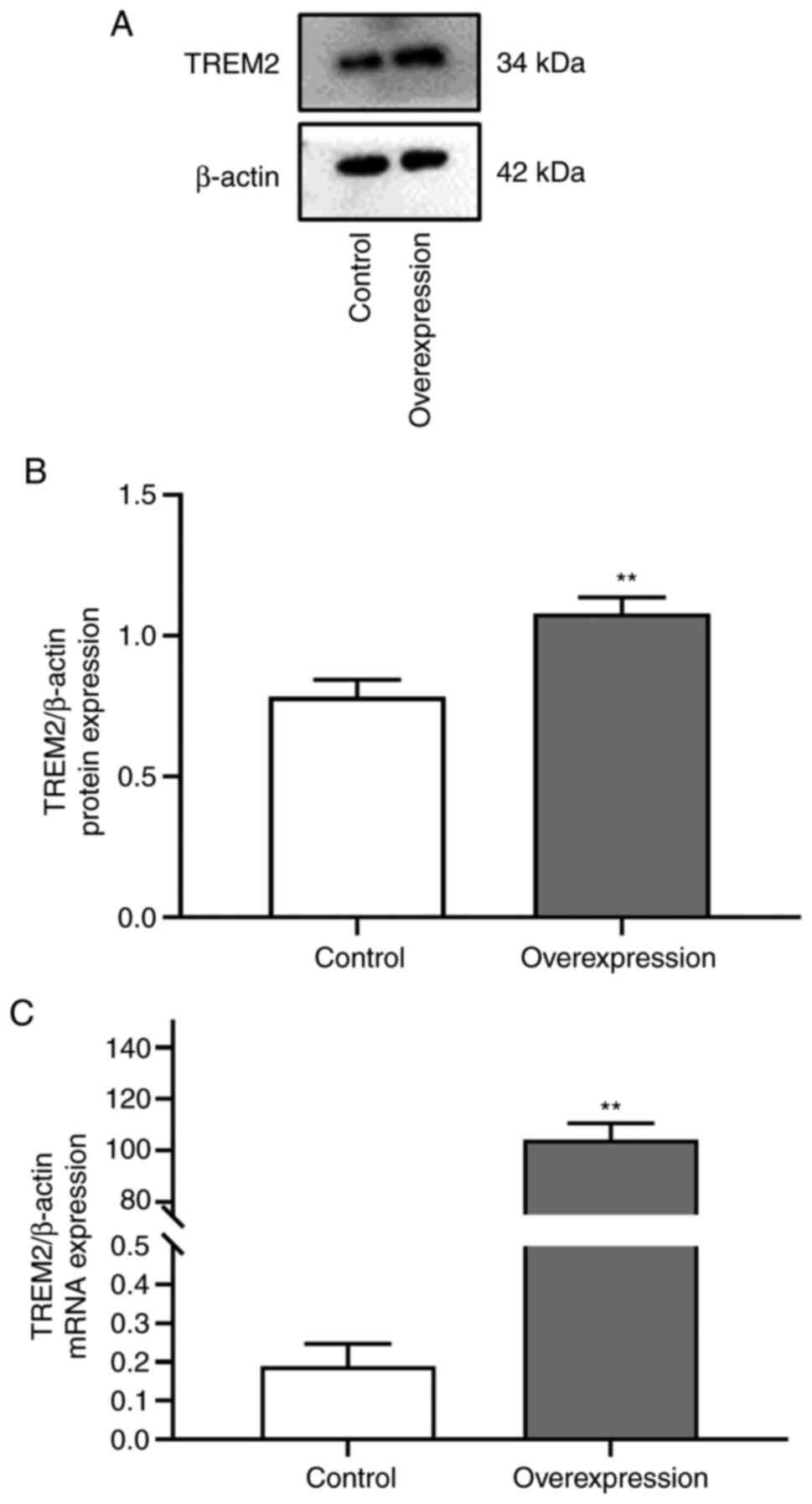
Effect of TREM2 overexpression on
dysregulation of BDNF, Copine-6, TREM1, and TREM2 protein
expression induced by high cholesterol in SH-SY5Y cells
The mRNA (t=16.188, P=0.004) and protein (t=9.849,
P=0.010) expression levels of TREM2 after transfection of the
TREM2-overexpressing plasmid were significantly increased (Fig. 6A-C) compared with those of the
control group. These results indicated the successful establishment
of the TREM2 overexpression model.
As shown in Fig. 7A
and B, the cholesterol-stimulated
SH-SY5Y cells exhibited lower protein expression levels of BDNF
(F0.05, 5=7.497, P=0.033) and Copine-6 (F0.05,
5=4.565, P=0.044) and higher expression of TREM1 (F0.05,
5=5.597, P=0.024) compared with the control cells. However,
this imbalance in expression was reversed after overexpression of
TREM2 as shown in Fig. 7B.
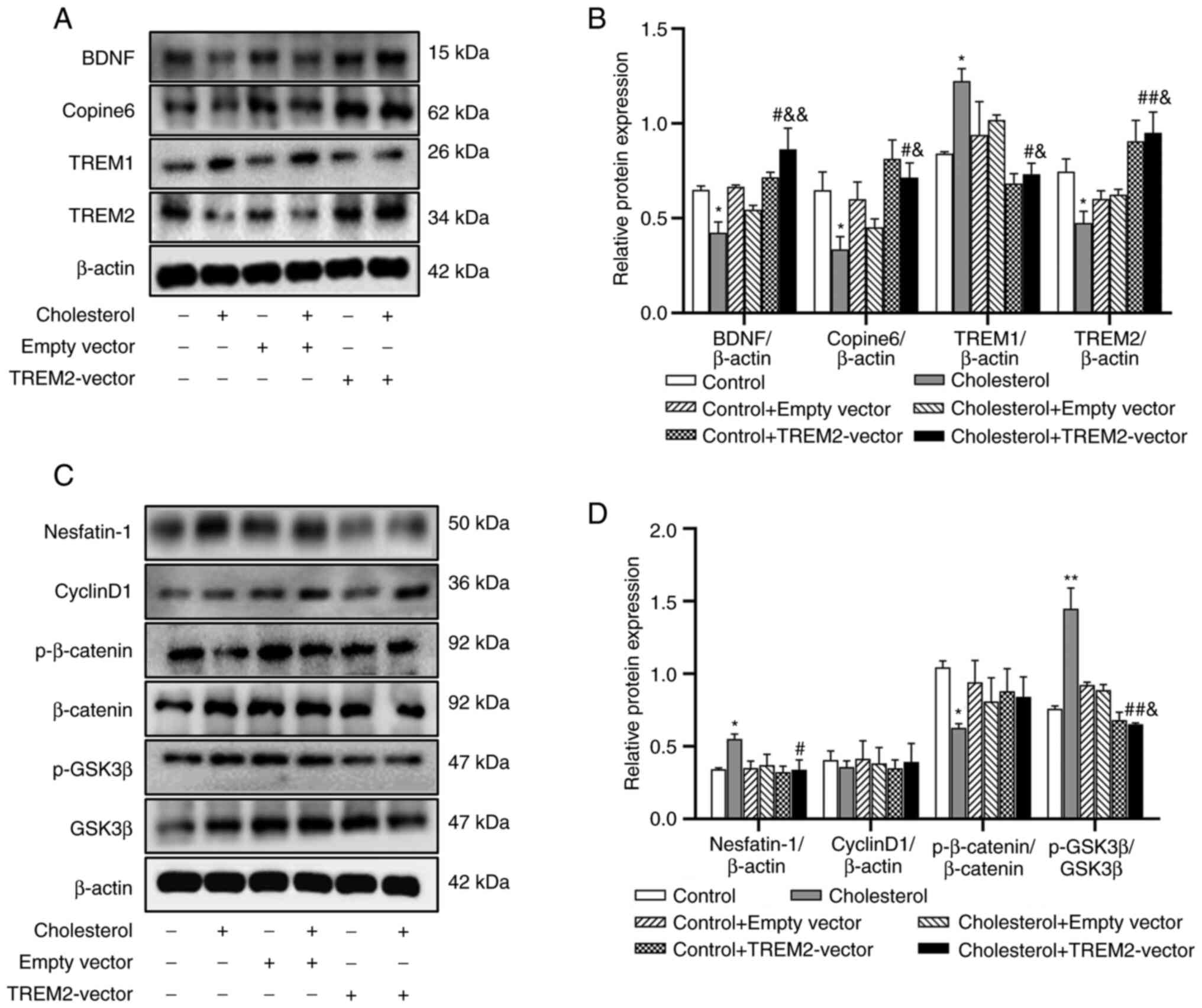 | Figure 7Effects of TREM2 overexpression on
the protein expression levels of BDNF, Copine-6, TREM1, TREM2,
nesfatin-1, and key molecules in the Wnt/β-catenin signaling
pathway following treatment with 100 µM cholesterol in SH-SY5Y
cells. Data are presented as the mean ± SEM of three repeats each
from two separate passages (n=6). (A) Representative blots of BDNF,
Copine-6, TREM1, and TREM2 expression in SH-SY5Y cells. (B)
Densitometry analysis of the western blotting results. (C)
Representative blots of nesfatin-1, CyclinD1, p-β-catenin,
β-catenin, p-GSK3β, GSK3β expression in SH-SY5Y cells. (D)
Densitometry analysis of the western blotting results.
*P<0.05, **P<0.01 vs. control.
#P<0.05, ##P<0.01 vs. cholesterol.
&P<0.05, &&P<0.01 vs.
cholesterol + empty vector. |
Effects of TREM2 overexpression on the
protein expression levels of nesfatin-1 and key molecules in the
Wnt/β-catenin signaling pathway induced by high cholesterol in
SH-SY5Y cells
As shown in Fig. 7C
and D, cholesterol-stimulated
SH-SY5Y cells showed higher expression levels of nesfatin-1
(F0.05, 5=2.824, P=0.015) and p-GSK3β (F0.05,
5=19.835, P≤0.0001) and lower expression levels of
p-β-catenin (F0.05, 5=1.222, P=0.046) compared with the
control cells. Overexpression of TREM2 reversed the increase in
expression of nesfatin-1, and p-GSK3β induced by cholesterol
treatment, and no significant change in the expression of CyclinD1
and p-β-catenin was observed in the TREM2 overexpression group.
Discussion
In the present study, the potential role of TREM2 in
cell injury and metabolic dysfunction induced by high cholesterol
in SH-SY5Y cells was investigated and the potential mechanism was
explored. The results showed that cholesterol could induce cell
injury and lipid accumulation in SH-SY5Y cells, as indicated by the
decrease in cell viability, changes in cell morphology from a
spindle to an oval shape, an increase in the number of cells in the
G0/G1 phase, an increase in the number of Oil-red O-positive cells,
and higher protein expression levels of the SREBPs. Moreover,
cholesterol-stimulated SH-SY5Y cells showed marked decreases in the
protein expression levels of BDNF, nesfatin-1, Copine-6, TREM2, and
p-β-catenin and increases in the expression of TREM1 and p-GSK3β.
However, the imbalanced expression of BDNF, Copine-6, nesfatin-1
and p-GSK3β induced by cholesterol in SH-SY5Y cells was reversed
after the overexpression of TREM2. These results suggested that
TREM2 was associated with the neurotoxicity of cholesterol in
SH-SY5Y cells.
As the basic structural component of the cell plasma
membrane, cholesterol participates in the formation of dense myelin
and plays an extremely important role in the structure and function
of the central nervous system (47,48).
An increasing body of evidence has demonstrated that
hypercholesterolemia is an important risk factor for
neuropsychiatric diseases, including AD (49,50).
The results of a 13-year follow-up study (49) showed that higher levels of TC and
LDL-C increased the risk of AD and accelerated the accumulation of
β-amyloid molecules by 20-fold (51), the latter of which is a key factor
in neuropathological damage in patients with AD (52,53).
Consistent with the findings that excessive cholesterol levels can
cause nerve cell injury and apoptosis (54), the results of the present study
showed that stimulation with high cholesterol resulted in a
decrease in the viability of SH-SY5Y cells in a dose- and
time-dependent manner accompanied by morphological changes from a
spindle-like shape to an oval-like shape, and a significant
increase in the proportion of cells in the G0/G1 phase. Moreover,
after stimulation with cholesterol, SH-SY5Y cells exhibited
significantly higher lipid accumulation, as indicated by the
increase in the number of Oil red O-positive cells, and in the
protein expression levels of SREBP1 and SREBP2. These results
indicated that cholesterol could induce dyslipidemia resulting in
cell injuries and intracellular lipid deposition.
Synaptic plasticity is an important characteristic
of the neural system that plays a significant role in maintaining
the structure and function of the neural system. BDNF is a member
of the neurotrophic factor family, and Copine-6 is a brain-specific,
calcium-dependent protein. These molecules play important roles in
regulating synaptic plasticity (55,56),
and their secretion and abundance are involved in the activity of
the Wnt/β-catenin signaling pathway (57). Moreover, BDNF participates in the
regulation of cholesterol homeostasis (58,59)
and improves nerve damage induced by high cholesterol levels
(58). Consistently, the
BDNF-related imbalance in the expression of Copine-6 and
synapse-associated proteins in the hippocampus and PFC of stressed
(60) or hyperlipidemic (14,46)
rats has been demonstrated in our previous studies. In line with
these findings, the results from the present study showed that
cholesterol-induced downregulation of BDNF and Copine-6 protein
expression in SH-SY5Y cells, and the expression of these molecules
was positively correlated with each other. These results again
suggest the role of BDNF-related Copine-6 expression in high
cholesterol-related neural injury.
Nesfatin-1 may be involved in mediating metabolic
disorders and neuropsychiatric injury. Our previous study showed
that the plasma concentration of nesfatin-1 was markedly increased
in high-fat diet-induced nonalcoholic fatty liver disease rats and
was significantly correlated with impaired learning and memory
abilities and imbalanced protein expression of BDNF in the
hippocampus (46). In line with
this finding, the protein expression levels of nesfatin-1 in
SH-SY5Y cells were upregulated after stimulation with high
cholesterol. In addition, SH-SY5Y cells stimulated with high
cholesterol showed decreased expression of p-β-catenin, a key
molecule in the Wnt signaling pathway, and increased expression of
p-GSK3β. The results from the correlation analysis showed that the
expression of nesfatin-1 was positively correlated with the
expression of p-GSK3β and Cyclin-D1 and negatively correlated with
the expression of p-β-catenin. This result was consistent with the
previous finding that cholesterol metabolism is closely related to
the activity and function of the Wnt signaling pathway (61).
Given the findings that the variants of TREM2 could
markedly increase the risk of AD, this receptor has been a major
focus in the neuroscience (62-64).
Although studies have shown that TREM2 is exclusively expressed in
microglia (62), double
immunofluorescence staining has demonstrated that TREM2 is
expressed in microglia (ionized calcium-binding adapter molecule
1), astrocytes (glial fibrillary acidic protein), and neurons
(neuronal nucleus) in the brain (35,65).
TREM2 is required by microglia to respond to Aβ
deposition and to limit neuronal degeneration, and microglia fail
to colocalize with Aβ plaques in TREM2-/- mice (9). Moreover, TREM2 could regulate
microglial cholesterol metabolism upon chronic phagocytic
challenge, and loss of TREM2 or apolipoprotein E may result in
dysregulated cholesterol transport and metabolism in microglia
(13,66). TREM2 deletion not only causes
cholesterol ester overload in microglia but also abrogates the
recruitment of macrophages to enlarged adipocytes, leading to
massive adipocyte hypertrophy, systemic hypercholesterolemia,
inflammation, and glucose intolerance (67). These findings suggest that brain
cholesterol metabolism and lipid signaling through TREM2 play major
roles in age-related neurodegeneration processes (18,67).
However, little is known regarding the role of TREM2 in cholesterol
metabolism in neurons. Using SH-SY5Y cells, Huang et al
(54) demonstrated that
cholesterol overload in neuronal cells could result in an imbalance
in cholesterol homeostasis and increase protein expression levels
of truncated tyrosine kinase B, flotillin-2, Beta-Secretase (BACE)
and β-amyloid (Aβ) and thereby inducing cell apoptosis, and the
underlying mechanism involved the downregulation of BDNF.
Consistent with these findings, the results of the present study
showed that in addition to decreased viability, increased lipid
accumulation, and imbalanced synaptic plasticity in SH-SY5Y cells,
high cholesterol induced a decrease in the protein expression
levels of TREM2. Moreover, the dysregulated expression of BDNF,
Copine-6, and p-GSK3 in SH-SY5Y cells induced by cholesterol could
be reversed by overexpression of TREM2. Taken together with the
findings that microglia in the brains of TREM2-/- mice
could phagocytose myelin debris but failed to clear myelin
cholesterol, resulting in cholesteryl ester accumulation (66), and the significant correlation
between TREM2 protein expression and BDNF, Copine-6 and nesfatin-1,
these results suggested that TREM2 also plays a major role in
cholesterol metabolism in neuronal cells.
This study has several limitations that should be
addressed in future studies. First, only one cell line was used in
this study, and the role of TREM2 in nerve injury induced by high
cholesterol should be verified using additional types of neuronal
cells, particularly primary neurons. Second, the effect of high
cholesterol on the protein expression levels of BDNF, Copine-6, and
key proteins in the Wnt/β-catenin signaling pathway has been
detected regardless of the overexpression of TREM2. In addition, an
explanation for the fact that TREM1 and TREM2 are inversely
correlated with each, as in other experimental models, could not be
provided, and the accurate mechanism should be explored in detail
in the future.
In conclusion, the results of the present study
suggest that a high concentration of cholesterol can induce cell
injury in SH-SY5Y cells, and that an imbalance expression in TREM2
may play an important role in these injuries. A summary schematic
is shown in Fig. 8.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the National Natural
Science Foundation of China (grant nos. 81870403 and 81701327), the
Top Talents in University of Anhui Province (grant no.
gxbjZD2022013), the Scientific Research Promotion Plan of Anhui
Medical University (grant no. 2022xkjT009), and the Comprehensive
Reform Pilot Project of ‘San Quan Education’ of Anhui Medical
University (grant no. 2021xsqyr03).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JG, ZQ and YH conceived and designed the study. YH
and QZ performed the experiments, analyzed the data, and drafted
the manuscript. MF, MM, and JX analyzed and interpreted the data.
XG and SL performed cell culture. JG, YH and QZ confirm the
authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sun L, Clarke R, Bennett D, Guo Y, Walters
RG, Hill M, Parish S, Millwood IY, Bian Z, Chen Y, et al: Causal
associations of blood lipids with risk of ischemic stroke and
intracerebral hemorrhage in Chinese adults. Nat Med. 25:569–574.
2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Yusuf S, Bosch J, Dagenais G, Zhu J,
Xavier D, Liu L, Pais P, Lopez-Jaramillo P, Leiter LA, Dans A, et
al: Cholesterol lowering in intermediate-risk persons without
cardiovascular disease. N Engl J Med. 374:2021–2031.
2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Alenghat FJ and Davis AM: Management of
blood cholesterol. JAMA. 321:800–801. 2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Grundy SM, Stone NJ, Bailey AL, Beam C,
Birtcher KK, Blumenthal RS, Braun LT, de Ferranti S,
Faiella-Tommasino J, Forman DE, et al: 2018
AHA/ACC/AACVPR/AAPA/ABC/ACPM/ADA/AGS/APhA/ASPC/NLA/PCNA Guideline
on the Management of Blood Cholesterol: A Report of the American
College of Cardiology/American Heart Association Task Force on
Clinical Practice Guidelines. Circulation. 139:e1082–e1143.
2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Wilson PWF, Polonsky TS, Miedema MD, Khera
A, Kosinski AS and Kuvin JT: Systematic Review for the 2018
AHA/ACC/AACVPR/AAPA/ABC/ACPM/ADA/AGS/APhA/ASPC/NLA/PCNA Guideline
on the Management of Blood Cholesterol: A Report of the American
College of Cardiology/American Heart Association Task Force on
Clinical Practice Guidelines. Circulation. 139:e1144–e1161.
2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Holt RI, Phillips DI, Jameson KA, Cooper
C, Dennison EM and Peveler RC: Hertfordshire Cohort Study Group.
The relationship between depression, anxiety and cardiovascular
disease: Findings from the Hertfordshire Cohort Study. J Affect
Disord. 150:84–90. 2013.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Stough C, Pipingas A, Camfield D, Nolidin
K, Savage K, Deleuil S and Scholey A: Increases in total
cholesterol and low density lipoprotein associated with decreased
cognitive performance in healthy elderly adults. Metab Brain Dis.
34:477–484. 2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Song Y, Liu J, Zhao K, Gao L and Zhao J:
Cholesterol-induced toxicity: An integrated view of the role of
cholesterol in multiple diseases. Cell Metab. 33:1911–1925.
2021.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Zhu X, Tang HD, Dong WY, Kang F, Liu A,
Mao Y, Xie W, Zhang X, Cao P, Zhou W, et al: Distinct
thalamocortical circuits underlie allodynia induced by tissue
injury and by depression-like states. Nat Neurosci. 24:542–553.
2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zou Y, Zhu Q, Deng Y, Duan J, Pan L, Tu Q,
Dai R, Zhang X, Chu LW and Lu Y: Vascular risk factors and mild
cognitive impairment in the elderly population in Southwest China.
Am J Alzheimers Dis Other Demen. 29:242–247. 2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Reed B, Villeneuve S, Mack W, DeCarli C,
Chui HC and Jagust W: Associations between serum cholesterol levels
and cerebral amyloidosis. JAMA Neurol. 71:195–200. 2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Liu AH, Chu M and Wang YP: Up-Regulation
of Trem2 inhibits hippocampal neuronal apoptosis and alleviates
oxidative stress in epilepsy via the PI3K/Akt pathway in mice.
Neurosci Bull. 35:471–485. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Roca-Agujetas V, de Dios C, Abadin X and
Colell A: Upregulation of brain cholesterol levels inhibits
mitophagy in Alzheimer disease. Autophagy. 17:1555–1557.
2021.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Gao XR, Chen Z, Fang K, Xu JX and Ge JF:
Protective effect of quercetin against the metabolic dysfunction of
glucose and lipids and its associated learning and memory
impairments in NAFLD rats. Lipids Health Dis.
20(164)2021.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Leritz EC, McGlinchey RE, Salat DH and
Milberg WP: Elevated levels of serum cholesterol are associated
with better performance on tasks of episodic memory. Metab Brain
Dis. 31:465–473. 2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zhou F, Deng W, Ding D, Zhao Q, Liang X,
Wang F, Luo J, Zheng L, Guo Q and Hong Z: High Low-density
lipoprotein cholesterol inversely relates to dementia in
community-dwelling older adults: The shanghai aging study. Front
Neurol. 9(952)2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Banach M, Rizzo M, Nikolic D, Howard G,
Howard V and Mikhailidis D: Intensive LDL-cholesterol lowering
therapy and neurocognitive function. Pharmacol Ther. 170:181–191.
2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Bertolio R, Napoletano F, Mano M,
Maurer-Stroh S, Fantuz M, Zannini A, Bicciato S, Sorrentino G and
Del Sal G: Sterol regulatory element binding protein 1 couples
mechanical cues and lipid metabolism. Nat Commun.
10(1326)2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Oh IS, Shimizu H, Satoh T, Okada S, Adachi
S, Inoue K, Eguchi H, Yamamoto M, Imaki T, Hashimoto K, et al:
Identification of nesfatin-1 as a satiety molecule in the
hypothalamus. Nature. 443:709–712. 2006.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhang Z, Li L, Yang M, Liu H, Boden G and
Yang G: Increased plasma levels of nesfatin-1 in patients with
newly diagnosed type 2 diabetes mellitus. Exp Clin Endocrinol
Diabetes. 120:91–95. 2012.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Catak Z, Aydin S, Sahin I, Kuloglu T,
Aksoy A and Dagli AF: Regulatory neuropeptides (ghrelin, obestatin
and nesfatin-1) levels in serum and reproductive tissues of female
and male rats with fructose-induced metabolic syndrome.
Neuropeptides. 48:167–177. 2014.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Prinz P and Stengel A: Nesfatin-1: Current
status as a peripheral hormone and future prospects. Curr Opin
Pharmacol. 31:19–24. 2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Yin Y, Li Z, Gao L, Li Y, Zhao J and Zhang
W: AMPK-dependent modulation of hepatic lipid metabolism by
nesfatin-1. Mol Cell Endocrinol. 417:20–26. 2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Stengel A, Goebel M, Wang L, Rivier J,
Kobelt P, Mönnikes H, Lambrecht NW and Taché Y: Central nesfatin-1
reduces dark-phase food intake and gastric emptying in rats:
Differential role of corticotropin-releasing factor2 receptor.
Endocrinology. 150:4911–4919. 2009.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Yoshida N, Maejima Y, Sedbazar U, Ando A,
Kurita H, Damdindorj B, Takano E, Gantulga D, Iwasaki Y, Kurashina
T, et al: Stressor-responsive central nesfatin-1 activates
corticotropin-releasing hormone, noradrenaline and serotonin
neurons and evokes hypothalamic-pituitary-adrenal axis. Aging
(Albany NY). 2:775–784. 2010.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Han YX, Tao C, Gao XR, Wang LL, Jiang FH,
Wang C, Fang K, Chen XX, Chen Z and Ge JF: BDNF-related imbalance
of copine 6 and synaptic plasticity markers couples with
depression-like behavior and immune activation in CUMS rats. Front
Neurosci. 12(731)2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Xu YY, Ge JF, Qin G, Peng YN, Zhang CF,
Liu XR, Liang LC, Wang ZZ, Chen FH and Li J: Acute, but not
chronic, stress increased the plasma concentration and hypothalamic
mRNA expression of NUCB2/nesfatin-1 in rats. Neuropeptides.
54:47–53. 2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Ge JF, Xu YY, Qin G, Pan XY, Cheng JQ and
Chen FH: Nesfatin-1, a potent anorexic agent, decreases exploration
and induces anxiety-like behavior in rats without altering learning
or memory. Brain Res. 1629:171–181. 2015.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Ge JF, Xu YY, Qin G, Peng YN, Zhang CF,
Liu XR, Liang LC, Wang ZZ and Chen FH: Depression-like behavior
induced by nesfatin-1 in rats: Involvement of increased immune
activation and imbalance of synaptic vesicle proteins. Front
Neurosci. 9(429)2015.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Liu J, Xiao Q, Xiao J, Niu C, Li Y, Zhang
X, Zhou Z, Shu G and Yin G: Wnt/β-catenin signalling: function,
biological mechanisms, and therapeutic opportunities. Signal
Transduct Target Ther. 7(3)2022.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Schlupf J and Steinbeisser H: IGF
antagonizes the Wnt/β-Catenin pathway and promotes differentiation
of extra-embryonic endoderm. Differentiation. 87:209–219.
2014.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Xu X, Wang L, Liu B, Xie W and Chen YG:
Activin/Smad2 and Wnt/β-catenin up-regulate HAS2 and ALDH3A2 to
facilitate mesendoderm differentiation of human embryonic stem
cells. J Biol Chem. 293:18444–18453. 2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Logan CY and Nusse R: The Wnt signaling
pathway in development and disease. Annu Rev Cell Dev Biol.
20:781–810. 2004.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Yi H, Hu J, Qian J and Hackam AS:
Expression of brain-derived neurotrophic factor is regulated by the
Wnt signaling pathway. Neuroreport. 23:189–194. 2012.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Chen J, Park CS and Tang SJ:
Activity-dependent synaptic Wnt release regulates hippocampal long
term potentiation. J Biol Chem. 281:11910–11916. 2006.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Spagnuolo MS, Donizetti A, Iannotta L,
Aliperti V, Cupidi C, Bruni AC and Cigliano L: Brain-derived
neurotrophic factor modulates cholesterol homeostasis and
Apolipoprotein E synthesis in human cell models of astrocytes and
neurons. J Cell Physiol. 233:6925–6943. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Ge JF, Xu YY, Qin G, Cheng JQ and Chen FH:
Resveratrol ameliorates the anxiety- and depression-like behavior
of subclinical hypothyroidism rat: Possible Involvement of the HPT
Axis, HPA Axis, and Wnt/beta-Catenin pathway. Front Endocrinol
(Lausanne). 7(44)2016.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Ford JW and McVicar DW: TREM and TREM-like
receptors in inflammation and disease. Curr Opin Immunol. 21:38–46.
2009.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Ulland TK and Colonna M: TREM2-a key
player in microglial biology and Alzheimer disease. Nat Rev Neurol.
14:667–675. 2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Zheng H, Jia L, Liu CC, Rong Z, Zhong L,
Yang L, Chen XF, Fryer JD, Wang X, Zhang YW, et al: TREM2 Promotes
Microglial Survival by Activating Wnt/β-Catenin Pathway. J
Neurosci. 37:1772–1784. 2017.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Fan Y, Ma Y, Huang W, Cheng X, Gao N, Li G
and Tian S: Up-regulation of TREM2 accelerates the reduction of
amyloid deposits and promotes neuronal regeneration in the
hippocampus of amyloid beta1-42 injected mice. J Chem Neuroanat.
97:71–79. 2019.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Jiang T, Tan L, Zhu XC, Zhang QQ, Cao L,
Tan MS, Gu LZ, Wang HF, Ding ZZ, Zhang YD and Yu JT: Upregulation
of TREM2 ameliorates neuropathology and rescues spatial cognitive
impairment in a transgenic mouse model of Alzheimer's disease.
Neuropsychopharmacology. 39:2949–2962. 2014.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Parhizkar S, Arzberger T, Brendel M,
Kleinberger G, Deussing M, Focke C, Nuscher B, Xiong M,
Ghasemigharagoz A, Katzmarski N, et al: Loss of TREM2 function
increases amyloid seeding but reduces plaque-associated ApoE. Nat
Neurosci. 22:191–204. 2019.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Jiang T, Tan L, Zhu XC, Zhou JS, Cao L,
Tan MS, Wang HF, Chen Q, Zhang YD and Yu JT: Silencing of TREM2
exacerbates tau pathology, neurodegenerative changes, and spatial
learning deficits in P301S tau transgenic mice. Neurobiol Aging.
36:3176–3186. 2015.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Jiang T, Wan Y, Zhang YD, Zhou JS, Gao Q,
Zhu XC, Shi JQ, Lu H, Tan L and Yu JT: TREM2 Overexpression has No
Improvement on Neuropathology and Cognitive Impairment in Aging
APPswe/PS1dE9 Mice. Mol Neurobiol. 54:855–865. 2017.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Chen Z, Xu YY, Wu R, Han YX, Yu Y, Ge JF
and Chen FH: Impaired learning and memory in rats induced by a
high-fat diet: Involvement with the imbalance of nesfatin-1
abundance and copine 6 expression. J Neuroendocrinol.
(29)2017.PubMed/NCBI View Article : Google Scholar : doi:
10.1111/jne.12462.
|
|
47
|
Arenas F, Garcia-Ruiz C and
Fernandez-Checa JC: Intracellular cholesterol trafficking and
impact in neurodegeneration. Front Mol Neurosci.
10(382)2017.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Dietschy JM and Turley SD: Thematic review
series: Brain Lipids. Cholesterol metabolism in the central nervous
system during early development and in the mature animal. J Lipid
Res. 45:1375–1397. 2004.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Schilling S, Tzourio C, Soumare A,
Kaffashian S, Dartigues JF, Ancelin ML, Samieri C, Dufouil C and
Debette S: Differential associations of plasma lipids with incident
dementia and dementia subtypes in the 3C Study: A longitudinal,
population-based prospective cohort study. PLoS Med.
14(e1002265)2017.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Chen H, Du Y, Liu S, Ge B, Ji Y and Huang
G: Association between serum cholesterol levels and Alzheimer's
disease in China: A case-control study. Int J Food Sci Nutr.
70:405–411. 2019.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Knowles TP, Vendruscolo M and Dobson CM:
The amyloid state and its association with protein misfolding
diseases. Nat Rev Mol Cell Biol. 15:384–396. 2014.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Habchi J, Chia S, Galvagnion C, Michaels
TCT, Bellaiche MMJ, Ruggeri FS, Sanguanini M, Idini I, Kumita JR,
Sparr E, et al: Cholesterol catalyses Abeta42 aggregation through a
heterogeneous nucleation pathway in the presence of lipid
membranes. Nat Chem. 10:673–683. 2018.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Di Scala C, Chahinian H, Yahi N, Garmy N
and Fantini J: Interaction of Alzheimer's β-amyloid peptides with
cholesterol: Mechanistic insights into amyloid pore formation.
Biochemistry. 53:4489–4502. 2014.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Huang YN, Lin CI, Liao H, Liu CY, Chen YH,
Chiu WC and Lin SH: Cholesterol overload induces apoptosis in
SH-SY5Y human neuroblastoma cells through the up regulation of
flotillin-2 in the lipid raft and the activation of BDNF/Trkb
signaling. Neuroscience. 328:201–209. 2016.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Reinhard JR, Kriz A, Galic M, Angliker N,
Rajalu M, Vogt KE and Ruegg MA: The calcium sensor Copine-6
regulates spine structural plasticity and learning and memory. Nat
Commun. 7(11613)2016.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Burk K, Ramachandran B, Ahmed S,
Hurtado-Zavala JI, Awasthi A, Benito E, Faram R, Ahmad H,
Swaminathan A, McIlhinney J, et al: Regulation of Dendritic Spine
Morphology in Hippocampal Neurons by Copine-6. Cereb Cortex.
28:1087–1104. 2018.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Zhang W, Shi Y, Peng Y, Zhong L, Zhu S,
Zhang W and Tang SJ: Neuron activity-induced Wnt signaling
up-regulates expression of brain-derived neurotrophic factor in the
pain neural circuit. J Biol Chem. 293:15641–15651. 2018.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Motamedi S, Karimi I and Jafari F: The
interrelationship of metabolic syndrome and neurodegenerative
diseases with focus on brain-derived neurotrophic factor (BDNF):
Kill two birds with one stone. Metab Brain Dis. 32:651–665.
2017.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Sharma D, Barhwal KK, Biswal SN,
Srivastava AK, Bhardwaj P, Kumar A, Chaurasia OP and Hota SK:
Hypoxia-mediated alteration in cholesterol oxidation and raft
dynamics regulates BDNF signalling and neurodegeneration in
hippocampus. J Neurochem. 148:238–251. 2019.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Sayed FA, Telpoukhovskaia M, Kodama L, Li
Y, Zhou Y, Le D, Hauduc A, Ludwig C, Gao F, Clelland C, et al:
Differential effects of partial and complete loss of TREM2 on
microglial injury response and tauopathy. Proc Natl Acad Sci USA.
115:10172–10177. 2018.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Sheng R, Kim H, Lee H, Xin Y, Chen Y, Tian
W, Cui Y, Choi JC, Doh J, Han JK and Cho W: Cholesterol selectively
activates canonical Wnt signalling over non-canonical Wnt
signalling. Nat Commun. 5(4393)2014.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Damisah EC, Hill RA, Rai A, Chen F,
Rothlin CV, Ghosh S and Grutzendler J: Astrocytes and microglia
play orchestrated roles and respect phagocytic territories during
neuronal corpse removal in vivo. Sci Adv.
6(eaba3239)2020.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Wang S, Mustafa M, Yuede CM, Salazar SV,
Kong P, Long H, Ward M, Siddiqui O, Paul R, Gilfillan S, et al:
Anti-human TREM2 induces microglia proliferation and reduces
pathology in an Alzheimer's disease model. J Exp Med.
217(e20200785)2020.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Zhou Y, Song WM, Andhey PS, Swain A, Levy
T, Miller KR, Poliani PL, Cominelli M, Grover S, Gilfillan S, et
al: Human and mouse single-nucleus transcriptomics reveal
TREM2-dependent and TREM2-independent cellular responses in
Alzheimer's disease. Nat Med. 26:131–142. 2020.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Diaz-Lucena D, Kruse N, Thüne K, Schmitz
M, Villar-Piqué A, da Cunha JEG, Hermann P, López-Pérez Ó,
Andrés-Benito P, Ladogana A, et al: TREM2 expression in the brain
and biological fluids in prion diseases. Acta Neuropathol.
141:841–859. 2021.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Nugent AA, Lin K, van Lengerich B,
Lianoglou S, Przybyla L, Davis SS, Llapashtica C, Wang J, Kim DJ,
Xia D, et al: TREM2 regulates microglial cholesterol metabolism
upon chronic phagocytic challenge. Neuron. 105:837–854.e839.
2020.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Jaitin DA, Adlung L, Thaiss CA, Weiner A,
Li B, Descamps H, Lundgren P, Bleriot C, Liu Z, Deczkowska A, et
al: Lipid-Associated macrophages control metabolic homeostasis in a
Trem2-Dependent manner. Cell. 178:686–698.e14. 2019.PubMed/NCBI View Article : Google Scholar
|