Introduction
Itching is ‘an unpleasant sensation that evokes a
desire to scratch’ (1). ~10-20% of
individuals will cope with itching in their lifetime, which
seriously affects the patients' quality of life, mood, and sleep
(2-4).
According to a survey of global disease, the economic burden caused
by chronic pruritus ranks it among the top 50 most serious diseases
(5). However, the itch has been
severely underestimated in the past several decades, resulting in
vague mechanisms of itch development. Without effective drugs in
clinical application, pruritus has become an urgent clinical
problem to be solved.
Women have unique hormonal changes over time, which
may be correlated with the periodic changes in the elemental
composition of the skin throughout the age range. Notably, it has
been also reported that the incidence of pruritus increases in
postmenopausal women (6), and up
to 55% suffer from pruritus and dryness associated with chronic
skin diseases including psoriasis, dermatitis and infection,
causing severe discomfort (7,8).
Changes in estrogen levels may affect the hydration, collagen
content and glycosaminoglycan concentration in the skin (9,10),
and the number and types of estrogen receptors expressed by
epithelial keratinocytes (11).
Notably, it was shown that estrogen receptors are also widely
expressed in the dorsal root ganglion (DRG) and superficial dorsal
horn (12,13). The application of estrogen causes
changes in intracellular Ca2+ concentrations in DRG
cells of sensory neurons by binding to receptors and is involved in
the transduction of nociception (14). However, whether and how estrogen is
involved in developing chronic itch in females still needs to be
understood in an improved way.
Unmyelinated small-diameter C fibres are critically
involved in the generation of itch sensation (15-17).
Among them, three pruriceptive small-diameter DRG neuron subtypes
have been identified by both functional studies and RNA-sequencing
(RNA-seq) analysis. NP1 neurons are classified by the expression of
Mas-related G-protein coupled receptor (Mrgpr)D, NP2 neurons are
classified by the expression of MrgprA3 and MrgprC11, and NP3
neurons are rich in other unique markers, including the
neuropeptides natriuretic polypeptide b (Nppb), neurotensin and
somatostatin. To further characterize the relationship between
estrogen and the expression of those itch-related genes, RNA-seq
analysis was employed and the key components in hormonal
change-related itch sensation development were determined.
In summary, it was aimed to decipher the function of
estrogen in acute and chronic itch. In a behavioural study,
estrogen application rescued the increased scratching bouts in
ovariectomized mice in both acute and chronic itch models. RNA-seq
analysis indicated that loss of estrogen expression in the
ovariectomized mice resulted in significantly increased levels of
itch signalling molecules, and this effect could be reversed by the
implantation of estrogen sustained-release tablets. The present
study uncovered the critical role of estrogen in itch modulation
and provided a reliable theoretical basis for developing safe and
effective drugs for treating pruritus.
Materials and methods
Animals
A total of 50 healthy C57BL/6J female mice (9~10
weeks-old; weight, 20~25 g) were purchased from Beijing Vital River
Laboratory Animal Technology Co., Ltd. Mice were housed in an SPF
environment, maintained at room temperature at 23~25˚C with a
diurnal 12 h rhythm, and had free access to water and food.
Experiments were performed one week after adaptive feeding.
All animal procedures conformed to the Guide for the
Care and Use of Laboratory Animals of the National Institutes of
Health, and all experiments complied with the Animal Research:
Reporting in vivo Experiments (ARRIVE) guidelines. The
present study was approved (approval no. ZX201898) by the Animal
Experimental Ethics Committee of the Plastic Surgery Hospital,
Chinese Academy of Medical Sciences (Beijing, China).
Experimental design
The aim of the present experiment was to investigate
the effect of estrogen on pruritus at a slightly higher than
physiological level. Embedding estrogen sustained-release tablets
can result in a relatively stable estrogen level in mice. By
observing the state of the mice, measuring the mouse body weight
(Fig. S1), detecting the activity
of the mice with the rotarod test (Fig. S2), and synthesizing the previous
research, it was considered that this dose of estrogen will not
affect the health of the mice. The mice exhibited favorable
recovery, and their health status was also favorable. To achieve
the purpose of the experiment, an appropriate dose of estrogen
sustained-release tablets should be selected. According to previous
studies (11,18), three doses of estrogen
sustained-release tablets were used in the preliminary experiment:
0.36, 0.1 and 0.01 mg per tablet. The levels of estrogen in the
body were 402±86.39, 109±2.89, and 24.64±12.10 pg/l, respectively.
Finally, 0.1 mg was selected as the experimental dose.
Estrogen replacement
Ovariectomy via the dorsal route bilaterally.
Anesthesia was induced with 80 mg/kg ketamine plus 8 mg/kg xylazine
(19). The hair was removed, and a
smooth incision ~1 cm long was made in the skin. The skin and
subcutaneous tissue were separated. The color of the ovary is
slightly lighter than that of the surrounding muscle. An incision
of ~0.5 cm was made in that location, which exposed the adipose
tissue surrounding the ovary and the uterine horn closely connected
to the ovary. Curved forceps were used to gently clamp the adipose
tissue to pull it away from the wound. Then, the fallopian tube was
ligated at the upper and lower uterine horns, the uterine horn was
cut off, and the ovary was removed and dissected (20). Then, estrogen sustained-release
tablets or placebo tablets were implanted into the ovary position
simultaneously. The adipose tissue was put back into its original
position, and 7.5 mg/kg Rimadyl was diluted in saline injection
subcutaneously before the mouse woke up (21,22).
The subcutaneous tissue and skin were sutured, and the sutured
wound surface was disinfected with iodophor. After surgery, the
mice were placed in a cage at 30˚C separately. A total of 80% of
the mice could wake up within 1-2 h, the remaining other mice could
wake up within 3 h at the latest, and the wounds could heal in 3
days. Absorbable sutures were used for intradermal sutures, and the
sutures of the skin fell off within 4-6 days as the mice moved, and
new hair gradually grew at the sutures. No abnormalities were
observed after one week. The cage was kept dry and the wounds were
checked every day.
Slow-release pellet of 17β-estradiol. Based
on our preliminary test results and previous studies (11,18),
a slow-release pellet of 17β-estradiol (0.1 mg/pellet, 60-day
release, Innovative Research of America) or the placebo was placed
back in the ovary position, respectively. These are tablets with a
diameter of 3 mm and a release cycle of 60 days, or placebo tablets
of the same texture and size. The pellets produce a slow release of
the active drug over 60 days (1.7 µg per day) and have been shown
to maintain estrogen plasma levels of ~125 pg/ml for the duration
of the pellet. The results of the plasma estradiol concentrations
are shown in Fig. S3.
Itch behavioral tests
Acute itch behavioral tests. The acute itch
behavior tests were performed similarly to a previous study
(23). Intradermal injection with
different pruritogens was used to evaluate the acute itch behavior.
Pruritogens included 200 µg histamine, 100 µg chloroquine (CQ), 50
µg compound 48/80, 50 nmol 5-hydroxytryptamine (5-HT) and 100 nmol
SLIGRL-NH2 dissolved in phosphate buffered saline, respectively.
The pruritogens doses used in the acute itch experiments were
examined in the preliminary experiments, and the data are
demonstrated in Fig. S4.
Protocols of estrogen effect on acute itch. A
total of 50 mice were used in the acute itch tests, which included
five experiments. In every experiment, 10 littermate mice were
randomly assigned to 2 groups, namely the estrogen group (E group,
n=5) and placebo group (P group, n=5); the E group was implanted
with estrogen sustained-release tablets, and the P group was
implanted with the placebo tablets. The acute behavioural tests
were completed within one week.
Chronic itch behavioral tests. The
acetone/ether/water (AEW) model was established as the chronic itch
model (24,25). The mice were shaved (2x2 cm) to
expose the skin, and the exposed skin was covered with
acetone-ether mixed liquid (AE)-soaked cotton for 15 sec, followed
by water-soaked cotton for 30 sec twice a day at 9 am and 5 pm for
seven days. The skin was observed and images were captured to
assess the dry-skin-evoked changes, including excoriation and dry
scales. Under sterile conditions, the skin of each group was
collected, H&E staining was performed at room temperature
(stain with haematoxylin 5 min and stain with eosin 1 min), the
samples were observed under a light microscope (DM3000; Leica
Microsystems), and images were recorded. The relative thickness of
the epidermis was analyzed by Image-Pro Plus 6.0 (Media
Cybernetics).
Protocols of estrogen effect on chronic itch.
A total of 10 days after the ovariectomy and implanted surgery, 16
littermate mice were randomized into two groups, the E group (n=8)
and the P group (n=8); the grouping method was the same as the
acute itch behavioural test. The AEW chronic itch model was
completed within ten days when the chronic itch model was
successfully established. A total of six mice in each group were
used for the RNA-seq analysis, and two mice in each group were
subjected to H&E staining.
How animal count was decided. Behavioral
experiments were performed by recording the number of bouts of
scratching. The hair on the nape of the neck was shaved three days
prior to the itch behavior test. The mice were placed in behavior
chambers for 30 min for acclimation before the acute or chronic
itch behavior test. A scratching bout is defined as follows
(26): The mice raise the hind leg
towards the treated sites (chemical-injected site in acute itch and
AEW-treated site in chronic itch), scratching once or continuously
scratching for several times and then put down the leg. This series
of movements was counted as one scratching bout. Scratching towards
other parts is omitted.
Microarray analyses of the DRG
A total of 15 days after ovariectomy and estrogen
replacement surgery, DRG neurons on both sides of the lower
cervical and upper thoracic segments (C4~T2) of the mice were
chosen. According to the manufacturer's instructions, total RNA was
extracted from the mice using the RNeasy Mini Kit (cat. no. 74104;
Qiagen) and RNA-sequencing was analyzed by Novogene Co., Ltd.
FeatureCounts v1.5.0-p3 was used to count the reads numbers mapped
to each gene and then FPKM of each gene was calculated based on the
length of the gene and reads count mapped to this gene.
Differential expression analysis of two conditions was performed
using the DESeq2 package (1.16.1). The P-values were adjusted using
the Benjamini & Hochberg method (27). All differentially expressed genes
were determined by Padj <0.05.
Calcium imaging test. Isolation and
preparation of DRG cells
Nerve fibers were cut off, and the DRG was minced
and placed in calcium-free and magnesium-free (Hanks' balanced salt
solution; HBSS) buffer followed by 5 ml of digestive enzymes, which
contained 0.5 mg/ml trypsin (Type III; Thermo Fisher Scientific,
Inc.), 1 mg/ml collagenase (Type IA, Sigma-Aldrich, USA.) and 0.1
mg/ml DNase (Type III, Sigma-Aldrich, USA.), and placed in a 37˚C
incubator for rough stirring at a speed of 10~11 rpm for no more
than 30 min. Then, the samples were centrifuged at 4˚C (60 x g; 2
min), and the digestive enzymes were aspirated. Next, 1 ml NBM
culture medium was added, the samples were gently stirred and
aspirated and allowed to stand for 1~2 min, and the supernatant was
aspirated and placed into a 15 ml centrifuge tube. The samples were
repeatedly stirred and aspirated three times, after which the
precipitate was discarded. The samples were centrifuged at 800 rpm
for 4 min, and the supernatant was discarded. NBM culture medium
was added, the samples were stirred evenly and aspirated, the
dispersed cells were added to the cell climbing sheet coated with
poly-lysine, and the samples were precipitated in incubators at
37˚C and 5% CO2 for 1 h. Another 2 ml of NBM culture
medium was added to the culture dish, and the samples were cultured
overnight at 37˚C in a 5% CO2 incubator. The cells were
divided into a control group (C group) and an estrogen group (E
group), and 100 nM β-estradiol was added to the estrogen group.
Calcium imaging. Before calcium imaging
experiments, cultured DRG neurons were loaded with 4 µM Fura-2 AM
(Life Technologies) for 60 min at 37˚C. Cells were washed three
times and placed in HBSS for 30 min at room temperature. During the
experiment, fluorescence at 340 and 380 nm excitation was imaged
using an inverted Nikon Ti-E microscope controlled by NIS-elements
imaging software (Naikon Instruments, Inc.). The Fura-2 ratio
(F340/F380) was used to reflect the change in
[Ca2+]i after stimulation, and the activation
threshold was defined as 3 SD above the mean (~20% above
baseline).
Plasma estradiol concentrations. Plasma
estradiol concentrations were tested by Radioimmunoassay by the
Beijing North Institute of Biotechnology company. Mice in the E and
P group were enucleated bilaterally under sevoflurane anesthesia,
blood was instilled into an EP tube, allowed to stand at room
temperature for more than 1 h, centrifuged speed at x1,900 x g for
10 min, and serum was removed and rapidly frozen at -70˚C. This kit
uses competitive radioimmunoassay to perform competitive binding
reactions between radiolabeled antigen and non-labeled antigen to
be tested and a limited amount of specific antibody simultaneously.
By separating unbound labeled antigens, the radioactivity count of
the labeled antigen-antibody complex is determined. The content of
the substance to be exmined in the sample is calculated using the
standard curve and the mathematical model of RIA (28). Plasma estradiol concentrations were
tested twice on 10th day after implanted surgery and when all the
experiments were finished.
Euthanasia
After mice were anaesthetized with 7-8% sevoflurane
by a delivery machine at an airflow rate of 1 l/min for 5-7 min
when the experiments were completed, reflexes disappeared,
respiratory arrest and pupil dilation occurred, and the mice were
euthanized by cervical dislocation.
Statistical methods
Statistical analysis was performed with GraphPad
Prism 8 (GraphPad Software, Inc.). All data are expressed as the
mean ± SEM. Unpaired two-tailed Student's t-test or one-way
analysis of variance followed by Dunnett's was used to evaluate
differences. P<0.05 was considered to indicate a statistically
significant difference for all tests.
Results
Estrogen mitigates chemical-induced
acute itch sensation in mice
The ovariectomy was first performed and tablets were
implanted, either the estrogen sustained-release or placebo
pallets. Then, the acute itch model was established by intradermal
injection of histamine, CQ, SLIGRL, compound 48/80 and 5-HT in the
P group and E group. The number of scratches was counted by
individuals unaware of the grouping.
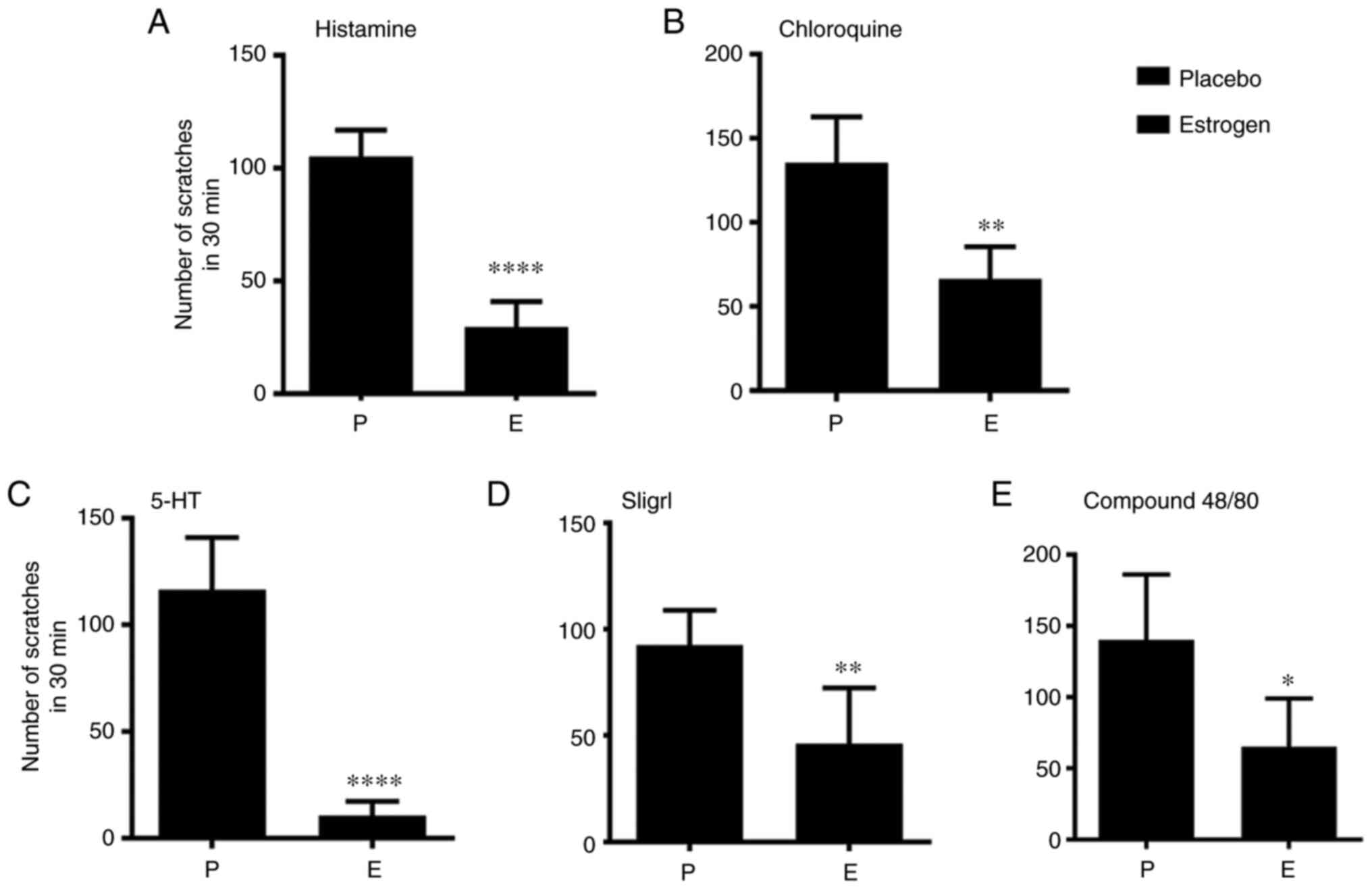
Estrogen treatment significantly reduced the number
of scratches in mice injected with histamine (P<0.05) (Fig. 1A), CQ (P<0.01) (Fig. 1B), 5-HT (P<0.01) (Fig. 1C), SLIGRL (P<0.05) (Fig. 1D) and compound 48/80 (P<0.01)
(Fig. 1E), suggesting that
estrogen treatment mitigates chemical-induced acute itch
sensation.
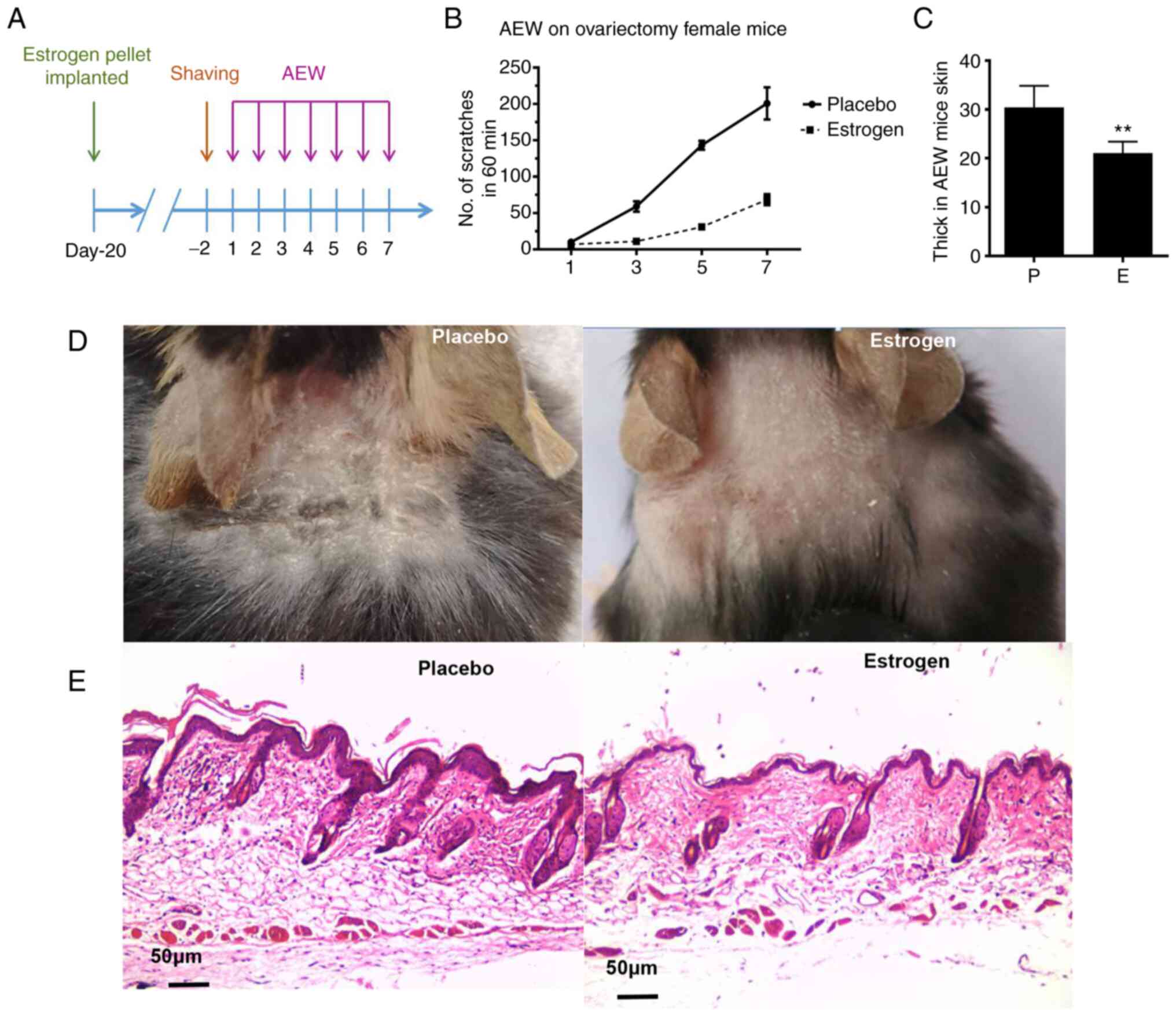
Estrogen attenuates chronic itch in a
mouse model of AEW
Next, the effects of estrogen on scratching behavior
in the AEW mouse model were evaluated; a schematic experimental
protocol is presented in Fig. 2A.
Notably, the number of scratches was reduced in the estrogen
treatment group compared with the placebo group over the 7-day
period. (P<0.001); (Fig.
2B). Moreover, AEW-induced skin hyperproliferation was also
reduced, as the thickness of the epidermal layers was significantly
thinner in the E group than that in the P group (Fig. 2C and E) and photo displaying the area of
treatment in the cervical back of mice, hyperplasia and dryness is
milder in the estrogen group than in the placebo group (Fig. 2D).
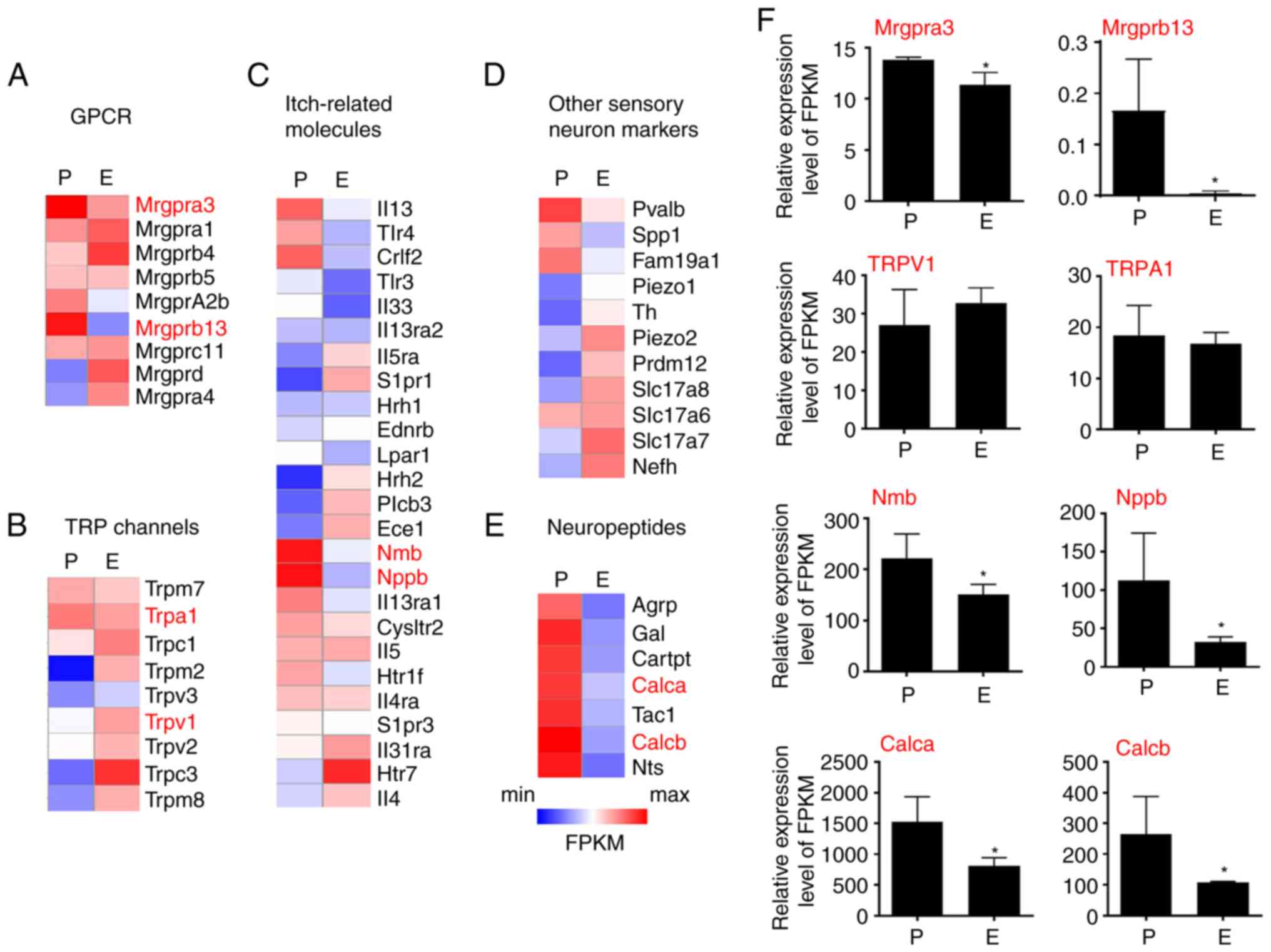
Estrogen suppresses itch-related gene
expression in the DRG
To exclude the endogenous expression of estrogen,
the ovariectomy was first performed and sustained-release tablets
filled with either vehicle control or estrogen were implanted in
mice. Then, the overall effect of estrogen on gene expression was
assessed by RNA-seq analysis in the placebo group (P) and the
estrogen group (E). The gene expression pattern of known genes
involved in itch sensation was examined. The first group analyzed
was GRCR family members. Strikingly, MrgprA3, a well-known
pruritogen receptor in both acute and chronic itch, was highly
enriched in the P group compared with the E group. The expression
of some other Mrgprs with unknown functions, such as MrgprB13, was
also reduced in the E group (Fig.
3A).
A total of 25 known genes that are essential to itch
sensation were then analyzed. Although the expression of two
histamine receptors (Hrh1 and Hrh2) was not affected by estrogen,
neuromedin B and Nppb, the neuropeptides required for scratching
behaviors induced by histamine, compound 48/80, and 5-HT were
significantly reduced in the E group compared with the P group,
suggesting that estrogen treatment may be involved in itch signal
transduction (Fig. 3C).
Transient Receptor Potential (TRP) channels are a
class of cationic channels that act as signal transducers by
altering membrane potential or intracellular calcium concentration
and function as important initiators in itch sensation. The current
data showed that neither TRPA1 nor TRPV1 was affected by estrogen
treatment. Notably, TRPC3 expression was significantly upregulated
in the E group. However, its role in itch remains to be determined
(Fig. 3B).
Numerous other sensory neuro markers were also
analyzed but did not provide any informative results (Fig. 3D). In addition, it was found that
several neuropeptides, including Calca, TAC1 and Calcb (Fig. 3E), were reduced in the E group,
suggesting a potential role of estrogen in the modulation of
somatic sensation and neurogenic inflammation (Fig. 3F).
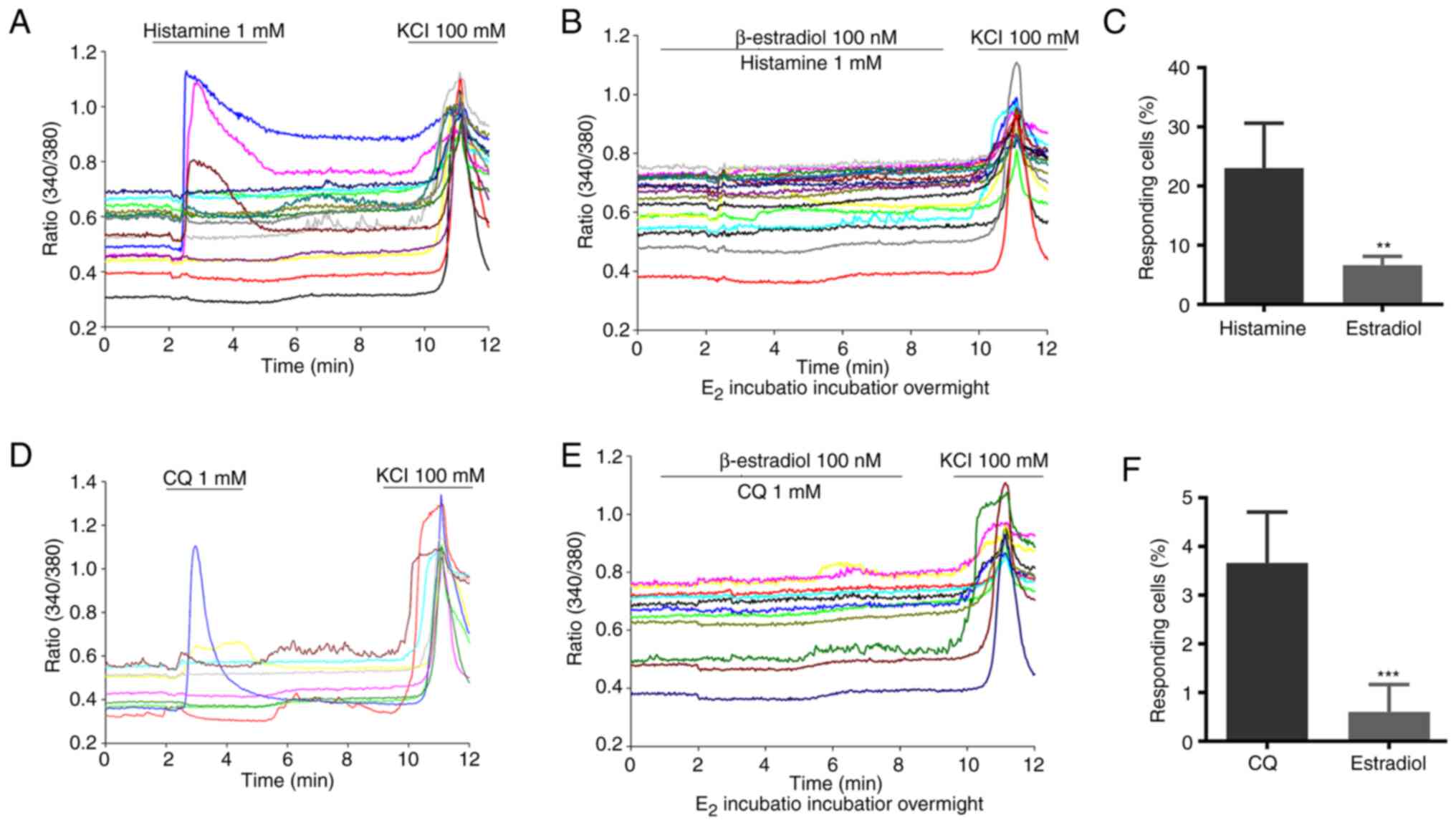
Estradiol attenuates chemical-induced
calcium influx in mouse DRG neurons
To elucidate the cellular basis of the inhibitory
effect of estrogen on itch sensation, cultured mouse DRG neurons
were first incubated in 100 nM β-estradiol overnight. As revealed
in Fig. 4, histamine or CQ
elicited a significant neuronal Ca2+ influx in the
vehicle-treated DRG neurons. By marked contrast, 100 nM β-estradiol
treatment abolished histamine or CQ-induced calcium influx DRG
neurons. Taken together, these data suggested that estrogen
treatment could reduce pruritogen-induced DRG neuron
activation.
Discussion
The acute chemical itch can be relieved by
scratching or antihistamine therapy, but the chronic itch is
challenging to cure because the mechanisms are unclear, and the
chronic itch is resistant to conventional treatments (e.g.,
antihistamines) (29,30). Numerous patients suffer from
chronic itch, and the development of effective antipruritic drugs
is imminent (31). Numerous
studies have shown that estrogen is involved in the regulation of
pruritus, including clinical research, animal experiments and
cellular experiments (8,32), but its specific mechanism remains
unknown. In the present study, it was revealed that estrogen could
suppress the expression of itch-related receptors and neuropeptides
and inhibit pruritogen-induced calcium influx in DRG neurons,
contributing to both acute and chronic itch relief in
estrogen-treated mice.
To determine the role of estrogen in modulating the
expression of itch-related genes, spinal DRG neurons were isolated
for high-throughput sequencing. It was found that high
concentrations of estrogen caused a large number of gene expression
changes in mice. Among the numerous genes with changes, MrgprA3,
but not TRPA1 or TRPV1, displayed significantly different
expression levels between the P group and E group, which was
consistent with behavioral results. MrgprA3 is a receptor for the
antimalarial drug CQ that directly activates DRG neurons, causing
site-directed scratching in response to CQ (33). Since multiple itch receptors are
highly enriched in MrgprA3-positive neurons, MrgprA3-positive
neurons play critical roles in the itch responses induced by at
least four different pruritogens, including CQ, histamine, BAM8-22
and SLIGRL-NH2, as well as in AEW-induced chronic itch (34). Although it was shown that estrogen
induced upregulation of TRPA1 and TRPV1 in the rat endometrium
(35), no change was detected in
mouse DRG neurons, suggesting that estrogen may modulate TRP
channel expression with regional specificity.
To explore the role of estrogen in itch sensation,
ovariectomies were performed in mice and the in vivo
estrogen concentration was manipulated by embedding β-estradiol
sustained-release tablets or a placebo. With this approach, it was
surprisingly found that high estrogen levels can reduce pruritic
sensation in not only the acute itch model but also the chronic
itch model. This result is consistent with numerous clinical
observations that ~50% of elderly women develop vulvar pruritus
(36), which is closely correlated
with the reduction of estrogen levels in serum, and the local
application of low-dose estrogen can safely and effectively treat
vulvar pruritus (37). A
multicenter, randomized, double-blind study of 550 female patients
for 12 weeks showed that topical estrogen could also effectively
relieve vulvar dryness and reduce pruritic sensation in patients
(7).
The DRG is an important site of visceral afferent
convergence and cross-sensitization (38,39)
and mediates somatic sensation, including pain, touch and itching
(40). The present experimental
results revealed that estrogen inhibits histamine-induced calcium
influx in neuronal cells supplemented with estrogen in calcium
imaging buffer. These results indicated that estrogen reduces the
response of neuronal cells. Estrogen binding to different receptors
in different parts of the body can produce a variety of biological
effects (41,42). A large number of studies support
that estrogen attenuates nociceptive responses by regulating the
intracellular calcium concentration [Ca2+]i
in DRG neurons (43). For example,
ATP/capsaicin-induced nociceptive responses can cause increased
intracellular calcium concentrations in DRG cells and cause
significant [Ca2+]i responses, and
β-estradiol (E2) inhibits nociceptive responses caused by
TRPV1-induced calcium influx in adult rat nociceptor neurons by
regulating estrogen receptor β-signaling (44); it has also been shown that E2 can
gate primary afferent responses to increase or decrease
nociception, depending on the input of nociceptive signals
(45). These results indicate
various functions of estrogen in different sensory
neuron-innervated areas, which are related to the cell type,
location and proportion of estrogen receptors expressed by the
innervating neurons (46), further
illustrating the complexity of estrogen regulation in the
peripheral nervous system.
The present study revealed that estrogen could
modulate the expression of itch-related signaling and inhibit both
acute and chronic itch in mice. Notably, Takanami et al
(32) showed that ovariectomy
females treated with 17β-estradiol had a serum concentration
estradiol of 60.18±13.54 pg/ml. These researchers further
demonstrated that estradiol modulated the GRP-GRPR signalling at
the spinal cord level. By marked contrast, in the present study the
dose of 17β-estradiol was further increased up to 100 pg/ml in the
serum. Notably, with the RNA-seq technique, it was identified that
a higher concentration of 17β-estradiol suppresses the expression
of itch-related genes in the peripheral DRG level. Combined with
the aforementioned results, these data together revealed that the
inhibitory or excitatory roles of 17β-estradiol are tightly
correlated with their serum concentration, implicating the
complexity of 17β-estradiol in itch modulation.
Moreover, estrogen acts in vivo mainly
through three receptors, ERα, ERβ and GPER, and estrogen receptor
expression has been found in various systems throughout the body
(46,47), which may be caused by estrogen
acting on different receptors. More in-depth subsequent studies are
needed to clarify this issue. Furthermore, different animal
models\interventions\observation methods affect the effects of
estrogen (48,49), and there are some differences in
the present and previous studies (30): i) The experimental animals in
previous studies were rats, but mice were used in the current
study; ii) The previous researchers used estradiol from 15 mm
Silastic capsules containing crystalline 17β-estradiol with the
blood concentrations of estradiol within the normal range of the
estrus cycle in adult female rats from the previous studies. One to
two months after surgery, rats were used for behavioural analysis.
However, slow-release matrix pellets were used in the present
experiment, 0.1 mg/pellet, 60-day release, and plasma estradiol
concentrations were slightly higher than the normal range of the
estrus cycle. Behavioural analysis was performed 10 days after
surgery. Moreover, there are differences between the two kinds of
pellets according to a previous study (18). In the aforementioned study,
histamine was used at a dose of 3 g/100 µl, but the current dose
was 200 µg/50 µl, which is different. In summary, the effect of
estrogen on pruritus needs to be further explored, and
understanding the role of estrogen in pruritus can provide
theoretical support for understanding the generation and regulation
of pruritus and can also provide a theoretical basis for developing
personalized drugs for the treatment of pruritus.
Supplementary Material
Weight of female mice before and after
ovariectomy surgery in E group or P group. There was no significant
difference of the weight of mice between the two groups before and
after ovariectomy surgery on Days 0, 7, 12 and 15. n=5 mice for
each group. one-way ANOVA. Error bars represent SEM. E, estrogen;
P, placebo.
Rotrod test after ovariectomy surgery
in E group or P group. There was no significant difference of the
Motor coordination between the two groups after ovariectomy
surgery. n=5 mice for each group. one-way ANOVA. Error bars
represent SEM. E, estrogen; P, placebo.
Plasma estradiol concentrations in the
P and E group in AEW model female mice. After completed AEW model
in ovariectomized female mice, E group implanted with estrogen
pellets exhibited higher concentrations of plasma E compared with
that in ovariectomized female mice implanted with placebo pellets.
***P<0.001, n=5, Student's t-test. P, placebo; E,
estrogen; AEW, acetone/ether/water.
Dose dependency of pruritogen-induced
scratching number in wild-type C57 female mice. (A) The doses of
chloroquine groups were: 200 μg; 100 μg; 50
μg/50 μl, respectively, n=5. (B) The doses of
histamine groups were: 200 μg; 100 μg; 50 μg/
50 μl, respectively, n=5. (C) The doses of Sliglr groups
were: 100 nmol, 50 nmol and 20 nmol/50 μl, respectively,
n=5. (D) The doses of compound 48/80 groups were: 100 μg; 50
μg and 10 μg/50 μl, respectively, n=5. (E) The
doses of serotonin (5-HT) groups were: 100 nmol, 50 nmol and 30
nmol/50 μl, respectively, n=5. 5-HT,
5-hydroxytryptamine.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported in part by the Fund of
Plastic Surgery Hospital, Chinese Academy of Medical Sciences
(grant nos. Q2017001 and Z2018010) and the Fundamental Research
Funds for the Central Universities (grant no. 3332019059).
Availability of data and materials
The related RNA-seq data have been deposited to GEO
(accession number: GSE222059) and datasets used and/or analyzed
during the current study are available from the corresponding
author upon reasonable request.
Authors' contributions
FX, GY, JJ and LL designed the project. FX, JJ, LL
and GY wrote the manuscript. JJ and LL performed the experiments
and analyzed the data. JJ, KL, CW and LL performed behavioural
tests. YW, QuL and QiL assisted with Ca2+ imaging. WL
and AQ assisted with data analysis. YW, LL and KL assisted with
cell isolation experiments. All authors discussed the manuscript,
commented on the project and contributed to manuscript preparation.
JJ and LL confirm the authenticity of all the raw data. All authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved (approval no.
ZX201898) by The Animal Experimental Ethics Committee of the
Plastic Surgery Hospital, Chinese Academy of Medical Science
(Beijing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Meng J and Steinhoff M: Molecular
mechanisms of pruritus. Curr Res Transl Med. 64:203–206.
2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Dreno B, Amici JM, Demessant-Flavigny AL,
Wright C, Taieb C, Desai SR and Alexis A: The impact of acne,
atopic dermatitis, skin toxicities and scars on quality of life and
the importance of a holistic treatment approach. Clin Cosmet
Investig Dermatol. 14:623–632. 2021.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Drucker AM, Wang AR, Li WQ, Sevetson E,
Block JK and Qureshi AA: The burden of atopic dermatitis: Summary
of a report for the National eczema association. J Invest Dermatol.
137:26–30. 2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Mann C, Dreher M, Weeß HG and Staubach P:
Sleep disturbance in patients with urticaria and atopic dermatitis:
An underestimated burden. Acta Derm Venereol.
100(adv00073)2020.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Carroll CL, Balkrishnan R, Feldman SR,
Fleischer AB Jr and Manuel JC: The burden of atopic dermatitis:
Impact on the patient, family, and society. Pediatr Dermatol.
22:192–199. 2005.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Rimoin LP, Kwatra SG and Yosipovitch G:
Female-specific pruritus from childhood to postmenopause: Clinical
features, hormonal factors, and treatment considerations. Dermatol
Ther. 26:157–167. 2013.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Kroll R, Archer DF, Lin Y, Sniukiene V and
Liu JH: A randomized, multicenter, double-blind study to evaluate
the safety and efficacy of estradiol vaginal cream 0.003% in
postmenopausal women with dyspareunia as the most bothersome
symptom. Menopause. 25:133–138. 2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Constantine GD, Simon JA, Pickar JH,
Archer DF, Kushner H, Bernick B, Gasper G, Graham S and Mirkin S:
REJOICE Study Group. The REJOICE trial: A phase 3 randomized,
controlled trial evaluating the safety and efficacy of a novel
vaginal estradiol soft-gel capsule for symptomatic vulvar and
vaginal atrophy. Menopause. 24:409–416. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Caraffa A, Spinas E, Kritas SK, Lessiani
G, Ronconi G, Saggini A, Antinolfi P, Pizzicannella J, Toniato E,
Theoharides TC and Conti P: Endocrinology of the skin: Intradermal
neuroimmune network, a new frontier. J Biol Regul Homeost Agents.
30:339–343. 2016.PubMed/NCBI
|
|
10
|
Farage MA, Neill S and MacLean AB:
Physiological changes associated with the menstrual cycle: A
review. Obstet Gynecol Surv. 64:58–72. 2009.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Lephart ED and Naftolin F: Menopause and
the Skin: Old favorites and new innovations in cosmeceuticals for
estrogen-deficient skin. Dermatol Ther (Heidelb). 11:53–69.
2021.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Amandusson A and Blomqvist A: Estrogen
receptor-alpha expression in nociceptive-responsive neurons in the
medullary dorsal horn of the female rat. Eur J Pain. 14:245–248.
2010.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Dun SL, Brailoiu GC, Gao X, Brailoiu E,
Arterburn JB, Prossnitz ER, Oprea TI and Dun NJ: Expression of
estrogen receptor GPR30 in the rat spinal cord and in autonomic and
sensory ganglia. J Neurosci Res. 87:1610–1619. 2009.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Chaban VV and Micevych PE: Estrogen
receptor-alpha mediates estradiol attenuation of ATP-induced
Ca2+ signaling in mouse dorsal root ganglion neurons. J
Neurosci Res. 81:31–37. 2005.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Sutaria N, Adawi W, Goldberg R, Roh YS,
Choi J and Kwatra SG: Itch: Pathogenesis and treatment. J Am Acad
Dermatol. 86:17–34. 2022.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Hashimoto T and Yosipovitch G: Itchy body:
Topographical difference of itch and scratching and C Nerve fibres.
Exp Dermatol. 28:1385–1389. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Andoh T, Asakawa Y and Kuraishi Y:
Non-myelinated C-fibers, but not myelinated A-fibers, elongate into
the epidermis in dry skin with itch. Neurosci Lett. 672:84–89.
2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Gérard C, Gallez A, Dubois C, Drion P,
Delahaut P, Quertemont E, Noël A and Pequeux C: Accurate control of
17β-Estradiol long-term release increases reliability and
reproducibility of preclinical animal studies. J Mammary Gland Biol
Neoplasia. 22:1–11. 2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Doty P, Picker MJ and Dykstra LA:
Differential cross-tolerance to opioid agonists in
morphine-tolerant squirrel monkeys responding under a schedule of
food presentation. Eur J Pharmacol. 174:171–180. 1989.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ström JO, Theodorsson A, Ingberg E,
Isaksson IM and Theodorsson E: Ovariectomy and 17β-estradiol
replacement in rats and mice: A visual demonstration. J Vis Exp.
(e4013)2012.PubMed/NCBI View
Article : Google Scholar
|
|
21
|
Lee JY, Kim JH, Hong SH, Lee JY, Cherny
RA, Bush AI, Palmiter RD and Koh JY: Estrogen decreases zinc
transporter 3 expression and synaptic vesicle zinc levels in mouse
brain. J Biol Chem. 279:8602–8607. 2004.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kendall LV, Bailey AL, Singh B and McGee
W: Toxic Effects of High-dose meloxicam and carprofen on female CD1
Mice. J Am Assoc Lab Anim Sci. 61:75–80. 2022.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Liu T, He Z, Tian X, Kamal GM, Li Z, Liu
Z, Liu H, Xu F, Wang J and Xiang H: Specific patterns of spinal
metabolites underlying α-Me-5-HT-evoked pruritus compared with
histamine and capsaicin assessed by proton nuclear magnetic
resonance spectroscopy. Biochim Biophys Acta Mol Basis Dis.
1863:1222–1230. 2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Miyamoto T, Nojima H, Shinkado T,
Nakahashi T and Kuraishi Y: Itch-associated response induced by
experimental dry skin in mice. Jpn J Pharmacol. 88:285–292.
2002.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Liu BW, Li ZX, He ZG, Wang Q, Liu C, Zhang
XW, Yang H and Xiang HB: Altered expression of itch-related
mediators in the lower cervical spinal cord in mouse models of two
types of chronic itch. Int J Mol Med. 44:835–846. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Yamanoi Y, Kittaka H and Tominaga M: Cheek
Injection Model for Simultaneous Measurement of Pain and
Itch-related Behaviors. J Vis Exp: Sep 27, 2019 (Epub ahead of
print). doi: 10.3791/58943.
|
|
27
|
Luo C, Zhao S, Peng C, Wang C, Hu K, Zhong
X, Luo T, Huang J and Lu D: Mammography radiomics features at
diagnosis and progression-free survival among patients with breast
cancer. Br J Cancer. 127:1886–1892. 2022.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Wu C, Lin F, Qiu S and Jiang Z: The
characterization of obese polycystic ovary syndrome rat model
suitable for exercise intervention. PLoS One.
9(e99155)2014.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Pereira MP, Mittal A and Ständer S:
Current treatment strategies in refractory chronic pruritus. Curr
Opin Pharmacol. 46:1–6. 2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Brasileiro LE, Barreto DP and Nunes EA:
Psychotropics in different causes of itch: Systematic review with
controlled studies. An Bras Dermatol. 91:791–798. 2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Chiricozzi A, Faleri S, Lanti A, Adorno G,
Lorè B, Chimenti S and Saraceno R: Apheresis in the treatment of
recalcitrant atopic dermatitis: Case series and review of the
literature. Eur J Dermatol. 24:545–550. 2014.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Takanami K, Uta D, Matsuda KI, Kawata M,
Carstens E, Sakamoto T and Sakamoto H: Estrogens influence female
itch sensitivity via the spinal gastrin-releasing peptide receptor
neurons. Proc Natl Acad Sci USA. 118(e2103536118)2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Liu Q, Tang Z, Surdenikova L, Kim S, Patel
KN, Kim A, Ru F, Guan Y, Weng HJ, Geng Y, et al: Sensory
neuron-specific GPCR Mrgprs are itch receptors mediating
chloroquine-induced pruritus. Cell. 139:1353–1365. 2009.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Xing Y, Chen J, Hilley H, Steele H, Yang J
and Han L: Molecular signature of pruriceptive MrgprA3+
neurons. J Invest Dermatol. 140:2041–2050. 2020.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Pohóczky K, Kun J, Szalontai B, Szőke É,
Sághy É, Payrits M, Kajtár B, Kovács K, Környei JL, Garai J, et al:
Estrogen-dependent up-regulation of TRPA1 and TRPV1 receptor
proteins in the rat endometrium. J Mol Endocrinol. 56:135–149.
2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Mac Bride MB, Rhodes DJ and Shuster LT:
Vulvovaginal atrophy. Mayo Clin Proc. 85:87–94. 2010.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Smith P, Heimer G, Lindskog M and Ulmsten
U: Oestradiol-releasing vaginal ring for treatment of
postmenopausal urogenital atrophy. Maturitas. 16:145–154.
1993.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Martínez-Lavín M: Dorsal root ganglia:
Fibromyalgia pain factory? Clin Rheumatol. 40:783–787.
2021.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Wang Q, Yang J, Wang H, Shan B, Yin C, Yu
H, Zhang X, Dong Z, Yu Y, Zhao R, et al: Fibroblast growth factor
13 stabilizes microtubules to promote Na+ channel
function in nociceptive DRG neurons and modulates inflammatory
pain. J Adv Res. 31:97–111. 2021.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Dong X and Dong X: Peripheral and Central
Mechanisms of Itch. Neuron. 98:482–494. 2018.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Fuentes N and Silveyra P: Estrogen
receptor signaling mechanisms. Adv Protein Chem Struct Biol.
116:135–170. 2019.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Eyster KM: The Estrogen receptors: An
overview from different perspectives. Methods Mol Biol. 1366:1–10.
2016.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Xu S, Cheng Y, Keast JR and Osborne PB:
17beta-estradiol activates estrogen receptor beta-signalling and
inhibits transient receptor potential vanilloid receptor 1
activation by capsaicin in adult rat nociceptor neurons.
Endocrinology. 149:5540–5548. 2008.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Sun LH, Zhang WX, Xu Q, Wu H, Jiao CC and
Chen XZ: Estrogen modulation of visceral pain. J Zhejiang Univ Sci
B. 20:628–636. 2019.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Cho T and Chaban VV: Interaction between
P2X3 and oestrogen receptor (ER)α/ERβ in ATP-mediated calcium
signalling in mice sensory neurones. J Neuroendocrinol. 24:789–797.
2012.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Oyola MG, Thompson MK, Handa AZ and Handa
RJ: Distribution and chemical composition of estrogen receptor β
neurons in the paraventricular nucleus of the female and male mouse
hypothalamus. J Comp Neurol. 525:3666–3682. 2017.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Contoreggi NH, Mazid S, Goldstein LB, Park
J, Ovalles AC, Waters EM, Glass MJ and Milner TA: Sex and age
influence gonadal steroid hormone receptor distributions relative
to estrogen receptor β-containing neurons in the mouse hypothalamic
paraventricular nucleus. J Comp Neurol. 529:2283–2310.
2021.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Sanoja R and Cervero F: Estrogen-dependent
abdominal hyperalgesia induced by ovariectomy in adult mice: A
model of functional abdominal pain. Pain. 118:243–253.
2005.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Multon S, Pardutz A, Mosen J, Hua MT,
Defays C, Honda S, Harada N, Bohotin C, Franzen R and Schoenen J:
Lack of estrogen increases pain in the trigeminal formalin model: A
behavioural and immunocytochemical study of transgenic ArKO mice.
Pain. 114:257–265. 2005.PubMed/NCBI View Article : Google Scholar
|