Introduction
Abnormal fat accumulation owing to an imbalance
between energy intake and consumption causes obesity (1). The rapid increase in the obese
population has emerged as a serious problem worldwide because
obesity causes serious metabolic diseases, such as diabetes,
arthritis, hyperlipidemia, and high blood pressure (1-3).
Thus, many drugs have been developed to treat obesity, including
orlistat (a lipase inhibitor) and sibutramine (an appetite
suppressant) (4). However, the
long-term clinical use of these anti-obesity drugs is limited
because they cause various side effects, including digestive
disorders, diarrhea, and insomnia (5). Therefore, the search for and
development of natural anti-obesity agents with few side effects is
urgently required (1).
As a medicinal plant belonging to the Solanaceae
family, Solanum nigrum has been used for the treatment of
tuberculosis, diuresis, neurological disorders, ulcers, liver
disorders, seizures, and inflammation (6). Numerous studies have shown that
Solanum nigrum relieves inflammation, protects the liver,
lowers blood pressure, and improves glucose tolerance (7-9).
In addition, Solanum nigrum reportedly reduced the
accumulation of hepatic and body fat in mice fed a high-fat diet
(10), indicating that it exerts
anti-obesity activity. However, since the mechanism of action
related to the anti-obesity activity of Solanum nigrum has
not been elucidated, in this study, we report that Solanum
nigrum inhibits adipogenesis and activates lipolysis, browning,
and autophagy in adipocytes and 3T3-L1 cells.
Materials and methods
Chemicals and antibodies. Dexamethasone
(D4902), 3-isobutyl-1-methylxanthine (IBMX) (I5879), insulin
(I6634), and Oil Red O (O0625) were purchased from Sigma-Aldrich
(St. Louis, MO, USA). The antibodies against PPARγ (#2430), CEBPα
(#8178), FABP4 (#2120), adiponectin (#2789), ATGL (#2138), HSL
(#4107), Perilipin-1 (#9349), p-AMPK (#2535), AMPK (#5831), UCP-1
(#14670), LC3 (#2775), β-actin (#5125), anti-rabbit IgG, HRP-linked
antibody (#7074) and anti-mouse IgG, HRP-linked antibody (#7076)
were purchased from Cell Signaling (Bervely, MA, USA). Antibodies
against PGC-1α (sc-518025) and PRDM16 (ab106410) were purchased
from Santa Cruz Biotechnology (Dallas, TX, USA) and Abcam
(Cambridge, United Kingdom), respectively.
Sample preparation. Aerial parts and fruit of
Solanum nigrum were purchased from a local market in
Dongdaemun-gu, Seoul. After grinding, 10 g of the powder was
immersed in 200 ml of distilled water and left at 80˚C
for 6 h for extraction, after which the extracts were centrifuged
at 15,000 rpm at 4˚C for 10 min, and the clear
supernatant was freeze-dried. The extraction yields of SNAP and SNF
were 21.35 and 19.26%, respectively. The freeze-dried extracts from
aerial parts (SNAP) or fruits (SNF) of Solanum nigrum were
dissolved in distilled water for treatment of the cells and stored
at -80˚C.
Culture and differentiation of 3T3-L1 cells.
Mouse adipocytes, 3T3-L1 cells, were used in this study because
they are mainly used to verify in vitro anti-obesity
activity (11). The 3T3-L1 cells
(American Type Culture Collection, Manassas, VA, USA, Cat. NO.:
CL-173) were maintained in a DMEM/F-12 medium containing 10% bovine
calf serum, 100 U/ml penicillin, and 100 µg/ml streptomycin at
37˚C under a humidified atmosphere of 5% CO2.
All experiments performed in this study used 3T3-L1 cells at
passage numbers 10 or lower. For the differentiation of 3T3-L1
cells, 2 days (D0) after reaching 100% confluence, the cells were
cultured in a DMI medium (DMEM/F-12 medium, 10% fetal bovine serum,
1 µM dexamethasone, 0.5 mM IBMX, and 10 µg/ml insulin) for 2 days
(D2). Then, the cells were cultured in an insulin medium (DMEM/F-12
medium, 10% fetal bovine serum, and 10 µg/ml insulin) for 2 days
(D4). The culture medium (DMEM/F-12 containing 10% fetal bovine
serum) was changed every two days (D6-D8).
Measurement of cell viability. The 3T3-L1
cells were further cultured for 2 days in a 6-well plate at full
confluence at 37˚C with 5% CO2. After 2 days,
the 3T3-L1 cells were treated with SNAP without DMI and insulin
(undifferentiated) or with DMI and insulin (differentiated) and
then cultured from D2 to D8. On D8 after SNAP treatment, cell
viability was measured using a NucleoCounter NC-250 (Chemometec,
Allerod, Denmark) according to the manufacturer's protocol.
Oil Red O staining
The 3T3-L1 cells were fixed in 10% formalin for 1 h
at room temperature. They were then dehydrated with 60% isopropanol
for 5 min and stained with Oil Red O solution (isopropanol:
water=6:4, v/v) for 20 min. After washing the 3T3-L1 cells with
distilled water, the stained lipid droplets of 3T3-L1 cells were
photographed using a light microscope (Olympus, Tokyo, Japan) and
eluted with 100% isopropanol for quantitative analysis. The
absorbance of the eluted solution was measured at 500 nm using a
microplate reader (Human Co., Xma-3000PC, Seoul, Korea).
Measurement of glycerol content. The 3T3-L1
cells were differentiated for 6 days without SNAP treatment, after
which they were treated with SNAP for 48 h, and the free glycerol
content was measured using a cell-based glycerol assay kit (Cayman
Chemical, Ann Arbor, MI, USA) according to the manufacturer's
protocol. The cell culture medium was mixed with the reconstituted
free glycerol assay reagent in a 1:4 ratio and incubated at room
temperature for 15 min. After the reaction, absorbance was measured
at 540 nm using a microplate reader (Human Cop., Xma-3000PC, Seoul,
Korea).
Western blot analysis. Protein changes in
SNAP-treated 3T3-L1 cells were analyzed by Western blotting. The
cells were collected using RIPA buffer and incubated at 4˚C for 30
min. After centrifugation at 15,000 rpm for 30 min the protein
extract was obtained. The total protein extracted from the 3T3-L1
cells was quantified using the BCA protein assay kit (Thermo Fisher
Scientific, Waltham, MA, USA). For SDS-PAGE, an equal amount of
protein (30 g/well) was subjected to electrophoresis on 12 or 8%
acrylamide gels at 150 V and 400 A for 1 h. Proteins separated on
the acrylamide gel were transferred onto a nitrocellulose membrane
(Thermo Fisher Scientific) for 2 h at 100 and 300 A. After blocking
at room temperature for 1 h, the membranes were incubated with the
primary antibodies (1:1,000) at 4˚C overnight. The
membranes were then incubated with secondary antibodies (1:1,000)
at room temperature for 1 h. After treating the membrane with ECL
Western blotting substrate, the protein bands were visualized using
an LI-COR C-DiGit Blot Scanner (LI-COR, NE, USA). Quantitative
analysis of the visualized protein bands was performed using
UN-SCAN-IT gel software version 5.1 (Silk Scientific Inc. Orem, UT,
USA).
Statistical analysis. All experiments were
repeated at least three times. Statistical analyses were verified
using GraphPad Prism version 5.0 (GraphPad Software, Inc.) and data
are presented as mean ± standard deviation. Results were considered
statistically significant at P-<0.05. Each data point was
analyzed using a one-way analysis of variance (ANOVA), and the data
were analyzed using the Bonferroni post hoc test. During ANOVA
analysis, a normality test (Shapiro-Wilk Test) was performed.
Results
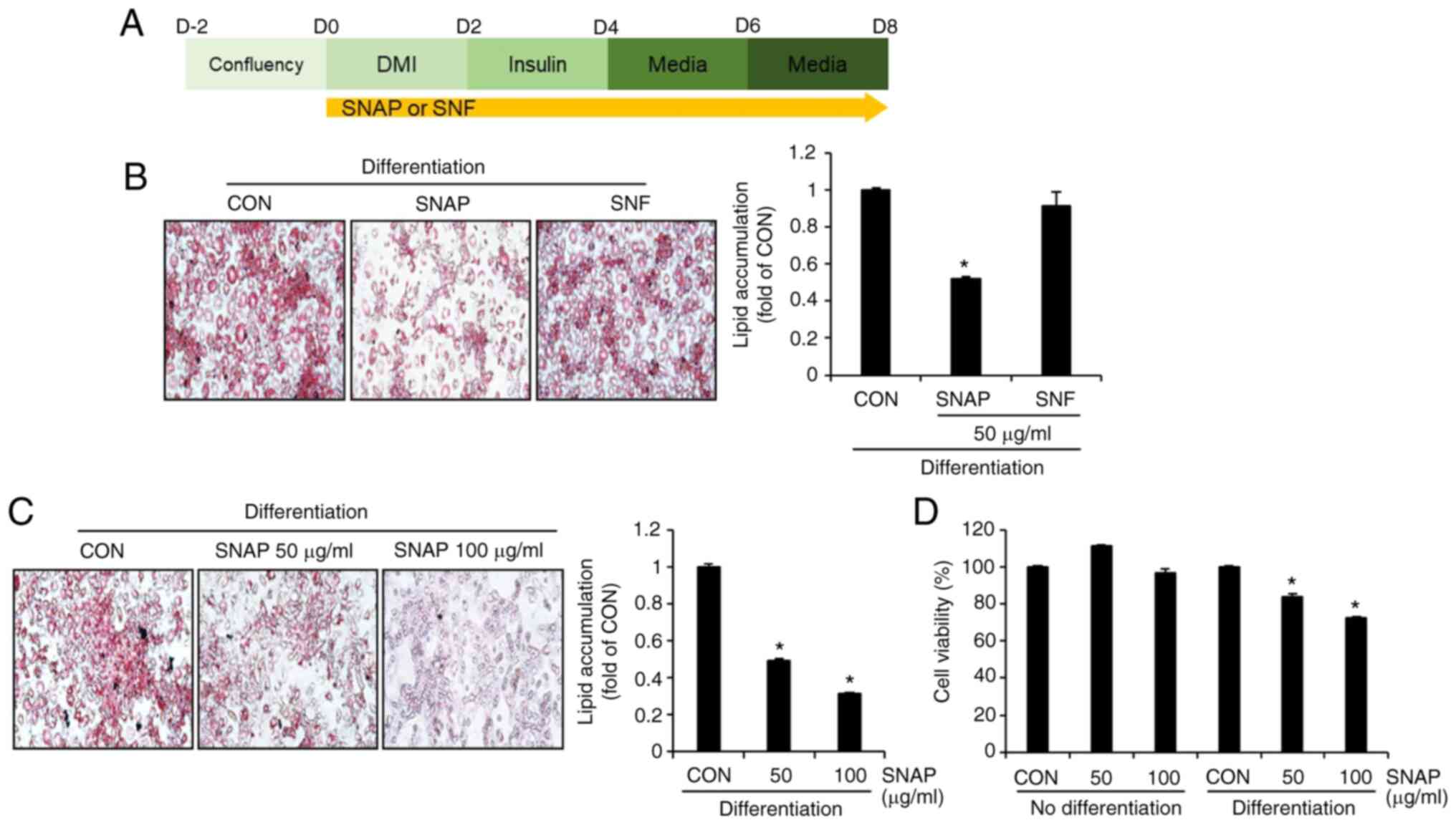
The inhibitory effect of SNAP on lipid droplet
accumulation in 3T3-L1 cells is higher than that of SNF.
Because obesity is closely related to the accumulation of lipid
droplets within adipocytes (12),
we investigated whether SNAP and SNF inhibit the accumulation of
lipid droplets in 3T3-L1 cells. In previous studies (10), treatment of 3T3-L1 cells with
polyphenol extract from aerial parts of Solanum nigrum
resulted in a rapid decrease in cell viability at concentrations
higher than 100 µg/ml. Therefore, in this study, a concentration of
100 µg/ml was designated as a high concentration, and a
concentration of 50 µg/ml was designated as a low concentration.
SNAP and SNF were administered from the beginning (D0) to the end
(D8) of 3T3-L1 cell differentiation, and their inhibitory activity
on lipid droplet accumulation was measured using Oil Red O staining
(Fig. 1A). As shown in Fig. 1B, SNAP effectively reduced lipid
droplet accumulation compared to that in the untreated control
group (CON), but SNF did not show such an effect. Because this
result demonstrates that only SNAP has anti-obesity activity, we
used SNAP in subsequent studies. Furthermore, SNAP inhibited lipid
droplet accumulation in a concentration-dependent manner (Fig. 1C). Evaluation of the effect of SNAP
on cell viability in undifferentiated or differentiated 3T3-L1
cells showed that SNAP had no effect on cell viability in
undifferentiated 3T3-L1 cells, but slightly decreased cell
viability in differentiated 3T3-L1 cells (Fig. 1D).
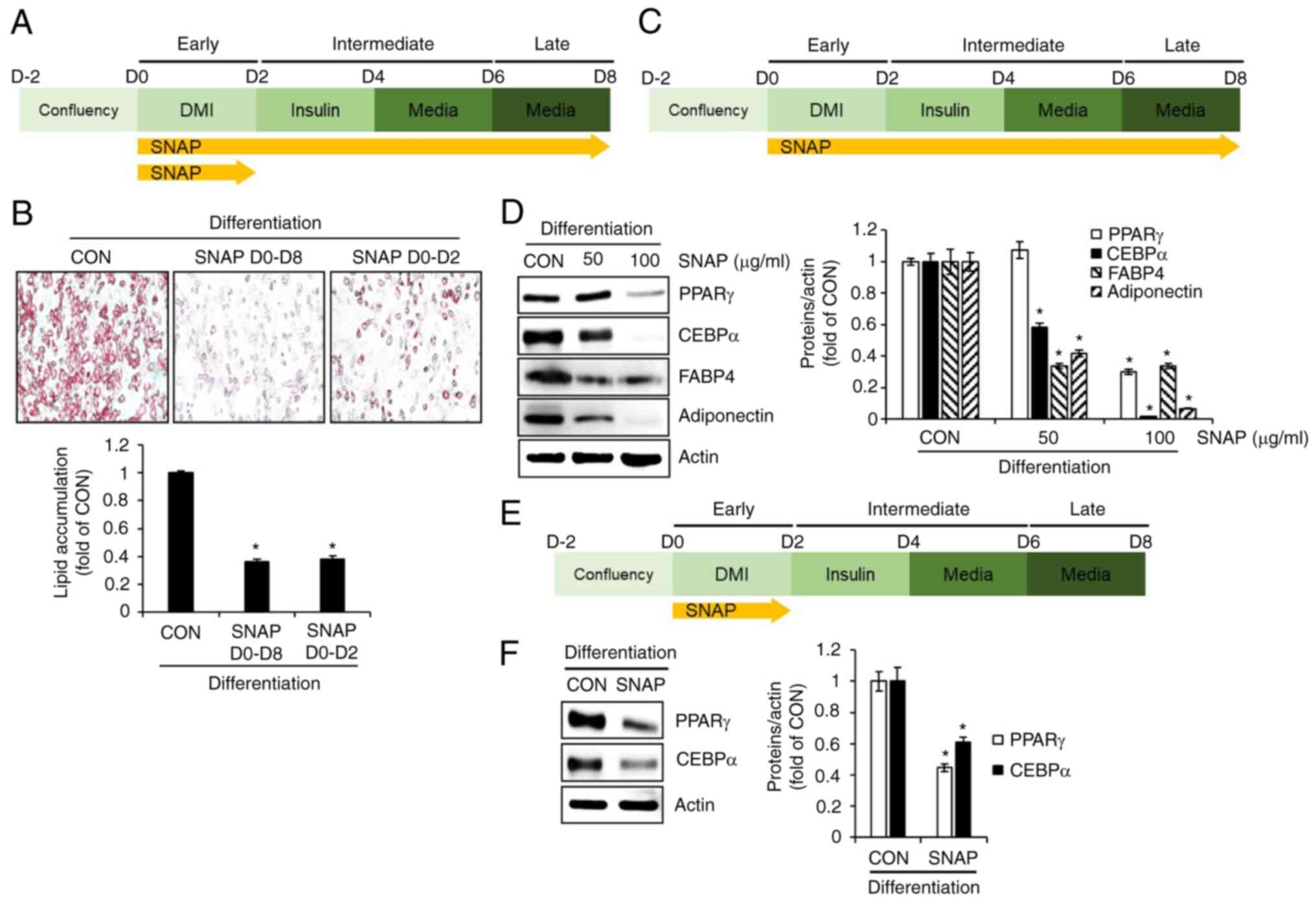
SNAP inhibits adipogenesis in 3T3-L1 cells.
To verify whether the inhibition of adipogenesis by SNAP was
involved in the reduction in lipid droplet accumulation, we treated
cells with SNAP from D0 to D8 (during all phases of the
differentiation process) or from D0 to D2 (at the early phase of
the differentiation process) and investigated the extent of lipid
droplet accumulation using Oil Red O staining (Fig. 2A). As shown in Fig. 2B, SNAP treatment from D0 to D8 or
from D0 to D2 resulted in a significant reduction in lipid droplet
accumulation compared to the untreated control group (CON). Thus,
we analyzed the expression changes of adipogenesis-related proteins
such as PPARγ, CEBPα, FABP4, and adiponectin by SNAP in 3T3-L1
cells treated with SNAP from D0 to D8 using Western blot analysis
(Fig. 2C). As shown in Fig. 2D, SNAP dose-dependently
downregulated the expression levels of PPARγ, CEBPα, FABP4, and
adiponectin. We also investigated the effects of SNAP on the
expression of adipogenesis-related proteins during the early stages
of differentiation. In 3T3-L1 cells treated with SNAP from D0 to D2
(Fig. 2E), SNAP significantly
decreased protein expression levels of PPARγ and CEBPα (Fig. 2F).
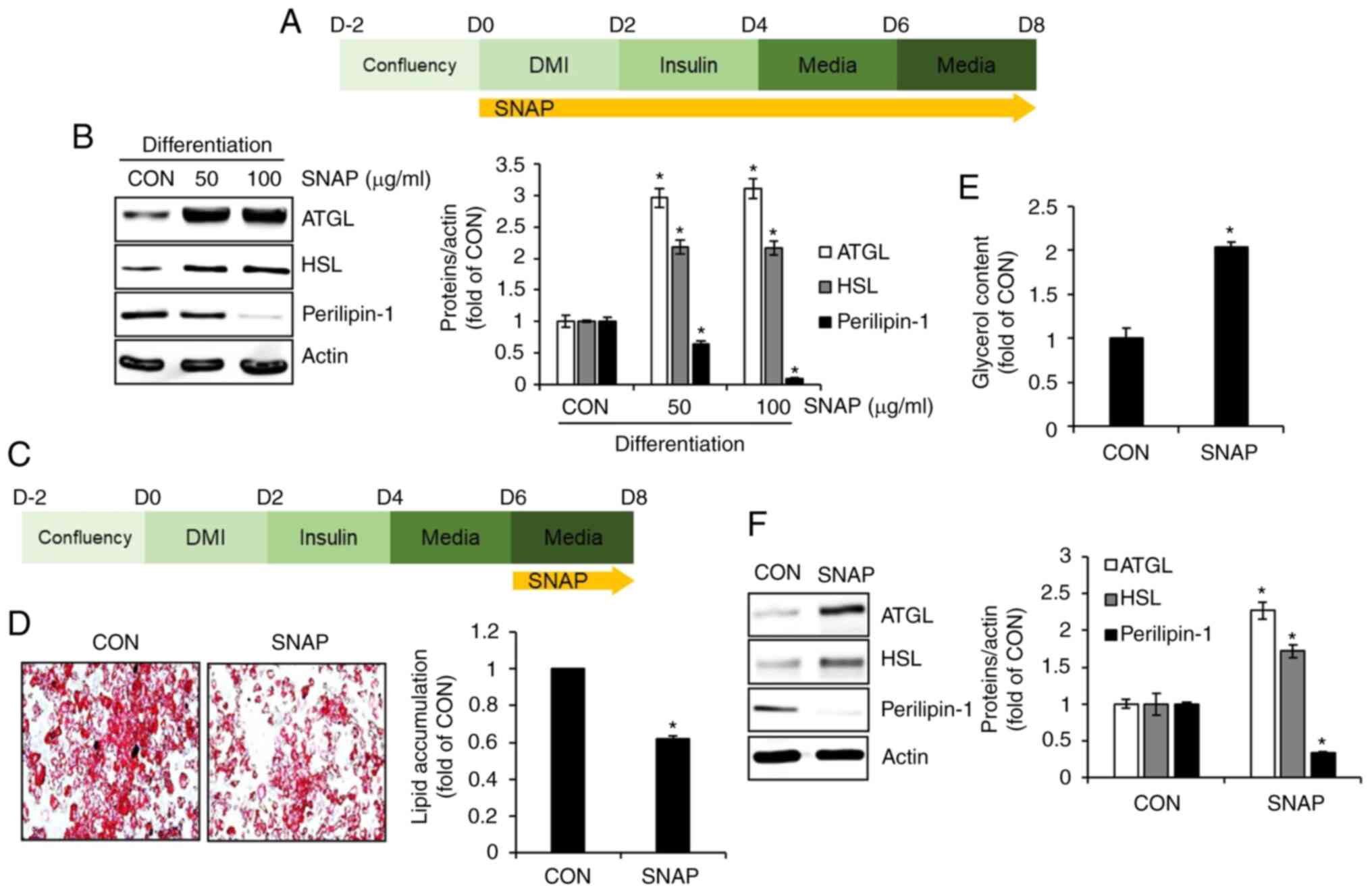
SNAP induces lipolysis in 3T3-L1 cells. To
investigate whether lipolysis is involved in SNAP-mediated
inhibition of lipid droplet accumulation, 3T3-L1 cells undergoing
differentiation were treated with SNAP from D0 to D8, and changes
in the protein expression of ATGL, HSL, and perilipin-1, which are
involved in lipolysis, were measured by Western blot analysis
(Fig. 3A). As shown in Fig. 3B, the protein expression of ATGL
and HSL increased, whereas that of perilipin-1 decreased in
SNAP-treated 3T3-L1 cells compared to untreated cells (CON). To
further demonstrate whether SNAP induces lipolysis, 3T3-L1 cells
were allowed to accumulate lipid droplets from D0 to D6, followed
by treatment with SNAP from D6 to D8 (Fig. 3C). Changes in lipid droplet
accumulation, free glycerol content, and protein expression of
ATGL, HSL, and perilipin-1 were investigated. SNAP treatment from
D6 to D8 led to a decrease in lipid droplet accumulation (Fig. 3D) and an increase in free glycerol
content (Fig. 3E) in 3T3-L1 cells
compared to the untreated cells (CON). In addition, the protein
expression of ATGL and HSL was upregulated, whereas that of
perilipin-1 was downregulated in SNAP-treated 3T3-L1 cells compared
to untreated cells (CON).
SNAP induces thermogenesis and autophagy in
3T3-L1 cells. To verify whether thermogenesis and autophagy
contribute to the SNAP-mediated inhibition of lipid droplet
accumulation, 3T3-L1 cells undergoing differentiation were treated
with SNAP from D0 to D8 (Fig. 4A),
and the changes in the expression of the thermogenesis-related
proteins (p-AMPK, AMPK, UCP-1, PGC-1α, and PRDM16) and the
autophagy-related protein (LC3-II) were measured by Western blot
analysis. As shown in Fig. 4B,
treatment with SNAP did not affect the protein expression of AMPK
but effectively phosphorylated AMPK and significantly increased the
protein expression of UCP-1, PGC-1α, and PRDM16 in 3T3-L1 cells.
Furthermore, the protein expression of LC3-II was significantly
increased in 3T3-L1 cells treated with SNAP (Fig. 4C).
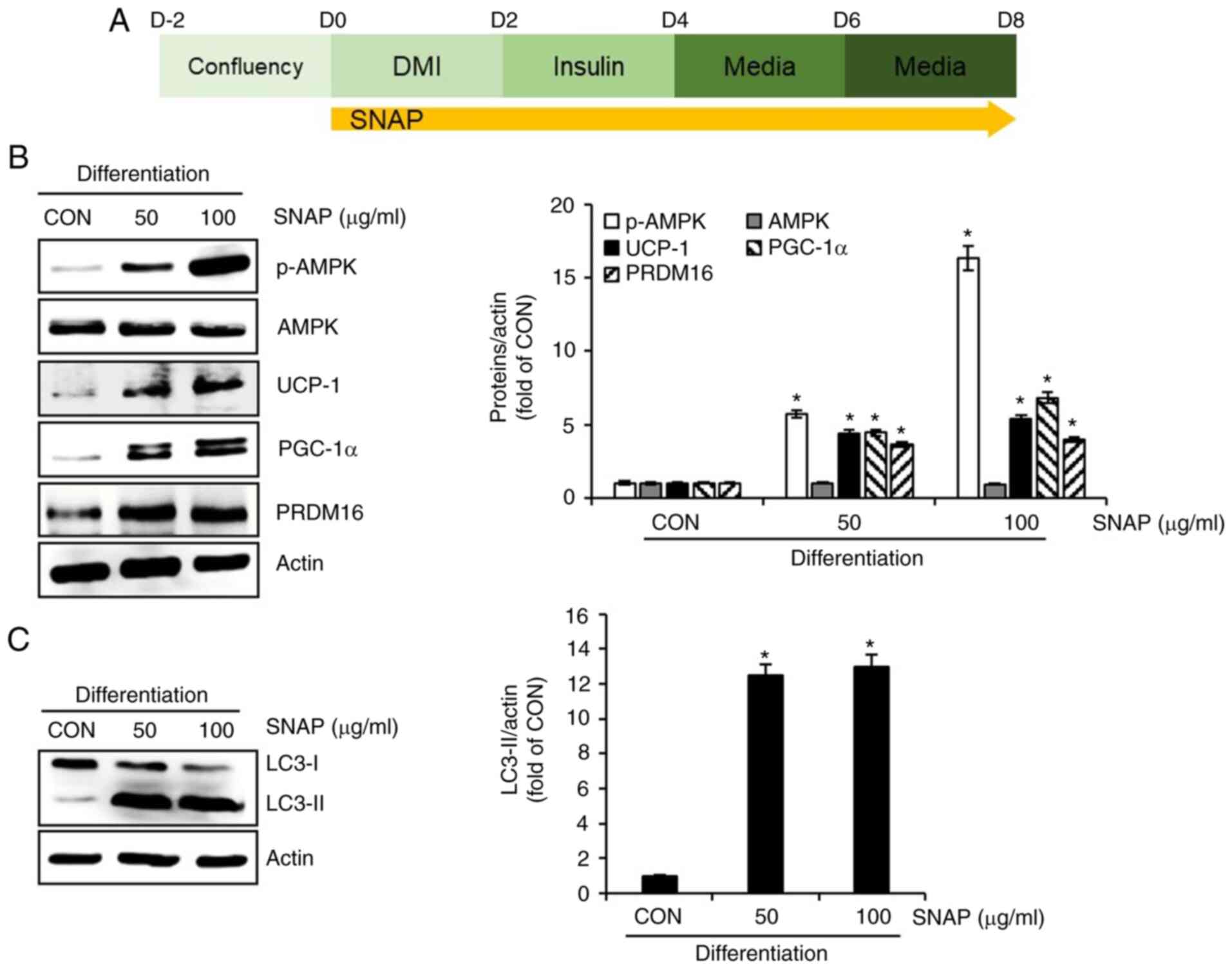 | Figure 4Effect of SNAP on thermogenesis and
autophagy in 3T3-L1 cells. (A) Experimental design. (B) Western
blot analysis in 3T3-L1 cells treated with SNAP from D0 to D8. (C)
Western blot analysis of LC3-I and -II in 3T3-L1 cells treated with
SNAP from D0 to D8. *P<0.05 vs. CON group. CON,
control group without the sample treatment; DMI, mixture of 0.05 mM
IBMX, 1 µM dexamethasone and 10 µg/ml insulin; SNAP, Solanum
nigrum aerial part extracts; p, phosphorylated; AMPK,
AMP-activated protein kinase; PGC-1α, peroxisome
proliferator-activated receptor-γ coactivator 1α; PRDM16, PR
domain-containing 16; UCP-1, uncoupling protein 1; LC3,
microtubule-associated protein 1A/1B-light chain 3. |
Discussion
The pursuit of a comfortable lifestyle and unhealthy
eating habits leads to obesity, a global health problem (13). Obesity occurs when there is
excessive accumulation of fat in adipose tissue (13). White adipose tissue (WAT) is known
to increase because of excessive lipid accumulation within
adipocytes, which is a major characteristic of obesity (14). Adipogenesis, the process by which
preadipocytes differentiate into mature adipocytes, increases the
number of mature adipocytes in WAT (15-17).
Therefore, agents that inhibit adipogenesis can be developed as
anti-obesity agents (14). PPARγ,
CEBPα, FABP4, and adiponectin are major factors that promote
adipogenesis in adipocytes (18-22).
In this study, we demonstrated that SNAP downregulates the protein
expressions of PPARγ, CEBPα, FABP4, and adiponectin in
differentiating 3T3-L1 cells. Downregulation of PPARγ and CEBPα
expression inhibits weight gain by reducing adipogenesis in an
obese mouse model (18,19). FABP4 is overexpressed during
adipogenesis and autonomously accounts for 6% of total cytoplasmic
proteins in mature adipocytes (20). FABP4 overexpression promotes the
progression of obesity, obesity-induced type 2 diabetes,
hypertension, and cardiovascular diseases (21). Adiponectin promotes adipogenesis
and lipid accumulation in adipocytes (22). While it is a limitation of this
study that we did not analyze the expression of other proteins
involved in adipogenesis beyond PPARγ, CEBPα, FABP4, and
adiponectin, considering the previous reports (18-22)
and our results, we believe that SNAP may reduce lipid droplet
accumulation by inhibiting adipogenesis.
Lipolysis is the process by which triacylglycerol
(TAG) in the adipose tissue sequentially decomposes into
diacylglycerol (DAG), monoacylglycerol (MAG), fatty acids (FA), and
glycerol (23,24). As a decrease in lipolysis is
frequently observed in obesity, reducing stored fat by stimulating
hydrolysis through lipolysis is considered a major target for
obesity treatment (25). In the
present study, we present two pieces of evidence that SNAP induces
lipolysis in 3T3-L1 cells. The first piece of evidence suggests
that SNAP increases the protein expression of ATGL and HSL and
decreases the protein expression of perilipin-1 in 3T3-L1 cells.
ATGL and HSL are involved in lipid breakdown during lipolysis.
Specifically, ATGL breaks down TAG into DAG and FA, whereas HSL
breaks down TAG into DAG and further into MAG (24). Perilipin-1 surrounding lipid
droplets in adipocytes inhibits lipolysis by limiting the access of
ATGL and HSL to lipid droplets (23). These reports provided the first
evidence that SNAP induce lipolysis. Secondly, the amount of free
glycerol increased in SNAP-treated 3T3-L1 cells. Because TAG are
ultimately hydrolyzed via lipolysis into three molecules of free FA
and one molecule of free glycerol (23,24),
the increase in free glycerol in SNAP-treated 3T3-L1 cells
indicated that lipolysis was induced. These two pieces of evidence
demonstrated that SNAP inhibited the accumulation of lipid droplets
by inducing lipolysis in adipocytes.
Brown adipose tissue (BAT) generates heat by
dissipating excess energy stored in the form of lipids in WAT, a
phenomenon referred to as thermogenesis (26). Therefore, as the dissipation of
excess energy in the form of heat is effective for weight loss, an
increase in BAT is highly effective in combating obesity (27). In this study, we propose that SNAP
induces thermogenesis by presenting evidence that SNAP increases
the phosphorylation of AMPK and protein expression of UCP-1,
PGC-1α, and PRDM16. AMPK and PRDM16 activate BAT through WAT
browning (26,28), and UCP-1 and PGC-1α predominantly
expressed in BAT are involved in the dissipation of excess energy
as heat (29,30). These previous reports support the
evidence for SNAP-mediated thermogenesis presented in this study.
Therefore, we propose that thermogenesis may be involved in
SNAP-mediated inhibition of lipid droplet accumulation in
adipocytes.
Recently, continuous activation of autophagy was
reported to inhibit adipocyte maturation via a lipolytic mechanism
(31). In adipocytes, autophagy
degrades lipid droplets in the cytoplasm and reduces
triacylglycerol stored in the lipid droplets (32). In the present study, we observed a
significant increase in LC3-II protein expression in SNAP-treated
3T3-L1 cells. Because LC3-II is closely associated with the
formation of autophagosomes and autolysosomes, it is commonly used
as a key indicator of autophagy induction (33). Furthermore, it has been reported
that the increase in ATGL and decrease in perilipin-1 promote
autophagic lipolysis (34,35). Based on these previous reports, it
has been suggested that SNAP can induce autophagy in adipocytes and
that SNAP-induced autophagic lipolysis may contribute to the
inhibition of lipid droplet accumulation.
Because this study was conducted in vitro to
elucidate the anti-obesity mechanisms of SNAP, the lack of in
vivo studies is a limitation. However, it has recently been
reported that Solanum nigrum exhibit effective anti-obesity
activity in an animal model of HFD-induced obesity (10). Moreover, they reported that
Solanum nigrum induces lipolysis through the upregulation of
PPARα and CPT-1 and inhibits lipogenesis through the downregulation
of FAS and HMG-CoR (10). They
also confirmed an increase in AMPK phosphorylation in the livers of
mice treated with Solanum nigrum (10). Although there were differences in
the anti-obesity-related proteins analyzed between previous in
vivo studies and our in vitro study, a previous in
vivo study suggested that SNAP may also exhibit effective
anti-obesity activity in an in vivo model. Therefore, it is
necessary to confirm whether SNAP exhibits anti-obesity activity in
an in vivo model and whether the changes in
anti-obesity-related proteins analyzed in this study are also
observed in vivo. Furthermore, while previous studies showed
that polyphenol extracts from Solanum nigrum (SNPE)
inhibited lipid accumulation at concentrations of 300-500 µg/ml,
this study demonstrated noteworthy inhibition of lipid accumulation
at concentrations as low as 50-100 µg/ml of SNAP. Such differences
could be attributed to the differences in the extraction solvents,
as SNPE is an ethanol extract and SNAP is a water extract.
Additionally, differences in growing environments could also be a
factor, as SNPE was derived from Solanum nigrum grown in
Taiwan, whereas SNAP was derived from Solanum nigrum grown
in Korea.
Although an accurate analysis of the
anti-obesity-related components of SNAP is crucial, this study has
a limitation in that we could not analyze the specific components
of SNAP that inhibit adipogenesis and induce lipolysis,
thermogenesis, and autophagy. However, when Solanum nigrum
was extracted with distilled water, various components such as
miquelianin, catechin, rutin, gallic acid, isoquercetin,
peltatoside, nuciferine, quercetin, and epigallocatechin gallate
were detected (10). Among the
various compounds detected in water extracts of Solanum
nigrum, catechin (36), rutin
(37), nuciferine (38), quercetin (39), and epigallocatechin gallate
(40) have been reported to
exhibit anti-obesity activity. Although the precise ingredients
involved in the anti-obesity activity of SNAP have not been
identified, based on preliminary reports (36-40),
it can be inferred that catechin, rutin, nuciferine, quercetin, and
epigallocatechin gallate are involved in the anti-obesity activity
of SNAP. However, it is necessary to precisely analyze the
anti-obesity ingredients in SNAP using activity-tracing-based
ingredient analysis.
In this study, we observed that SNAP exhibited
higher inhibitory activity against lipid accumulation in 3T3-L1
cells than SNF at the same concentration. Although this study did
not conduct a quantitative analysis of the specific components
responsible for inhibiting lipid accumulation in SNAP and SNF, the
results suggest that SNAP may contain more components that inhibit
lipid accumulation than SNF. Therefore, it is necessary to compare
the components that inhibit lipid accumulation in SNAP and SNF in
future studies to overcome the limitations of this study.
In this study, we demonstrated that SNAP suppresses
the excessive accumulation of lipid droplets by inhibiting
adipogenesis and inducing lipolysis, thermogenesis, and autophagy
in 3T3-L1 cells. Although the anti-obesity activity of SNAP has
already been verified, the novelty of this study lies in its newly
identified mechanism of action, which provides new evidence for its
anti-obesity activity.
Acknowledgements
Not applicable.
Funding
Funding: This work was supported by the Basic Science Research
Program of the National Research Foundation of Korea (NRF) and
funded by the Ministry of Education (grant nos.
NRF-2018R1A6A1A03024862 and NRF-2022R1I1A3055428).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JWC, HJC, GHR and JWL performed the experiments and
analyzed the data. JWC and HJC drafted the manuscript. GHR and JWL
edited the manuscript. JBJ designed the experiments and wrote and
edited the manuscript. JWC, HJC, GHR, JWL and JBJ confirm the
authenticity of all the raw data. All the authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Park YH, An M, Kim JK and Lim YH:
Antiobesity effect of ethanolic extract of Ramulus mori in
differentiated 3T3-L1 adipocytes and high-fat diet-induced obese
mice. J Ethnopharmacol. 251(112542)2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Haslam DW and James WPT: Obesity. Lancet.
366:1197–1209. 2005.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Visscher TL and Seidell JC: The public
health impact of obesity. Ann Rev Public Health. 22:55–375.
2001.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Hofbauer KG, Nicholson JR and Boss O: The
obesity epidemic: Current and future pharmacological treatments.
Ann Rev Pharmacol Toxicol. 47:565–592. 2007.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Cheung BMY, Cheung TT and Samaranayake NR:
Safety of antiobesity drugs. Ther Adv Drug Saf. 4:171–181.
2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Javed T, Ashfaq UA, Riza S, Rehman S and
Riazuddin S: In-vitro antiviral activity of Solanum nigrum
against Hepatitis C Virus. Virol J. 8(26)2011.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zakaria ZA, Gopalan HK, Zainal H, Pojan
NHM, Morsid NA, Aris A and Sulaiman MR: Antinociceptive,
anti-inflammatory and antipyretic effects of Solanum nigrum
chloroform extract in animal models. Yakugaku Zasshi.
126:1171–1178. 2006.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Lin HM, Tseng HC, Wang CJ, Chyau CC, Liao
KK, Peng PL and Chou FP: Induction of autophagy and apoptosis by
the extract of Solanum nigrum Linn in HepG2 cells. J Agric
Food Chem. 55:3620–3628. 2007.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Hsieh CC, Fang HL and Lina WC: Inhibitory
effect of Solanum nigrum on thioacetamide-induced liver
fibrosis in mice. J Ethnopharmacol. 119:117–121. 2008.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Peng CH, Cheng JJ, Yu MH, Chung DJ, Huang
CN and Wang CJ: Solanum nigrum polyphenols reduce body
weight and body fat by affecting adipocyte and lipid metabolism.
Food Funct. 11:483–492. 2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Lee ES, Choi SJ, Kim HD, Kang MH, Ji YJ,
Kim GS and Jang GY: Anti-obesity activity in 3T3-L1 cells of
Cornus officinalis fruits harvested at different times.
Processes. 10(2008)2020.
|
|
12
|
Getiye Y, Rice TA, Phillips BD, Carrillo
DF and He G: Dysregulated lipolysis and lipophagy in lipid droplets
of macrophages from high fat diet-fed obese mice. J Cell Mol Med.
26:4825–4836. 2022.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Krahmer N Jr, Farese RV and Walther TC:
Balancing the fat: Lipid droplets and human disease. EMBO Mol Med.
5:905–915. 2013.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Yang L, Jia X, Fang D, Cheng Y, Zhai Z,
Deng W, Du B, Lu T, Wang L, Yang C and Gao Y: Metformin inhibits
lipid droplets fusion and growth via reduction in cidec and its
regulatory factors in rat adipose-derived stem cells. Int J Mol
Sci. 23(5986)2022.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Lefterova MI and Lazar MA: New
developments in adipogenesis. Trends Endocrinol Metab. 20:107–114.
2009.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Rosen ED and MacDougald OA: Adipocyte
differentiation from the inside out. Nat Rev Mol Cell Biol.
7:885–896. 2006.PubMed/NCBI View
Article : Google Scholar
|
|
17
|
Tang QQ and Lane MD: Adipogenesis: From
stem cell to adipocyte. Ann Rev Biochem. 81:715–736.
2012.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Guo L, Kang JS, Kang NJ, Je BI, Lee YJ,
Park YH and Choi YW: Pelargonidin suppresses adipogenesis in 3T3-L1
cells through inhibition of PPAR-γ signaling pathway. Arch Biochem
Biophys. 686(108365)2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Millward CA, Heaney JD, Sinasac DS, Chu
EC, Bederman IR, Gilge DA, Previs SF and Croniger CM: Mice with a
deletion in the gene for CCAAT/enhancer-binding protein beta are
protected against diet-induced obesity. Diabetes. 56:161–167.
2007.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Spiegelman BM and Green H: Control of
specific protein biosynthesis during the adipose conversion of 3T3
cells. J Biol Chem. 255:8811–8818. 1980.PubMed/NCBI
|
|
21
|
Nakamura R, Okura T, Fujioka Y, Sumi K,
Matsuzawa K, Izawa S, Ueta E, Kato M, Taniguchi SI and Yamamoto K:
Serum fatty acid-binding protein 4 (FABP4) concentration is
associated with insulin resistance in peripheral tissues, A
clinical study. PLoS One. 12(e0179737)2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Fu Y, Luo N, Klein RL and Garvey WT:
Adiponectin promotes adipocyte differentiation, insulin
sensitivity, and lipid accumulation. J Lipid Res. 46:1369–1379.
2005.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Duncan RE, Ahmadian M, Jaworski K,
Sarkadi-Nagy E and Sul HS: Regulation of lipolysis in adipocytes.
Ann Rev Nutr. 27:79–101. 2007.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Lass A, Zimmermann RZ, Oberer M and
Zechner R: Lipolysis-A highly regulated multi-enzyme complex
mediates the catabolism of cellular fat stores. Prog Lipid Res.
50:14–27. 2011.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Langin D: Adipose tissue lipolysis as a
metabolic pathway to define pharmacological strategies against
obesity and the metabolic syndrome. Pharmacol Res. 53:482–491.
2006.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Lo KA and Sun L: Turning WAT into BAT: A
review on regulators controlling the browning of white adipocytes.
Biosci Rep. 33(e00065)2013.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Kuryłowicz A and Puzianowska-Kuźnicka M:
Induction of adipose tissue browning as a strategy to combat
obesity. Int J Mol Sci. 21(6241)2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
van der Vaart JI, Boon MR and Houtkooper
RH: The role of AMPK signaling in brown adipose tissue activation.
Cells. 10(1122)2021.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Cannon B and Nedergaard J: Brown adipose
tissue: Function and physiological significance. Physiol Rev.
84:277–359. 2004.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Seale P, Kajimura S, Yang W, Chin S, Rohas
LM, Uldry M, Tavernier G, Langin D and Spiegelman BM:
Transcriptional control of brown fat determination by PRDM16. Cell
Metab. 6:38–54. 2007.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Zhang X, Wu D, Wang C, Luo Y, Ding X, Yang
X, Silva F, Arenas S, Weaver JM, Mandell M, et al: Sustained
activation of autophagy suppresses adipocyte maturation via a
lipolysis-dependent mechanism. Autophagy. 16:1668–1682.
2020.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Singh R, Xiang Y, Wang Y, Baikati K,
Cuervo AM, Luu YK, Tang Y, Pessin JE, Schwartz GJ and Czaja MJ:
Autophagy regulates adipose mass and differentiation in mice. J
Clin Invest. 119:3329–3339. 2009.PubMed/NCBI View
Article : Google Scholar
|
|
33
|
Runwal G, Stamatakou E, Siddiqi FH, Puri
C, Zhu Y and Rubinsztein DC: LC3-positive structures are prominent
in autophagy-deficient cells. Sic Rep. 9(10147)2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Sathyanarayan A, Mashek MT and Mashek DG:
ATGL promotes autophagy/lipophagy via SIRT1 to control hepatic
lipid droplet catabolism. Cell Rep. 49:1–9. 2017.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Sztalryd C and Brasaemile DL: The
perilipin family of lipid droplet proteins: Gatekeepers of
intracellular lipolysis. Biochim Biophys Acta. 1862:1221–1232.
2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Jiang Y, Ding S, Li F, Zhang C,
Sun-Waterhouse D, Chen Y and Li D: Effects of (+)-catechin on the
differentiation and lipid metabolism of 3T3-L1 adipocytes. J Funct
Foods. 62(103558)2019.
|
|
37
|
Choi I, Park Y, Choi H and Lee EH:
Anti-adipogenic activity of rutin in 3T3-L1 cells and mice fed with
high-fat diet. Biofactors. 26:273–281. 2006.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Xu H, Wang L, Yan K, Zhu H, Pan H, Yang H,
Liu M and Gong F: Nuciferine iinhibited the differentiation and
lipid accumulation of 3T3-L1 preadipocytes by regulating the
expression of lipogenic genes and adipokines. Front Pharmacol.
12(632236)2021.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Hong SY, Ha AW and Kim W: Effects of
quercetin on cell differentiation and adipogenesis in 3T3-L1
adipocytes. Nutr Res Pract. 15:444–455. 2021.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Kim H, Hiraishi A, Tsuchiya K and Sakamoto
K: (-) Epigallocatechin gallate suppresses the differentiation of
3T3-L1 preadipocytes through transcription factors FoxO1 and
SREBP1c. Cytotechnology. 62:245–255. 2010.PubMed/NCBI View Article : Google Scholar
|