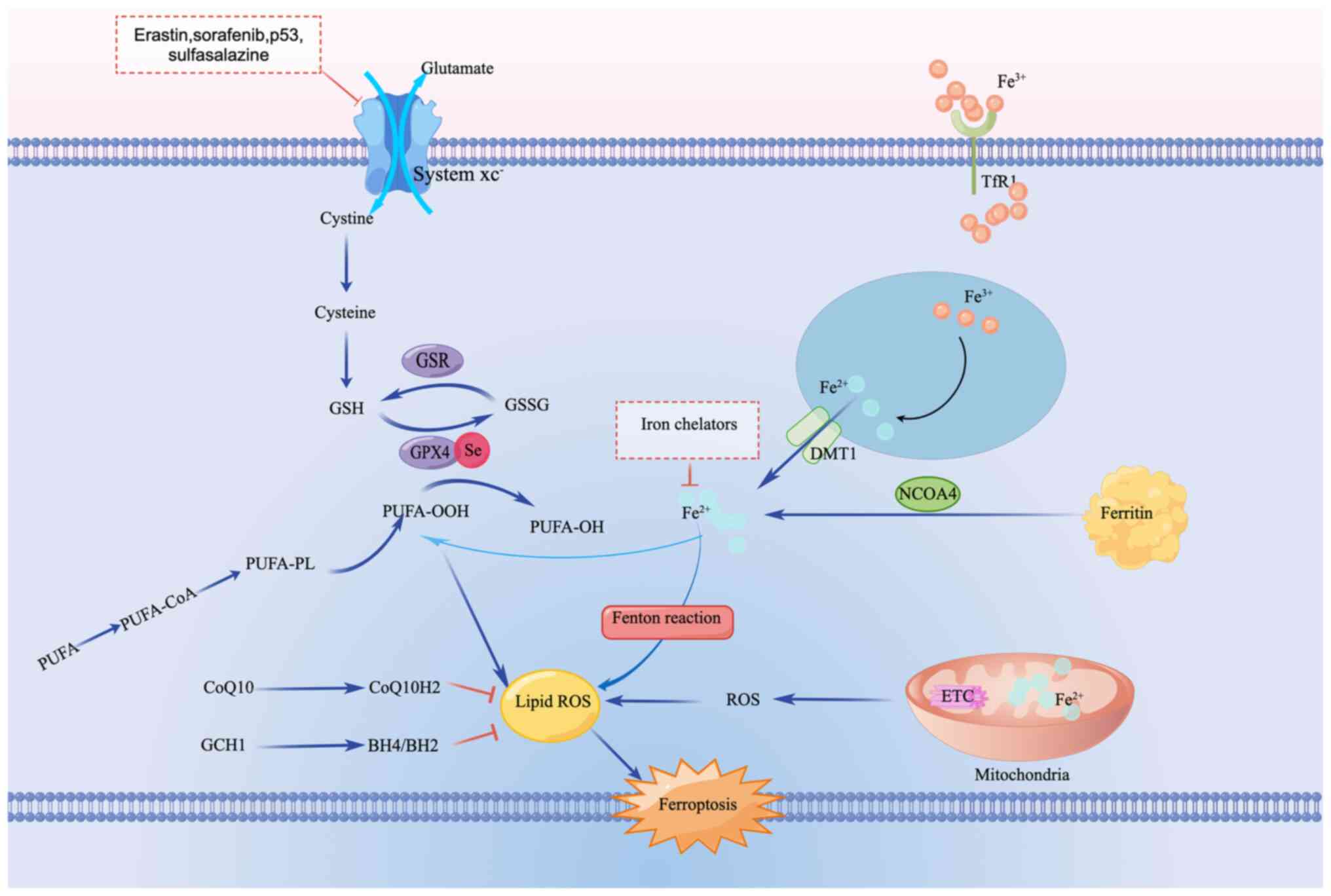
Ferroptosis was proposed in 2012 as a distinctive
type of non-apoptotic cell death (1) characterized by iron overload and
lipid peroxidation. This differs from other forms of regulated cell
death. The primary feature of ferroptosis is mitochondrial
shrinkage, which occurs alongside an increase in mitochondrial
membrane density and degeneration of mitochondrial crista but with
no changes in morphology of the nucleus (1).
Several pathways, including the metabolism of iron,
acid and lipid, have been implicated in ferroptosis (13). Excessive cytosolic Fe2+
catalyzes the Fenton reaction and activates iron-dependent
metabolic enzymes, leading to production of highly reactive
hydroxyl radicals and oxidized polyunsaturated fatty acids (PUFAs),
which results in the promotion of the accumulation of lipid
reactive oxygen species (ROS) and ferroptosis. The metabolism of
amino acids, especially the system Xc/glutathione (GSH)/GSH
peroxidase 4 (GPX4) axis, is key to eliminating lipid ROS, with
GPX4 regarded as a key regulator of ferroptosis. Additionally,
GPX4-independent pathways, such as the NADPH/ferroptosis suppressor
protein 1 (FSP1)/coenzyme Q10 and the GTP
cyclohydrolase-1/tetrahydrobiopterin/dihydrobiopterin axes, have
also been implicated in the ferroptosis process in the past few
years (14-16)
(Fig. 1).
Two types of immunity exist in the body, innate and
adaptive. The innate immune system detects invading pathogens,
while the adaptive immune system promotes a specific and
long-lasting protection against infection. Innate immune cells
mainly include dendritic cells, macrophages and neutrophils, while
the adaptive immune system generally contains T and B lymphocytes
and nature killer (NK) cells (17).
In the last few years, more evidence has revealed a
close association between ferroptosis and the immune system
(18,19). Notably, autoimmune disease is
initiated and propagated by the activation of self-antigen-reactive
T cells (20), pointing to the
crucial role of T cells in autoimmunity. In a study investigating
ferroptosis in immunity, both antigen-specific CD8+ and
CD4+ T cells failed to expand and protect infection in T
cell-specific GPX4-deficient mice (TΔGpx4/ΔGpx4),
whereas GPX4-deficient T cells rapidly accumulated lipid peroxides
and underwent ferroptosis in vitro (21). Ferroptosis was found to be involved
in immunotherapy-activated CD8+ T cells, with increased
ferroptosis contributing to the anti-tumor efficacy of
immunotherapy (22). Recently, the
homeostasis of follicular helper T cells, a specialized subset of
CD4+ T cells, was also shown to be regulated by
ferroptosis (23).
Key features of ferroptosis, iron overload and lipid
peroxidation, participate in immunity. Iron overload increases
oxidative stress and DNA damage in T cells, leading to immune
dysfunction (24), while lipid
peroxidation is associated with intracellular ROS in regulatory T
cells (25). Ferroptosis affects
the viability of B cells, with Muri et al (26) demonstrating that GPX4 is key to the
development, maintenance and responses of B1 and marginal zone B
cells via suppression of ferroptosis. Moreover, the ferroptosis
inducer erastin increases lipid peroxidation and promotes
peripheral blood mononuclear cell proliferation and differentiation
into B and NK cells (27).
In addition to its links to the adaptive immune
system, ferroptosis also plays a key role in innate immunity. In
tumor cells, exogenous circularly polarized magnetic field-induced
ferroptosis leads to the maturation of dendritic cells (28) and in immune-competent mice,
ferroptosis promotes phenotypical maturation of bone-marrow-derived
dendritic cells (29). With regard
to other innate immune cells, ferroptosis is associated with the
infiltration of macrophages and neutrophils (30), while also regulating polarization
of macrophages (31) and the
recruitment of neutrophils (32).
Although the pathology of autoimmune disease is
complex, it is hypothesized that inflammation serves a key role in
autoimmunity (33). DAMPs,
endogenous molecules released by damaged tissue or dying cells,
have been proved to be detrimental in inflammatory response and
lead to the development of inflammatory disorders (34,35).
Autoimmune diseases, such as SLE (36) and IBD (37), are among the inflammatory disorders
initiated by DAMPs. In the host, DAMPs either activate innate
immune cells, leading to release of various cytokines and
chemokines and activation of adaptive immune responses, or
stimulate adaptive immune cells directly (35).
As a key part of regulated cell death, ferroptosis
can stimulate the release of DAMPs. Adenosine triphosphate (ATP)
and high mobility group box 1 (HMGB1), two well-characterized
DAMPs, are released along during ferroptosis in murine fibrosarcoma
MCA205 or glioma GL261(29) and
p53 R273H-expressing non-small cell lung cancer cells (38). Cotreatment with erastin and
celastrol initiates expression of heat shock proteins (HSPs)
(39). Using the
immunoprecipitation assay, an interaction between HSP90 and GPX4
has been demonstrated in a model of acute kidney injury (AKI)
(40). Another DAMP, calreticulin,
also participates in ferroptosis. In the head and neck squamous
cell carcinoma, Zhao et al (41) found ferroptosis reverses
immunosuppressive microenvironments by releasing calreticulin and
HMGB1, while Van Loenhout et al (42) demonstrated that auranofin and
plasma-treated PBS mixture-induced ferroptosis led to a significant
increase in calreticulin, ATP and HMGB1. The aforementioned reports
point to a close link between ferroptosis and DAMPs, which may
partly explain the mechanism of ferroptosis-mediated
autoimmunity.
Autoimmune diseases are also associated with
cytokines and chemokines. In a mouse AKI model, ferrostatin-1 was
shown to prevent upregulation of IL-33(43). Additionally, liproxstatin-1
alleviates radiation-induced lung fibrosis via the downregulation
of TGF-β1(44). Ferroptosis,
therefore, could have an intimate relationship with autoimmune
disease.
Autoimmune diseases are complicated and
characterized by the development of specific autoantibodies and the
presence of autoreactive T cells, leading to the impairment of
sustained immune responses and organs (45). Recent studies have highlighted the
association between ferroptosis and autoimmune disease (Table I).
AIH is an immune-mediated inflammatory liver
disorder characterized by histological abnormality, as well as
elevated aspartate aminotransferase, alanine aminotransferase and
total IgG and the presence of autoantibodies (46).
In concanavalin A (ConA)-induced hepatitis,
redox-active iron accumulation and malondialdehyde (MDA) are
detected in the hepatic tissues of mice. Moreover, the expression
of GPX4 and system xc- is markedly decreased in the
liver of ConA-treated mice and is accompanied by the downregulation
of caveolin-1(47). In LO2
hepatocyte cell line, the overexpression of caveolin-1 results in
the upregulation of the expression of system xc-,
suggesting that cavelolin-1 protects against ConA-induced AIH by
inhibiting ferroptosis. In another study, levels of cyclooxygenase2
and acyl-coenzyme A synthase long-chain family member 4 (ACSL4)
were shown to be upregulated in the liver tissue of S100-induced
AIH model mice, while the levels of GPX4 and ferritin heavy chain 1
(FTH1) are downregulated (48). In
addition, GPX4 knockdown via adeno-associated virus injection
aggravates severity of S100-induced AIH. The aforementioned studies
suggested that ferroptosis is a possible mediator of AIH.
RA is the most common autoimmune inflammatory
arthritis in adults and is characterized by chronic destructive
synovitis and multisystem disorder (49,50).
In the rheumatoid synovium and synovial fluid of
patients with RA, the levels of iron accumulation and lipid
peroxidation increase, while in a collagen-induced arthritis (CIA)
mouse model, selectively targeting fibroblasts in vivo to
induce ferroptosis attenuates arthritis progression, indicating
that ferroptosis inducers serve as candidates for RA treatment
(51). Sulfasalazine, a U.S. Food
and Drug Administration-approved RA drug, is an effective inducer
of ferroptosis (52). As shown by
Ling et al (53), the
expression of ACSL4 declines, while the expression of FTH1, GPX4,
and cystine/glutamate antiporter solute carrier family 7 member 11
are increased in the RA synovium of CIA model mouse compared with
healthy control (53). Luo and
Zhang (54) showed that in a
lipopolysaccharide-induced synovitis cell model, MDA levels are
increased, whereas GPX4 levels are decreased, representing an
increase in ferroptosis in human synoviocytes; inhibition of
ferroptosis may be a new therapeutic strategy for synovitis
(54). Further studies are
required to identify the exact role of ferroptosis in RA.
SLE is a multisystem autoimmune disease
characterized by formation of autoantibodies, deposition of immune
complexes and inflammation that primarily presents in women of
reproductive age. The pathogenetic mechanisms of SLE are complex
and this disorder is prone to relapse and remissions, leading to
considerable morbidity and mortality (55,56).
IBD, including Crohn's disease (CD) and ulcerative
colitis (UC), is a complex chronic inflammation disorder that
arises due to dysregulated immune response (61). Smoking, diet, lifestyle and
behavior, as well as gut microbiota, are all key contributors to
disease pathogenesis (62).
MS is considered an autoimmune disorder of the
central nervous system that is characterized by inflammation,
demyelination and degeneration (68,69).
mRNA levels of GPX4 in the brain of patients with MS are decreased,
while the levels of GPX4 mRNA and protein are decreased and lipid
peroxidation is enhanced in an experimental autoimmune
encephalomyelitis mouse model (70). Jhelum et al (71) investigated the underlying mechanism
of cuprizone (CZ), a copper chelator, used to induce
oligodendrocyte (OL) cell loss and demyelination, revealing that CZ
treatment resulted in an increase in mRNA expression of nuclear
receptor coactivator 4, transferrin receptor 1 and PTGS2, as well
as lipid peroxidation, and a decrease in the expression of GPX4 and
system xc- in the brain tissue of experimental mice.
Additionally, the CZ-induced loss of OL and demyelination was
prevented by ferrostatin-1(71).
These results indicated that ferroptosis is a potential therapeutic
target for MS.
Psoriasis is a chronic immune-mediated inflammatory
skin disease characterized by hyperproliferation of keratinocytes
and excessive infiltration of immune cells. Currently, it is
considered a systemic disease associated with metabolic, arthritic
and cardiovascular comorbidities (80).
A previous study showed a significant reduction in
GPX4 and elevation in Nrf2 downstream targets in psoriatic skin
lesions compared with samples from healthy patients (81). Additionally, the mRNA levels of
ACSL4, PTGS2 and TFR are much higher in psoriasis lesions than in
healthy controls. Furthermore, in an imiquimod (IMQ)-induced mouse
model of psoriasis, immunohistochemical analysis uncovered notably
increased ACSL4 levels and markedly decreased GPX4 levels in the
basal epidermal layer and ferrostatin-1 treatment attenuated
IMQ-induced psoriasis-like dermatitis (82). Ferroptosis is, therefore, a
potential physiological mechanism for eliminating inflammatory
response in psoriasis.
IDDM is a chronic disorder stemming from autoimmune
damage of pancreatic β cells (83). While ferroptosis is involved in
cell death of the myocardium and renal tubules during diabetes
(84-87),
the role of ferroptosis in the death of β cells is unknown. In
2018, Bruni et al reported massive ferroptosis in pancreatic
islets isolated from IDDM patients, whereas the transplantable
number of islet equivalents increased following addition of
ferrostatin (88). Along with
evidence that ferroptosis is induced in rat pancreatic β cells
after exposure to tert-butyl hydroperoxide (89), ferroptosis can be considered a
possible mode of β cell destruction. However, more studies are
required to determine the link between ferroptosis and β cell
death.
As aforementioned, there is an association between
ferroptosis and autoimmune disease. Therefore, targeting
ferroptosis is a promising therapeutic option for autoimmune
disease. Ferroptosis can primarily be inhibited by iron chelators
and lipophilic antioxidants (1).
The present review summarizes anti-ferroptosis agents and their
potential benefits in the treatment of autoimmune disorder
(Table II).
Deferoxamine (DFO) has been investigated in the
treatment of several types of autoimmune disease. In patients with
RA, DFO prevents synovial injury (90) and improves anemia (91). A pilot study showed that patients
with MS tolerate a short course of DFO therapy relatively well
(92), however, no effect on
disease progression has been noted (93). In addition to its effect against RA
and MS, DFO has been found to ameliorate motor defects and
pathology in a PD rat model (94).
Deferiprone (DFP), another iron chelator, suppresses disease
activity in a mouse model of MS (95). Additionally, DFP reportedly
improves motor performance of patients with PD in a phase II
clinical trial (96). DFP can
ameliorate DSS-induced UC in a mouse model by suppressing
ferroptosis (67). The
aforementioned studies indicate that iron chelators are promising
therapeutic options for autoimmune disease. However, larger clinal
trials are needed to determine the value of iron chelators in the
therapy of autoimmune disease.
Ferrostatin-1 and liproxstatin-1 are well-known
inhibitors of ferroptosis. Numerous studies have investigated these
ferroptosis inhibitors in autoimmune diseases, including AIH, IBD
and PD (48,97). To the best of our knowledge,
however, examinations have yet to be conducted in models other than
experimental mouse models. Hence, clinical trials must be performed
to explore their roles in patients.
Vitamin E is a key lipid soluble antioxidant that
can suppress ferroptosis by inhibiting 15-lipoxygenase (98). Reports show that supplementation
with vitamin E relieves joint pain in patients with RA (99). In patients with SLE, vitamin E is
said to suppress autoantibody production. Moreover, vitamin E
improves functional capacity and gait parameters in patients with
relapsing-remitting MS (100) and
improves clinical signs and metabolic status in patients with PD
(101). Furthermore,
supplementation with vitamin E is a feasible option for the
management of patients with severe forms of psoriasis as it
decreases the markers of oxidative stress (102). Vitamin E treatment, therefore,
may be a therapeutic option for autoimmune diseases.
Selenium, an essential trace element with
antioxidant properties, has been assessed as a potential treatment
for autoimmune diseases. Supplementation with selenium relieves
inflammatory reaction in patients with MS (103), IBD (104) and psoriasis (102). However, it has shown no
significant clinical benefit against RA (105,106). In preclinical studies, selenium
decreases loss of dopamine and slows the progression of
neurodegeneration during PD (107).
N-acetylcysteine, a pharmaceutical drug with an
anti-ferroptosis property, has been investigated in the treatment
of RA and SLE. In patients with RA, the oral administration of
N-acetylcysteine relieves severity of joint pain and improves
physical performance (108). In
patients with SLE, N-acetylcysteine inhibits lupus disease activity
(109).
Polyphenols are natural antioxidants that prevent
ferroptosis owing to their ROS scavenging property (110). Resveratrol, a well-studied
polyphenol, decreases disease activity score assessment for 28
joints in patients with RA (111), decreases the clinical colitis
activity index score and improves quality of life in patients with
UC (112,113) and exerts antidiabetic and
antioxidant effects in patients with IDDM (114). Other polyphenols, such as
pomegranate juice could alleviate disease activity of patients with
RA (115), and licorice could
improve symptoms in patients with PD (116).
Although the aforementioned experiments posited
lipophilic antioxidants as having potential role in the therapy of
autoimmune disease, larger and more in-depth studies are required
to determine the exact impact of various polyphenols in the
treatment of autoimmune disorder.
The present review summarized research on
ferroptosis in autoimmune disorders and discussed ferroptosis as a
promising therapeutic target. Although autoimmune diseases are
heterogeneous in manifestation, there are commonalities between
these disorders with respect to ferroptosis. Among these key
commonalities is inflammation (117,118). As an important part of regulated
cell death, ferroptosis stimulates release of DAMPs and
inflammatory cytokines, leading to activation of immune response
and eventually promoting the development of autoimmune disease.
Even though recent evaluations have investigated
ferroptosis in autoimmune disorders, the association between this
cell death type and autoimmune diseases is relatively undeveloped.
Therefore, more studies are required to determine the association
between ferroptosis and autoimmune disease, including Graves'
disease, Hashimoto thyroiditis, coeliac disease, Addison disease
and autoimmune myocarditis and polyendocrine syndrome type 2.
Additionally, although numerous ferroptosis-related reagents have
been investigated in the treatment of various autoimmune diseases,
the reported efficacy pertains mainly to basic studies, patient
sample sizes and follow-up periods were relatively limited. Hence,
larger clinical trials must be performed to highlight and confirm
the treatment values of ferroptosis-associated regents.
In summary, ferroptosis plays a critical role in the
pathogenesis of autoimmune diseases and is a promising therapeutic
target for autoimmune diseases.
Funding: The present study was supported by the Zhejiang
Provincial Science Foundation of China (grant no. LGF21H020006),
the Pioneer Innovation Team of Jiaxing Institute of Atherosclerotic
Diseases (grant no. XFCX-DMYH), Program of the First Hospital of
Jiaxing (grant no. 2021-YA-001), the Jiaxing Key Laboratory of
Arteriosclerotic Diseases (grant no. 2020-dmzdsys) and Jiaxing Key
Laboratory of Virus-mediated Infectious Diseases (grant no.
2021-bdzdsys).
Not applicable.
LS and XW drafted the manuscript. CZ edited the
manuscript. YC designed the study and revised the manuscript. All
authors have read and approved the final manuscript. Data
authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta
R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS,
et al: Ferroptosis: An iron-dependent form of nonapoptotic cell
death. Cell. 149:1060–1072. 2012.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Alvarez SW, Sviderskiy VO, Terzi EM,
Papagiannakopoulos T, Moreira AL, Adams S, Sabatini DM, Birsoy K
and Possemato R: NFS1 undergoes positive selection in lung tumours
and protects cells from ferroptosis. Nature. 551:639–643.
2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Lee JY, Nam M, Son HY, Hyun K, Jang SY,
Kim JW, Kim MW, Jung Y, Jang E, Yoon SJ, et al: Polyunsaturated
fatty acid biosynthesis pathway determines ferroptosis sensitivity
in gastric cancer. Proc Natl Acad Sci USA. 117:32433–32442.
2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Friedmann Angeli JP, Schneider M, Proneth
B, Tyurina YY, Tyurin VA, Hammond VJ, Herbach N, Aichler M, Walch
A, Eggenhofer E, et al: Inactivation of the ferroptosis regulator
Gpx4 triggers acute renal failure in mice. Nat Cell Biol.
16:1180–1191. 2014.PubMed/NCBI View
Article : Google Scholar
|
|
5
|
Fang X, Wang H, Han D, Xie E, Yang X, Wei
J, Gu S, Gao F, Zhu N, Yin X, et al: Ferroptosis as a target for
protection against cardiomyopathy. Natl Acad Sci USA.
116:2672–2680. 2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Park MW, Cha HW, Kim J, Kim JH, Yang H,
Yoon S, Boonpraman N, Yi SS, Yoo ID and Moon JS: NOX4 promotes
ferroptosis of astrocytes by oxidative stress-induced lipid
peroxidation via the impairment of mitochondrial metabolism in
Alzheimer's diseases. Redox Biol. 41(101947)2021.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Do Van B, Gouel F, Jonneaux A, Timmerman
K, Gelé P, Pétrault M, Bastide M, Laloux C, Moreau C, Bordet R, et
al: Ferroptosis, a newly characterized form of cell death in
Parkinson's disease that is regulated by PKC. Neurobiol Dis.
94:169–178. 2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Hayter SM and Cook MC: Updated assessment
of the prevalence, spectrum and case definition of autoimmune
disease. Autoimmun Rev. 11:754–765. 2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Cho JH and Gregersen PK: Genomics and the
multifactorial nature of human autoimmune disease. N Engl J Med.
365:1612–1623. 2011.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Lai B, Wu CH, Wu CY, Luo SF and Lai JH:
Ferroptosis and autoimmune diseases. Front Immunol.
13(916664)2022.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Fan J, Jiang T and He D: Emerging insights
into the role of ferroptosis in the pathogenesis of autoimmune
diseases. Front Immunol. 14(1120519)2023.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Ohl K, Rauen T and Tenbrock K:
Dysregulated neutrophilic cell death in SLE: A spotlight on
ferroptosis. Signal Transduct Target Ther. 6(392)2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Hu H, Chen Y, Jing L, Zhai C and Shen L:
The link between ferroptosis and cardiovascular diseases: A novel
target for treatment. Front Cardiovasc Med.
8(710963)2021.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Bersuker K, Hendricks JM, Li Z, Magtanong
L, Ford B, Tang PH, Roberts MA, Tong B, Maimone TJ, Zoncu R, et al:
The CoQ oxidoreductase FSP1 acts parallel to GPX4 to inhibit
ferroptosis. Nature. 575:688–692. 2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Kraft VAN, Bezjian CT, Pfeiffer S,
Ringelstetter L, Müller C, Zandkarimi F, Merl-Pham J, Bao X,
Anastasov N, Kössl J, et al: GTP cyclohydrolase
1/tetrahydrobiopterin counteract ferroptosis through lipid
remodeling. ACS Cent Sci. 6:41–53. 2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Doll S, Freitas FP, Shah R, Aldrovandi M,
da Silva MC, Ingold I, Goya Grocin A, Xavier da Silva TN, Panzilius
E, Scheel CH, et al: FSP1 is a glutathione-independent ferroptosis
suppressor. Nature. 575:693–698. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Iwasaki A and Medzhitov R: Control of
adaptive immunity by the innate immune system. Nat Immunol.
16:343–353. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
18
|
Yang Y, Wang Y, Guo L, Gao W, Tang TL and
Yan M: Interaction between macrophages and ferroptosis. Cell Death
Dis. 13(355)2022.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Stockwell BR and Jiang X: A physiological
function for ferroptosis in tumor suppression by the immune system.
Cell Metab. 30:14–15. 2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Akirav EM, Ruddle NH and Herold KC: The
role of AIRE in human autoimmune disease. Nat Rev Endocrinol.
7:25–33. 2011.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Matsushita M, Freigang S, Schneider C,
Conrad M, Bornkamm GW and Kopf M: T cell lipid peroxidation induces
ferroptosis and prevents immunity to infection. J Exp Med.
212:555–568. 2015.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wang W, Green M, Choi JE, Gijón M, Kennedy
PD, Johnson JK, Liao P, Lang X, Kryczek I, Sell A, et al:
CD8+ T cells regulate tumour ferroptosis during cancer
immunotherapy. Nature. 569:270–274. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Yao Y, Chen Z, Zhang H, Chen C, Zeng M,
Yunis J, Wei Y, Wan Y, Wang N, Zhou M, et al: Selenium-GPX4 axis
protects follicular helper T cells from ferroptosis. Nat Immunol.
22:1127–1139. 2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Shaw J, Chakraborty A, Nag A, Chattopadyay
A, Dasgupta AK and Bhattacharyya M: Intracellular iron overload
leading to DNA damage of lymphocytes and immune dysfunction in
thalassemia major patients. Eur J Haematol. 99:399–408.
2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Anupam K, Kaushal J, Prabhakar N and
Bhatnagar A: Effect of redox status of peripheral blood on immune
signature of circulating regulatory and cytotoxic T cells in
streptozotocin induced rodent model of type I diabetes.
Immunobiology. 223:586–597. 2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Muri J, Thut H, Bornkamm GW and Kopf M: B1
and marginal zone B cells but not follicular B2 cells require Gpx4
to prevent lipid peroxidation and ferroptosis. Cell Rep.
29:2731–2744.e4. 2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Wang D, Xie N, Gao W, Kang R and Tang D:
The ferroptosis inducer erastin promotes proliferation and
differentiation in human peripheral blood mononuclear cells.
Biochem Biophys Res Commun. 503:1689–1695. 2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Yu B, Choi B, Li W and Kim DH: Magnetic
field boosted ferroptosis-like cell death and responsive MRI using
hybrid vesicles for cancer immunotherapy. Nat Commun.
11(3637)2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Efimova I, Catanzaro E, Van der Meeren L,
Turubanova VD, Hammad H, Mishchenko TA, Vedunova MV, Fimognari C,
Bachert C, Coppieters F, et al: Vaccination with early ferroptotic
cancer cells induces efficient antitumor immunity. J Immunother
Cancer. 8(e001369)2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Zhang Z, Qiu X, Yan Y, Liang Q, Cai Y,
Peng B, Xu Z and Xia F: Evaluation of ferroptosis-related gene
AKR1C1 as a novel biomarker associated with the immune
microenvironment and prognosis in breast cancer. Int J Gen Med.
14:6189–6200. 2021.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Dai E, Han L, Liu J, Xie Y, Kroemer G,
Klionsky DJ, Zeh HJ, Kang R, Wang J and Tang D: Autophagy-dependent
ferroptosis drives tumor-associated macrophage polarization via
release and uptake of oncogenic KRAS protein. Autophagy.
16:2069–2083. 2020.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Li W, Feng G, Gauthier JM, Lokshina I,
Higashikubo R, Evans S, Liu X, Hassan A, Tanaka S, Cicka M, et al:
Ferroptotic cell death and TLR4/Trif signaling initiate neutrophil
recruitment after heart transplantation. J Clin Invest.
129:2293–2304. 2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Hall SW and Cooke A: Autoimmunity and
inflammation: Murine models and translational studies. Mamm Genome.
22:377–389. 2011.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Roh JS and Sohn DH: Damage-associated
molecular patterns in inflammatory diseases. Immune Netw.
18(e27)2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Gong T, Liu L, Jiang W and Zhou R:
DAMP-sensing receptors in sterile inflammation and inflammatory
diseases. Nat Rev Immunol. 20:95–112. 2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Schaper F, Westra J and Bijl M: Recent
developments in the role of high-mobility group box 1 in systemic
lupus erythematosus. Mol Med. 20:72–79. 2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Boyapati RK, Dorward DA, Tamborska A,
Kalla R, Ventham NT, Doherty MK, Whitfield PD, Gray M, Loane J,
Rossi AG, et al: Mitochondrial DNA is a pro-inflammatory
damage-associated molecular pattern released during active IBD.
Inflamm Bowel Dis. 24:2113–2122. 2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Freire Boullosa L, Van Loenhout J,
Flieswasser T, De Waele J, Hermans C, Lambrechts H, Cuypers B,
Laukens K, Bartholomeus E, Siozopoulou V, et al: Auranofin reveals
therapeutic anticancer potential by triggering distinct molecular
cell death mechanisms and innate immunity in mutant p53 non-small
cell lung cancer. Redox Biol. 42(101949)2021.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Liu M, Fan Y, Li D, Han B, Meng Y, Chen F,
Liu T, Song Z, Han Y, Huang L, et al: Ferroptosis inducer erastin
sensitizes NSCLC cells to celastrol through activation of the
ROS-mitochondrial fission-mitophagy axis. Mol Oncol. 15:2084–2105.
2021.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Chen C, Wang D, Yu Y, Zhao T, Min N, Wu Y,
Kang L, Zhao Y, Du L, Zhang M, et al: Legumain promotes tubular
ferroptosis by facilitating chaperone-mediated autophagy of GPX4 in
AKI. Cell Death Dis. 12(65)2021.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Zhao YY, Lian JX, Lan Z, Zou KL, Wang WM
and Yu GT: Ferroptosis promotes anti-tumor immune response by
inducing immunogenic exposure in HNSCC. Oral Dis. 29:933–941.
2023.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Van Loenhout J, Freire Boullosa L,
Quatannens D, De Waele J, Merlin C, Lambrechts H, Lau HW, Hermans
C, Lin A, Lardon F, et al: Auranofin and cold atmospheric plasma
synergize to trigger distinct cell death mechanisms and immunogenic
responses in glioblastoma. Cells. 10(2936)2021.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Martin-Sanchez D, Ruiz-Andres O, Poveda J,
Carrasco S, Cannata-Ortiz P, Sanchez-Niño MD, Ruiz Ortega M, Egido
J, Linkermann A, Ortiz A and Sanz AB: Ferroptosis, but not
necroptosis, is important in nephrotoxic folic acid-induced AKI. J
Am Soc Nephrol. 28:218–229. 2017.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Li X, Duan L, Yuan S, Zhuang X, Qiao T and
He J: Ferroptosis inhibitor alleviates radiation-induced lung
fibrosis (RILF) via down-regulation of TGF-β1. J Inflamm (Lond).
16(11)2019.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Zhernakova A, Withoff S and Wijmenga C:
Clinical implications of shared genetics and pathogenesis in
autoimmune diseases. Nat Rev Endocrinol. 9:646–659. 2013.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Mack CL, Adams D, Assis DN, Kerkar N,
Manns MP, Mayo MJ, Vierling JM, Alsawas M, Murad MH and Czaja AJ:
Diagnosis and management of autoimmune hepatitis in adults and
children: 2019 Practice guidance and guidelines from the American
association for the study of liver diseases. Hepatology.
72:671–722. 2020.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Deng G, Li Y, Ma S, Gao Z, Zeng T, Chen L,
Ye H, Yang M, Shi H, Yao X, et al: Caveolin-1 dictates ferroptosis
in the execution of acute immune-mediated hepatic damage by
attenuating nitrogen stress. Free Radic Biol Med. 148:151–161.
2020.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Zhu L, Chen D, Zhu Y, Pan T, Xia D, Cai T,
Lin H, Lin J, Jin X, Wu F, et al: GPX4-regulated ferroptosis
mediates S100-induced experimental autoimmune hepatitis associated
with the Nrf2/HO-1 signaling pathway. Oxid Med Cell Longev.
2021(6551069)2021.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Singh JA, Saag KG, Bridges SL Jr, Akl EA,
Bannuru RR, Sullivan MC, Vaysbrot E, McNaughton C, Osani M,
Shmerling RH, et al: 2015 American college of Rheumatology
guideline for the treatment of rheumatoid arthritis. Arthritis
Rheumatol. 68:1–26. 2016.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Fraenkel L, Bathon JM, England BR, St
Clair EW, Arayssi T, Carandang K, Deane KD, Genovese M, Huston KK,
Kerr G, et al: 2021 American college of Rheumatology guideline for
the treatment of rheumatoid arthritis. Arthritis Rheumatol.
73:1108–1123. 2021.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Wu J, Feng Z, Chen L, Li Y, Bian H, Geng
J, Zheng ZH, Fu X, Pei Z, Qin Y, et al: TNF antagonist sensitizes
synovial fibroblasts to ferroptotic cell death in collagen-induced
arthritis mouse models. Nat Commun. 13(676)2022.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Floros KV, Cai J, Jacob S, Kurupi R,
Fairchild CK, Shende M, Coon CM, Powell KM, Belvin BR, Hu B, et al:
MYCN-amplified neuroblastoma is addicted to iron and vulnerable to
inhibition of the system Xc-/glutathione axis. Cancer Res.
81:1896–1908. 2021.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Ling H, Li M, Yang C, Sun S, Zhang W, Zhao
L, Xu N, Zhang J, Shen Y, Zhang X, et al: Glycine increased
ferroptosis via SAM-mediated GPX4 promoter methylation in
rheumatoid arthritis. Rheumatology (Oxford). 61:4521–4534.
2022.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Luo H and Zhang R: Icariin enhances cell
survival in lipopolysaccharide-induced synoviocytes by suppressing
ferroptosis via the Xc-/GPX4 axis. Exp Ther Med.
21(72)2021.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Gordon C, Amissah-Arthur MB, Gayed M,
Brown S, Bruce IN, D'Cruz D, Empson B, Griffiths B, Jayne D,
Khamashta M, et al: The British Society for Rheumatology guideline
for the management of systemic lupus erythematosus in adults.
Rheumatology (Oxford). 57:e1–e45. 2018.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Tsokos GC, Lo MS, Costa Reis P and
Sullivan KE: New insights into the immunopathogenesis of systemic
lupus erythematosus. Nat Rev Rheumatol. 12:716–730. 2016.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Indrakanti DL, Alvarado A, Zhang X,
Birmingham DJ, Hinton A and Rovin BH: The
interleukin-6-hepcidin-hemoglobin circuit in systemic lupus
erythematosus flares. Lupus. 26:200–203. 2017.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Chen W, Li W, Zhang Z, Tang X, Wu S, Yao
G, Li K, Wang D, Xu Y, Feng R, et al: Lipocalin-2 exacerbates lupus
nephritis by promoting Th1 cell differentiation. J Am Soc Nephrol.
31:2263–2277. 2020.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Scindia Y, Wlazlo E, Ghias E, Cechova S,
Loi V, Leeds J, Ledesma J, Helen C and Swaminathan S: Modulation of
iron homeostasis with hepcidin ameliorates spontaneous murine lupus
nephritis. Kidney Int. 98:100–115. 2020.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Li P, Jiang M, Li K, Li H, Zhou Y, Xiao X,
Xu Y, Krishfield S, Lipsky PE, Tsokos GC and Zhang X: Glutathione
peroxidase 4-regulated neutrophil ferroptosis induces systemic
autoimmunity. Nat Immunol. 22:1107–1117. 2021.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Ananthakrishnan AN: Epidemiology and risk
factors for IBD. Nat Rev Gastroenterol Hepatol. 12:205–217.
2015.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Guan Q: A comprehensive review and update
on the pathogenesis of inflammatory bowel disease. J Immunol Res.
2019(7247238)2019.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Mayr L, Grabherr F, Schwärzler J,
Reitmeier I, Sommer F, Gehmacher T, Niederreiter L, He GW, Ruder B,
Kunz KTR, et al: Dietary lipids fuel GPX4-restricted enteritis
resembling Crohn's disease. Nat Commun. 11(1775)2020.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Xu J, Liu S, Cui Z, Wang X, Ning T, Wang
T, Zhang N, Xie S, Min L, Zhang S, et al: Ferrostatin-1 alleviated
TNBS induced colitis via the inhibition of ferroptosis. Biochem
Biophys Res Commun. 573:48–54. 2021.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Xu M, Tao J, Yang Y, Tan S, Liu H, Jiang
J, Zheng F and Wu B: Ferroptosis involves in intestinal epithelial
cell death in ulcerative colitis. Cell Death Dis.
11(86)2020.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Chen Y, Wang J, Li J, Zhu J, Wang R, Xi Q,
Wu H, Shi T and Chen W: Astragalus polysaccharide prevents
ferroptosis in a murine model of experimental colitis and human
Caco-2 cells via inhibiting NRF2/HO-1 pathway. Eur J Pharmacol.
911(174518)2021.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Chen Y, Zhang P, Chen W and Chen G:
Ferroptosis mediated DSS-induced ulcerative colitis associated with
Nrf2/HO-1 signaling pathway. Immunol Lett. 225:9–15.
2020.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Montalban X, Gold R, Thompson AJ,
Otero-Romero S, Amato MP, Chandraratna D, Clanet M, Comi G, Derfuss
T, Fazekas F, et al: ECTRIMS/EAN guideline on the pharmacological
treatment of people with multiple sclerosis. Eur J Neurol.
25:215–237. 2018.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Hemmer B, Archelos JJ and Hartung HP: New
concepts in the immunopathogenesis of multiple sclerosis. Nat Rev
Neurosci. 3:291–301. 2002.PubMed/NCBI View
Article : Google Scholar
|
|
70
|
Hu CL, Nydes M, Shanley KL, Morales
Pantoja IE, Howard TA and Bizzozero OA: Reduced expression of the
ferroptosis inhibitor glutathione peroxidase-4 in multiple
sclerosis and experimental autoimmune encephalomyelitis. J
Neurochem. 148:426–439. 2019.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Jhelum P, Santos-Nogueira E, Teo W,
Haumont A, Lenoël I, Stys PK and David S: Ferroptosis mediates
cuprizone-induced loss of oligodendrocytes and demyelination. J
Neurosci. 40:9327–9341. 2020.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Stewart DA: NICE guideline for Parkinson's
disease. Age Ageing. 36:240–242. 2007.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Sulzer D, Alcalay RN, Garretti F, Cote L,
Kanter E, Agin-Liebes J, Liong C, McMurtrey C, Hildebrand WH, Mao
X, et al: T cells from patients with Parkinson's disease recognize
α-synuclein peptides. Nature. 546:656–661. 2017.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Thapa K, Khan H, Kanojia N, Singh TG, Kaur
A and Kaur G: Therapeutic insights on ferroptosis in Parkinson's
disease. Eur J Pharmacol. 930(175133)2022.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Zhang P, Chen L, Zhao Q, Du X, Bi M, Li Y,
Jiao Q and Jiang H: Ferroptosis was more initial in cell death
caused by iron overload and its underlying mechanism in Parkinson's
disease. Free Radic Biol Med. 152:227–234. 2020.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Dexter DT, Wells FR, Lees AJ, Agid F, Agid
Y, Jenner P and Marsden CD: Increased nigral iron content and
alterations in other metal ions occurring in brain in Parkinson's
disease. J Neurochem. 52:1830–1836. 1989.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Dexter DT, Carter CJ, Wells FR, Javoy-Agid
F, Agid Y, Lees A, Jenner P and Marsden CD: Basal lipid
peroxidation in substantia nigra is increased in Parkinson's
disease. J Neurochem. 52:381–389. 1989.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Sofic E, Lange KW, Jellinger K and
Riederer P: Reduced and oxidized glutathione in the substantia
nigra of patients with Parkinson's disease. Neurosci Lett.
142:128–130. 1992.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Zuo Y, Xie J, Li X, Li Y, Thirupathi A,
Zhang J, Yu P, Gao G, Chang Y and Shi Z: Ferritinophagy-mediated
ferroptosis involved in paraquat-induced neurotoxicity of
dopaminergic neurons: Implication for neurotoxicity in PD. Oxid Med
Cell Longev. 2021(9961628)2021.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Parisi R, Iskandar IYK, Kontopantelis E,
Augustin M, Griffiths CEM and Ashcroft DM: Global Psoriasis Atlas.
National, regional, and worldwide epidemiology of psoriasis:
Systematic analysis and modelling study. BMJ.
369(m1590)2020.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Arbiser JL, Bonner MY, Ward N, Elsey J and
Rao S: Selenium unmasks protective iron armor: A possible defense
against cutaneous inflammation and cancer. Biochim Biophys Acta Gen
Subj. 1862:2518–2527. 2018.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Shou Y, Yang L, Yang Y and Xu J:
Inhibition of keratinocyte ferroptosis suppresses psoriatic
inflammation. Cell Death Dis. 12(1009)2021.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Atkinson MA and Maclaren NK: The
pathogenesis of insulin-dependent diabetes mellitus. N Engl J Med.
331:1428–1436. 1994.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Zang H, Wu W, Qi L, Tan W, Nagarkatti P,
Nagarkatti M, Wang X and Cui T: Autophagy inhibition enables Nrf2
to exaggerate the progression of diabetic cardiomyopathy in mice.
Diabetes. 69:2720–2734. 2020.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Ni T, Huang X, Pan S and Lu Z: Inhibition
of the long non-coding RNA ZFAS1 attenuates ferroptosis by sponging
miR-150-5p and activates CCND2 against diabetic cardiomyopathy. J
Cell Mol Med. 25:9995–10007. 2021.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Li S, Zheng L, Zhang J, Liu X and Wu Z:
Inhibition of ferroptosis by up-regulating Nrf2 delayed the
progression of diabetic nephropathy. Free Radic Biol Med.
162:435–449. 2021.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Kim S, Kang SW, Joo J, Han SH, Shin H, Nam
BY, Park J, Yoo TH, Kim G, Lee P and Park JT: Characterization of
ferroptosis in kidney tubular cell death under diabetic conditions.
Cell Death Dis. 12(160)2021.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Bruni A, Bornstein S, Linkermann A and
Shapiro AMJ: Regulated cell death seen through the lens of islet
transplantation. Cell Transplant. 27:890–901. 2018.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Krümmel B, Plötz T, Jörns A, Lenzen S and
Mehmeti I: The central role of glutathione peroxidase 4 in the
regulation of ferroptosis and its implications for pro-inflammatory
cytokine-mediated beta-cell death. Biochim Biophys Acta Mol Basis
Dis. 1867(166114)2021.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Fudman EJ, Till GO and Fox IH:
Deferoxamine induced decreases of lipid peroxides in rheumatoid
arthritis. J Rheumatol. 14:686–691. 1987.PubMed/NCBI
|
|
91
|
Salvarani C, Baricchi R, Lasagni D,
Boiardi L, Piccinini R, Brunati C, Macchioni P and Portioli I:
Effects of desferrioxamine therapy on chronic disease anemia
associated with rheumatoid arthritis. Rheumatol Int. 16:45–48.
1996.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Lynch SG, Peters K and LeVine SM:
Desferrioxamine in chronic progressive multiple sclerosis: A pilot
study. Mult Scler. 2:157–160. 1996.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Lynch SG, Fonseca T and LeVine SM: A
multiple course trial of desferrioxamine in chronic progressive
multiple sclerosis. Cell Mol Biol (Noisy-le-grand). 46:865–869.
2000.PubMed/NCBI
|
|
94
|
Febbraro F, Andersen KJ, Sanchez-Guajardo
V, Tentillier N and Romero-Ramos M: Chronic intranasal deferoxamine
ameliorates motor defects and pathology in the α-synuclein rAAV
Parkinson's model. Exp Neurol. 247:45–58. 2013.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Sweeney ME, Slusser JG, Lynch SG, Benedict
SH, Garcia SL, Rues L and LeVine SM: Deferiprone modulates in vitro
responses by peripheral blood T cells from control and
relapsing-remitting multiple sclerosis subjects. Int
Immunopharmacol. 11:1796–1801. 2011.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Devos D, Moreau C, Devedjian JC, Kluza J,
Petrault M, Laloux C, Jonneaux A, Ryckewaert G, Garçon G, Rouaix N,
et al: Targeting chelatable iron as a therapeutic modality in
Parkinson's disease. Antioxid Redox Signal. 21:195–210.
2014.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Yang B, Hou S, Huang S, Li H and Li Y:
Ferroptosis inhibitor regulates the disease progression of
systematic lupus erythematosus mice model through Th1/Th2 ratio.
Curr Mol Med: May 25, 2022 (Epub ahead of print).
|
|
98
|
Hinman A, Holst CR, Latham JC, Bruegger
JJ, Ulas G, McCusker KP, Amagata A, Davis D, Hoff KG, Kahn-Kirby
AH, et al: Vitamin E hydroquinone is an endogenous regulator of
ferroptosis via redox control of 15-lipoxygenase. PLoS One.
13(e0201369)2018.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Edmonds SE, Winyard PG, Guo R, Kidd B,
Merry P, Langrish-Smith A, Hansen C, Ramm S and Blake DR: Putative
analgesic activity of repeated oral doses of vitamin E in the
treatment of rheumatoid arthritis. Results of a prospective placebo
controlled double blind trial. Ann Rheum Dis. 56:649–655.
1997.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Aristotelous P, Stefanakis M, Pantzaris M,
Pattichis CS, Calder PC, Patrikios IS, Sakkas GK and Giannaki CD:
The effects of specific omega-3 and omega-6 polyunsaturated fatty
acids and antioxidant vitamins on gait and functional capacity
parameters in patients with relapsing-remitting multiple sclerosis.
Nutrients. 13(3661)2021.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Taghizadeh M, Tamtaji OR, Dadgostar E,
Daneshvar Kakhaki R, Bahmani F, Abolhassani J, Aarabi MH, Kouchaki
E, Memarzadeh MR and Asemi Z: The effects of omega-3 fatty acids
and vitamin E co-supplementation on clinical and metabolic status
in patients with Parkinson's disease: A randomized, double-blind,
placebo-controlled trial. Neurochem Int. 108:183–189.
2017.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Kharaeva Z, Gostova E, De Luca C, Raskovic
D and Korkina L: Clinical and biochemical effects of coenzyme
Q(10), vitamin E, and selenium supplementation to psoriasis
patients. Nutrition. 25:295–302. 2009.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Odinak MM, Bisaga GN and Zarubina IV: New
approaches to antioxidant therapy in multiple sclerosis. Zh Nevrol
Psikhiatr Im S S Korsakova. (Suppl):S72–S75. 2002.PubMed/NCBI(In Russian).
|
|
104
|
Miroliaee AE, Esmaily H, Vaziri-Bami A,
Baeeri M, Shahverdi AR and Abdollahi M: Amelioration of
experimental colitis by a novel nanoselenium-silymarin mixture.
Toxicol Mech Methods. 21:200–208. 2011.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Peretz A, Siderova V and Nève J: Selenium
supplementation in rheumatoid arthritis investigated in a double
blind, placebo-controlled trial. Scand J Rheumatol. 30:208–212.
2001.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Tarp U, Overvad K, Thorling EB, Graudal H
and Hansen JC: Selenium treatment in rheumatoid arthritis. Scand J
Rheumatol. 14:364–368. 1985.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Zafar KS, Siddiqui A, Sayeed I, Ahmad M,
Salim S and Islam F: Dose-dependent protective effect of selenium
in rat model of Parkinson's disease: Neurobehavioral and
neurochemical evidences. J Neurochem. 84:438–446. 2003.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Batooei M, Tahamoli-Roudsari A, Basiri Z,
Yasrebifar F, Shahdoust M, Eshraghi A, Mehrpooya M and Ataei S:
Evaluating the effect of oral N-acetylcysteine as an adjuvant
treatment on clinical outcomes of patients with rheumatoid
arthritis: A randomized, double blind clinical trial. Rev Recent
Clin Trials. 13:132–138. 2018.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Lai ZW, Hanczko R, Bonilla E, Caza TN,
Clair B, Bartos A, Miklossy G, Jimah J, Doherty E, Tily H, et al:
N-acetylcysteine reduces disease activity by blocking mammalian
target of rapamycin in T cells from systemic lupus erythematosus
patients: A randomized, double-blind, placebo-controlled trial.
Arthritis Rheum. 64:2937–2946. 2012.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Zheng K, Dong Y, Yang R, Liang Y, Wu H and
He Z: Regulation of ferroptosis by bioactive phytochemicals:
Implications for medical nutritional therapy. Pharmacol Res.
168(105580)2021.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Khojah HM, Ahmed S, Abdel-Rahman MS and
Elhakeim EH: Resveratrol as an effective adjuvant therapy in the
management of rheumatoid arthritis: A clinical study. Clin
Rheumatol. 37:2035–2042. 2018.PubMed/NCBI View Article : Google Scholar
|
|
112
|
Samsami-Kor M, Daryani NE, Asl PR and
Hekmatdoost A: Anti-inflammatory effects of resveratrol in patients
with ulcerative colitis: A randomized, double-blind,
placebo-controlled pilot study. Arch Med Res. 46:280–285.
2015.PubMed/NCBI View Article : Google Scholar
|
|
113
|
Samsamikor M, Daryani NE, Asl PR and
Hekmatdoost A: Resveratrol supplementation and
oxidative/anti-oxidative status in patients with ulcerative
colitis: A randomized, double-blind, placebo-controlled pilot
study. Arch Med Res. 47:304–309. 2016.PubMed/NCBI View Article : Google Scholar
|
|
114
|
Movahed A, Raj P, Nabipour I, Mahmoodi M,
Ostovar A, Kalantarhormozi M and Netticadan T: Efficacy and safety
of resveratrol in type 1 diabetes patients: A two-month preliminary
exploratory trial. Nutrients. 12(161)2020.PubMed/NCBI View Article : Google Scholar
|
|
115
|
Ghavipour M, Sotoudeh G, Tavakoli E, Mowla
K, Hasanzadeh J and Mazloom Z: Pomegranate extract alleviates
disease activity and some blood biomarkers of inflammation and
oxidative stress in rheumatoid arthritis patients. Eur J Clin Nutr.
71:92–96. 2017.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Petramfar P, Hajari F, Yousefi G, Azadi S
and Hamedi A: Efficacy of oral administration of licorice as an
adjunct therapy on improving the symptoms of patients with
Parkinson's disease, A randomized double blinded clinical trial. J
Ethnopharmacol. 247(112226)2020.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Lin D, Zhang M, Luo C, Wei P, Cui K and
Chen Z: Targeting ferroptosis attenuates inflammation, fibrosis,
and mast cell activation in chronic prostatitis. J Immunol Res.
2022(6833867)2022.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Zhao T, Yang Q, Xi Y, Xie Z, Shen J, Li Z,
Li Z and Qin D: Ferroptosis in rheumatoid arthritis: A potential
therapeutic strategy. Front Immunol. 13(779585)2022.PubMed/NCBI View Article : Google Scholar
|