Introduction
Enterovirus 71 (EV71) belongs to the small
ribonucleovirus family of the enterovirus genus, and is a
non-envelope single-stranded RNA virus (1). Patients who are <6 years old with
EV71 infection may suffer from brain stem encephalitis, neurogenic
pulmonary edema, cardiac arrest and other fatal clinical symptoms,
including death (2,3).
Previous studies have demonstrated that viral
infections, such as severe acute respiratory syndrome coronavirus 2
(SARS-CoV-2) (4), influenza virus
(5), hepatitis B virus (6) and West Nile virus (7), can induce pyroptosis, and our
previous study has also indicated that EV71 infection could induce
pyroptosis (8). Pyroptosis is a
type of inflammatory programmed cell death. NLR family, pyrin
domain-containing 3 (NLRP3) activation triggers NLRP3-inflammasome
assembly, which results in pyroptosis initiation when viral
infections occur. Caspase-1 is a downstream molecule of the
inflammasome and activated caspase-1 initiates pyroptosis by
cleaving gasdermin D (GSDMD). Activated caspase-1 also cleaves
Pro-IL-1β into mature IL-1β. When pyroptosis occurs, the cleaved
GSDMD forms a pore channel on the cell surface, which causes
membrane rupture, cell death and release of the inflammatory
cytokines IL-1β and IL-18. Therefore, IL-1β, which can induce the
inflammatory reaction, is a sensitive marker of the activation of
GSDMD (9-14).
It has been demonstrated that excessive inflammatory
activation is one of the main causes of severe hand, foot and mouth
disease (HFMD) (2,3). We hypothesized that the suppression
of virus-induced cytotoxicity and pyroptosis with drugs can be a
novel therapeutic strategy for EV71 infection. At present, certain
researchers have tried to explore potential inhibitors to reduce
the formation of GSDMD pore channels; for example, combination of
dimethyl fumarate and cysteine residues limits the oligomerization
of GSDMD in cells (15). In
addition, the inhibition of diabetes-induced pyroptosis by
inhibiting the thioredoxin interacting protein (TXNIP)/NLRP3
signaling pathway has been observed (16). In another study, paeoniflorin has
been used to inhibit the activation of caspase-11, thereby aiming
to inhibit the activation of the inflammatory caspase cascade to
suppress inflammation (17).
Metformin was originally extracted from goat beans
to treat diabetes (18). It has
been demonstrated that metformin reduces liver glucose generation
via the AMP-activated protein kinase (AMPK) signaling pathway
(19). A previous study has
revealed that metformin inhibits the production of IL-1β by
macrophages (20), and that it
interferes with the AMPK signaling pathway to suppress inflammation
(21). Metformin has also been
revealed to inhibit NLRP3(22).
Another study has demonstrated that metformin inhibits cardiac
muscle cell pyroptosis by inhibiting the TXNIP-NLRP3-GSDMD
signaling pathway (23).
A recent study has indicated that metformin
treatment can lower the mortality rate of diabetic patients
infected with SARS-CoV-2(24). At
present, to the best of our knowledge, there is no report on the
effect of metformin treatment against EV71 infection or the effect
of metformin on EV71-induced pyroptosis.
Previous studies suggest that metformin can increase
DEP domain-containing mTOR-interacting protein (DEPTOR) levels in
hepatoma cells, ureteral epithelial cells and rats with unilateral
ureteral obstruction (25,26). DEPTOR is a component of the mTOR
complex (mTORC)1 and mTORC2 and binds to mTOR to suppress its
kinase activity (27). A study has
demonstrated that DEPTOR serves an important role in immune
regulation and that DEPTOR is higher in T cells before inflammatory
activation (28). Zhai et
al (29) revealed that DEPTOR
deficiency in T cells is associated with NLRP3-activated IL-1β
release. Another study has revealed that decreased DEPTOR levels
induce the release of a large number of chemokines in endothelial
cells, such as chemokine (C-X-Cmotif) ligand 9 (CXCL9), CXCL10,
CXCL11, CX3CL1, chemokine (C-C motif) ligand 5 (CCL5) and
CCL20(30). It has been revealed
that DEPTOR is also regulated by the nuclear receptors of steroids,
which can increase the levels of DEPTOR mRNA in cells (31). An androgen receptor repressor,
prochloramide, is used in clinical treatment to control SARS-CoV-2
infection (32). It has been
demonstrated that DEPTOR is regulated by the androgen receptor,
which serves a negative role in the regulation of DEPTOR mRNA
levels (33). Therefore the
present study investigated whether EV71 could induce a change in
the DEPTOR protein levels and what consequences this had on
EV71-induced pyroptosis. In addition, in terms of inhibiting
EV71-induced pyroptosis, the present study aimed to find a
commercial drug that may be used in the treatment of EV71 infection
and investigate its anti-pyroptosis mechanism.
Materials and methods
Reagents and antibodies
DMEM (cat. no. PYG0073), penicillin and streptomycin
(cat. no. PYG0016), trypsin, BCA protein detection kit (cat. no.
AR0146), ECL western blot detection kit (cat. no. AR1172), SDS-PAGE
loading buffer (5X) (cat. no. AR1112-10), horseradish
peroxidase-conjugated goat anti-rabbit IgG (H+L) (cat. no. BA1054)
and GSDMD antibody (cat. no. A02842) were purchased from Boster
Biological Technology. Cell Counting Kit (CCK-8) (cat. no.
40203ES76) was purchased from Yeasen Biotechnology (Shanghai) Co.,
Ltd.. Bovine serum albumin (cat. no. abs9157) was purchased from
Absin Bioscience Inc., Fetal bovine serum (cat. no. 04-001-1A) were
purchased from Biological Industries, metformin (cat. no. IM0140)
were obtained from Beijing Solarbio Science & Technology Co.,
Ltd. DAPI (cat. no. C1002), Lipofectamine 8000 (cat. no.
C0533-0.5ml), propidium iodide (PI; cat. no. ST511) and Alexa Fluor
488 goat anti-rabbit IgG antibody (cat. no. A0423) were purchased
from Beyotime Biotechnology Company. The caspase-1 antibody (cat.
no. A0964) and caspase 1 p20 (cat. no. A23429) were purchased from
ABclonal Biotech Co., Ltd. The lactate dehydrogenase (LDH)
detection kit (cat. no. WLA072a) and Pro-IL-1β (cat. no. WL02257)
and NLRP3 (cat. no. WL02635) antibodies were purchased from
Wanleibio Co., Ltd. The GAPDH (cat. no. BS65656) antibody was
obtained from Bioworld Technology, Inc. The β-actin (cat. no.
AB0061) antibody was purchased from Shanghai Abways Biotechnology
Co., Ltd. The DEPTOR/DEPDC6 (D9F5) antibody (cat. no.
#11816) was purchased from Cell Signaling Technology,
Inc. pcDNA 3.1 and pcDNA 3.1-DEPTOR were purchased from Shanghai
GenePharma Co., Ltd.
Cells and virus
GES-1 human normal gastric epithelial cells were
purchased from Boster Biological Technology and RD human
rhabdomyosarcoma cells were purchased from the National Collection
of Authenticated Cell Cutures, and cultured in DMEM containing 10%
fetal bovine serum and 1% penicillin and streptomycin in an
incubator with 5% CO2 at 37˚C. EV71 was obtained from
Professor Zhendong Zhao (Institute of Pathogen Biology, Chinese
Academy of Medical Sciences & Peking Union Medical College).
GES-1 cells were infected with EV71 at various multiplicity of
infections (MOIs) of 1, 2, 3, 4 or 5 for different times (12, 18 or
24 h) at 37˚C, and cells cultured in full culture medium without
EV71 were used as the mock group (mock infection).
Cell culture infectious dose 50 (CCID
50) assay
RD cells (1x104/well) were seeded into
96-well plates and cultured in an incubator at 37˚C with 5%
CO2 for 12 h. Subsequently, RD cells were infected with
EV71 that was serially diluted from 1x101 to
1x1011 times, and cells in a total of eight wells per
96-well plate were infected with EV71 at the same dilution and
cultured at 37˚C. The 96-well plates were observed once per day for
5 days. The wells with ~50% cytopathic effect of RD cells were
observed under a microscope and recorded, and the CCID 50 results
were calculated using the Reed-Münch method (34).
Measurement of LDH release
GES-1 cells were infected with EV71 at various MOIs
of 1, 2, 3, 4 or 5 or infected with EV71 at MOI 5 and treated with
metformin (1, 3 or 5 µM) for 24 h and then the cell supernatant was
collected at 400 x g for 5 min at 4˚C. The LDH release of the cell
supernatant was measured using an LDH detection kit. LDH activity
(%) was calculated as follows:
(LDHtreatment-LDHcontrol)/(LDHmax-LDHcontrol)
x100.
Western blotting
GES-1 cells from different groups were collected in
1.5-ml centrifuge tubes and lysed with 60 µl RIPA lysis buffer
(Beyotime Institute of Biotechnology). The total protein
concentration was measured using a BCA kit. A total of 30 µg total
extracted protein from each group was separated by 12% SDS-PAGE and
then transferred to a nitrocellulose (NC) membrane. The NC membrane
was blocked with 5% skimmed milk for 2 h at room temperature and
then incubated with NLRP3 (1:1,000), GSDMD (1:1,000), caspase-1
(1:1,000), caspase-1 P20 (1:1,000), Pro-IL-1β (1:5,000), DEPTOR
(1:1,000), GAPDH (1:5,000) and β-actin (1:5,000) antibodies
overnight at 4˚C. Subsequently, the NC membrane was washed three
times with TBST (0.1% Tween) and incubated with horseradish
peroxidase-conjugated goat anti-rabbit IgG secondary antibody
(1:5,000) at room temperature for 2 h. Finally, immunoblots were
incubated with an ECL chromogenic kit and observed using a
ChemiDoc™ MP Imaging System (Bio-Rad Laboratories, Inc.). ImageJ
software (version 6.0; National Institutes of Health) was used for
densitometry.
Cell Counting Kit-8 (CCK-8) assay
GES-1 cells seeded onto 96-well plates were infected
with EV71 at MOI 1, 2, 3, 4, or 5 or infected with EV71 at MOI 5
and treated with metformin (1, 3 or 5 µM) for 24 h at 37˚C with 5%
CO2. At 2 h before the end of the culture, 20 µl CCK-8
reagent was added to the 96-well plates. Subsequently, the
absorption of each sample was detected at 450 nm using a microplate
reader (BioTek Instruments, Inc.). The cell viability was
calculated using the following formula: Cell viability
(%)=(Absorbancetreatment-Absorbanceblank)/(Absorbancecontrol-Absorbanceblank)
x100%.
Immunofluorescence staining
GES-1 cells were seeded into 24-well plates
(1x105 cells/well). When the EV71 infected-GES-1 cell
density reached 60-70%, GES-1 cells were fixed with 4%
paraformaldehyde at 4˚C for 20 min. Subsequently, the cells were
permeated with 0.5% Triton X-100 at room temperature for 15 min and
blocked with 5% bovine serum albumin at room temperature for 1 h.
Then the cells were incubated with the NLRP3 antibody (1:200) at
4˚C overnight, followed by incubation with Alexa Fluor 488 goat
anti-rabbit IgG (1:200) at room temperature for 1 h, and then
protected from light and stained with 1 µg/ml DAPI solution in PBS
for 5 min at room temperature. The cells were again protected from
light and washed three times with PBS after each staining step. All
images were observed under an inverted fluorescence microscope
(x200 magnification2; Nikon Corporation).
PI staining
GES-1 cells in the logarithmic phase were seeded
into 24-well plates (1.2x105/well) for 12 h and then
treated with EV71, metformin (5 µM) or EV71 and metformin (5 µM)
for 24 h at 37˚C. When the density of the treated cells reached
~80%, GES-1 cells were stained with 4 µM PI for 30 min at 37˚C. The
images were captured using an inverted fluorescence microscope
(magnification, x200; Nikon Corporation).
DNA transfection
GES-1 cells were plated into 6-well plates
(50x104/well) for 12 h. Subsequently, 2.5 µg pcDNA 3.1
(negative control with empty vector) or pcDNA 3.1-DEPTOR and 5 µl
Lipofectamine 8000 were added to 125 µl fresh DMEM (without bovine
serum and antibiotics) and gently mixed. Then, the mixture was
added to the 6-well plate. Cells were cultured at 37˚C, and
subsequent experiments were carried out 48 h later.
Statistical analysis
GraphPad Prism 8 software (GraphPad Software;
Dotmatics) was used for statistical tests. All experiments were
independently repeated three times and data are presented as the
mean ± SD. Multiple groups were analyzed through one-way ANOVA
followed by Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
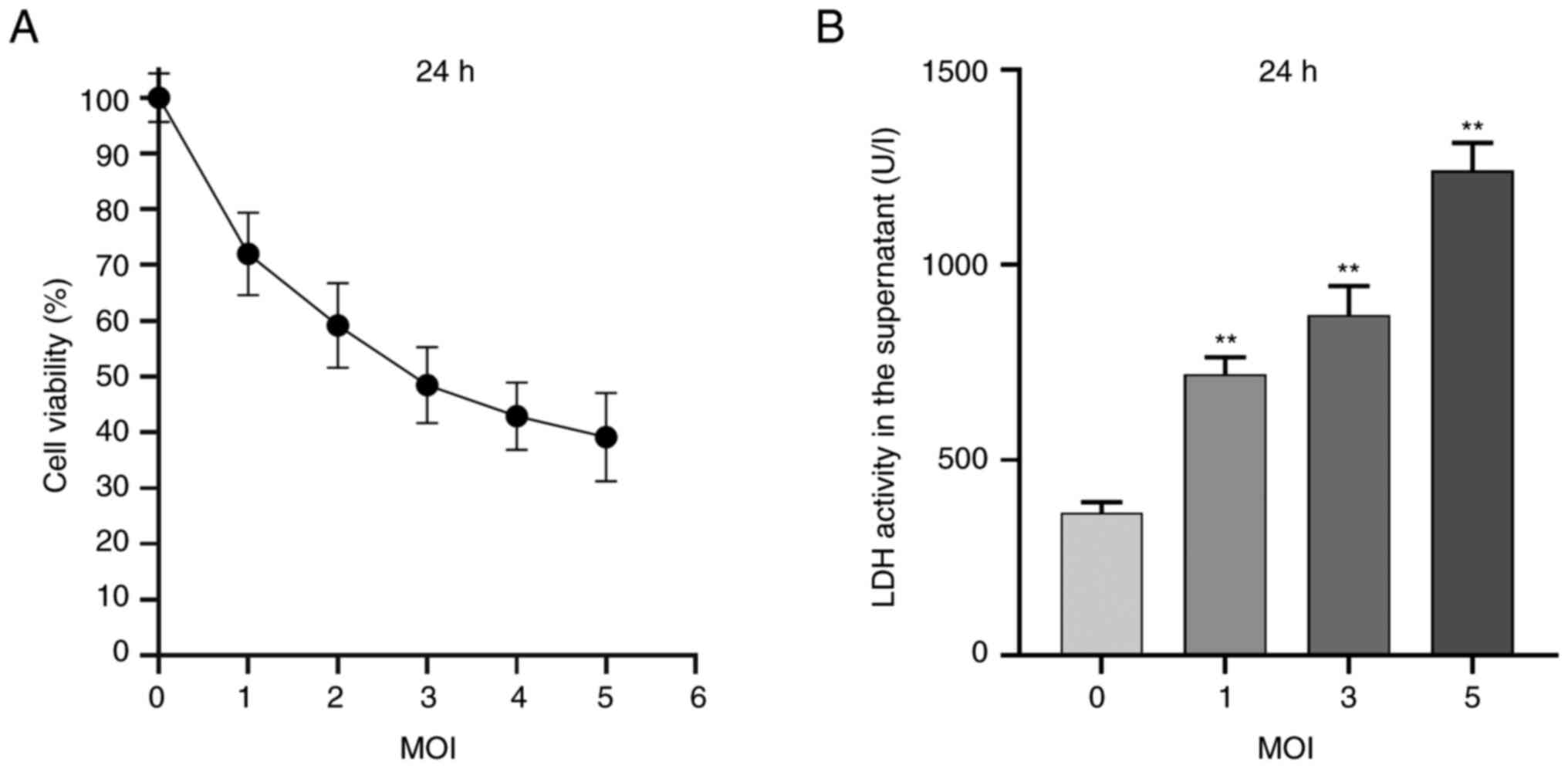
EV71 infection causes cell damage and
reduces cell viability
GES-1 cells were treated with EV71 at the different
MOIs (0, 1, 2, 3, 4 and 5) for 24 h. The cell viability of each
group was detected using a CCK-8 assay (Fig. 1A). Subsequently, GES-1 cells were
treated with EV71 at a MOI of 1, 3 and 5 for 24 h, and the cell
damage level was measured using an LDH release assay (Fig. 1B). The results demonstrated that
EV71 decreased cell viability and increased cell damage in a
dose-dependent manner.
EV71 infection induces pyroptosis
The present study further examined whether
pyroptosis is involved in EV71-induced cell damage. GES-1 cells
were infected with EV71 at different MOIs (0, 1, 2, 3, 4 and 5) for
24 h. The protein levels of NLRP3, GSDMD, caspase-1 and Pro-IL-1β
in cells were detected by western blotting. The results
demonstrated that EV71 increased the levels of NLRP3, and decreased
the levels of GSDMD, caspase-1 and Pro-IL-1β in a dose-dependent
manner (Fig. 2A-C). Subsequently,
GES-1 cells were infected with EV71 at a MOI of 5 for 12, 18 and 24
h. Compared to those of the mock group, the protein levels of
GSDMD, and Pro-IL-1β were reduced in a time-independent manner, and
caspase-1 was decreased significantly at 24 h (Fig. 2D and E). GES-1 cells were infected with EV71 at
a MOI of 5 for 24 h, and the NLRP3 levels were examined using
immunofluorescence staining. The results indicated that
intracellular NLRP3 was upregulated and showed intracellular
aggregation (Fig. 2F). These
suggested that EV71 infection could induce cell pyroptosis.
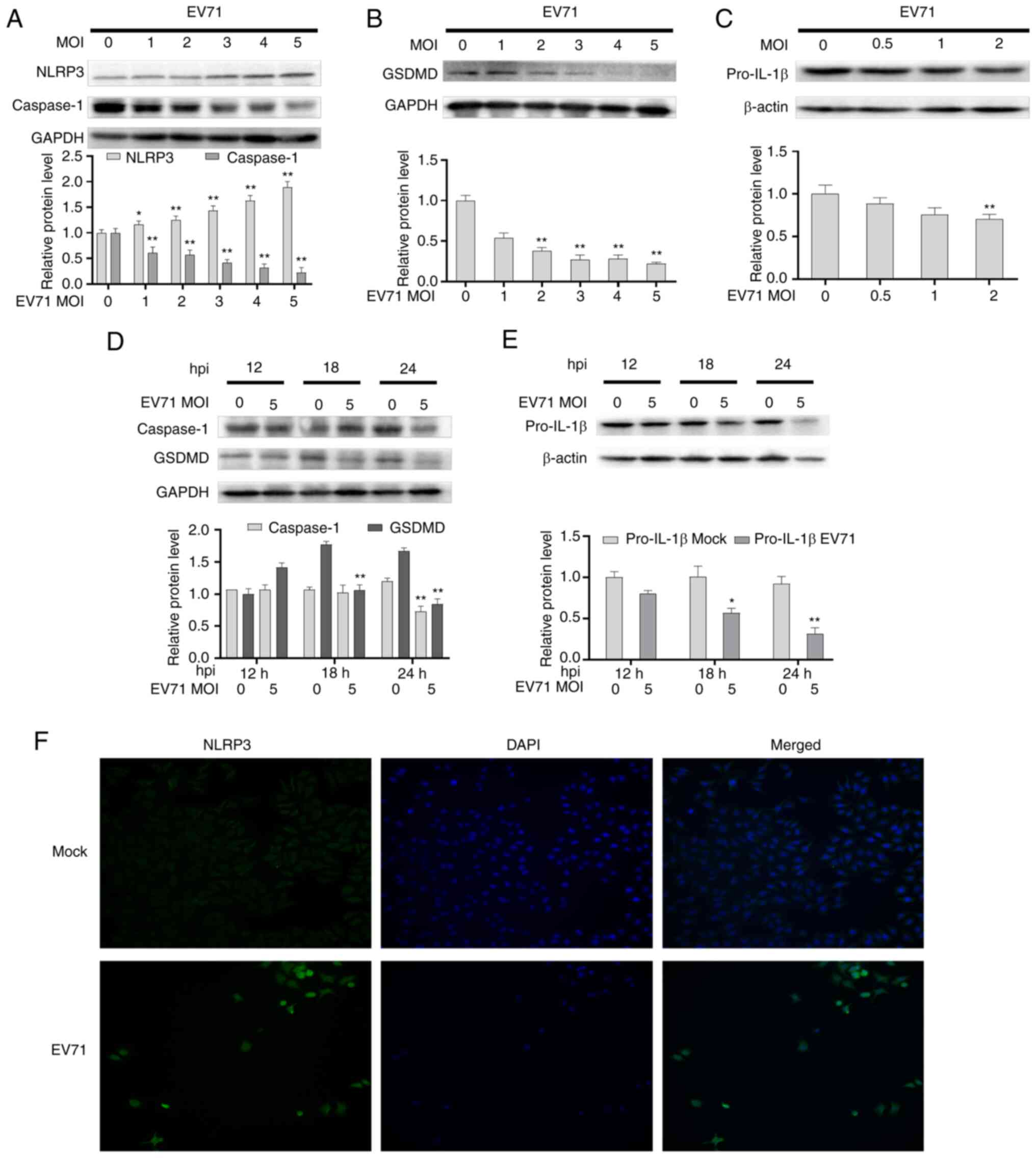 | Figure 2EV71 induces cell pyroptosis. Western
blotting was performed to detect the levels of (A) NLRP3 and
caspase-1, (B) GSDMD and (C) Pro-IL-1β, with β-actin or GAPDH as an
internal reference. Western blot analysis of the levels of (D)
caspase-1 and GSDMD and (E) Pro-IL-1β, with β-actin or GAPDH as
internal references. (F) NLRP3 staining in GES-1 cells was detected
using an immunofluorescence assay (magnification, x200).
*P<0.05 and **P<0.01 compared with the
mock group. EV71, enterovirus 71; GSDMD, gasdermin D; NLRP3, NLR
family, pyrin domain-containing 3; MOI, multiplicity of infection;
hpi, hours post inoculation. |
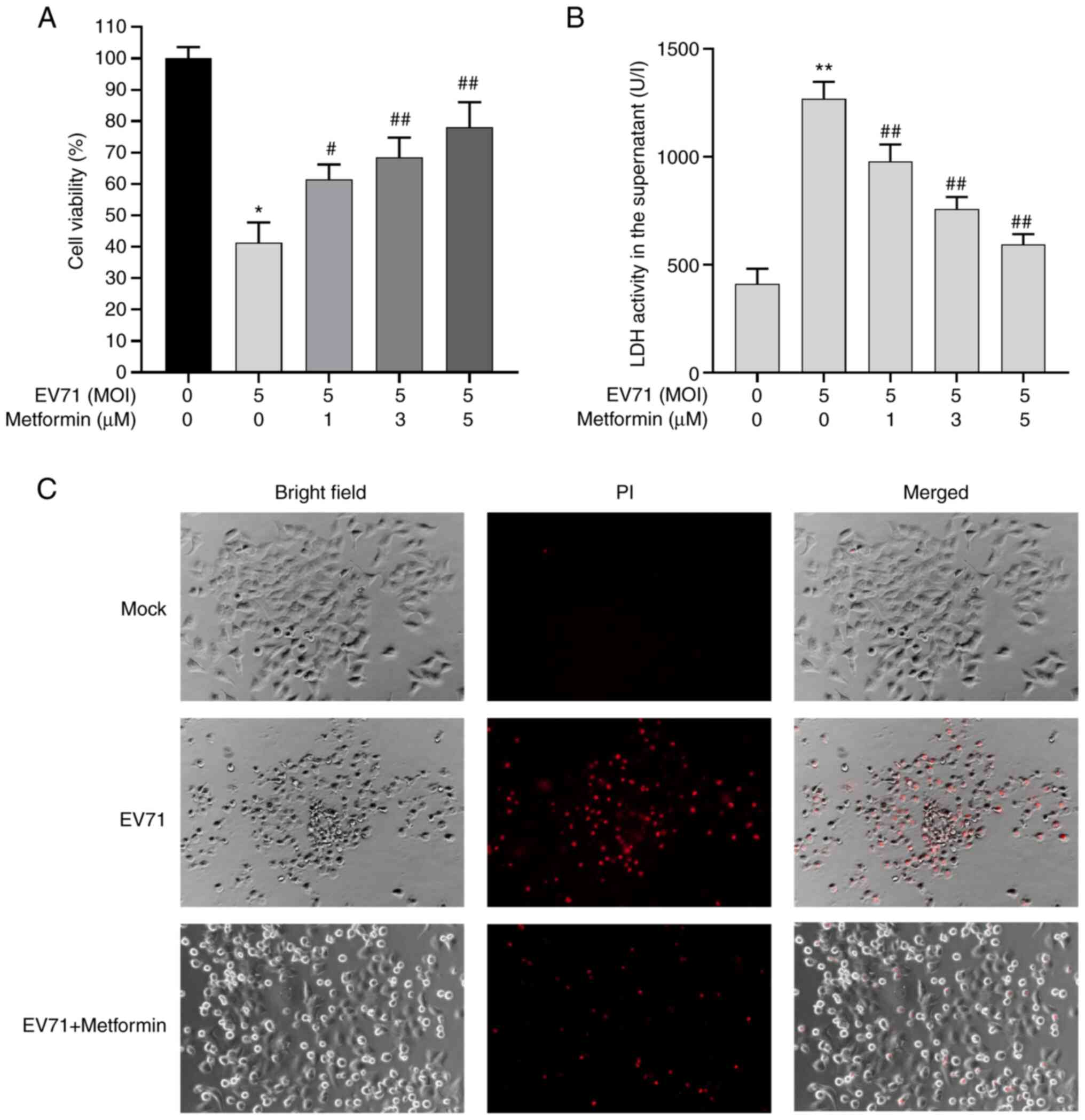
Metformin reverses EV71-induced
cytotoxicity and cell viability inhibition
The present study evaluated whether metformin was
able to affect the viability of EV71-infected cells and reverse
EV71-induced cytotoxicity. Shifts in cell viability and cell damage
were detected using CCK-8 and LDH release assays. The cells were
treated with EV71 (MOI=5) alone or combined with metformin at
different concentrations (1, 3 and 5 µM) for 24 h. The cell
viability was increased and cell damage was inhibited by 5 µM
metformin treatment in EV71-infected cells compared with cells
infected with EV71 only (Fig. 3A
and B). Next, The GES-1 cells were
stained with PI (red fluorescence), which was used to stain the
dead and dying cells. EV71-treated cells exhibited cell injury and
the number of GES-1 cells were decreased compared with that in the
uninfected group, while treatment with metformin reversed the
EV71-induced cell damage (Fig.
3C).
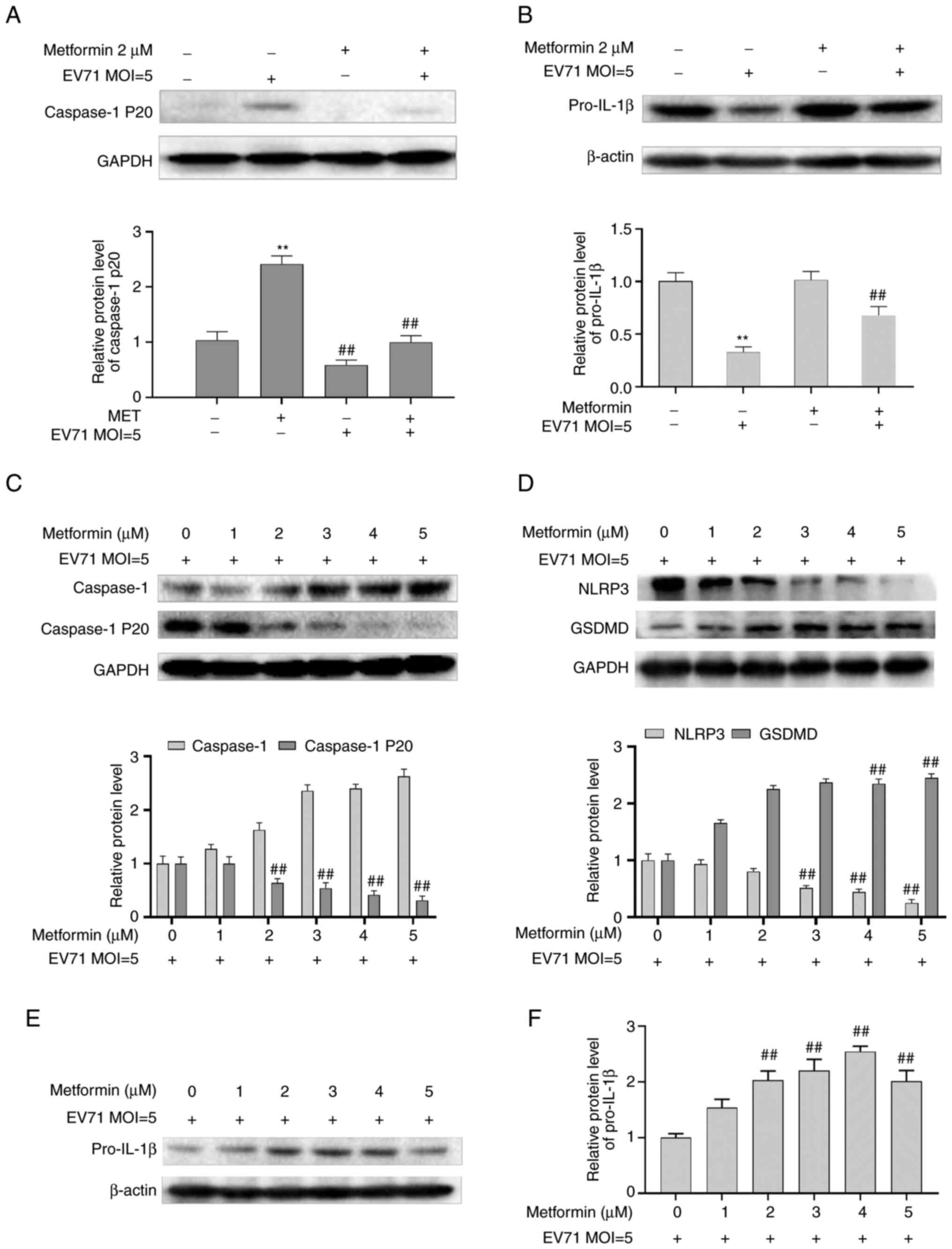
Metformin inhibits EV71-induced
pyroptosis in GES-1 cells
In order to define whether metformin against EV71
induced cell pyroptosis, GES-1 cells were treated with EV71 and 2
µM metformin. Metformin upregulated Pro-IL-1β and downregulated
caspase-1 P20 in EV71-infected cells compared with the EV71
infected group (P<0.01; Fig. 4A
and B). EV71-infected GES-1 cells
were treated with different concentrations (0, 1, 2, 3, 4 and 5 µM)
of metformin for 24 h. The results revealed that NLRP3 and
caspase-1 P20 were downregulated, and GSDMD, caspase-1 and
Pro-IL-1β were upregulated (P<0.01; Fig. 4C-F) compared with the EV71 group.
This indicated that metformin exerted substantial inhibitory
effects on EV71-induced pyroptosis.
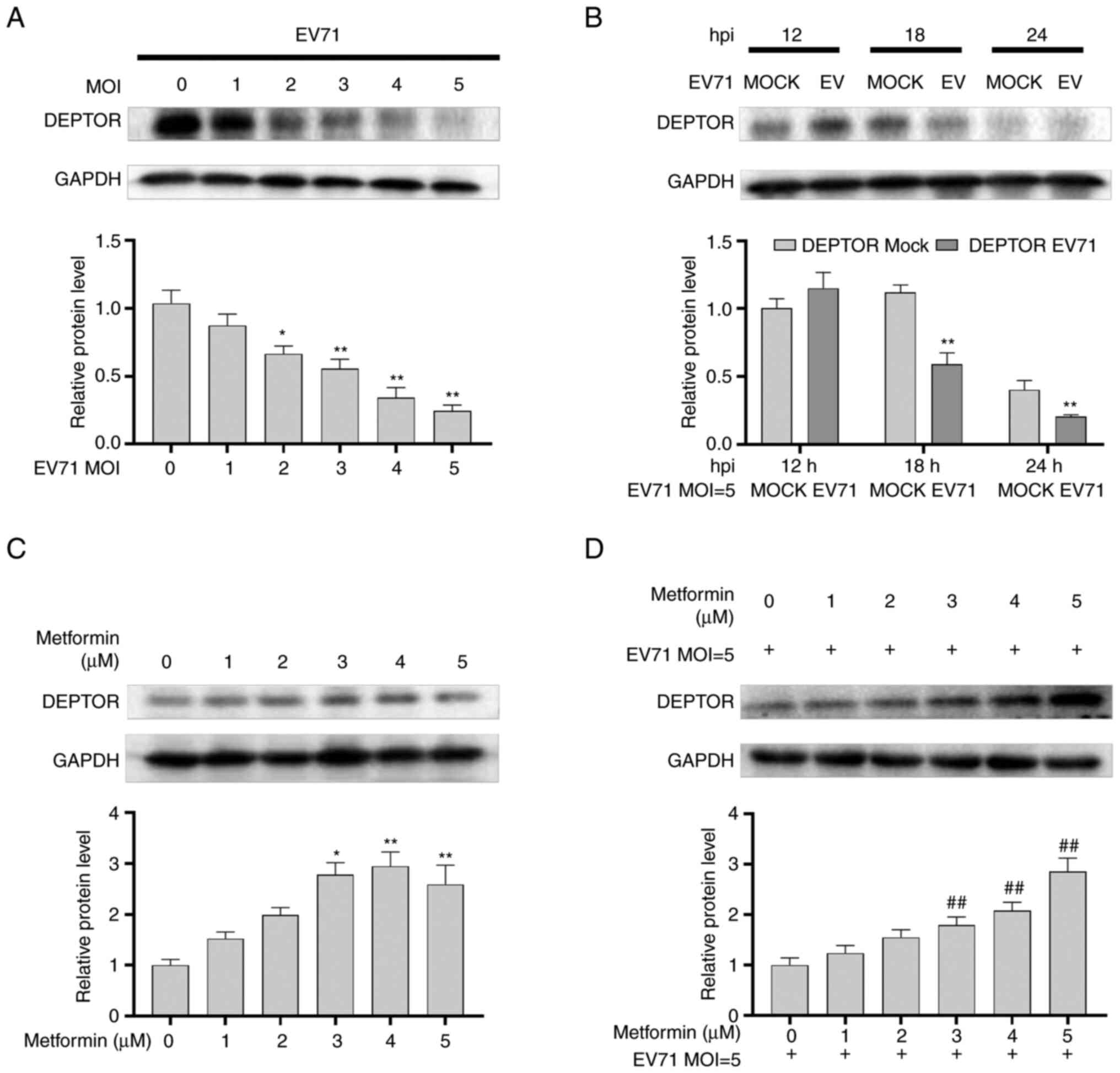
Metformin upregulates DEPTOR protein
expression in GES-1 cells
GES-1 cells were infected with EV71 at various MOIs
(0, 1, 2, 3, 4 and 5) for 24 h, and the results indicated that the
protein levels of DEPTOR were decreased in a dose-dependent manner
in EV71-infected GES-1 cells (P<0.01; Fig. 5A). Subsequently, GES-1 cells were
infected with EV71 (MOI=5) at varying times (12, 18 and 24 h), and
western blot analysis revealed the degradation of DEPTOR in a
time-dependent manner compared with the mock group at the same time
(P<0.01; Fig. 5B). Next, GES-1
cells were incubated with varying concentrations (0, 1, 2, 3, 4 and
5 µM) of metformin for 24 h, and the results indicated that
metformin increased DEPTOR levels in a dose-dependent manner
(P<0.01; Fig. 5C). Finally,
DEPTOR protein levels were upregulated in EV71-infected and
metformin-treated (0, 1, 2, 3, 4 and 5 µM) GES-1 cells compared
with EV71-infected alone GES-1 cells (P<0.01; Fig. 5D).
The results demonstrated that metformin could
reverse the downregulating effect of EV71 on DEPTOR. The
alterations of DEPTOR were accompanied by changes in pyroptosis
that were regulated by EV71 or metformin, as aforementioned. These
findings suggested a potential association between pyroptosis and
DEPTOR expression.
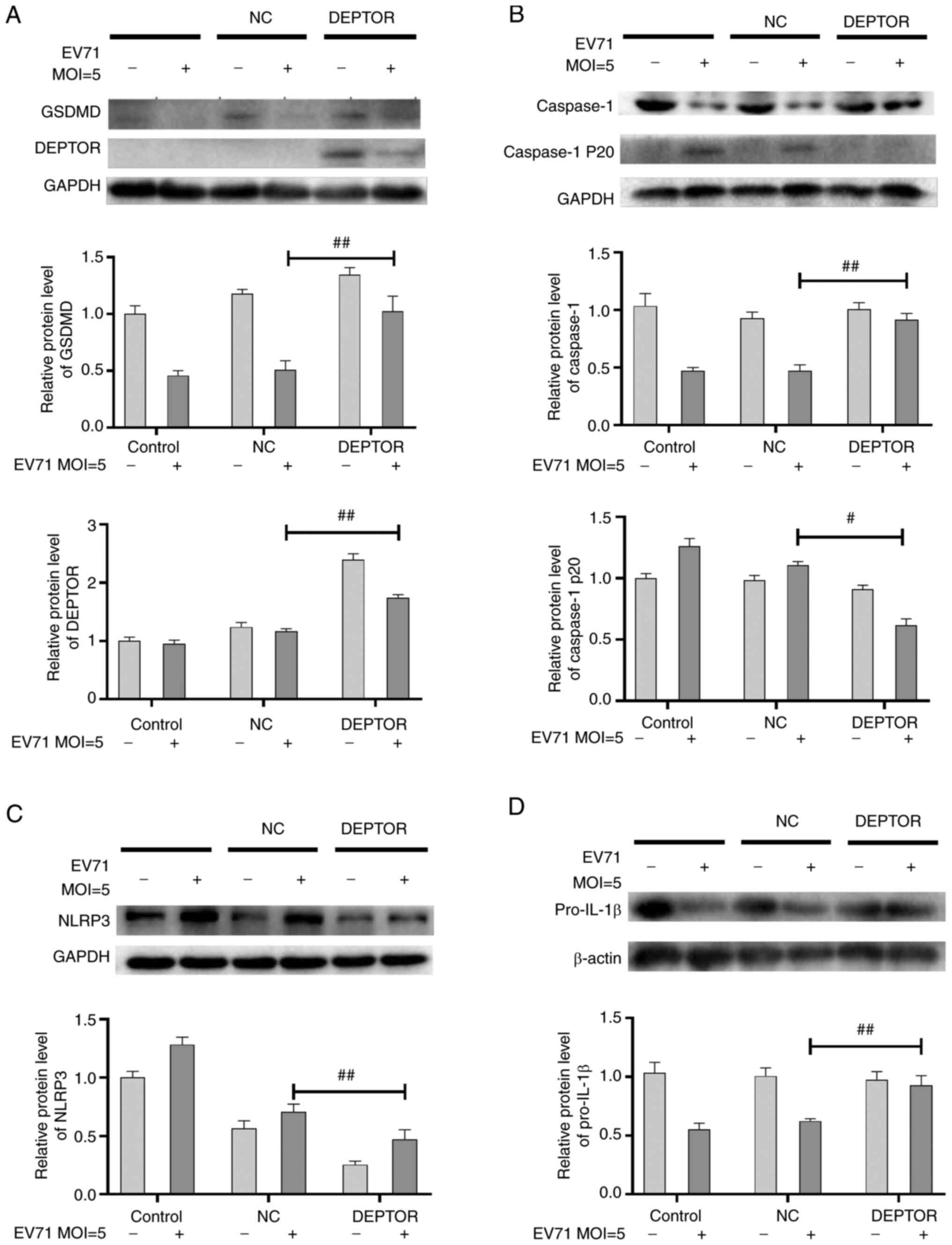
Overexpression of DEPTOR inhibits
EV71-induced pyroptosis
EV71 infection induced pyroptosis and the
degradation of DEPTOR, which were reversed by metformin treatment.
Next, the present study aimed to further investigate the
association between pyroptosis and DEPTOR. pcDNA 3.1 and pcDNA
3.1-DEPTOR were transfected into GES-1 cells, which were then
infected with EV71. The changes of pyroptosis-related proteins were
observed. The results showed that after EV71 infection, compared
with that of the empty vector expression group, the level of GSDMD
and DEPTOR was increased (P<0.01; Fig. 6A), caspase-1 level was increased
(P<0.01; Fig. 6B), caspase-1
P20 level was decreased (P<0.05; Fig. 6B), the expression of NLRP3 was
reduced (P<0.01; Fig. 6C) and
the level of Pro-IL-1β was increased (P<0.01; Fig. 6D) in the DEPTOR overexpression
group. The results demonstrated that DEPTOR overexpression led to
inhibition of pyroptosis and indicated that metformin inhibited
EV71-induced pyroptosis via upregulating DEPTOR.
Discussion
EV71 infection causes HFMD in infants and children,
particularly severe infections with pulmonary edema and
neurological complications, which pose a serious threat to the life
and health of children. There is no specific drug to control EV71
infection at present, thus it is important to investigate
anti-infection mechanisms and find effective antiviral drugs
(35). Viral infection induces
pyroptosis and causes an inflammation response. We hypothesized
that targeting the virus-induced pyroptosis and its initiated
inflammation can be an innovative strategy for antiviral treatment.
In the present study, the inhibitory effect of metformin on
EV71-induced pyroptosis and the upregulating effect of metformin on
DEPTOR levels were revealed. It was demonstrated that DEPTOR was
involved in the inhibitory process of EV71-induced pyroptosis,
which provided a reference for discovering effective anti-EV71
infection drugs.
The present study first confirmed that EV71
infection increased cell damage and decreased cell viability, and
that 5 µM metformin treatment reversed this cytotoxicity. This
finding suggests that metformin may play a protective role in EV71
infections. EV71 infected GES-1 cells exhibited pyroptosis,
accompanied by caspase-1 activation, GSDMD increase and IL-1β
maturation. This finding is consistent with the work of Yogarajah
et al (36) showing that
EV71 induced host cell pyroptosis. Other studies have demonstrated
that EV71 3C protease inhibits the activation of GSDMD (36-38).
It is possible that the viral protease prolongs the proliferation
stage of the virus in cells by inhibiting pyroptosis in the early
stage of infection (39). However,
a large number of aggregating progeny viruses eventually lead to
the occurrence of pyroptosis in the later stage of infection, which
causes the release of mature viruses and inflammatory cytokines,
increases infection and pathogenesis, and causes further serious
pathological damage to the body. This hypothesis is supported by
the marked intracellular elevation of viral structure protein VP1
and the pyroptosis-related proteins NLRP3, GSDMD, Caspase-1 and Pro
IL-1β 24 h after infection (40).
The present study subsequently tried to find a marketed drug that
could suppress pyroptosis to shorten its clinical trial period. It
was revealed that metformin could inhibit EV71-induced cell damage
and pyroptosis in a dose-dependent manner. This finding
demonstrated the anti-pyroptosis effect of metformin in a novel
model, which further supplemented the pharmacological effects of
metformin.
The anti-pyroptosis mechanism of metformin was
investigated further, and it was revealed that metformin could
regulate DEPTOR. DEPTOR, as a metabolic pathway inhibitor protein,
is upregulated by corticosteroids (31). It was explored whether the levels
of DEPTOR changed in the EV71-infection model and whether metformin
could regulate DEPTOR. The results demonstrated that EV71 infection
reduced, while metformin increased the cellular levels of DEPTOR
which is consistent with the findings of Obara et al
(25). In the present cell model
of EV71 infection, metformin reversed EV71-induced DEPTOR reduction
in a dose-dependent manner, suggesting that metformin may exert its
effect by upregulating the expression levels of DEPTOR.
The present study subsequently aimed to determine
whether DEPTOR is involved in the regulation of virus-induced
pyroptosis. The effects of DEPTOR overexpression on NLRP3,
Pro-IL-1β, caspase-1, caspase-1 P20 and GSDMD were detected. The
results demonstrated that DEPTOR overexpression could suppress
EV71-induced pyroptosis. Zhai et al (29) indicated that the reduced level of
DEPTOR can activate NLRP3 and mature IL-1β in macrophages. DEPTOR
knockdown experiments were conducted in metformin-treated cells in
the present study with an aim to observe whether the effect of
metformin on pyroptosis can be inhibited. However, knockdown
experiments require prolonged cell cultivation that make the
expression level of DEPTOR in cells too low to detect. This is a
limitation of the present study and an aim in future research to
improve upon. The present study demonstrated that metformin could
inhibit EV71-induced pyroptosis by upregulating DEPTOR. However,
the mechanism via which metformin upregulates DEPTOR and how DEPTOR
regulates pyroptosis should be investigated in the future.
In future studies on this topic, the first
consideration should be to further investigate the association
between DEPTOR and the pyroptosis pathway. Peterson et al
(41) revealed that overexpression
of DEPTOR activates the mTORC2/AKT signaling pathway, leading to an
increase in phosphorylation at the Ser-473 site of AKT. Zhao et
al (42) revealed that
activation of AKT inhibits NLRP3 degradation, thereby suppressing
NLRP3 oligomerization and activation. Considering the relationship
between AKT and mTORC2, it may be hypothesized that DEPTOR may
affect AKT activity through mTORC2 to regulate NLRP3
oligomerization and inhibit pyroptosis.
In conclusion, the findings of the present study
suggested that EV71 infection induced GES-1 cell pyroptosis, while
metformin suppressed EV71-induced pyroptosis, alleviated
cytotoxicity and the inflammatory response. The implicated
mechanism may involve the upregulation of DEPTOR by metformin.
Acknowledgements
The authors would like to thank Professor Zhendong
Zhao (Institute of Pathogen Biology, Chinese Academy of Medical
Sciences & Peking Union Medical College, Beijing, China) for
his gift of cell lines and virus.
Funding
Funding: The present study was supported by the National Natural
Science Foundation of China (grant no. 81301426), the Provincial
Natural Science Foundation of Shanxi (grant no. 201901D111329), the
Mega Research and Development Projects of Lüliang (grant no.
2020SHFZ38), the Program of Fenyang College, Shanxi Medical
University (grant no. 2020B01) and the Key Laboratory Platform
Construction Projects of Lüliang (grant no. 2020ZDSYS17).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XiaZ and CS conceived and designed the experiments.
CS, XiaZ, JH, LC and XinZ performed the experiments. CS, JD, QH
analyzed the data. XiaZ and CS wrote the manuscript. XiaZ and JH
revised the manuscript. All authors have read and approved the
final manuscript. XiaZ and CS confirm the authenticity of all the
raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Huang SW, Kiang D, Smith DJ and Wang JR:
Evolution of re-emergent virus and its impact on enterovirus 71
epidemics. Exp Biol Med (Maywood). 236:899–908. 2011.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Chen Y, Hu L, Xu F, Liu C and Li J: A case
report of a teenager with severe hand, foot, and mouth disease with
brainstem encephalitis caused by enterovirus 71. BMC Pediatr.
19(59)2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Xing J, Wang K, Wei H and Wei D:
Pathologic and molecular studies of enterovirus 71 infection in a
fatal case from a recent epidemic in China. Medicine (Baltimore).
97(e13447)2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Sun X, Liu Y, Huang Z, Xu W, Hu W, Yi L,
Liu Z, Chan H, Zeng J, Liu X, et al: SARS-CoV-2 non-structural
protein 6 triggers NLRP3-dependent pyroptosis by targeting ATP6AP1.
Cell Death Differ. 29:1240–1254. 2022.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Allen IC, Scull MA, Moore CB, Holl EK,
McElvania-TeKippe E, Taxman DJ, Guthrie EH, Pickles RJ and Ting JP:
The NLRP3 inflammasome mediates in vivo innate immunity to
influenza A virus through recognition of viral RNA. Immunity.
30:556–565. 2009.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Xie WH, Ding J, Xie XX, Yang XH, Wu XF,
Chen ZX, Guo QL, Gao WY, Wang XZ and Li D: Hepatitis B virus X
protein promotes liver cell pyroptosis under oxidative stress
through NLRP3 inflammasome activation. Inflamm Res. 69:683–696.
2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ramos HJ, Lanteri MC, Blahnik G, Negash A,
Suthar MS, Brassil MM, Sodhi K, Treuting PM, Busch MP, Norris PJ
and Gale M Jr: IL-1β signaling promotes CNS-intrinsic immune
control of West Nile virus infection. PLoS Pathog.
8(e1003039)2012.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zhang X, Hao J, Sun C, Du J, Han Q and Li
Q: Total astragalosides decrease apoptosis and pyroptosis by
inhibiting enterovirus 71 replication in gastric epithelial cells.
Exp Ther Med. 23(237)2022.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Kayagaki N, Stowe IB, Lee BL, O'Rourke K,
Anderson K, Warming S, Cuellar T, Haley B, Roose-Girma M, Phung QT,
et al: Caspase-11 cleaves gasdermin D for non-canonical
inflammasome signalling. Nature. 526:666–671. 2015.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Fink SL and Cookson BT:
Caspase-1-dependent pore formation during pyroptosis leads to
osmotic lysis of infected host macrophages. Cell Microbiol.
8:1812–1825. 2006.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Kesavardhana S, Malireddi RKS and
Kanneganti T: Caspases in cell death, inflammation, and
gasdermin-induced pyroptosis. Annu Rev Immunol. 38:567–595.
2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Shi J, Zhao Y, Wang K, Shi X, Wang Y,
Huang H, Zhuang Y, Cai T, Wang F and Shao F: Cleavage of GSDMD by
inflammatory caspases determines pyroptotic cell death. Nature.
526:660–665. 2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
He WT, Wan H, Hu L, Chen P, Wang X, Huang
Z, Yang ZH, Zhong CQ and Han J: Gasdermin D is an executor of
pyroptosis and required for interleukin-1β secretion: Gasdermin D
is an executor of pyroptosis and required for interleukin-1beta
secretion. Cell Res. 25:1285–1298. 2015.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Lamkanfi M and Dixit VM: Mechanisms and
functions of inflammasomes. Cell. 157:1013–1022. 2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Humphries F, Shmuel-Galia L,
Ketelut-Carneiro N, Li S, Wang B, Nemmara VV, Wilson R, Jiang Z,
Khalighinejad F, Muneeruddin K, et al: Succination inactivates
gasdermin D and blocks pyroptosis. Science. 369:1633–1637.
2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
An X, Zhang Y, Cao Y, Chen J, Qin H and
Yang L: Punicalagin protects diabetic nephropathy by inhibiting
pyroptosis based on TXNIP/NLRP3 pathway. Nutrients.
12(1516)2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Tian DD, Wang M, Liu A, Gao MR, Qiu C, Yu
W, Wang WJ, Zhang K, Yang L, Jia YY, et al: Antidepressant effect
of paeoniflorin is through inhibiting pyroptosis CASP-11/GSDMD
pathway. Mol Neurobiol. 58:761–776. 2021.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Bailey CJ: Metformin: Historical overview.
Diabetologia. 60:1566–1576. 2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Agius L, Ford BE and Chachra SS: The
metformin mechanism on gluconeogenesis and AMPK activation: The
metabolite perspective. Int J Mol Sci. 21(3240)2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Qing L, Fu J, Wu P, Zhou Z, Yu F and Tang
J: Metformin induces the M2 macrophage polarization to accelerate
the wound healing via regulating AMPK/mTOR/NLRP3 inflammasome
singling pathway. Am J Transl Res. 11:655–668. 2019.PubMed/NCBI
|
|
21
|
Chen X, Li X, Zhang W, He J, Xu B, Lei B,
Wang Z, Cates C, Rousselle T and Li J: Activation of AMPK inhibits
inflammatory response during hypoxia and reoxygenation through
modulating JNK-mediated NF-κB pathway. Metabolism. 83:256–270.
2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Yan J, Ding D, Feng G, Yang Y, Zhou Y, Ma
L, Guo H, Lu Z and Jin Q: Metformin reduces chondrocyte pyroptosis
in an osteoarthritis mouse model by inhibiting NLRP3 inflammasome
activation. Exp Ther Med. 23(222)2022.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Jia Y, Cui R, Wang C, Feng Y, Li Z, Tong
Y, Qu K, Liu C and Zhang J: Metformin protects against intestinal
ischemia-reperfusion injury and cell pyroptosis via
TXNIP-NLRP3-GSDMD pathway. Redox Biol. 32(101534)2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Samuel SM, Varghese E and Büsselberg D:
Therapeutic potential of metformin in COVID-19: Reasoning for its
protective role. Trends Microbiol. 29:894–907. 2021.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Obara A, Fujita Y, Abudukadier A,
Fukushima T, Oguri Y, Ogura M, Harashima S, Hosokawa M and Inagaki
N: DEPTOR-related mTOR suppression is involved in metformin's
anti-cancer action in human liver cancer cells. Biochem Biophys Res
Commun. 460:1047–1052. 2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Wang Y, Wang Y, Li Y, Lu L, Peng Y, Zhang
S and Xia A: Metformin attenuates renal interstitial fibrosis
through upregulation of Deptor in unilateral ureteral obstruction
in rats. Exp Ther Med. 20(1)2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Caron A, Briscoe DM, Richard D and
Laplante M: DEPTOR at the nexus of cancer, metabolism, and
immunity. Physiol Rev. 98:1765–1803. 2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Wedel J, Bruneau S, Liu K, Kong SW, Sage
PT, Sabatini DM, Laplante M and Briscoe DM: DEPTOR modulates
activation responses in CD4+ T cells and enhances immunoregulation
following transplantation. Am J Transplant. 19:77–88.
2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Zhai Y, Lin P, Feng Z, Lu H, Han Q, Chen
J, Zhang Y, He Q, Nan G, Luo X, et al: TNFAIP3-DEPTOR complex
regulates inflammasome secretion through autophagy in ankylosing
spondylitis monocytes. Autophagy. 14:1629–1643. 2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Bruneau S, Nakayama H, Woda CB, Flynn EA
and Briscoe DM: DEPTOR regulates vascular endothelial cell
activation and proinflammatory and angiogenic responses. Blood.
122:1833–1842. 2013.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Laplante M, Horvat S, Festuccia WT, Birsoy
K, Prevorsek Z, Efeyan A and Sabatini DM: DEPTOR cell-autonomously
promotes adipogenesis, and its expression is associated with
obesity. Cell Metab. 16:202–212. 2012.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Cadegiani FA, McCoy J, Gustavo Wambier C,
Vaño-Galván S, Shapiro J, Tosti A, Zimerman RA and Goren A:
Proxalutamide significantly accelerates viral clearance and reduces
time to clinical remission in patients with mild to moderate
COVID-19: Results from a randomized, double-blinded,
placebo-controlled trial. Cureus. 13(e13492)2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Kanno Y, Zhao S, Yamashita N, Yanai K,
Nemoto K and Inouye Y: Androgen receptor functions as a negative
transcriptional regulator of DEPTOR, mTOR inhibitor. J Toxicol Sci.
40:753–758. 2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Reed LJ and Muench H: A simple method for
estimating fifty percent endpoints. Am J Hyg. 27:493–497. 1938.
|
|
35
|
Nayak G, Bhuyan SK, Bhuyan R, Sahu A, Kar
D and Kuanar A: Global emergence of Enterovirus 71: A systematic
review. Beni Suef Univ J Basic Appl Sci. 11(78)2022.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Yogarajah T, Ong KC, Perera D and Wong KT:
AIM2 inflammasome-mediated pyroptosis in enterovirus A71-infected
neuronal cells restricts viral replication. Sci Rep.
7(5845)2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Bai J, Chen X, Liu Q, Zhou X and Long JE:
Characteristics of enterovirus 71-induced cell death and genome
scanning to identify viral genes involved in virus-induced cell
apoptosis. Virus Res. 265:104–114. 2019.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Wang H, Lei X, Xiao X, Yang C, Lu W, Huang
Z, Leng Q, Jin Q, He B, Meng G and Wang J: Reciprocal regulation
between enterovirus 71 and the NLRP3 inflammasome. Cell Rep.
12:42–48. 2015.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Chen H, Ning X and Jiang Z: Caspases
control antiviral innate immunity. Cell Mol Immunol. 14:736–747.
2017.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Wang SM, Lei HY and Liu CC: Cytokine
immunopathogenesis of enterovirus 71 brain stem encephalitis. Clin
Dev Immunol. 2012(876241)2012.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Peterson TR, Laplante M, Thoreen CC,
Sancak Y, Kang SA, Kuehl WM, Gray NS and Sabatini DM: DEPTOR is an
mTOR inhibitor frequently overexpressed in multiple myeloma cells
and required for their survival. Cell. 137:873–886. 2009.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Zhao W, Shi CS, Harrison K, Hwang IY,
Nabar NR, Wang M and Kehrl JH: AKT regulates NLRP3 inflammasome
activation by phosphorylating NLRP3 serine 5. J Immunol.
205:2255–2264. 2020.PubMed/NCBI View Article : Google Scholar
|