Introduction
Alzheimer's disease (AD) is a progressive,
persistent, and degenerative disease of the central nervous system
(CNS), with clinical manifestations of cognitive impairment and
memory impairment (1,2). The pathogenesis of AD is not clear,
and there is a lack of effective treatments. Currently,
cholinesterase inhibitors are commonly used in the clinical
treatment of AD, but these drugs can only improve and relieve
symptoms. Natural medicines, including polysaccharides (3,4),
phenylpropanoids (5,6), flavonoids (7,8),
alkaloids (9), saponins (10,11),
and polyphenols (12,13), have also been considered for the
treatment of AD.
Tripterygium glycoside (TG), also known as
Tripterygium, is the total glycoside extracted from the peeled
roots of Tripterygium wilfordii Hook.f. It has been used in
the treatment of rheumatoid arthritis (RA) (14), lupus nephritis (LN) (15), diabetes mellitus (DM) (16), and Guillain-Barre syndrome (GBS)
(17). Animal experiments have
shown that TG has protective effects on the CNS. It has been
suggested that TG can significantly improve the inflammatory damage
to astrocytes induced by lipopolysaccharide (LPS) by decreasing the
expression of TNF-α, iNOS, and IL-6(18). In our previous study, TG suppressed
the release of inflammatory factors and inhibited the
phosphorylation of IκBα and p38 MAPK in Aβ25-35-induced
AD mice (19). However, the
mechanism of action of TG in AD remains to be determined.
Noncoding RNAs, including long noncoding RNAs
(lncRNAs), circular RNAs (circRNAs), and microRNAs (miRNAs), have
been studied due to the extent of their expression and their
involvement in several biological processes (20). LncRNAs, such as BACE1- AS (21), 17A (22), NDM29(23), 51A (24), BC200(25), and NAT-RAD18(26), have been reported to be involved in
the formation of senile plaques, DNA repair or synaptic formation,
as well as in the pathogenesis of AD. CircRNAs are derived from
mRNA precursors, which may affect normal cell differentiation,
maintain tissue homeostasis, and influence the progression of
various diseases (27).
Additionally, circRNAs have been shown to play an important role in
the occurrence and development of AD by influencing neuronal
genesis and injury, Aβ deposition, neuroinflammation, autophagy,
and synaptic function through the function of microRNA (miRNA/miR)
sponges (28-30).
Thus, lncRNAs and circRNAs may be considered risk factors,
progression biomarkers, and therapeutic targets for AD.
However, the lncRNAs and circRNAs regulated by TG in
AD treatment have not been determined. In the present study, the
expression profiles of the lncRNAs and circRNAs of an AD mouse
model treated with TG were determined using microarrays.
Materials and methods
Animal model of AD
The study was performed in accordance with the
ARRIVE guidelines (https://arriveguidelines.org/), and the protocols
followed the National Institutes of Health Guide for the Care and
Use of Laboratory Animals (31).
The research procedures were approved by the Ethics Committee of
Changsha Medical University (approval no. EC20190114). A total of
Twenty-four C57BL/6 J mice (male, 28±5 g, 6 months old) were
purchased from Silaike Jingda Biotechnology Co., Ltd. (12th Sep
2021; Changsha, China). The mice were fed standard chow, and housed
under controlled conditions (temperature, 23±0.6˚C; relative
humidity, 55±8%; 12 h light: dark cycle). Animal health and
behavior of all mice were monitored weekly. The neuroprotective
effects of TG on AD mouse models were identified in our previous
study (19). A normal control
group was excluded from the present study. Thus, the mice were
divided into two groups: AD+TG group and AD+normal saline (NS)
group. The AD model was constructed as described in our previous
study (19). Except for 8 mice
that died, a total of 16 AD mice models were included in further
study. Mice in the AD+TG group (n=8) were treated with TG (0.25
mg/10 g.d, 1 mg/ml). The dosing and duration of TG followed the
study conducted by Wang et al (18). Mice in the AD+NS group (N=8) were
treated with NS (0.9%) 0.5 ml/d. The treatments were administered
by intraperitoneal injection once per day for 4 weeks. At the end
of treatment, a total of 6 and 7 mice were obtained in the AD+TG
group and AD+NS group, respectively. From each group, 3 samples
were selected randomly for lncRNA and mRNA microarray analysis. Any
surviving animals (n=3 in the AD+TG group and n=4 in the AD+NS
group) at the end of the experiment were euthanized by exposure to
carbon dioxide (CO2) overdose with the CO2
displacement rate of 30-70% of the cage volume per min. Death was
confirmed based on a lack of heartbeat and brain death (no
environmental response, pupil reflex to light or spontaneous
breathing).
Tissue collection and RNA
extraction
General anesthesia was performed by intraperitoneal
injection of 0.2% sodium pentobarbital (40 mg/kg). Hippocampal
tissues were isolated from mouse brains and stored at -80˚C until
required for further analysis. TRIzol® (Invitrogen;
Thermo Fisher Scientific, Inc.) was used to extract total RNA from
hippocampal tissues. Then, DNAse I was used to remove DNA
contamination. The quality of RNA was quantified using a NanoDrop
spectrophotometer (Thermo Fisher Scientific, Inc.).
Microarray analysis
PCR amplification and fluorescence labeling of total
RNA from 6 samples [AD+NS (n=3) and AD+TG (n=3)] were performed
using an Agilent expression spectrum chip kit (Agilent, California,
USA). The labeled cRNA was purified with an RNA extraction and
purification kit (Sigma-Aldrich, St. Louis, USA). A total of 600 µg
cRNA was hybridized using an Agilent-085631 microarray (Agilent
Technologies, Inc.) with the following conditions: 65˚C for 17 h at
3.354 x g. Feature Extraction Software version 12.0 (Agilent
Technologies, Inc.) was used to read the data. The Limma package
(release 3.16; https://www.bioconductor.org/packages/release/bioc/html/limma.html)
(32) in R-4.2.3 software
(https://cran.r-project.org/bin/windows/base/)
(33,34) was used for normalization. The
differentially expressed lncRNAs, circRNAs, and mRNAs with a fold
change ≥2 and P<0.05 were selected and identified. Clustering
analyses were performed with hierarchical and average linkage
algorithms using Mev version 4.9.0(35).
Reverse transcription-quantitative
(RT-q)PCR
Four lncRNAs, mRNAs, and circRNAs were randomly
selected and identified by qPCR. Trans-script II First-strand cDNA
Synthesis SuperMix Kit (Sigma-Aldrich, St. Louis, USA) was used for
reverse transcription according to the manufacturer's protocol. The
TB Green Premix ExTaq™ kit (Takara Bio, Inc.) was used for qPCR.
The reaction conditions were as follows: 95˚C for 5 min, followed
by 38 cycles of 94˚C for 30 sec, 58˚C for 30 sec and 72˚C for 60
sec. The relative expression levels of lncRNAs and mRNAs were
analyzed using the 2-ΔΔCq method (36) and normalized to GAPDH.
Amplification was performed on an ABI7500 quantitative PCR
instrument (Applied Biosystems; Thermo Fisher Scientific, Inc.).
The sequences of the primers are listed in Table I.
 | Table ISequences of the primers used for
quantitative PCR detection of the selected lncRNAs, circRNAs, and
mRNAs. |
Table I
Sequences of the primers used for
quantitative PCR detection of the selected lncRNAs, circRNAs, and
mRNAs.
| Gene names | Forward, 5'-3' | Reverse, 5'-3' |
|---|
|
NONMMUG042458.2 |
GACACTGGCGTGAAAAAGGAAC |
GACAATCAAGGCAGGAGGAG |
|
NONMMUG004800.3 |
CCTCCACCCCTACAGGAAGAG |
GACAGGAGCAAAAGTGACTT |
|
NONMMUG007415.2 |
GAAATGGCAACTCAGCGGAGAG |
GAAATTATCCAGGATGAGAAAG |
|
NONMMUT141368.1 |
GTCTCGGTGGGCGGGCATG |
GTGTCTGAGACAAAAGGGAG |
| Skint8 | ACCACTCCCA
CAAGACACCT |
GAAGGAGGCCATTGGAGAAG |
| Lce1b | AGATGTCCTG
CCAGCAGAAC |
GGTGGTTGCTGCAGTTCTGG |
| Padi4 |
GGTGAAAGCAGCCAGCAGCAG |
GAATGGACTTTGAGGATGAC |
| Slc4a9 |
CTTCATTCAACTAAATGAGC TG |
AGCCCAGAACTGAGAGGACA GCT |
|
MMU_CIRCpedia_20654 |
ACATGAGCCTTCAGAGATAC |
CAGAGGCAACAACTACCC |
|
mmu_circ_0010693 |
GGAACATTTCCATCAACATT | CTCTGAATTACTGC |
|
MMU_CIRCpedia_214399 |
TGATGTCATCCTGATAGTTG |
GGTTGACATCGACCAA |
|
mmu_circ_0010830 |
AGGATATTCACAGACATGC |
GAACATTGAGCCTACTCAAG |
Co-expression network analysis
Pearson's correlation analysis was used to calculate
the correlation coefficients (r) and P-values. The screening
criteria were r>|0.85| and P<0.05. The co-expression network
was generated using Circos software (version 0.69-6; http://circos.ca/software/download/) (37).
PPI network of the differentially
expressed mRNAs
The PPI network of differentially expressed mRNAs
was generated using STRING (version 11.5) (38) (https://cn.string-db.org/) with a threshold score
>0.4.
Functional classification and pathway
analysis
Gene Ontology (GO) analysis (39,40)
(http://geneontology.org/page/go-database) from three
aspects [biological process (BP), cellular component (CC), and
molecular function (MF)], and Kyoto Encyclopedia of Genes and
Genomes (KEGG) pathway analysis (http://www.kegg.jp/) (41) were used to determine the roles of
the differentially expressed mRNAs. P<0.01 was used as the
selection criterion to analyze the difference in mRNAs involved in
the pathways.
Cis/trans target gene prediction of
lncRNA
All coding genes within 100k bp upstream and
downstream of differentially expressed lncRNAs were selected as the
target gene for cis-regulation. Cis-regulation was assessed using
the FEELNC software version 0.2.1(42). Trans-regulation was predicted using
RIsearch version 2.0 (Center for non-coding RNA in Technology and
Health, Department of Veterinary and Animal Sciences, Faculty for
Health and Medical Sciences, University of Copenhagen
Frederiksberg, Denmark) (43,44)
(https://rth.dk/resources/risearch/)
with the following parameters: Number of interacting bases between
lncRNA and gene, <10; binding free energy, <-100.
lncRNA-target-transcription factor
(TF) network analysis
Potential lncRNA binding TFs were predicted based on
the JASPAR database (http://jaspar.genereg.net/). The gene-TF pairs
provided by the GTRD database (http://gtrd.biouml.org/) and the co-expression
relationship of lncRNA-mRNA pairs were used to construct the
3-element regulatory network of the lncRNA-TF-mRNA. The top 500
relationship pairs were extracted, and the regulatory network
diagram of lncRNA-TF-mRNA was drawn using the network software.
Prediction of circRNA-miRNA
interactions
The interaction between circRNA and related miRNAs
was predicted using miRDB database (http://www.mirdb.org/). The miRNA response element
(MRE) of different circRNAs was studied. Target miRNAs were
selected based on complementary pairing sequences. Cytoscape
software (version.3.9.1) (45)
(http://circos.ca/software/download/)
was used to construct the circRNA-miRNA network.
Statistical analysis
Data were analyzed using SPSS 22.0 (IBM Corp.). Data
are presented as the mean ± SD, and a Student's t-test was used to
compare differences between the two groups. All tests were
two-sided, and P<0.05 was considered to indicate a statistically
significant difference.
Results
Differentially expressed lncRNAs,
mRNAs, and circRNAs in the Aβ25-35-induced AD mouse
model treated with TG
In total, 108,510 lncRNAs, 15,321 circRNAs, and
43,093 mRNAs were obtained by high-throughput sequencing. Compared
with the control group, 661 lncRNAs, 64 circRNAs, and 503 mRNAs
were significantly differentially expressed following TG treatment,
including 422, 341, and 34 upregulated, as well as 81, 320, and 30
downregulated mRNAs, lncRNAs, and circRNAs, respectively. Amongst
them, NONMMUG090228.1 (FC=268.27) and NONMMUG011123.2 (FC=-36.51)
were the most upregulated and downregulated lncRNAs, respectively,
Pou4f1 (FC=26.01) and Gm14781 (FC=-10.75) were the most upregulated
and downregulated mRNAs, respectively, and MMU_CIRCpedia_35174
(FC=5.47) and mmu_circ_0007187 (FC=-4.04) were the most upregulated
and downregulated circRNAs, respectively. The lncRNA, mRNA, and
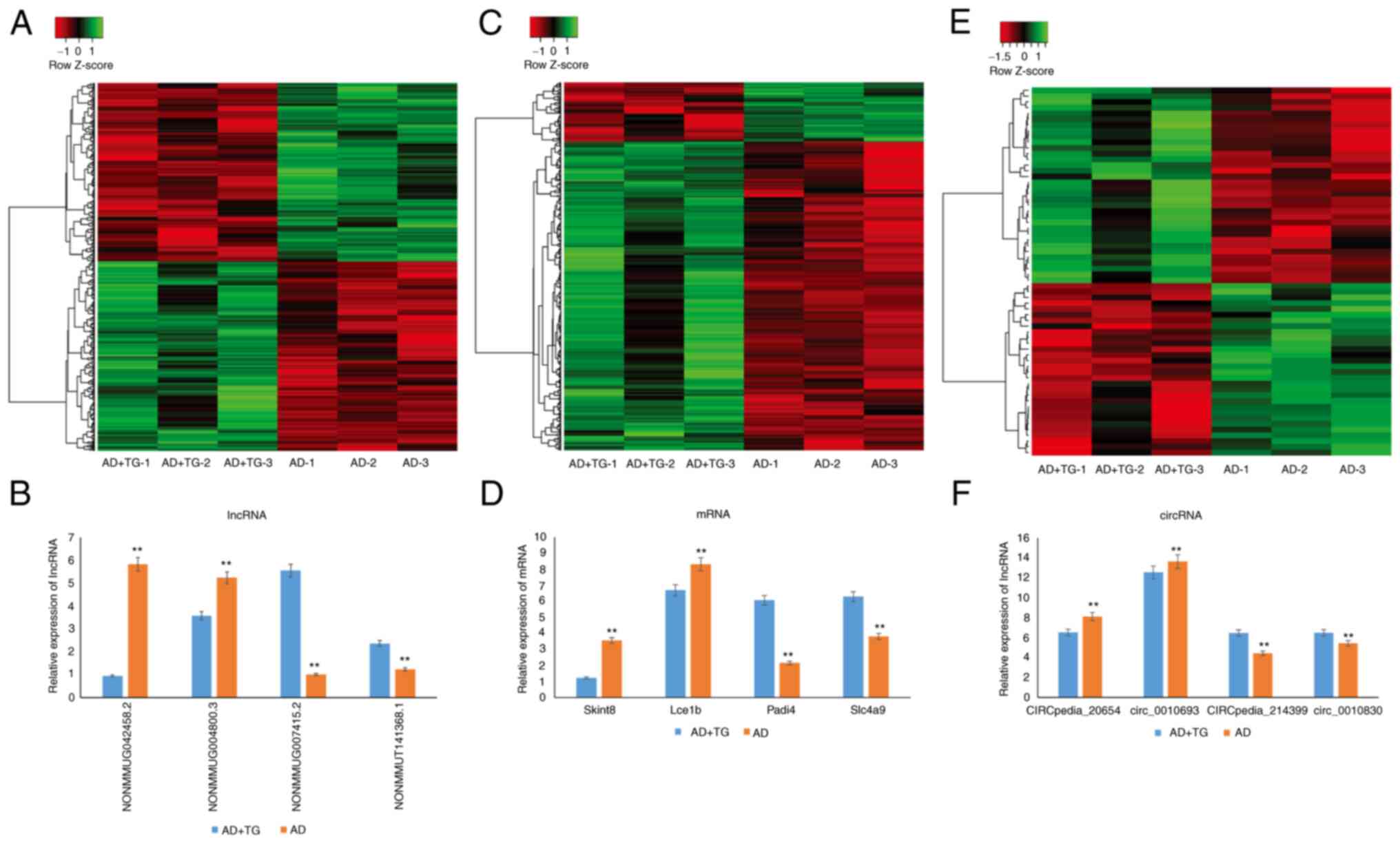
circRNA expression patterns in different samples are shown in
Fig. 1A-C.
To verify the microarray results, the expression of
4 randomly selected lncRNAs (2 upregulated and 2 downregulated), 4
mRNAs (2 upregulated and 2 downregulated), and 4 circRNAs (2
upregulated and 2 downregulated) using qPCR. The results showed
that the expression trend of the selected genes was consistent with
the microarray analysis (Fig.
1D-F).
Co-expression of lncRNAs with mRNAs,
and circRNAs with mRNAs
Overall, 648 network nodes and 152,453 connections
(64,141 negative and 88,312 positive interactions) were identified
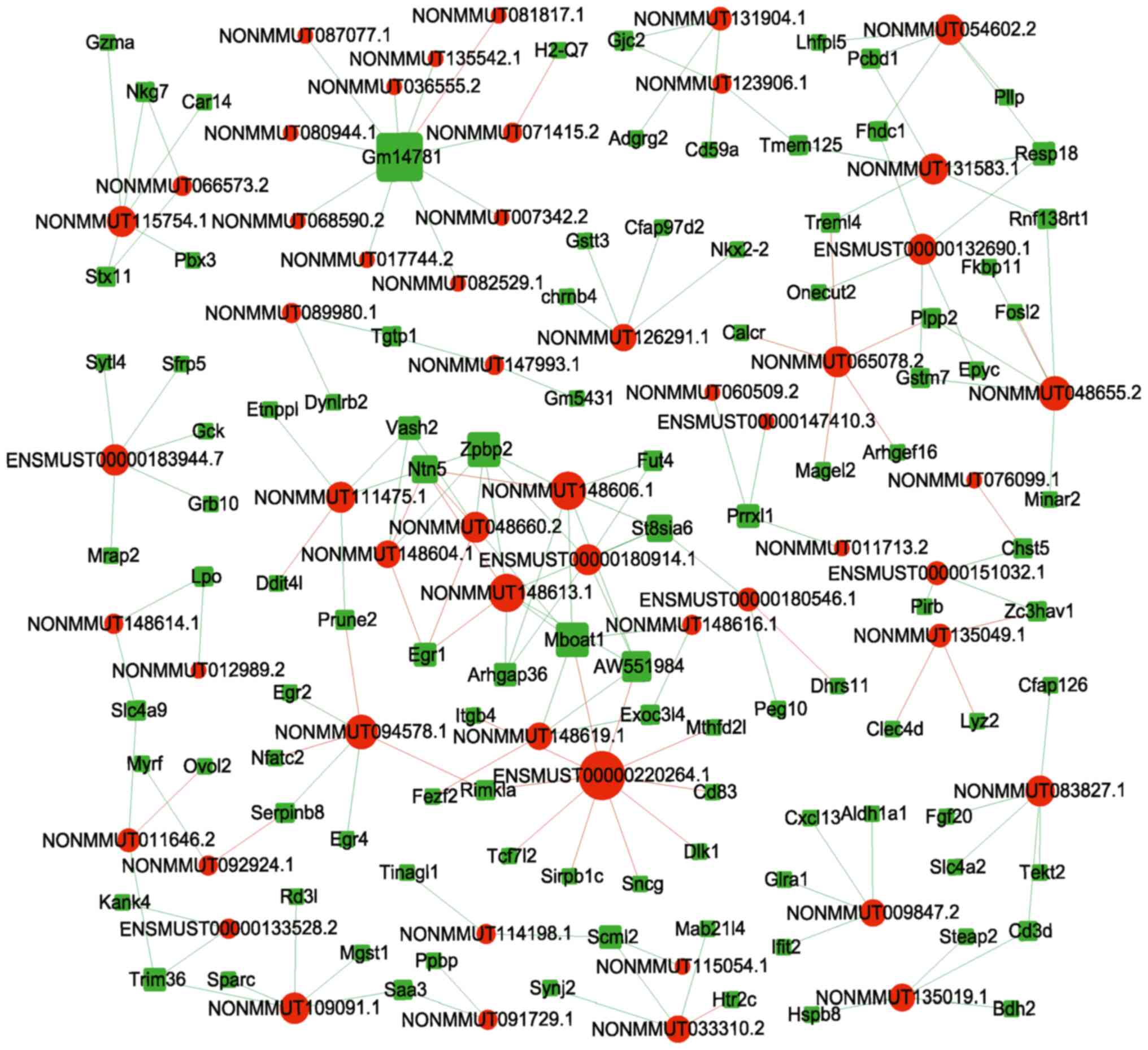
in the lncRNA-mRNA co-expression network. The correlations between
the top 20 (10 up and 10 downregulated) dysregulated lncRNAs and
mRNAs are shown in Fig. 2. In
total, 19,553 pairs of mRNA-circRNA interactions were identified
(8,871 negative and 10,682 positive interactions) in the
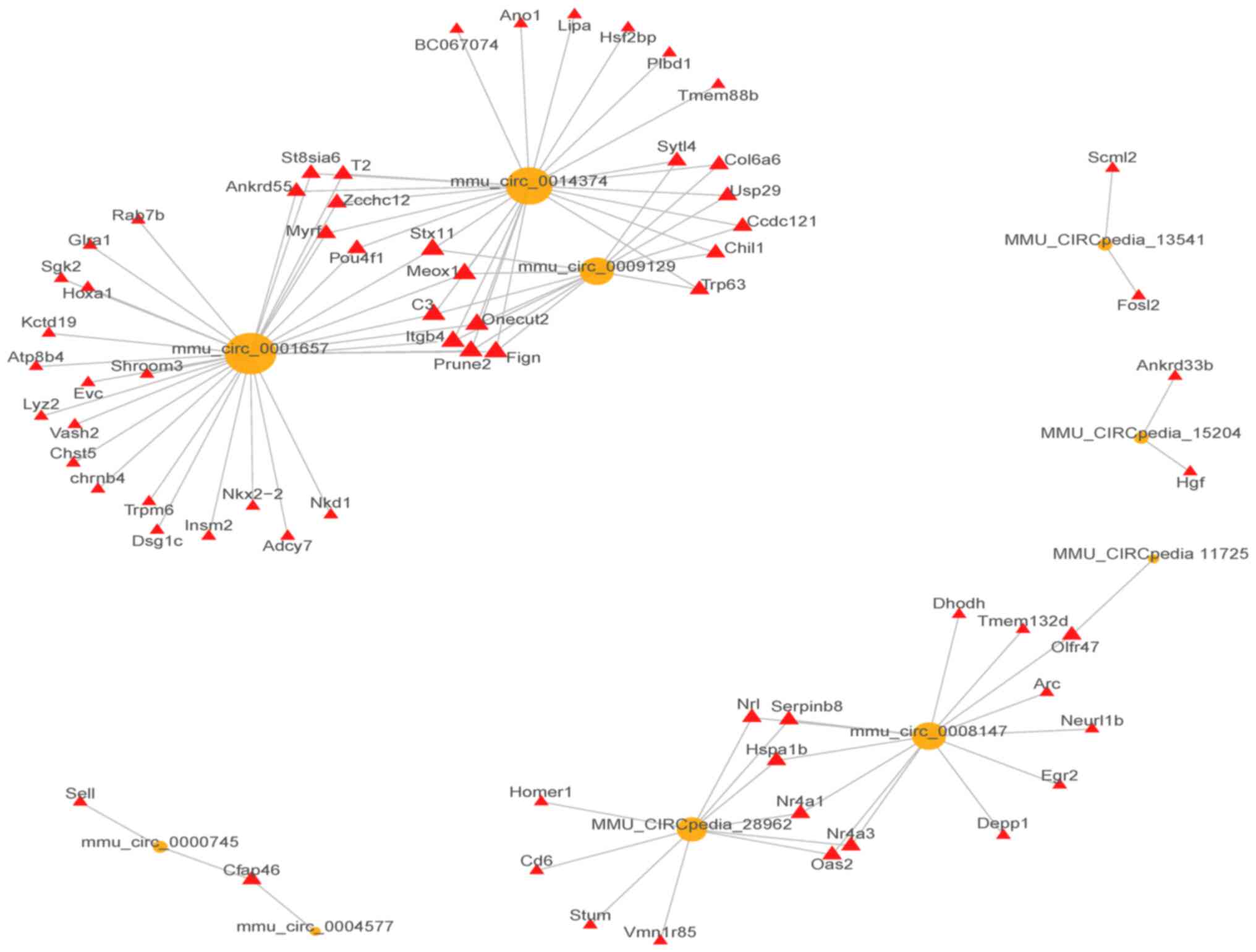
circRNA-mRNA co-expression network. The correlations between the
top 10 (5 up and 5 downregulated) dysregulated lncRNAs and mRNAs
are shown in Fig. 3.
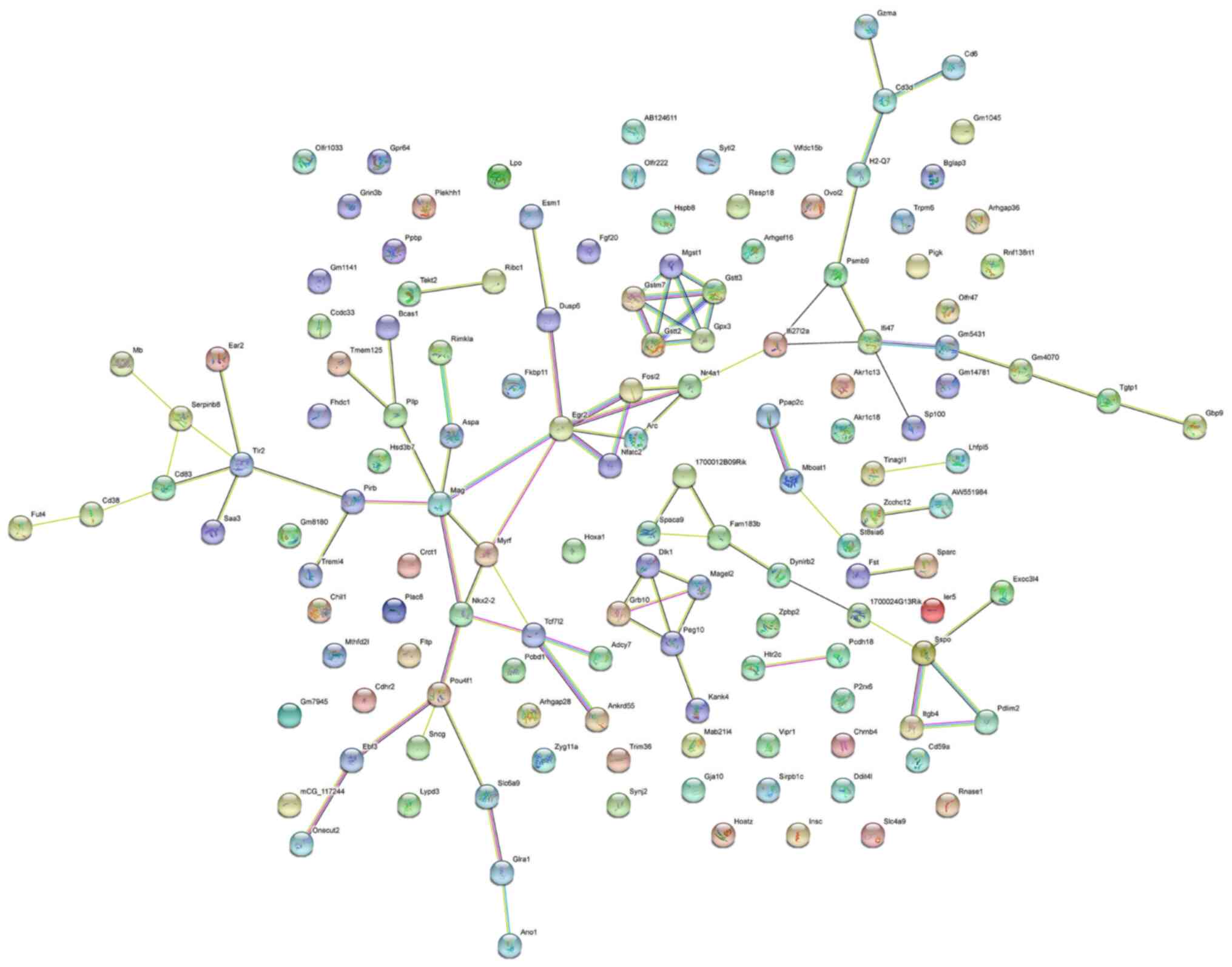
PPI network of differentially
expressed mRNAs
The PPI network analysis conducted on the top 200
differentially expressed mRNAs consisted of 139 nodes and 88 edges,
with an average node degree of 1.27 (Fig. 4; Table SI). Amongst the mRNAs, Pou4f1,
Egr2, Mag, and Nr4a1 were the hub genes in the PPI network. The
protein-encoding genes in the PPI network were primarily enriched
in ‘Oligodendrocyte development’ (GO: 0014003), ‘Myelination’ (GO:
0042552), and ‘Glial cell development’ (GO: 0021782), and were
involved in the ‘Glutathone metabolism’ pathway (mmu00480).
GO and KEGG analysis of lncRNAs
co-expressed with mRNAs
Overall, 499 genes were identified, including 396
genes annotated by BP, 402 genes annotated by CC, and 398 genes
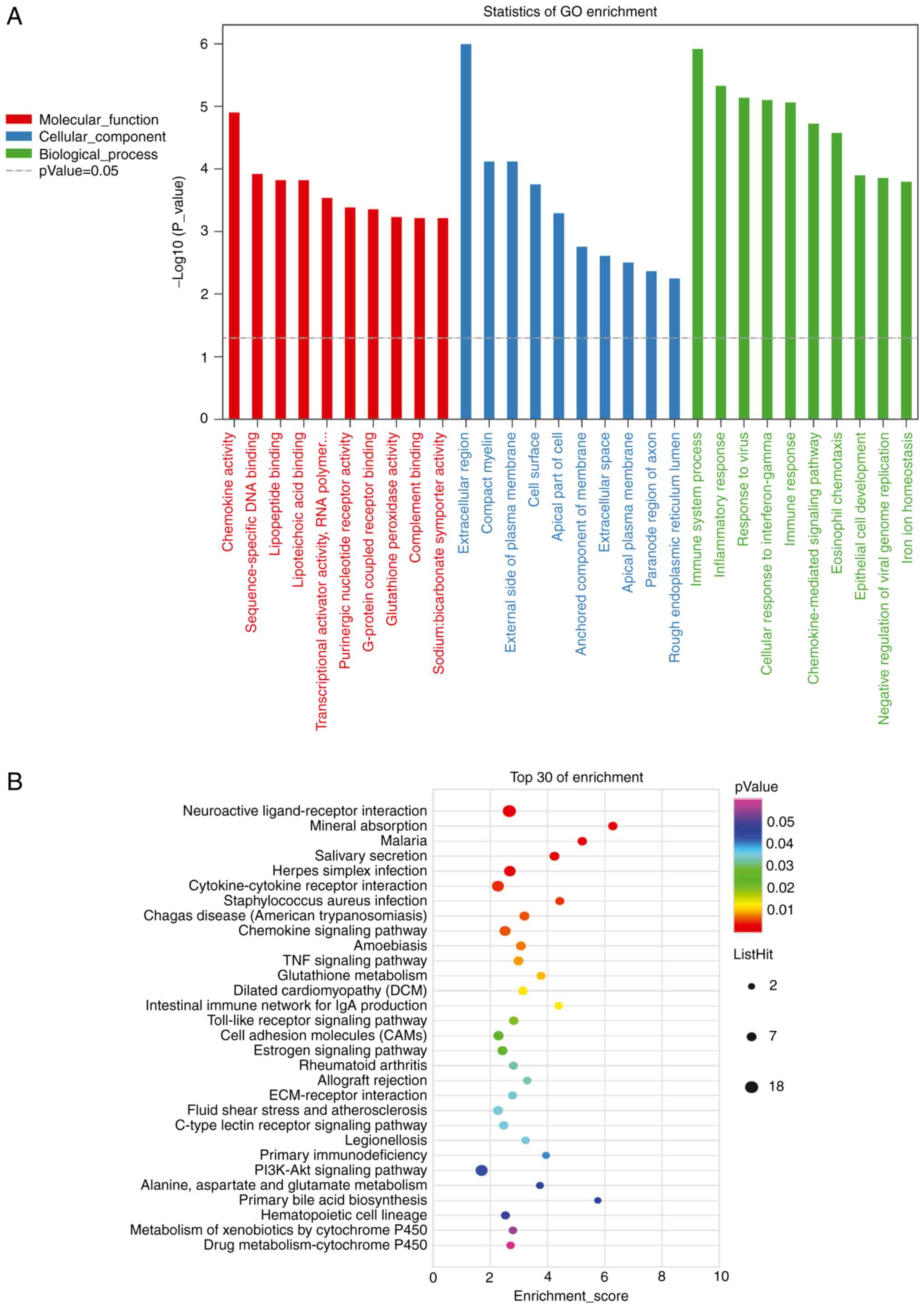
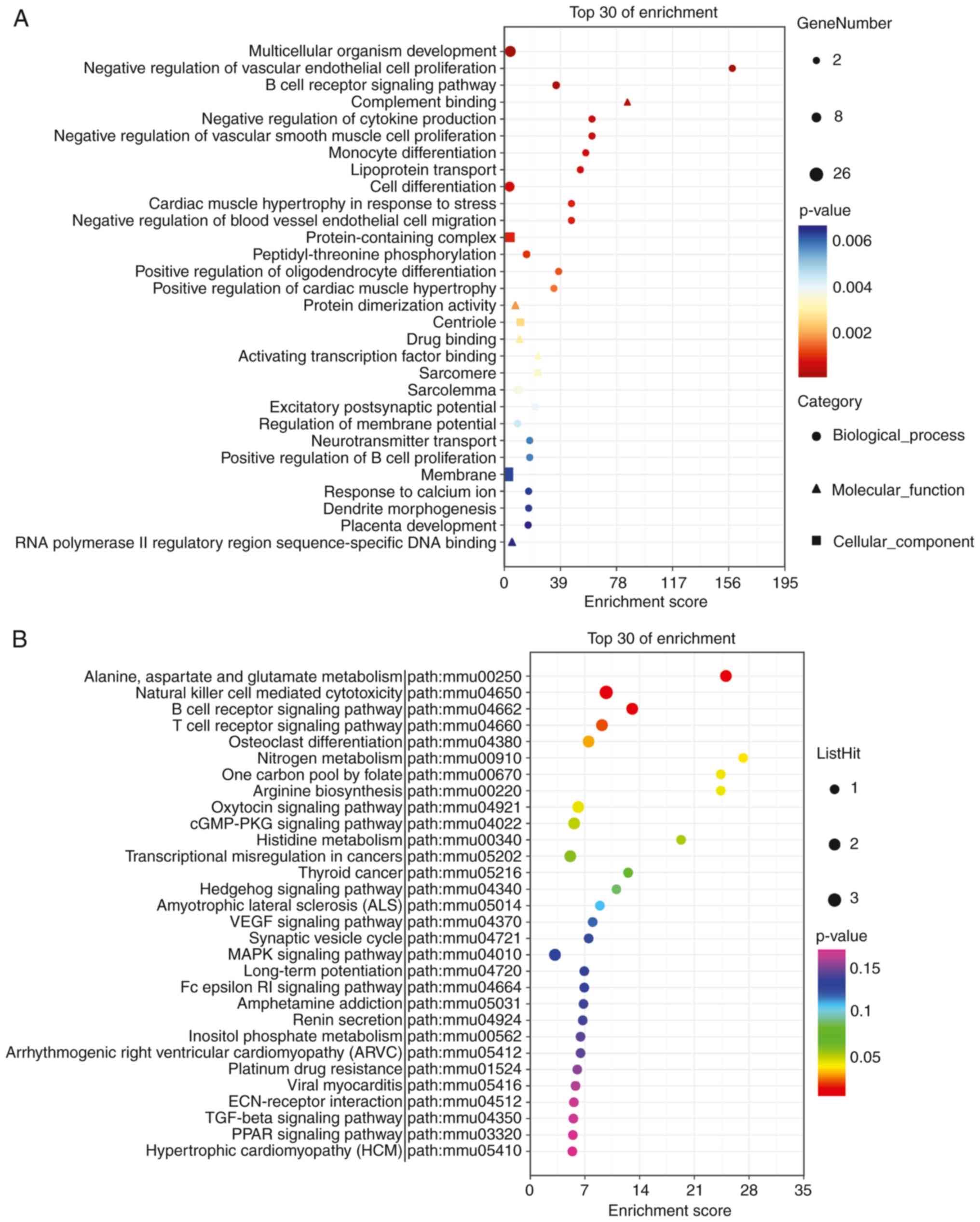
annotated by MF. There were 284 terms with P-values ≤0.05.
‘Extracellular region’ (GO:0005576, P=9.9x10-7), ‘immune
system process’ (GO:0002376, P=1.2x10-6), and
‘inflammatory response’ (GO:0006954, P=4.6x10-6) had the
lowest P-values (Fig. 5A; Table SII).
In total, 171 genes were annotated using KEGG
pathway analysis, with 28 pathways having P-values ≤0.05. The top
three pathways with minimum P-values were ‘Neuroactive
ligand-receptor interaction’ (path: MMU04080, P=0.00013), ‘Mineral
absorption’ (path: mmu04978, P=0.00034), and ‘Malaria’ (path:
mmu05144, P=0.00095). KEGG pathway analysis suggested that lncRNAs
were also involved in the ‘TNF signaling pathway’, ‘PI3K-Akt
signaling pathway’, and ‘Wnt signaling pathway’ (Fig. 5B; Table SIII).
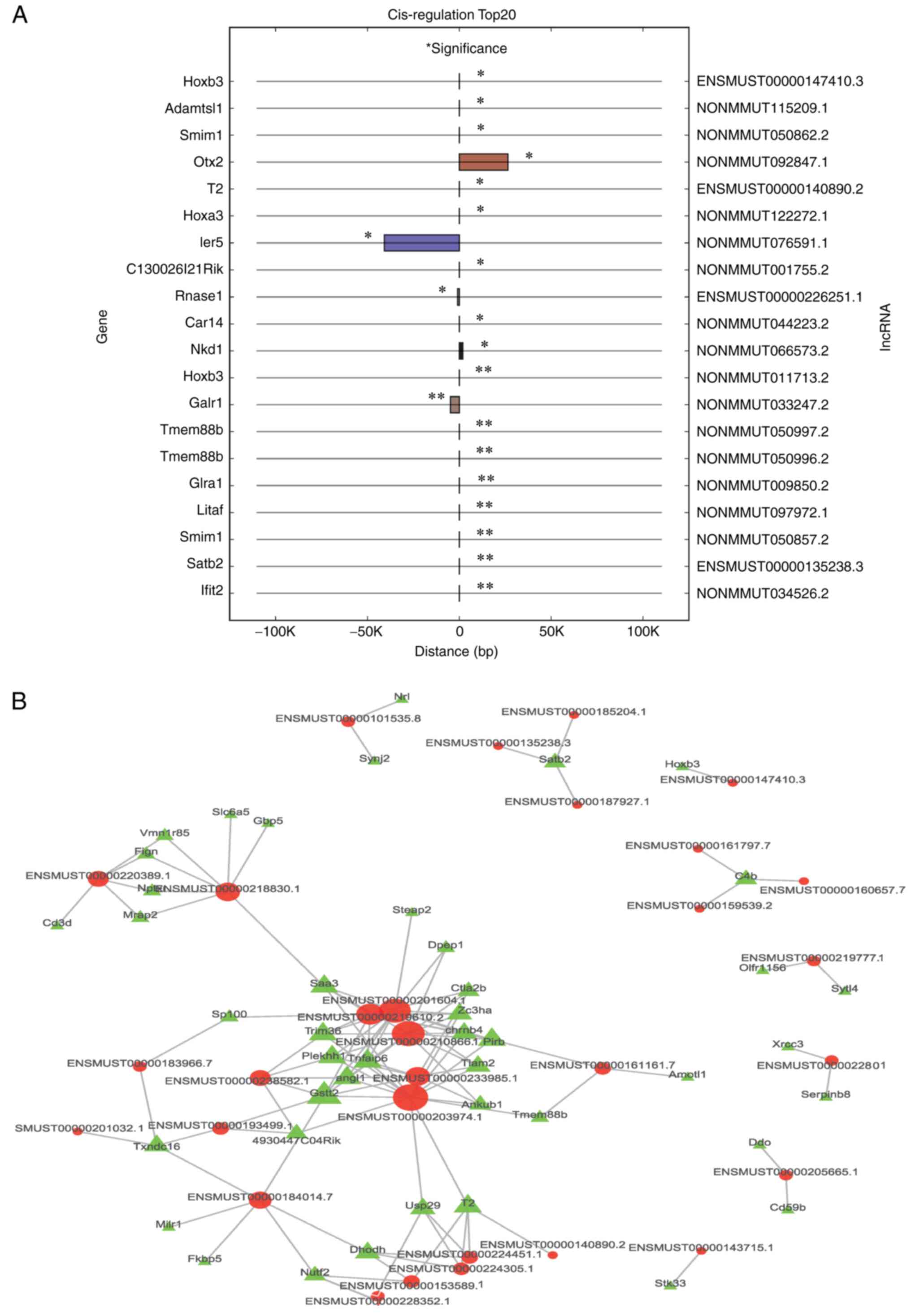
Cis/trans target genes of lncRNAs
The top 20 cis-regulatory results are shown in
Fig. 6A, and 46 cis-regulated
pairs were identified, most of which were positive regulation pairs
(91.3%). Additionally, the top 500 trans-regulated results are
shown in Fig. 6B, where 748
trans-regulation pairs were identified.
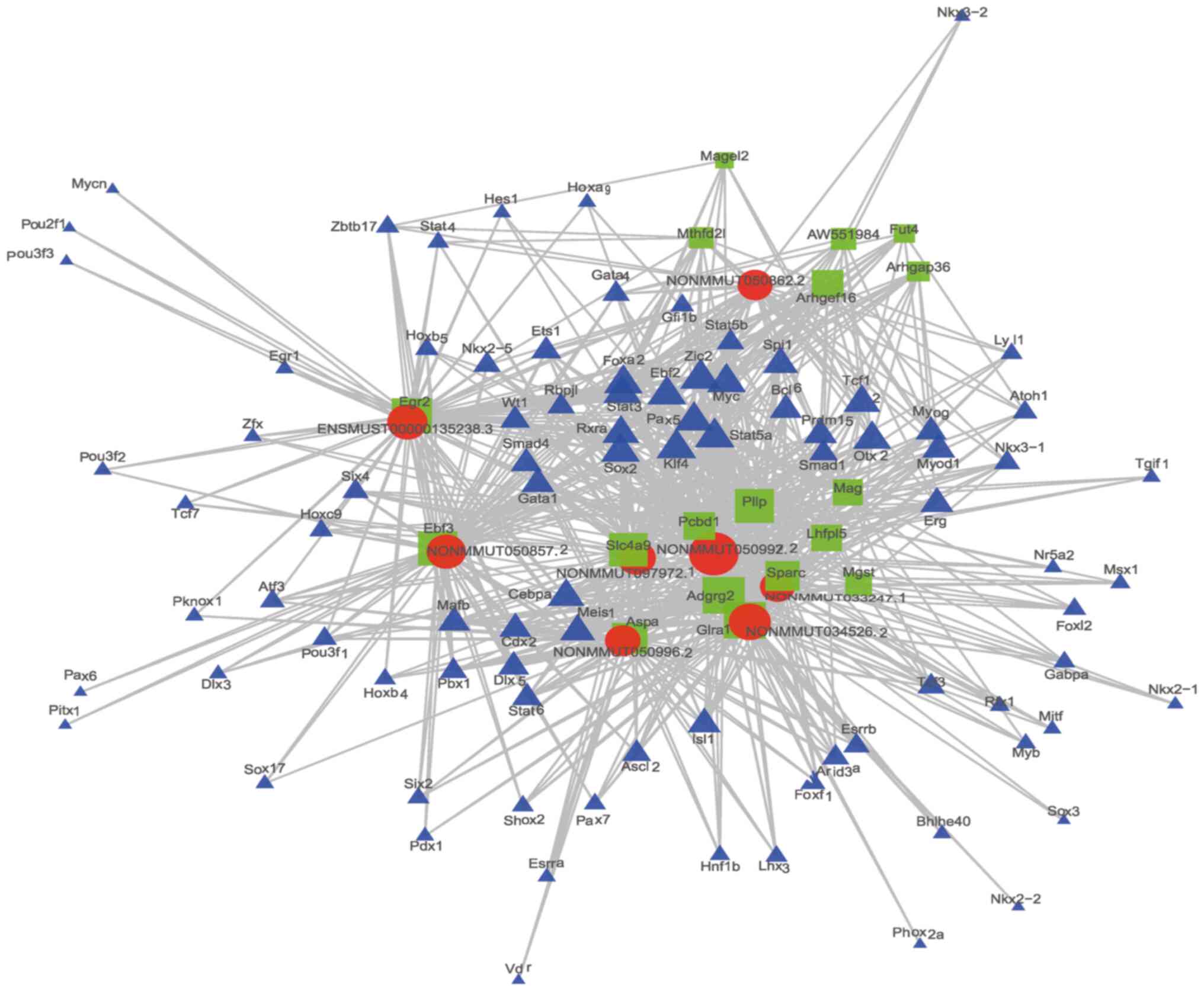
LncRNA-Target-TF network
In total, 98 TFs and 75 mRNAs were predicted to
regulate or be the target of the top 20 cis-lncRNAs. Among the TFs,
Cebpa (n=170), Zic2 (n=166), and Rxra (n=159) were predicted to
regulate most of the lncRNAs. The 2 most related lncRNA-mRNA and
lncRNA-TF pairs according to the P-value were used to construct the
lncRNA-target-TF network (Fig.
7).
Enrichment analysis of circRNA host
genes
Overall, 61 circRNA host genes were analyzed. Of
these, 51 were annotated by BP, 53 genes were annotated by CC, and
52 genes were annotated for MF. ‘Multicellular organism
development’ (GO: 0007275, P=2.1x10-5), ‘negative
regulation of vascular endothelial cell proliferation’ (GO:
1905563, P=6.2x10-5), and ‘B-cell receptor signaling
pathway’ (GO: 0050853, P=7.8x10-5) had the smallest
P-values (Fig. 8A). In addition,
61 genes were identified in this study, and 17 genes were annotated
as KEGG pathways. The top three pathways with minimum P-values were
‘Alanine, aspartate, and glutamate metabolism’ (path: mmu00250,
P=0.0028), ‘Natural killer cell-mediated cytotoxicity’ (path:
mmu04650, P=0.0033), and ‘B-cell receptor signaling pathway’ (path:
mmu04662, P=0.01, Fig. 8B).
CircRNA-miRNA interactions
Overall, 275 miRNAs that bind to the 64 circRNAs
were predicted. The top 4 differentially expressed circRNAs (2
upregulated and 2 downregulated) were mmu_circ_0007187,
mmu_circpedia_35174, mmu_circpedia_35014, and mmu_circpedia_15204,
and they were shown to bind to 5 miRNAs. The top 2 circRNA-miRNA
interaction pairs are shown in Table
II.
| Table IIThe circRNA-miRNA interaction pairs
of mmu_circ_0007187, mmu _circpedia_35174, mmu_circpedia_35014, and
mmu_circpedia_15204. |
Table II
The circRNA-miRNA interaction pairs
of mmu_circ_0007187, mmu _circpedia_35174, mmu_circpedia_35014, and
mmu_circpedia_15204.
| miRNA | circRNA | Total score | Total energy | Max score | Max energy | miRNA length | circRNA length | Positions |
|---|
|
mmu_circ_0007187 |
mmu-miR-92a-2-5p | 881 | -144.01 | 165 | -27.79 | 22 | 1,816 | 461, 1,337, 76,
1,301 1,373, 1,310 |
| |
mmu-miR-7006-5p | 753 | -129.77 | 167 | -30.12 | 25 | 1,816 | 1,329, 1,127,
1,409, 1,295, 1,300 |
| |
mmu-miR-7092-3p | 706 | -84.66 | 145 | -22.43 | 24 | 1,816 | 618, 269, 207,
1,470, 1,786 |
| |
mmu-miR-6931-5p | 620 | -123.24 | 169 | -36.94 | 21 | 1,816 | 1,343, 1,303, 464,
1,330 |
| |
mmu-miR-6914-5p | 615 | -146.74 | 167 | -40.26 | 24 | 1,816 | 1,281, 1,415, 781,
947 |
|
MMU_CIRCpedia_35314 |
mmu-miR-669h-5p | 326 | -35.45 | 165 | -19.12 | 25 | 378 | 273, 93 |
| |
mmu-miR-7048-3p | 314 | -39.08 | 168 | -23.15 | 24 | 378 | 274, 96 |
| |
mmu-miR-1966-5p | 302 | -31.78 | 159 | -17.38 | 22 | 378 | 1, 73 |
| |
mmu-miR-1a-2-5p | 295 | -44.5 | 154 | -22.58 | 22 | 378 | 209, 63 |
| |
mmu-miR-1a-1-5p | 292 | -43.52 | 148 | -23.17 | 22 | 378 | 130, 74 |
|
MMU_CIRCpedia_35174 |
mmu-miR-7680-5p | 288 | -30.18 | 144 | -16.93 | 20 | 393 | 86, 310 |
| | mmu-miR-205-5p | 180 | -29.63 | 180 | -29.63 | 22 | 393 | 63 |
| |
mmu-miR-669h-3p | 168 | -15.87 | 168 | -15.87 | 22 | 393 | 170 |
| | mmu-miR-8118 | 166 | -18.86 | 166 | -18.86 | 22 | 393 | 1 |
| | mmu-miR-669i | 163 | -14.74 | 163 | -14.74 | 21 | 393 | 169 |
|
MMU_CIRCpedia_35014 | mmu-miR-500-5p | 326 | -35.45 | 165 | -19.12 | 25 | 378 | 273, 93 |
| | mmu-miR-362-5p | 314 | -39.08 | 168 | -23.15 | 24 | 378 | 274, 96 |
| |
mmu-miR-7028-3p | 302 | -31.78 | 159 | -17.38 | 22 | 378 | 1, 73 |
| |
mmu-miR-3074-5p | 295 | -44.5 | 154 | -22.58 | 22 | 378 | 209, 63 |
| | mmu-miR-1903 | 292 | -43.52 | 148 | -23.17 | 22 | 378 | 130, 74 |
|
MMU_CIRCpedia_15204 | mmu-miR-203-5p | 448 | -54.12 | 163 | -27.81 | 22 | 1,332 | 361, 515, 185 |
| |
mmu-miR-7028-5p | 438 | -70.06 | 153 | -25.95 | 23 | 1,332 | 211, 594, 1303 |
| | mmu-miR-432 | 437 | -57.75 | 151 | -20.8 | 23 | 1,332 | 599, 314, 953 |
| | mmu-miR-1291 | 333 | -38.53 | 180 | -21.55 | 26 | 1,332 | 534, 358 |
| | mmu-miR-761 | 319 | -45.01 | 168 | -23.52 | 22 | 1,332 | 431, 1,124 |
Discussion
In the present study, hippocampal tissues of AD+NS
mice and AD+TG mice were used for microarray analysis. In total,
661 differentially expressed lncRNAs, 503 differentially expressed
mRNAs, and 64 differentially expressed circRNAs were screened using
bioinformatics analysis. A total of 12 RNAs (four lncRNAs, four
mRNAs and four circRNAs) were randomly selected and analyzed using
RT-qPCR, and the results were consistent with the microarray
analyses.
There were 661 differentially expressed lncRNAs (341
upregulated and 320 downregulated) identified between the AD+NS
mouse group and the AD mouse model treated with TG. Abnormal
regulation of lncRNAs may cause cancer, epilepsy, heart disease,
and neurodegenerative diseases (46-48).
In addition, an increasing number of lncRNAs have been associated
with the pathogenesis of AD. The innate immune system and
inflammatory signaling are critical for homeostasis, repair, and
neuroprotection. Excess oxygen free radicals and proinflammatory
cytokines trigger an inflammatory cascade that ultimately leads to
neurodegeneration (49).
LncRNA-cox-2 is located downstream of cyclo-oxygenase 2 (COX2) and
was reported to activate and inhibit the expression of various
immune genes in macrophages and regulate NF-κB, which in turn
affects aging and age-related diseases, including AD (50). In addition, the accumulation of
mitochondrial superoxide free radicals and transformation to
hydrogen peroxide can cause oxidative stress, the release of
cytochrome C and apoptosis, which are also important mechanisms of
AD (51). Several mitochondrial
lncRNAs, including LNCND5, LNCND6, and LNCCYTB, may be involved in
the regulation of mitochondrial genes and in maintaining normal
mitochondrial function, which has been shown to be associated with
neurodegenerative diseases (52).
In our previous study, multiple lncRNAs associated with the
pathogenesis of AD induced by LPS were detected. NONMMUT034127.2
and NONMMUT079254.1 were the most differentially expressed lncRNAs
in the LPS-induced AD mouse model (53). In the present study, it was found
that NONMMUG090228.1 and NONMMUG011123.2 were the most
differentially expressed lncRNAs in an AD mouse model treated with
TG, which indicated that these two lncRNAs may participate in the
development of AD. However, to confirm this hypothesis, functional
identification of the two lncRNAs in AD is necessary.
Four hub genes, Pou4f1, Egr2, Mag, and Nr4a1, were
identified in the PPI network. Notably, these genes were shown to
be involved in the formation, differentiation, maturation,
apoptosis, and autophagy of neurons. POU4F1 (POU Class 4 Homeobox
1) is an important molecule in the POU TF family and is expressed
in central neural precursor cells in the early stage of embryonic
development (54). The TF POU4F1
can bind to the promoter region of the Bcl-2 gene to regulate its
expression and interact with the P53 protein to regulate its
transcriptional activity (55).
Thus, the TF POU4F1 can promote the differentiation of the nervous
system and inhibit the apoptosis of nerve cells. In the present
study, Pou4f1 expression was significantly increased in the AD+TG
group, which indicated that TG may exert a neuroprotective effect
by inhibiting neuronal apoptosis via the regulation of Pou4f1.
The NR4A1 nuclear receptor belongs to the orphan
nuclear receptor subfamily NR4As. Studies have shown that NR4A1 is
involved in long-term memory formation (56). In the central nervous system, NR4A1
expression in the hippocampus increases after learning tasks
related to hippocampal memory (57). The NR4A1 receptor is related to a
variety of signaling molecules involved in memory formation, such
as ERK, CREB, and BDNF (58).
NR4A1 deficiency leads to late-phase long-term potentiation (L-LTP)
and impaired long-term memory formation (59). In the present study, Nr4a1
expression was significantly increased in the AD+TG group, which
indicated that TG may also improve memory in AD by regulating
Nr4a1.
EGR2 plays an important role in peripheral nerve
myelination, T-cell maturation, posterior brain segmentation, and
lipid biosynthesis (60), whereas
MAG plays an important role in maintaining myelin integrity and
inhibiting axon regeneration in the central nervous system
(61). Therefore, the results
indicate that TG may ameliorate AD by regulating Pou4f1, Egr2, Mag,
and Nr4a1 expression.
Furthermore, 64 differentially expressed circRNAs
(34 upregulated and 30 downregulated) were identified between the
AD+NS and AD+TG groups. circRNAs tend to accumulate in the aging
brain, as well as in cells with low proliferation rates (such as
neurons), especially neurons with synaptic development and
differentiation (30,62). Zhang et al (63) found that there were several
differentially expressed circRNAs in the brains of patients with
AD. Knockdown/overexpression of some of these differentially
expressed circRNAs was also assessed in AD cells and animal models
to reduce AD-like pathological manifestations, suggesting that
circRNAs may be involved in the regulation of AD pathology
(64). Neuronal injury and
apoptosis are the most intuitive pathological manifestations of
neurodegenerative diseases, such as AD (65). Abnormal deposition of Aβ,
neuroinflammation, oxidative stress injury, abnormal autophagy
levels, and other factors can lead to neuronal injury and apoptosis
(66,67). circRNAs are hypothesized to be
involved in the above pathogenesis, as well as in AD. In previous
studies, circHIPK2(68),
circ-0002468(69), circ
HDAC9(70), and
circ-0000950(71) were shown to be
involved in neuronal injury and to affect the pathogenesis and
pathological process of AD. Circ-7(72), hsa_circ RNA-405619, and hsa_circ
RNA-000843(73) were shown to
participate in Aβ metabolism. In addition, circNF1-419(74) and circHECTD1(75) were shown to participate in
autophagy and to affect the occurrence and development of AD. In
the present study, 64 differentially expressed circRNAs were
identified between the hippocampus of AD+NS mice and AD mice
treated with TG. mmu_CIRCpedia_35174 and mmu_circ_0007187 were the
most upregulated and downregulated circRNAs, respectively, which
indicated that TG may treat AD by regulating the expression of
these two circRNAs. However, the relationships of
mmu_CIRCpedia_35174 and mmu_circ_0007187 with AD remain to be
determined. Therefore, it is necessary to investigate the functions
of these two circRNAs in AD treated with TG in the future.
In summary, several dysregulated lncRNAs, mRNAs, and
circRNAs that may serve as potential biomarkers or targets in AD
treated with TG were identified. Future studies should elucidate
the detailed mechanisms underlying the regulation of the identified
differentially expressed lncRNAs, mRNAs, and circRNAs.
Supplementary Material
Network stats for the protein-protein
interaction network.
The top 10 enriched functional GO
terms.
The top 10 enriched Kyoto Encyclopedia
of Genes and Genomes pathways.
Acknowledgements
Not applicable.
Funding
Funding: This work was supported by the National Natural Science
Foundation of China (grant no. 81873780), The Changsha Outstanding
Innovative Young People Training Scheme (grant nos. kq2206058 and
kq2206056), The Foundation of Project of Hunan Health and Family
Planning Commission (grant no. 202202082739), The Foundation of the
Education Department of Hunan Province (grant nos. 21A0586 and
22A0662), The Foundation of the Education Department of Guangxi
Province (grant no. 2021KY1959); The Hunan Key Laboratory
Cultivation Base of the Research and Development of Novel
Pharmaceutical Preparations (grant no. 2016TP1029), and the
Application Characteristic Discipline of Hunan Province.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request. The relevant data can also be accessed via a public
repository (GEO series accession no. GSE204817; http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE204817).
Author's contributions
LT and JML designed the study, collected the data,
performed the initial analysis, and drafted the manuscript. YW and
QX aided in data acquisition, data analysis, and statistical
analysis. JX, YZ and DWY performed the literature search, assisted
with data acquisition, and edited the manuscript. YZ, JX and JML
reviewed the manuscript. QX, DWY and YZ confirm the authenticity of
all the raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The research procedures were approved by the Ethics
Committee of the Changsha Medical University, China (approval no.
EC20190114).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mucke L: Alzheimer's disease. Nature.
461:895–897. 2009.PubMed/NCBI View
Article : Google Scholar
|
|
2
|
Wenk GL: Neuropathologic changes in
Alzheimer's disease. J Clin Psychiatry. 64:7–10. 2003.PubMed/NCBI
|
|
3
|
Du Q, Zhu X and Si J: Angelica
polysaccharide ameliorates memory impairment in Alzheimer's disease
rat through activating BDNF/TrkB/CREB pathway. Exp Biol Med
(Maywood). 245:1–10. 2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Zhou B, Tan J, Zhang C and Wu Y:
Neuroprotective effect of polysaccharides from Gastrodia elata
blume against corticosterone-induced apoptosis in PC12 cells via
inhibition of the endoplasmic reticulum stress-mediated pathway.
Mol Med Rep. 17:1182–1190. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Wei M, Liu Y, Pi Z, Li S, Hu M, He Y, Yue
K, Liu T and Liu Z, Song F and Liu Z: Systematically characterize
the anti-Alzheimer's disease mechanism of lignans from S. chinensis
based on in-vivo ingredient analysis and target-network
pharmacology strategy by UHPLC-Q-TOF-MS. Molecules.
24(1203)2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Guo X, Ye YJ, Song K, An LP and Sheng Y:
Mechanism of schisandrae chinensis fructus lignans in alleviating
learning and memory ability in D-galactose aging mice. Chin J Exp
Trad Med Formulae. 26:85–91. 2020.
|
|
7
|
Wu J, Qu JQ, Zhou YJ, Zhou YJ, Li YY,
Huang NQ, Deng CM and Luo Y: Icariin improves cognitive deficits by
reducing the deposition of β-amyloid peptide and inhibition of
neurons apoptosis in SAMP8 mice. Neuroreport. 31:663–671.
2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Li Z, Zhang XB, Gu JH, Zeng YQ and Li JT:
Breviscapine exerts neuroprotective effects through multiple
mechanisms in APP/PS1 transgenic mice. Mol Cell Biochem. 468:1–11.
2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Chen Y, Chen Y, Liang Y, Chen H, Ji X and
Huang M: Berberine mitigates cognitive decline in an Alzheimer's
disease mouse model by targeting both tau hyperphosphorylation and
autophagic clearance. Biomed Pharmacother.
121(109670)2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Quan QK, Li X, Feng JJ, Hou JX, Li M and
Zhang BW: Ginsenoside Rg1 reduces β-amyloid levels by inhibiting
CDΚ5-induced PPARγ phosphorylation in a neuron model of Alzheimer's
disease. Mol Med Rep. 22:3277–3288. 2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Zhang B, Li Q, Chu X, Sun S and Chen S:
Salidroside reduces tau hyperphosphorylation via up-regulating
GSK-3β phosphorylation in a tau transgenic Drosophila model of
Alzheimer's disease. Transl Neurodegener. 5:1–6. 2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Bao J, Liu W, Zhou HY, Gui YR, Yan YH, Wu
MJ, Xiao YF, Shang JT, Long GF and Shu XJ:
Epigallocatechin-3-gallate alleviates cognitive deficits in APP/PS1
mice. Curr Med Sci. 40:18–27. 2020.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Hase T, Shishido S, Yamamoto S, Yamashita
R, Nukima H, Taira S, Toyoda T, Abe K, Hamaguchi T, Ono K, et al:
Rosmarinic acid suppresses Alzheimer's disease development by
reducing amyloid β aggregation by increasing monoamine secretion.
Sci Rep. 9:1–13. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Luo J, Song W, Xu Y, Chen GY, Hu Q and Tao
QW: Benefits and safety of tripterygium glycosides and total
glucosides of paeony for rheumatoid arthritis: An overview of
systematic reviews. Chin J Integr Med. 25:696–703. 2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Wu X, Huang Y, Zhang Y, He C, Zhao Y, Wang
L and Gao J: Efficacy of tripterygium glycosides combined with ARB
on diabetic nephropathy: A meta-analysis. Biosci Rep.
40(BSR20202391)2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Li T, Zhou HC and Chen J: Effects of
tripterygium wilfordii polyglycosides on the proliferation,
apoptosis and PI3K/AKT signaling pathway of human oral cancer KB
cells. J Guangdong Coll Pharm. 35:653–657. 2019.(In Chinese).
|
|
17
|
Ma CL, Zhang BL and Liu XM: Effects of
tripterygium glycosides on migration and angiogenesis of lung
adenocarcinoma cell by inhibiting PI3K/AKT signaling. Anhui Med
Pharm J. 26:235–238. 2022.(In Chinese).
|
|
18
|
Wang M, Chen TG, Yang XL, Zhang DL, Zhou
KS, Nan W and Zhang HH: Effect of tripterygium glycosides on
inflammatory factors induced by lipopolysaccharide in rat
astrocytes. Chin J Clin Pharmacol. 35:154–158. 2019.(In
Chinese).
|
|
19
|
Tang L, Xiang Q, Xiang J, Zhang Y and Li
J: Tripterygium glycoside ameliorates neuroinflammation in a mouse
model of Aβ25-35-induced Alzheimer's disease by
inhibiting the phosphorylation of IκBα and p38. Bioengineered.
12:8540–8554. 2021.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Sun X and Malhotra A: Noncoding RNAs
(ncRNA) in hepato cancer: A review. J Environ Pathol Toxicol Oncol.
37:15–25. 2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Faghihi M, Modarresi F, Khalil AM, Wood
DE, Sahagan BG, Morgan TE, Finch CE, Laurent GS III, Kenny PJ and
Wahlestedt C: Expression of a noncoding RNA is elevated in
Alzheimer's disease and drives rapid feed-forward regulation of
β-secretase. Nat Med. 14:723–730. 2008.PubMed/NCBI View
Article : Google Scholar
|
|
22
|
Massone S, Vassallo I, Fiorino G,
Castelnuovo M, Barbieri F, Borghi R, Tabaton M, Robello M, Gatta E,
Russo C, et al: 17A, a novel non-coding RNA, regulates GABA B
alternative splicing and signaling in response to inflammatory
stimuli and in Alzheimer disease. Neurobiol Dis. 41:308–317.
2011.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Massone S, Ciarlo E, Vella S, Nizzari M,
Florio T, Russo C, Cancedda R and Pagano A: NDM29, a RNA polymerase
III-dependent non coding RNA, promotes amyloidogenic processing of
APP and amyloid β secretion. Biochim Biophys Acta. 1823:1170–1177.
2012.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Ciarlo E, Massone S, Penna I, Nizzari M,
Gigoni A, Dieci G, Russo C, Florio T, Cancedda R and Pagano A: An
intronic ncRNA-dependent regulation of SORL1 expression affecting
Aβ formation is upregulated in post-mortem Alzheimer's disease
brain samples. Dis Model Mech. 6:424–433. 2013.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Mus E, Hof PR and Tiedge H: Dendritic
BC200 RNA in aging and in Alzheimer's disease. Proc Natl Acad Sci
USA. 104:10679–10684. 2007.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Parenti R, Paratore S, Torrisi A and
Cavallaro S: A natural antisense transcript against Rad18,
specifically expressed in neurons and upregulated during
beta-amyloid-induced apoptosis. Eur J Neurosci. 26:2444–2457.
2007.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Verduci L, Tarcitano E, Strano S, Yarden Y
and Blandino G: CircRNAs: Role in human diseases and potential use
as biomarkers. Cell Death Dis. 12:1–12. 2021.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Dube U, Del-Aguila JL, Li Z, Budde JP,
Jiang S, Hsu S, Ibanez L, Fernandez MV, Farias F, Norton J, et al:
An atlas of cortical circular RNA expression in Alzheimer disease
brains demonstrates clinical and pathological associations. Nat
Neurosci. 22:1903–1912. 2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Ma N, Pan J, Ye X, Yu B, Zhang W and Wan
J: Whole-transcriptome analysis of APP/PS1 mouse brain and
identification of circRNA-miRNA-mRNA networks to investigate AD
pathogenesis. Mol Ther Nucleic Acids. 18:1049–1062. 2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Gruner H, Cortes-Lopez M, Cooper DA, Bauer
M and Miura P: CircRNA accumulation in the aging mouse brain. Sci
Rep. 6(38907)2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
National Research Council (US) Institute
for Laboratory Animal Research. Guide for the Care and Use of
Laboratory Animals. National Academies Press, Washington, DC,
1996.
|
|
32
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43(e47)2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
R Core Team: R: A language and environment
for statistical computing. R Foundation for Statistical Computing,
Vienna, 2012. http://www.R-project.org/.
|
|
34
|
RStudio Team: RStudio, Integrated
Development for R. RStudio, Inc., Boston MA, 2015. http://www.rstudio.com/.
|
|
35
|
Saeed AI, Sharov V, White J, Li J, Liang
W, Bhagabati N, Braisted J, Klapa M, Currier T, Thiagarajan M, et
al: TM4: A free, open-source system for microarray data management
and analysis. Biotechniques. 34:374–378. 2003.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Krzywinski M, Schein J, Birol I, Connors
J, Gascoyne R, Horsman D, Jones SJ and Marra MA: Circos: An
information aesthetic for comparative genomics. Genome Res.
19:1639–1645. 2009.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Szklarczyk D, Gable AL, Lyon D, Junge A,
Wyder S, Huerta-Cepas J, Simonovic M, Doncheva NT, Morris JH, Bork
P, et al: STRING v11: protein-protein association networks with
increased coverage, supporting functional discovery in genome-wide
experimental datasets. Nucleic Acids Res. 47:D607–D613.
2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Ashburner M, Ball C, Blake J, Botstein D,
Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, et
al: Gene ontology: Tool for the unification of biology. The gene
ontology consortium. Nat Genet. 25:25–29. 2000.PubMed/NCBI View Article : Google Scholar
|
|
40
|
The Gene Ontology Consortium. The gene
ontology resource: 20 years and still GOing strong. Nucleic Acids
Res. 47:D330–D338. 2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Kanehisa M: Post-genome Informatics.
Oxford University press Inc., New York, NY, 2000.
|
|
42
|
Wucher V, Legeai F, Hedan B, Rizk G,
Lagoutte L, Leeb T, Jagannathan V, Cadieu E, David A, Lohi H, et
al: FEELnc: A tool for long non-coding RNA annotation and its
application to the dog transcriptome. Nucleic Acids Res.
45(e57)2017.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Alkan F, Wenzel A, Palasca O, Kerpedjiev
P, Rudebeck AF, Stadler PF, Hofacker IL and Gorodkin J: RIsearch2:
Suffix array-based large-scale prediction of RNA-RNA interactions
and siRNA off-targets. Nucleic Acids Res. 45(e60)2017.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Wenzel A, Akbasli E and Gorodkin J:
RIsearch: Fast RNA-RNA interaction search using a simplified
nearest-neighbor energy model. Bioinformatics. 28:2738–2746.
2012.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Shen Y, Peng X and Shen C: Identification
and validation of immune-related lncRNA prognostic signature for
breast cancer. Genomics. 112:2640–2646. 2020.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Luo ZH, Walid AA, Xie Y, Long H, Xiao W,
Xu L, Fu Y, Feng L and Xiao B: Construction and analysis of a
dysregulated lncRNA-associated ceRNA network in a rat model of
temporal lobe epilepsy. Seizure. 69:105–114. 2019.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Wei CW, Luo T, Zou SS and Wu AS: The role
of long noncoding RNAs in central nervous system and
neurodegenerative diseases. Front Behav Neurosci.
12(175)2018.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Singh A, Kukreti R, Saso L and Kukreti S:
Oxidative stress: A key modulator in neurodegenerative diseases.
Molecules. 24(1583)2019.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Yang Z, Jiang S, Shang J, Jiang Y, Dai Y,
Xu B, Yu Y, Liang Z and Yang Y: LncRNA: Shedding light on
mechanisms and opportunities in fibrosis and aging. Ageing Res Rev.
52:17–31. 2019.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Swerdlow RH: Mitochondria and
mitochondrial cascades in Alzheimer's disease. J Alzheimers Dis.
62:1403–1416. 2018.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Hezroni H, Perry RBT and Ulitsky I: Long
noncoding RNAs in development and regeneration of the neural
lineage. Cold Spring Harb Symp Quant Biol. 84:165–177.
2019.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Tang L, Liu L, Li G, Jiang P, Wang Y and
Li J: Expression profiles of long noncoding RNAs in intranasal
LPS-mediated Alzheimer's disease model in mice. BioMed Res Int.
2019(9642589)2019.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Deng M, Yang H, Xie X, Liang G and Gan L:
Comparative expression analysis of POU4F1, POU4F2 and ISL1 in
developing mouse cochleovestibular ganglion neurons. Gene Expr
Patterns. 15:31–37. 2014.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Yadav R and Srivastava P: Establishment of
resveratrol and its derivatives as neuroprotectant against
monocrotophos-induced alteration in NIPBL and POU4F1 protein
through molecular docking studies. Environ Sci Pollut Res Int.
27:291–304. 2020.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Jeanneteau F, Barrère C, Vos M, De Vries
CJM, Rouillard C, Levesque D, Dromard Y, Moisan MP, Duric V,
Franklin TC, et al: The stress-induced transcription factor NR4A1
adjusts mitochondrial function and synapse number in prefrontal
cortex. J Neurosci. 38:1335–1350. 2018.PubMed/NCBI View Article : Google Scholar
|
|
57
|
McNulty SE, Barrett RM, Vogel-Ciernia A,
Malvaez M, Hernandez N, Davatolhagh MF, Matheos DP, Schiffman A and
Wood MA: Differential roles for Nr4a1 and Nr4a2 in object location
vs. object recognition long-term memory. Learn Mem. 19:588–592.
2012.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Zhang Z and Yu J: NR4A1 promotes cerebral
ischemia reperfusion injury by repressing Mfn2-mediated mitophagy
and inactivating the MAPK-ERK-CREB signaling pathway. Neurochem
Res. 43:1963–1977. 2018.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Chen YL, Wang Y, Erturk A, Kallop D, Jiang
Z, Weimer RM, Kaminker J and Sheng M: Activity-induced Nr4a1
regulates spine density and distribution pattern of excitatory
synapses in pyramidal neurons. Neuron. 83:431–443. 2014.PubMed/NCBI View Article : Google Scholar
|
|
60
|
LeBlanc SE, Srinivasan R, Ferri C, Mager
GM, Gillian-Daniel AL, Wrabetz L and Svaren J: Regulation of
cholesterol/lipid biosynthetic genes by Egr2/Krox20 during
peripheral nerve myelination. J Neurochem. 93:737–748.
2005.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Llorens F, Gil V and del Río JA: Emerging
functions of myelin-associated proteins during development,
neuronal plasticity, and neurodegeneration. FASEB J. 25:463–475.
2011.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Wang Z, Xu P, Chen B, Zhang Z, Zhang C,
Zhan Q, Huang S, Xia ZA and Peng W: Identifying
circRNA-associated-ceRNA networks in the hippocampus of
Aβ1-42-induced Alzheimer's disease-like rats using
microarray analysis. Aging (Albany NY). 10:775–788. 2018.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Zhang Y, Yu F, Bao S and Sun J: Systematic
characterization of circular RNA-associated CeRNA network
identified novel circRNA biomarkers in Alzheimer's disease. Front
Bioeng Biotechnol. 7(222)2019.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Castillo E, Leon J, Mazzei G, Abolhassani
N, Haruyama N, Saito T, Saido T, Hokama M, Iwaki T, Ohara T, et al:
Comparative profiling of cortical gene expression in Alzheimer's
disease patients and mouse models demonstrates a link between
amyloidosis and neuroinflammation. Sci Rep. 7(17762)2017.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Fielder E, Von Zglinicki T and Jurk D: The
DNA damage response in neurons: Die by apoptosis or survive in a
senescence-like state? J Alzheimer's Dis. 60:S107–S131.
2017.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Coimbra-Costa D, Alva N, Duran M, Carbonel
T and Rama R: Oxidative stress and apoptosis after acute
respiratory hypoxia and reoxygenation in rat brain. Redox Biol.
12:216–225. 2017.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Galluzzi L, Pedro JMBS, Blomgren K and
Kroemer G: Autophagy in acute brain injury. Nat Rev Neurosci.
17:467–484. 2016.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Wang G, Han B, Shen L, Wu S, Yang L, Liao
J, Wu F, Li M, Leng S, Zang F, et al: Silencing of circular RNA
HIPK2 in neural stem cells enhances functional recovery following
ischaemic stroke. EBioMedicine. 52(102660)2020.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Yang M, Xiang G, Yu D, Yang G, He W, Yang
S, Zhou G and Liu A: Hsa_circ_0002468 regulates the neuronal
differentiation of SH-SY5Y cells by modulating the MiR-561/E2F8
axis. Med Sci Monit. 25:2511–2519. 2019.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Zhang N, Gao Y, Yu S, Sun XH and Shen K:
Berberine attenuates Aβ42-induced neuronal damage through
regulating circHDAC9/miR-142-5p axis in human neuronal cells. Life
Sci. 252(117637)2020.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Yang H, Wang H, Shang H, Chen X, Yang S,
Qu Y, Ding J and Li X: Circular RNA circ_0000950 promotes neuron
apoptosis, suppresses neurite outgrowth and elevates inflammatory
cytokines levels via directly sponging miR-103 in Alzheimer's
disease. Cell Cycle. 18:2197–2214. 2019.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Shi Z, Chen T, Yao Q, Zheng L, Zhang Z,
Wang J, Hu Z, Cui H, Han Y, Han X, et al: The circular RNA cirs-7
promotes APP and BACE 1 degradation in an NF-κB-dependent manner.
FEBS J. 284:1096–1109. 2017.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Li Y, Lv Z, Zhang J, Ma Q, Li Q, Song L,
Gong L, Zhu Y, Li X, Hao Y and Yang Y: Profiling of differentially
expressed circular RNAs in peripheral blood mononuclear cells from
Alzheimer's disease patients. Metab Brain Dis. 35:201–213.
2020.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Chen D, Guo Y, Qi L, Tang X, Liu Y, Yang
X, Hu GY, Shuai Q, Yong Y, Wang D, et al: Circular RNA NF1-419
enhances autophagy to ameliorate senile dementia by binding
Dynamin-1 and adaptor protein 2 B1 in AD-like mice. Aging (Albany
NY). 11:12002–12031. 2019.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Han B, Zhang Y, Zhang Y, Bai Y, Chen XF,
Huang R, Wu FF, Shou L, Chao J, Zhang J, et al: Novel insight into
circular RNA HECTD1 in astrocyte activation via autophagy by
targeting MIR142-TIPARP: Implications for cerebral ischemic stroke.
Autophagy. 14:1164–1184. 2018.PubMed/NCBI View Article : Google Scholar
|