Introduction
Hemangioma is the most common type of benign tumor
in infants and children and can affect physical appearance and
quality of life and may cause functional impairment (1). The incidence of hemangioma in infants
is 2-3% (statistics from China, the US, and Germany), and this
condition is more prevalent in females compared with males
(approximately 2 to 3 times higher) (2-5).
The first-line treatments for hemangioma include propranolol,
corticosteroids, β-blockers, topical timolol and pulsed dye laser
therapy (6-8).
However, nearly 10-15% of patients do not respond to treatments and
sclerotherapy can be offered as a subsequent treatment due to its
efficacy, safety, affordability and non-invasive nature in patients
with hemangioma (6,9-14).
Bleomycin (BLM) and pingyangmycin (PYM) are drugs used in
sclerotherapy that act by preventing DNA replication to interfere
with cell division and proliferation (15-17).
However, treatment outcomes of BLM and PYM may not be effective in
certain patients with hemangioma, which may be due to BLM and PYM
resistance in these patients (18,19).
For example, a previous study reported that 26% of patients with
hemangioma receiving PYM do not achieve a cure, defined as the
lesion disappearing completely without recurrence at least 1 year
after treatment (19). In
addition, another study reported that 45.5% of patients with
hemangioma do not exhibit a complete response to BLM and 18.2% of
these patients show no response to BLM treatment (18). Thus, exploring potential strategies
to improve BLM and PYM sensitivity is key to enhance the management
of symptoms of patients with hemangioma.
microRNA (miR)-203a-3p is located on the 14q32.33
chromosome and contributes to the progression of certain types of
malignant tumor, such as cervical, colorectal and thyroid papillary
cancer (20-22).
Our recent studies reported the involvement of miR-203a-3p in
hemangioma (23,24). miR-203a-3p silencing regulates the
long non-coding RNA maternally expressed 8-mediated Notch pathway
to increase hemangioma cell proliferation and invasion whilst
decreasing apoptosis (23).
Furthermore, miR-203a-3p overexpression inactivates vascular
endothelial growth factor A (VEGFA)-regulated PI3K/AKT pathway
signaling to inhibit hemangioma cell proliferation and invasion, as
well as increase apoptosis (24).
A number of studies have reported the involvement of the PI3K/AKT
pathway in sensitizing hemangioma cells to BLM and PYM (25,26).
For example, the PI3K/AKT pathway inhibits the anti-tumor effect of
PYM on hemangioma cell viability, apoptosis and invasion (26). In addition, inhibition of AKT
activity increases sensitivity to BLM, which attenuates hemangioma
progression (25). Considering the
regulatory role of miR-203a-3p on the PI3K/AKT pathway in
hemangioma progression and the involvement of the PI3K/AKT pathway
in BLM and PYM resistance (24-26),
miR-203a-3p may have the potential to improve sensitivity to BLM
and PYM.
The present study aimed to explore the influence of
miR-203a-3p on hemangioma cell sensitivity to BLM and PYM
treatment, in addition to analyzing potential effects on the
PI3K/AKT pathway.
Materials and methods
Cell culture
Human hemangioma endothelial cells (HemECs) were
purchased from the BeNa Culture Collection, sourced the cells from
Otwo Biotech (cat. no. HTX2171). HemECs were cultured in
endothelial cell medium (Gibco; Thermo Fisher Scientific, Inc.)
with 10% fetal bovine serum (HyClone; Cytiva) and 1%
penicillin/streptomycin (Sangon Biotech Co., Ltd.) in 5%
CO2 at 37˚C. The present study was approved by the
Ethics Committee of the Affiliated Hospital of Hebei University of
Engineering [Handan, China; approval no. 2019(K)016].
Cell transfection
miR-203a-3p (5'-GTGAAATGTTTAGGACCACTAG-3') and
negative control (NC) mimics (5'-CAGTACTTTTGTGTAGTACAA-3') were
purchased from Changchun Changsheng Gene Pharmaceutical Co., Ltd.
HemECs were seeded in 6-well plates (3x105 cells/well)
and incubated overnight at 37˚C, and then incubated with 50 nM
miR-mimic or NC-mimic for 48 h using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) at 37˚C.
Reverse transcription-quantitative PCR
(RT-qPCR)
Following transfection, HemECs were collected for
detection of miR-203a-3p expression with RT-qPCR as previously
described (24). In brief, total
RNA was isolated using the RNeasy Mini kit (Qiagen GmbH). Then, the
High-Capacity cDNA Reverse Transcription kit (Thermo Fisher
Scientific, Inc.) was used for RT according to the kit's
instructions. qPCR was performed using SYBR® Green PCR
Master Mix (Thermo Fisher Scientific, Inc.). The thermocycling
conditions were as follows: Initial denaturation at 95˚C for 3 min,
followed by 40 cycles of 95˚C for 10 sec and 61˚C for 30 sec (this
was a two-step PCR, and the annealing and extension steps were
combined). The primer sequences were as follows: miR-203a-3p
forward (F), 5'-ACACTCCAGCTGGGGTGAAATGTTTAGGAC-3' (adapter
sequence, ACACTCCAGCTGGG) and reverse (R),
5'-TGTCGTGGAGTCGGCAATTC-3'; and U6 F, 5'-GCTCGCTTCGGCAGCACATA-3'
and R 5'-AATATGGAACGCTTCACGAATTTGC-3' (23,27,28).
The result was analyzed with 2-ΔΔCq method (29).
BLM and PYM treatment
Sensitivity of HemECs to BLM and PYM (both
MedChemExpress) was measured using Cell Counting Kit-8
(MilliporeSigma). In brief, transfected HemECs were seeded into
96-well plates (3x103 cells/well) and incubated with 0,
4, 8, 12, 16 or 20 µM BLM or PYM for 24 h at 37˚C. The
concentrations of BLM and PYM were determined according to
preliminary experiments. Next, HemECs were incubated with CCK-8
buffer for 2 h at 37˚C. The optical density (450 nm) of samples was
recorded using an enzyme immunoassay analyzer (BioTek China) and
used for calculating cell viability at various treatment
concentrations. Half-maximal inhibitory concentration
(IC50) of BLM and PYM was calculated using sigmoidal
dose-response function (30).
Transfected HemECs were seeded in 6-well plates
(3x105 cells/well) and incubated overnight at 37˚C. BLM
(14 µM) and PYM (12 µM) were added, then cells were incubated for
24 h at 37˚C. The concentration of BLM and PYM was determined
according to the IC50. EdU staining, annexin V-FITC/PI
apoptosis assay and western blotting assays of apoptotic proteins
were subsequently performed.
740 Y-P treatment
HemECs were transfected with miR-mimic or NC-mimic
as aforementioned, then stimulated with 740 Y-P (20 µM; APeXBIO
Technology LLC), a cell-permeable phosphopeptide activator of PI3K
(31), for 24 h at 37˚C to
investigate the potential effects of miR-203a-3p on the PI3K/AKT
pathway. The phosphorylated level of PI3K and AKT were assessed
with western blotting and the drug sensitivity of cells to BLM and
PYM were measured as aforementioned. In addition, after stimulating
with 740 Y-P, HemECs were incubated with BLM (14 µM) and PYM (12
µM) for 24 h at 37˚C and EdU staining, annexin V-FITC/PI apoptosis
assays and western blotting assays of apoptotic proteins were
performed.
EdU staining
EdU staining was performed to analyze cell
proliferation using the BeyoClick™ EdU kit (Beyotime Institute of
Biotechnology). Briefly, treated HemECs (as described in ‘BLM and
PYM treatment’ and ‘740 Y-P treatment’ subsections) were seeded in
6-well plates (3x105 cells/well) and incubated overnight
at 37˚C. EdU (10 µM) was added to each well and cells were
incubated for 2 h at 25˚C. HemECs were fixed in 4% paraformaldehyde
at room temperature, permeabilized by 0.3% Triton X-100 and stained
with Click Additive Solution (Beyotime Institute of Biotechnology)
according to the manufacturer's protocol. Cells were imaged using a
fluorescence microscope (Olympus Corp.) under a magnification of
x200. The proportion of EdU-positive cells was calculated as
follows: (Red fluorescent spot number/blue fluorescence spot
number) x100.
Apoptosis assay
The apoptosis rate of HemECs was measured using
Annexin V-FITC/PI kit (Beyotime Institute of Biotechnology). In
brief, treated HemECs were washed and resuspended in Annexin V-FITC
binding buffer (5x105 cells/195 µl). Annexin V-FITC (5
µl) and PI (10 µl) were added for 10 min at 37˚C. Flow cytometric
detection was performed within 1 h using a FACSCanto II (BD
Biosciences). The data was analyzed by FlowJo 10.0.7 (Becton,
Dickinson and Co.).
Western blot analysis
HemECs were lysed using precooled RIPA reagent
(MilliporeSigma) for protein extraction. The proteins were
quantified by bicinchoninic acid kit (Beyotime Institute of
Biotechnology), 20 µg of which were separated by 4-20% SDS-PAGE and
transferred onto nitrocellulose membranes (Pall Life Sciences).
Following blocking with 5% BSA (MilliporeSigma) at 37˚C for 1.5 h,
membranes were incubated with primary antibodies overnight at 4˚C,
then incubated with secondary antibodies for 1 h at 37˚C. ECL-PLUS
reagent (Thermo Fisher Scientific, Inc.) was used for visualization
of protein bands. The protein bands were analyzed by Image J 1.8
(National Institutes of health). The specific antibodies were as
follows: BCL2, cleaved caspase 3, phosphorylated (p)-PI3K, PI3K,
p-AKT, AKT (all 1:1,000 dilution; cat. nos. PA5-22209, PA5-114687,
PA5-105113, MA5-14870, 44-621G and MA5-14916; all Invitrogen;
Thermo Fisher Scientific, Inc.), GAPDH (1:5,000 dilution; cat. no.
GB15004-100) and Horseradish Peroxidase conjugated goat anti-rabbit
secondary antibodies (1:20,000 dilution; cat. no. GB23303; both
Wuhan Servicebio Technology Co., Ltd.).
Statistical analysis
The data in triplet were presented as mean ±
standard deviation and analyzed using GraphPad Prism 7.0
(Dotmatics). One-way ANOVA with Tukey's post-hoc test was used to
analyze statistical significance between groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-203a-3p enhanced HemEC sensitivity
to BLM and PYM
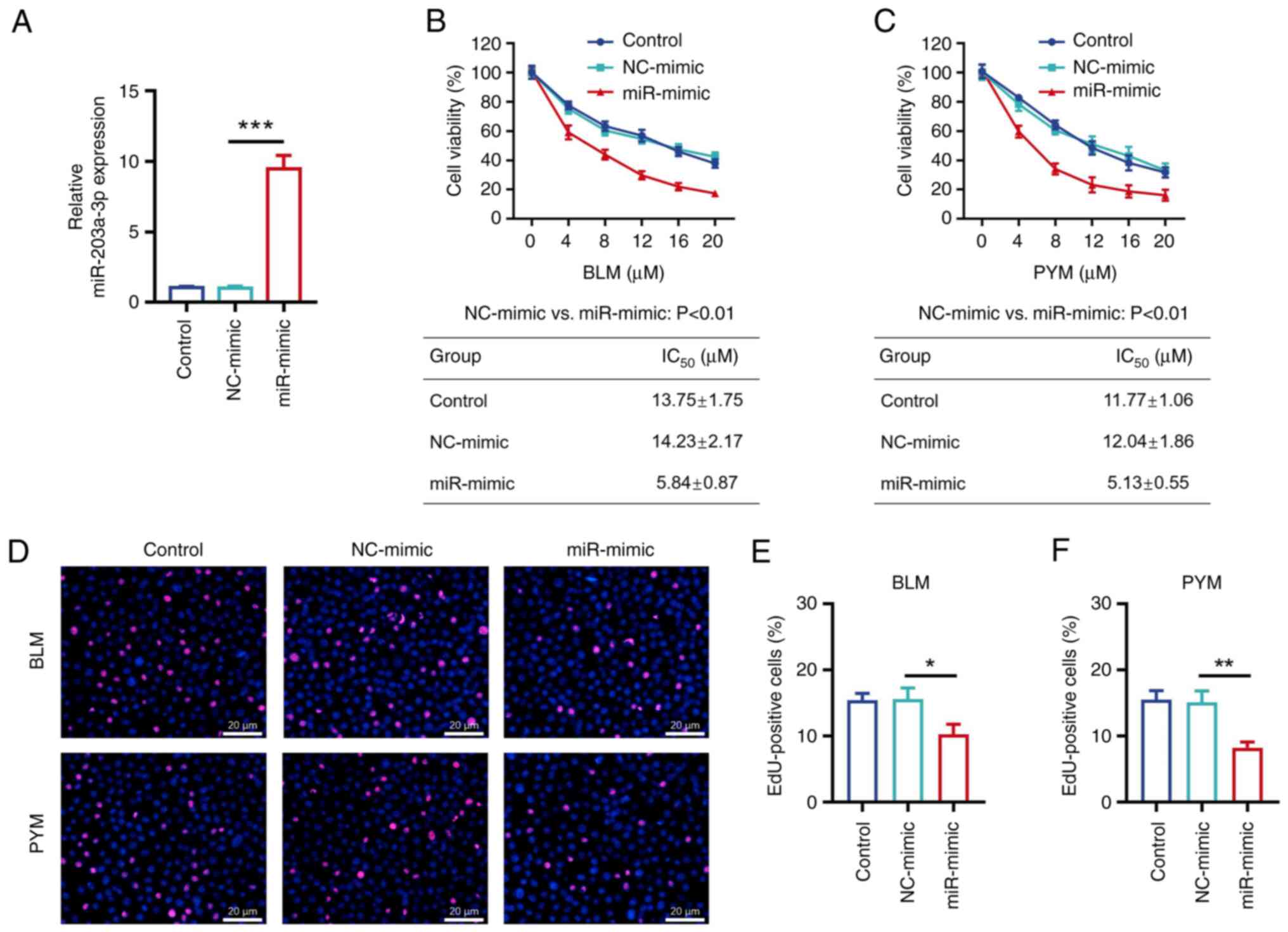
The relative miR-203a-3p expression levels were
significantly increased in the miR-mimic group compared with the
NC-mimic group (P<0.001; Fig.
1A), which demonstrated that the transfection was
successful.
CCK-8 assay demonstrated that relative cell
viability was decreased by BLM or PYM treatment in a dose-dependent
manner (Fig. 1B and C). IC50 values of BLM
(5.84±0.87 vs. 14.23±2.17 µM; P<0.01; Fig. 1B) and PYM (5.13±0.55 vs. 12.04±1.86
µM; P<0.01; Fig. 1C) were
significantly decreased in the miR-mimic group compared with the
NC-mimic group.
EdU staining was performed after cells were treated
with the IC50 dose of BLM (14 µM) or PYM (12 µM;
Fig. 1D); proportion of
EdU-positive cells were significantly decreased in the miR-
compared with the NC-mimic cells treated with BLM (P<0.05;
Fig. 1E) or PYM (P<0.01;
Fig. 1F).
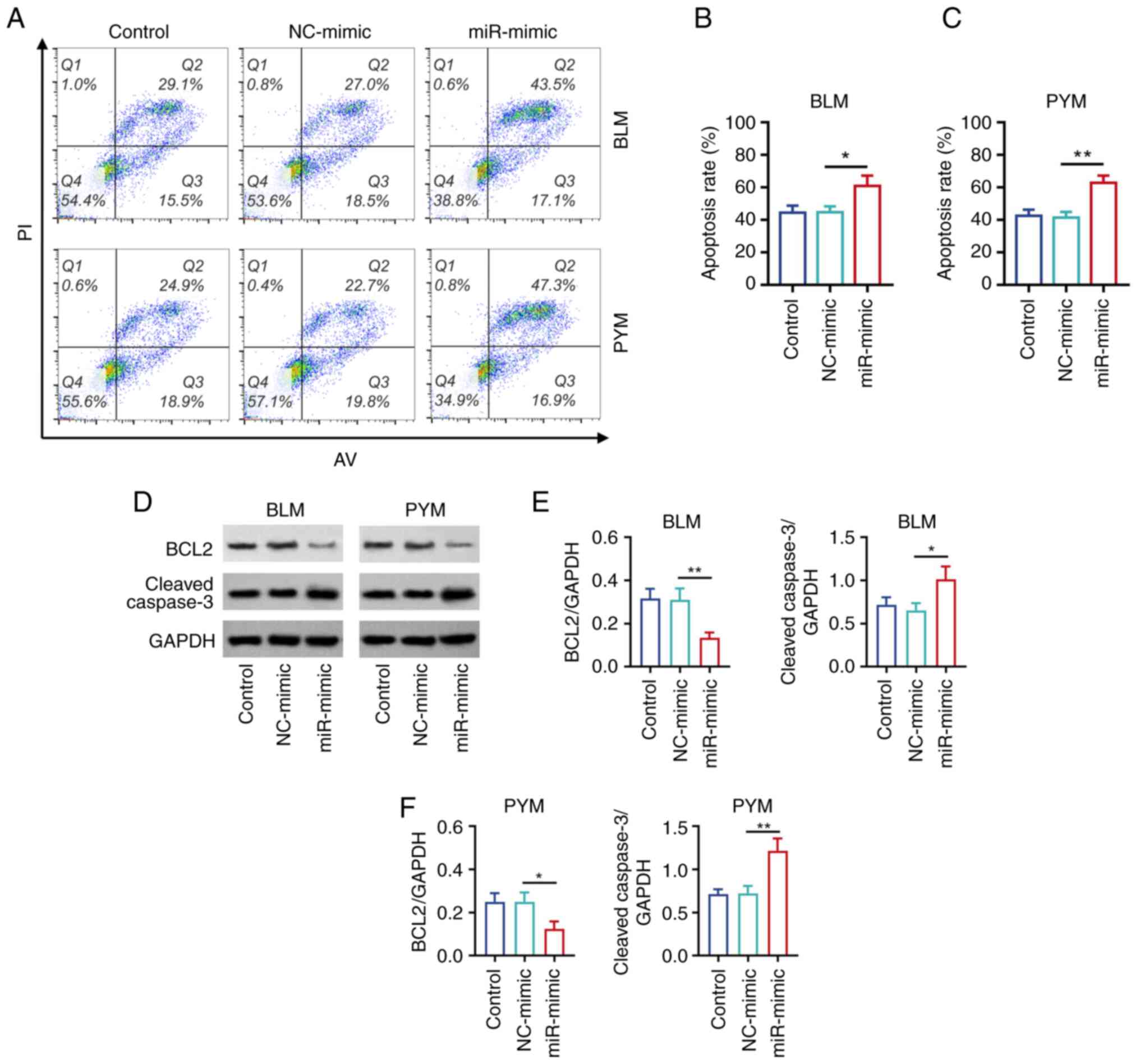
Annexin V-FITC/PI kit was used to detect HemEC
apoptosis in cells treated with BLM or PYM (Fig. 2A). The apoptosis rate was
significantly increased in the miR-mimic group compared with the
NC-mimic group treated with BLM (P<0.05; Fig. 2B) or PYM (P<0.01; Fig. 2C). Western blotting (Fig. 2D) demonstrated that protein
expression levels of apoptotic marker BCL2 were significantly
decreased, but cleaved caspase 3 protein expression levels were
significantly increased in the miR- compared with the NC-mimic
groups of cells treated with BLM (Fig.
2E) or PYM (all P<0.05; Fig.
2F).
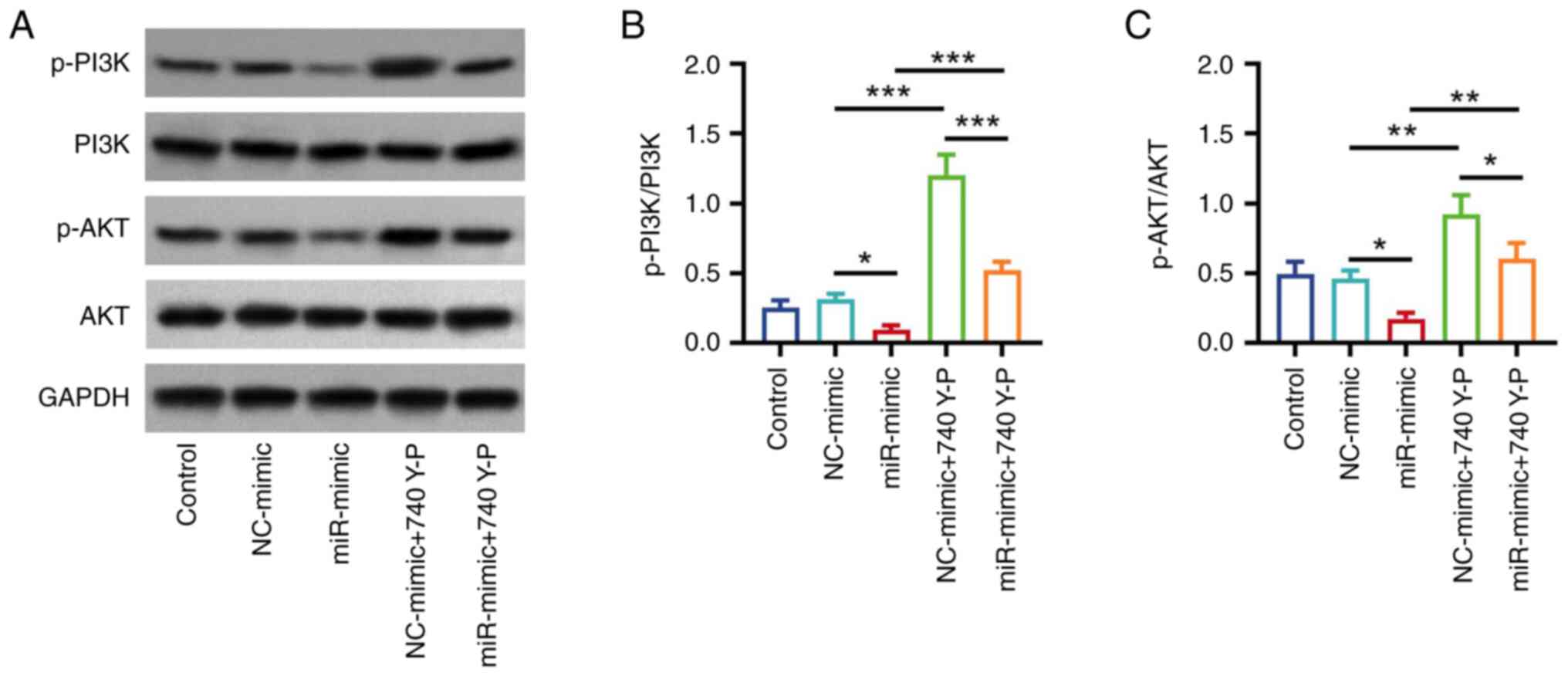
740 Y-P attenuated the inhibition of
miR-203a-3p on the PI3K/AKT pathway in HemECs
Our previous study reported that miR-203a-3p
inhibits the PI3K/AKT pathway to facilitate hemangioma progression
(24). Therefore, the effect of
740 Y-P, an activator of PI3K, was analyzed in the present study.
Western blotting (Fig. 3A)
demonstrated that p-PI3K/PI3K and p-AKT/AKT protein expression
levels were significantly elevated in the NC-mimic + 740 Y-P group
compared with the NC-mimic group (both P<0.01; Fig. 3B). Furthermore, p-PI3K/PI3K and
p-AKT/AKT protein expression levels were significantly increased in
the miR-mimic + 740 Y-P group compared with the miR-mimic group
(P<0.01; Fig. 3C).
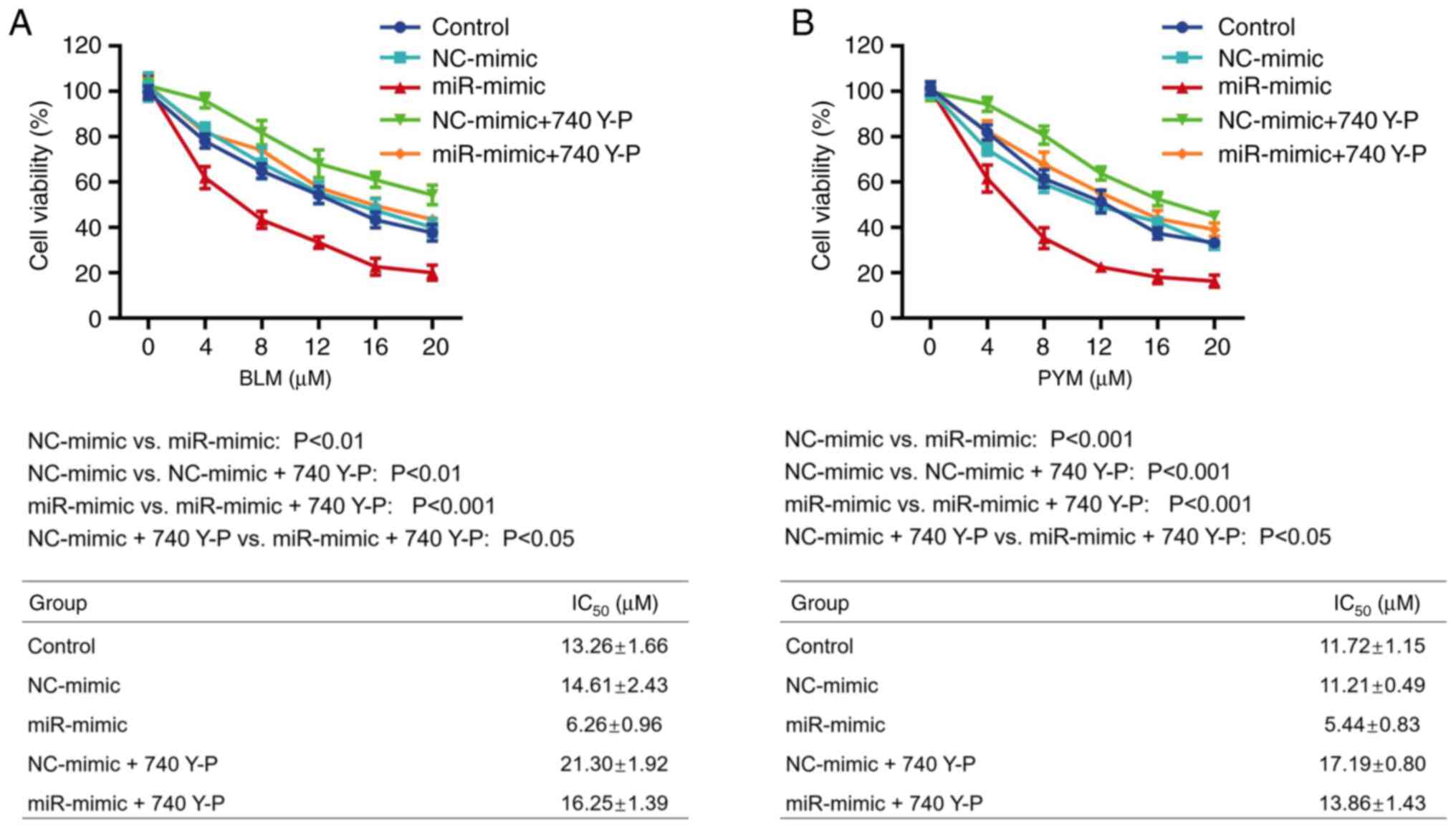
740 Y-P attenuated the role of
miR-203a-3p in enhancing HemEC sensitivity to BLM and PYM
CCK-8 assay demonstrated that the IC50
value of BLM was significantly increased in the NC-mimic + 740 Y-P
compared with the NC-mimic group (P<0.01). Furthermore,
IC50 value of BLM was significantly increased in the
miR-mimic + 740 Y-P group compared with the miR-mimic group
(P<0.001; Fig. 4A). The same
trend was demonstrated in HemECs treated with PYM (both P<0.001;
Fig. 4B).
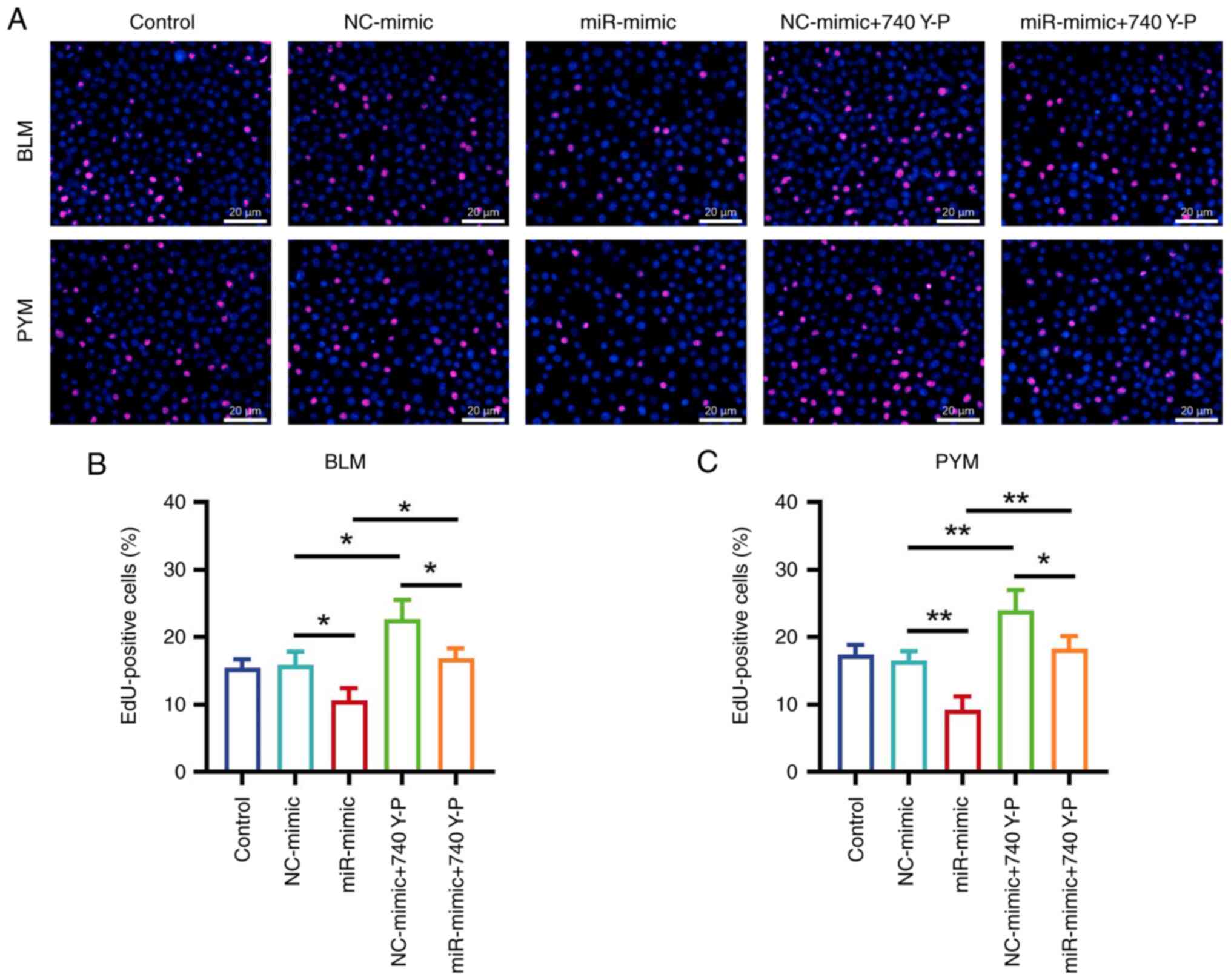
EdU staining (Fig.
5A) demonstrated that, when treated with BLM, the proportion of
EdU-positive cells significantly increased in the NC-mimic + 740
Y-P group compared with the NC-mimic group (P<0.05; Fig. 5B). Additionally, the proportion of
EdU-positive cells was significantly elevated in the miR-mimic +
740 Y-P group compared with the miR-mimic group (P<0.05). The
same trend was demonstrated in HemECs treated with PYM (both
P<0.01; Fig. 5C).
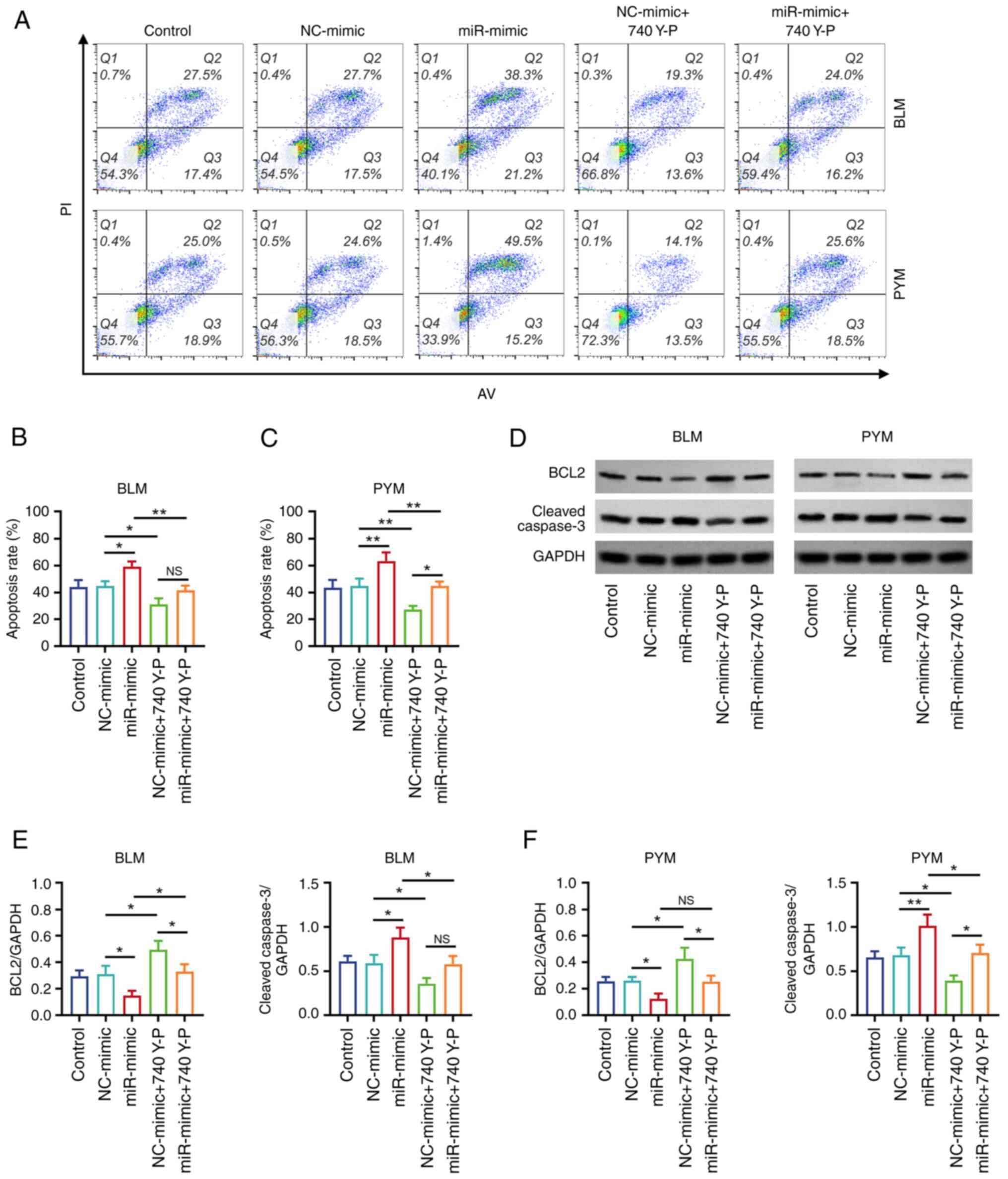
Annexin V-FITC/PI staining (Fig. 6A) demonstrated that, in cells
treated with BLM, the apoptosis rate was significantly decreased in
the NC-mimic + 740 Y-P group compared with the NC-mimic group
(P<0.05; Fig. 6B). In addition,
the apoptosis rate was significantly decreased in the miR-mimic +
740 Y-P group compared with the miR-mimic group (P<0.01). The
same trend was demonstrated in HemECs treated with PYM (both
P<0.01; Fig. 6C).
Western blotting (Fig.
6D) demonstrated that, in cells treated with BLM, BCL2 protein
expression levels were significantly increased but protein
expression of cleaved caspase 3 was decreased in the NC-mimic + 740
Y-P group compared with the NC-mimic group (both P<0.05;
Fig. 6E). Additionally, BCL2
protein expression was significantly increased and cleaved caspase
3 protein expression levels were decreased in the miR-mimic + 740
Y-P group compared with the miR-mimic group (both P<0.05). The
same trend of increased protein expression of BCL2 and decreased
protein expression levels of cleaved caspase 3 was demonstrated in
PYM-treated HemECs (P<0.05), with the exception of a marked
increase in BCL2 protein expression levels in the miR-mimic + 740
Y-P group compared with the miR-mimic group (P>0.05; Fig. 6F).
Discussion
A number of miRs, such as miR-556-3p, miR-139-5p and
miR-382-5p, participate in the progression of hemangioma (32-35).
For example, miR-556-3p targets VEGF C to suppress hemangioma cell
proliferation and increase apoptosis (32). miR-206 promotes extracellular
matrix accumulation and decreases hemangioma cell proliferation by
downregulating DNA methyltransferase 3A expression (36). In addition, miR-497-5p inhibits
hemangioma cell proliferation and induces ferroptosis by regulating
Notch receptor 2(37). miR-200c-3p
facilitates hemangioma cell proliferation by targeting the Notch
pathway (35). Our previous study
reported that miR-203a-3p knockdown regulates the Notch pathway to
facilitate progression of hemangioma (23). Furthermore, our previous study
reported that miR-203a-3p inhibits hemangioma cell proliferation
and invasion but facilitates apoptosis by inactivating the PI3K/AKT
pathway (24).
The present study hypothesized that miR-203a-3p may
influence the drug sensitivity of hemangioma cells. The present
study demonstrated that miR-203a-3p increased the sensitivity of
hemangioma cells to BLM and PYM. This may be due to regulation of
the PI3K/AKT pathway, phosphodiesterase 4D or snail family
transcriptional repressor 2 gene (21,24,38).
The latter factors (the PI3K/AKT pathway, phosphodiesterase 4D and
snail family transcriptional repressor 2 gene) are involved in the
regulation of BLM and PYM sensitivity (26,39,40).
However, this hypothesis requires further experimental validation.
In addition, the impact of miR-203a-3p on drug sensitivity in
certain types of cancer has been previously reported (41-43)
and the findings of the present study were in accordance with
aforementioned studies.
The PI3K/AKT pathway is a crucial regulator in
hemangioma (24,44,45).
Notably, a number of studies have reported the involvement of the
PI3K/AKT pathway in BLM or PYM sensitivity in hemangioma (25,26).
For example, the PI3K/AKT pathway is involved in PYM mediation of
hemangioma cell proliferation, invasion and apoptosis (26). To explore the downstream effects of
miR-203a-3p regulation of BLM and PYM sensitivity in hemangioma,
the impact of 740 Y-P was analyzed in the present study. It was
demonstrated that miR-203-3p negatively regulated the PI3K/AKT
pathway in HemECs and 740 Y-P diminished the impact of miR-203a-3p
on the PI3K/AKT pathway in HemECs. This finding was in line with
our previous study (24).
Additionally, 740 Y-P attenuated the effect of miR-203a-3p on BLM
or PYM sensitivity in hemangioma cells. This may have been due to
miR-203a-3p modulation of the expression of genes, such as VEGFA
and PTEN, to inhibit the PI3K/AKT pathway, which further
contributed to increased sensitivity of hemangioma cells to BLM and
PYM (24,46).
A number of studies have reported the involvement of
the PI3K/AKT pathway in BLM and PYM resistance in hemangioma
(25,26). The methods used by the
aforementioned studies were different compared with those used in
the present study. For example, the cell viability was analyzed
using MTT assay in a previous study (26), whereas a CCK-8 assay was used in
the present study. In addition, expression of the apoptotic marker
BCL2 was studied using immunofluorescence in a previous study
(25), while BCL2 protein
expression levels were detected by western blotting here. Overall,
the findings demonstrated in the present study were in line with
those of aforementioned studies (25,26),
which indicated the reliability.
The present study demonstrated that miR-203a-3p
downregulated PI3K/AKT pathway signaling to improve the BLM and PYM
sensitivity in hemangioma; however, whether the regulation of
miR-203a-3p on the PI3K/AKT pathway is direct or indirect should be
validated. Furthermore, miR-203a-3p may also regulate other
cellular pathways to sensitize hemangioma cells to BLM and PYM.
To conclude, miR-203a-3p improved hemangioma
sensitivity to BLM and PYM by inactivating the PI3K/AKT pathway.
This demonstrated the potential of miR-203a-3p as a future
co-treatment option for patients with hemangioma receiving BLM or
PYM treatment.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LZ and ZH contributed to study conception and
design. Material preparation and data collection were performed by
LZ, ZH and JC. Data analysis was performed by LZ, ZH, JC, QG and
JG. The manuscript was written by LZ, ZH and JC, and the manuscript
was revised by JC, QG and JG. LZ and ZH confirm the authenticity of
all the raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rodriguez Bandera AI, Sebaratnam DF,
Wargon O and Wong LF: Infantile hemangioma. Part 1: Epidemiology,
pathogenesis, clinical presentation and assessment. J Am Acad
Dermatol. 85:1379–1392. 2021.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Lin PF, Chen FC, Chen JY, Hu LH, Xie WJ,
Liu TY, Guo SB, Lin XM, Liu XW, Ye XH, et al: Incidence and
familial clustering of infantile haemangiomas: A multicentre study.
J Eur Acad Dermatol Venereol. 36:1641–1647. 2022.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Anderson KR, Schoch JJ, Lohse CM, Hand JL,
Davis DM and Tollefson MM: Increasing incidence of infantile
hemangiomas (IH) over the past 35 years: Correlation with
decreasing gestational age at birth and birth weight. J Am Acad
Dermatol. 74:120–126. 2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Seiffert A, Schneider M, Roessler J,
Larisch K and Pfeiffer D: Incidence, treatment patterns, and health
care costs of infantile hemangioma: Results of a retrospective
german database analysis. Pediatr Dermatol. 34:450–457.
2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Leaute-Labreze C, Harper JI and Hoeger PH:
Infantile haemangioma. Lancet. 390:85–94. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Sebaratnam DF, Rodriguez Bandera AL, Wong
LF and Wargon O: Infantile hemangioma. Part 2: Management. J Am
Acad Dermatol. 85:1395–1404. 2021.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Lee JC, Modiri O, England RW, Shawber CJ
and Wu JK: Propranolol therapy in infantile hemangioma: It is not
just about the beta. Plast Reconstr Surg. 147:875–885.
2021.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Colmant C and Powell J: Medical management
of infantile hemangiomas: An update. Paediatr Drugs. 24:29–43.
2022.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Wang C, Sun J, Guo L, Song D, Zhang X, Liu
Z and Wang L: Low-dose sclerotherapy with lauromacrogol in the
treatment of infantile hemangiomas: A retrospective analysis of 368
cases. Front Oncol. 12(1014465)2022.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Hassan Y, Osman AK and Altyeb A:
Noninvasive management of hemangioma and vascular malformation
using intralesional bleomycin injection. Ann Plast Surg. 70:70–73.
2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Ji Y, Chen S, Yang K, Zhang X, Zhou J, Li
L, Xiang B, Qiu T, Dai S, Jiang X, et al: Efficacy and safety of
propranolol vs atenolol in infants with problematic infantile
hemangiomas: A randomized clinical trial. JAMA Otolaryngol Head
Neck Surg. 147:599–607. 2021.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Kim KH, Choi TH, Choi Y, Park YW, Hong KY,
Kim DY, Choe YS, Lee H, Cheon JE, Park JB, et al: Comparison of
efficacy and safety between propranolol and steroid for infantile
hemangioma: A randomized clinical trial. JAMA Dermatol.
153:529–536. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Chen T, Gudipudi R, Nguyen SA, Carroll W
and Clemmens C: Should propranolol remain the gold standard for
treatment of infantile hemangioma? A systematic review and
meta-analysis of propranolol versus atenolol. Ann Otol Rhinol
Laryngol. 132:332–340. 2023.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Wu HW, Wang X, Zhang L, Zheng JW, Liu C
and Wang YA: Topical timolol vs. oral propranolol for the treatment
of superficial infantile hemangiomas. Front Oncol.
8(605)2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
McGahan JP and Goldman RE: Percutaneous
sclerotherapy with bleomycin and ethiodized oil: A welcomed
minimally invasive treatment for giant liver hemangiomas.
Radiology. 301:472–473. 2021.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Chen ZY, Wang QN, Zhu YH, Zhou LY, Xu T,
He ZY and Yang Y: Progress in the treatment of infantile
hemangioma. Ann Transl Med. 7(692)2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Ayoobi Yazdi N, Mehrabinejad MM, Dashti H,
Pourghorban R, Nassiri Toosi M and Rokni Yazdi H: Percutaneous
Sclerotherapy with bleomycin and ethiodized oil: A promising
treatment in symptomatic giant liver hemangioma. Radiology.
301:464–471. 2021.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Tiwari P, Pandey V, Bera RN, Tiwary N,
Mishra A and Sharma SP: Sandwich therapy in the management of
propranolol resistant infantile hemangioma of the lip. J Stomatol
Oral Maxillofac Surg. 123:e499–e505. 2022.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Hou J, Wang M, Tang H, Wang Y and Huang H:
Pingyangmycin sclerotherapy for infantile hemangiomas in oral and
maxillofacial regions: an evaluation of 66 consecutive patients.
Int J Oral Maxillofac Surg. 40:1246–1251. 2011.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhang C, Liu P, Huang J, Liao Y, Pan C,
Liu J, Du Q, Liu T, Shang C, Ooi S, et al: Circular RNA
hsa_circ_0043280 inhibits cervical cancer tumor growth and
metastasis via miR-203a-3p/PAQR3 axis. Cell Death Dis.
12(888)2021.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Chen L, Gao H, Liang J, Qiao J, Duan J,
Shi H, Zhen T, Li H, Zhang F, Zhu Z and Han A: miR-203a-3p promotes
colorectal cancer proliferation and migration by targeting PDE4D.
Am J Cancer Res. 8:2387–2401. 2018.PubMed/NCBI
|
|
22
|
Dai L, Zhang W, Wu X and Zhou S:
MicroRNA-203a-3p may prevent the development of thyroid papillary
carcinoma via repressing MAP3K1 and activating autophagy. J Clin
Lab Anal. 36(e24470)2022.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Hu Z, Liu X, Guo J, Zhuo L, Chen Y and
Yuan H: Knockdown of lncRNA MEG8 inhibits cell proliferation and
invasion, but promotes cell apoptosis in hemangioma, via
miR-203-induced mediation of the Notch signaling pathway. Mol Med
Rep. 24(872)2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Hu Z, Zhuo L, Li Y, Duan D and Guo J:
MicroRNA-203a-3p suppresses endothelial cell proliferation and
invasion, and promotes apoptosis in hemangioma by inactivating the
VEGF-mediated PI3K/AKT pathway. Exp Ther Med.
24(644)2022.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Mabeta P and Pepper MS: Inhibition of
hemangioma development in a syngeneic mouse model correlates with
bcl-2 suppression and the inhibition of Akt kinase activity.
Angiogenesis. 15:131–139. 2012.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Peng LX, Zhao P, Zhao HS, Pan E, Yang BB
and Li Q: Phosphoinositide 3-kinase/Akt pathway is involved in
pingyangmycin-induced growth inhibition, apoptosis and reduction of
invasive potential in EOMA mouse hemangioendothelioma cells. Mol
Med Rep. 12:8275–8281. 2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Zhou B, Li L, Qiu X, Wu J, Xu L and Shao
W: Long non-coding RNA ANRIL knockdown suppresses apoptosis and
pro-inflammatory cytokines while enhancing neurite outgrowth via
binding microRNA-125a in a cellular model of Alzheimer's disease.
Mol Med Rep. 22:1489–1497. 2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Zhao N, Du L, Ma Y, Wang Y, Ma J and Fang
Z: LncRNA NEAT1/microRNA-124 regulates cell viability, inflammation
and fibrosis in high-glucose-treated mesangial cells. Exp Ther Med.
24(507)2022.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Tian S, Lou L, Tian M, Lu G, Tian J and
Chen X: MAPK4 deletion enhances radiation effects and triggers
synergistic lethality with simultaneous PARP1 inhibition in
cervical cancer. J Exp Clin Cancer Res. 39(143)2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Lin F, Yang Y, Wei S, Huang X, Peng Z, Ke
X, Zeng Z and Song Y: Hydrogen sulfide protects against high
glucose-induced human umbilical vein endothelial cell injury
through activating PI3K/Akt/eNOS pathway. Drug Des Devel Ther.
14:621–633. 2020.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Jin W, Chen L, Gao F, Yang M, Liu Y and
Wang B: Down-regulation of miR-556-3p inhibits hemangioma cell
proliferation and promotes apoptosis by targeting VEGFC. Cell Mol
Biol (Noisy-le-grand). 66:204–207. 2020.PubMed/NCBI
|
|
33
|
Wu Y, Li H, Xie J, Wang F, Cao D and Lou
Y: miR-139-5p affects cell proliferation, migration and
adipogenesis by targeting insulin-like growth factor 1 receptor in
hemangioma stem cells. Int J Mol Med. 45:569–577. 2020.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Yuan X, Xu Y, Wei Z and Ding Q: CircAP2A2
acts as a ceRNA to participate in infantile hemangiomas progression
by sponging miR-382-5p via regulating the expression of VEGFA. J
Clin Lab Anal. 34(e23258)2020.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Hu X, Bai S, Li L, Tian P, Wang S, Zhang
N, Shen B, Du J and Liu S: MiR-200c-3p increased HDMEC
proliferation through the notch signaling pathway. Exp Biol Med
(Maywood). 246:897–905. 2021.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Wu M, Chen Y, Feng L, Dai H, Fang S and Xu
J: MiR-206 promotes extracellular matrix accumulation and relieves
infantile hemangioma through targeted inhibition of DNMT3A. Cell
Cycle. 20:978–992. 2021.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Ma Q, Dai X, Lu W, Qu X, Liu N and Zhu C:
Silencing long non-coding RNA MEG8 inhibits the proliferation and
induces the ferroptosis of hemangioma endothelial cells by
regulating miR-497-5p/NOTCH2 axis. Biochem Biophys Res Commun.
556:72–78. 2021.PubMed/NCBI View Article : Google Scholar
|
|
38
|
An N and Zheng B: MiR-203a-3p inhibits
pancreatic cancer cell proliferation, EMT, and apoptosis by
regulating SLUG. Technol Cancer Res Treat.
19(1533033819898729)2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Martin M, Zhang J, Miao Y, He M, Kang J,
Huang HY, Chou CH, Huang TS, Hong HC, Su SH, et al: Role of
endothelial cells in pulmonary fibrosis via SREBP2 activation. JCI
Insight. 6(e125635)2021.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Udalov S, Dumitrascu R, Pullamsetti SS,
Al-tamari HM, Weissmann N, Ghofrani HA, Guenther A, Voswinckel R,
Seeger W, Grimminger F and Schermuly RT: Effects of
phosphodiesterase 4 inhibition on bleomycin-induced pulmonary
fibrosis in mice. BMC Pulm Med. 10(26)2010.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Xiao Z, Qu Z, Chen Z, Fang Z, Zhou K,
Huang Z, Guo X and Zhang Y: LncRNA HOTAIR is a prognostic biomarker
for the proliferation and chemoresistance of colorectal cancer via
MiR-203a-3p-mediated Wnt/ss-catenin signaling pathway. Cell Physiol
Biochem. 46:1275–1285. 2018.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Shen M, Dong C, Ruan X, Yan W, Cao M,
Pizzo D, Wu X, Yang L, Liu L, Ren X and Wang SE:
Chemotherapy-induced extracellular vesicle miRNAs promote breast
cancer stemness by targeting ONECUT2. Cancer Res. 79:3608–3621.
2019.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Santangelo A, Rossato M, Lombardi G,
Benfatto S, Lavezzari D, De Salvo GL, Indraccolo S, Dechecchi MC,
Prandini P, Gambari R, et al: A molecular signature associated with
prolonged survival in glioblastoma patients treated with
regorafenib. Neuro Oncol. 23:264–276. 2021.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Yang K, Qiu T, Zhou J, Gong X, Zhang X,
Lan Y, Zhang Z and Ji Y: Blockage of glycolysis by targeting PFKFB3
suppresses the development of infantile hemangioma. J Transl Med.
21(85)2023.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Zhao Y, Li D, Han Y, Wang H, Du R and Yan
Z: The ester derivatives obtained by C-ring modification of
podophyllotoxin-induced apoptosis and inhibited proliferation in
hemangioma endothelial cells via downregulation of PI3K/Akt
signaling pathway. Chem Biol Drug Des. 99:828–838. 2022.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Zhang Y, Zheng Y and Zhu G: MiR-203a-3p
targets PTEN to promote hepatocyte proliferation by regulating
PI3K/Akt pathway in BRL-3A cells. Biosci Biotechnol Biochem.
84:725–733. 2020.PubMed/NCBI View Article : Google Scholar
|