Introduction
Pancreatic cancer (PC) is a deadly and aggressive
disease, accounting for 1.8% of all types of cancer worldwide, and
is characterized by increasing morbidity and mortality rates.
Pancreatic ductal adenocarcinoma (PDAC), arising from non-invasive
precancerous lesions, accounts for ~90% of all PC cases (1). PC is characterized by poor prognosis,
with a 5-year survival rate of <5% and an average survival time
without prompt treatment of no more than 6 months (2,3).
Currently, surgical resection combined with
neoadjuvant therapy is considered as the mainstay therapy approach
for PC. However, due to the lack of early symptoms and screening
strategies, the majority of patients with PC are initially
diagnosed at an advanced stage of the disease, presented with
unresectable or metastatic PC (4).
Chemotherapy is the most common strategy for treating metastatic
PC. However, prevailing resistance to chemotherapy greatly
restricts its utilization (5).
Therefore, exploring the intrinsic mechanism of PC to overcome
chemotherapy resistance and prevent cancer metastasis is of great
importance.
Glutathione peroxidase 3 (GPX3), located on
chromosome 5q33.1, is a glycosylated tetramer composed of four
subunits of 226 amino acids. It is the only exocrine member of the
GPX family, which catalyzes the detoxification of hydro- and
soluble lipid hydroperoxides by reducing glutathione and protects
cells from oxidative stress-related damage (6,7). In
recent years, the effect of GPX3 in cancer has attracted increasing
attention (8,9). Emerging evidence has suggested that
GPX3 has a diverse role in different types of human cancers,
serving as a pro-survival protein in myeloid leukemia and clear
cell renal cell carcinoma, and as a tumor suppressor in lung,
ovarian and gastric cancer (10-14).
Consistently, a previous study demonstrated that GPX3 was
downregulated in human cancers, but it was positively associated
with poor outcomes (15), thus
supporting the controversial role of GPX3 in cancer. It has been
also reported that GPX3 is highly involved in cancer metastasis and
chemotherapy resistance (15).
Nevertheless, the particular role of GPX3 in PC has not been
extensively investigated.
Therefore, the present study aimed to explore the
substantial effect of GPX3 on the tumorigenesis and metastasis of
PC and to elucidate its underlying molecular mechanism of action,
thus providing novel insights into the development of effective
therapeutic strategies against PC.
Materials and methods
Application of bioinformatics
databases
The expression pattern of GPX3 in pancreatic
adenocarcinoma (PAAD; n=178) and the normal tumor-adjacent tissues
(n=4) was downloaded from UALCAN database (https://ualcan.path.uab.edu/index.html) (16). The association between GPX3
expression (cut-off value, 50%; to determine high and low GPX3
expression levels) and prognosis [overall survival (OS),
disease-free survival (DFS) and relapse-free survival (RFS)] in
PAAD was obtained from the GEPIA (http://gepia.cancer-pku.cn/) (17) and Kaplan-Meier Plotter (http://kmplot.com/analysis/) databases (18).
Cell lines
The human pancreatic ductal epithelial cell line
HPDE6c7 (cat. no. BNCC359453) was obtained from BeNa Culture
Collection. The PC cell lines BxPC-3 (cat. no. CL-0042), SW1990
(cat. no. CL-0448B) and PANC-1 (cat. no. CL-0184) were purchased
from Wuhan Procell Life Science & Technology Co., Ltd. HPDE6c7
and BxPC-3 cells were cultured in RPMI-1640 medium, while SW1990
and PANC-1 cells were cultured in DMEM supplemented with 10% fetal
bovine serum (FBS; MilliporeSigma) and 1% penicillin/streptomycin
solution (Life Technologies; Thermo Fisher Scientific, Inc.). All
cells were cultured at 37˚C in a humidified incubator with 5%
CO2.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from cells utilizing a
TRIzol reagent (MilliporeSigma), according to the manufacturer's
instructions. After purity verification and concentration
measurement, 1 µg total RNA was reverse transcribed into
complementary DNA using the riboScript Reverse Transcription Kit
(Guangzhou RiboBio Co., Ltd.) according to the manufacturer's
instructions. Subsequently, qPCR was carried out using the SYBR
Premix Ex Taq II kit (Takara Bio Inc.) on a real-time PCR
instrument (Bio-Rad Laboratories, Inc.). The reaction protocol was
as follows: Initial denaturation at 95˚C for 30 sec; 40 cycles at
95˚C for 5 sec and 60˚C for 20 sec, and a final extension step at
72˚C for 10 min. The primer sequences used in this study were as
follows: GPX3 forward, 5'-AGCAGTATGCTGGCAAATATGTCC-3' and reverse,
5'-CAGACCGAATGGTGCAAGCTCTTC-3'; β-actin forward,
5'-AGCGAGCATCCCCCAAAGTT-3' and reverse, 5'-GGGCACGAAGGCTCATCATT-3'.
The expression levels of GPX3 were calculated using the
2-ΔΔCq method (19) and
β-actin served as an endogenous control.
Western blot analysis
Protein samples were prepared following cell lysis
with radioimmunoprecipitation assay (RIPA) lysis buffer (Beyotime
Institute of Biotechnology) containing protease and phosphatase
inhibitors (Roche Diagnostics). Following protein concentration
assessment using a bicinchoninic acid protein assay kit (Pierce;
Thermo Fisher Scientific, Inc.), equal amounts of protein extracts
(30 µg/lane) were separated by 15% SDS-PAGE and were then
transferred onto polyvinylidene fluoride membranes
(MilliporeSigma). Non-specific binding was blocked following
membrane incubation with 5% skimmed milk for 2 h at room
temperature. Subsequently, membranes were incubated with primary
antibodies, including anti-GPX3 (1:1,000; cat. no. ab275965,
Abcam), anti-E-cadherin (1:1,000; cat. no. ab40772, Abcam),
anti-N-cadherin (1:5,000; cat. no. ab76011, Abcam), anti-Snail
(1:1,000; cat. no. ab216347, Abcam), anti-Bax (1:1,000; cat. no.
ab32503, Abcam), anti-Bcl-2 (1:1,000; cat. no. ab32124, Abcam),
anti-p-JNK (1:1,000; cat. no. 9251, Cell Signaling Technology,
Inc.), anti-JNK (1:1,000; cat. no. 9252, Cell Signaling Technology,
Inc.), anti-p-c-Jun (1:1,000; cat. no. ab32385, Abcam), anti-c-Jun
(1:1,000; cat. no. ab40766, Abcam), anti-p21 (1:1,000; cat. no.
ab109520, Abcam), anti-c-Myc (1:1,000; cat. no. ab32072, Abcam),
and anti-GAPDH (1:2,500; cat. no. ab9485, Abcam) at 4˚C overnight
followed by incubation with HRP-conjugated secondary antibody
(1:2,000; cat. no. ab6721, Abcam) for 2 h at room temperature.
Finally, the protein bands were visualized utilizing an Enhanced
Chemiluminescence System Reagent (Nanjing KeyGen Biotech, Co.,
Ltd.) and quantified using ImageJ software version 1.48 (National
Institutes of Health).
Cell transfection and anisomycin
treatment
The coding sequences of GPX3 (accession no.
NM_002084.5) were cloned into pcDNA3.1 (Shanghai Genechem Co.,
Ltd.) to construct GPX3-overexpressing vector (Oe-GPX3), and the
empty pcDNA3.1 vector served as the negative control (Oe-NC). Upon
achieving 60-70% confluency, PANC-1 cells were transfected with 5
µg/ml Oe-NC or Oe-GPX3 using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.) at 37˚C for 48 h,
according to the manufacturer's protocol. At 48 h following
transfection, the cell transfection efficiency was assessed by
RT-qPCR and western blot analysis. For the rescue experiment, the
GPX3-overexpressing PANC-1 cells were treated with 0.01 µM
anisomycin (Shanghai Yuan Ye Bio-Technology Co., Ltd.), an
activator of JNK, for another 24 h at 37˚C.
Cell viability assay
Cell viability was assessed using a Cell Counting
Kit-8 (CCK-8) assay. Briefly, PANC-1 cells were seeded into 96-well
plates at a density of 2x103 cells/well and cultured at
37˚C in an incubator with 5% CO2. Following incubation
for 24, 48 and 72 h, each well was supplemented with 10 µl CCK-8
reagent (Dojindo Laboratories, Inc.) and cells were incubated for
an additional 3 h. The absorbance at a wavelength of 450 nm was
measured in each well using a microplate reader (BioTek
Instruments, Inc.).
Colony formation assay
A total of 2x103 PANC-1 cells were
inoculated into each well of 6-well plates and cultured at 37˚C in
an incubator with 5% CO2 for 2 weeks. The culture medium
was changed with fresh one every 3 days. The formed colonies were
fixed with 4% paraformaldehyde for 10 min at room temperature and
then stained with 1% crystal violet for 30 min at room temperature.
The colonies (>50 cells) were observed under a microscope and
counted using ImageJ software version 1.48.
5-Ethynyl-2'-deoxyuridine (EdU)
incorporation assay
An EdU assay was carried out using the Cell-Light
EdU DNA Cell Proliferation Kit (Guangzhou RiboBio Co., Ltd.).
Briefly, PANC-1 cells were labeled with EdU for 2 h at 37˚C.
Subsequently, cells were stained with DAPI solution
(MilliporeSigma) for 5 min. The fluorescence signal was visualized
under an inverted fluorescence microscope (Olympus Corporation),
and then quantified using ImageJ software version 1.48.
Wound healing assay
A wound healing assay was conducted to assess the
migration ability of PC cells. When PANC-1 cells reached 100%
confluency in 6-well plates, a wound was created on the cell
monolayer using a 200-µl sterile pipette tip. Following washing
with PBS for three times to remove detached cells, cells were
incubated in serum-free medium for 24 h. Images of the wound at 0
and 24 h were captured under a light microscope (Olympus
Corporation). The cell migration rate was calculated depending on
the shortened wound distance of each group: (wound distance at 0
h-wound distance at 24 h)/wound distance at 0 h x100.
Transwell assay
To evaluate the invasion ability of PC cells, a
Transwell assay was performed in a 24-well plate using a Transwell
chamber (Corning; Corning, Inc.) pre-coated with Matrigel (BD
Biosciences) at 37˚C for 30 min. Briefly, PANC-1 cells
(5x105 cells/ml) were resuspended in serum-free medium
and were then added onto the upper chamber of the Transwell insert.
The lower chamber was supplemented with complete medium containing
10% FBS. Following incubation at 37˚C for 48 h, the remaining cells
on the upper surface of the membrane were removed using cotton
swabs. The invasive cells were fixed in 4% paraformaldehyde and
were then stained with 1% crystal violet for 30 min at room
temperature. Finally, the invasive cells were observed and counted
under a light microscope (Olympus Corporation). The cell invasion
rate was calculated depending on the number of invasive cells of
each group.
Gemcitabine (GEM) chemosensitivity
assay
PANC-1 cells were treated with increasing
concentrations of GEM (0, 1, 2, 5, 10 and 20 µM; APeXBIO Technology
LLC) for 48 h. Meanwhile, GEM-resistant PANC-1 cells (PANC-1/GEM;
cat. no. IMD-015; Xiamen Immocell Biotechnology Co., Ltd.) were
treated with 0, 5, 10, 20, 40, 60, 80 and 100 µM GEM for 48 h.
Subsequently, the cell viability of each group was assessed as
aforementioned. The half maximal inhibitory concentration
(IC50) values were calculated using the
concentration-response data in GraphPad Prism 8.0 software
(GraphPad Software, Inc.).
Flow cytometric analysis
Cell apoptosis was assessed using an Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) kit (BD
Biosciences). PANC-1/GEM cells were seeded into 6-well plates
(5x105 cells/per well). Following incubation with GEM
(80 µM) or PBS at 37˚C for 48 h, PANC-1/GEM cells were washed with
ice-cold PBS and resuspended in binding buffer, followed by the
addition of Annexin V-FITC and PI, according to the manufacturer's
protocol. Following incubation at 37˚C for 15 min in the dark, the
apoptotic cells, including early apoptotic cells and late apoptotic
cells, were analyzed with the FACScan flow cytometer (BD
Biosciences) and CellQuest Pro software version 3.3 (BD
Biosciences).
Statistical analysis
The experimental data were statistically analyzed
using GraphPad Prism 8.0 software (GraphPad Software, Inc.;
Dotmatics). All experiments were repeated at least three times. The
experimental data were normally distributed and were expressed as
the mean ± standard deviation (SD). The differences among multiple
groups were compared using one-way ANOVA, followed by Tukey's post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
GPX3 is downregulated in PC
Bioinformatics analysis was performed to assess the
effect of GPX3 on PC. As shown in Fig.
1A, the expression levels of GPX3 in tumor tissues were lower
compared with those in normal ones, thus suggesting that GPX3 was
downregulated in PC. In addition, the increased expression levels
of GPX3 were positively associated with enhanced OS, DFS and RFS
(Fig. 1B and C). Additionally, the expression levels of
GPX3 were also detected in PC cell lines and the results
consistently disclosed that both the mRNA and protein expression
levels of GPX3 were markedly decreased in the PC cell lines BxPC-3,
SW1990 and PANC-1 compared with HPDE6c7 cells (Fig. 1D). Since the lowest expression
levels of GPX3 were observed in PANC-1 cells, this cell line was
selected for the subsequent in vitro experiments.
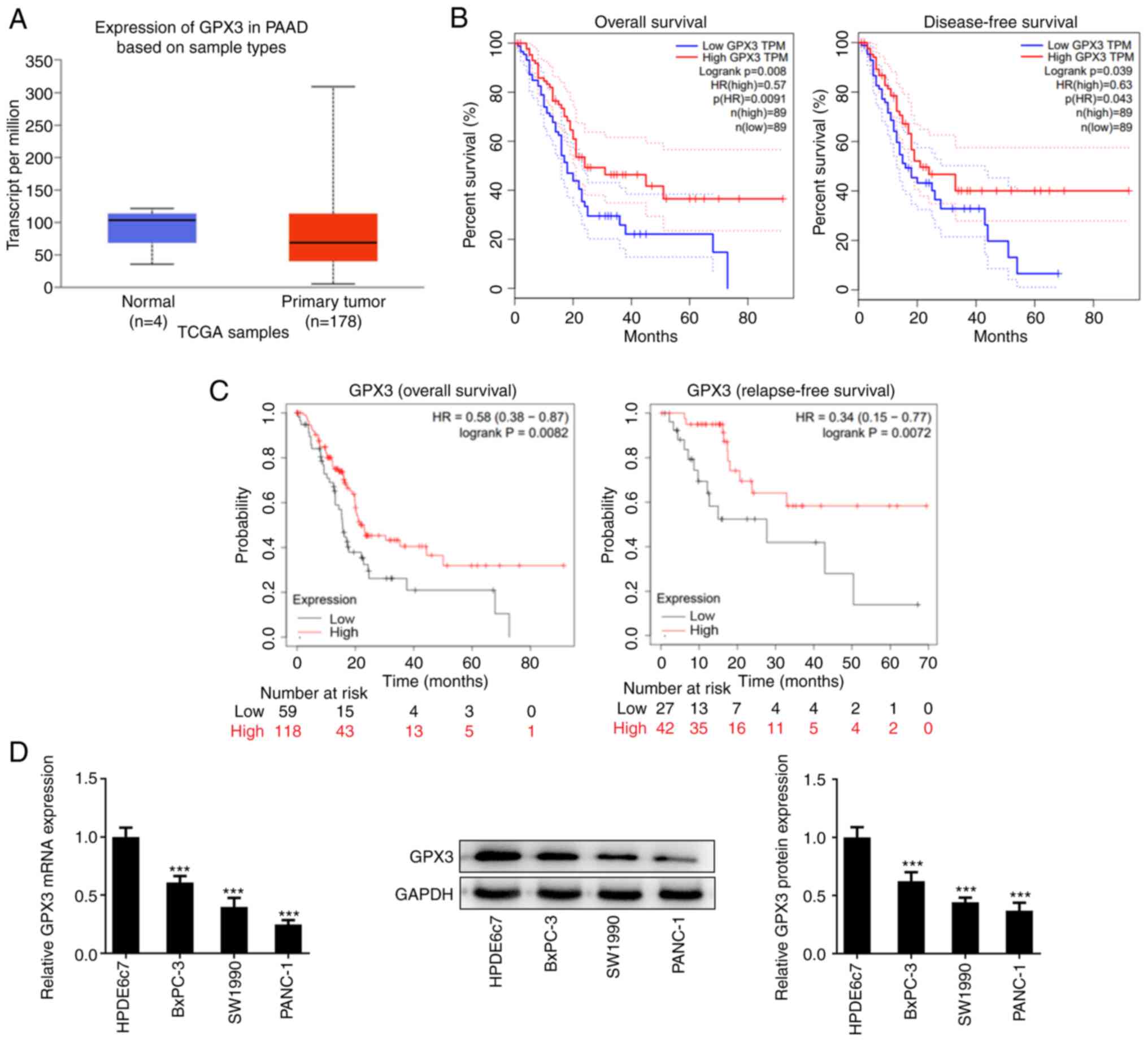 | Figure 1GPX3 is downregulated in PC. (A) The
expression of GPX3 in PAAD and normal-adjacent tissues was
presented by UALCAN database (https://ualcan.path.uab.edu/index.html). (B) The
association between GPX3 expression and OS and disease-free
survival in PAAD was obtained from GEPIA database (http://gepia.cancer-pku.cn/). (C) The association
between GPX3 expression and OS and relapse-free survival in PAAD
was obtained from Kaplan-Meier Plotter database (http://kmplot.com/analysis/). (D) The mRNA and protein
expression levels of GPX3 in the PC cell lines, BxPC-3, SW1990 and
PANC-1, and human pancreatic ductal epithelial cell line, HPDE6c7
cells, were determined using reverse transcription-quantitative PCR
and western blot analysis, respectively. ***P<0.001
vs. HPDE6c7 cells. GPX3, glutathione peroxidase 3; PC, pancreatic
cancer; PAAD, pancreatic adenocarcinoma; OS, overall survival; TPM,
transcript per million. |
GPX3 inhibits the proliferation,
migration and invasion of PANC-1 cells
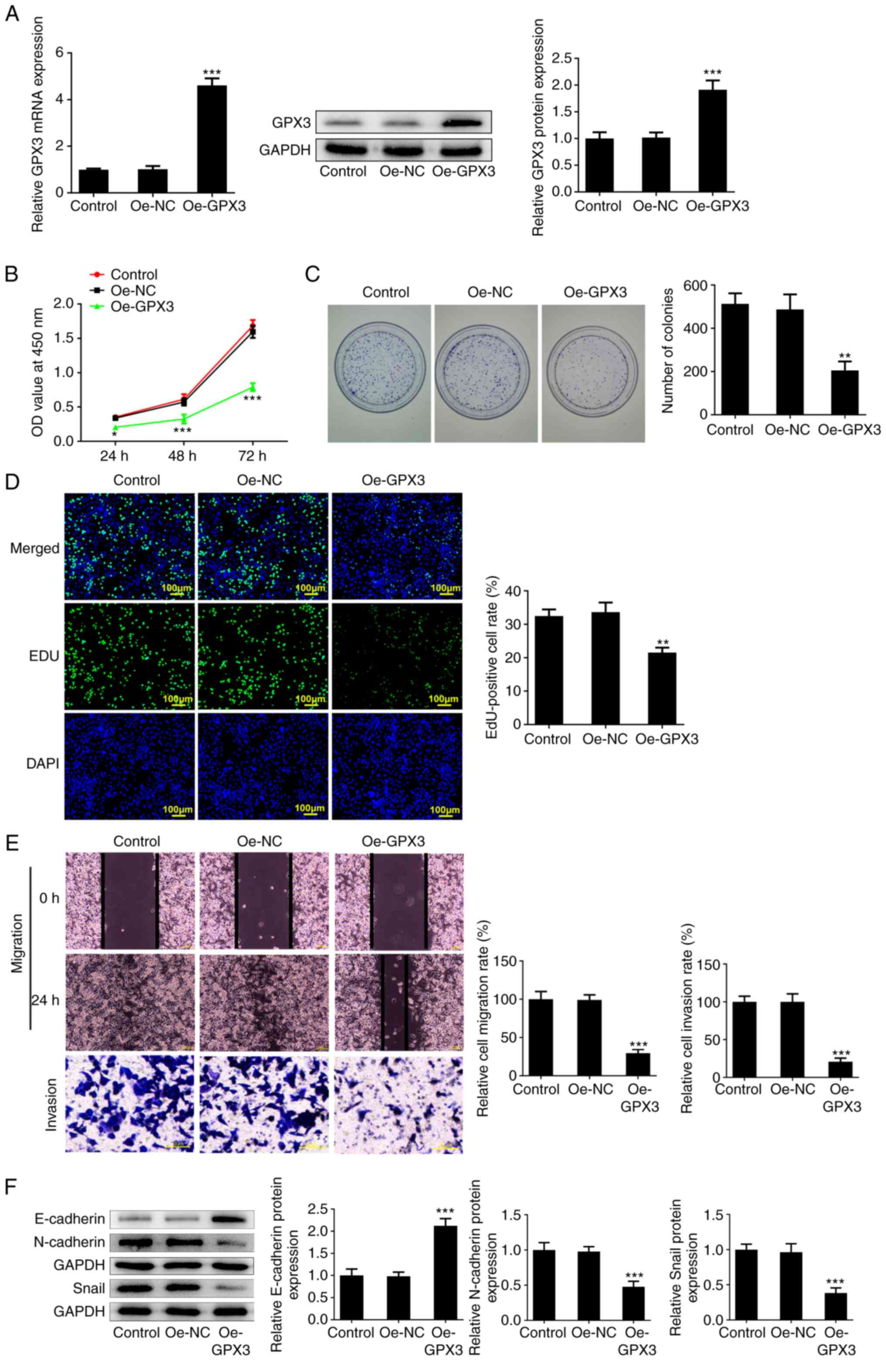
To explore the regulatory role of GPX3 in PC,
gain-of-function experiments were carried out. PANC-1 cells were
first transfected with Oe-GPX3. The markedly increased mRNA and
protein expression levels of GPX3 in the Oe-GPX3 group confirmed
that the cells were successfully transfected (Fig. 2A). As shown in Fig. 2B and C, GPX3 overexpression significantly
inhibited cell viability and reduced the colony formation ability
of PC cells, thus supporting the anti-proliferative capacity of
GPX3 in PANC-1 cells. The above finding was further verified by the
reduced number of EdU-positive cells in the Oe-GPX3 group (Fig. 2D). Furthermore, wound healing and
Transwell assays revealed that GPX3 overexpression not only
inhibited the healing velocity of PANC-1 cells within 48 h, but
also reduced their invasion ability (Fig. 2E), thus suggesting that GPX3 could
inhibit the migration and invasion of PANC-1 cells. The results
also demonstrated that GPX3 notably increased the protein
expression levels of E-cadherin and decreased those of N-cadherin
and Snail (Fig. 2F), thus
indicating that GPX3 attenuated the epithelial-mesenchymal
transition (EMT) of PANC-1 cells.
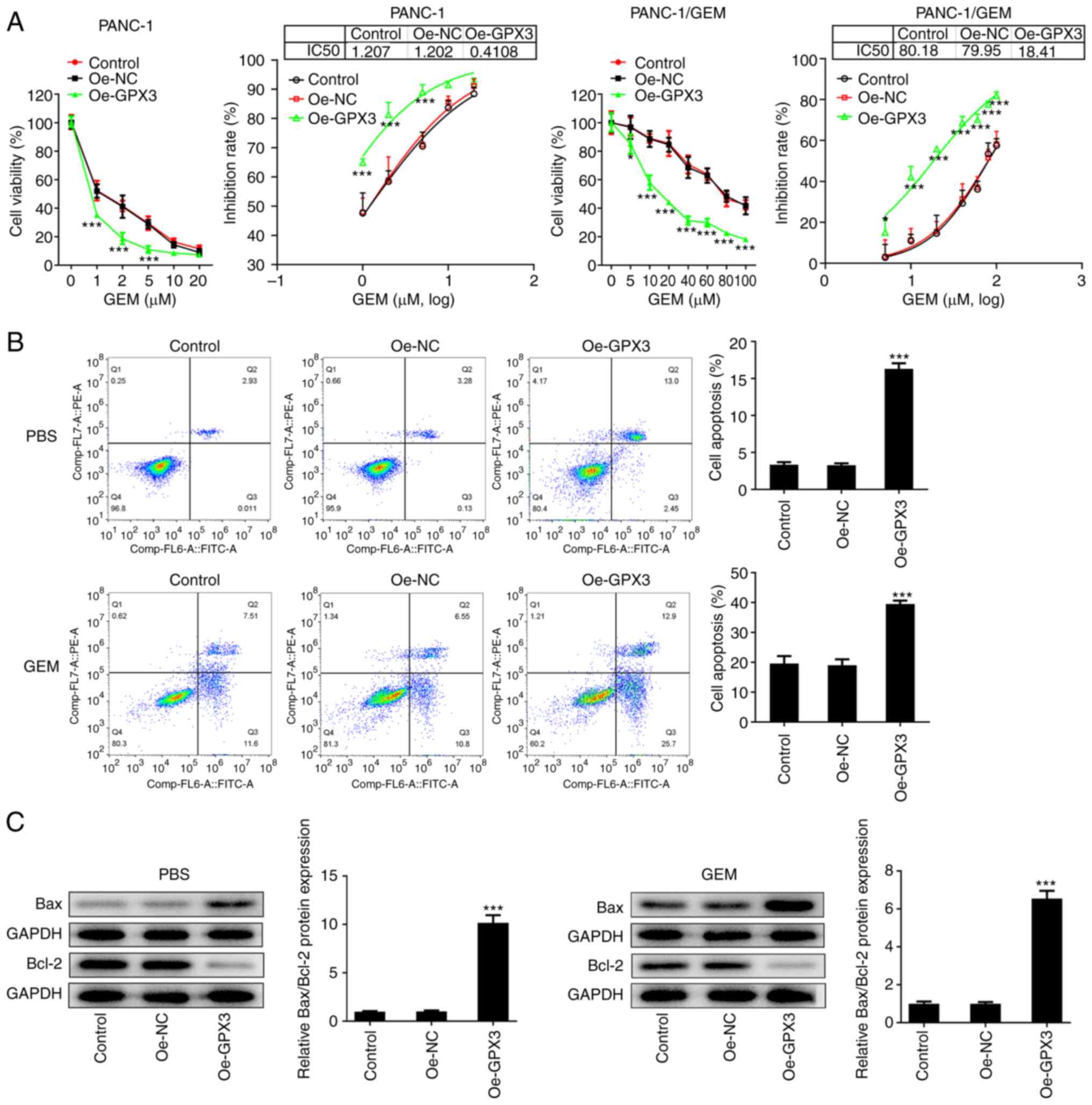
GPX3 sensitizes PANC-1 and PANC-1/GEM
cells to GEM
In addition to the enhanced proliferation and
invasion abilities of cancer cells, chemo-resistance is also
significantly associated with poor prognosis in PC (20). Therefore, the present study also
aimed to investigate the effect of GPX3 on the chemo-resistance of
PANC-1 cells to GEM. The results revealed that PANC-1/GEM cells
possessed a higher GEM IC50 value compared with PANC-1
cells, while GPX3 overexpression markedly reduced the
IC50 value of GEM in PANC-1 and PANC-1/GEM cells
(Fig. 3A). The flow cytometric
analysis showed that cell treatment with GEM promoted the apoptosis
of PANC-1/GEM cells, which was further enhanced by GPX3
overexpression (Fig. 3B).
Consistently, the western blot analysis results showed that Bax was
upregulated and Bcl-2 was downregulated in the Oe-GPX3 group, thus
further verifying the anti-apoptotic activity of GPX3 in PANC-1/GEM
cells (Fig. 3C). The
aforementioned findings indicated that GPX3 overexpression could
robustly improve the chemo-sensitivity of PC and GEM-resistant PC
cells to GEM.
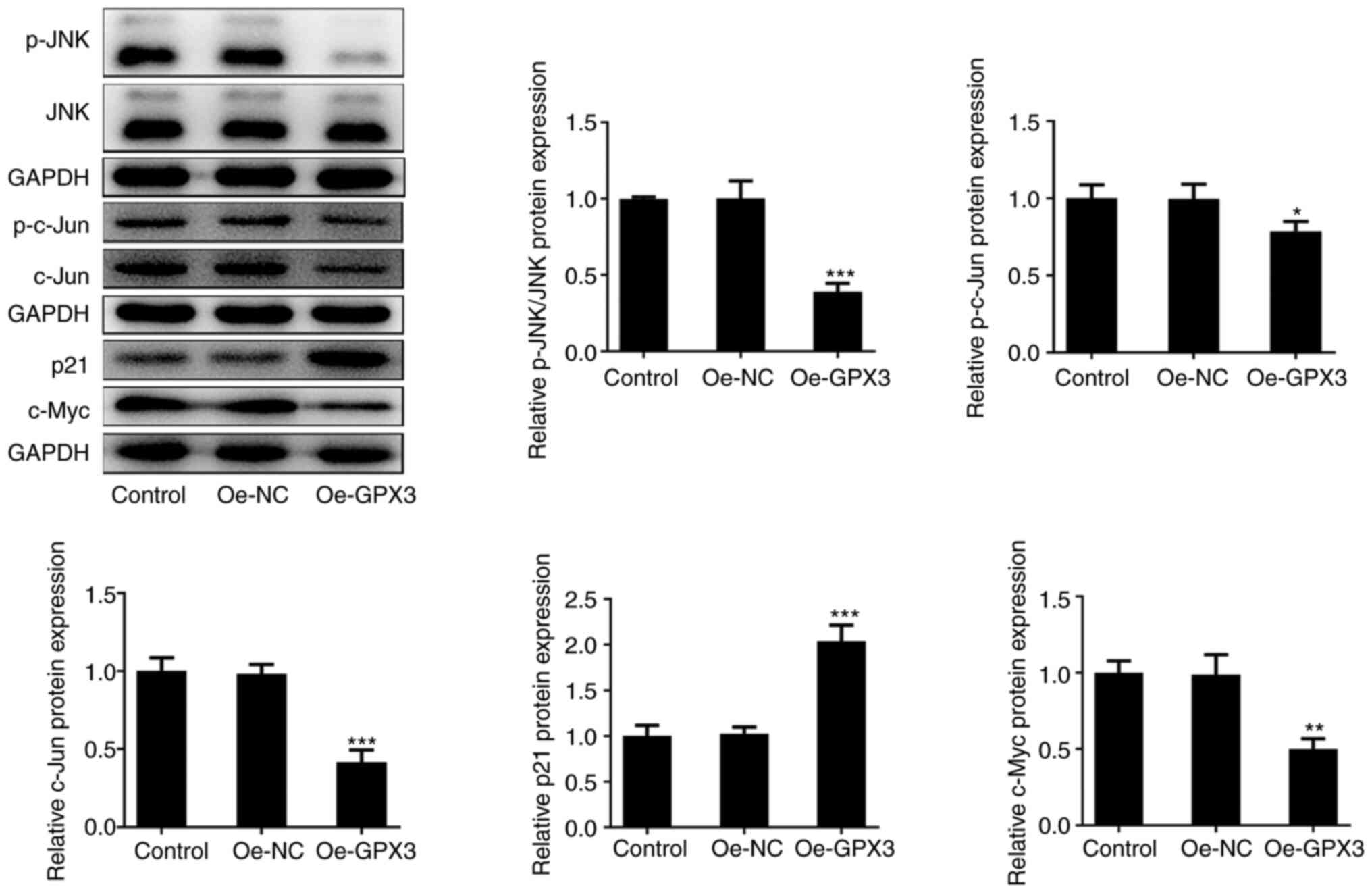
GPX3 inhibits the activity of JNK
signaling in PANC-1 cells
Subsequently, the present study aimed to uncover the
detailed mechanism underlying the effect of GPX3 on antagonizing
the malignant behavior of PC cells. Therefore, western blot
analysis showed that compared with the Oe-NC group, the protein
expression levels of phosphorylated (p)-JNK, p-c-Jun, c-Jun and
c-Myc were markedly decreased, while those of p21 were robustly
elevated in the Oe-GPX3 group (Fig.
4). The aforementioned finding indicated that GPX3
significantly inhibited JNK/c-Jun signaling in PANC-1 cells.
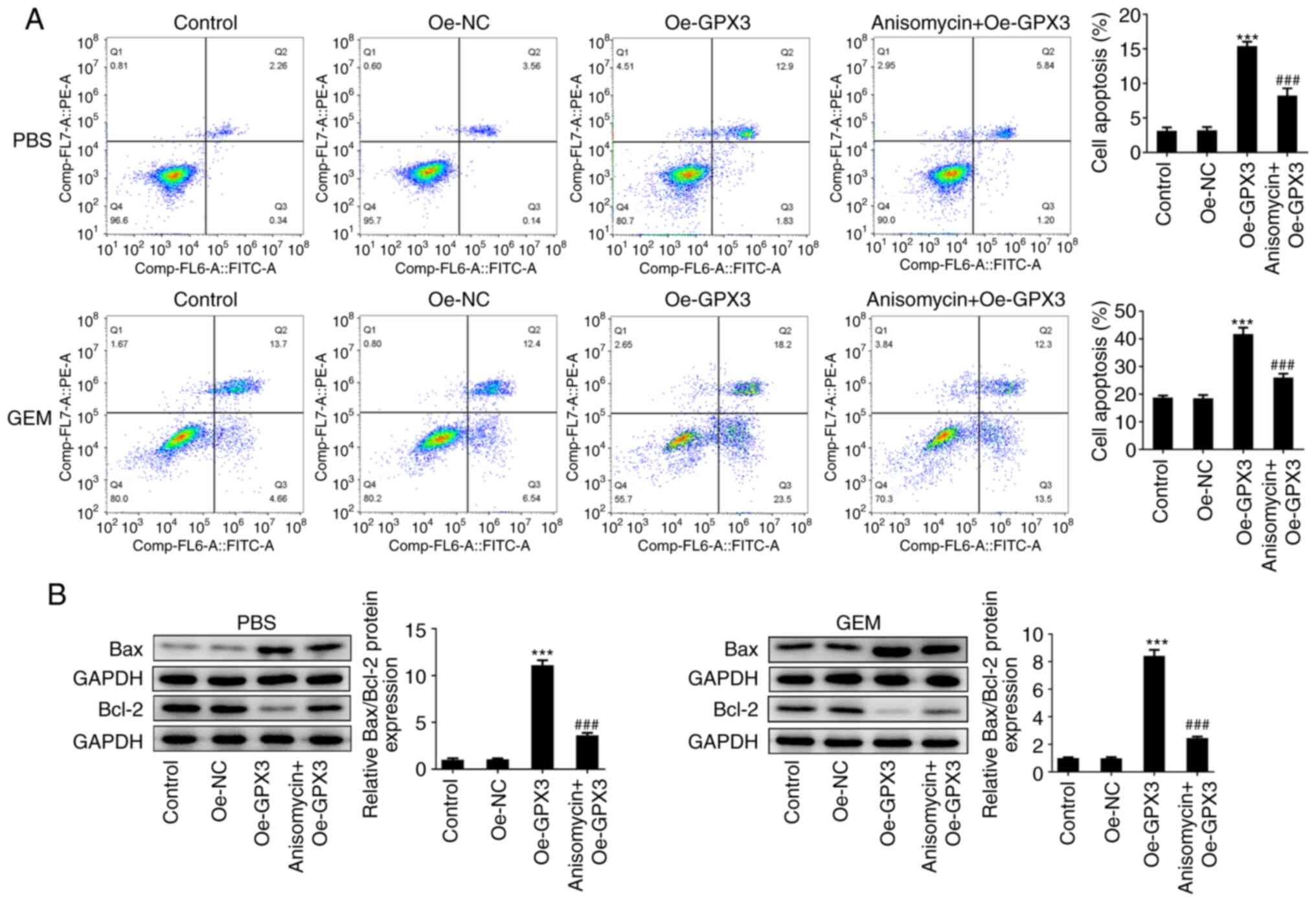
Anisomycin reverses the inhibitory
effects of GPX3 on the proliferation, invasion, EMT and
chemo-resistance of PC cells
Finally, to clarify the significance of JNK/c-Jun
signaling in the antitumor activity of GPX3 in PC,
GPX3-overexpressing PANC-1 cells were treated with 0.01 µM
anisomycin, an activator of JNK, and then a series of in
vitro experiments were performed. As shown in Fig. 5A-C, anisomycin significantly
weakened the anti-proliferative effect of GPX3 on PANC-1 cells, as
evidenced by the enhanced cell viability and cell colony formation
ability, and the increased number of EdU-positive cells in the
anisomycin + Oe-GPX3 group compared with the Oe-GPX3 group. In
addition, compared with the Oe-GPX3 group, cell treatment with
anisomycin enhanced the migration and invasion rates of PC cells
(Fig. 5D). Furthermore, E-cadherin
downregulation and N-cadherin and Snail upregulation following cell
treatment with anisomycin indicated that the inhibitory effect of
GPX3 overexpression on EMT was partially restored by anisomycin
treatment (Fig. 5E). In addition,
compared with the Oe-GPX3 group, the reduced cell apoptosis rate
and Bax/Bcl-2 ratio in the anisomycin + Oe-GPX3 group suggested
that cell treatment with anisomycin abrogated the beneficial effect
of GPX3 on the chemo-sensitivity of PC cells to GEM (Fig. 6A and B). These fidings indicated that GPX3
inhibited cell proliferation, migration, invasion and
chemo-resistance in PC cells via inhibiting the JNK/c-Jun signaling
pathway.
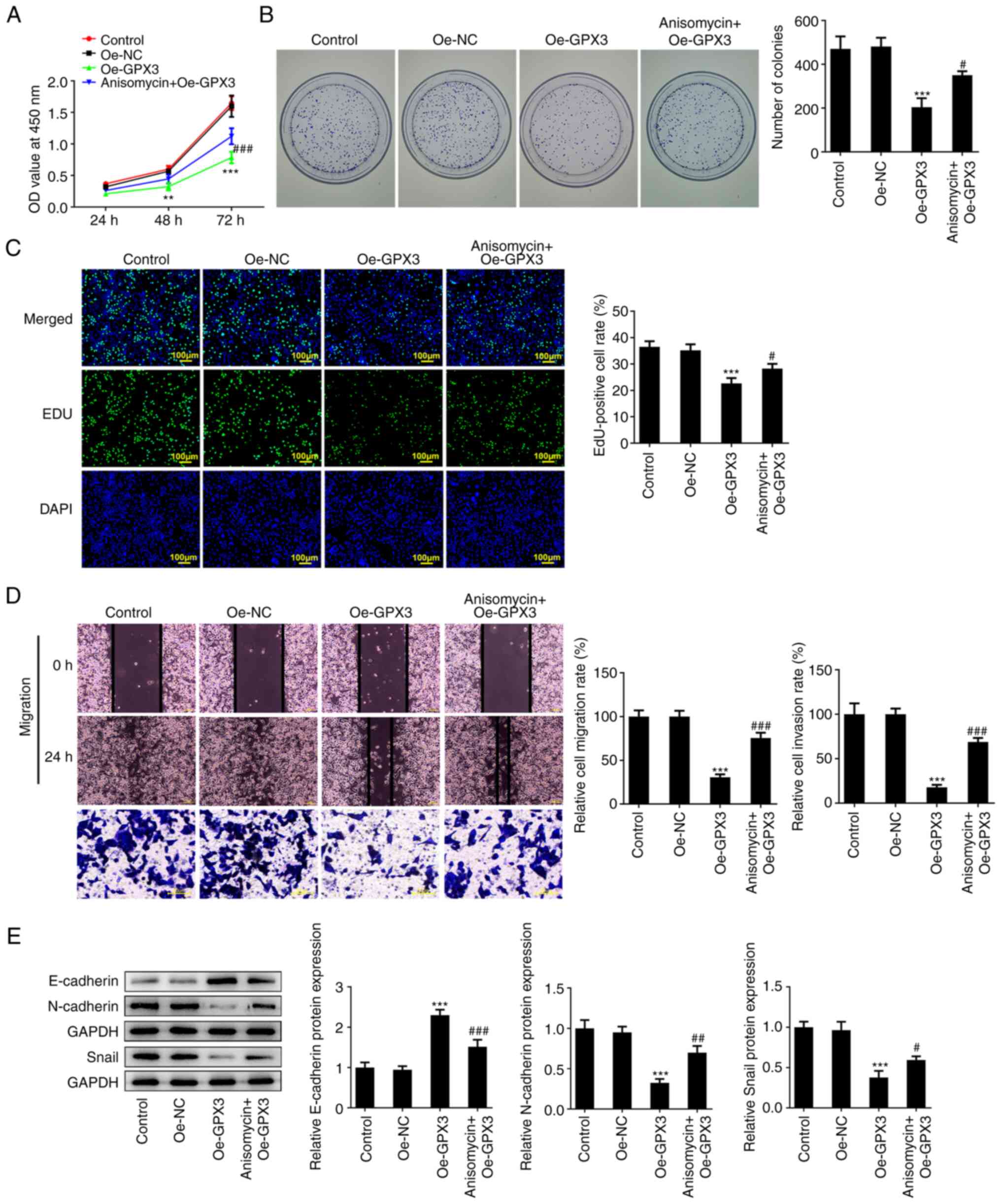 | Figure 5Anisomycin reverses the inhibitory
effects of GPX3 on the proliferation, invasion and
epithelial-mesenchymal transition of pancreatic cancer cells. (A)
PANC-1 cells were transfected with Oe-NC or Oe-GPX3 and
GPX3-overexpressing PANC-1 cells were then treated with 0.01 µM
anisomycin, a JNK activator. Cell viability was measured at 24, 48
and 72 h using Cell Counting Kit-8 assay. (B) A colony formation
assay was performed and the formed colonies were counted under a
microscope. (C) 5-Ethynyl-2'-deoxyuridine incorporation assay was
performed to assess cell proliferation. (D) Wound healing and
Transwell assays were carried out to evaluate the cell migration
and invasion abilities, respectively. (E) The protein expression
levels of E-cadherin, N-cadherin and Snail were detected using
western blot analysis. **P<0.01 and
***P<0.001 vs. the Oe-NC group;
#P<0.05, ##P<0.01 and
###P<0.001 vs. the Oe-GPX3 group. GPX3, glutathione
peroxidase; Oe-NC, negative control overexpression vector; Oe-GPX3,
GPX3-overexpressing vector; Oe-GPX3, GPX3-overexpressing
vector. |
Discussion
In the present study, GPX3 was discovered to be
downregulated in PC and positively associated with the prognosis of
patients with PC. More particularly, in vitro experiments
revealed that GPX3 overexpression could not only markedly suppress
the proliferation, migration and invasion of PANC-1 cells, but it
could also enhance the chemo-sensitivity of PANC-1/GEM cells to
GEM, suggesting that GPX3 exhibited tumor suppressive activity
during the malignant metastasis and chemo-resistance of PC. In
addition, the results demonstrated that the antitumor activity of
GPX3 in PC was partially mediated by JNK/c-Jun signaling
inhibition.
Chemotherapy is the most common strategy for
treating PC metastasis. GEM, as a standard first-line
chemotherapeutic agent, is widely utilized for the palliative
treatment of patients with PC. However, the effect of GEM on
prolonging the prognosis of patients with PC is limited due to drug
resistance (21). Emerging
evidence has suggested that multiple genes and proteins are
involved in regulating the chemo-resistance of PC cells to GEM. For
instance, a previous study showed that HEAT repeat containing 1
(HEATR1) was closely associated with the prognosis of patients with
PC, while HEATR1 depletion could greatly enhance the proliferation
of PC cells and their resistance to GEM, thus indicating that
HEATR1 may be a promising therapeutic target for PC (22). In addition, cancerous inhibitor of
protein phosphatase 2A (CIP2A) was found to be highly expressed in
PC tissues and CIP2A depletion could significantly repress the
proliferation and increase the chemo-sensitivity of PC cells to
GEM, thus attenuating the progression of PC (23). In the present study, the results
showed that GPX3 was downregulated in PC cell lines. Furthermore,
the gain-of-function experiments revealed that GPX3 overexpression
could not only notably attenuate the malignant behavior of PC
cells, including cell proliferation, migration and invasion, but it
could also markedly sensitize PC and GEM-resistant PC cells to GEM.
The aforementioned findings suggested that GPX3 could function as a
tumor suppressor in PC and may possibly serve as a biomarker to
guide the GEM chemotherapy of PC.
It is widely recognized that the MAPK signaling
pathway is one of the most attractive targets for cancer therapy.
JNK, one of the two major MAPK pathways, can regulate the
expression of target genes involved in modulating cell survival and
apoptosis via activating c-Jun (24,25).
Emerging evidence has also suggested that JNK/c-Jun signaling is
involved in the development of PC and the chemosensitivity of PC
cells (25,26). Previous studies revealed that the
phosphorylation of JNK induced the migration and invasion of PC
cells. c-Jun, the main downstream molecule of JNK, was expansively
expressed in pancreatic tumor lesions and it was closely associated
with PC progression (27-30).
Liu et al (31) reported
that the Zrt-Irt-like protein 4/zinc finger E-box-binding homeobox
1 axis mediated the resistance of pancreatic tumors to GEM via
regulating JNK/c-Jun signaling. Shi et al (26) also demonstrated that IX, a JNK
inhibitor, restrained PC via regulating p53 and p21. Therefore,
inhibiting JNK/c-Jun signaling may be a practicable approach for
alleviating the metastasis and chemoresistance of PC (26,32).
In the present study, GPX3 overexpression significantly inhibited
the activity of JNK/c-Jun signaling. To verify the aforementioned
regulatory mechanism, rescue experiments were carried out using
anisomycin, a JNK activator, and the results revealed that the
inhibitory effects of GPX3 on PC cell proliferation, invasion and
chemo-resistance were partially restored by anisomycin, thus
confirming that GPX3 exerted its antitumor effect in PC partly via
inhibiting JNK/c-Jun signaling.
However, there are some limitations in the present
study. It would be beneficial to perform animal experiments to
verify the in vitro findings. Furthermore, all data were
obtained from PANC-1 cells, and the experiments in other PC cell
lines may be beneficial for further validation. In addition, the
present study preliminarily revealed the regulatory role of GPX3
and the potential mechanism focusing on JNK/c-Jun signaling during
the progression of PC; however, other specific players involved and
the molecular mechanisms in PC require further investigation. These
limitations need to be addressed and may be future directions of
subsequent research.
To the best of the authors' knowledge, the present
study, for the first time, clarified the precise role of GPX3 in
PC, and elucidated the regulatory mechanism. The findings of the
present study revealed that GPX3 may serve as a tumor suppressor in
PC via inhibiting the malignant behavior of PC cells and improving
their chemosensitivity to GEM, which could be partially triggered
via inhibiting JNK/c-Jun signaling. The present study suggested
that GPX3 may be considered as a potentially valuable target for
improving PC treatment, thus providing novel insights into the
development of more effective therapeutic strategies for treating
PC.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
DZ designed the experiments. YM, LZ and XG obtained,
analyzed and interpreted the data. YM and LZ drafted the manuscript
and DZ revised the manuscript. YM and DZ confirm the authenticity
of all the raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lippi G and Mattiuzzi C: The global burden
of pancreatic cancer. Arch Med Sci. 16:820–824. 2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Bengtsson A, Andersson R and Ansari D: The
actual 5-year survivors of pancreatic ductal adenocarcinoma based
on real-world data. Sci Rep. 10(16425)2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Doleh Y, Lal LS, Blauer-Petersen C, Antico
G and Pishvaian M: Treatment patterns and outcomes in pancreatic
cancer: Retrospective claims analysis. Cancer Med. 9:3463–3476.
2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Kolbeinsson HM, Chandana S, Wright GP and
Chung M: Pancreatic cancer: A review of current treatment and novel
therapies. J Invest Surg. 36(2129884)2023.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Dauer P, Nomura A, Saluja A and Banerjee
S: Microenvironment in determining chemo-resistance in pancreatic
cancer: Neighborhood matters. Pancreatology. 17:7–12.
2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Yang Z, Wang S, Yin K, Zhang Q and Li S:
MiR-1696/GPx3 axis is involved in oxidative stress mediated
neutrophil extracellular traps inhibition in chicken neutrophils. J
Cell Physiol. 236:3688–3699. 2021.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Takahashi K, Avissar N, Whitin J and Cohen
H: Purification and characterization of human plasma glutathione
peroxidase: A selenoglycoprotein distinct from the known cellular
enzyme. Arch Biochem Biophys. 256:677–686. 1987.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Chang C, Worley BL, Phaeton R and Hempel
N: Extracellular glutathione peroxidase GPx3 and its role in
cancer. Cancers (Basel). 12(2197)2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Nirgude S and Choudhary B: Insights into
the role of GPX3, a highly efficient plasma antioxidant, in cancer.
Biochem Pharmacol. 184(114365)2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Miess H, Dankworth B, Gouw AM, Rosenfeldt
M, Schmitz W, Jiang M, Saunders B, Howell M, Downward J, Felsher
DW, et al: The glutathione redox system is essential to prevent
ferroptosis caused by impaired lipid metabolism in clear cell renal
cell carcinoma. Oncogene. 37:5435–5450. 2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Wei J, Xie Q, Liu X, Wan C, Wu W, Fang K,
Yao Y, Cheng P, Deng D and Liu Z: Identification the prognostic
value of glutathione peroxidases expression levels in acute myeloid
leukemia. Ann Transl Med. 8(678)2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Liu Q, Bai W, Huang F, Tang J and Lin X:
Downregulation of microRNA-196a inhibits stem cell self-renewal
ability and stemness in non-small-cell lung cancer through
upregulating GPX3 expression. Int J Biochem Cell Biol.
115(105571)2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Worley BL, Kim YS, Mardini J, Zaman R,
Leon KE, Vallur PG, Nduwumwami A, Warrick JI, Timmins PF, Kesterson
JP, et al: GPx3 supports ovarian cancer progression by manipulating
the extracellular redox environment. Redox Biol.
25(101051)2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Cai M, Sikong Y, Wang Q, Zhu S, Pang F and
Cui X: Gpx3 prevents migration and invasion in gastric cancer by
targeting NFкB/Wnt5a/JNK signaling. Int J Clin Exp Pathol.
12:1194–1203. 2019.PubMed/NCBI
|
|
15
|
Hu Q, Chen J, Yang W, Xu M, Zhou J, Tan J
and Huang T: GPX3 expression was down-regulated but positively
correlated with poor outcome in human cancers. Front Oncol.
13(990551)2023.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Chandrashekar DS, Karthikeyan SK, Korla
PK, Patel H, Shovon AR, Athar M, Netto GJ, Qin ZS, Kumar S, Manne
U, et al: UALCAN: An update to the integrated cancer data analysis
platform. Neoplasia. 25:18–27. 2022.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Li C, Tang Z, Zhang W, Ye Z and Liu F:
GEPIA2021: Integrating multiple deconvolution-based analysis into
GEPIA. Nucleic Acids Res. 49 (W1):W242–W246. 2021.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Győrffy B: Discovery and ranking of the
most robust prognostic biomarkers in serous ovarian cancer.
Geroscience. 45:1889–1898. 2023.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zeng S, Pöttler M, Lan B, Grützmann R,
Pilarsky C and Yang H: Chemoresistance in pancreatic cancer. Int J
Mol Sci. 20(4504)2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Song Y, Zou L, Li J, Shen ZP, Cai YL and
Wu XD: LncRNA SNHG8 promotes the development and chemo-resistance
of pancreatic adenocarcinoma. Eur Rev Med Pharmacol Sci.
22:8161–8168. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zhou Y, Wang K, Zhou Y, Li T, Yang M, Wang
R, Chen Y, Cao M and Hu R: HEATR1 deficiency promotes pancreatic
cancer proliferation and gemcitabine resistance by up-regulating
Nrf2 signaling. Redox Biol. 29(101390)2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Xu P, Yao J, He J, Zhao L, Wang X, Li Z
and Qian J: CIP2A down regulation enhances the sensitivity of
pancreatic cancer cells to gemcitabine. Oncotarget. 7:14831–14840.
2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wagner EF and Nebreda AR: Signal
integration by JNK and p38 MAPK pathways in cancer development. Nat
Rev Cancer. 9:537–549. 2009.PubMed/NCBI View
Article : Google Scholar
|
|
25
|
Kawasaki S, Ohtsuka H, Sato Y, Douchi D,
Sato M, Ariake K, Masuda K, Fukase K, Mizuma M, Nakagawa K, et al:
Silencing of LRRFIP1 enhances the sensitivity of gemcitabine in
pancreatic cancer cells by activating JNK/c-Jun signaling.
Pancreatology. 21:771–778. 2021.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Shi J, Yang X, Kang Q, Lu J, Denzinger M,
Kornmann M and Traub B: JNK inhibitor IX restrains pancreatic
cancer through p53 and p21. Front Oncol. 12(1006131)2022.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Cai J, Du S, Wang H, Xin B, Wang J, Shen
W, Wei W, Guo Z and Shen X: Tenascin-C induces migration and
invasion through JNK/c-Jun signalling in pancreatic cancer.
Oncotarget. 8:74406–74422. 2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Meggiato T, Calabrese F, De Cesare CM,
Baliello E, Valente M and Del Favero G: C-JUN and CPP32 (CASPASE 3)
in human pancreatic cancer: Relation to cell proliferation and
death. Pancreas. 26:65–70. 2003.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Kalli M, Li R, Mills GB, Stylianopoulos T
and Zervantonakis IK: Mechanical stress signaling in pancreatic
cancer cells triggers p38 MAPK- and JNK-dependent cytoskeleton
remodeling and promotes cell migration via Rac1/cdc42/Myosin II.
Mol Cancer Res. 20:485–497. 2022.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Ichimaru Y, Sano M, Kajiwara I, Tobe T,
Yoshioka H, Hayashi K, Ijichi H and Miyairi S: Indirubin 3'-oxime
inhibits migration, invasion, and metastasis invivo in mice bearing
spontaneously occurring pancreatic cancer via blocking the RAF/ERK,
AKT, and SAPK/JNK pathways. Transl Oncol. 12:1574–1582.
2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Liu M, Zhang Y, Yang J, Cui X, Zhou Z,
Zhan H, Ding K, Tian X, Yang Z, Fung KA, et al: ZIP4 increases
expression of transcription factor ZEB1 to promote integrin α3β1
signaling and inhibit expression of the gemcitabine transporter
ENT1 in pancreatic cancer cells. Gastroenterology. 158:679–692.e1.
2020.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Takahashi R, Hirata Y, Sakitani K, Nakata
W, Kinoshita H, Hayakawa Y, Nakagawa H, Sakamoto K, Hikiba Y,
Ijichi H, et al: Therapeutic effect of c-Jun N-terminal kinase
inhibition on pancreatic cancer. Cancer Sci. 104:337–344.
2013.PubMed/NCBI View Article : Google Scholar
|