Introduction
Despite the successful production of cloned animals,
including sheep (1,2), cattle (3–7),
goats (8), pigs (9–11)
and mice (12–14), somatic cell nuclear transfer
(SCNT) is still hampered by extremely low efficiency. Early
embryonic and fetal loss, stillbirth, and postnatal loss and
phenotypic abnormities are commonly reported (12,15–18). The fetal losses and abnormalities
may be associated with abnormal placental development (17). Most cloned cows and mice lost
prenatally and postnatally are associated with enlarged and
dysfunctional placentas (17,19). It was recently proposed that the
high rates of developmental failure and abnormalities may be caused
by epigenetic alterations (20,21). After SCNT, the donor nucleus
requires epigenetic reprogramming to re-enter a totipotent ground
state (22), suggesting that the
donor nucleus must cease its own program of gene expression and
restore the particular program of embryonic expression necessary
for normal development. If the reprogramming process is
inefficient, embryogenesis will be abnormal (23). Poor epigenetic reprogramming in
early-cleavage embryos results in dysregulation of gene expression
and abnormal proteins accumulate that can disrupt normal embryonic
or fetal development. Indeed, along with the low survival rate,
various disease phenotypes have been observed in cloned animals,
including circulatory distress, placental edema, hydrallantois,
respiratory problems, immune dysfunction and brain malformation
(17,18,24). In theory, therefore, partial
reprogramming can result in a range of outcomes, including abnormal
phenotypes or lethality at various stages of development.
Epigenetic modifications of the genome include
changes in genomic methylation, the acetylation state of histones
and the remodeling of chromatin. DNA methylation regulates the
function of the genome by affecting tissue-specific gene
expression, cell differentiation, genomic imprinting, X chromosome
inactivation, regulation of chromatin structure, carcinogenesis and
aging (25). Recently, Kang et
al (26) reported that
methylation levels of repeated regions and unique sequences were
much higher in cloned morulae and blastocysts than in normal
embryos and resembled the methylation levels in the donor cell
genome. Interphase nuclei in cloned bovine morulae also contain
high methylation levels (27),
indicating that reprogramming is deficient in most cloned
preimplantation embryos and demethylation seems to be particularly
inefficient. The selective demethylation pattern of developmentally
important genes was confirmed in SCNT embryos. Unequal methylation
was maintained between the inner cell mass and trophectoderm
regions, thereby resulting in placental dysfunction in cloned
animals (28).
Genomic imprinting is an epigenetic mechanism by
which related genes are inherited in a parental-specific manner
(29). Imprinted genes have roles
in prenatal growth, development of the germ line and embryo and
behavior, as well as being implicated in human disease (30). The evolution of genomic imprinting
must have required oocytes to create and maintain the epigenetic
asymmetry of donor cells and the importance of the oocyte cytoplasm
can be seen in the parent-of-origin effects on interspecific
hybrids of the deer mouse (31).
The disruption of imprinting may be due to nuclear to
nuclear-cytoplasmic incompatibility between the nucleus of the
donor and the cytoplasm of the recipient in these crosses. Cloning
inefficiency also contributes to another incompatibility (32): reduced fetal methylation and
repression of ovine IGF2R is associated with fetal overgrowth.
Preimplantation embryo procedures may thus be vulnerable to
epigenetic alterations in imprinted genes. Kato et al
(33) has shown that
transplantation of imprint-free primordial germ cell nuclei into
oocytes results in embryonic lethality (33) and partially abnormal
extraembryonic tissues resulting from the inappropriate silencing
or activation of imprinted genes.
Cloning by SCNT is a multiple-step procedure and
many factors affect cloning efficiency, such as donor cell type and
age, the phase of the cell cycle that the donor cell is in and the
SCNT procedure itself. Although the SCNT procedure has been
improved, developmental defects are still consistently observed in
cloned animals. These defects may be related to methodological
faults including squeezing to remove nuclear, activation and
fusion, which cause the instability of critical reprogramming.
We have generated three cloned pigs that developed
wrinkles on their bodies, a phenotype typical of aged animals, at a
relatively young age. Here, we report that this phenotype is due to
a genetic abnormality of the donor cells rather than to senescence
or epigenetic abnormalities during reprogramming.
Materials and methods
Preparation of donor cells
Two types of fetal fibroblasts [named s-pig fetal
fibroblasts (sPFFs) and w-pig fetal fibro-blasts (wPFFs)] were
isolated from pig fetuses (on Day 30 of gestation) obtained from a
slaughterhouse on different days as it has been described before
(34). Briefly, collected fetuses
were washed 3 times with Ca2+- and Mg2+-free
phosphate buffered saline (PBS). The head and internal organs were
discarded by scooping them out with two watchmaker’s forceps. After
2 washes with PBS (pH 7.4), the carcass was minced with a surgical
blade in a 100π culture dish (Becton-Dickinson, Lincoln Park, NJ).
The minced fetal tissues were dissociated in Dulbecco’s modified
Eagle’s medium (DMEM) supplemented with 0.25% (w/v) trypsin and 1
mM EDTA for 1–2 h. The trypsinized cells were then washed once in
PBS by centrifugation at 300 x g for 10 min and subsequently seeded
into 100-mm culture dishes. The cells were cultured for 6–8 days in
DMEM supplemented with 10% (v/v) fetal bovine serum (FBS), 1% (v/v)
nonessential amino acids and 10 μg/ml penicillin-streptomycin
(Sigma-Aldrich, St. Louis, MO) in a humidified atmosphere of 5% CO2
and 95% air. After the unattached clumps of cells and explants were
removed, the attached cells were further cultured until confluent,
subcultured at intervals of 5–7 days by trypsinization and stored
after 2 passages in liquid nitrogen at −196°C. The freezing medium
consisted of 80% (v/v) DMEM, 10% (v/v) dimethylsulfoxide (DMSO;
Sigma-Aldrich) and 10% (v/v) FBS. Before SCNT, frozen fetal
fibroblasts (cultured for 3–5 passages) were thawed for 3–5 days in
DMEM supplemented with 0.5% FBS. Individual cells were retrieved
from the monolayer by trypsinization for 30 sec and used as donor
cells for SCNT.
Production of cloned pigs
The nuclear transfer of donor cells to enucleated
porcine oocytes was performed to produce cloned piglets as
previously described (35,36).
wPFFs produced normal cloned pigs when used as donor nuclei and
sPFFs produced the wrinkled cloned pigs. A parentage analysis was
performed on the piglets obtained by SCNT and the surrogate
recipient females to confirm the identity of the donor cells used
for nuclear transfer (35,36).
The DNA was extracted from ear punches or tail clippings obtained
from each newborn piglet and recipient female. DNA was also
extracted from donor cells. Eight porcine DNA microsatellite
markers (S0086, S0230, SW902, S0007, S0313, SW61, S0005 and S0164)
were used to confirm the genetic identity.
Measurement of telomere length
Telomere length was determined by mean terminal
restriction fragment (TRF) length analysis using a Telo TAGGG
Telomere Length Assay kit (Roche Diagnostics GmbH, Mannheim,
Germany). The genomic DNA (10 μg) isolated from donor cells or ear
tissues of cloned pigs was digested with the restriction enzymes
HinfI (New England Biolabs, Inc., Beverly, MA) and
RsaI (New England Biolabs, Inc.). Undigested or digested
genomic DNA samples were fractionated by 0.6% agarose gel
electrophoresis at 30 V/cm for 5 h. Gels were denatured,
neutralized and transferred to a positively charged nylon membrane
(Hybond+; Amersham Pharmacia Biotech, Oakville, Canada). The
membranes were prehybridized in 40 ml of DIG Easy Hyb (Roche
Diagnostics GmbH) for 2 h at 42°C and then hybridized in 10 ml of
DIG Easy Hyb containing 50 pmol of an end-labeled telomere-specific
probe for 16 h at 42°C. Membranes were washed 3 times in 50 ml of
0.5X standard saline citrate (SSC; 1X SSC: 0.15 M NaCl, 0.015 M
sodium citrate) for 15 min at room temperature. The signals were
visualized by chemiluminescence using a DIG Luminescent Detection
kit (Roche Diagnostics GmbH) and exposed to X-ray film (Hyperfilm;
Amersham Pharmacia Biotech). The signals were scanned and analyzed
using Gel Doc software (Bio-Rad, Hercules, CA) by taking the mean
value of the largest and smallest values.
Cell culture and induction of cellular
senescence
sPFFs were routinely cultured until replicative
senescence as previously described (38). The slowly growing cultures were
replenished with fresh DMEM supplemented with 10% FBS every 4 days.
Confluent sPFFs were exposed to 700 μM H2O2
in DMEM containing 10% FBS for 5 days as a stressor. After
H2O2 stress for 5 days, the sPFFs were rinsed
with PBS and incubated in DMEM with 10% FBS.
Galactosidase (Gal) staining in skin
tissues
Tissue was collected from the ear skin of cloned
pigs, rapidly frozen in liquid nitrogen and mounted in OCT compound
(Fisher Scientific). Thin sections (10 μm) were cut and mounted
onto glass slides. The sections were washed in PBS, fixed for 3–5
min at room temperature in 3% formaldehyde, washed again and
incubated at 37°C with fresh senescence associated Gal (SA-Gal)
stain solution [1 mg of 5-bromo-4-chloro-3-indoly-D-galactoside
(X-Gal)/ml of solution containing 40 mM citric acid/sodium
phosphate (pH 6.0), 5 mM potassium ferrocyanide, 5 mM potassium
ferricyanide, 150 mM NaCl, 2 mM MgCl2]. Stain signals
were evident after 2–4 h and maximal after 12–16 h.
Bisulfite treatment of genomic DNA
All procedures for genomic DNA preparation have been
described previously (37). The
genomic DNA was digested with HindIII enzyme in a 20-μl
reaction volume for 16 h, then denatured with 0.3 N NaOH. Bisulfite
modification (38) was initiated
by adding 235 μl freshly made 5 M sodium bisulfite (pH 5, Sigma)
and 13.5 μl of 10 mM hydroquinone. The reaction mixture was
incubated at 55°C for 16 h in the dark. The bisulfite-treated
genomic DNA was recovered using a Wizard DNA purification kit
(Promega). Desulfonation was performed by adding 0.3 N NaOH and
incubating the solution at 37 °C for 30 min. Following
precipitation, the DNA was resuspended in 20 μl of distilled
water.
Polymerase chain reaction (PCR)
amplification, cloning and sequencing
To amplify the satellite region (GenBank™
Z75640) and the PRE-1 sequence (GenBankTM X64127, Y00104 and
AJ251914), we performed PCR with ear tissues using 2 μl of the
bisulfite-converted genomic DNA as a template. The primer set used
for the satellite region was 5′-TTTGTAGAATGTAGTTTTTAGAAG-3′ and
5′-AAAATCT AAACTACCTCTAACTC-3′. The amplification cycle was 45
cycles at 94 °C for 60 sec, 55°C for 60 sec and 72°C for 20 sec and
the amplification was finished by incubation at 72°C for 10 min.
For amplification of the PRE-1 sequence (41), a primer set of
5′-TTAACRAATCCRACTAAAAACCAT A-3′ and
5′-GTTGGTTTATMTTAGAGTTATAGTAA-3′ was designed. The amplification
cycle was 45 cycles at 94°C for 60 sec, 52°C for 60 sec and 72°C
for 20 sec, and the amplification was finished with one cycle of
72°C for 10 min. The PCR products after elution were cloned into a
TOPO TA cloning vector (Invitrogen Life Technologies). Individual
clones were sequenced using an automatic sequencer (ABI PRISM 377;
Applied BioSystems).
RNA isolation and semiquantitative
reverse transcriptase PCR (RT-PCR)
Tissues from normally fertilized pigs and cloned
pigs were collected and minced. Total-RNAs were extracted using
TRIzol Reagent (Invitrogen Life Technologies) and used for the
synthesis of a first-strand cDNA using a First-Strand cDNA
Synthesis kit (Amersham Pharmacia Biotech) after heating to 60°C
for 10 min. The RT-PCR was semiquantitative in the linear portion
of the exponential amplification for each gene. The oligonucleotide
primers for the imprinted genes are shown in Table I.
 | Table IOligo sequences of the primers used
for RT-PCR of imprinting genes. |
Table I
Oligo sequences of the primers used
for RT-PCR of imprinting genes.
| Genes | Primer
sequences | Tm (°C) | Size (bp) |
|---|
| COPG2 |
5′-CAAGAAGGACGAGGAGTCTGGTAG-3′
5′-CACTTGATGCAGCTTCTTGGGCTT-3′ | 55 | 544 |
| SNRPN |
5′-CGGGTTTTGGGTCTGGTGTTG-3′
5′-ATAATGCCTGGAGGTGGGGTTGC-3′ | 60 | 380 |
| MEST |
5′-CCTTGATTTCTTAGGCTTTGGCTT-3′
5′-CAGCGTTTTCCTGTACAGCTCCAA-3′ | 55 | 600 |
| GNAS1 |
5′-GCCAACAAAAAGATCGAGAAGCAG-3′
5′-CCACCTGGAACTTGGTCTCAAAGA-3′ | 55 | 582 |
| UBE3A |
5′-GCATCTAATAGAACGCTACTACCA-3′
5′-TTCCAGATATTCAGGACTGTGGAG-3′ | 50 | 818 |
Statistical analysis
The experiments were repeated at least 3 times with
different sets of nuclear donor cells, ear tissue samples, in
vitro fertilized or SCNT blastocysts (a total of 90
blastocysts). The data were subjected to one-way analysis of
variance and the protected least significant different test using
general linear models in a statistical analysis system (SAS,
version 8.1) program. The data are presented as means ± SD.
Differences among the treatments were determined to be significant
where P-value <0.05.
Results
We produced cloned pigs using two types of fetal
fibroblasts (sPFFs and wPFFs, which differed by the day on which
the fetuses were obtained) as nuclear donor cells (35). At birth, the cloned pigs were
healthy and were of normal weight, and the surface of skin and
color appeared normal. At 2 years, however, we observed many
wrinkles on the bodies of three cloned pigs the only ones obtained
from sPFFs, which did not appear on age-matched normal pigs
(Fig. 1B). The face around the
nose and forehead was most wrinkled, the neck and ears were easily
bent, and a line of wrinkles was present around the neck, ears,
tail and legs. In contrast, age-matched normal pigs and normal
cloned pigs showed normal smooth skin without wrinkles on these
parts of the body (Fig. 1).
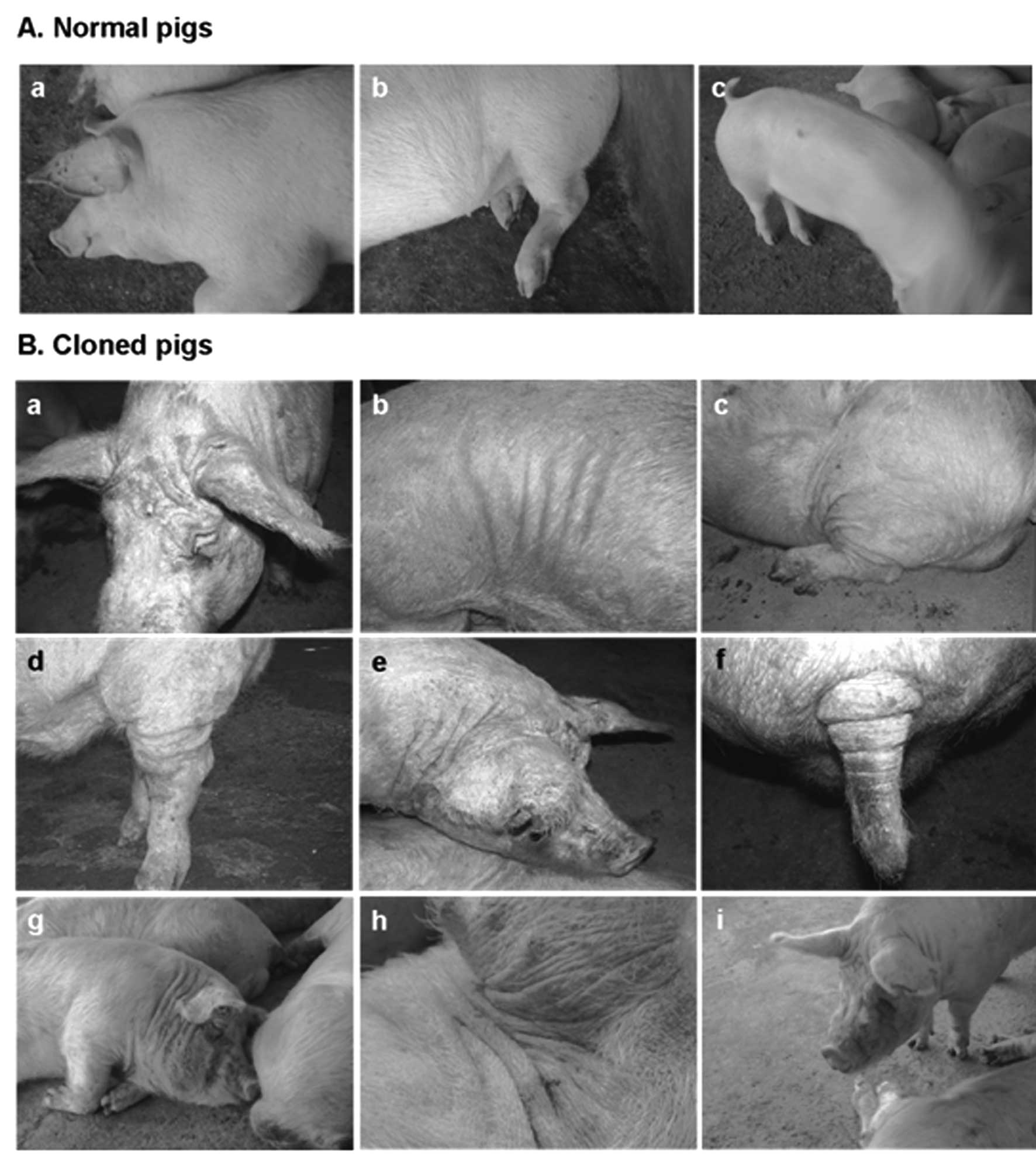 | Figure 1Phenotypic abnormalities found in 3
cloned pigs. To produce cloned piglets, we transferred the nuclei
of donor cells to enucleated porcine oocytes. The normal cloned
pigs were generated using wPFFs as donor cells and the wrinkled
cloned-pigs were produced from sPFFs. Compared to age-matched
normal (naturally fertilized) pigs, 3 pigs cloned by SCNT from
sPFFs had many wrinkles on their bodies, including face, neck, legs
and tail. (A) Age-matched normal pigs used as controls. At 2 years
old, the normal pigs had smooth skin without wrinkles on the parts
of the body that were wrinkled in the wrinkled cloned pigs. (B)
Abnormal cloned pigs. To show distinct phenotypes, pictures taken
(a–f) without or (g–i) with a flash are shown. (a, b, d, f and h)
Also, the specific parts of the body are shown at high
magnification to show the details of the specific
abnormalities. |
One of the important indications of senescence and
successful reprogramming in SCNT animals is the telomere length. In
our previous report (39), we
measured telomere length in 15 cloned piglets, including the three
wrinkled piglets and normal cloned pigs, and compared the lengths
with those in donor somatic cells. The telomeres were longer in
1-day-old cloned piglets than in the nuclear donor cells and
age-matched controls, suggesting that nuclear reprogramming resets
the cellular age of donor cells after SCNT. Here, to determine
whether the wrinkled phenotype is associated with the status of the
genome or the age of the pigs, we compared the telomere lengths of
the wrinkled and normal cloned pigs with those of age-matched
normal pigs. First, telomere length was measured in adult pigs and
newborn piglets to determine the normal reduction in length that
occurs with biological and chronological aging. As expected, the
2-year-old adult pigs had shorter telomeres than the 1-day-old
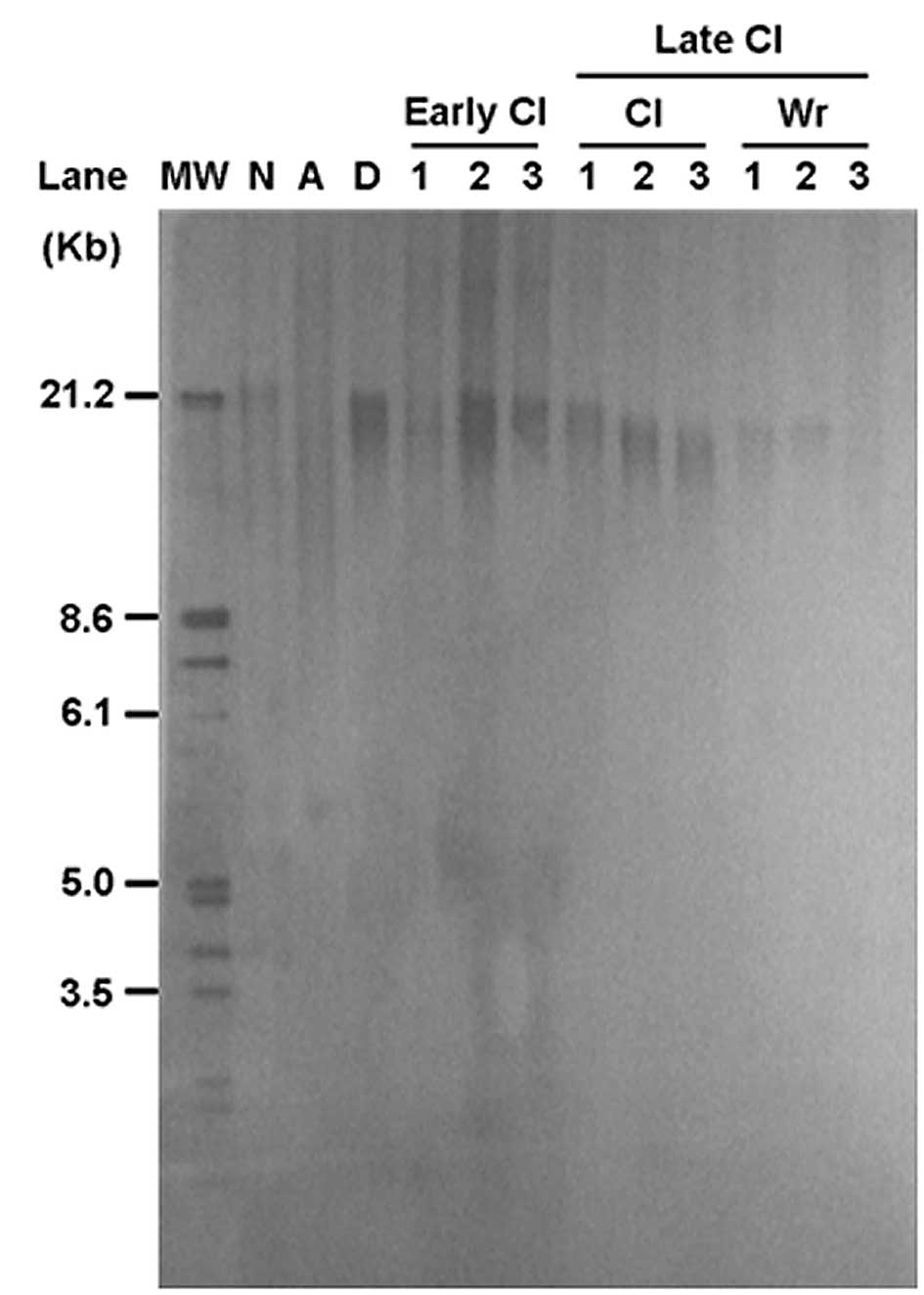
newborn piglets and the donor fibroblast cells. The telomeres were
longer in the early cloned piglets (mean TRF length = 23.5±1.6 kb)
than in donor fetal fibroblasts (mean TRF length = 21.3±1.1 kb;
Fig. 2); however, telomere length
decreased over time in both normal and wrinkled cloned pigs. Normal
cloned pigs >1.5-years-old had a mean telomere length of
19.6±2.3 kb, whereas wrinkled cloned pigs at the same age had
shorter telomeres (mean TRF length = 18.7±0.6 kb) (P<0.05).
Cloned pigs with or without wrinkles had shorter telomeres than
age-matched normal pigs.
Along with telomere length analysis, we examined
whether SA-β-gal activity, a biomarker of senescence, was present
in our wrinkled cloned pigs. We first confirmed SA-β-gal staining
in cells with replicative senescence (RS) or stress-induced
premature senescence (SIPS). The SIPS-associated morphology of PFFs
was obtained by treating cells with 700 μM
H2O2 for 5 days. For RS morphology, the cells
were passaged every 7 days for >15 passages. The SIPS and RS
cells had larger cell surfaces than the normal and early-passage
PFFs (Fig. 3A). After 10
passages, the cell growth declined markedly (data not shown). The
senescent pig fibroblasts expressed SA-β-Gal (shown as blue
staining), while the early-passage cells did not show SA-β-Gal
activity.
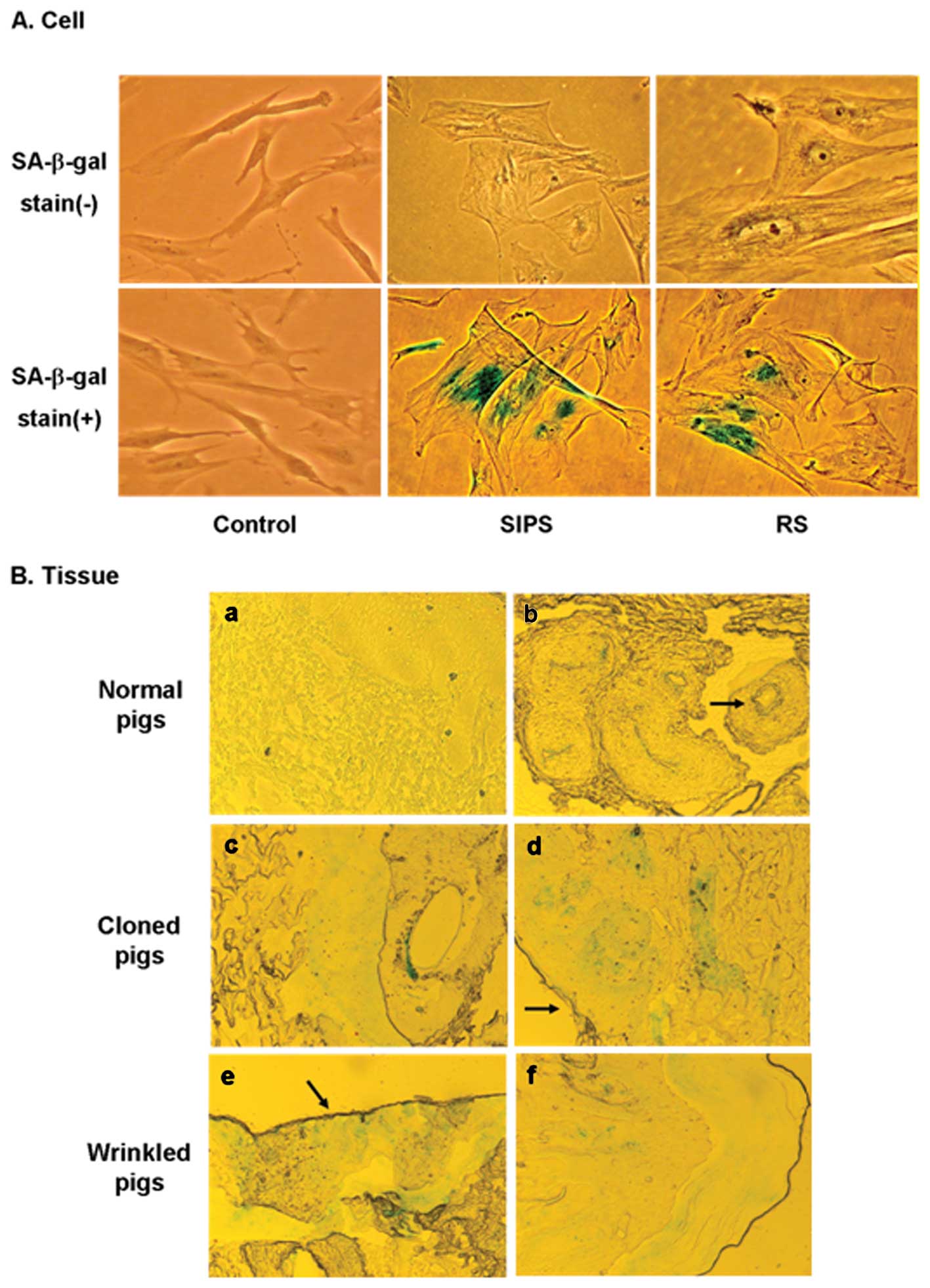 | Figure 3Senescence-associated β-galactosidase
activity in cells and the tissues of cloned pigs. (A) SA-β-gal in
cells. SA-β-gal stain (−) refers to unlabeled cells and SA-β-gal
stain (+) refers to cells labeled with SA-β-gal staining.
Replicative senescence (RS) of pig fetal fibroblast cells (PFFs)
was induced after long-term cell culture, and stress-induced
premature senescence (SIPS) was induced by treatment with
subcytotoxic doses of HB2BOB2B for 5 days.
Early-passage PFF cells were used as a control. The
SA-β-gal–positive cells are shown in the SIPS and RS cells in the
lower panel. The cells were photographed at x100 magnification. (B)
SA-β-gal staining in the ear tissues of cloned pigs. Ear tissues
were sectioned, stained for SA-β-gal and photographed at
magnification x40. All 6 cloned pigs were positive for SA-β-gal
activity in the dermis and epidermis (c,d, normal cloned pigs; e,f,
wrinkled cloned pigs). Keratinous layers were also positive for
SA-β-gal in both kinds of cloned pigs (arrows in d,e). Keratinous
layers contain many dead and aged cells. Moreover, SA-β-gal
activity was always seen in the hair follicles and is frequently
associated with sebaceous glands and eccrine glands and ducts. (a)
However, no staining of SA-β-gal was observed in age-matched normal
pigs, (arrow in b) except in the hair follicles stained as a
positive control. |
To examine whether senescent cells accumulate in
vivo after SCNT, we stained ear tissues from the wrinkled and
normal cloned pigs, as well as age-matched normal pigs, for
SA-β-Gal activity when they were >1.5-years-old. We examined the
sections for both the frequency and identity of the positive cells.
The hair follicles and associated sebaceous glands and ducts were
used as positive controls for SA-β-Gal activity because these
structures were stained with consistent frequency and intensity
independent of age (Fig. 3Bb and
c) (40). In the dermis and
epidermis of both the wrinkled and normal cloned pigs, SA-β-Gal
staining was sparsely and randomly distributed (Fig. 3Bc–f). The keratinous layers in the
most peripheral region of sectioned tissue were also positive in
both types of cloned pigs (Fig. 3Bd
and e). In contrast, except for hair follicles, no blue
precipitation was detected in the age-matched normal pigs (Fig. 3Ba and b).
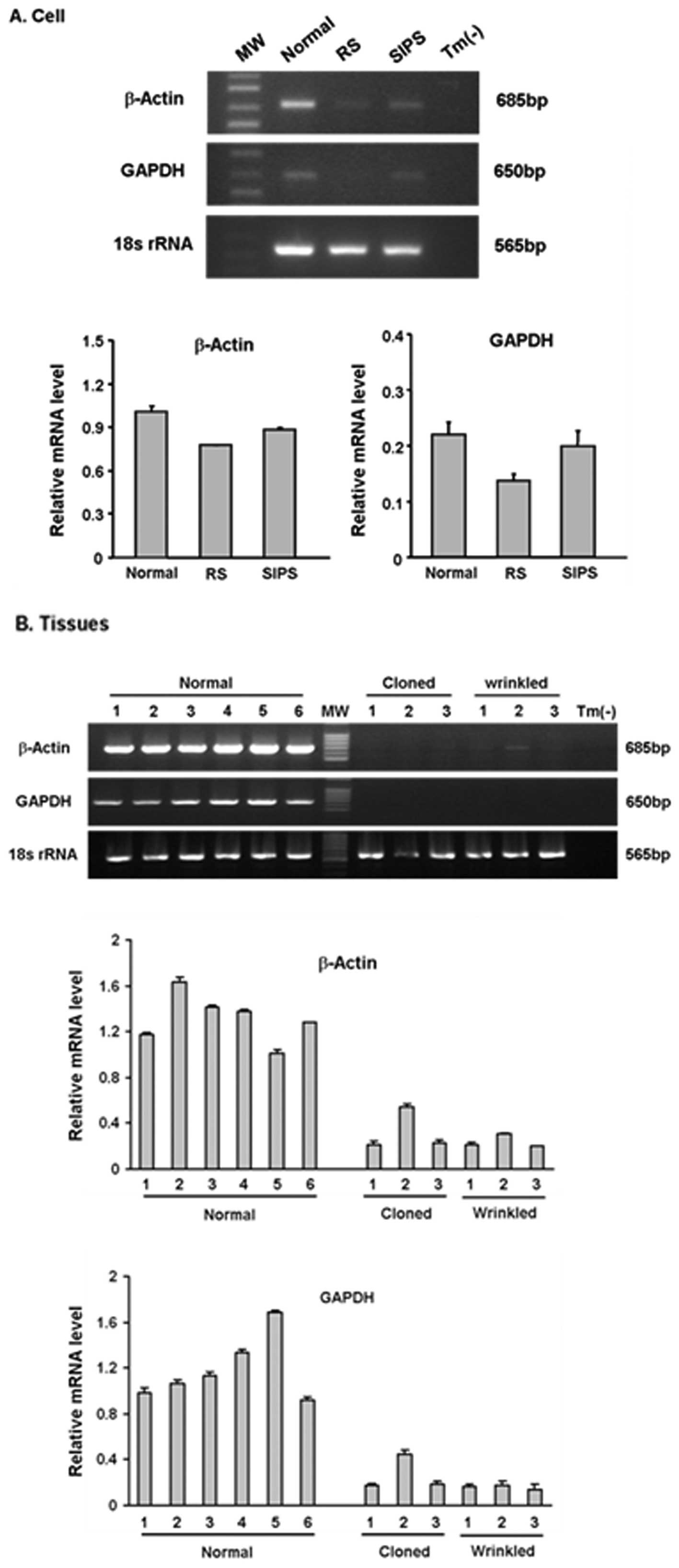
Expression levels of GAPDH and β-actin change with
age in several cell and tissue types (41,42); accordingly, they are not always a
proper internal control for mRNA analyses regarding aging. We
therefore used the level of 18S rRNA as a reference and the mRNA
levels of GAPDH and β-actin as biomarkers for senescence. We first
performed semiquantitative RT-PCR with SIPS- and RS-associated
cells. Less β-actin mRNA and GAPDH mRNA was observed in the RS and
SIPS cells than in the control cells (Fig. 4A). We then determined the mRNA
levels of these genes in the wrinkled and normal cloned pigs.
Tissues from age-matched normal pigs were used as a control. The
normal and wrinkled cloned pigs displayed significantly less
β-actin mRNA than the age-matched pigs (Fig. 4B). GAPDH expression was also
diminished in the cloned pigs. Although there were individual
differences within each group, similar expression patterns were
detected within the normal and wrinkled cloned groups.
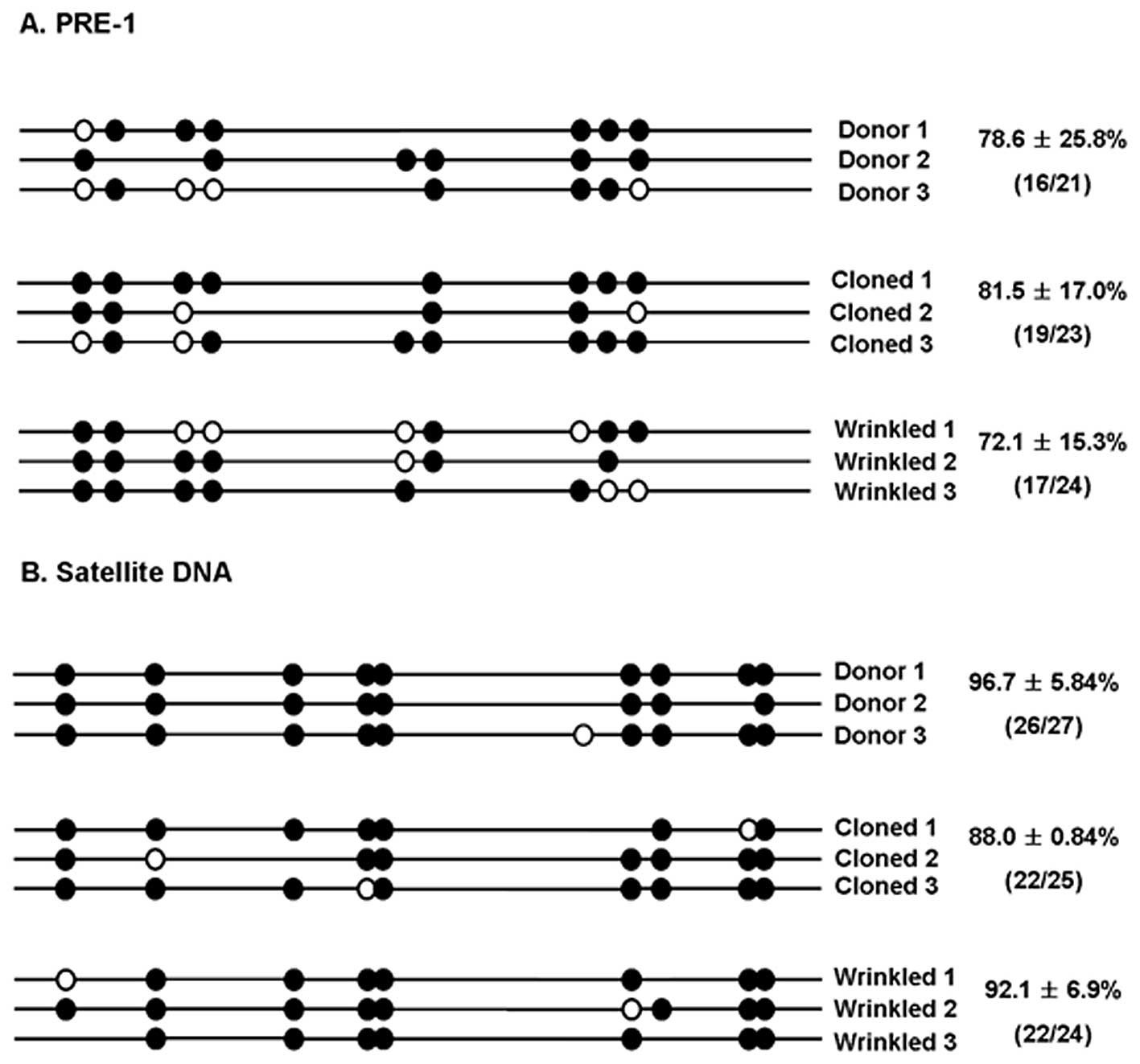
We examined the methylation status of normal and
wrinkled cloned pigs in order to determine whether the
reprogramming process occurred during SCNT and whether the
methylation pattern could contribute to the phenotypic abnormality.
The pig euchromatic PRE-1 repetitive sequence, a subfamily of pig
short interspersed element sequences, was chosen as a target for
the methylation analysis. We characterized methylation patterns of
the PRE-1 sequence using the bisulfite method. Bisulfite causes
deamination of unmethylated cytosines to uridine, thereby allowing
the discrimination of unmethylated and methylated cytosine residues
through sequencing. Sixteen of the 21 CpG sites (78.6±25.8%) in the
PRE-1 sequence were methylated in the donor cells (Fig. 5A). Hypomethylation was detected in
the three wrinkled cloned pigs, where only 17/24 sites (72.1±15.3%)
were methylated. Then we examined the methylation patterns of
satellite DNA sequences in pigs. A tiny difference in the
methylation status of the satellite region was observed between the
donor cells (96.7±5.84%) and wrinkled cloned pigs (92.1±6.9%).
Since the donor cells used to produce normal cloned pigs were not
available in this study, the methylation status in normal cloned
pigs could not be directly compared with the donor cells. However,
in normal cloned pigs, methylation was observed in 81.5±17.0% and
88.0±0.84% in both eucromatic PRE-1 repetitive sequences and
centromeric satellite DNA. The status of the wrinkled cloned pigs
was similar to that of the donor cells (Fig. 5).
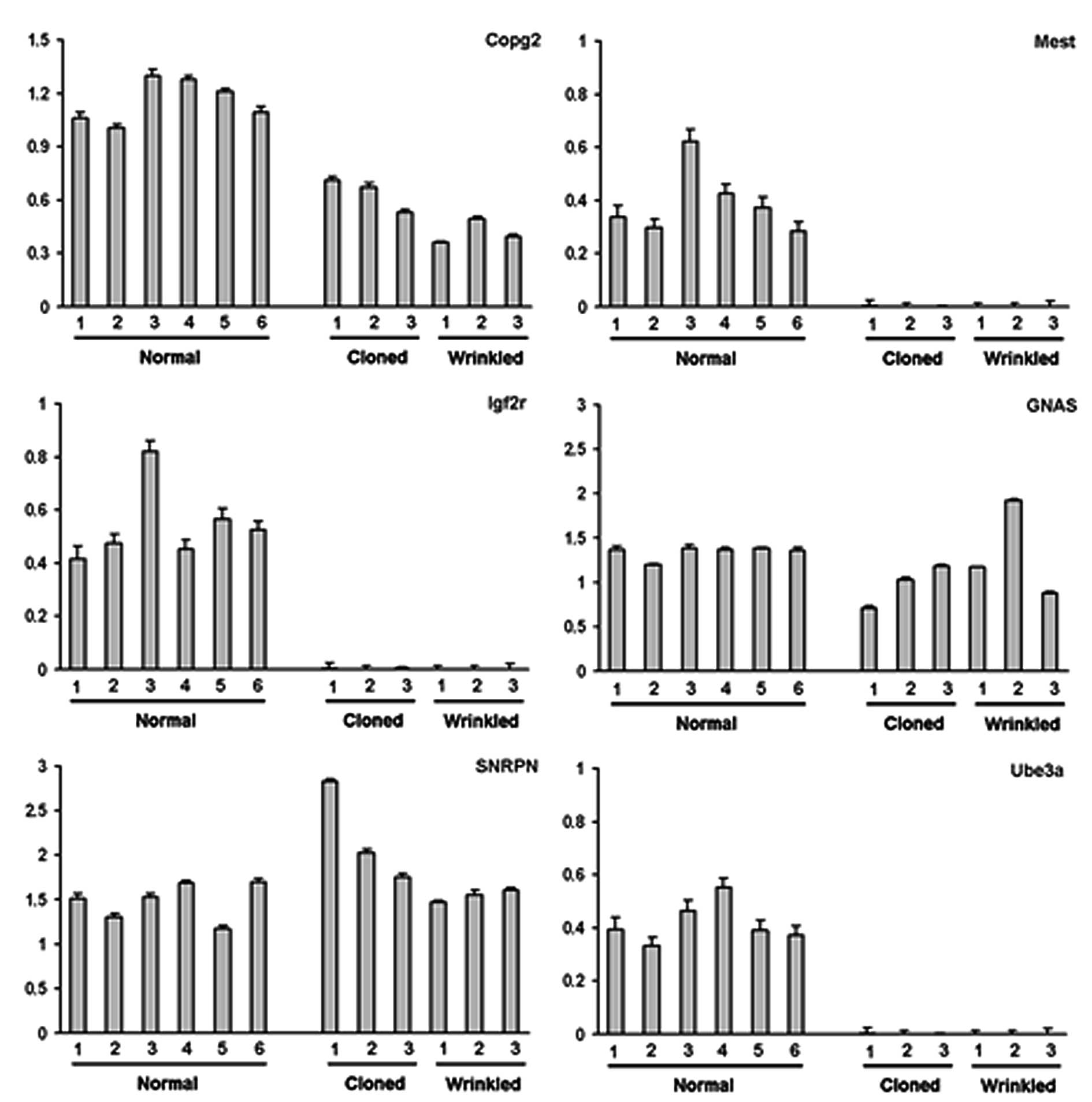
Because parent-specific monoallelic expression of
imprinted genes is critically important for normal embryo
development (43), we determined
the allelic pattern of imprinted genes in the wrinkled clones as
well as in normal clones. However, few sequences of imprinted genes
have been reported in the pig genome database. Therefore, we
selected several imprinted genes whose DNA sequences are homologous
between human and mouse. For PCR amplification, the primer set for
each imprinted gene was designed from the consensus sequence and
PCR products were directly sequenced. The PCR conditions, including
annealing temperature, size of PCR products, primer sequences and
the names of newly selected genes, are shown in Table I. Of the six imprinted genes,
COPG2, SNRPN, MEST, GNAS1, UBE3A and IGF2R, all but IGF2R were
newly identified in the pig. Using this information, relative
transcript levels in the tissues of normal (control) pigs and the
normal and wrinkled cloned pigs were estimated by quantitative
RT-PCR (P<0.05). Four of the six imprinted genes had
considerably reduced transcript levels in the two cloned groups
compared to the normal age-matched pigs; however, the mRNA levels
of SNRPN and GNAS were within the range of control levels. Although
there were individual variations within each group, it appeared
that there were lower amounts of Igf2R, Mest and Ube3a transcripts
in the wrinkled and normal cloned pigs than in the normal pigs.
Discussion
We used SCNT to derive cloned pigs from two sets of
donor cells, sPFFs and wPFFs. Although at birth the cloned pigs
appeared normal, the pigs cloned from sPFFs started to show many
deep wrinkles in their faces and bodies 2 years after birth. In a
previous study, we reported on a cloned pig derived from fetal
fibroblasts of an inbred miniature pig that had anal atresia
(44). However, we could not
determine the cause of this malformation with just one case. Many
studies have reported that developmental aberrancies are due to
fully unsuccessful reprogramming from somatic cell nuclei to
embryonic genomes (20) or to the
importance of donor cell selection and its preparation (1,6,14,45–47). Here, we investigated whether the
causes of the abnormal skin phenotype (body wrinkles) were derived
from a genetic problem caused by donor specificity or from an
epigenetic problem during reprogramming. First, we compared
telomere length in wrinkled pigs with that of normal pigs and
normal (unwrinkled) cloned pigs. In our previous studies, telomere
length increased significantly in cloned pigs at birth. However,
there was a dramatic shortening of the telomeres in the cloned pigs
during growth and the time spent for SCNT (Fig. 2). These data suggest that aging
may accelerate more quickly in the wrinkled cloned pigs and that,
although there is no visible phenotype at early stages after birth,
some error from the nuclear transfer is expressed during growth.
Because telomere length is strongly influenced by many genetic and
other factors, by itself it can be a poor indicator of aging or
cell viability (48–50). To confirm that the wrinkled
phenotype is a phenotype of aging, we further examined biomarkers
for senescence (aging), including senescence-associated
β-galactosidase (SA-β-gal) activity and the expression of GAPDH and
β-actin. This specific type of the β-galactosidase enzyme is not
expressed in young proliferating fibroblasts (54,55). In our wrinkled cloned pigs and
normal cloned pigs, positive SA-β-gal staining was clearly detected
in the dermis and the epidermis. The outermost layer and hair
follicles of sectioned tissue, known as the keratinous layers, were
always positive for SA-β-gal in the age-matched normal pigs
(Fig. 3Bb–e, black arrow). GAPDH
and β-actin expression were also used as biomarkers for aging. Lowe
et al (41) reported that
GAPDH expression in skeletal muscles diminishes with age. In
addition, Moe et al (42)
suggested that GAPDH, β-actin and HPRT, but not 18S rRNA, are
differentially expressed at different developmental processes. The
decreased mRNA levels of GAPDH and β-actin provide genetic evidence
for senescence in our cloned pigs, in addition to the morphological
evidence of the abnormal wrinkled phenotype. However, because these
markers were observed in both kinds of cloned pigs, the appearance
of these senescence markers are not due to abnormal aging that is
expressed in wrinkling of the skin but are due to the SCNT
procedures and genetic material from donor cells.
It is conceivable that an aberrant sequence of
reprogramming events, especially DNA methylation, leads to
developmental problems in cloned animals. Previously, Kang et
al (26) reported that the
methylation status of cloned donor genomes during cleavage was
similar to that detected in fertilized counterparts (26). Interestingly, depending on their
confirmed sequences, the methylation status of the PRE-1 and
satellite DNA sequences was maintained in our wrinkled cloned pigs
and showed few changes from those of the donor cells and complete
reconstruction of genome status from donor. However, this is in
contrast to observations in many previous studies. McKay et
al (51) implicated aberrant
DNA methylation in the pathogenesis of a number of diseases
associated with aging, cancer and diseases of the cardiovascular
and neurological systems (51).
Accordingly, incomplete epigenetic changes in the donor genome
alone might not be sufficient for the wrinkled phenotype of our
cloned pigs and other complex factors may be required for this
abnormality.
Given that the donor cells had identical methylation
patterns, we examined whether the wrinkled phenotype of the cloned
pigs was caused by abnormal imprinting. As shown in Fig. 6, the reduction in the mRNA levels
of Mest, Igf2R and Ube3a may be associated with the developmental
failure or abnormal phenotypes of the resultant cloned animals. In
previous reports, mutation of imprinted genes resulted in fetal
growth abnormalities in cloned animals and human patients (52,53). Importantly, a substantial
correlation was observed between the abnormal expression of any
single imprinted gene and the degree of methylation in the promoter
regions of the gene. However, the abnormalities observed in most
clones result from the cumulative dysregulation of several
imprinted genes (54). Consistent
with the idea that a single imprinted gene could be insufficient to
produce a substantial correlation with an abnormality, we showed
that the repression of several imprinted genes does not make a
critical contribution to the deep wrinkles in our cloned pigs. In
conclusion, we showed that the abnormal wrinkled phenotype in our
pigs cloned by SCNT was caused by a genetic abnormality of the
donor cells, suggesting that the differentiation status of the
donor cell affects the developmental potential of cloned animals.
Therefore, although it is also important that donor cells be
accurately reprogrammed to early-stage cells, the selection of
good-quality donor cells is the ultimate challenge for successful
cloning by SCNT.
Acknowledgements
This study was supported by a grant
from the Next-Generation BioGreen 21 Program (no. PJ008323), Rural
Development Administration, Republic of Korea.
References
|
1.
|
I WilmutAE SchniekeJ McWhirAJ KindKH
CampbellViable offspring derived from fetal and adult mammalian
cellsNature385810813199710.1038/385810a09039911
|
|
2.
|
KH CampbellJ McWhirWA RitchieI WilmutSheep
cloned by nuclear transfer from a cultured cell
lineNature3806466199610.1038/380064a08598906
|
|
3.
|
JB CibelliSL SticePJ GoluekeCloned
transgenic calves produced from nonquiescent fetal
fibroblastsScience28012561258199810.1126/science.280.5367.12569596577
|
|
4.
|
Y KatoT TaniY SotomaruEight calves cloned
from somatic cells of a single
adultScience28220952098199810.1126/science.282.5396.20959851933
|
|
5.
|
K ShigaT FujitaK HiroseY SasaeT
NagaiProduction of calves by transfer of nuclei from cultured
somatic cells obtained from Japanese black
bullsTheriogenology52527535199910.1016/S0093-691X(99)00149-110734386
|
|
6.
|
Y KK GotoS KobayashiK ImaiM Shin-nohT
TsujinoT NakanoS MatsudaS NakaneT KojimaBirth of cloned calves
derived from cultured oviductal epithelial cells of a dairy cowAnim
Sci J702432451999
|
|
7.
|
C KubotaH YamakuchiJ TodorokiSix cloned
calves produced from adult fibroblast cells after long-term
cultureProc Natl Acad Sci
USA97990995200010.1073/pnas.97.3.99010655472
|
|
8.
|
A BaguisiE BehboodiDT MelicanProduction of
goats by somatic cell nuclear transferNat
Biotechnol17456461199910.1038/863210331804
|
|
9.
|
A OnishiM IwamotoT AkitaPig cloning by
microinjection of fetal fibroblast
nucleiScience28911881190200010.1126/science.289.5482.118810947985
|
|
10.
|
IA PolejaevaSH ChenTD VaughtCloned pigs
produced by nuclear transfer from adult somatic
cellsNature4078690200010.1038/3502408210993078
|
|
11.
|
J BetthauserE ForsbergM
AugensteinProduction of cloned pigs from in vitro systemsNat
Biotechnol1810551059200010.1038/8024211017042
|
|
12.
|
T WakayamaR YanagimachiCloning of male
mice from adult tail-tip cellsNat
Genet22127128199910.1038/963210369248
|
|
13.
|
T WakayamaY ShinkaiKL TamashiroCloning of
mice to six
generationsNature407318319200010.1038/3503030111014179
|
|
14.
|
T WakayamaAC PerryM ZuccottiKR JohnsonR
YanagimachiFull-term development of mice from enucleated oocytes
injected with cumulus cell
nucleiNature394369374199810.1038/286159690471
|
|
15.
|
LE YoungKD SinclairI WilmutLarge offspring
syndrome in cattle and sheepRev
Reprod3155163199810.1530/ror.0.00301559829550
|
|
16.
|
P Chavatte-PalmerY HeymanJP RenardCloning
and associated physiopathology of gestationGynecol Obstet
Fertil28633642200011075501
|
|
17.
|
JR HillAJ RousselJB CibelliClinical and
pathologic features of cloned transgenic calves and fetuses (13
case
studies)Theriogenology5114511465199910.1016/S0093-691X(99)00089-810729073
|
|
18.
|
JR HillRC BurghardtK JonesEvidence for
placental abnormality as the major cause of mortality in
first-trimester somatic cell cloned bovine fetusesBiol
Reprod6317871794200010.1095/biolreprod63.6.178711090450
|
|
19.
|
H NiemannC WrenzyckiAlterations of
expression of developmentally important genes in preimplantation
bovine embryos by in vitro culture conditions: implications for
subsequent
developmentTheriogenology532134200010.1016/S0093-691X(99)00237-X
|
|
20.
|
WM Rideout IIIK EgganR JaenischNuclear
cloning and epigenetic reprogramming of the
genomeScience29310931098200110.1126/science.106320611498580
|
|
21.
|
XC TianReprogramming of epigenetic
inheritance by somatic cell nuclear transferReprod Biomed
Online8501508200410.1016/S1472-6483(10)61095-415151710
|
|
22.
|
JB GurdonA ColmanThe future of
cloningNature402743746199910.1038/4542910617195
|
|
23.
|
TH BestorCytosine methylation and the
unequal developmental potentials of the oocyte and sperm genomesAm
J Hum Genet6212691273199810.1086/3018919585619
|
|
24.
|
RP LanzaJB CibelliC BlackwellExtension of
cell life-span and telomere length in animals cloned from senescent
somatic
cellsScience288665669200010.1126/science.288.5466.66510784448
|
|
25.
|
W ReikER MaherImprinting in clusters:
lessons from Beckwith-Wiedemann syndromeTrends
Genet13330334199710.1016/S0168-9525(97)01200-69260520
|
|
26.
|
YK KangDB KooJS ParkTypical demethylation
events in cloned pig embryos. Clues on species-specific differences
in epigenetic reprogramming of a cloned donor genomeJ Biol
Chem2763998039984200110.1074/jbc.M10651620011524426
|
|
27.
|
W DeanF SantosM StojkovicConservation of
methylation reprogramming in mammalian development: aberrant
reprogramming in cloned embryosProc Natl Acad Sci
USA981373413738200110.1073/pnas.24152269811717434
|
|
28.
|
YK KangJS ParkDB KooLimited demethylation
leaves mosaic-type methylation states in cloned bovine
pre-implantation embryosEMBO
J2110921100200210.1093/emboj/21.5.109211867537
|
|
29.
|
SM TilghmanThe sins of the fathers and
mothers: genomic imprinting in mammalian
developmentCell96185193199910.1016/S0092-8674(00)80559-09988214
|
|
30.
|
AC Ferguson-SmithMA SuraniImprinting and
the epigenetic asymmetry between parental
genomesScience29310861089200110.1126/science.106402011498578
|
|
31.
|
PB VranaJA FossellaP MattesonT del RioMJ
O’NeillSM TilghmanGenetic and epigenetic incompatibilities underlie
hybrid dysgenesis in PeromyscusNat
Genet25120124200010.1038/7551810802670
|
|
32.
|
LE YoungK FernandesTG McEvoyEpigenetic
change in IGF2R is associated with fetal overgrowth after sheep
embryo cultureNat Genet27153154200110.1038/8476911175780
|
|
33.
|
Y KatoWM Rideout 3rdK HiltonSC BartonY
TsunodaMA SuraniDevelopmental potential of mouse primordial germ
cellsDevelopment12618231832199910101117
|
|
34.
|
GS LeeSH HyunHS KimImprovement of a
porcine somatic cell nuclear transfer technique by optimizing donor
cell and recipient oocyte
preparationsTheriogenology5919491957200310.1016/S0093-691X(02)01294-312600732
|
|
35.
|
S HyunG LeeD KimProduction of nuclear
transfer-derived piglets using porcine fetal fibroblasts
transfected with the enhanced green fluorescent proteinBiol
Reprod6910601068200310.1095/biolreprod.102.014886
|
|
36.
|
GS LeeHS KimSH HyunProduction of
transgenic cloned piglets from genetically transformed fetal
fibroblasts selected by green fluorescent
proteinTheriogenology63973991200510.1016/j.theriogenology.2004.04.017
|
|
37.
|
YK KangDB KooJS ParkAberrant methylation
of donor genome in cloned bovine embryosNat
Genet28173177200110.1038/8890311381267
|
|
38.
|
PM WarneckeJR MannM FrommerSJ
ClarkBisulfite sequencing in preimplantation embryos: DNA
methylation profile of the upstream region of the mouse imprinted
H19 geneGenomics51182190199810.1006/geno.1998.53719722940
|
|
39.
|
HY JeonSH HyunGS LeeThe analysis of
telomere length and telomerase activity in cloned pigs and cowsMol
Reprod Dev71315320200510.1002/mrd.2027915806556
|
|
40.
|
GP DimriX LeeG BasileA biomarker that
identifies senescent human cells in culture and in aging skin in
vivoProc Natl Acad Sci
USA9293639367199510.1073/pnas.92.20.93637568133
|
|
41.
|
DA LoweH DegensKD ChenSE
AlwayGlyceraldehyde-3-phosphate dehydrogenase varies with age in
glycolytic muscles of ratsJ Gerontol A Biol Sci Med
Sci55B160B164200010.1093/gerona/55.3.B16010795720
|
|
42.
|
TK MoeJ ZiliangA BarathiRW
BeuermanDifferential expression of glyceraldehyde-3-phosphate
dehydrogenase (GAPDH), beta actin and hypoxanthine
phosphoribosyltransferase (HPRT) in postnatal rabbit scleraCurr Eye
Res234450200110.1076/ceyr.23.1.44.5420
|
|
43.
|
W ReikW DeanJ WalterEpigenetic
reprogramming in mammalian
developmentScience29310891093200110.1126/science.106344311498579
|
|
44.
|
GS LeeHS KimSH LeeSuccessful surgical
correction of anal atresia in a transgenic cloned pigletJ Vet
Sci6243245200516131829
|
|
45.
|
SJ UhmHM ChungC KimIn vitro development of
porcine enucleated oocytes reconstructed by the transfer of porcine
fetal fibroblasts and cumulus
cellsTheriogenology54559570200010.1016/S0093-691X(00)00371-X11071130
|
|
46.
|
P ChesnePG AdenotC VigliettaM BaratteL
BoulangerJP RenardCloned rabbits produced by nuclear transfer from
adult somatic cellsNat
Biotechnol20366369200210.1038/nbt0402-36611923842
|
|
47.
|
T ShinD KraemerJ PryorA cat cloned by
nuclear
transplantationNature415859200210.1038/nature72311859353
|
|
48.
|
EH BlackburnTelomere states and cell
fatesNature4085356200010.1038/3504050011081503
|
|
49.
|
RC AllsoppH VaziriC PattersonTelomere
length predicts replicative capacity of human fibroblastsProc Natl
Acad Sci USA891011410118199210.1073/pnas.89.21.101141438199
|
|
50.
|
CB HarleyHuman ageing and telomeresCiba
Found Symp21112914419979524755
|
|
51.
|
JA McKayEA WilliamsJC MathersFolate and
DNA methylation during in utero development and agingBiochem Soc
Trans3210061007200410.1042/BST032100615506948
|
|
52.
|
MR MannYG ChungLD NolenRI VeronaKE
LathamMS BartolomeiDisruption of imprinted gene methylation and
expression in cloned preimplantation stage mouse embryosBiol
Reprod69902914200310.1095/biolreprod.103.01729312748125
|
|
53.
|
L YangP Chavatte-PalmerC KubotaExpression
of imprinted genes is aberrant in deceased newborn cloned calves
and relatively normal in surviving adult clonesMol Reprod
Dev71431438200510.1002/mrd.2031115895469
|
|
54.
|
K InoueT KohdaJ LeeFaithful expression of
imprinted genes in cloned
miceScience295297200210.1126/science.295.5553.29711786635
|
|
55.
|
JM MilnerTE CawstonMatrix
metalloproteinase knockout studies and the potential use of matrix
metalloproteinase inhibitors in the rheumatic diseasesCurr Drug
Targets Inflamm
Allergy4363375200510.2174/156801005402214116101546
|