Introduction
Allergic asthma is conventionally characterized by
an increase in serum immunoglobulin E and the accumulation and
activation of Th2 cells, eosinophils and mast cells. Recently,
T-helper 17 (Th17) cells, a CD4+ helper T cell subset
that produces interleukin-17A (IL-17A) and IL-17F, have been
discovered to play important roles in allergic responses such as
delayed-type hypersensitivity, contact hypersensitivity, and
allergic airway inflammation (1).
IL-17 promotes inflammation by inducing different proinflammatory
cytokines and chemokines, recruiting neutrophils, enhancing
antibody production, and activating T cells. Overexpression of
IL-17A and IL-17F in the lungs leads to increased proinflammatory
cytokine and chemokine expression, causing inflammation associated
with neutrophil infiltration (2–5).
IL-17 mRNA and/or protein was reported to be increased in the
lungs, sputum, bronchoalveolar lavage fluids and sera from
asthmatics, and the levels of IL-17 were correlated with the degree
of severity of airway hypersensitivity (6).
Accumulating evidence suggests that certain
probiotics may contribute to the development of immune function in
the gastrointestinal tract. In particular, there is growing
interest in the therapeutic potential of these organisms in
allergic disorders. Clinical trials have indicated that oral
administration of certain organisms can modulate immune responses
in the airway (7–9). Live Lactobacillus reuteri
significantly attenuated the development of experimental allergic
asthma in one study (9). The
development of mucosal and systemic tolerance seems to rely on
immunosuppressive mechanisms orchestrated by regulatory T-cell
classes that attenuate both Th1 and Th2 responses (10). However, the involvement of Th17
cells and anti-allergic action of certain bacteria remains
obscure.
To seek evidence for an association between the
suppression of allergic airway responses induced by certain
microorganisms and the presence of Th17 cells, we investigated the
expression of Th17 cells in a murine allergic asthma model. A
product of lysozyme and heat-treated Enterococcus faecalis
FK-23, LFK was orally administered to asthmatic mice with a view to
decreasing airway inflammation. We had previously shown that LFK
inhibited active cutaneous anaphylaxis and allergen-induced
peritoneal accumulation of eosinophils in murine models (11,12). The potential role of Th17 cells in
LFK-mediated attenuation of allergic airway inflammation is
described in the present study.
Materials and methods
Preparation of LFK
LFK was prepared as previously described (11,12). In brief, Enterococcus
faecalis FK-23 was cultured for 24 h at 37°C, in a broth medium
containing 2% glucose, 2% yeast extract and 4%
KH2PO4. The cells were harvested by
centrifugation, washed three times with distilled water and then
treated with lysozyme (1 mg/ml) at 37°C for 2 h, heated to 105°C
for 10 min, then lyophilized.
Other compounds
Methacholine chloride and ISOGEN were purchased from
Wako Pure Chemical Industries (Osaka, Japan). Ovalbumin (OVA),
Percoll, phorbol myristate acetate (PMA), ionmycin and DNase I were
obtained from Sigma Chemical Co. (St. Louis, MO, USA). Collagenase
D and dispase were from Roche Diagnostics GmbH (Mannheim,
Germany).
Animal
Male BALB/c mice (20–25 g) (Japan SLC, Hamamatsu,
Japan) were used. They were housed at a constant temperature of
22±2°C with a humidity of 55±10% on an automatically controlled
12:12 h light-dark cycle, and given food and water ad
libitum. Animal care and research protocols were in accordance
with the principles and guidelines adopted by the Animal Care
Committee of Ehime University and approved by the University
Committee for Animal Research.
Administration of LFK and
sensitization
Mice were given LFK (60 mg) in 0.5 ml saline every
day for 28 days. Mice given only saline were used as controls
(Fig. 1). Mice were sensitized by
subcutaneous injection of 20 μg OVA adsorbed to 500
μg aluminum hydroxide suspension in saline on Day 14. A
booster injection of OVA was given on the Day 21. Sensitized mice
were exposed to 1% OVA for 30 min by inhalation on Days 25, 26 and
27, followed by inhalation of 5% OVA on Day 28. Mice challenged
with saline were used as controls (Fig. 1).
Measurement of airway
hypersensitivity
Twenty-four hours after the final OVA challenge,
bronchial reactivity to aerosolized methacholine was measured using
a whole-body plethysmographic chamber (Buxco Electronics, Sharon,
CT, USA) to determine changes in enhanced respiratory pause (PenH).
After a 10-min stabilization period, increasing concentrations of
methacholine (2–32 mg/ml) or saline were aerosolized for 1 min
each, and mean PenH values were obtained over 5-min periods. PenH
(= pause × PEP/PIP) describes airway resistance, using the peak
expiratory pressure (PEP) and the peak inspiratory pressure (PIP).
The pause was defined as (Tc-Tr)/Tc, where Tc is the time of
expiration and Tr is the relaxation time. The Tr was the time of
pressure decay to 30% of the total expiratory pressure signal.
Constriction of the airway is shown as the increased PenH
value.
Leukocyte accumulation in bronchoalveolar
fluid (BALF)
Twenty-four hours after challenge with OVA or
saline, BALF was collected via tracheal cannulation using 250
μl phosphate-buffered saline solution (PBS, pH 7.20).
Collections were repeated 3 times on each mouse. Routinely, >90%
of the lavage fluid was recovered from the lungs. Cells were
removed from BALF by centrifugation at 200 × g for 15 min and BALF
cell pellets were pooled and resuspended in PBS for total and
differential cell counts.
The morphology of BALF cell preparations was
analyzed using light microscopy. Air-dried preparations were fixed
and stained with hematoxylin and eosin (H&E) to determine the
total cell number. The other preparations were stained with Alcian
blue and nuclear fast to identify mast cells. Results are expressed
as the number of cells per lung.
Histological analysis
Lung sections were stained with H&E after
fixation with 4% paraformaldehyde in 0.1 M phosphate buffer (pH
7.20) for 12 h at 4°C. All sample slides were examined using a
Nikon C1 microscope (Nikon, Japan) and compared at the same
magnification.
Analysis of Th17 cells in lungs, spleens
and lamina propria of the intestine
Twenty-four hours after the final OVA challenge,
tissues were removed and cells were isolated. Splenocytes were
isolated from spleens using a standard dissection technique and
resuspended in RPMI-1640 for staining (13). For isolation of lung cells,
briefly, lungs were cut into small pieces and digested in 10 ml of
medium containing 300 U/ml collagenase D and 1.5 mg/ml DNase I at
37°C for 1 h. The digested lung tissue was filtered through a 70
μm cell strainer, after red blood cells had been lysed
(14). For isolation of
intestinal lamina propria lymphocytes, the intestines were opened
longitudinally and shaken in Hanks’ balanced salt solution
containing 5 mM EDTANa2 for 20 min at 37°C. Small pieces
of intestine were incubated with RPMI-1640 containing 4% fetal
bovine serum, 1 mg/ml collagenase D, 1 mg/ml dispase and 40
μg/ml DNase I for 1 h at 37°C in a shaking water bath. Then
the digest was resuspended in 40% Percoll and overlayed on 80%
Percoll. After centrifugation, the lamina propria lymphocytes from
the interface of the Percoll gradients were collected immediately
for intracellular cytokine staining (15). Dispersed cell suspensions from
lungs, spleens and lamina propria were resuspended at
1×106 cells/ml and incubated with 50 ng/ml PMA, 500
ng/ml ionomycin in RPMI-1640 at 37°C for 6 h. After extracellular
staining with PE anti-mouse CD4, the cells were permeabilized with
Fix/Perm solution for 20 min at 4°C. They were then stained with
Alexa Fluor 488 anti-mouse IL-17A. Data were acquired on a BD
FACSAria (Becton-Dickinson, Oakville, ON, Canada) and analyzed
using FlowJo (Tree Star Inc., Ashland, OR, USA)
Quantitative real-time PCR
Total-RNA was extracted with ISOGEN from lungs,
spleens and intestines of mice 24 h after the final challenge. The
expression of TGF-β, IL-6 and β-actin mRNA was assessed according
to the manufacturer’s instruction using the One Step SYBR
PrimeScript Plus RT-PCR kit (Takara Bio, Inc., Shiga, Japan). The
fluorescence emission of the probe was monitored and analyzed using
an Applied Biosystems 7500 Fast Real-Time PCR system (Applied
Biosystems). Specific oligonucleotide primers were designed
according to published sequences as shown in Table I. The expression levels of IL-6
and TGF-β were corrected by reference to β-actin, and the relative
amount of each mRNA in each sample was calculated by the
comparative ΔCt method.
 | Table ISequences of PCR primers. |
Table I
Sequences of PCR primers.
| Target | Forward sequence
(5′→3′) | Reverse sequence
(5′→3′) |
|---|
| TGF-β |
GACTCTCCACCTGCAAGACCA |
GGGACTGGCCGAGCCTTAGTT |
| IL-6 |
TTCCATCCAGTTGCCTTCTTG |
TTGGGAGTGGTATCCTCTGTGA |
| β-actin |
AGAGGGAAATCGTGCGTGAC |
CAATAGTGATGACCTGGCCGT |
Statistical analyses
All experiments were designed as completely
randomized multifactorials with 3–15 mice/group. Results are
expressed as the mean ± SEM. The significance of the differences
among all groups were analyzed using repeated measures analysis of
variances (ANOVA) followed by the Scheffe’s F-test. Significant
differences between treatments were assessed using Student’s
t-test. P-values <0.05 were taken to indicate significant
differences.
Results
Suppression of allergic airway
inflammation by oral administration of LFK
We determined the effect of LFK on airway
hyperresponsiveness, one of the most important characteristics of
the allergic airway response. Suppression of airway
hyperresponsiveness, reduced accumulation of leukocytes in BALF and
less lung inflammation were seen following oral administration of
LFK. Thus, the final OVA challenge to OVA-sensitized mice resulted
in an enhanced response to methacholine as determined by an
enhanced pause index, but this was significantly attenuated by oral
administration of LFK for 28 days (Fig. 2A). Administration of LFK alone did
not have any effects on airway resistance in non-sensitized
mice.
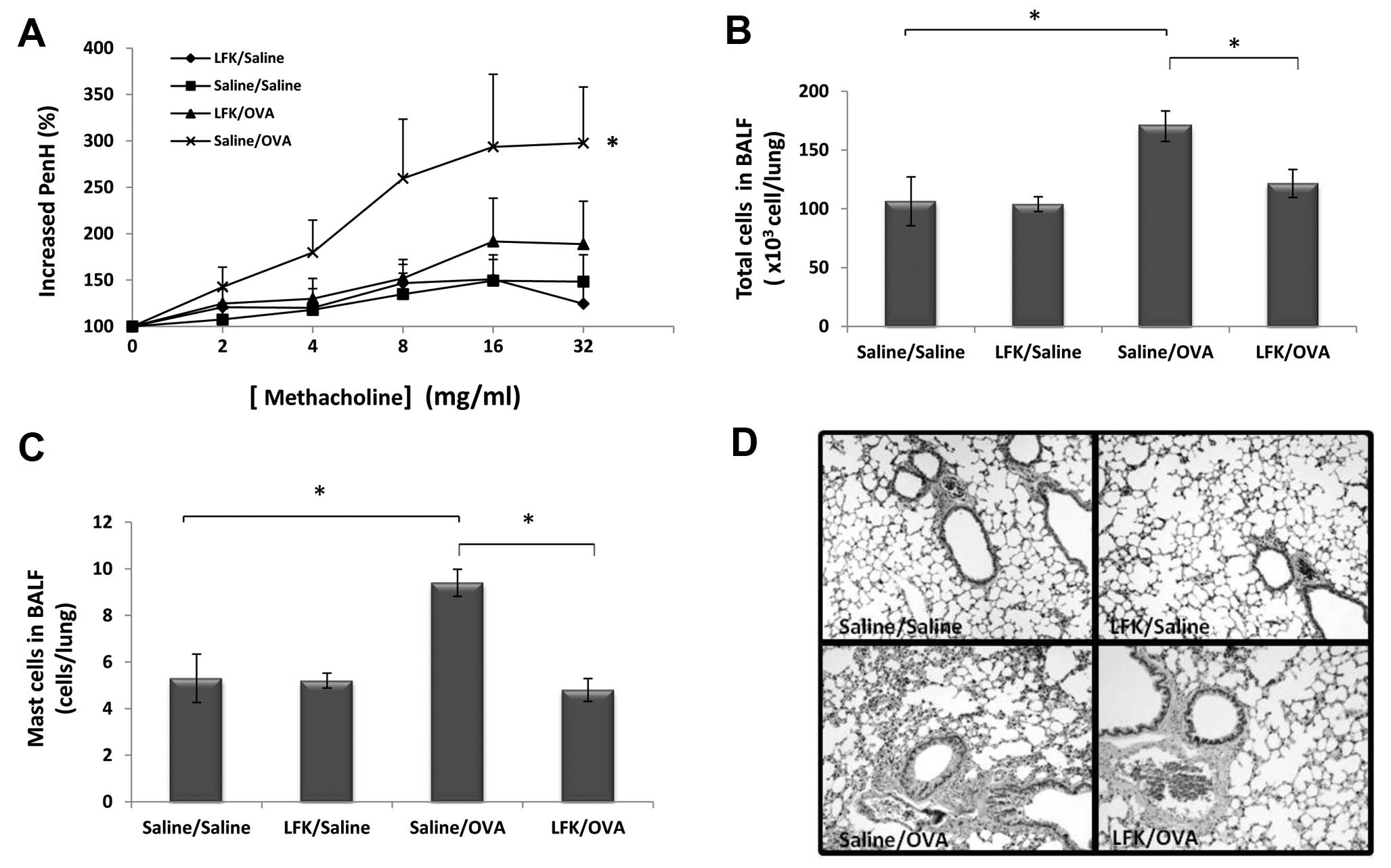 | Figure 2Suppression of allergic airway
inflammation by administration of LFK. (A) Changes in airway
hypersensitivity were quantified using the enhanced pause index,
which was measured using a whole-body plethysmograph. For each
experiment, increasing concentrations of methacholine (2–32 mg/ml)
or saline were aerosolized for 1 min each, and mean PenH values
were obtained over 5-min periods. The results are expressed as the
mean ± SEM. *P<0.05, ANOVA followed by Scheffe’s test
for differences in saline-treated/OVA-challenged mice (x-marks),
LFK-treated/OVA-challenged mice (triangles),
LFK-treated/saline-challenged mice (diamonds) and
saline-treated/saline-challenged mice (squares). (B) Total number
of cells in bronchoalveolar lavage fluid (BALF) taken from mice 24
h after challenge by inhalation of 5% OVA. The BALF cells were
analyzed by H&E staining. Mice challenged with saline were used
as controls. *P<0.05, Student’s t-test, compared with
each group. (C) The number of mast cells in BALF. Mast cells were
detected by Alcian blue/nuclear fast staining.
*P<0.05, Student’s t-test, compared with each group.
(D) Histological analysis of the lung. The lung sections were
stained by H&E. All sample slides were examined using a Nikon
C1 microscope (Nikon, Japan) and compared at the same magnification
(n=5–15 mice/group). |
Oral treatment with LFK also attenuated inflammatory
cell influx into the airway. The total number of cells in BALF
increased by 1.74-fold (P<0.05) 24 h after the final antigen
challenge in OVA-sensitized (saline-treated/OVA-challenged) mice
compared with the saline-treated/saline-challenged controls
(Fig. 2B).
LFK-treated/OVA-challenged mice showed a significant reduction in
total cells recovered in BALF to 71.8% (P<0.05) compared to
saline-treated/OVA-challenged mice.
The OVA challenge also caused a marked increase in
the proportion of mast cells present (Fig. 2C), one of the most important
functional cell populations in the allergic airway response. In the
LFK-treated/OVA-challenged group, the number of mast cells was
significantly decreased to 50.9% of the
saline-treated/OVA-challenged mice. This result is in line with the
marked decrease in total numbers following administration of
LFK.
The anti-allergic effect of LFK on airway
inflammation was also observed in histological analysis. The
administration of LFK to OVA-challenged mice resulted in decreased
perivascular and peribronchiolar infiltration by inflammatory cells
(Fig. 2D). Thus, suppression of
airway inflammation as a result of oral administration of LFK was
clearly demonstrated in this model.
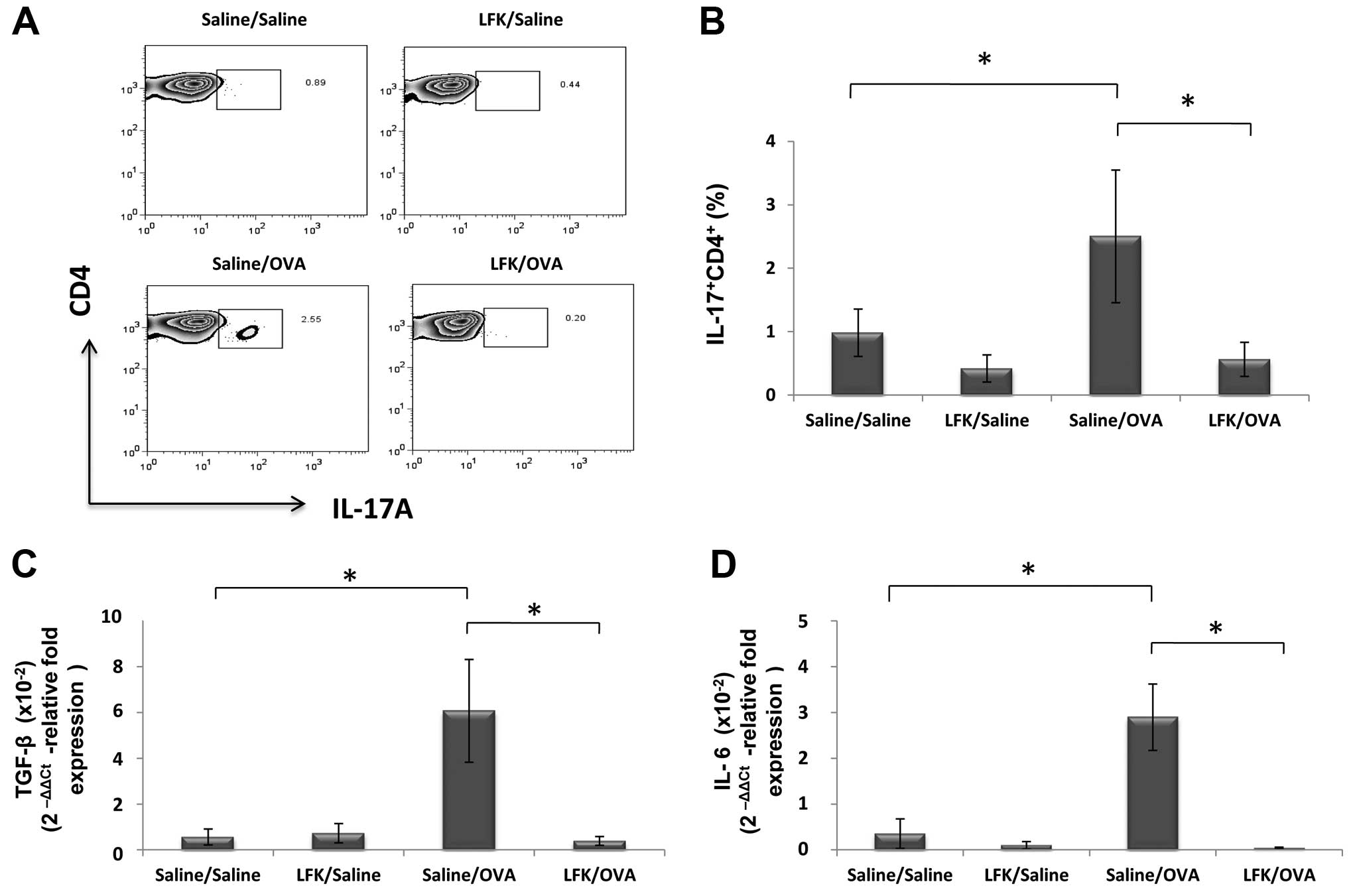
Effect of LFK on the presence of Th17
cells in lung
To investigate the effect of oral administration of
LFK on the development of Th17 cells, we assessed IL-17 expression
in isolated lymphocytes from the lung after oral treatment with LFK
for 28 days. In the LFK-treated/saline-challenged mice, the
percentage of IL-17-expressing CD4+ cells in the lung
was reduced to 43.5% (P<0.05) compared to
saline-treated/saline-challenged mice. OVA sensitization and
challenge dramatically increased IL-17-expressing CD4+
cells in the lung, showing a 2.4-fold increase compared with the
saline-sensitized and challenged mice. However, this robust
development of Th17 cells was attenuated by LFK administration,
which resulted in a significant decrease of the percentage of
IL-17-expressing CD4+ cells in
LFK-treated/OVA-challenged mice, to 23.8% (P<0.05) compared to
saline-treated/OVA-challenged mice (Fig. 3A and B).
Oral treatment with LFK also markedly reduced TGF-β
expression to 8.7% (P<0.05) and IL-6 to 1.3% (P<0.05) of that
seen in saline-treated/OVA-challenged mice (Fig. 3C and D). Thus, LFK had marked
effects on several related cytokines and reduced the development of
Th17 cells in allergic airway inflammation.
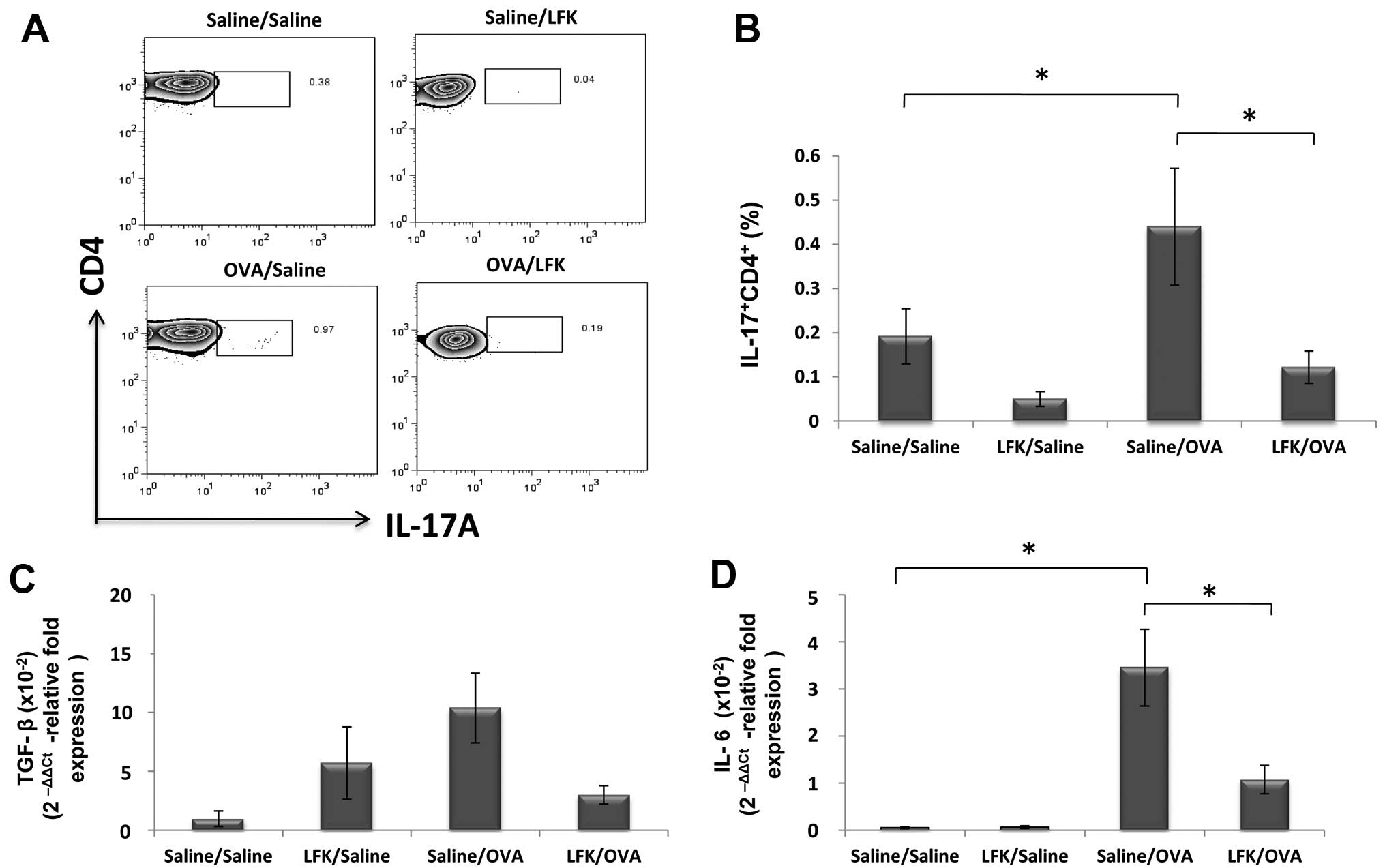
Effect of LFK on the presence of Th17
cells in splenocytes
The changes in the percentage of IL-17-expressing
CD4+ cells in splenocytes showed a similar tendency as
in isolated lung cells. Oral administration of LFK reduced the
percentage of IL-17-expressing CD4+ cells to 26.2%
(P<0.05) compared with saline-treated/OVA-challenged mice
(Fig. 4A and B). Again, the
expression of IL-6 was also decreased to 33.3% (P<0.05), and the
reduction of TGF-β did not achieve significance.
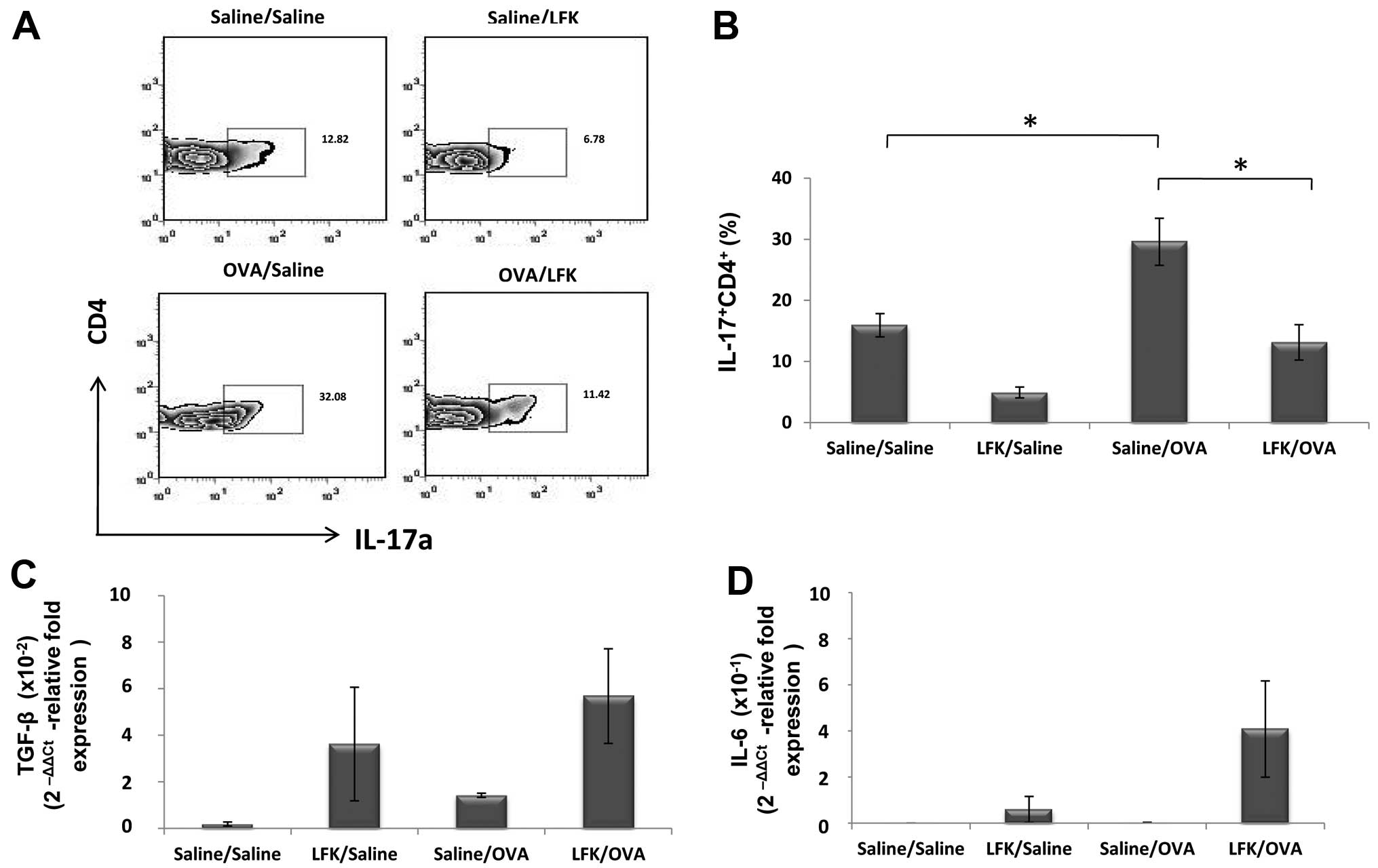
Effect of LFK on the presence of Th17
cells in the intestine
To test whether LFK suppresses Th17 cells in a
crucial location, IL-17-expression of CD4+ T cells in
the intestinal lamina propria was investigated. IL-17-expressing
CD4+ cells in the intestine was increased 1.97-fold
(P<0.05) in saline-treated/OVA-sensitized mice compared with the
saline-treated/saline-challenged control. LFK reduced the
percentage of IL-17-expressing CD4+ cells to 44.8%
(P<0.05) compared with saline-treated/OVA-challenged mice
(Fig. 5A and B). No significant
change was detected in IL-6 expression in these mice. The
expression of TGF-β showed a tendency to increase in LFK-treated
mice, but this was not significant.
Discussion
In the present study, we clearly showed that oral
administration of LFK attenuated allergic airway inflammation along
with significantly suppressing Th17 cell development in mice. That
may help us to understand the anti-inflammatory effects of
probiotics in the prevention of allergic airway inflammation.
Studies using other specific strains of probiotic organisms such as
Lactobacillus reuteri reported the importance of live but
not heat-killed bacteria in maintaining Th2 functional tolerance
(9). The reasons for choosing the
lysed heat-killed probiotics for this study were the following:
first, in the form of lysed heat-killed Enterococcus
faecalis FK-23, the effects of LFK had been analyzed in several
immunological disorders, such as active cutaneous anaphylaxis and
allergen-induced peritoneal eosinophil accumulation. It is
suggested that heat-killed probiotics also have potential
therapeutic effects as live bacterias. Second, the specific strains
of probiotics have anti-inflammatory activities but the
relationships between the probiotics and IL-17 suppression have not
been discussed. Third, use of lysed heat-killed probiotics rather
than live bacteria assisted in maintaining consistency between
experiments in mice. Here we show unequivocally that oral
administration of LFK to mice attenuated lung inflammation and
airway hyperresponsiveness. These findings are in line with the
study of Hunt et al (16),
who demonstrated that the intragastric administration of
heat-killed Mycobacterium vaccae significantly reduced
pulmonary inflammation after antigen challenge in OVA-sensitized
mice.
We next asked why the pathological features of
airway inflammation are inhibited by LFK. Despite clear evidence
for strong immunomodulatory properties of LFK, the mechanisms
underlying these effects are poorly understood. Here we
demonstrated one of the most important effects of probiotics on
allergic airway inflammation in mice. That is that oral
administration of LFK could suppress Th17 cell development in the
intestinal lamina propria, spleen and lung of the asthmatic mice.
The importance of IL-17-producing T (Th17) cells rather than Th1
cells or Th2 cells for the development of allergic diseases has
been demonstrated in IL-17-deficent mice (17). The clinical observations also
showed that the concentration of IL-17 was significantly increased
in BALF, sputum and blood from patients with asthma (18,19). Exploration of the mechanisms by
which Th17 cells differentiate from naïve T cells have shown that
IL-6, in synergy with TGF-β have a critical role in inducing the
differentiation of Th17 cells.
Our data have demonstrated that oral administration
of LFK significantly decreased mRNA expression of TGF-β as well as
IL-6 in lungs (Fig. 3C and D),
which may explain the inhibitory effects of oral LFK treatment on
Th17 cell development in the local site. In spleens, LFK treatment
decreased mRNA expression of IL-6 (Fig. 4D) while suppressing Th17 cell
development (Fig. 4A and B).
Indeed, IL-6 has key functions in programming naïve CD4+
T cells to become Th17 cells. IL-6 induced expression of IL-21 that
amplified an autocrine loop to induce more IL-21 and IL-23 receptor
in naïve and CD4+ cells. Furthermore, IL-6 through the
STAT3-RORγt pathway in concert with TGF-β are involved in
differentiation of Th17 cells (20). Ghoreschi et al (21) showed that Th17 differentiation can
occur in the absence of TGF-β signaling in mice. Neither IL-6 nor
IL-23 alone efficiently generated pathogenic Th17 cells. On the
other hand, IL-6-deficient mice have been shown to have a deficit
in Th17 development (22).
In contrast, although the TGF-β and IL-6 mRNA
expression levels in intestinal lamina propria were not
significantly different between the asthmatic mice and LFK-treated
asthmatic mice, both TGF-β and IL-6 showed a tendency to increase
in LFK-treated models of asthma. IL-6 is one of the main cytokines
produced by intestinal lamina propria cells after infection. For
example, overproduction of IL-6 has been found in many types of
colitis. Further analyses are needed to clarify whether the
expression of IL-6 in intestinal lamina propria cells could be
suppressed by LFK. TGF-β, has a dual function in directing the
differentiation of Treg cells and proinflammatory Th17 cells.
Whether increased production of TGF-β by intestinal lamina propria
cells contributed to a shift in the Treg/Th17 balance toward Treg
resulting in suppression of Th17 cell development (Fig. 5A and B) needs to be further
clarified. It has been reported that Foxp3 can directly bind to
RORγt to then inhibit RORγt-dependent transcriptional activation of
its key target genes such as IL-17 (23).
Examination of cytokines in the present study was
performed in vivo. To mimic the in vivo interaction
between probiotics and target cells using an in vitro
experiment would be beneficial to investigate the precise
mechanism. In fact, in vitro experiments have been
extensively performed. Tanabe et al (24) and Jan et al (25) analyzed cytokine production after
splenocytes were cultured with Bifidobacterium infantis and
Lactobacillus gasseri and demonstrated that these selected
probiotics are able to suppress IL-17 production (24,25). Foligne and colleagues demonstrated
that mouse bone marrow-derived dendritic cells (DCs) stimulated
in vitro with selected Lactobacillus strains, could
attenuate colitis via a Tregs-dependent mechanism (26). In vivo studies using DCs
have shown that the functional changes in DCs following interaction
with probiotics are critical for immune regulation. These selected
probiotics can stimulate DC regulatory functions by targeting a
specific pattern-recognition receptor (PRR) family, such as
Toll-like receptors (TLRs), through DCs expression of various
cytokines, such as IL-10, TGF-β and indoleamine 2,3-dioxygease
(IDO), which drive the generation of
CD4+25+Foxp3+ Tregs (27–29). As described above, Foxp3 can
inhibit Th17 cells differentiation by antagonizing RORγt, while,
these probiotic-induced Tregs can also inhibit the differentiation
of Th1 and Th2 cells (30), which
may explain how some probiotic strains could suppress the Th2
response in asthma model (31).
Although we showed evidence for the attenuation of
Th17 cells and related cytokines produced by lung cells and
splenocytes following oral administration of LFK, this does not
prove the precise mechanism and the LFK component(s) which
responded with target cells. Further studies are required to
elucidate the nature of the probiotic-host cell interactions, how
these interactions induce immunomodulation and to identify the
specific component(s) of probiotic and host responsible for these
interactions. In addition, the mechanism by which the
immunomodulatory effects of oral probiotics treatment influence
sites distant to the intestine, such as respiratory tract need to
be examined. To address this issue, a gut-lung axis of probiotic
action was proposed by Forsythe (32). A literature review shows that the
anti-allergic effects of probiotics are strain-dependent and
mediated by different effects (33). For example, in contrast to the
suppression of IL-17/Th17 by selected probiotics it has been
reported that a specific component of the macrobiota, Candidatus
arthromitus, a subset of the Firmicute-Clostridiae group
commonly termed segmented filamentous bacteria (SFB), promotes Th17
cell induction in the intestine (15).
In conclusion, our results have shown that oral
administration of LFK resulted in significant attention of allergic
airway response in OVA-sensitized mice. Our data also indicated
that oral administration of LFK could suppress the in vivo
development of Th17, which may be caused by LFK-mediated
restoration of the Treg/Th17 balance though the TGF-β and IL-6
cytokine-dependent signaling pathways. Increased knowledge of the
regulatory activities of probiotics will stimulate the development
of alternative approaches for the treatment of immune disorders via
the modulation of T cell-balance.
References
|
1.
|
Y IwakuraS NakaeS SaijoH IshigameThe roles
of IL-17A in inflammatory immune responses and host defense against
pathogensImmunol
Rev2265779200810.1111/j.1600-065X.2008.00699.x19161416
|
|
2.
|
SD HurstT MuchamuelDM GormanNew IL-17
family members promote Th1 or Th2 responses in the lung: in vivo
function of the novel cytokine IL-25J
Immunol169443453200210.4049/jimmunol.169.1.44312077275
|
|
3.
|
N OdaPB CanelosDM EssayanBA PlunkettAC
MyersSK HuangInterleukin-17F induces pulmonary neutrophilia and
amplifies antigen-induced allergic responseAm J Respir Crit Care
Med1711218200510.1164/rccm.200406-778OC15477493
|
|
4.
|
H ParkZ LiXO YangA distinct lineage of CD4
T cells regulates tissue inflammation by producing interleukin
17Nat Immunol611331141200510.1038/ni126116200068
|
|
5.
|
XO YangSH ChangH ParkRegulation of
inflammatory responses by IL-17FJ Exp
Med20510631075200810.1084/jem.2007197818411338
|
|
6.
|
K ObokiT OhnoH SaitoS NakaeTh17 and
allergyAllergol
Int57121134200810.2332/allergolint.R-07-16018427165
|
|
7.
|
N BlumerS SelS VirnaPerinatal maternal
application of Lactobacillus rhamnosus GG suppresses
allergic airway inflammation in mouse offspringClin Exp
Allergy37348357200717359385
|
|
8.
|
W FeleszkoJ JaworskaRD
RhaProbiotic-induced suppression of allergic sensitization and
airway inflammation is associated with an increase of T
regulatory-dependent mechanisms in a murine model of asthmaClin Exp
Allergy37498505200710.1111/j.1365-2222.2006.02629.x
|
|
9.
|
P ForsytheMD InmanJ BienenstockOral
treatment with live Lactobacillus reuteri inhibits the
allergic airway response in miceAm J Respir Crit Care
Med1755615692007
|
|
10.
|
K KarimiMD InmanJ BienenstockP
ForsytheLactobacillus reuteri-induced regulatory T cells
protect against an allergic airway response in miceAm J Respir Crit
Care Med179186193200910.1164/rccm.200806-951OC
|
|
11.
|
T ShimadaL ChengM IdeS FukudaT EnomotoT
ShirakawaEffect of lysed Enterococcus faecalis FK-23 (LFK)
on allergen-induced peritoneal accumulation of eosinophils in
miceClin Exp Allergy336846872003
|
|
12.
|
T ShimadaL ChengA YamasakiEffects of lysed
Enterococcus faecalis FK-23 on allergen-induced serum
antibody responses and active cutaneous anaphylaxis in miceClin Exp
Allergy34178417882004
|
|
13.
|
MH CatoIW YauRC RickertMagnetic-based
purification of untouched mouse germinal center B cells for ex vivo
manipulation and biochemical analysisNat
Protoc6953960201110.1038/nprot.2011.34421720310
|
|
14.
|
KA SauerP ScholtesR KarwotS
FinottoIsolation of CD4+ cells from murine lungs: a
method to analyze ongoing immune responses in the lungNat
Protoc128702875200617406546
|
|
15.
|
K AtarashiT TanoueK HondaInduction of
lamina propria Th17 cells by intestinal commensal
bacteriaVaccine2880368038201010.1016/j.vaccine.2010.09.02620849872
|
|
16.
|
JR HuntR MartinelliVC AdamsGA RookLR
BrunetIntragastric administration of Mycobacterium vaccae
inhibits severe pulmonary allergic inflammation in a mouse
modelClin Exp Allergy35685690200515898994
|
|
17.
|
S NakaeY KomiyamaA NambuAntigen-specific T
cell sensitization is impaired in IL-17-deficient mice, causing
suppression of allergic cellular and humoral
responsesImmunity17375387200210.1016/S1074-7613(02)00391-612354389
|
|
18.
|
S MoletQ HamidF DavoineIL-17 is increased
in asthmatic airways and induces human bronchial fibroblasts to
produce cytokinesJ Allergy Clin
Immunol108430438200110.1067/mai.2001.11792911544464
|
|
19.
|
A BarczykW PierzchalaE
SozanskaInterleukin-17 in sputum correlates with airway
hyperresponsiveness to methacholineRespir
Med97726733200310.1053/rmed.2003.150712814161
|
|
20.
|
L ZhouIvanov IIR SpolskiIL-6 programs
T(H)-17 cell differentiation by promoting sequential engagement of
the IL-21 and IL-23 pathwaysNat
Immunol8967974200710.1038/ni148817581537
|
|
21.
|
K GhoreschiA LaurenceXP YangGeneration of
pathogenic T(H)17 cells in the absence of TGF-beta
signallingNature467967971201010.1038/nature0944720962846
|
|
22.
|
E BettelliY CarrierW GaoReciprocal
developmental pathways for the generation of pathogenic effector
TH17 and regulatory T
cellsNature441235238200610.1038/nature04753
|
|
23.
|
L ZhouJE LopesMM ChongTGF-beta-induced
Foxp3 inhibits T(H)17 cell differentiation by antagonizing
RORgammat
functionNature453236240200810.1038/nature0687818368049
|
|
24.
|
S TanabeY KinutaY SaitoBifidobacterium
infantis suppresses proinflammatory interleukin-17 production
in murine splenocytes and dextran sodium sulfate-induced intestinal
inflammationInt J Mol Med221811852008
|
|
25.
|
RL JanKC YehMH HsiehLactobacillus
gasseri suppresses Th17 pro-inflammatory response and
attenuates allergen-induced airway inflammation in a mouse model of
allergic asthmaBr J NutrOct142011(Epub ahead of print).
|
|
26.
|
B FoligneG ZoumpopoulouJ DewulfA key role
of dendritic cells in probiotic functionalityPLoS
One2e313200710.1371/journal.pone.000031317375199
|
|
27.
|
HK KwonCG LeeJS SoGeneration of regulatory
dendritic cells and CD4+Foxp3+ T cells by
probiotics administration suppresses immune disordersProc Natl Acad
Sci USA10721592164201020080669
|
|
28.
|
P PuccettiU GrohmannIDO and regulatory T
cells: a role for reverse signalling and non-canonical NF-kappaB
activationNat Rev Immunol7817823200710.1038/nri216317767193
|
|
29.
|
AL HartK LammersP BrigidiModulation of
human dendritic cell phenotype and function by probiotic
bacteriaGut5316021609200410.1136/gut.2003.03732515479680
|
|
30.
|
Y BelkaidThe role of CD4(+)CD25(+)
regulatory T cells in Leishmania infectionExpert Opin Biol
Ther38758852003
|
|
31.
|
E SchiaviB BarlettaC ButteroniS CorintiM
BoirivantG Di FeliceOral therapeutic administration of a probiotic
mixture suppresses established Th2 responses and systemic
anaphylaxis in a murine model of food
allergyAllergy66499508201110.1111/j.1398-9995.2010.02501.x21058959
|
|
32.
|
P ForsytheProbiotics and lung
diseasesChest139901908201110.1378/chest.10-186121467057
|
|
33.
|
S MichailThe role of probiotics in
allergic diseasesAllergy Asthma Clin
Immunol55200910.1186/1710-1492-5-5
|