Introduction
The major pathological changes in osteoarthritis
(OA) include synovial proliferation and cartilage degeneration that
involve several inflammatory factors. Cartilage damage is one of
the primary pathological characteristics of OA. Cartilage fragments
enter the synovial fluid, irritating the synovial membrane and
leading to synovitis. The inflamed synovial membrane subsequently
releases inflammatory mediators, which further degenerate
cartilage, thereby forming a vicious circle. Thus, synovitis is the
main cause for knee cartilage damage and chronic degeneration in OA
(1). Elevated expression of
interleukin-1 (IL-1), a crucial pro-inflammatory cytokine, is often
observed in the synovial membrane in patients with OA. The
imbalance between the IL-1 receptor (IL-1R) and the IL-1 receptor
antagonist (IL-1Ra) is correlated with the severity of OA (2). IL-1β has the potential to stimulate
chondrocytes to produce several inflammatory factors, including
nitric oxide (NO), inducible nitric oxide synthase (iNOS),
prostaglandin-E2 (PGE2) or matrix
metalloproteases (MMPs) (3),
leading to the inflammatory response of chondrocytes as well as the
degradation of the cartilage matrix (4–6).
IL-18 has a structure similar to that of IL-1 and is
therefore acknowledged as a member of the IL-1 super family
(7). Initially, T-cell-produced
IL-18 was found to be able to induce interferon-γ (IFN-γ)
production, and was subsequently named IFN-γ-inducing factor. IL-18
is derived from an inactive precursor. The mature and functional 18
kDa IL-18 protein molecule is generated through enzymatic cleavage
(7,8). IL-18 has multiple biological
activities, while being crucial for the pathogenesis of rheumatoid
arthritis, especially synovitis. IL-18 can be detected in the
synovial fluid in the knees in patients with rheumatic arthritis
(RA). IL-18 receptor (IL-18R) is also expressed in the synovial
membrane in RA-affected joints (9–11).
A continuous low-level IL-18 expression, tumor necrosis factor-α
(TNF-α), granulocyte macrophage-colony stimulating factor (GM-CSF)
and IL-1β were reported in fibroblast-like synoviocytes (FLS) in
the synovial membrane of RA and OA patients as well as in the
synovial fluid of spondyloarthropathy (SpA) patients (12). A study analyzing the impact of
high-level IL-18 on synoviocytes and peripheral blood monocytes in
RA patients demonstrated that IL-18 had a more prominent
pro-inflammatory effect on peripheral blood monocytes (12).
The response of synoviocytes in OA patients to IL-18
remains unclear. In OA, destruction of articular cartilage mainly
results from the imbalance between the synthesis and the
degradation of cartilage. IL-1 and TNF-α are essential in
stimulating chondrocyte catabolism (13). IL-1 triggers PGE2
production as well as the expression of COX-2 and iNOS (14). However, IL-18 also induces the
expression of genes, such as iNOS, COX-2 and IL-6, leading to the
degeneration and inflammation of chondrocytes (14). IL-18 inhibited the in vitro
proteoglycan production in the patella of young rats in the
presence of IL-1 (15).
Nevertheless, the effect of IL-18 on OA chondrocytes and the role
of IL-1β and IL-18 in these processes remains unclear.
In this study, we investigated the effect of IL-18
on the synoviocytes and chondrocytes of patients with OA. We also
detected the expression of IL-18 receptors in the two types of
cells, and examined the relationship between IL-18 and IL-1 in the
pro-inflammatory process.
Materials and methods
Cell culture
Synoviocytes and chondrocytes were collected from 8
patients with OA (5 females and 3 males; average age, 62.3 years).
The patients provided informed consent prior to sample collection.
This study was monitored and approved by the Medical Ethics
Committee of the Nanfang Hospital.
Synoviocytes were cultured in vitro in
accordance with the methods described by Zimmermann et al
(16). The synovial tissue was
washed twice using sterile phosphate-buffered saline (PBS). The
sample was cut into sections in PBS and digested with 0.1% trypsin
at 37°C for 30 min. The tissue was then placed in Dulbecco’s
modified Eagle’s medium (DMEM) containing 0.1% type I collagenase
(Sigma, St. Louis, MO, USA) and 10% fetal bovine serum (FBS) for 2
h. After filtration and centrifugation, the cells were collected
and cultured in medium with penicillin and streptomycin, for 7 days
at 37°C in a 5% carbon dioxide environment and transferred to a
6-well plate. Synoviocytes were classified as described in the
study by Franke et al (17). Identical numbers of primary,
secondary and tertiary cells were starved in DMEM for 24 h. The
cells were then transferred and cultured in fresh medium containing
IL-18 (10 mM/ml) for an additional 24 h. The cultured cells and
supernatants were harvested for additional analysis.
Articular cartilage collected from the femur condyle
and tibial plateau in the surgeries of total knee joint
arthroplasty was cut into 2–3 mm3-sized pieces and then
washed with DMEM three times. The sample was digested in 10%
trypsin at 37°C for 15 min. The digested tissue was then collected
and mixed with 0.2% type II collagenase (Gibco, Carlsbad, CA, USA)
and 0.1% hyaluronidase (Sigma) at 37°C in a shaker for 6 h. Cells
were washed with DMEM to remove proteases and filtered through a
sterile cell strainer. Primary cells were suspended and cultured in
DMEM supplemented with 10% FBS. The expression of type II collagen
was then analyzed. Selected primary cells were seeded onto 6-well
plates and cultured in medium with different concentrations of
IL-18 (0, 50, 100 and 200 ng/ml) for 48 h.
Immunohistochemistry
Sterilized coverslips (8x8 mm) were placed onto
6-well plates. Cells were seeded and cultured for 24 h, allowing
cells to attach to coverslips. After incubation, coverslips were
washed three times with PBS, for 3 min/wash, and treated with 3%
hydrogen peroxide and normal serum to block intrinsic peroxidases
and proteins possibly generating non-specific cross reactions.
After the removal of residual serum, coverslips were incubated with
rabbit polyclonal antibodies against type I and type II collagen
(Boster, China) in a wet box at 4°C overnight. The following day,
polyclonal goat anti-rabbit secondary antibodies (Boster) were
applied to the coverslips at 37°C for 30 min. After washing, the
coverslips were incubated with horseradish peroxidase substrates
and diaminobenzidine (DAB) and then examined under an inverted
microscope.
Cytokine detection
Supernatants from cultured cells were collected and
centrifuged in 1.5 ml microcentrifuge tubes at 3,000 x g for 10 min
and then stored in a −80°C freezer. The concentration of cytokines
in the samples was assayed simultaneously. Standard curves were
plotted following the manufacturer’s instructions (PGE2;
Uscn Life Science, Inc., Wuhan, China) (TNF-α; Cusabio Biotech Co.,
Ltd., Wuhan, China) and the PGE2 and TNF-α
concentrations in the samples were then determined using
enzyme-linked immunosorbent assay (ELISA).
RT-PCR analysis
Total-mRNA was extracted from synoviocytes and
chondrocytes using TRIzol (Invitrogen, Carlsbad, CA, USA). The
concentration mRNA concentration was determined using a NanoDrop
Spectrophotometer (Wilmington, NC, USA). Reverse transcription was
performed using a reverse transcription kit (Toyobo, Japan) The
reaction conditions were as follows: 30°C for 10 min, 42°C for 20
min and 95°C for 5 min. The ΔΔCt method was applied for the
quantitative analysis (18).
Primer sequences used in this study were: aggrecan, forward: 5′-TGA
GTC CTC AAG CCT CCT GT-3′ and reverse: 5′-GTC CCT CTG TCT CCT TGC
AG-3′; TNF-α, forward: 5′-TCT CTT CAA GGG ACA AGG CTG-3′ and
reverse: 5′-ATA GCA AAT CGG CTG ACG GT-3′; COX-2, forward: 5′-TGA
ACC CAC TCC AAA CAC A-3′ and reverse: 5′-CAG CAA ACC GTA GAT GCT
CA-3′. PCR products were analyzed using agarose gel
electrophoresis. The levels of IL-18Rα and IL-18Rβ expressions were
determined as described in the study by Moller et al
(19).
Statistical analysis
Statistical analysis was performed using the SPSS
13.0 software. Cytokine concentrations in the supernatant from
cultured cells were shown as the mean ± SE. P<0.05 was
considered to indicate a statistically significant difference. Two
independent-sample t-tests were conducted to analyze the effects of
IL-18 on synoviocytes. The effects of different IL-18
concentrations on synoviocytes were analyzed using one-way analysis
of variance (ANOVA). When statistically significant difference was
obtained, multiple comparisons were carried out, using the LSD test
or Dunnetts’ T3 test (unequal variances).
Results
Characterization and morphology of
synoviocytes and chondrocytes
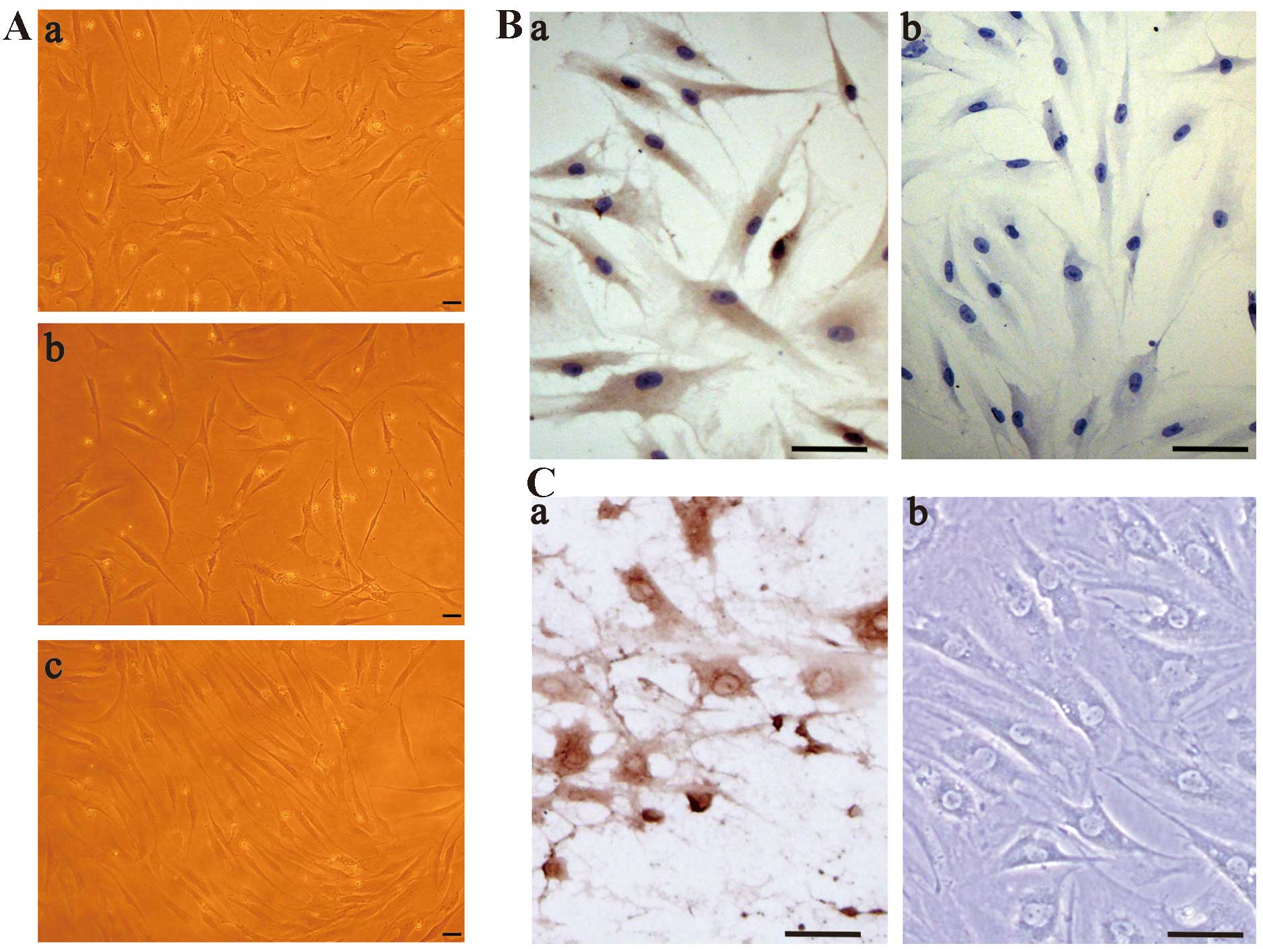
There were 2 morphological forms of primary
synoviocytes (G1-synoviocytes): spindle-shaped fibroblast-like
synoviocytes and polygonal macrophage-like synoviocytes (Fig. 1Aa). The number of polygonal
macrophage-like synoviocytes decreased in the secondary generation
of synoviocytes (G2-synoviocytes) (Fig. 1Ab). The majority of cells in the
third generation (G3-synoviocytes) were spindle-shaped, i.e., they
were fibroblast-like synoviocytes (Fig. 1Ac). The numbers of the
macrophage-like cells decreased with increasing passage numbers.
Type I collagen expression in synoviocytes was detected by
immunohistochemistry (Fig. 1B).
The cytosol of the synoviocytes was stained brown, indicating a
high level of type I collagen expression. Type I collagen
expression in cells with the two forms was similar. Chondrocytes
were stained with an antibody against type II collagen, while
reticular type II collagen was visible in the intracellular space
(Fig. 1Ca).
IL-18-induced inflammatory response in
synoviocytes
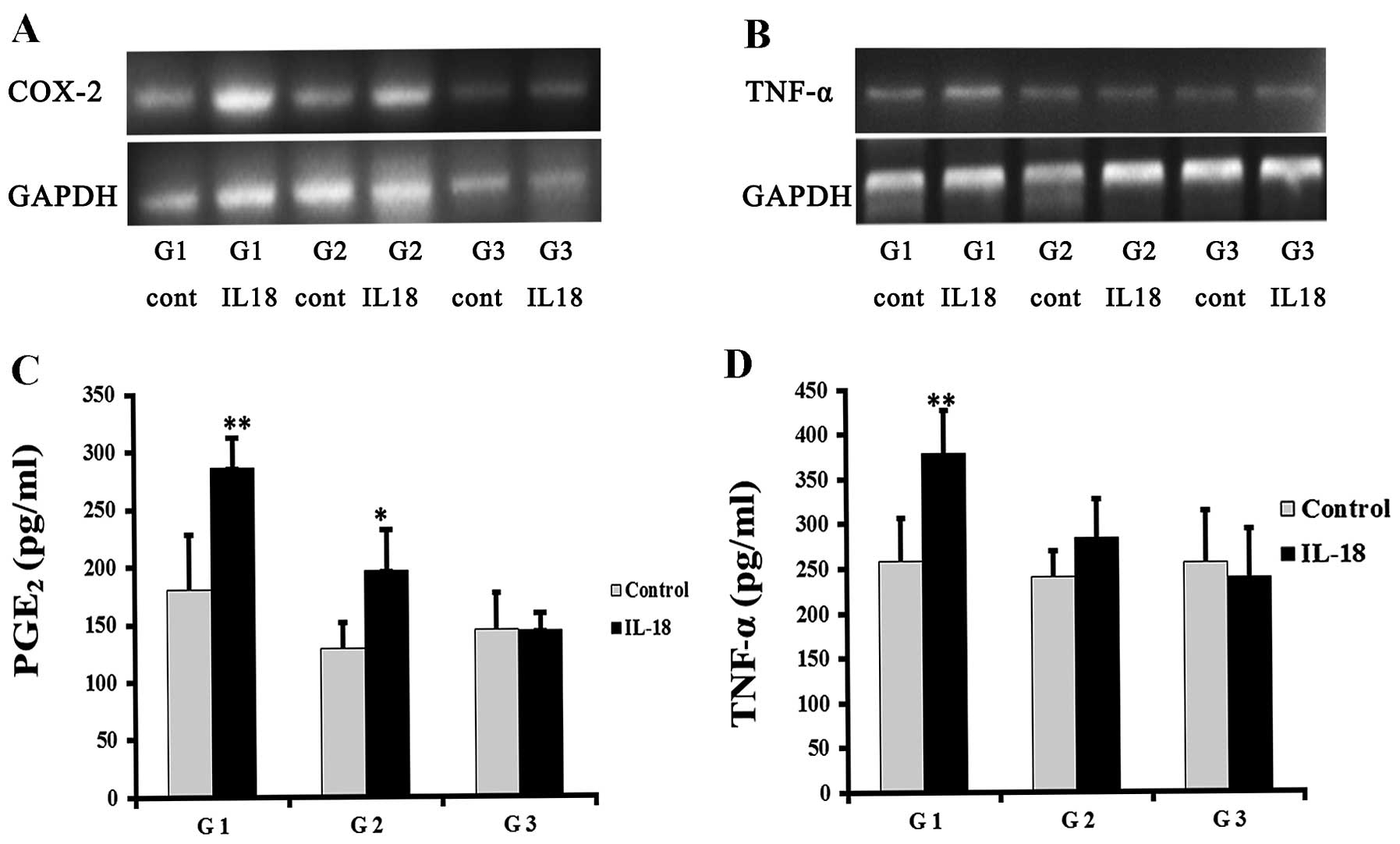
G1- and G2-synoviocytes demonstrated an elevated
COX-2 expression subsequent to IL-18 induction, while
G3-synoviovytes showed no significant change (Fig. 2A). All the synoviocytes
demonstrated in vitro PGE2 production. G1- and
G2-synoviocytes in the IL-18-treated groups had markedly higher
levels of PGE2 expression, compared to the control group
(286.21±26.24 vs. 180.31±47.69 pg/ml, 196.60±34.79 vs. 128.72±22.37
pg/ml; P=0.005 and P=0.011, respectively) (Fig. 2C). Similarly, IL-18 induced mRNA
expression in G1-synoviocytes, although not in G2-synoviocytes
(Fig. 2B). The results were
consistent with the protein assay results. There was a
statistically significant difference in the TNF-α concentration in
the G1-synoviocyte supernatant in the IL-18-treated and the control
group (379.20±47.60 vs. 257.90±48.76 pg/ml; P= 0.007) (Fig. 2D). However, G3-synoviocytes were
less sensitive to IL-18 induction, whereas the TNF-α and
PGE2 levels were not increased (P>0.05).
Response of the chondrocytes to IL-18
stimulation
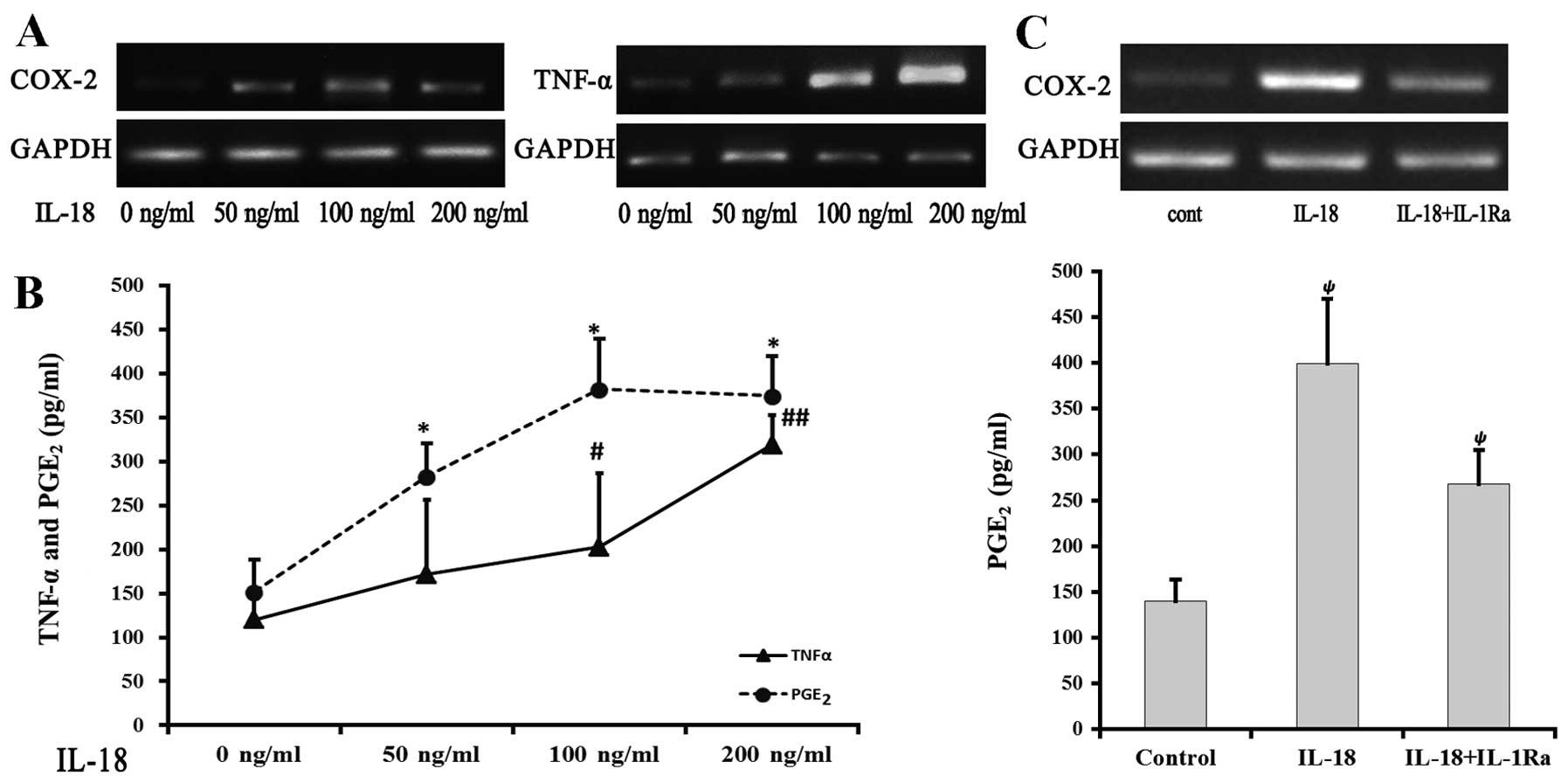
An increase was observed in COX-2 and TNF-α mRNA
expression with increasing concentrations of IL-18 (Fig. 3A). PGE2 levels in the
50, 100 and 200 ng/ml IL-18 groups were 282.83±52.16, 381.85±84.15
and 374.51±34.14 pg/ml, respectively, which were significantly
higher compared to the control group (151.72±23.76 pg/ml; P=0.003,
P<0.001 and P<0.001, respectively). In addition, the 100 and
200 ng/ml IL-18 groups showed significantly higher TNF-α levels
compared to the control group (202.48±57.46 and 318.63±45.23 pg/ml;
P=0.002 and P<0.001, respectively) (Fig. 3B). The induction of
PGE2 and TNF-α by IL-18 was dose-dependent.
To elucidate the correlation between IL-18 and
IL-1β-induced inflammatory responses in chondrocytes, chondrocytes
in the IL-18 group were treated with 100 ng/ml IL-18, while those
in the IL-1Ra group were treated with 100 ng/ml IL-18 and IL-1Ra.
After 24-h incubation, the IL-18 group demonstrated a significantly
higher COX-2 mRNA expression, compared to the other 2 groups
(Fig. 3C). The IL-18 group also
had a markedly higher level of PGE2, compared to the
control group (P=0.003). The PGE2 level decreased
following addition of the inhibitor, although this level was still
higher when compared to the control group (P=0.002).
Expression of IL-18R
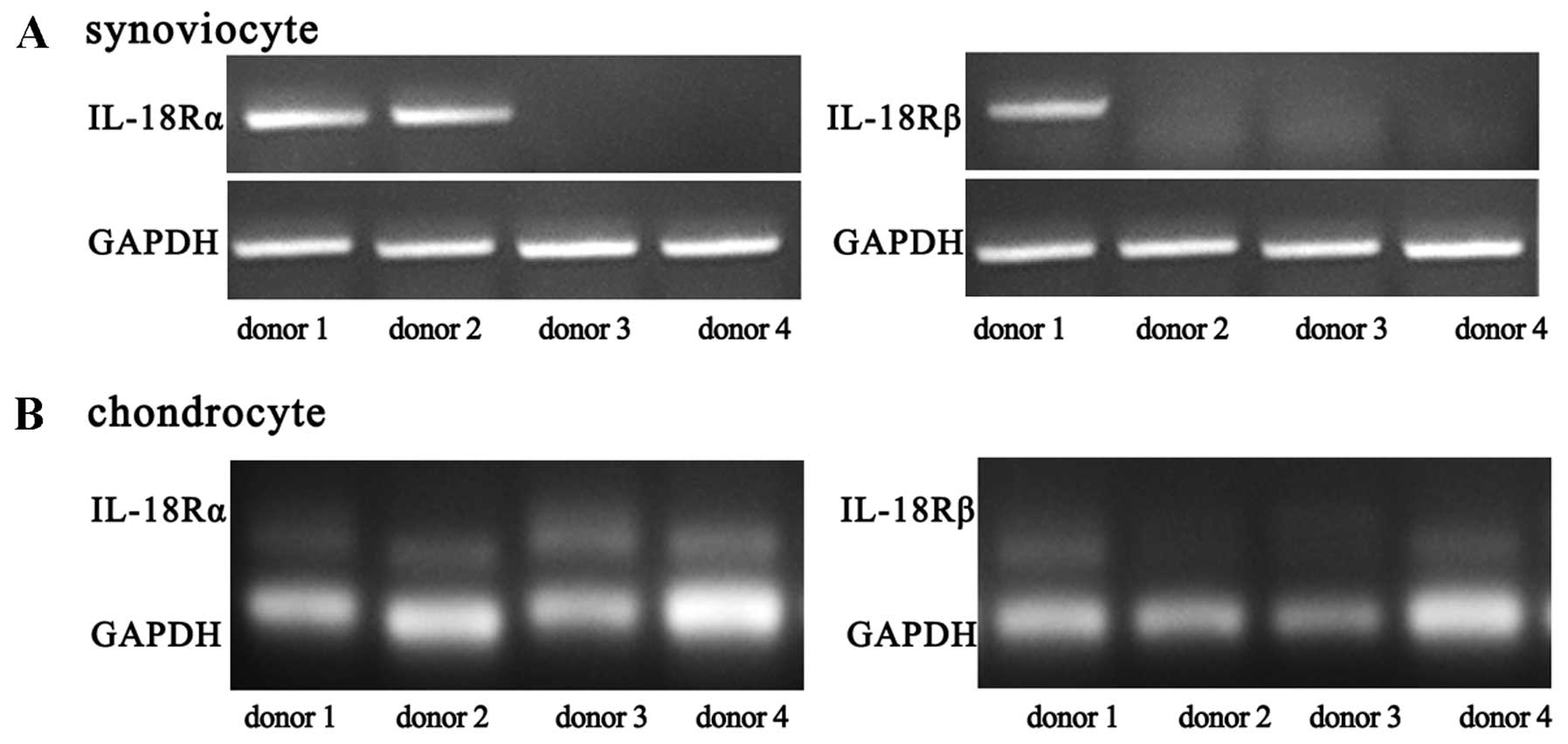
The primary synoviocytes of 8 patients were obtained
and analyzed. The synoviocytes of two of the 8 donors had
detectable IL-18Rα expression, and those of one donor had IL-18Rβ
expression (Fig. 4A). After
passage, no receptor expression was detected. Chondrocytes from 4
donors expressed IL-18Rα, whereas those from 2 donors expressed
IL-18Rβ (Fig. 4B).
Discussion
The present study investigated the impact of IL-18
on synoviocytes and chondrocytes. The results showed that high
levels of IL-18 induced the expression of inflammatory factors,
such as TNF-α, PGE2 and COX2 in primary synoviocytes.
With the increase of passage numbers, fibroblast-like cells became
more dominant, IL-18R expression decreased, while the synoviocytes
became less responsive to IL-18 stimulation. IL-18 had the
potential to induce the expression of PGE2, TNF-α and
COX-2 in human OA chondrocytes, which express IL-18Rα and
IL-18Rβ.
IL-18 induced inflammatory responses in OA
synoviocytes. The knee synovial membrane is composed of two types
of cells: the macrophage-like synoviocytes known as A cells, and
the fibroblast-like synoviocytes known as B cells. Goto et
al (21) have differentiated
and purified these two types of cells, using cell cloning (20). Type A cells proliferate slowly,
are less active and lose activity after 10 days of culture
(22). Primary synoviocytes from
OA patients contained two cell types expressing IL-18. They
produced high TNF-α, PGE2 and COX-2 levels when induced
by IL-18. Such a pro-inflammatory response is also noted in RA
patient-derived synoviocytes. In a mouse model of collagen-induced
arthritis (CIA), mature IL-18 induced synovial proliferation,
synovial inflammation and cartilage damage (23). It has even been attempted to
neutralize intrinsic IL-18 in RA patients with a view to treat the
disease (24). Synoviocytes from
OA patients express higher levels of IL-18 than those from healthy
individuals. The elevated levels of typical inflammatory factors,
such as PGE2 and TNF in OA in the joint fluid is closely
associated with the clinical presentations of OA (25).
However, not all the synoviocytes are equally
sensitive to IL-18. Primary cells secrete more inflammatory factors
upon IL-18 stimulation. As passage numbers increased, the
production of PGE2 and TNF-α was less affected by IL-18
treatment. In the present study, only IL-18Rα was detected in
primary OA synoviocytes, while IL-18Rβ was virtually absent. This
finding suggested that IL-18 targets A cells, instead of B cells.
Since the number of A cells decreased or even disappeared with
increasing culture time, IL-18R expression disappeared along with
the pro-inflammatory effects of IL-18 on synoviocytes. This is
similar to RA synoviocytes: IL-18 induces inflammation and joint
destruction through the stimulation of T-cells and macrophages,
instead of fibroblast-like synoviocytes, which lack IL-18R.
Excessive IL-18 triggers inflammatory responses in
chondrocytes. In this study, IL-18 was found to be able to induce
TNF-α and PGE2 expressions and to trigger excessive
COX-2 production in chondrocytes. COX-2 degrades arachidonic acid
to generate PGE2. An elevated level of PGE2
has the potential to generate the metabolic imbalance and
degradation of cartilage (26).
TNF-α is a contributing factor of inflammation in OA, and its level
in the joint fluid is closely correlated with the progression of OA
(27–29). In addition, in this study, all the
OA chondrocytes expressed IL-18Rα, while certain cells expressed
IL-18Rβ. IL-18 stimulates cells through the two receptors,
triggering signal transduction via the NF-κB pathway thus inducing
cellular inflammatory responses (30).
IL-18 and IL-1β affect chondrocytes in an
interactive yet independent way. IL-1β promotes the secretion of
inflammatory factors and MMPs in chondrocytes, leading to the
degradation of cartilage matrix. IL-1β is considered to be a
typical pro-inflammatory factor in OA pathogenesis (31,32). IL-18 has similar functions to
IL-1β, while both promote the production of inflammatory factors in
chondrocytes. Our results have demonstrated that IL-18 induced
inflammatory responses in chondrocytes despite the blockage of the
IL-1β function by IL-1Ra, although the PGE2 and TNF-α
levels were lowered, indicating that the effect of IL-18 is only
partially dependent on IL-1β, possibly due to their distinct
signaling transduction pathways. IL-18 induces COX-2 through the
MAPK-p38-AP1 and NF-κB pathways (33,34), whereas IL-1β induces MMP through
the p38 APK/c-Fos/AP-1 and JAK2/STAT1/2 pathways. The utilization
of different signaling pathways by IL-18 and IL-1β led to their
disparate effects on chondrocytes.
In conclusion, synoviocytes and chondrocytes from OA
patients express IL-18R. IL-18 were able to induce inflammatory
responses in the two types of cells. This pro-inflammatory effect
was partially independent on IL-1 in chondrocytes.
Acknowledgements
This study was supported by research
grants from the Guangdong Natural Science Foundation of China
(S2011010003871) and the Research Fund for the Doctoral Program of
Higher Education (20114433110016).
References
|
1.
|
MK HaynesEL HumeJB SmithPhenotypic
characterization of inflammatory cells from osteoarthritic synovium
and synovial fluidsClin
Immunol105315325200210.1006/clim.2002.528312498813
|
|
2.
|
MD SmithS TriantafillouA ParkerPP YoussefM
ColemanSynovial membrane inflammation and cytokine production in
patients with early osteoarthritisJ
Rheumatol2436537119979034998
|
|
3.
|
CA DinarelloThe IL-1 family and
inflammatory diseasesClin Exp Rheumatol20S1S13200214989423
|
|
4.
|
GL BjornssonL ThorsteinssonKO GudmundssonH
Jonsson JrS GudmundssonB GudbjornssonInflammatory cytokines in
relation to adrenal response following total hip replacementScand J
Immunol6599105200710.1111/j.1365-3083.2006.01872.x17212773
|
|
5.
|
SM DaiZZ ShanH XuK NishiokaCellular
targets of interleukin-18 in rheumatoid arthritisAnn Rheum
Dis6614111418200710.1136/ard.2006.06779317502360
|
|
6.
|
M MohtaiMK GuptaB DonlonExpression of
interleukin-6 in osteoarthritic chondrocytes and effects of
fluid-induced shear on this expression in normal human chondrocytes
in vitroJ Orthop Res146773199610.1002/jor.11001401128618168
|
|
7.
|
H OkamuraH TsutsiT KomatsuCloning of a new
cytokine that induces IFN-gamma production by T
cellsNature3788891199510.1038/378088a07477296
|
|
8.
|
T GhayurS BanerjeeM HuguninCaspase-1
processes IFN-gamma-inducing factor and regulates LPS-induced
IFN-gamma productionNature386619623199710.1038/386619a09121587
|
|
9.
|
K MatsuiH TsutsuiK
NakanishiPathophysiological roles for IL-18 in inflammatory
arthritisExpert Opin Ther
Targets7701724200310.1517/14728222.7.6.70114640907
|
|
10.
|
A PawlikM KurzawskiM DrozdzikV
DziedziejkoK SafranowM HerczynskaInterleukin-18 gene (IL-18)
promoter polymorphisms in patients with rheumatoid arthritisScand J
Rheumatol38159165200910.1080/0300974080260074819229765
|
|
11.
|
XT ShaoL FengLJ GuExpression of
interleukin-18, IL-18BP, and IL-18R in serum, synovial fluid, and
synovial tissue in patients with rheumatoid arthritisClin Exp
Med9215221200910.1007/s10238-009-0036-219225717
|
|
12.
|
B MollerN Kukoc-ZivojnovU
KesslerExpression of interleukin-18 and its monokine-directed
function in rheumatoid arthritisRheumatology
(Oxford)40302309200110.1093/rheumatology/40.3.30211285378
|
|
13.
|
M LotzFJ BlancoJ von KempisCytokine
regulation of chondrocyte functionsJ Rheumatol
Suppl431041081995
|
|
14.
|
T OleeS HashimotoJ QuachM LotzIL-18 is
produced by articular chondrocytes and induces proinflammatory and
catabolic responsesJ Immunol1621096110019999916738
|
|
15.
|
LA JoostenRL SmeetsMI
KoendersInterleukin-18 promotes joint inflammation and induces
interleukin-1-driven cartilage destructionAm J
Pathol165959967200410.1016/S0002-9440(10)63357-315331419
|
|
16.
|
T ZimmermannE KunischR PfeifferIsolation
and characterization of rheumatoid arthritis synovial fibroblasts
from primary culture - primary culture cells markedly differ from
fourth-passage cellsArthritis Res37276200110.1186/ar142
|
|
17.
|
S FrankeM SommerC RusterAdvanced glycation
end products induce cell cycle arrest and proinflammatory changes
in osteoarthritic fibroblast-like synovial cellsArthritis Res
Ther11R136200910.1186/ar280719735566
|
|
18.
|
KJ LivakTD SchmittgenAnalysis of relative
gene expression data using real-time quantitative PCR and the
2(-Delta Delta C(T))
methodMethods25402408200110.1006/meth.2001.126211846609
|
|
19.
|
B MollerU KesslerS RehartExpression of
interleukin-18 receptor in fibroblast-like synoviocytesArthritis
Res4139144200210.1186/ar39011879550
|
|
20.
|
WN UengCC ChuangCH ShihDouble-rib
composite free transfer to reconstruct a single-spared lower
extremity defectJ
Trauma38210212199510.1097/00005373-199502000-000107869437
|
|
21.
|
M GotoM SasanoH YamanakaSpontaneous
production of an interleukin 1-like factor by cloned rheumatoid
synovial cells in long-term cultureJ Clin
Invest80786796198710.1172/JCI1131352442197
|
|
22.
|
SW UengDC ChuangSL ChengCH ShihManagement
of large infected tibial defects with radical debridement and
staged double-rib composite free transferJ
Trauma40345350199610.1097/00005373-199603000-000038601847
|
|
23.
|
JA GracieRJ ForseyWL ChanA proinflammatory
role for IL-18 in rheumatoid arthritisJ Clin
Invest10413931401199910.1172/JCI731710562301
|
|
24.
|
C Plater-ZyberkLA JoostenMM
HelsenTherapeutic effect of neutralizing endogenous IL-18 activity
in the collagen-induced model of arthritisJ Clin
Invest10818251832200110.1172/JCI20011209711748266
|
|
25.
|
MB GoldringM OteroDA PlumbRoles of
inflammatory and anabolic cytokines in cartilage metabolism:
signals and multiple effectors converge upon MMP-13 regulation in
osteoarthritisEur Cell Mater21202220201121351054
|
|
26.
|
X LiM EllmanP MuddasaniProstaglandin E2
and its cognate EP receptors control human adult articular
cartilage homeostasis and are linked to the pathophysiology of
osteoarthritisArthritis
Rheum60513523200910.1002/art.2425819180509
|
|
27.
|
BY ChanES FullerAK RussellIncreased
chondrocyte sclerostin may protect against cartilage degradation in
osteoarthritisOsteoarthritis
Cartilage19874885201110.1016/j.joca.2011.04.01421619935
|
|
28.
|
VA MaganevRA DavletshinGK DavletshinaGS
IapparovDynamics of tumour necrosis factor-alpha and clinical signs
of osteoarthrosis during the treatment with alflutop in combination
with peloid applications under conditions of health resortVopr
Kurortol Fizioter Lech Fiz Kult18202011(In Russian)
|
|
29.
|
S OritaT KoshiT MitsukaAssociations
between proinflammatory cytokines in the synovial fluid and
radiographic grading and pain-related scores in 47 consecutive
patients with osteoarthritis of the kneeBMC Musculoskeletal
Disord12144201110.1186/1471-2474-12-144
|
|
30.
|
M KawashimaP MiossecHeterogeneity of
response of rheumatoid synovium cell subsets to interleukin-18 in
relation to differential interleukin-18 receptor
expressionArthritis Rheum48631637200310.1002/art.1082512632414
|
|
31.
|
H LimHP KimMatrix metalloproteinase-13
expression in IL-1β-treated chondrocytes by activation of the p38
MAPK/c-Fos/AP-1 and JAK/STAT pathwaysArch Pharm Res341091172011
|
|
32.
|
R SinghS AhmedCJ MalemudVM GoldbergTM
HaqqiEpigallocatechin-3-gallate selectively inhibits
interleukin-1beta-induced activation of mitogen activated protein
kinase subgroup c-Jun N-terminal kinase in human osteoarthritis
chondrocytesJ Orthop
Res21102109200310.1016/S0736-0266(02)00089-X
|
|
33.
|
B ChandrasekarS MummidiL
MahimainathanInterleukin-18-induced human coronary artery smooth
muscle cell migration is dependent on NF-kappaB- and AP-1-mediated
matrix metalloproteinase-9 expression and is inhibited by
atorvastatinJ Biol Chem2811509915109200610.1074/jbc.M600200200
|
|
34.
|
JK LeeSH KimEC LewisT AzamLL ReznikovCA
DinarelloDifferences in signaling pathways by IL-1beta and
IL-18Proc Natl Acad Sci
USA10188158820200410.1073/pnas.040280010115161979
|