Introduction
Prostate cancer (PCa) is a leading cause of cancer
mortality and is among the most common tyeps of cancer in Western
countries (1). In PCa, recurrent
gene fusions occurring between the transmembrane protease, serine 2
(TMPRSS2) gene, an androgen-regulated prostate-specific
serine protease and members of the erythroblast
transformation-specific (ETS) family of transcription factors [ETS
variant gene (ETV)-1, ETV-4, ETV-5 and most
commonly, ETS-related gene ERG] resulting in the increased
expression of the latter rearranged ETS members in response to
androgens, are frequently found (2–4).
The ETS family of transcription factors which is
comprised of 27 members in humans (5) has been reported to be involved in
many processes such as development, differentiation, proliferation,
apoptosis, migra tion, tissue remodeling, invasion and angiogenesis
in a wide range of cells (such as B-cells, endothelial cells,
fibroblasts, as well as neoplastic cells) (6–13).
The ETS family is defined by the presence of an evolutionary
conserved DNA-binding domain, termed the ETS domain, which is
comprised of approximately 80 amino acids with four tryptophan
repeats and recognizes DNA sequences with a GGAA/T core motif
(14,15). Phylogenetic analysis of the human
ETS domains has led to the identification of subfamilies of more
highly related members (16).
Even though different ETS family members may have
different functions due to their binding preferences for distinct
flanking sequences around the GGAA/T core motif enabling them to
bind more specifically (16),
overlapping func tions of ETS members and redundant occupancy at
gene regions have been described (16,17). Furthermore, even though the ETS
transcription factors may have independent activities, it is likely
that they are part of an integrated network, in which gene
regulation may be influenced by the binding equilibrium and the
activity of the ETS transactivation domains, as well as the complex
formation of different ETS members and other factors (18,19).
While a number of studies have focused on single ETS
factors within a single tissue and within the context of specific
promoters, the functional impact of multiple ETS members present
within a specific cell type have not yet been investigated
(18), particularly in PCa. While
multiple ETS factors may be able to regulate the same sets of
genes, the magnitude or the directions may be different for each of
the ETS factors (18).
As the most prominent gene rearrangement in PCa
leads to the overexpression of ERG (20–23), combined with functional studies
showing that ERG knockdown induces morphological changes,
inhibits cell growth in both culture and mice, and that ERG
overexpression leads to an increase in cell invasion (24), the aim of the present study was to
investigate whether ERG is part of a complex integrated
transcriptional network that involves other ETS factors which are
highly likely to cooperate or influence the activity of ERG in
PCa.
More specifically, as the ETS family of
transcription factors consists of 27 members (5), we decided to focus our efforts
initially on investigating whether ERG is associated with three
well-known members of the family, ETS-1, ETS-2 and ETV-4, in PCa as
a proof of principle. The rationale behind choosing the latter ETS
members was that ETS-1, the prototype of the ETS family, is
overexpressed in latent as well as clinically manifest PCa
(25), ETS-2 is also
overexpressed in PCa (18), ETS-2
and ETS-1 play redundant roles (17), ETS-2 is associated with in
vitro synthesized ETS-1 (26), ETS-2 interacts with ERG in
vivo demonstrated by the two-hybrid system (26) and that ETV-4 is rearranged in PCa,
similar to ERG (2–4). The results from a previous study
were also taken into account, namely that the occurrence of
multiple ETS rearrangements within one prostate gland, within the
same tumor focus and within the same nucleus (27).
Materials and methods
Western blot analysis
The expression of ERG, ETS-1, ETS-2 and ETV-4 in PC3
cell nuclear extracts (Santa Cruz Biotechnology, Inc., Santa Cruz,
CA, USA) was determined by western blot analysis using a mouse
monoclonal anti-ERG antibody (Bio-Care, Holt, MI, USA), a mouse
monoclonal anti-ETS-1 antibody (Transduction Laboratories,
Lexington, KY, USA), a rabbit polyclonal anti-ETS-2 antibody
(Sigma-Aldrich, Munich, Germany) and a mouse monoclonal anti-ETV-4
antibody (BioCat, Heidelberg, Germany), respectively.
In protein lysates prepared from human PCa
prostatectomy specimens of five patients, the expression of ERG,
ETS-1, ETS-2 and ETV-4 was determined by western blot analysis
using a mouse monoclonal anti-ERG antibody (Biocare), a mouse
monoclonal anti-ETS-1 antibody (Transduction Laboratories) and a
rabbit polyclonal C-20 anti-ETS-1 antibody (Santa Cruz
Biotechnology, Inc.), a rabbit polyclonal anti-ETS-2 antibody
(Sigma-Aldrich) and a mouse monoclonal anti-ETV-4 antibody
(BioCat), respectively.
Human PCa prostatectomy specimens and
protein lysate preparations
Briefly, fresh tissue samples from five patients
with prostate carcinomas (Gleason scores, 6, 6 7, 7 and 8) were
taken immediately after radical prostatectomy. The tissue samples
were then shock-frozen in liquid nitrogen with ice-cold isopentane
as described previously (28).
Thereafter, 6-μm-thick frozen sections were cut from the samples
using a cryotome (Leica, Berlin, Germany) and mounted on
conventional slides followed by staining with hematoxylin and eosin
(H&E) for diagnostic evaluation by an experienced pathologist.
The preparation of protein lysates from the latter prostate
carcinoma tissue samples was carried out as previously described
(29).
Immunoprecipitation (IP)
To investigate whether ERG is associated with ETS-1,
ETS-2 and ETV-4 in PC3 cell nuclear extracts (Santa Cruz
Biotechnology, Inc.), we performed IP using a rabbit polyclonal
anti-ERG antibody (Gentex, Zeeland, MI, USA) as it exhibited the
best compatibility with our IP compared to all the other
commercially available antibodies that we tested. Briefly, PC3 cell
nuclear extracts were pre-cleared with protein A agarose beads
(Sigma-Aldrich) by rotation at 4°C for 2 h. An additional tube
containing protein A agarose beads (Sigma-Aldrich) and the rabbit
polyclonal anti-ERG antibody (Gentex) was incubated by rotation at
4°C for 2 h. Both tubes were then centrifuged at 2,000 x g for 2
min and the pre-cleared supernatant was then added to the ERG-bound
protein A agarose beads. The sample was then incubated by rotation
for 1 h at 4°C, collected by centrifugation at 2,000 x g for 2 min,
and then washed three times with a nuclear extract buffer (20 mM
Hepes, 25% glycerol, 2 M KCL, 1 mM MgCl2, 1%
Nonidet-p40, 0.5 mM EDTA). The beads were then re-suspended in a
loading buffer, boiled and loaded on an SDS/PAGE gel, followed by
western blot analysis using a mouse monoclonal anti-ERG antibody
(Biocare), a mouse monoclonal anti-ETS-1 antibody (Transduction
Laboratories), a rabbit polyclonal anti-ETS-2 antibody
(Sigma-Aldrich) and a mouse monoclonal anti-ETV-4 antibody
(BioCat), respectively.
In order to investigate whether ERG is associated
with ETS-1, ETS-2 and ETV-4 in the protein lysates prepared from
the above five PCa prostatectomy specimens, we first pooled all the
five protein lysates and then performed IP as described above.
Western blot analysis was performed with the same antibodies used
for the above IP and additionally with a rabbit polyclonal C-20
anti-ETS-1 antibody (Santa Cruz Biotechnology, Inc.).
Results
Protein expression of ERG, ETS-1, ETS-2
and ETV-4 in PC3 cell nuclear extracts and in protein lysates
prepared from human PCa prostatectomy specimens
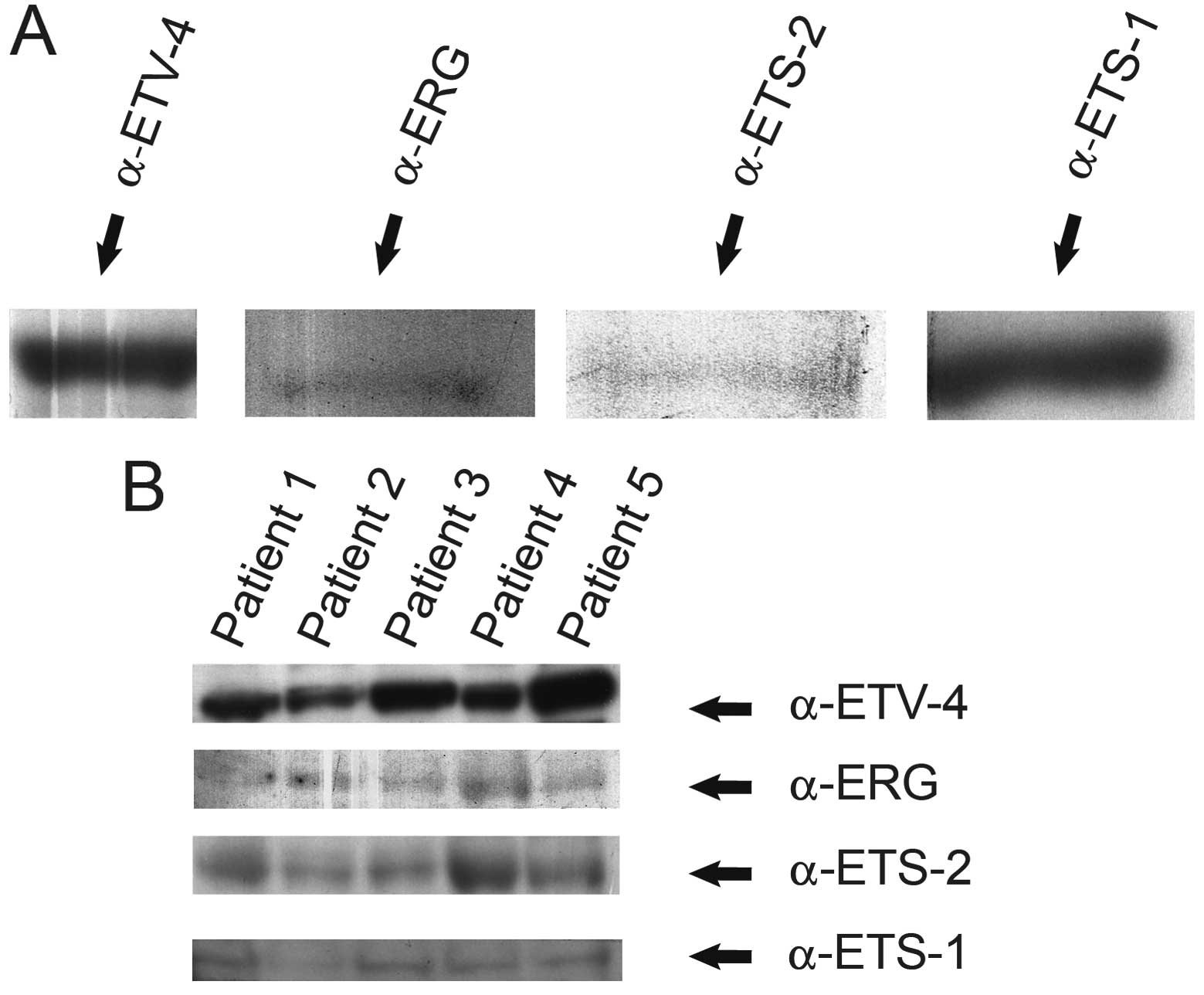
As shown by western blot analysis using specific
antibodies, ERG, ETS-1, ETS-2 and ETV-4 were expressed in PC3 cell
nuclear extracts (Santa Cruz Biotechnology, Inc.), as well as in
protein lysates prepared from the human PCa prostatectomy specimens
of five patients (Fig. 1).
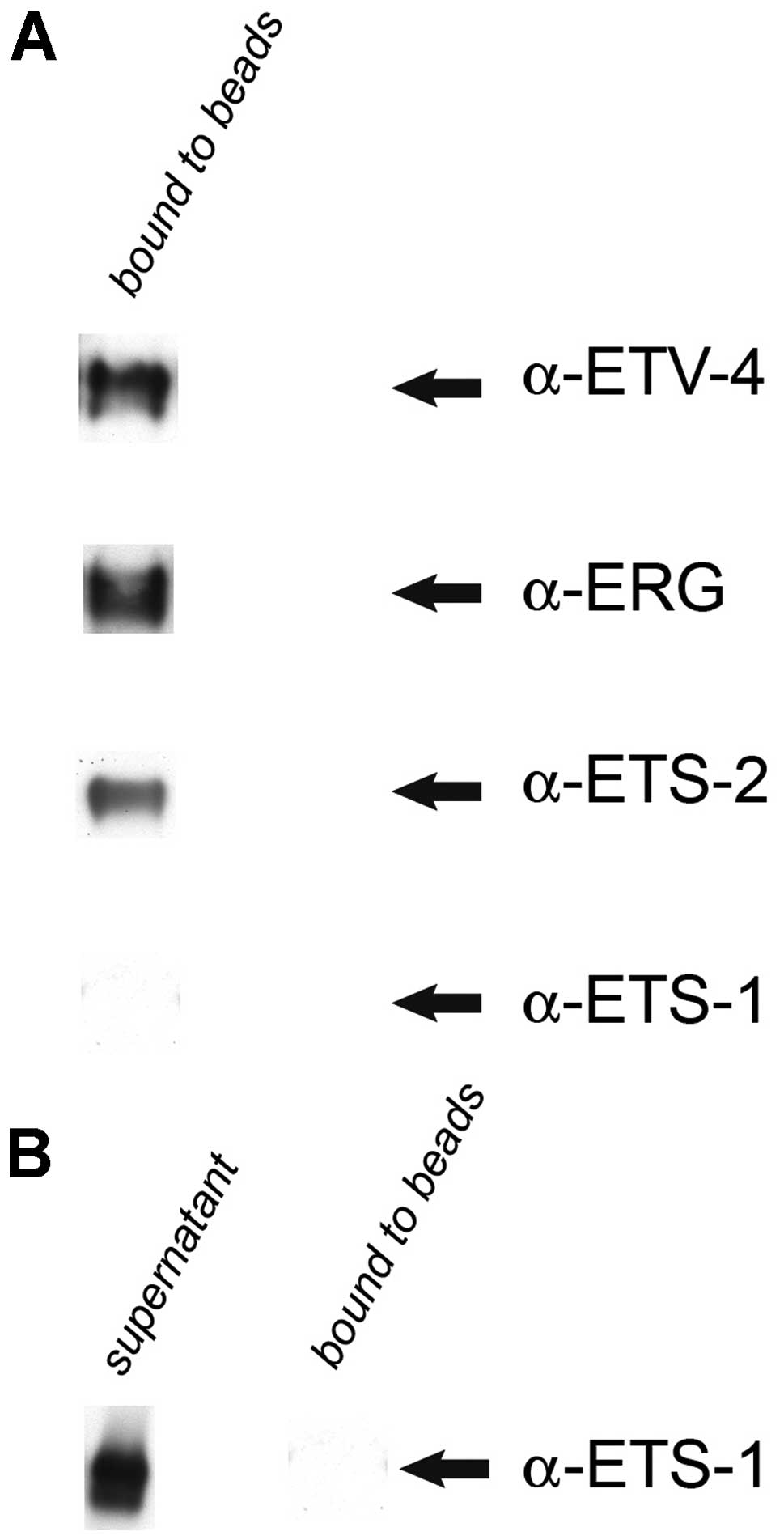
ERG is specifically associated with ETS-2
and ETV-4, but not with ETS-1, in PC3 cell nuclear extracts
To investigate whether ERG is associated with ETS-1,
ETS-2 and ETV-4 in PC3 cell nuclear extracts (Santa Cruz
Biotechnology, Inc.), IP was performed using a rabbit polyclonal
anti-ERG antibody (Gentex). Following western blot analysis, the
results revealed that ERG is specifically associated with ETS-2 and
ETV-4, but not with ETS-1 (Fig.
2).
ERG is specifically associated with ETS-2
and ETV-4, but not with ETS-1, in protein lysates prepared from
human PCa prostatectomy specimens
Using IP with a rabbit polyclonal anti-ERG antibody
(Gentex), we investigated whether ERG is associated with ETS-1,
ETS-2 and ETV-4 in a pooled protein lysate sample that was prepared
from the PCa prostatectomy specimens of five patients. Following
western blot analysis, the results revealed that ERG is
specifically associated with ETS-2 and ETV-4, but not with ETS-1
(Fig. 3).
Discussion
The most prominent gene rearrangement in PCa arises
between the androgen-regulated prostate-specific serine protease
TMPRSS2 gene and the ETS transcription factor ERG
gene, leading to the overexpression of ERG (20–23). The ERG rearrangement is
highly specific to PCa (30), is
maintained in advanced disease (2,31),
and depending on the cohort design and the histological subtypes of
PCa (20,32–34) approximately 15–80% of PCa patients
harbour the TMPRSS2-ERG fusions (2–4,35,36). Furthermore, the knockdown of
ERG has been reported to induce morphological changes and
the inhibition of cell growth in both cell cultures and mice,
whereas ERG overexpression has been shown to result in an
increase in cell invasion (24).
Even though ETS transcription factors, such as ERG
may have independent activities in PCa, ERG is highly likely to be
part of a complex integrated transcriptional network that involves
other ETS factors. In such a network, gene regulation may be
influenced by the binding equilibrium, the activity of ETS
transactivation domains and a complex formation of different ETS
members and other factors, and even if multiple ETS factors may be
able to regulate the same sets of genes, the magnitude or the
directions may be different for each of the ETS factors (18,19).
While the focus of many studies has been on single
ETS factors within a single tissue and within the context of
specific promoters, the functional impact of multiple ETS factors
present within a specific cell type has not yet been well
investigated (18), particularly
in PCa. Therefore, the aim of this study was to investigate whether
ERG is associated with specific ETS transcription factors in PCa.
More specifically, we investigated whether ERG is associated with
ETS-1, ETS-2 and ETV-4 in PC3 cell nuclear extracts, as well as in
PCa tissues.
As the ETS family of transcription factors consists
of 27 members in humans (5), we
decided to focus our efforts initially on investigating whether is
ERG associated with three well-known members of the family, ETS-1,
ETS-2 and ETV-4, in PCa as a proof of principle. The rationale
behind choosing the latter ETS members was as follows: ETS-1 is the
prototype of the ETS family and has been reported to be
overexpressed in latent as well as clinically manifest PCa and a
strong expression of ETS-1 has been associated with poor tumor
differentiation (25). As shown
in a previous study of our, the blockade of ETS-1 in PCa cell lines
results in a decrease in cell migration (11) and has a major effect upon genes
involved in the metastatic cascade (12). As ETS-1 and ETS-2 have been
reported to play redundant roles during embryonic development
(17), combined with reports that
ETS-2 is associated with in vitro synthesized ETS-1
(26) and that ETS-2 interacts
with ERG in vivo demonstrated by the two-hybrid system
(26), we decided to investigate
ETS-2 as well. ETS-2 has been reported to be overexpressed in PCa
(18), and a blockade of ETS-2
has been shown to reduce the transformed properties of PCa cells
(37), as well as growth
inhibition and apoptosis (38).
ETV-4 on the other hand, was chosen as ETV-4 is known to be
rearranged in PCa, similar to ERG (2–4),
combined with a report showing the occurrence of multiple ETS
rearrangements within one prostate gland, within the same tumor
focus and within the same nucleus (27). Moreover, we recently reported the
ETV-4 rearranged gene status in primary PCas and their
corresponding lymph nodes, and found the rearrangement in 6% of
both primary PCas and the corresponding lymph node metastases
(39). Furthermore, ETV-4 has
been shown to be required for anchorage-independent growth and cell
proliferation gene expression program in PCa cell lines (40).
Prior to investigating whether is ERG associated
with ETS-1, ETS-2 and ETV-4, we first examined whether these latter
proteins are expressed in PC3 cell nuclear extracts and in protein
lysates prepared from human PCa prostatectomy specimens. We found
by western blot analysis that ERG, ETS-1, ETS-2 and ETV-4 are
expressed in both PC3 cell nuclear extracts as well as in protein
lysates prepared from the PCa tissue samples of five patients
(Fig. 1).
Upon confirming the expression of ERG, ETS-1, ETS-2
and ETV-4 (Fig. 1), we performed
IP using an anti-ERG antibody to investigate whether ERG is
associated with ETS-1, ETS-2 and ETV-4 in PC3 cell nuclear
extracts, as well as in a pooled protein lysate sample prepared
from the PCa tissue samples of five patients.
Of note, our results revealed that ERG is
specifically associated with ETS-2 and ETV-4, but not with ETS-1,
in PC3 cell, nuclear extracts (Fig.
2) and even more remarkably, we found ERG to exhibit the same
specificity for ETS-2 and ETV-4, but not for ETS-1 in the pooled
protein lysate sample prepared from the PCa tissue samples of five
patients (Fig. 3).
Our observation that ERG is not associated with
ETS-1 and ETS-2, but rather only with ETS-2, may be due to the fact
that ETS-1 and ETS-2 have been reported to play redundant roles
(17), which would make it
counterproductive for ERG to be associated with ETS-1 and ETS-2
simultaneously. The association of ERG with ETS-2 based on our
findings is supported by a previous study reporting that ERG
interacts with ETS-2 in vivo using the two-hybrid system
(26). Our finding that ERG is
associated with ETV-4 is highly intriguing, as ETV-4 is
known to be rearranged in PCa, similar to ERG (2–4,39),
combined with a recent report showing the occurrence of multiple
ETS rearrangements within one prostate gland, within the same tumor
focus and within the same nucleus (27), which may imply a potential
cooperation or interaction between rearranged ETS genes in PCa.
Taken together, our findings that ERG is
specifically associated with ETS-2 and ETV-4, but not with ETS-1,
in PC3 cell nuclear extracts and PCa tissues strongly support the
notion that ERG is part of a complex integrated transcriptional
network that involves other ETS factors, such as ETS-2 and ETV-4,
which are likely to cooperate or influence the activity of ERG in
PCa. The functional impact of multiple ETS factors associating with
ERG in PCa should be investigated in further studies as it may
provide insights into the mechanism in which ERG exerts its
influence in PCa, and subsequently contribute to our understanding
of the molecular basis of PCa.
Acknowledgements
The present study was supported by a
grant from the German Research Foundation [Deutsche
Forschungsgemeinschaft (DFG)] (WE1104/11-1) and the German Cancer
Aid (Deutsche Krebshilfe, 107827) to N.W. and by a grant from the
German Research Foundation [Deutsche Forschungsgemeinschaft (DFG)],
Emmy-Noether-Program, (PE1179/2-1), the Rudolf-Becker-Foundation
and the Wilhelm-Sander-Foundation (2011.077.1) to S.P.
References
|
1.
|
A JemalR SiegelJ XuE WardCancer
statistics, 2010CA Cancer J Clin60277300201010.3322/caac.20073
|
|
2.
|
SA TomlinsDR RhodesS PernerRecurrent
fusion of TMPRSS2 and ETS transcription factor genes in prostate
cancerScience310644648200510.1126/science.111767916254181
|
|
3.
|
MA RubinCA MaherAM ChinnaiyanCommon gene
rearrangements in prostate cancerJ Clin
Oncol2936593668201110.1200/JCO.2011.35.191621859993
|
|
4.
|
C Kumar-SinhaSA TomlinsAM
ChinnaiyanRecurrent gene fusions in prostate cancerNat Rev
Cancer8497511200810.1038/nrc240218563191
|
|
5.
|
PC HollenhorstDA JonesBJ GravesExpression
profiles frame the promoter specificity dilemma of the ETS family
of transcription factorsNucleic Acids
Res3256935702200410.1093/nar/gkh90615498926
|
|
6.
|
T OikawaETS transcription factors:
possible targets for cancer therapyCancer
Sci95626633200410.1111/j.1349-7006.2004.tb03320.x15298723
|
|
7.
|
N HashiyaN JoM AokiIn vivo evidence of
angiogenesis induced by transcription factor Ets-1: Ets-1 is
located upstream of angiogenesis
cascadeCirculation10930353041200410.1161/01.CIR.0000130643.41587.DB15173033
|
|
8.
|
T RothhammerJC HahneA FlorinThe Ets-1
transcription factor is involved in the development and invasion of
malignant melanomaCell Mol Life
Sci61118128200410.1007/s00018-003-3337-814704859
|
|
9.
|
JC HahneAF OkuducuA KaminskiA FlorinF
SoncinN WernertEts-1 expression promotes epithelial cell
transformation by inducing migration, invasion and
anchorage-independent
growthOncogene2453845388200510.1038/sj.onc.120876115940256
|
|
10.
|
A SahinC VercamerA
KaminskiDominant-negative inhibition of Ets 1 suppresses tumor
growth, invasion and migration in rat C6 glioma cells and reveals
differentially expressed Ets 1 target genesInt J
Oncol343773892009
|
|
11.
|
Z ShaikhibrahimB LangerA LindstrotEts-1 is
implicated in the regulation of androgen co-regulator FHL2 and
reveals specificity for migration, but not invasion, of PC3
prostate cancer cellsOncol Rep2511251129201121258770
|
|
12.
|
Z ShaikhibrahimA LindstrotB LangerR
BuettnerN WernertComprehensive gene expression microarray analysis
of Ets-1 blockade in PC3 prostate cancer cells and correlations
with prostate cancer tissues: Insights into genes involved in the
metastatic cascadeInt J Mol Med278118192011
|
|
13.
|
Z ShaikhibrahimN WernertETS transcription
factors and prostate cancer: The role of the family prototype ETS-1
(Review)Int J Oncol4017481754201222366814
|
|
14.
|
J DittmerThe biology of the Ets1
proto-oncogeneMol Cancer229200310.1186/1476-4598-2-2912971829
|
|
15.
|
B WasylykSL HahnA GiovaneThe Ets family of
transcription factorsEur J
Biochem211718199310.1111/j.1432-1033.1993.tb19864.x8425553
|
|
16.
|
PC HollenhorstAA ShahC HopkinsBJ
GravesGenome-wide analyses reveal properties of redundant and
specific promoter occupancy within the ETS gene familyGenes
Dev2118821894200710.1101/gad.156170717652178
|
|
17.
|
G WeiR SrinivasanCZ Cantemir-StoneEts1 and
Ets2 are required for endothelial cell survival during embryonic
angiogenesisBlood11411231130200910.1182/blood-2009-03-21139119411629
|
|
18.
|
DK WatsonDP TurnerMN ScheiberVJ FindlayPM
WatsonETS transcription factor expression and conversion during
prostate and breast cancer progressionOpen Cancer
J32439201010.2174/1874079001003010024
|
|
19.
|
GH WeiG BadisMF BergerGenome-wide analysis
of ETS-family DNA-binding in vitro and in vivoEMBO
J2921472160201010.1038/emboj.2010.10620517297
|
|
20.
|
SA TomlinsA BjartellAM ChinnaiyanETS gene
fusions in prostate cancer: from discovery to daily clinical
practiceEur
Urol56275286200910.1016/j.eururo.2009.04.03619409690
|
|
21.
|
G PetrovicsA LiuS ShaheduzzamanFrequent
overexpression of ETS-related gene-1 (ERG1) in prostate cancer
transcriptomeOncogene2438473852200510.1038/sj.onc.120851815750627
|
|
22.
|
K RostadM MannelqvistOJ HalvorsenERG
upregulation and related ETS transcription factors in prostate
cancerInt J Oncol301932200717143509
|
|
23.
|
N CerveiraFR RibeiroA PeixotoTMPRSS2-ERG
gene fusion causing ERG overexpression precedes chromosome copy
number changes in prostate carcinomas and paired HGPIN
lesionsNeoplasia8826832200610.1593/neo.0642717032499
|
|
24.
|
C SunA DobiA MohamedTMPRSS2-ERG fusion, a
common genomic alteration in prostate cancer activates C-MYC and
abrogates prostate epithelial
differentiationOncogene2753485353200810.1038/onc.2008.18318542058
|
|
25.
|
G AlipovT NakayamaM ItoOverexpression of
Ets-1 proto-oncogene in latent and clinical prostatic
carcinomasHistopathology46202208200510.1111/j.1365-2559.2005.02059.x15693893
|
|
26.
|
JP BasuyauxE FerreiraD StehelinG
ButticeThe Ets transcription factors interact with each other and
with the c-Fos/c-Jun complex via distinct protein domains in a
DNA-dependent and -independent mannerJ Biol
Chem2722618826195199710.1074/jbc.272.42.261889334186
|
|
27.
|
MA SvenssonCJ LaFargueTY MacDonaldTesting
mutual exclusivity of ETS rearranged prostate cancerLab
Invest91404412201110.1038/labinvest.2010.17920975660
|
|
28.
|
Z ShaikhibrahimA LindstrotJ EllingerS
RogenhoferR BuettnerN WernertIdentification of immunity-related
genes in prostate cancer and potential role of the ETS family of
transcription factors in their regulationInt J Mol
Med28799807201121833467
|
|
29.
|
V EspinaJD WulfkuhleVS
CalvertLaser-capture microdissectionNat
Protoc1586603200610.1038/nprot.2006.85
|
|
30.
|
VJ SchebleM BraunR BeroukhimERG
rearrangement is specific to prostate cancer and does not occur in
any other common tumorMod
Pathol2310611067201010.1038/modpathol.2010.8720473283
|
|
31.
|
JP ClarkCS CooperETS gene fusions in
prostate cancerNat Rev
Urol6429439200910.1038/nrurol.2009.12719657377
|
|
32.
|
B HanR MehraK SulemanCharacterization of
ETS gene aberrations in select histologic variants of prostate
carcinomaMod
Pathol2211761185200910.1038/modpathol.2009.7919465903
|
|
33.
|
CC GuoJY DancerY WangTMPRSS2-ERG gene
fusion in small cell carcinoma of the prostateHum
Pathol421117201110.1016/j.humpath.2010.05.02621040948
|
|
34.
|
VJ SchebleM BraunT WilbertzERG
rearrangement in small cell prostatic and lung
cancerHistopathology56937943201010.1111/j.1365-2559.2010.03564.x20636794
|
|
35.
|
AM JoshuaA EvansT Van der KwastProstatic
preneoplasia and beyondBiochim Biophys
Acta1785156181200818166163
|
|
36.
|
M BraunVJ SchebleR MenonRelevance of
cohort design for studying the frequency of the ERG rearrangement
in prostate
cancerHistopathology5810281036201110.1111/j.1365-2559.2011.03862.x21707704
|
|
37.
|
VI SementchenkoCW SchweinfestTS PapasDK
WatsonETS2 function is required to maintain the transformed state
of human prostate cancer
cellsOncogene1728832888199810.1038/sj.onc.12022209879994
|
|
38.
|
GM CarboneS NapoliA ValentiniF CavalliDK
WatsonCV CatapanoTriplex DNA-mediated downregulation of Ets2
expression results in growth inhibition and apoptosis in human
prostate cancer cellsNucleic Acids
Res3243584367200410.1093/nar/gkh74415314206
|
|
39.
|
Z ShaikhibrahimM BraunP
NikolovRearrangement of the ETS genes ETV-1, ETV-4, ETV-5, and
ELK-4 is a clonal event during prostate cancer progressionHum
PatholMay72012(Epub ahead of print)
|
|
40.
|
PC HollenhorstL PaulMW FerrisBJ GravesThe
ETS gene ETV4 is required for anchorage-independent growth and a
cell proliferation gene expression program in PC3 prostate
cellsGenes Cancer110441052201110.1177/194760191039557821373373
|