Introduction
Lung cancer is the leading cause of cancer death
throughout the world, causing ∼1.2 million deaths annually and an
estimated >250,000 deaths in China during 2010 (1,2).
Lung cancer is clinically divided into two subtypes, small cell
lung cancer (SCLC) and non-small cell lung cancer (NSCLC). The
latter is the most common type, accounting for 85–90% of total
cases (3). Although recent
studies have shown that adjuvant chemotherapy improves survival in
completely resected NSCLC, only 5–15% of treated individuals
ultimately benefit (4).
Resistance of NSCLC to current treatment protocols still remains a
major concern in cancer therapy (5).
Tumor necrosis factor-related apoptosis-inducing
ligand (TRAIL) is a highly promising anticancer agent with
pronounced pro-apoptotic activity towards various malignant cell
types, including lung cancer. Importantly, TRAIL essentially lacks
activity towards normal cells (6). Based on these characteristics,
recombinant soluble TRAIL (sTRAIL) preparations have recently
entered clinical trials, with promising preliminary studies on
anti-tumor activity and safety (7). TRAIL engages its receptors via the
extrinsic apoptotic pathway and recruits caspase-8, which is then
cleaved to its active form. Activated caspase-8 then cleaves the
BH3-only molecule, Bid, which then interacts with mitochondrial
anti- and pro-apoptotic molecules. Previous studies have
demonstrated that TRAIL modulates the apoptotic response by binding
to receptors, including the death receptor 4 (DR4, TRAIL-R1)
(8), KILLER/DR5 (TRAIL-R2)
(9), TRID (DcR1, TRAIL-R3)
(10), TRUNDD (DcR2, TRAIL-R4),
and osteoprotegerin (OPG) (11).
Both DR4 and DR5 have a C-terminal death domain that signals
through downstream caspase activation. However, it is not yet fully
understood how TRAIL induces tumor-cell apoptosis. Hence, a great
deal of research has been focused on improving the potency of
TRAIL-based therapeutic approaches, some by establishing new
modalities for combined treatment with TRAIL and other agents.
Bcl-2 (B-cell lymphoma 2) is the founding member of the Bcl-2
family of apoptosis regulator proteins encoded by the Bcl-2
gene. Bcl-2 is highly expressed in many cell lines and
primary tumors including lung tumors and its expression has been
associated with drug resistance (12,13). Thus, novel strategies are
necessary to target resistance of tumors with Bcl-2
overexpression.
In this study, focusing on the lung cancer line
A549, we constructed a recombinant adenovirus Ad5.TRAIL/siBcl2,
with overexpression of TRAIL and downregulation of Bcl-2. We
investigated the therapeutic gene of Ad5.TRAIL/siBcl2 and explored
its antitumor potential in experimental lung tumors.
Materials and methods
Cell culture and reagents
The human embryonic kidney cell line HEK293, the
human lung tumor cell lines A549 and HCC827 as well as the normal
lung tissue cell line WI-38, were obtained from the American Type
Culture Collection (Rockville, MD) and cultured in Dulbecco’s
modified Eagle’s medium (DMEM) supplemented with 10% FBS and 100
IU/ml penicillin/streptomycin in a 37°C humidified incubator with
5% CO2. The recombinant adenovector™ system was
purchased from Stratagene (Carlsbad, CA) for the construction of
adenoviruses. The anti-TRAIL, the Bcl-2 and the HRP-conjugated
anti-mouse IgG antibodies were obtained from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA).
MTT assay
The cytotoxic activity of TRAIL protein was
determined based on cycotoxicity to the human lung tumor cell line
A549, using an MTT assay. Briefly, cells were seeded in 96-well
tissue culture plates at a density of 5×103 cells/well
and then treated at the concentration of 0, 1, 10 and 100 ng/ml
TRAIL protein in the medium. The following day, the medium was
removed and 100 μl of fresh medium containing 0.5 mg/ml MTT (Roche
Diagnostics GmbH; Mannheim, Germany) was added to each well. The
cells were incubated at 37°C in a humidified atmosphere with 5%
CO2 for 4 h, followed by the addition of 150 μl of
solubilization solution [0.01 mol/l HCl in 100 g/l sodium dodecyl
(SDS)] to each well and the incubation of cells for further 10 min
at 37°C with gentle shaking. The optical density of the plates was
measured using the spectrophotometrical absorbance at 570 nm in the
Microplate Reader Model 550 (Bio-Rad, Hercules, CA).
Viability and caspase-3/7 assays
For the Apo-One® Homogeneous caspase-3/7
assay (Promega Corporation, Madison, WI), cells were seeded into
96-well plates at 2×104 cells/well, incubated overnight
and subsequently treated with TRAIL (Alexis Corporation, Lausanne,
Switzerland). The substrate for the caspase assay was added after
0–24 h of treatment (as indicated in the text) and plates were read
2 h later using a Tecan Plate Reader (Tecan, Group. Ltd.,
Switzerland). All treatments were done in triplicate. Background
absorbance was determined by incubating media with substrate alone
and subtracting the values from wells containing cells.
Preparation of cell extracts and western
blot analysis
Protein lysates were prepared on ice in RIPA buffer
[50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 0.1% SDS, 1% NP40 and 0.5%
sodium deoxycholate] with freshly added 0.1 mg/ml
phenylmethylsulfonyl fluoride, 1 mm sodium orthovanadate and 1
mg/ml aprotinin. Protein concentrations were determined using the
Bio-Rad protein assay system (Bio-Rad Laboratories, Inc., Richmond,
CA). Aliquots of cell extracts containing 20–50 mg of total protein
were resolved in 12% SDS-PAGE and transferred to 0.45 mm
nitrocellulose membranes (Osmonics, Westborough, MA). Filters were
blocked for 1 h at room temperature (RT) in Blotto A [5% nonfat
milk powder in TBS-T: 10 mm Tris-HCl (pH 8.0), 150 mm NaCl, 0.05%
Tween-20], and then incubated for 1 h at room temperature in Blotto
A containing a 1:1,000 dilution of rabbit anti-TRAIL, anti-Bcl-2
and anti-GAPDH monoclonal antibodies. After washing in TBS-T buffer
(5 min at RT), filters were incubated for 1 h at room temperature
in Blotto A containing a 1:10,000 dilution of peroxidase conjugated
anti-rabbit or anti-mouse secondary antibody (Amersham Life
Science, Arlington Heights, IL). After washing in TBS-T, ECL was
performed according to the manufacturer’s instructions. All results
were normalized to GAPDH protein expression.
Construction of gene silencing and
adenoviral vectors
Plasmid pGenesil-1 (Wuhan Genesil Biotechnology Co.,
Ltd., Wuhan, China) was used to express small hairpin RNA (shRNA)
targeting human Bcl-2. The following hairpin inserts were used,
Bcl2-1F, 5′-GATCCGGTACGATAACCGGGAGATA
GTTTCAAGACGACTATCTCCCGGTTATCGTACTTTTT TA-3′ and Bcl2-1R,
5′-AGCTTAAAAAAGTACGATAAC CGGGAGATAGTCGTCTTGAAACTATCTCCCGGTTATC
GTAC-3′; Bcl2-2F, 5′-GATCCGGGAGGATTGTGGCCTTC
TTTTTCAAGACGAAAGAAGGCCACAATCCTCCTTTT TTA-3′ and Bcl2-2R:
5′-AGCTTAAAAAAGGAGGATTG TGGCCTTCTTTCGTCTTGAAAAAGAAGGCCACAATC
CTCCCG-3′; shRNA control, Con-F, 5′-GATCCGCCACTTG
GACCAGTATTATTTCAAGACGATAATACTGGTCCAAG TGGTTTTTTA-3′ and Con-R,
5′-AGCTTAAAAAACCACT TGGACCAGTATTATCGTCTTGAAATAATACTGGTCCA
AGTGGCG-3′. The insert was cloned into BamHI and
HindIII site, which is downstream of the human U6 promoter
(hU6), an RNA polymerase III promoter. All the constructions were
confirmed by DNA sequencing. The correct plasmid, named
pGenesil-Bcl2-1, pGenesil-Bcl2-2 and pGenesil-con, was used for
screening the silence genes.
The sense primer (5′-GAAGATCTGTGAGAGAAAGAGG
TCCTCAGAGA-3′) and the antisense primer (5′-GGGGTA CCTTAGCCAACTA
AAAAGGCCCCGA-3′) were used for cloning the human TRAIL gene
directly from plate purchased from Sino Biological, Inc. (Beijing,
China) by using the polymerase chain reaction (PCR). The cloned
TRAIL cDNA fragment was ligated into the pShuttle-CMV vector to
form pShuttle-CMV/TRAIL for expressing TRAIL. The
pShuttle-CMV/TRAIL, which was cut by SalI and
HindIII, was ligased with the fragment hU6-Bcl2-2 from
pGenesil-Bcl2-2. The recombinant pAdeasy-1 backbone vector and
pShuttle-CMV/TRAIL, pShuttle-CMV/siBcl2 or
pShuttle-CMV/TRAIL/siBcl2 vector, DNA linealized with PmeI
digestion, were further co-transfected into the bacteria BJ5183
cells. The resultant pAd5.TRAIL, pAd5.siBcl2 and pAd5.TRAIL/siBcl2
plasmid vectors were purified from the above transfected BJ5183
cells, then linealized by PacI digestion and then
transfected into the HEK293 cells by lipofectamine 2000 (Invitrogen
Life Technologies, Carlsbad, CA), leading to the formation of the
recombinant adenoviruses Ad5.siBcl2, Ad5.TRAIL and Ad5.
TRAIL/siBcl2. The adenoviruses were amplified in HEK293 cells,
purified by cesium chloride ultracentrifugation and stored at −80°C
before use. Titers were determined by plaque assay on HEK293
cells.
Apoptotic cell assay
Cells seeded in 96-well tissue culture plates were
treated with PBS, Ad5.eGFP, Ad5.siBcl2, Ad5.TRAIL or
Ad5.TRAIL/siBcl2. After 48 h of treatment, cells were incubated
with MTT and followed by the MTT assay method.
Tumor studies
Athymic nude and BalB/C mice were obtained from the
Shanghai Experimental Animal Center (Shanghai, China) and
maintained according to the Animal Research Committee’s guidelines
at Qingdao University (Qingdao, China). Briefly, a total of
8×106 A549 cells were injected into the left dorsal
flank of mice (10 animals for each treatment) and then monitored
every other day for tumor growth. When tumors reached 40–50
mm3, animals were randomly divided and received
Ad5.eGFP, Ad5.siBcl2, Ad5.TRAIL or Ad5. TRAIL/siBcl2 treatment. All
treatments were given every other day for a total of 5 doses. Tumor
growth was monitored by measuring perpendicular tumor diameters
using an electronic digital caliper. Tumor volumes were calculated
by the formula: tumor size = ab2/2, where a is the
largest and b is the smallest of the 2 dimensions. The
tumor-bearing mice were sacrificed 10 days after injection with
different viruses and the tumors were removed, weighed and examined
by TUNEL assay.
TUNEL assay
In situ cell apoptosis of tumor sections
taken on Day 10 post-treatment was analyzed by using a commercially
available TUNEL kit (Promega Corporation). Briefly, tumor sections
were fixed with a freshly prepared paraformaldehyde solution (4% in
PBS, pH 7.4). After 30 min of incubation at room temperature, cells
were washed with PBS and resuspended in a permeabilization solution
(0.1% Triton X-100 in 0.1% sodium citrate) for 2 min on ice. After
an additional wash with PBS, cells were labeled using the TUNEL kit
according to the manufacturer’s instructions. The apoptotic index
was calculated by the formula: apoptotic index = (total number of
apoptotic cells/total number of cells) x 100%.
Statistical analysis
Student’s t-test and One-way ANOVA analysis were
performed for continuous variables. The χ2 test or
Fisher’s exact test were used for categorical variables. The error
bars represent the standard error of the mean. Statistical
significance for all the tests, assessed by calculating P-value,
was <0.05 from two-sided tests. The statistical analyses were
performed using SAS 9.0 software (SAS Institute, Inc., Cary,
NC).
Results
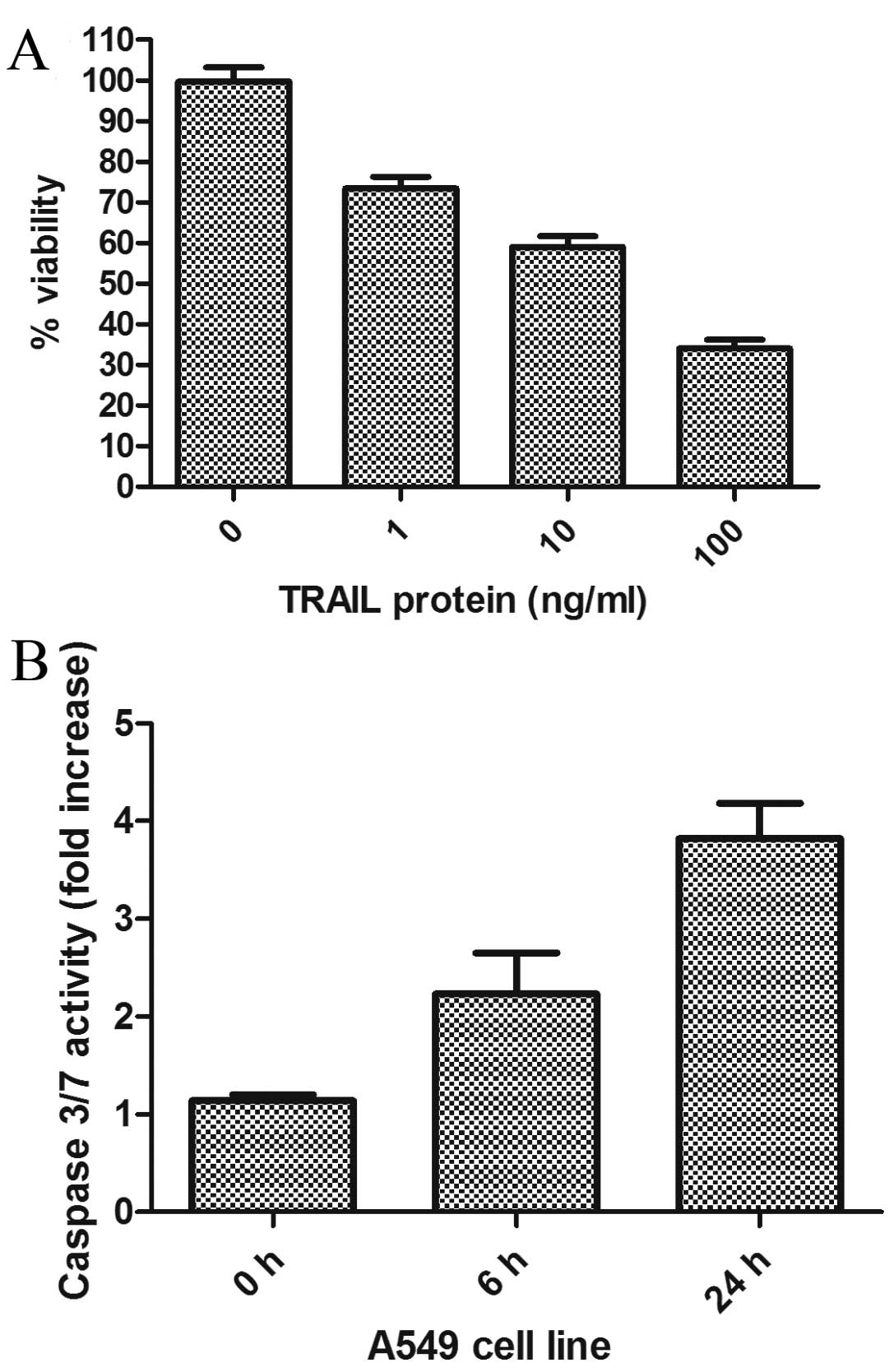
The susceptibility of A549 lung cancer
cell lines to TRAIL protein
The goal of this study was to determine the
sensitivity of lung cancer cells to TRAIL protein treatment. A549,
the common NSCLC cell line, was used for this purpose. As shown in
Fig. 1A, exposure of A549 cells
to increasing concentrations of recombinant TRAIL protein resulted
in a dose-dependent decrease in viability. To confirm that the
response to TRAIL was apoptotic, we performed caspase-3/7 activity
assays (Fig. 1B). In A549 cells,
a 2- to 3-fold increase in caspase activity was measured at 6 and
24 h respectively, following exposure to TRAIL protein. This level
of caspase activation may be sufficient to cause cell death.
However, there was some resistance to the TRAIL induced apoptosis.
TRAIL resistance in the cancer cells was due to differences in
expression of various proteins of the apoptotic pathway, including
TRAIL receptors, cFLIP, Bax, Bcl-2, caspase-3 or X-linked mammalian
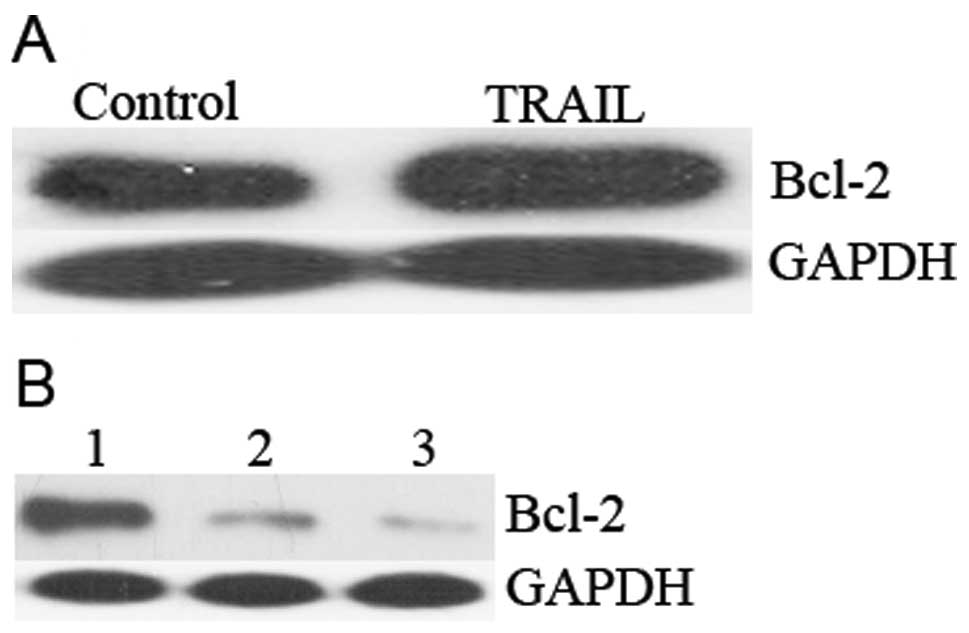
inhibitor of apoptosis protein (14). To explore whether Bcl-2 presents a
main resistance point in A549 cells, we analyzed Bcl-2 expression
with or without TRAIL treated cells. As presented in Fig. 2A, enhanced Bcl-2 expression was
observed in TRAIL treatment cells on the concentration of 100
ng/ml. Because Bcl-2 has been implicated in apoptotic regulation in
other organisms, we determined whether Bcl-2 deficiency affected
the in vitro apoptosis of mammalian cells using ‘knockdown’
(KD) technology. Two independent small hairpin RNA (shRNA) specific
for human Bcl-2 (siBcl2-1 and siBcl2-2) and a control (siCNT) were
introduced into the A549 cell line to verify their efficiency.
After 48 h, the level of Bcl-2 protein was reduced to <10% of
the control in A549 cells transfected with siBcl2-1 or siBcl2-2,
whereas introduction of siCNT had no effect on Bcl-2 protein levels
(Fig. 2B). To test the apoptotic
efficiency, siBcl2-2 was chosen for the downstream experiment in
vitro and in vivo.
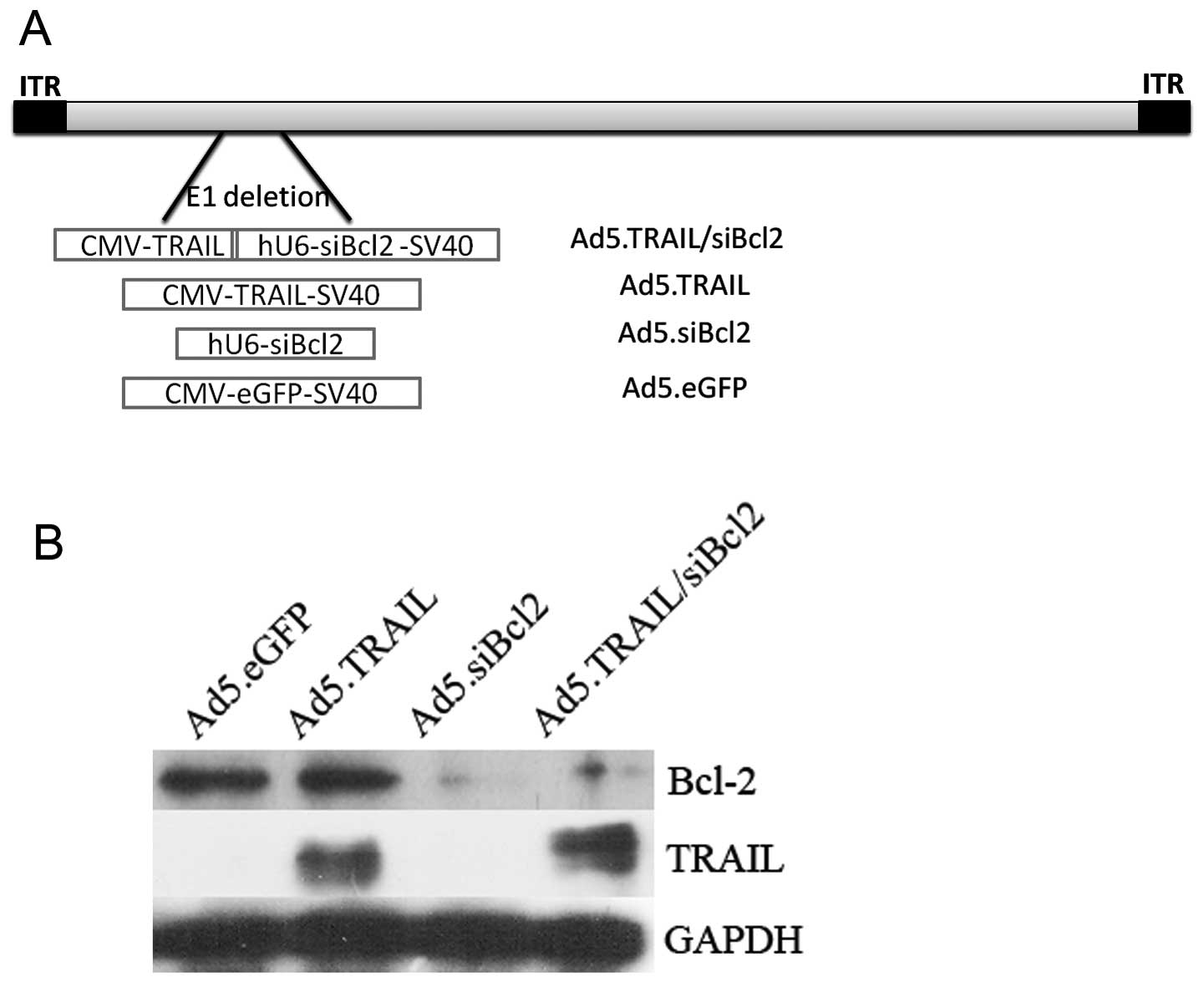
Characterization of adenoviral vectors
for co-expression of shRNA and transgene in vitro
To achieve maximum efficiency of Bcl-2 silencing and
TRAIL expression in lung cancer cells, adenoviral vectors
(Ad5.eGFP, Ad5.shBcl2, Ad5.TRAIL and Ad5.TRAIL/siBcl2) were
generated for co-expressing shRNA and TRAIL (Fig. 3A). To compare the efficiency of
shRNA-mediated silencing of Bcl-2 and expression of TRAIL, we
infected A549 cells with 4 adenoviral constructs and the expression
of Bcl-2 and TRAIL was measured. As shown in Fig. 3B, the level of Bcl-2 expression
was comparable in the cells infected with Ad5.eGFP, Ad5.TRAIL and
cells without infection. In contrast, infection with Ad5.siBcl2 and
Ad5.TRAIL/siBcl2 resulted in significant reduction of Bcl-2 gene
expression. The expression of Bcl-2 was almost undetectable in
cells infected with Ad5.siBcl2 and Ad5.TRAIL/siBcl2 (Fig. 3B). The high and comparable level
of TRAIL expression was only observed in the cells infected with
Ad5.TRAIL and Ad5.TRAIL/siBcl2. This indicated that
Ad5.TRAIL/siBcl2 could induce efficient silencing of Bcl-2 and
expression of TRAIL.
Ad5.TRAIL/siBcl2 induces tumor-cell
cytotoxicity in vitro
To evaluate whether Ad5.TRAIL/siBcl2 could impede
lung cancer cell proliferation, we performed cell viability assay
in the lung cancer cells after infection with adenoviral vectors.
As shown in Fig. 4, both
Ad5.TRAIL and Ad5.siBcl2 could reduce cell viability in lung cancer
cells. However, there was significant reduction in cell viability
in tumor cells infected with Ad5. TRAIL/siBcl2, comparing to cells
infected with Ad5.TRAIL and Ad5.shBcl2. These results suggested
that Bcl-2 silencing sensitized lung cancer cells to TRAIL. In
contrast, Ad5.siBcl2, Ad5.TRAIL or Ad5.TRAIL/siBcl2 exhibited low
cytotoxicity on normal lung cells WI-38. These data indicated that
Ad5. TRAIL/siBcl2 had enhanced antitumor activity in lung tumor
cells and maintained low toxicity to normal lung cells.
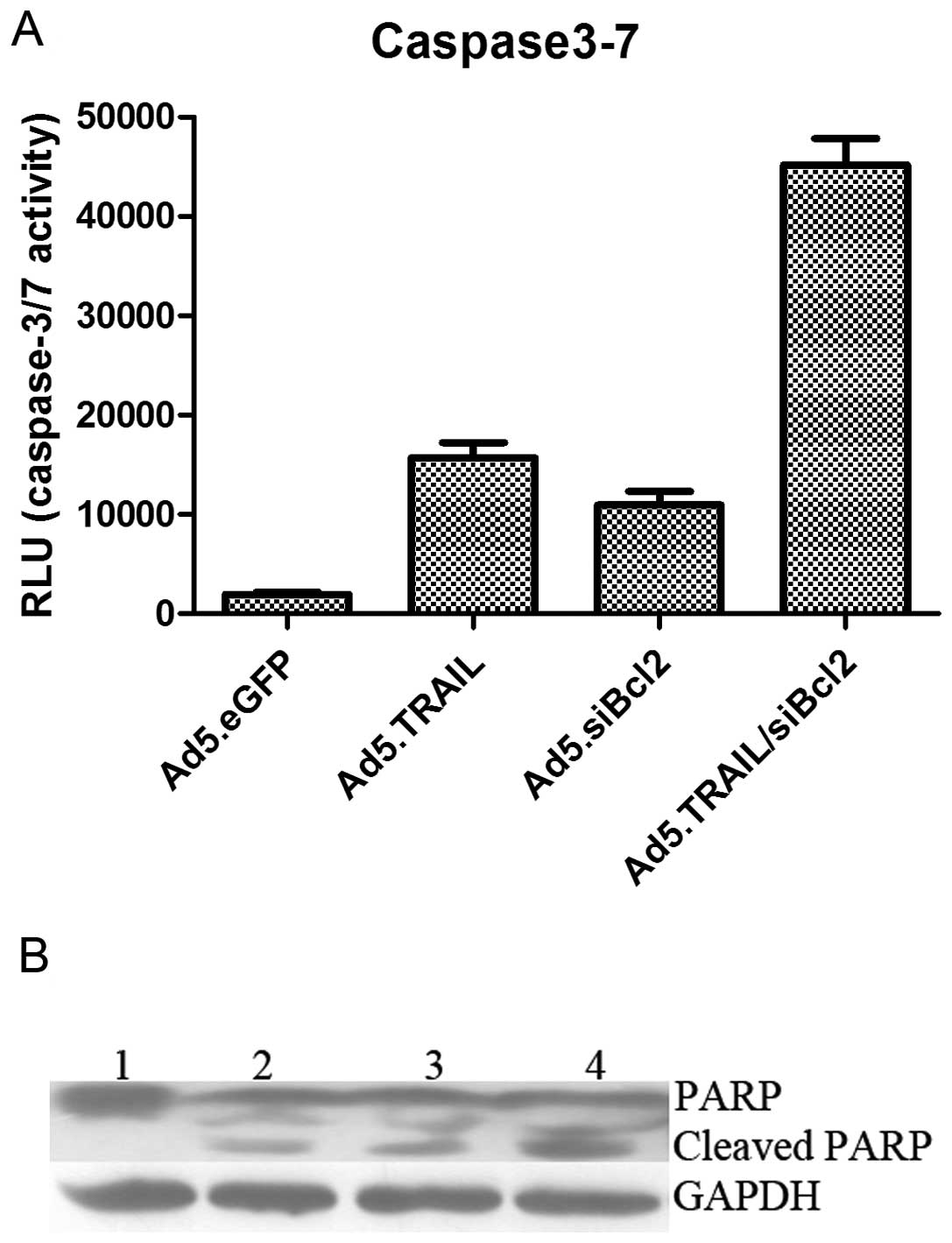
To investigate the mechanism of apoptosis, we
analyzed A549 cells infected with Ad5.eGFP, Ad5.TRAIL, Ad5.shBcl2
or Ad5.TRAIL/siBcl2 for 48 h by caspase-3/7 activity assay and
western blot analysis. Caspase-3/7 activity was increased in the
presence of Ad5.TRAIL/siBcl2 infection, indicating that TRAIL plus
siBcl-2 could sensitize NSCLC cells to the cytotoxic actions
(Fig. 5A). The analysis of
infected A549 cancer cells demonstrated the activation of
poly(ADP-ribose) polymerase (PARP) after infection with different
adenoviruses. The cleaved forms of PARP increased and the uncleaved
form of PARP decreased, which indicated the activation of caspase
cascade after treatment with adenoviruses. The most obvious effect
of caspase activation was observed in the cells treated with
Ad5.TRAIL/siBcl2 (Fig. 5B). These
results suggest that the combined use of TRAIL and siBcl-2 can
induce the caspase cascade activation in a synergistic manner.
Ad5.TRAIL/siBcl2 suppresses tumor growth
in vivo
To explore the in vivo antitumor effect of
adenoviruses, the athymic nude mice (10 mice/group), bearing A549
tumors (0.2–0.3 cm3) were intrathecally (i.t.) injected
with Ad5.eGFP, Ad5.siBcl2, Ad5. TRAIL and Ad5.TRAIL/siBcl2
(1x107 pfu/50 μl). The tumor growth was monitored daily.
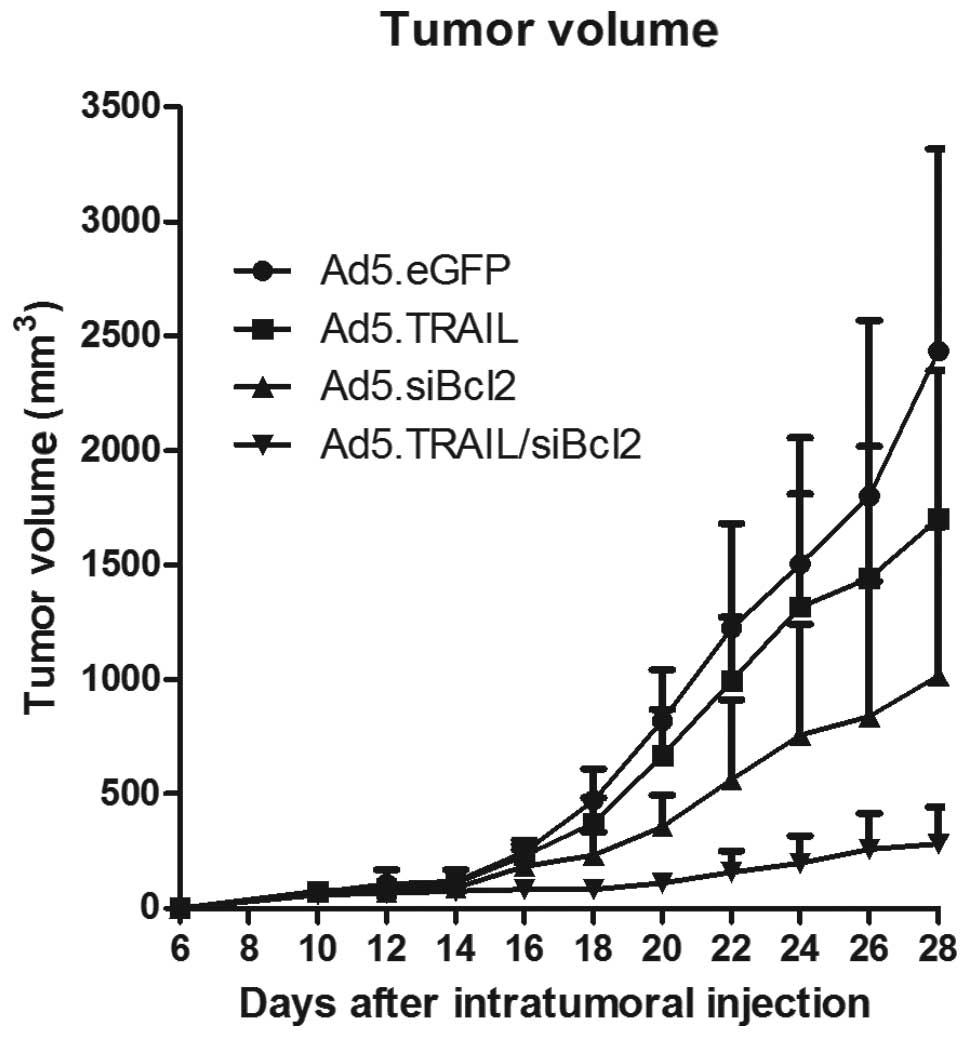
As shown in Fig. 6, the tumor
growth was significantly decreased in the groups of mice treated
with Ad5.TRAIL/siBcl2, compared with the tumor growth of mice
treated with Ad5.GFP (P<0.01), indicating that Ad5.TRAIL/siBcl2
can induce efficient tumor growth suppression in vivo. To
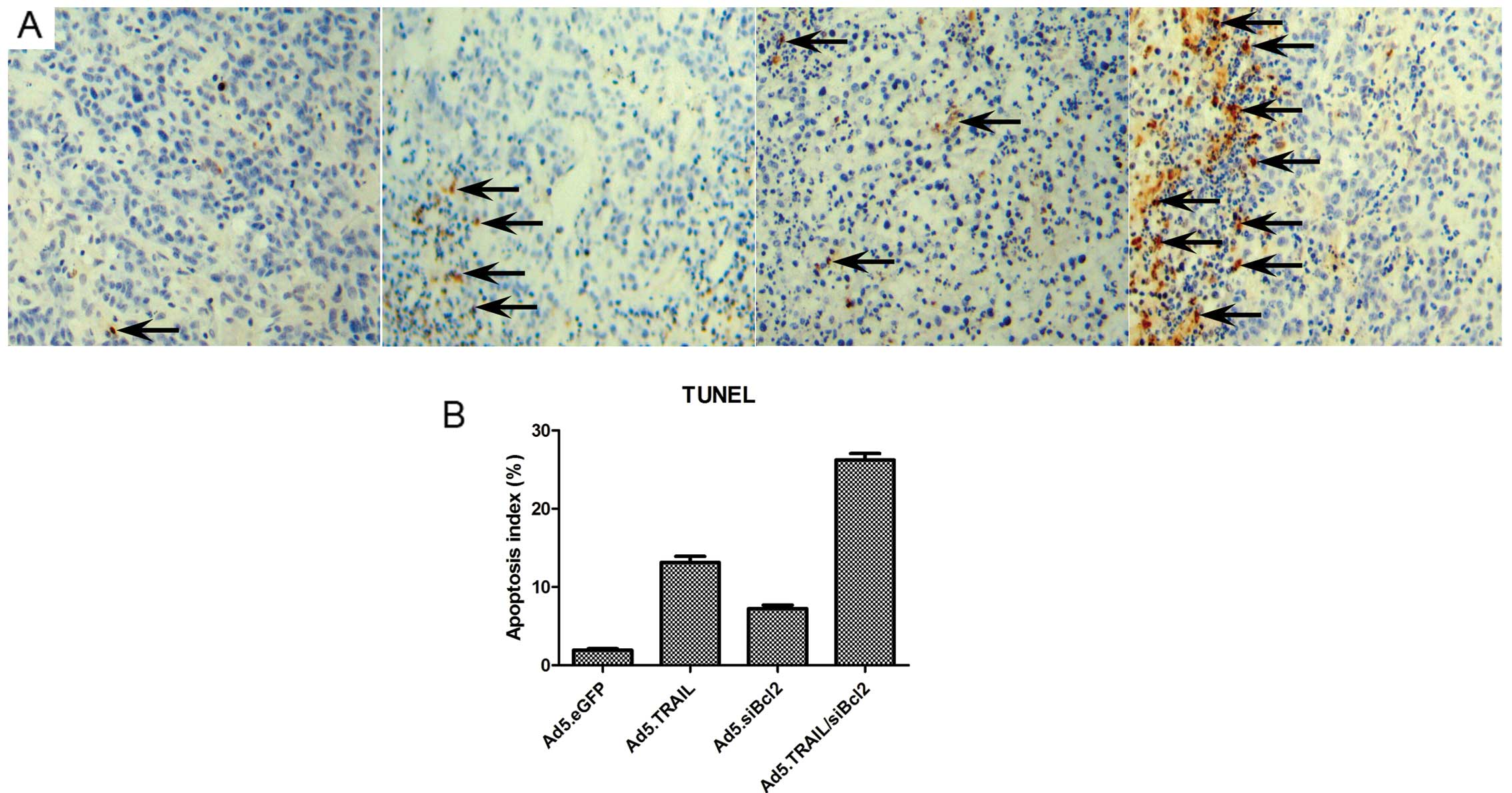
understand the mechanism of tumor inhibition, tumor sections were
analyzed for apoptosis by TUNEL assay. As shown in Fig. 7, tumors from the mice injected
with Ad5. TRAIL/siBcl2 demonstrated extensive apoptosis
26.24±2.62%, compared with the group treated with Ad5.eGFP,
Ad5.siBcl2 or Ad5.TRAIL. The apoptotic index, expressed as the
average percentages of TUNEL-positive cells from 10 random visual
fields, were 1.94±0.53, 7.25±1.42 and 13.15±2.38% in Ad5. eGFP,
Ad5.siBcl2 and Ad5.TRAIL, respectively.
Discussion
Despite the aggressive treatment regimens, patients
with diagnosis of NSCLC still have a poor prognosis (15). Current attempts to improve the
survival of these patients largely depend on novel strategies
designed to overcome the intrinsic resistance of NSCLC to the
established therapies (16,17). The idea to directly trigger
apoptosis in cancer cells by stimulating death receptors at the
cell surface has gained considerable attention, as these receptors
have a direct link to the cell’s intrinsic death machinery
(18). TRAIL, a member of the TNF
family of death-inducing ligands, is a prime candidate for cancer
therapy because it almost selectively induces apoptosis in a wide
spectrum of cancers with no or minimal toxicity on normal human
cells. Agents triggering TRAIL receptors, such as agonistic TRAIL
receptor antibodies or recombinant TRAIL, have been extensively
studied in preclinical models and are currently under investigation
in clinical trials. However, the potential application of TRAIL for
cancer therapy is resistant to the agents. Therefore, further
insights into the molecular determinants of TRAIL-induced apoptosis
are pivotal to improve outcome of TRAIL treatment in NSCLC
(19).
RNAi is a process of sequence-specific
post-transcriptional gene silencing mediated by double-stranded
RNA. Compared with conventional single-stranded antisense
oligonucleotides technology, RNAi is more efficient at silencing
the corresponding gene expression. At present, the main obstacle to
the development of therapeutic agents using RNAi technology is a
suitable delivery method. Although non-viral-based siRNA vectors
are regarded as the safest delivery systems, they still constitute
an essential challenge in therapeutics, limited by their poor
ability to escape from the endosomal compartment and to translocate
DNA into the nucleus. Another important consideration for
siRNA-mediated inhibition of gene expression is whether the
observed effects are free from potential IFN responses, as lipid
delivery of synthetic siRNAs can reportedly induce immune
activation in vivo (20).
In contrast, viral vectors have been used to deliver siRNA widely
and these methods tend to exhibit several advantages over nonviral
vectors, such as higher transduction efficacy and more stable gene
silencing. The nonreplicative viral vectors, used for siRNA
delivery, can be applied to overcome these limitations (21).
In aggressive therapies, resistance to current
treatment protocols constitutes a major obstacle in clinical
oncology. Most anticancer agents act through induction of apoptosis
in target cells and defects in apoptosis programs may confer
resistance (22). Apoptosis is
mediated through both the death receptor and the mitochondrial
pathway. At least in some cellular systems, mitochondria also seem
to be necessary for death receptor-induced apoptosis. The
mitochondrial pathway of apoptosis can be blocked by the
anti-apoptotic protein Bcl-2 which has been localized to cellular
membranes, e.g. the mitochondrial membrane. The Bcl-2 gene is the
prototype of a class of genes that contributes to neoplastic
progression by enhancing tumor cell survival through the inhibition
of apoptosis (23). Bcl-2 levels
are elevated in follicular lymphoma, melanomas, prostate and
gliomas suggesting that a decrease in Bcl-2 might sensitize cells
to apoptotic stimuli (24). Bcl-2
is highly expressed in many cell lines and primary tumors including
neuroblastoma or glioblastoma. High Bcl-2 expression has been
correlated with poor prognosis in some cases and it is also likely
to play a role in chemoresistance. Thus, novel strategies are
needed to target resistance, e.g. due to Bcl-2 overexpression. As
hypothesized, downregulation of Bcl-2 by siRNA sensitized NSCLC
cells to Apo2L/TRAIL-induced apoptosis (Fig. 4). Activation of the caspase
cascade was blocked at the level of complete activation of effector
caspases such as caspase-3/7 in NSCLC cell lines poorly responding
to TRAIL. TRAIL stimulated cleavage of PARP following
downregulation of Bcl-2 in A549 cells (Fig. 5B). Thus, downregulation of Bcl-2
may activate the mitochondrial arm (intrinsic pathway) to amplify
the effects of death receptor-mediated apoptosis.
To improve the therapeutic efficiency, we combined
this virotherapeutic agent with RNAi technology and conventional
gene therapy method. In this study, as proposed,
adenovirus-mediated siRNA can efficiently silence the expression of
Bcl-2 in vitro, leading to direct induction of cell death in
NSCLC cell line and increase in the sensitivity of both NSCLC cell
lines to Ad5.TRAIL mediated apoptosis, but with no apparent impact
on normal cells (WI-38). Bcl-2 may regulate caspase activation
through sequestration of unidentified caspase adaptors/activators.
Moreover, Bcl-2 could directly interact as substrate with different
subsets of caspases (25). These
factors might be important for A549 cells where Bcl-2 was
overexpressed providing protection against Ad.TRAIL-mediated
apoptosis, indicating that the caspase machinery activated by TRAIL
in A549 cells might be preferentially inhibited by Bcl-2. To
further prove the synergistic efficacy, our in vivo data
showed that Ad5.shBcl-2 could promote antitumor activity of Ad5.
TRAIL in NSCLC xenograft tumors. These phenomena potentially
suggest that adenoviral vectors are effectively infected the NSCLC
for long-term siRNA delivery.
In conclusion, our adenovirus vector has both a cell
cytotoxicity ability and siRNA delivery capability. Ad5.shBcl2
could knockdown Bcl-2 expression and enhance the antitumor activity
of Ad5.TRAIL both in vitro and in vivo. Our findings
may have important implications in the development of new
strategies in cancer therapy.
Acknowledgements
This study was carried out as part of
a project financed by the grant of Shandong Tackle Key Problems in
Science and Technology (2010GSF10245).
References
|
1.
|
A JemalR SiegelJ XuE WardCancer
statistics, 2010CA Cancer J Clin60277300201010.3322/caac.20073
|
|
2.
|
L HouwenState of the art: lung cancer in
ChinaAnn Thorac Cardiovasc Surg9147148200312882182
|
|
3.
|
DM vanHE Karim-KosS SieslingProgress in
standard of care therapy and modest survival benefits in the
treatment of non-small cell lung cancer patients in the Netherlands
in the last 20 yearsJ Thorac Oncol7291298201222157366
|
|
4.
|
MA BareschinoC SchettinoA RossiTreatment
of advanced non small cell lung cancerJ Thorac
Dis3122133201122263075
|
|
5.
|
BS NairV BhanderiSH JafriCurrent and
emerging pharmacotherapies for the treatment of relapsed small cell
lung cancerClin Med Insights Oncol5223234201121836818
|
|
6.
|
A AshkenaziRC PaiS FongSafety and
antitumor activity of recombinant soluble Apo2 ligandJ Clin
Invest104155162199910.1172/JCI692610411544
|
|
7.
|
JC SoriaE SmitD KhayatPhase 1b study of
dulanermin (recombinant human Apo2L/TRAIL) in combination with
paclitaxel, carboplatin, and bevacizumab in patients with advanced
non-squamous non-small-cell lung cancerJ Clin
Oncol2815271533201010.1200/JCO.2009.25.484720159815
|
|
8.
|
G PanRK OAM ChinnaiyanThe receptor for the
cytotoxic ligand
TRAILScience276111113199710.1126/science.276.5309.1119082980
|
|
9.
|
H WalczakMA Degli-EspostiRS
JohnsonTRAIL-R2: a novel apoptosis-mediating receptor for TRAILEMBO
J1653865397199710.1093/emboj/16.17.53869311998
|
|
10.
|
SA MarstersJP SheridanRM PittiA novel
receptor for Apo2L/TRAIL contains a truncated death domainCurr
Biol710031006199710.1016/S0960-9822(06)00422-29382840
|
|
11.
|
JG EmeryP McDonnellMB BurkeOsteoprotegerin
is a receptor for the cytotoxic ligand TRAILJ Biol
Chem2731436314367199810.1074/jbc.273.23.143639603945
|
|
12.
|
Y ZhangJH WangQ LuYJ WangBag3 promotes
resistance to apoptosis through Bcl-2 family members in non-small
cell lung cancerOncol Rep1109113201121971866
|
|
13.
|
G KroemerThe proto-oncogene Bcl-2 and its
role in regulating apoptosisNat
Med3614620199710.1038/nm0697-6149176486
|
|
14.
|
TS GriffithDH LynchTRAIL: a molecule with
multiple receptors and control mechanismsCurr Opin
Immunol10559563199810.1016/S0952-7915(98)80224-09794836
|
|
15.
|
LL CarrJH FiniganJA KernEvaluation and
treatment of patients with non-small cell lung cancerMed Clin North
Am9510411054201110.1016/j.mcna.2011.08.00122032426
|
|
16.
|
BD DalyRJ CerfolioMJ KrasnaRole of surgery
following induction therapy for stage III non-small cell lung
cancerSurg Oncol Clin N
Am20721732201110.1016/j.soc.2011.07.00621986268
|
|
17.
|
MH ColemanR BuenoRole of adjuvant
chemotherapy in NSCLC (stages I to III)Surg Oncol Clin N
Am20757767201110.1016/j.soc.2011.07.01121986270
|
|
18.
|
A AshkenaziTargeting death and decoy
receptors of the tumour-necrosis factor superfamilyNat Rev
Cancer2420430200210.1038/nrc82112189384
|
|
19.
|
EE McGrathThe tumor necrosis
factor-related apoptosis-inducing ligand and lung cancer: still
following the right TRAIL?J Thorac
Oncol6983987201110.1097/JTO.0b013e318217b6c821623278
|
|
20.
|
AJ BridgeS PebernardA DucrauxAL NicoulazR
IggoInduction of an interferon response by RNAi vectors in
mammalian cellsNat Genet34263264200310.1038/ng117312796781
|
|
21.
|
JE CaretteRM OvermeerFH
SchagenConditionally replicating adenoviruses expressing short
hairpin RNAs silence the expression of a target gene in cancer
cellsCancer
Res6426632667200410.1158/0008-5472.CAN-03-353015087375
|
|
22.
|
SH KaufmannWC EarnshawInduction of
apoptosis by cancer chemotherapyExp Cell
Res2564249200010.1006/excr.2000.483810739650
|
|
23.
|
A GrossJM McDonnellSJ KorsmeyerBcl-2
family members and the mitochondria in apoptosisGenes
Dev1318991911199910.1101/gad.13.15.189910444588
|
|
24.
|
B JansenU Zangemeister-WittkeAntisense
therapy for cancer - the time of truthLancet
Oncol3672683200210.1016/S1470-2045(02)00903-812424069
|
|
25.
|
S CoryJM AdamsThe Bcl-2 family: regulators
of the cellular life-or-death switchNat Rev
Cancer2647656200210.1038/nrc88312209154
|