Introduction
MicroRNAs (miRNAs or miRs) are small non-coding RNAs
that regulate both the mRNA and the protein expression of target
genes (1). miRNAs play a crucial
role in the regulation of genes involved in the control of
development, cell proliferation, apoptosis and stress response
(2). The expression profile
analysis of miRNAs is essential for understanding the complex
regulation of gene expression that involves miRNAs in addition to
characterizing miRNAs themselves (3). Some miRNAs exhibit tissue-specific
expression (4). The liver is a
crucial organ in which miRNAs may be involved in the regulation of
hepatocyte growth and development. The expression profiles of
miRNAs in fetal liver are different from those in the adult liver;
fetal miRNA expression has shown specificity in the developmental
stage (5). Global gene and miRNA
expression in embryonic and adult human livers has been analyzed,
revealing multiple regulated genes and demonstrating a change in
the expression patterns during the developmental process (6). A number of studies have focused on
accelerated aging and miRNAs, such as miRNAs in the aging mouse
brain (3,7). Previous studies have analyzed the
expression of miRNAs in the livers of young and elderly humans
(6) and rats (8), as well as in mice with delayed aging
(9,10). However, the association between
miRNA expression and the aging of the liver remains poorly
understood (11).
In the present study, we identified a gradual up-
and downregulation of miRNAs in rats at 7 different developmental
stages (fetal to 36 weeks old). We found that miR-29a, miR-29c,
miR-195 and miR-497 were gradually upregulated and that miR-301a,
miR-148b-3p, miR-7a, miR-93, miR-106b, miR-185, miR-450a, miR-539
and miR-301b were gradually downregulated in livers as the rats
aged.
Materials and methods
Animals
Inbred Wistar rats were bred under specific
pathogen-free conditions at the Institute for Animal
Experimentation of the Kagawa University School of Medicine Kagawa,
Japan. Eight male Wistar rats from the ages of post-natal day 3,
and post-natal week 1, 2, 4, 8 and 36 were used in this study.
Eight fetal livers were also obtained from pregnant rats at day 20.
We performed hepatectomy under ether anesthesia. For
immunohistochemistry, small sections of the liver tissue were
preserved in 10% formalin. The remaining sections of the liver
tissue were rapidly placed in liquid nitrogen, transferred
individually to pre-weighed tubes containing RNAlater (Ambion,
Tokyo, Japan) and stored at −80°C until analysis.
RNA isolation
The frozen tissue samples were thawed on ice and
dissolved in TRIzol reagent (Invitrogen Life Technologies,
Carlsbad, CA, USA). Total RNA was extracted from the tissues using
an miRNeasy Mini kit (Qiagen, Tokyo, Japan) according to the
manufacturer’s instructions. Briefly, 700 μl of TRIzol reagent
containing samples was mixed vigorously with 140 μl of chloroform,
incubated at room temperature for 3 min, followed by centrifugation
at 12,000 × g for 15 min at 4°C. The upper aqueous phase was
transferred to another tube containing 525 μl of 100% ethanol. The
mixture was loaded into RNeasy Mini columns, followed by serial
washing with solutions provided with the kit. Finally, RNA was
collected into RNase-free water for further experiments. All RNA
samples used in this study showed A260/280 ratios between 2.0 and
2.1. The integrity of RNA was determined using a NanoDrop 2000
spectrophotometer (Thermo Scientific, Rockford, IL, USA). The
quality of total RNA was determined using the RNA Nano 6000 chips
on an Agilent 2100 Bioanalyzer (Agilent Technologies Inc., Osaka,
Japan), and all RNA samples used for microarray analyses had RIN
values >8.2. These RNA samples were stored at −80°C.
miRNA arrays
Total RNA was labeled with Hy3 dye using the array
labeling kit (Exiqon, Vedbæk, Denmark). Total RNA (2 μg) was
incubated with a spike for 30 min at 37°C and then at 95°C for 5
min. Hy3 dye and labeling enzyme were added to the samples. The
enzyme was then heat-inactivated at 16°C for 1 h and 65°C for 15
min, protected from light and then received an addition of
hybridization buffer. The sample was loaded onto the arrays by
capillary force using 3D-Gene miRNA oligo chips (version 16; Toray
Industries, Inc., Kanagawa, Japan). The chips enabled the
examination of the expression of 679 miRNAs printed in duplicate
spots. The arrays were incubated at 32°C for 16 h, then briefly
washed in a 30°C wash buffer solution (0.5X SSC, 0.1% SDS), rinsed
in wash buffer solution (0.2X SSC, 0.1% SDS) followed by a wash in
another buffer solution (0.05X SSC), according to the
manufacturer’s instructions (Toray Industries, Inc.). The arrays
were spun for 1 min at 1,000 rpm for drying, followed by immediate
scanning using a Toray 3D-Gene 3000 miRNA microarray scanner (Toray
Industries, Inc.). The relative expression level of each miRNA was
calculated by comparing the average signal intensities of the valid
spots with their mean value throughout the microarray experiments
following normalization to their adjusted median values.
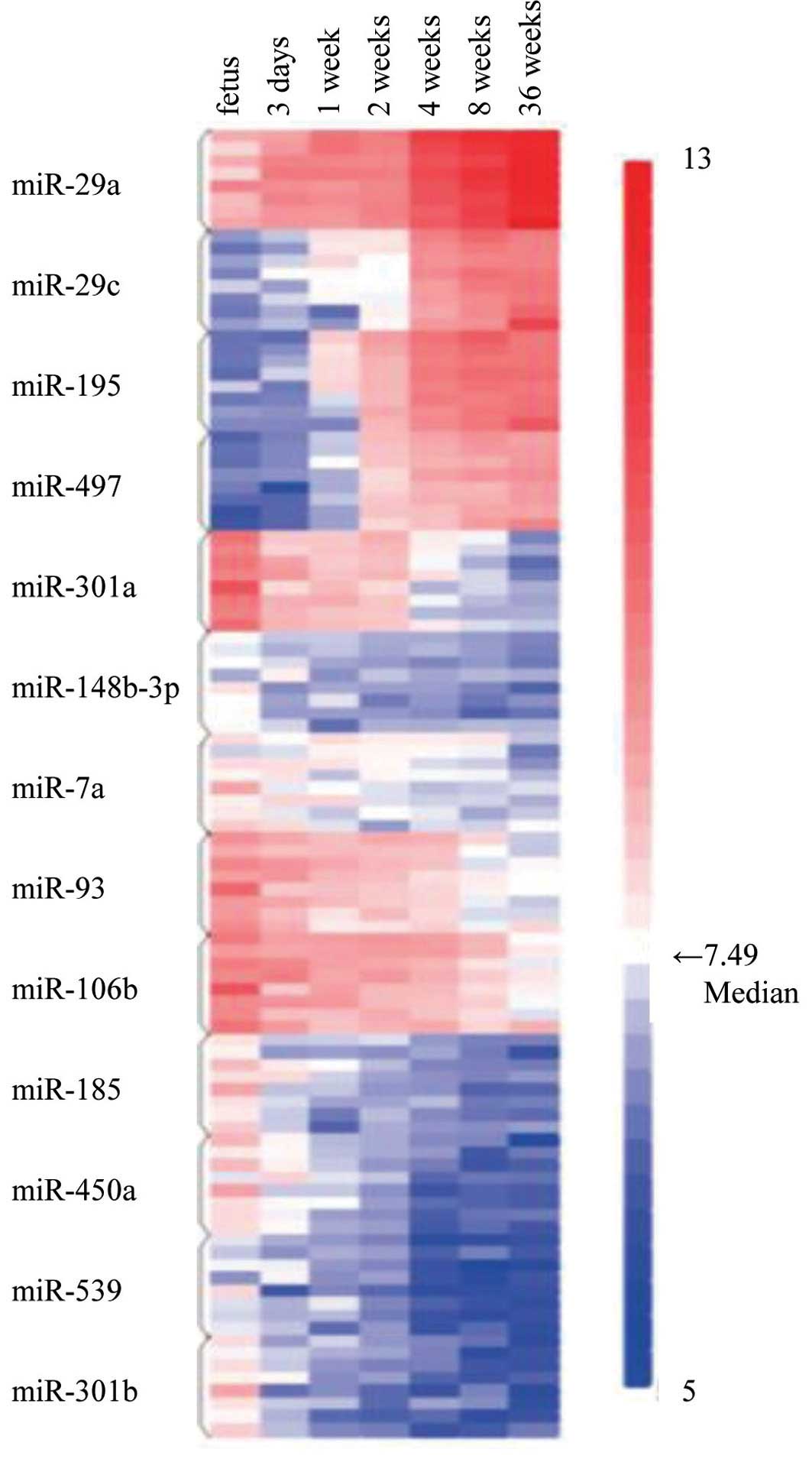
Heatmap
To illustrate the alterations in the expression
levels of the 13 upregulated or downregulated miRNAs, we created a
heatmap in which each cell represents the expression level of each
of the 13 miRNAs for each of 8 individual rats at different stages
of development, namely the fetal period, at post-natal day 3 and
post-natal week 1, 2, 4, 8 and 36. The heatmap was color-coded
according to the log2-transformed expression level. The center
level of the color code is set as the median value over all of the
values used in the heatmap. Briefly, in the heatmap, white color
represents mean values, red indicates gains and blue represents
losses.
Immunohistochemistry
The avidin-biotin complex (ABC) immunohistochemical
method was used. Following formalin fixation, the tissues were
dehydrated in a graded series of ethanol and embedded in paraffin.
The serial sections were mounted onto glass slides.
Immunohistochemical staining for proliferating cell nuclear antigen
(PCNA) was performed using the following procedure: the sections
were deparaffinized, hydrated and quenched for endogenous
peroxidase with 0.3% hydrogen peroxide in PBS at room temperature
for 30 min. The sections were permeabilized in PBS with 0.3%
polyoxyethylene (10) octyl
phenyl ether (Triton X-100) (Wako, Osaka, Japan) for 1 h before
quenching the endogenous peroxidase activity. The sections were
then incubated overnight at room temperature with a mouse anti-PCNA
antibody (1:100 dilution; Dako, Glostrup, Denmark). Antibody
binding was detected using a Vectastain Elite ABC kit (Vector
Laboratories, Burlingame, CA, USA) and 3,3′-diaminobenzidine as a
chromogen. After staining, all sections were counterstained with
Mayer’s hematoxylin. The specificity of immunostaining was examined
using non-immune mouse IgG as a negative control for the primary
antibody. Images were captured using an Olympus BX51 microscope and
Olympus DP72 camera (Olympus, Tokyo, Japan). The nuclear labeling
index for the PCNA-positive cells (positive nuclei/total counted)
was determined by evaluating at least 500 hepatocytes at random in
the microscopic field by 2 observers (T. Masaki and S. Mimura).
Western blot analysis
Frozen liver tissue samples were homogenized in a
protein extraction solution (PRO-PERP™; Intron Biotechnology, Inc.,
Gyeonggi, Korea), and cell lysis was then induced by incubating the
samples for 20 min on ice. The samples were prepared by
centrifugation at 13,000 rpm for 5 min at 4°C. The supernatant was
then collected. The protein concentration was measured by a
dye-binding protein assay performed according to the Bradford
method (12). Sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed
according to the method of Laemmli (13), and western blot analysis was
performed as previously described by Towbin et al (14) using anti-β-actin monoclonal
antibody (Sigma-Aldrich; A5442, used at 1:3,000) and cyclin D1
(Thermo Fisher Scientific, Waltham, MA, USA; RB-9041, used at
1:1,000) as primary antibodies and horseradish peroxidase-inked
anti-mouse and anti-rabbit IgG secondary antibodies (GE Healthcare
UK, Buckinghamshire, UK; used at 1:2,000).
Results
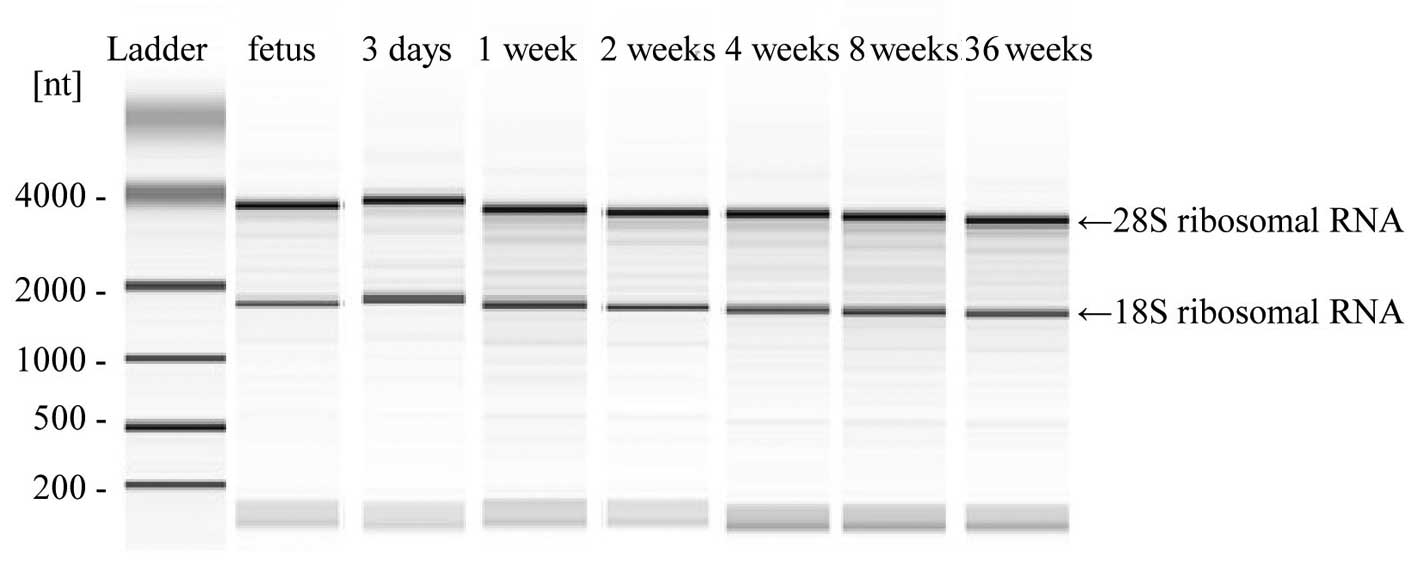
Quality assessment of the total RNA of
the sample using a miRNA array chip
Total RNA (20 μl) from all liver tissues used in
this study was loaded into each lane, and the bands of 18S and 28S
ribosomal RNA in the gel were detected using the 2100 Bioanalyzer
(Fig. 1). Based on these results,
RNA samples extracted from various liver tissues were shown to be
of adequate quality.
Developmental study of miRNA expression
in liver tissue
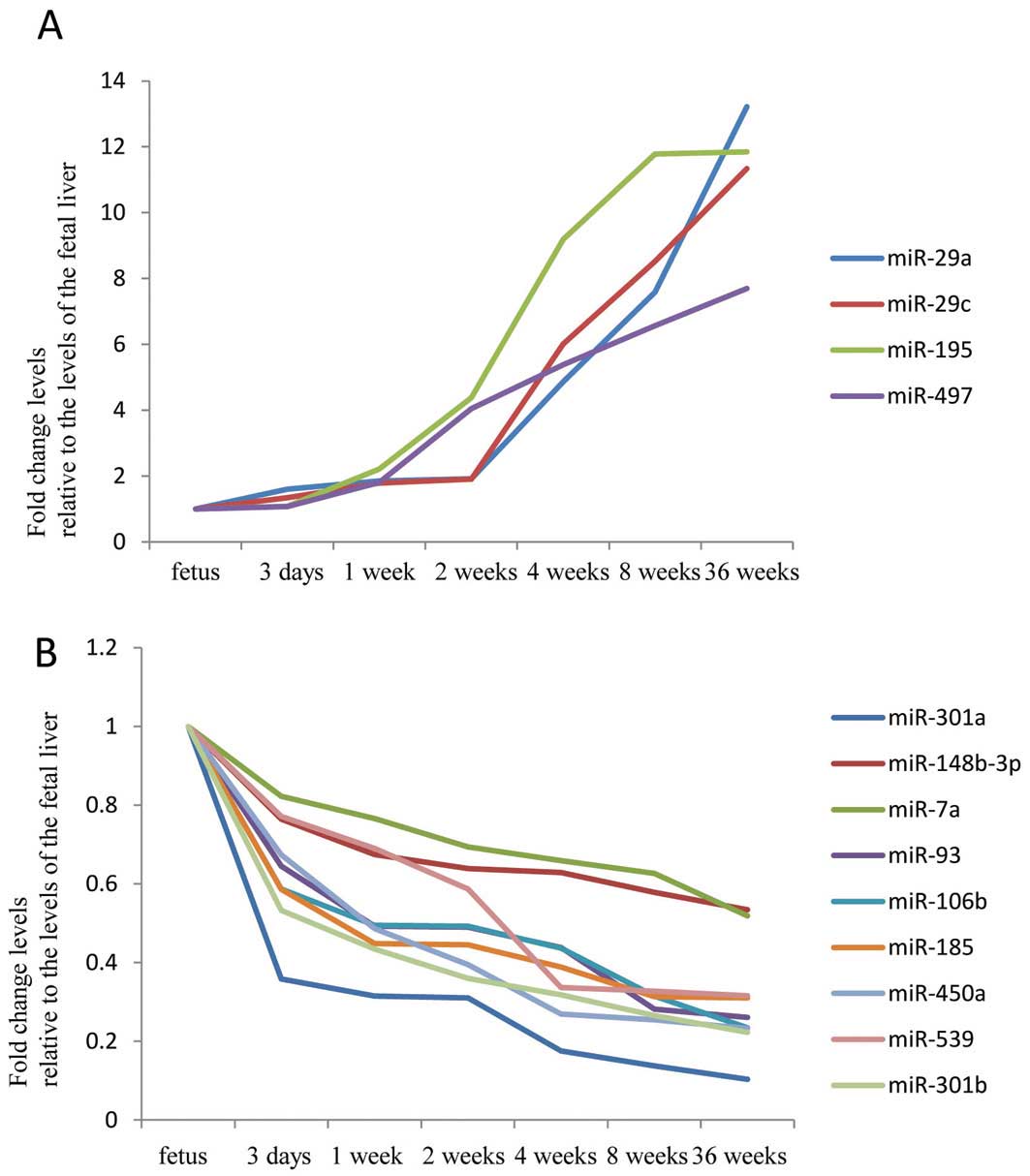
Using a custom microarray platform, we analyzed the
expression levels of 679 rat miRNA probes. Four miRNAs were found
to be gradually upregulated with age in the rat livers. The 4
upregulated miRNAs were miR-29a (spot no. 1), miR-29c (spot no. 2),
miR-195 (spot no. 3) and miR-497 (spot no. 4) (Figs. 2 and 3). By contrast, 9 miRNAs were gradually
downregulated. These downregulated miRNAs were miR-301a (spot no.
5), miR-148b-3p (spot no. 6), miR-7a (spot no. 7), miR-93 (spot no.
8), miR-106b (spot no. 9), miR-185 (spot no. 10), miR-450a (spot
no. 11), miR-539 (spot no. 12) and miR-301b (spot no. 13). The
heatmap (Fig. 4) clearly
demonstrates increasing trends for 4 miRNAs and decreasing trends
for 9 miRNAs in the expression levels among the age groups. It also
showed that the trends in miRNA expression were fairly consistent
among individual rats and were constant in both increasing and
decreasing directions.
 | Figure 2MicroRNA (miRNA) expression in rat
livers at 7 different stages of development (fetal, post-natal day
3 and post-natal weeks 1, 2, 4, 8 and 36) in miRNA chips. Spot
numbers 1–4 (1, miR-29a; 2, miR-29c; 3, miR-195; 4, miR-497) were
miRNAs upregulated with liver development. Spot numbers 5–13 (5,
miR-301a; 6, miR-148b-3p; 7, miR-7a; 8, miR-93; 9, miR-106b; 10,
miR-185; 11, miR-450a; 12, miR-539; 13, miR-301b) were miRNAs
downregulated with liver development. |
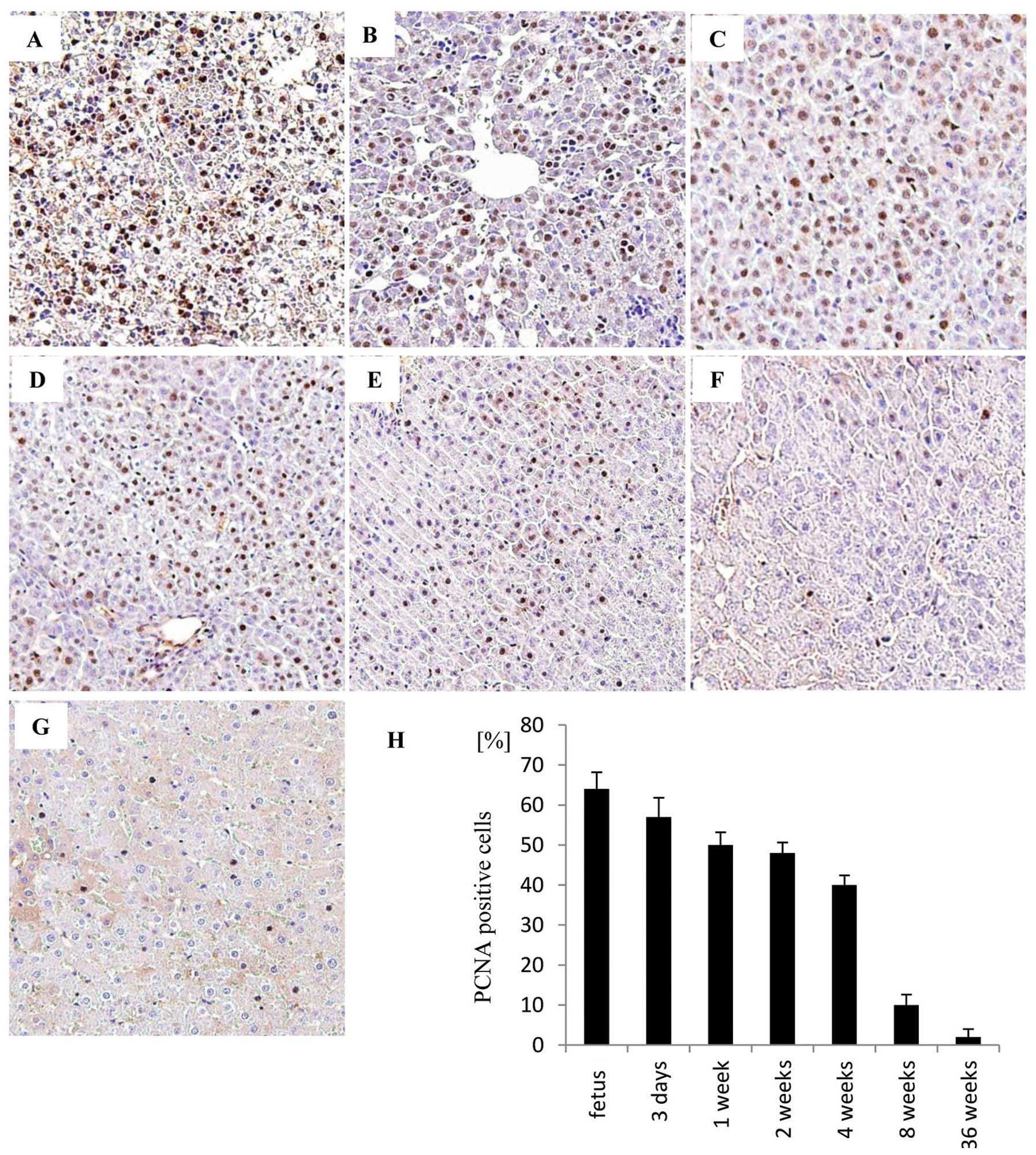
Immunohistochemical study of PCNA in
developing livers
The typical immunohistochemical staining pattern of
PCNA at various stages of development in the rat livers, including
fetal liver (Fig. 5A), post-natal
day 3 liver (Fig. 5B), post-natal
week 1 liver (Fig. 5C),
post-natal week 2 liver (Fig.
5D), postnatal week 4 liver (Fig.
5E), post-natal week 8 liver (Fig. 5F) and post-natal week 36 liver
(Fig. 5G). The labeling index of
PCNA in the hepatocytes in the developing livers decreased with age
(Fig. 5H).
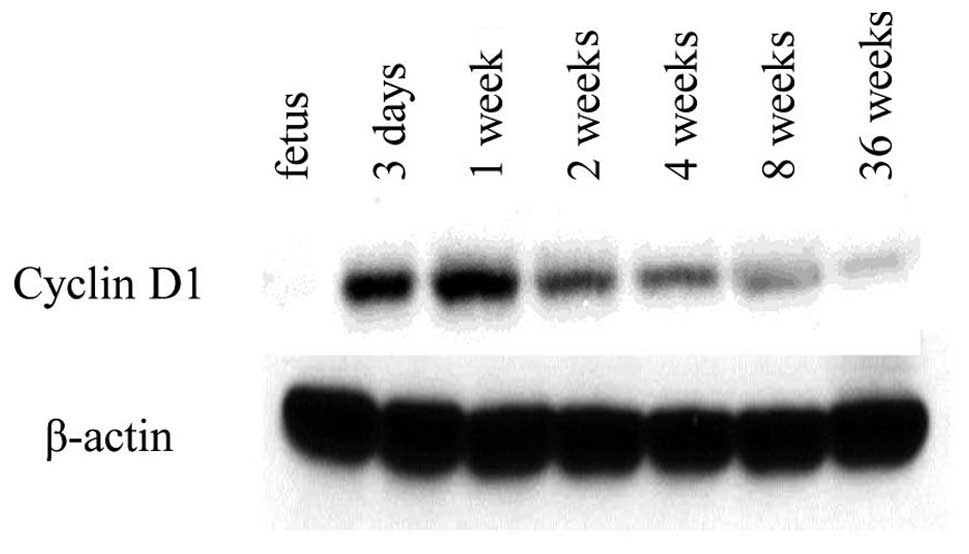
Western blot analysis of aging rat
liver
Western blot analysis was used to examine the cell
cycle of the aging rat liver by using an antibody against cyclin
D1. The cyclin D1 level was hardly detectable in the fetal liver
(Fig. 6). However, cyclin D1 was
detected in the liver at post-natal day 3. The highest level was
detected in the post-natal week 1 liver and the level slightly
decreased with age. The amount of β-actin (an internal control for
protein loading) was almost the same in all age groups.
Discussion
The dysregulation of miRNA expression may contribute
to numerous alterations present in very complex and multifactorial
processes, namely, proliferation, differentiation, inflammation,
carcinogenesis and aging (11).
The profiles of miRNAs have been reported by developmental studies
in various organs, such as the brain (15–18), kidneys (19), pancreas (20) and liver (21,22). However, the alterations in miRNA
expression in the rat liver during the process of aging remain
poorly understood. In the present study, in order to detect miRNAs
associated with aging in the rat liver, we investigated miRNAs with
a gradually increasing or decreasing expression in the livers of
rats at different stages of development. Such miRNAs appear to play
an important role in aging. To the best of our knowledge, to date,
no studies have established an association between miRNAs and aging
of the liver in a similar manner as we have done.
In the present study, we demonstrated that miR-29a,
miR-29c, miR-195 and miR-497 were upregulated with age, whereas
miR-301a, miR-148b-3p, miR-7a, miR-93, miR-106b, miR-185, miR-450a,
miR-539 and miR-301b were downregulated with age in the rat livers.
These data suggest that specific miRNAs are associated with aging
in the liver.
miR-29a and miR-29c were upregulated during early
development in the liver (rats at post-natal week 4 and 8) and
during late development in the liver (post-natal week 36). Notably,
the expression levels of miR-29a and miR-29c were >10-fold
higher in the livers from the 36-week-old rats than the fetal rat
livers. miR-29a and miR-29c have been shown to be age-related in
various organs, such as the aorta (23,24), the lungs (25,26), the kidneys (19) and the liver in mice (21,22). The upregulation of miR-29a and
miR-29c with age is consistent with a recent report showing an
increased expression of miR-29 family members in a model of
accelerated aging in mice (11).
Several studies have indicated that the cellular
replication capacity (proliferative activity) declines in various
tissues and may compromise the immune system during aging (27–29). In addition, the number of
senescent cells increases in various tissues with aging (30). Indeed, in this study, we also
demonstrated that the expression of PCNA in hepatocytes decreased
with age. These data suggest that the number of senescent cells in
the liver increased with age.
In the present study, the 4 miRNAs with a gradually
upregulated expression (miRNA-29a, miRNA29c, miRNA-195 and miR-497)
possess target genes to promote proliferative activity, whereas the
4 miRNAs (miR-148-3p, miR-93, miR-106b and miR-185) whose
expression was gradually reduced, possess genes to inhibit
proliferative activity, such as tumor suppressor gene and the
cyclin-dependent kinase inhibitor, p21WAF1 (Table I). Hepatocyte growth factor (HGF),
cyclin E, Cdk4, Cdk6 and cyclin D1 play important roles in the
progression of the cell cycle, and their molecules are the targets
of upregulated miRNAs (Table I).
By contrast, p21 (also known as p21CIP1/WAF1), sirtuin and fused in
sarcoma (FUS)1 play important roles in the arrest of the cell
cycle, and their molecules are targets of downregulated miRNAs
(Table I). Therefore, these data
suggest that numerous up- and downregulated miRNAs play an
important role in the decline of the proliferative activity of
hepatocytes with age.
 | Table IChromosomal locations and target
genes of the up- and downregulated miRNAs. |
Table I
Chromosomal locations and target
genes of the up- and downregulated miRNAs.
| Name | Genome context | | Target gene
(Ref.) |
|---|
| Upregulated |
| rno-miR-29a | 4:
58107760–58107847 [-] | 4q22 | p53 (11), hepatocyte growth factor (31), Bcl-2 (32), nuclear autoantigenic sperm protein
(33), TGF-β1 (34)
p42.3 (35), Arpc3 (36), monocarboxylate transporter 1
(37), maternally expressed gene
3 (38), Tcl1 (39), DNA methyltransferase 3 (40), phosphatase and tensin homolog
(41) |
| rno-miR-29c | 13:
110968048–110968135 [+] | 13q27 | p53 (11), MCT-1 (37), Bcl-2 (32), TGF-β1 (34), tumor necrosis factor α-induced
protein 3 (42), β-site APP
cleaving enzyme 1 (43), cyclin E
(44) |
| rno-miR-195 | 10:
57074170–57074256 [+] | 10q24 | Cyclin-dependent
kinase 4 (45), Bcl-2 (46–48), ethanol-mediated inhibition of
hepatic sirtuin 1 (47), cyclin
E1 (49), cyclin D1 (50,51), CDK6 (50), E2F3 (50,52)
Raf-1 (52) |
| rno-miR-497 | 10:
57073846–57073914 [+] | 10q24 | Bcl-2 (53,54), cyclin D2 (54) |
| Downregulated |
| rno-miR-301a | 10:
75386838–75386937 [+] | 10q26 | Plasminogen
activator inhibitor-1 (55) |
|
rno-miR-148b-3p | | | AMPKα1 (56), ITGA5, ROCK1, PIK3CA/p110α, NRAS,
CSF1 (57) |
| rno-miR-7a | | | Barx1 (58) |
| rno-miR-93 | 12:
17608173–17608259 [−] | 12q11 | Sirtuin 1 (8), S-transferase 1 (8), integrin-β8 (59), FUS1 (60), p21 (61) |
| rno-miR-106b | 12:
17608382–17608463 [−] | 12q11 | p21 (61–64), RB (65) |
| rno-miR-185 | 11:
84658785–84658864 [+] | 11q23 | DNA
methyltransferase 1 (66), Rho,
Cdc42 (67), Six1 (68) |
| rno-miR-450a | X:
139994947–139995037 [−] | Xq36 | DNA
methyltransferase 3a (69) |
| rno-miR-539 | 6:
134408917–134408992 [+] | 6q32 | Holocarboxylase
synthetase (HCS) (70,71) |
| rno-miR-301b | 11:
85885248–85885325 [+] | 11q23 | TP63 (72) |
In conclusion, in the present study, we identified
the upregulation of miR-29a, miR-29c, miR-195 and miR-497, and the
downregulation of miR-301a, miR-148b-3p, miR-7a, miR-93, miR-106b,
miR-185, miR-450a, miR-539 and miR-301b in the rat liver with age.
Our data also suggest that important changes in miRNA expression
occur during development, and one result of aging is likely to be
changes in miRNA expression. Our findings suggest that these up-
and downregulated miRNAs play important roles by regulating cell
cycles that are related to liver senescence. Further studies are
required to clarify additional miRNA targets, as well as the roles
of miRNAs in the development of the complex process of aging in the
liver.
Acknowledgements
We thank Madoka Seguchi, Yuuko Miyawaki, Fuyuko
Kokado and Ryoko Unose (Kagawa University) for their excellent
technical assistance.
Abbreviations:
|
miRNA
|
microRNA
|
|
PCNA
|
proliferating cell nuclear antigen
|
|
ABC
|
avidin-biotin complex
|
|
SDS-PAGE
|
sodium dodecyl sulfate-polyacrylamide
gel electrophoresis
|
|
HGF
|
hepatocyte growth factor
|
References
|
1
|
Lakner AM, Bonkovsky HL and Schrum LW:
MicroRNAs: Fad or future of liver disease. World J Gastroenterol.
17:2536–2542. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pogribny IP, Tryndyak VP, Boyko A, et al:
Induction of microRNAome deregulation in rat liver by long-term
tamoxifen exposure. Mutat Res. 619:30–37. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Eda A, Takahashi M, Fukushima T and Hohjoh
H: Alteration of microRNA expression in the process of mouse brain
growth. Gene. 485:46–52. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lagos-Quintana M, Rauhut R, Yalcin A, et
al: Identification of tissue-specific microRNAs from mouse. Curr
Biol. 12:735–739. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lagos-Quintana M, Rauhut R, Lendeckel W
and Tuschl T: Identification of novel genes coding for small
expressed RNAs. Science. 294:853–858. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tzur G, Israel A, Levy A, et al:
Comprehensive gene and microRNA expression profiling reveals a role
for microRNAs in human liver development. PLoS One. 4:75112009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li N, Bates DJ, An J, et al: Up-regulation
of key microRNAs, and inverse down-regulation of their predicted
oxidative phosphorylation target genes, during aging in mouse
brain. Neurobiol Aging. 32:944–955. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li N, Muthusamy S, Liang R, et al:
Increased expression of miR-34a and miR-93 in rat liver during
aging, and their impact on the expression of Mgst1 and Sirt1. Mech
Ageing Dev. 132:75–85. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bates DJ, Li N, Liang R, et al: MicroRNA
regulation in Ames dwarf mouse liver may contribute to delayed
aging. Aging Cell. 9:1–18. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Maes OC, An J, Sarojini H and Wang E:
Murine microRNAs implicated in liver functions and aging process.
Mech Ageing Dev. 129:534–541. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ugalde AP, Ramsay AJ, de la Rosa J, et al:
Aging and chronic DNA damage response activate a regulatory pathway
involving miR-29 and p53. EMBO J. 30:2219–2232. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bradford M: A rapid and sensitive method
for the quantitation of microgramof protein utilizing the principle
of protein-dye binding. Anal Biochem. 72:248–254. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bactriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Towbin H, Staehelin T and Gordon J:
Electrophoretic transfer of proteins from polyacrylamide gels to
nitrocellulose sheets: procedure and some applications. Proc Natl
Acad Sci USA. 76:4350–4354. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Aw S and Cohen SM: Time is of the essence:
microRNAs and age-associated neurodegeneration. Cell Res.
22:1218–1220. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Inukai S, de Lencastre A, Turner M and
Slack F: Novel microRNAs differentially expressed during aging in
the mouse brain. PLoS One. 7:e400282012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ungvari Z, Tucsek Z, Sosnowska D, et al:
Aging-induced dysregulation of dicer1-dependent microRNA expression
impairs angiogenic capacity of rat cerebromicrovascular endothelial
cells. J Gerontol A Biol Sci Med Sci. 68:877–891. 2013. View Article : Google Scholar
|
|
18
|
Rao YS, Mott NN, Wang Y and Chung WC:
MicroRNAs in the aging female brain: a putative mechanism for
age-specific estrogen effects. Endocrinology. 154:2795–2806. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ma L and Qu L: The function of microRNAs
in renal development and pathophysiology. J Genet Genomics.
40:143–152. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dumortier O and Van Obberghen E: MicroRNAs
in pancreas development. Diabetes Obes Metab. 14(Suppl 3): 22–28.
2012. View Article : Google Scholar
|
|
21
|
Ugalde AP, Español Y and López-Otín C:
Micromanaging aging with miRNAs: new messages from the nuclear
envelope. Nucleus. 2:549–555. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Roderburg C, Urban GW, Bettermann K, et
al: Micro-RNA profiling reveals a role for miR-29 in human and
murine liver fibrosis. Hepatology. 53:209–218. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Boon RA, Seeger T, Heydt S, et al:
MicroRNA-29 in aortic dilation: implications for aneurysm
formation. Circ Res. 109:1115–1119. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Milewicz DM: MicroRNAs, fibrotic
remodeling, and aortic aneurysms. J Clin Invest. 122:490–493. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Williams AE, Perry MM, Moschos SA and
Lindsay MA: MicroRNA expression in the aging mouse lung. BMC
Genomics. 8:1722007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang Y, Scheiber MN, Neumann C, et al:
MicroRNA regulation of ionizing radiation-induced premature
senescence. Int J Radiat Oncol Biol Phys. 81:839–848. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schneider EL, Sternberg H, Tice RR, et al:
Cellular replication and aging. Mech Ageing Dev. 9:313–324. 1979.
View Article : Google Scholar
|
|
28
|
Kuhn HG, Dickinson-Anson H and Gage FH:
Neurogenesis in the dentate gyrus of the adult rat: age-related
decrease of neuronal progenitor proliferation. J Neurosci.
16:2027–2033. 1996.PubMed/NCBI
|
|
29
|
Timchenko NA: Aging and liver
regeneration. Trends Endocrinol Metab. 20:171–176. 2009. View Article : Google Scholar
|
|
30
|
Salminen A, Ojala J and Kaarniranta K:
Apoptosis and aging: increased resistance to apoptosis enhances the
aging process. Cell Mol Life Sci. 68:1021–1031. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kwiecinski M, Noetel A, Elfimova N, et al:
Hepatocyte growth factor (HGF) inhibits collagen I and IV synthesis
in hepatic stellate cells by miRNA-29 induction. PLoS One.
6:e245682011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xiong Y, Fang JH, Yun JP, et al: Effects
of microRNA-29 on apoptosis, tumorigenicity, and prognosis of
hepatocellular carcinoma. Hepatology. 51:836–845. 2010.PubMed/NCBI
|
|
33
|
Ma W, Xie S, Ni M, et al: MicroRNA-29a
inhibited epididymal epithelial cell proliferation by targeting
nuclear autoantigenic sperm protein (NASP). J Biol Chem.
287:10189–10199. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang B, Komers R, Carew R, et al:
Suppression of microRNA-29 expression by TGF-β1 promotes collagen
expression and renal fibrosis. J Am Soc Nephrol. 23:252–265.
2012.
|
|
35
|
Cui Y, Su WY, Xing J, et al: MiR-29a
inhibits cell proliferation and induces cell cycle arrest through
the downregulation of p42.3 in human gastric cancer. PLoS One.
6:e258722011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lippi G, Steinert JR, Marczylo EL, et al:
Targeting of the Arpc3 actin nucleation factor by miR-29a/b
regulates dendritic spine morphology. J Cell Biol. 194:889–904.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pullen TJ, da Silva Xavier G, Kelsey G and
Rutter GA: miR-29a and miR-29b contribute to pancreatic
beta-cell-specific silencing of monocarboxylate transporter 1
(Mct1). Mol Cell Biol. 31:3182–3194. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Braconi C, Kogure T, Valeri N, et al:
microRNA-29 can regulate expression of the long non-coding RNA gene
MEG3 in hepatocellular cancer. Oncogene. 30:4750–4756. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pekarsky Y, Santanam U, Cimmino A, et al:
Tcl1 expression in chronic lymphocytic leukemia is regulated by
miR-29 and miR-181. Cancer Res. 66:11590–11593. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fabbri M, Garzon R, Cimmino A, et al:
MicroRNA-29 family reverts aberrant methylation in lung cancer by
targeting DNA methyltransferases 3A and 3B. Proc Natl Acad Sci USA.
104:15805–15810. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kong G, Zhang J, Zhang S, et al:
Upregulated microRNA-29a by hepatitis B virus X protein enhances
hepatoma cell migration by targeting PTEN in cell culture model.
PLoS One. 6:e195182011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang CM, Wang Y, Fan CG, et al: miR-29c
targets TNFAIP3, inhibits cell proliferation and induces apoptosis
in hepatitis B virus-related hepatocellular carcinoma. Biochem
Biophys Res Commun. 411:586–592. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zong Y, Wang H, Dong W, et al: miR-29c
regulates BACE1 protein expression. Brain Res. 1395:108–115. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ding DP, Chen ZL, Zhao XH, et al: miR-29c
induces cell cycle arrest in esophageal squamous cell carcinoma by
modulating cyclin E expression. Carcinogenesis. 32:1025–1032. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lin Y, Wu J, Chen H, et al:
Cyclin-dependent kinase 4 is a novel target in micoRNA-195-mediated
cell cycle arrest in bladder cancer cells. FEBS Lett. 586:442–447.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chen YQ, Wang XX, Yao XM, et al:
MicroRNA-195 promotes apoptosis in mouse podocytes via enhanced
caspase activity driven by BCL2 insufficiency. Am J Nephrol.
34:549–559. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhu H, Yang Y, Wang Y, et al: MicroRNA-195
promotes palmitate-induced apoptosis in cardiomyocytes by
down-regulating Sirt1. Cardiovasc Res. 92:75–84. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu L, Chen L, Xu Y, et al: microRNA-195
promotes apoptosis and suppresses tumorigenicity of human
colorectal cancer cells. Biochem Biophys Res Commun. 400:236–240.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sekiya Y, Ogawa T, Iizuka M, et al:
Down-regulation of cyclin E1 expression by microRNA-195 accounts
for interferon-β-induced inhibition of hepatic stellate cell
proliferation. J Cell Physiol. 226:2535–2542. 2011.PubMed/NCBI
|
|
50
|
Xu T, Zhu Y, Xiong Y, et al: MicroRNA-195
suppresses tumorigenicity and regulates G1/S transition of human
hepatocellular carcinoma cells. Hepatology. 50:113–21. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li D, Zhao Y, Liu C, et al: Analysis of
MiR-195 and MiR-497 expression, regulation and role in breast
cancer. Clin Cancer Res. 17:1722–1730. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhang QQ, Xu H, Huang MB, et al:
MicroRNA-195 plays a tumor-suppressor role in human glioblastoma
cells by targeting signaling pathways involved in cellular
proliferation and invasion. Neuro Oncol. 14:278–287. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhu W, Zhu D, Lu S, et al: miR-497
modulates multidrug resistance of human cancer cell lines by
targeting BCL2. Med Oncol. 29:384–391. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yadav S, Pandey A, Shukla A, et al:
miR-497 and miR-302b regulate ethanol-induced neuronal cell death
through BCL2 protein and cyclin D2. J Biol Chem. 286:37347–37357.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Patel N, Tahara SM, Malik P and Kalra VK:
Involvement of miR-30c and miR-301a in immediate induction of
plasminogen activator inhibitor-1 by placental growth factor in
human pulmonary endothelial cells. Biochem J. 434:473–482. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhao G, Zhang JG, Liu Y, et al: miR-148b
functions as a tumor suppressor in pancreatic cancer by targeting
AMPKα1. Mol Cancer Ther. 12:83–93. 2013.PubMed/NCBI
|
|
57
|
Cimino D, De Pittà C, Orso F, et al:
miR148b is a major coordinator of breast cancer progression in a
relapse-associated microRNA signature by targeting ITGA5, ROCK1,
PIK3CA, NRAS, and CSF1. FASEB J. 27:1223–1235. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kim BM, Woo J, Kanellopoulou C and
Shivdasani RA: Regulation of mouse stomach development and Barx1
expression by specific microRNAs. Development. 138:1081–1086. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Fang L, Deng Z, Shatseva T, et al:
MicroRNA miR-93 promotes tumor growth and angiogenesis by targeting
integrin-β8. Oncogene. 30:806–821. 2011.PubMed/NCBI
|
|
60
|
Du L, Schageman JJ, Subauste MC, et al:
miR-93, miR-98, and miR-197 regulate expression of tumor suppressor
gene FUS1. Mol Cancer Res. 7:1234–1243. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kim YK, Yu J, Han TS, et al: Functional
links between clustered microRNAs: suppression of cell-cycle
inhibitors by microRNA clusters in gastric cancer. Nucleic Acids
Res. 37:1672–1681. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Ivanovska I, Ball AS, Diaz RL, et al:
MicroRNAs in the miR-106b family regulate p21/CDKN1A and promote
cell cycle progression. Mol Cell Biol. 28:2167–2174. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li B, Shi XB, Nori D, et al:
Down-regulation of microRNA 106b is involved in p21-mediated cell
cycle arrest in response to radiation in prostate cancer cells.
Prostate. 71:567–574. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hu S, Dong TS, Dalal SR, et al: The
microbe-derived short chain fatty acid butyrate targets
miRNA-dependent p21 gene expression in human colon cancer. PLoS
One. 6:e162212011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Cai K, Wang Y and Bao X: MiR-106b promotes
cell proliferation via targeting RB in laryngeal carcinoma. J Exp
Clin Cancer Res. 30:732011. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zhang Z, Tang H, Wang Z, et al: MiR-185
targets the DNA methyltransferases 1 and regulates global DNA
methylation in human glioma. Mol Cancer. 10:1242011. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Liu M, Lang N, Chen X, et al: miR-185
targets RhoA and Cdc42 expression and inhibits the proliferation
potential of human colorectal cells. Cancer Lett. 301:151–160.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Imam JS, Buddavarapu K, Lee-Chang JS, et
al: MicroRNA-185 suppresses tumor growth and progression by
targeting the Six1 oncogene in human cancers. Oncogene.
29:4971–4979. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Weng Z, Wang D, Zhao W, et al:
microRNA-450a targets DNA methyltransferase 3a in hepatocellular
carcinoma. Exp Ther Med. 2:951–955. 2011.PubMed/NCBI
|
|
70
|
Bao B, Rodriguez-Melendez R, Wijeratne SS
and Zempleni J: Biotin regulates the expression of holocarboxylase
synthetase in the miR-539 pathway in HEK-293 cells. J Nutr.
140:1546–1551. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Bao B, Rodriguez-Melendez R and Zempleni
J: Cytosine methylation in miR-153 gene promoters increases the
expression of holocarboxylase synthetase, thereby increasing the
abundance of histone H4 biotinylation marks in HEK-293 human kidney
cells. J Nutr Biochem. 23:635–39. 2012. View Article : Google Scholar
|
|
72
|
Funamizu N, Lacy CR, Parpart ST, et al:
MicroRNA-301b promotes cell invasiveness through targeting TP63 in
pancreatic carcinoma cells. Int J Oncol. 44:725–734.
2014.PubMed/NCBI
|