Introduction
Seaweed is widely consumed throughout Asian
countries, such as China, Japan, Korea, Vietnam, Indonesia, the
Philippines and Hawaii, as a natural medicinal and food source
(1). Seaweed contains a variety
of nutrients, such as carbohydrates, vitamins, minerals, fatty
acids, dietary fiber, amino acids, iodine and essential
polysaccharides. Research on the chemical and nutritional
composition of seaweed has revealed that it has antioxidant,
antitumor and antibacterial activities (2,3).
The red seaweed, Pyropia yezoensis, is an economically
important seaweed in Asian countries, such as China, Japan and
Korea. Pyropia yezoensis is an important source of
physiologically active substances that contain 25–50% protein and
25–40% carbohydrates based on dry matter weight (4). Pyropia yezoensis has been
shown to exert biological effects, such as antitumor, anti-fatigue,
blood pressure-reducing, anti-inflammatory, antioxidant and
hepatoprotective effects (5,6).
In a previous study, we examined a peptide from Pyropia
yezoensis which stimulates the proliferation of IEC-6 cells by
activating the insulin-like growth factor I receptor signaling
pathway (7). In the present
study, we examined the effects of a Pyropia yezoensis
peptide [PYP1 (1–20)] on cell proliferation and related
EGFR signaling pathways in IEC-6 cells, a rat intestinal epithelial
cell line.
Cell proliferation is induced through intracellular
signal transduction mediated by receptor tyrosine kinases (RTKs),
such as epidermal growth factor receptor (EGFR) (8). RTKs are cell membrane receptors for
growth factors and other extracellular ligands. RTKs mediate
cellular tyrosine phosphorylation and regulate intracellular
signaling pathways, such as those involved in cell migration,
differentiation and proliferation (9–11).
RTKs of the EGFR family mediate essential cellular functions,
including the regulation of cellular proliferation, growth,
survival, migration, differentiation and development in normal and
pathological states (12,13). The binding of EGF to EGFR
initiates a number of molecular events. The EGF-EGFR molecular
interaction activates growth-promoting signals primarily through
the activation of Ras, leading to activation of the
Ras/Raf/mitogen-activated protein kinase (MAPK) and
phosphoinositide 3-kinase (PI3K)/Akt pathways, as well as many
others (14,15). EGFR is stimulated by the guanine
nucleotide-exchange factor, son of sevenless (SOS)-growth factor
receptor bound protein 2 (Grb2) complex. This activation of EGFR
leads to the activation of Ras. The Src homology 2 (SH2) domain of
Grb2 binds to autophosphorylation sites of EGFR, including Y1068,
and several other receptors. The SH3 domains of Grb2 bind to the
proline-rich C-terminal domain of SOS, a guanine nucleotide
exchange factor (GEF) for Ras. In this way, the SOS-Grb2
interaction plays a critical role in regulating the activation of
Ras (16,17). The small guanosine triphosphatase
protein Ras/MAPK signaling pathway is essential for the regulation
of a variety of biological processes, such as cell growth, cell
cycle, cell proliferation and cell senescence, all of which are
important for normal development (18). It consists of a core module of
three kinases comprising Raf, MEK and extracellular
signal-regulated kinase (ERK) that transmit signals downstream of
the small GTPase, Ras. GTP-loaded Ras triggers the sequential
activation of Raf, MEK and ERK to promote cell survival and various
cellular functions (19,20). Therefore, EGFR and related
proteins are attractive targets affecting cell proliferation.
The present study was carried out to confirm that
PYP1 (1–20) promotes the proliferation of IEC-6
cells and to determine the molecular mechanisms responsible for its
proliferative effects by investigating the involvement of the EGFR
signaling pathway. Our findings suggest that PYP1 (1–20)
affects EGFR-induced cell proliferation, a potential factor in
intestinal epithelial cell protection.
Materials and methods
Pyropia yezoensis peptide synthesis
The N-terminal 20 residues of PYP1 (1–20)
(A-L-E-G-G-K-S-S-G-G-G-E-A-T-R-D-P-E-P-T), designated as PYP1
(1–20), were synthesized by Peptron
(Daejeon, Korea) (7,21). The purification of PYP1 (1–20)
was performed using a Shimadzu Prominence high-performance liquid
chromatography (HPLC) apparatus and the software package Class-VP,
6.14 (Shimadzu, Kyoto, Japan), with a C18 column (Shiseido Capcell
Pak; Shiseido, Tokyo, Japan) in 0.1% TFA/water, a gradient of
10–70% acetonitrile in 0.1% TFA, a flow rate of 1 ml/min, and UV
detection at 220 nm. The molecular mass of PYP1 (1–20)
was confirmed to be 1,916 Da (matched with the sequence mass) by
mass spectrometric analysis (HP 1100 Series LC/MSD) (21).
Cell culture
The IEC-6 rat small intestinal epithelial cells
(ATCC CRL-1592; ATCC, Manassas, VA, USA) were cultured in
Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10%
fetal bovine serum (FBS; HyClone, Inc., South Logan, UT, USA), 100
U/ml penicillin and 100 mg/ml streptomycin, at a temperature of
37°C in a humidified atmosphere of 5% CO2. The cells
were cultured to 60% confluence in 100-mm dishes. The medium was
replaced every 2 days.
MTS assay
The effects of various PYP1 (1–20)
concentrations on cell proliferation were determined
colorimetrically after 24 h using the
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfonyl)-2H-tetrazolium
(MTS) assay with the Cell Titer 96 Aqueous One Solution reagent
(Promega, Madison, WI, USA). The cells were seeded in 96-well
plates at 1×104 cells/well. After 24 h of incubation,
the cells were maintained in serum-free medium (SFM) for 4 h. The
medium was replaced with fresh SFM containing PYP1 (1–20),
and the cells were incubated for an additional 24 h. The cells were
exposed to MTS assay solution at 37°C for 30 min, and the optical
density at 490 nm was measured using a microplate reader. The
OD490 values of the control cells were designated as
100%.
Western blot analysis
To prepare whole cell extracts, the IEC-6 cells in
100-mm dishes were cultured to 50–60% confluence and then incubated
in SFM for 4 h. Fresh SFM containing PYP1 (1–20)
(125, 250, 500 and 1,000 ng/ml) was added to the cells and
incubated another 24 h, after which the cells were washed with
phosphate-buffered saline (PBS) and suspended in extraction buffer
(50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 0.25% Na-deoxycholate, 1%
NP-40 and 1 mM EGTA) containing protease inhibitors (1 mM
Na3VO4, 1 μg/ml aprotinin, 1
μg/ml pepstatin, 1 μg/ml leupeptin, 1 mM NaF and 1 mM
PMSF) on ice. The extracts were centrifuged at 14,000 rpm for 10
min, and the supernatant was used in western blot analysis. Boiling
sample buffer (30 μg) was added to the total cell lysate,
and the samples were boiled for 10 min at 100°C. Proteins were
separated by 7.5–12.5% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) and transferred onto polyvinylidene
fluoride membranes (Millipore, Billerica, MA, USA). The membranes
were blocked for 1 h 40 min at room temperature in blocking buffer
[1% bovine serum albumin (BSA) in TBS-T]. The blots were probed
with primary antibodies [p-EGFR (sc-12351), EGFR (sc-03), Shc
(sc-1695), Grb2 (sc-255), SOS (sc-259), Ras (#3965), Raf (sc-227),
MEK (sc-219), p-ERK (sc-7383), ERK (sc-94), Cyclin D1 (sc-753),
Cyclin E (sc-481), Cdk2 (sc-163), Cdk4 (sc-601), Cdk6 (sc-177), pRb
(sc-16670), p21 (sc-397), p27 (sc-528), GAPDH (sc-25778) (1:1,000
and 1:2,000 in 1% BSA/TBS-T)] overnight at 4°C. The membranes were
then washed twice for 15 min in TBS-T. The secondary antibody was a
horseradish peroxidase (HRP)-conjugated goat anti-mouse or rabbit
antibody [goat anti-mouse IgG-HRP (sc-2031), goat anti-rat IgG-HRP
(sc-2032) (1:10,000 in 1% BSA/TBS-T)]. Signal bands were detected
using an enhanced chemiluminescence (ECL) western blotting kit
(Thermo Fisher Scientific, Inc., Rockford, IL, USA).
Reverse transcription-polymerase chain
reaction (RT-PCR)
The mRNA expression levels of specific genes were
evaluated by RT-PCR (22). The
IEC-6 cells were seeded into 100-mm dishes at a density of
2x104 cells/well and cultured for 24 h, after which the
medium was replaced with SFM containing PYP1 (1–20)
(125, 250, 500 and 1,000 ng/ml) for 24 h. Total RNA was isolated
from the cells using TRIzol reagent (Invitrogen Co., Carlsbad, CA,
USA) and converted to cDNA using oligo(dT) primers (Intron
Biotechnology Inc., Seongnam, Korea). For PCR amplification, the
cDNA and specific primers (Table
I) were added to 2X TOPsimple™ DyeMIX-nTaq (Enzynomics, Inc.,
Daejoen, Korea) and 0.1% diethylpyrocarbonate (DEPC) water. The
amplified products were analyzed on 1% agarose gels stained with
RedSafe™ nucleic acid staining solution (Intron Biotechnology,
Inc.).
 | Table IOligonucleotide sequences of the
primers used in RT-PCR. |
Table I
Oligonucleotide sequences of the
primers used in RT-PCR.
| Gene name | Primer sequences
(5′→3′) |
|---|
| EGFR | F:
CTC-ACG-CAG-TTG-GGC-ACT-TT
R: TCA-TGG-GCA-GCT-CCT-TCA-GT |
| SOS1 | F:
GCA-TCT-TAT-TGG-AAG-GAT-TT
R: CCT-CTC-AGG-TGA-GAC-TGC-TA |
| Grb2 | F:
CGG-GAT-CAT-GGA-AGC-CAT-GGC-CAA-A
R: CTA-GCT-AGC-TTA-GAC-GTT-CCG-GTT-CAC-TG |
| Ras | F:
CCC-GTC-CTC-ATG-TAC-TGG-TC
R: ATC-TTG-GAT-ACG-GCA-GGT-CA |
| Raf | F:
AAG-GCA-GTC-GTG-CAA-GCT-CA
R: GAT-GAT-GGC-AAA-CTC-ACG-GAT-TG |
| MEK | F:
CGA-TGG-ATC-CCC-CAA-GAA-GAA-GCC-GAC-G
R: CGA-TCT-CGA-GTT-AGA-CGC-CAG-CAG-CAT-G |
| GAPDH | F:
CAG-CCG-AGC-CAC-ATC-G
R: TGA-GGC-TGT-TGT-CAT-ACT-TCT-C |
Treatment with a MEK inhibitor
(PD98059)
The MEK inhibitor, PD98059 was obtained from Cell
Signaling Technology (Beverly, MA, USA) and stored as a 20 mM stock
solution at −20°C. The cells were pre-treated with 40 μM
PD98059 for 1 h and then incubated for 24 h as described above.
Cell cycle analysis
The cells were cultured in 6-well plates to 50–60%
confluence and treated with SFM or various doses of PYP1 (1–20)
(125, 250, 500 and 1,000 ng/ml) for 24 h. The cells were harvested
after trypsinization, washed with PBS, and treated with cold PI
solution (50 μg/ml) containing RNase A (0.1 mg/ml) in PBS
(pH 7.4) for 30 min in the dark. Flow cytometry was performed using
a FACSCalibur instrument (Becton-Dickinson, San Jose, CA, USA).
Statistical analysis
Multiple mean values were compared using analysis of
variance with SPSS (SPSS Inc., Chicago, IL, USA). Values are the
means ± SD. Different letters were used to indicate significant
values according to Duncan’s multiple range test.
Results
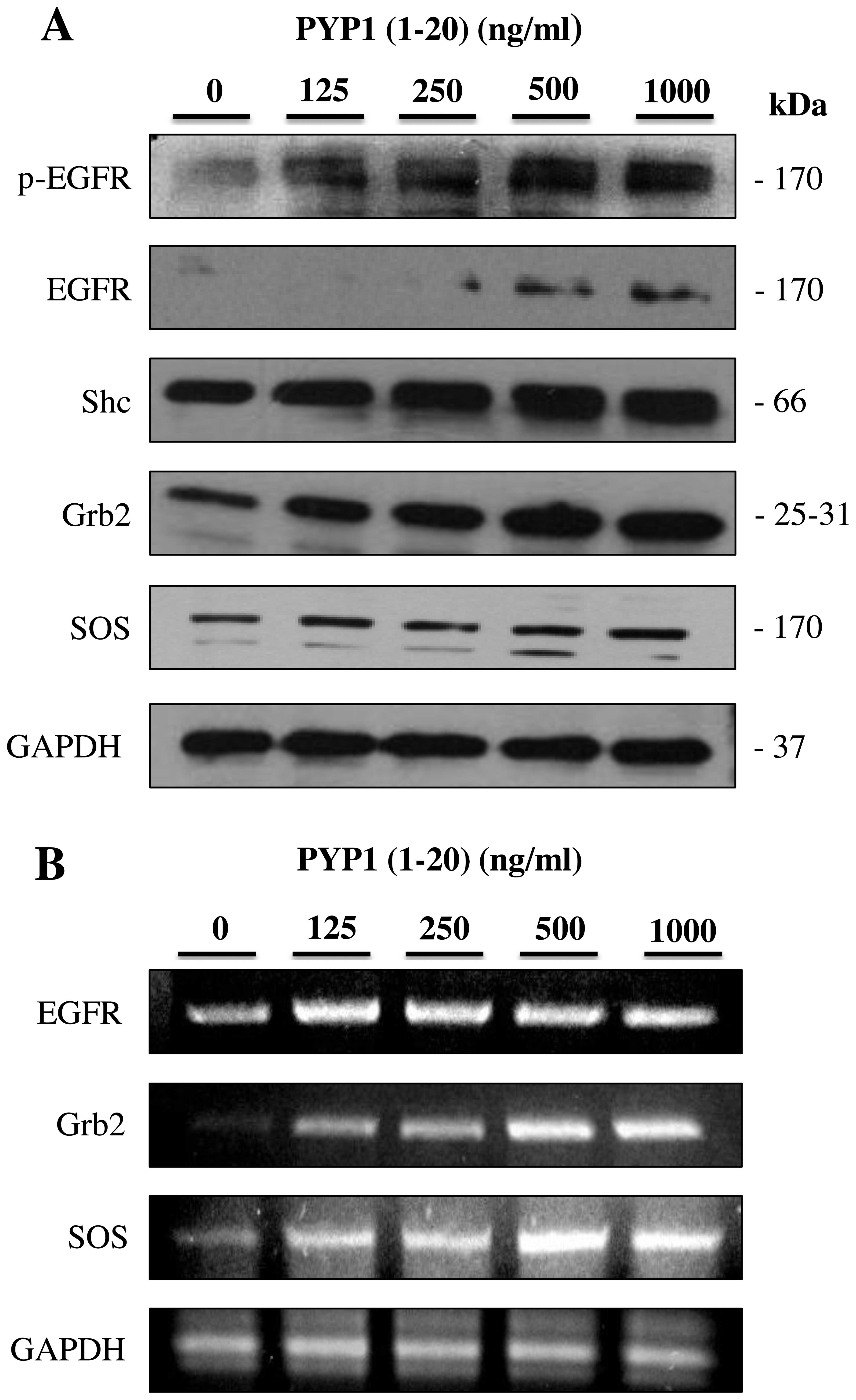
PYP1 (1–20)
increases the expression of EGFR and EGFR-related proteins
To investigate the mechanisms responsible for the
PYP1 (1–20)-induced proliferation of IEC-6
cells, we examined the effects of PYP1 (1–20)
on EGFR signaling-related proteins. The protein and mRNA expression
levels of phoshorylated (p-)EGFR, Shc, Grb2 and SOS in the IEC-6
cells treated with PYP1 (1–20)
(125, 250, 500 and 1,000 ng/ml) for 24 h were measured by western
blot analysis and RT-PCR. Treatment with PYP1 (1–20)
upregulated the protein (Fig. 1A)
and mRNA (Fig. 1B) expression
levels of p-EGFR, Shc, Grb2 and SOS in a dose-dependent manner.
These results indicate that PYP1 (1–20)
promotes the expression of EGFR signaling-related molecules.
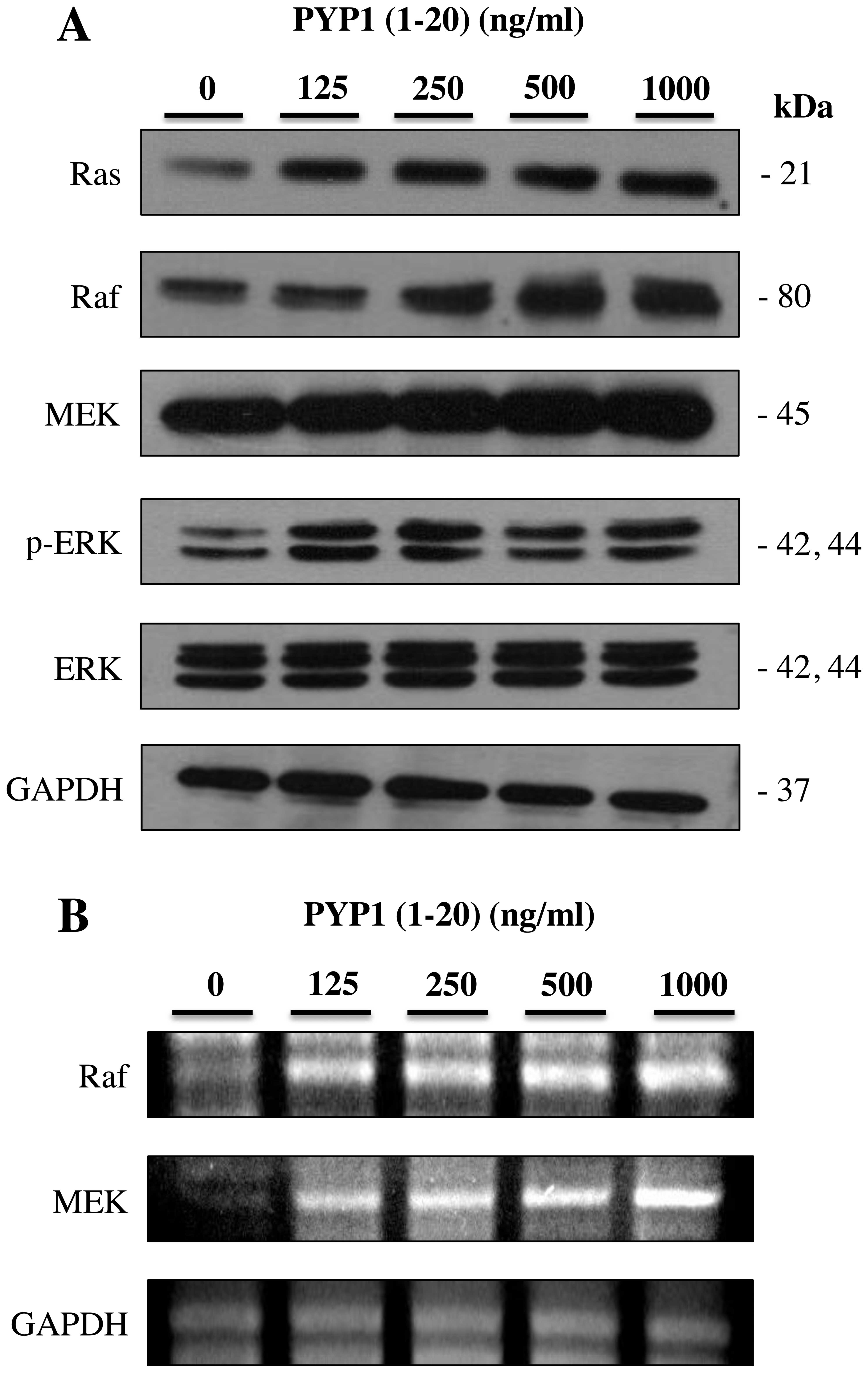
PYP1 (1–20)
induces the activation of the Ras-p42/p44 MAPK signaling
pathway
To further determine the downstream signals
regulated by EGFR activation, the protein and mRNA expression
levels of the Ras-p42/p44 signaling pathway members were measured
in the IEC-6 cells. The IEC-6 cells were treated with PYP1
(1–20) (125, 250, 500 and 1,000 ng/ml) for
24 h and then subjected to western blot analysis and RT-PCR.
Treatment with PYP1 (1–20) for 24 h resulted in increased
protein (Fig. 2A) and mRNA
(Fig. 2B) expression levels of
Ras, Raf, MEK and p-ERK compared with the untreated controls. These
results indicate that PYP1 (1–20)
activates the Ras-p42/p44 MAPK signaling pathway in IEC-6
cells.
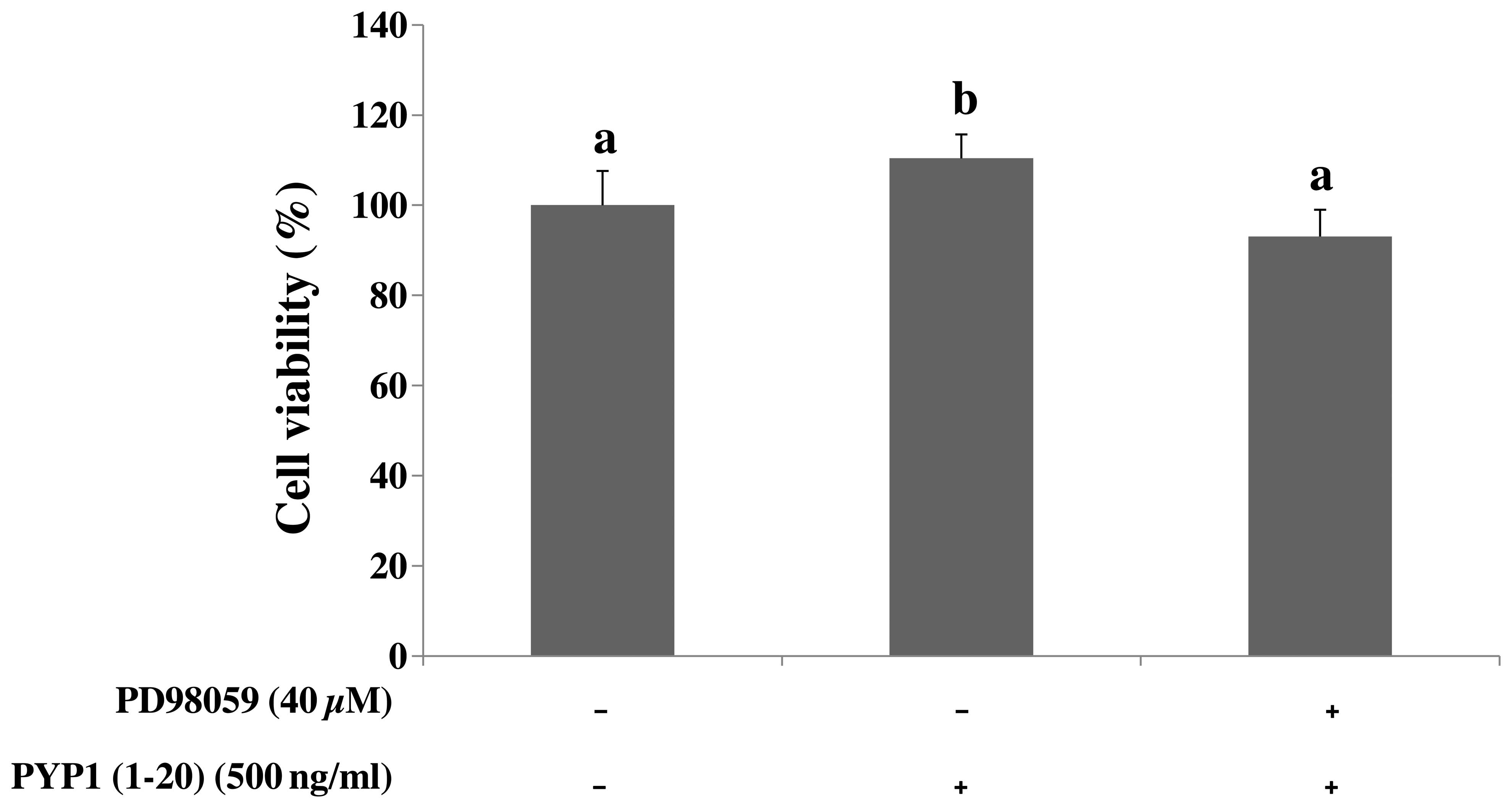
Pre-treatment with MEK inhibitor
suppresses the PYP1 (1–20)-induced cell proliferation
To investigate the suppressive effect of the MEK
inhibitor (PD98059) on PYP1 (1–20)-induced cell proliferation, an MTS
assay was performed. Pre-treatment with PD98059 for 1 h, followed
by the addition of PYP1 (1–20)
(500 ng/ml) for 24 h, induced a decrease in cell viability
identical to that of the controls (Fig. 3). Thus, EGFR is a target of PYP1
(1–20)-induced cell proliferation.
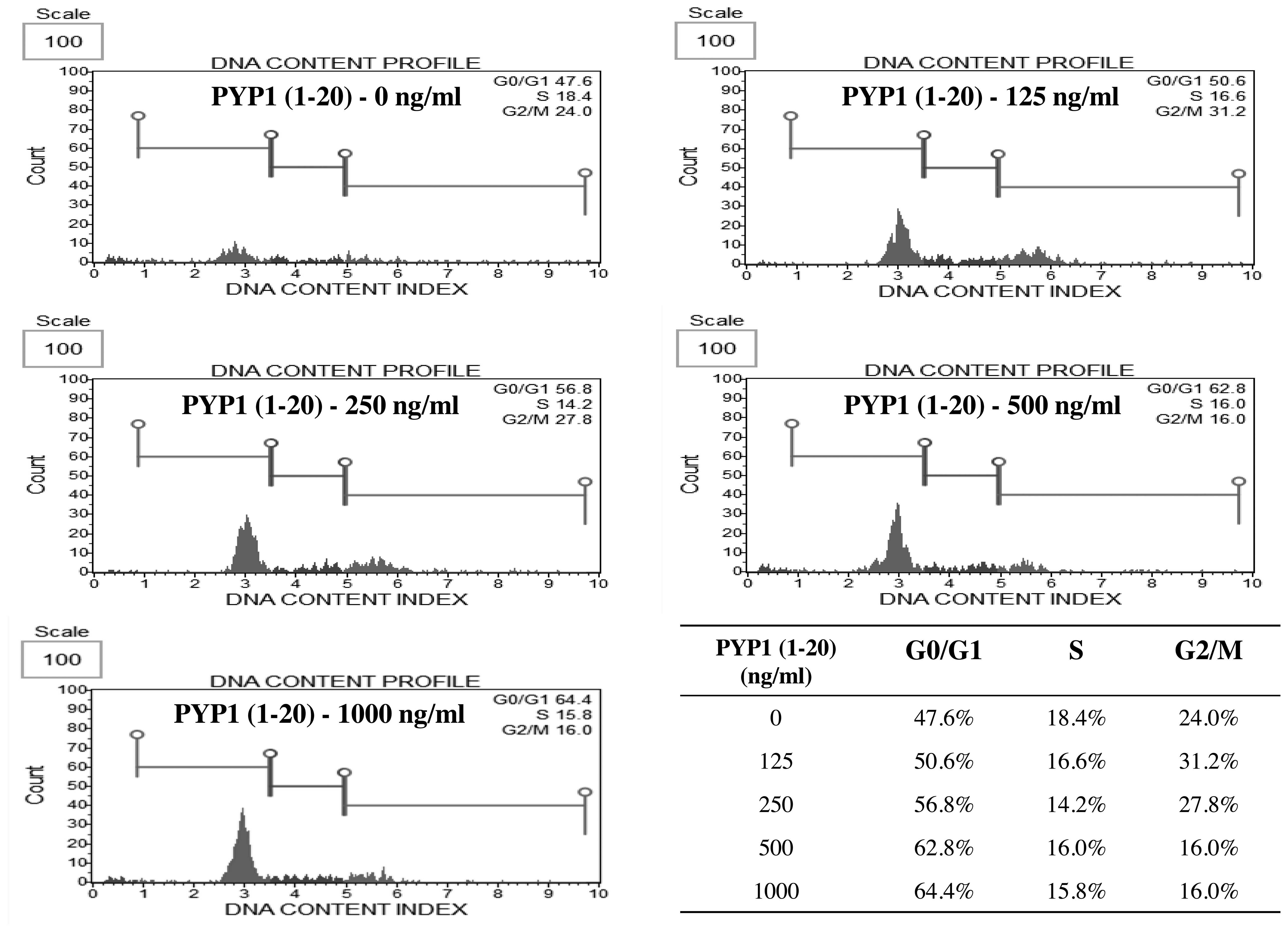
Effect of PYP1 (1–20)
on cell cycle progression
The percentage of cells at each phase of the cell
cycle was examined by flow cytometry. The cell cycle response was
determined in the cells treated with various concentration (125,
250, 500 and 1,000 ng/ml) of PYP1 (1–20)
for 24 h. Treatment with PYP1 (1–20)
increased the percentage of IEC-6 cells in the G1 phase (47.6,
50.6, 56.8, 62.8 and 64.4% following treatment with 0, 125, 250,
500 and 1,000 ng/ml PP-YE, respectively) in a dose-dependent manner
(Fig. 4). Therefore, treatment
with PYP1 (1–20) markedly increased the proportion of
cells in the G1 phase, suggesting that PYP1 (1–20)
promotes IEC-6 cell cycle progression.
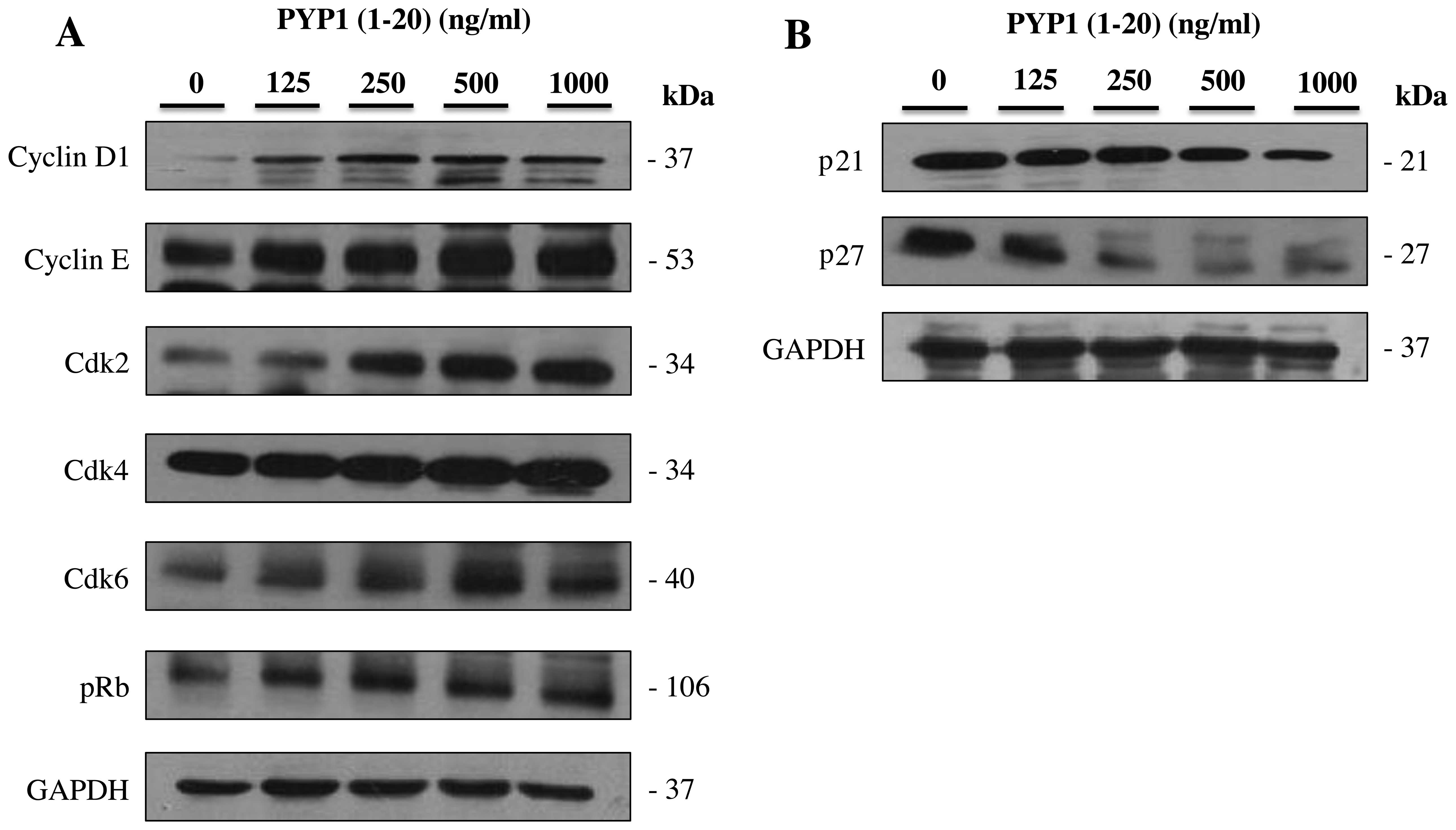
Effect of PYP1 (1–20)
on the expression of cell cycle-related proteins
The regulation of cell proliferation is defined as
the increase in the cell number resulting from the completion of
the cell cycle (23). To confirm
the cell proliferation mechanisms through which PYP1 (1–20)
promotes cell cycle progression, we examined the cell cycle-related
protein content. The expression levels of cyclin D1, cyclin E,
Cdk2, Cdk4, Cdk6, pRb, p21 and p27 were measured by western blot
analysis using specific antibodies. The IEC-6 cell cycle response
was determined following treatment with PYP1 (1–20)
at various concentrations (125, 250, 500 and 1,000 ng/ml). The
protein expression levels of cyclin D1, cyclin E, Cdk2, Cdk4, Cdk6
and pRb increased, whereas those of p21 and p27 decreased following
treatment with PYP1 (1–20) for 24 h (Fig. 5). These results suggest that PYP1
(1–20) promotes IEC-6 cell proliferation by
modulating the cell cycle-related proteins.
Discussion
Many types of seaweed have received a great deal of
attention from researchers in recent years for their high levels of
nutrients, such as proteins, minerals, vitamins and
polysaccharides. In particular, the anti-inflammatory and antitumor
activities of seaweed have been studied extenstively (24,25).
In a previous study, we demonstrated that treatment
with PYP1 (1–20) promotes the proliferation of IEC-6
cells through insulin-like growth factor I receptor (IGF-IR)
signaling pathways (7). In the
present study, we investigated whether PYP1 (1–20)
promotes IEC-6 cell proliferation and cell cycle progression
through the EGFR signaling pathway.
The activation of EGFR has been detected in many
different cell types, such as epithelial, nerve and mesenchymal
cells (14,15). EGFR activity induced by EGF
binding has been implicated in essential cellular functions,
including migration, differentiation, survival and proliferation
(12,23). Activated EGFR leads to the
activation of downstream signaling pathways, such as the
Ras-p42/p44 MAPK pathway. The Ras-p42/p44 MAPK signaling cascade is
a key mediator of growth factor-dependent cell survival,
proliferation and differentiation (24). The activation of Ras occurs mostly
via adaptor complex proteins containing Shc, Grb2 and SOS (16). In this study, PYP1 (1–20)
increased the protein and mRNA expression levels of EGFR, Shc, Grb2
and SOS (Fig. 1). Therefore, we
confirmed the effects of PYP1 (1–20)
on the Ras-p42/p44 MAPK signaling pathway. Ras/Raf/MEK/ERK
signaling commences at the cell surface, leading to the regulation
of gene expression within the cell nucleus (6). In this study, in accordance with
PYP1 (1–20)-induced cell proliferation, Ras,
Raf, MEK and ERK, important mediators that regulate cell survival,
growth and proliferation, were activated by exposure to PYP1
(1–20) (Fig.
2).
PYP1 (1–20)-induced cell proliferation was
examined by cell cycle analysis (Fig.
4). Treatment with PYP1 (1–20)
markedly increased the proportion of cells in the G0/G1 phase from
47.6 to 64.4%, suggesting that PYP1 (1–20)
promotes cell cycle progression (Fig.
4). The PYP1 (1–20)-induced cell cycle progression
resulted in cell proliferation and was related to the expression of
cell cycle-related proteins, such as cyclin and Cdk. The expression
levels of cyclin D1, cyclin E, Cdk2, Cdk4, Cdk6, pRb, p21 and p27
were measured by western blot analysis. The expression levels of
cyclin D1, cyclin E, Cdk2, Cdk4, Cdk6 and pRb were increased in a
dose-dependent manner, whereas the expression levels of p21 and p27
decreased in a dose-dependent manner (Fig. 5). Thus, the PYP1 (1–20)-induced cell cycle progression
resulted in IEC-6 cell proliferation.
In the present study, we demonstrate that PYP1
(1–20) mediates cell proliferation through
an EGFR signaling pathway in IEC-6 cells. These findings suggest
the significant role of EGFR in intestinal epithelial cell
proliferation, as well as the potential role of PYP1 (1–20)
as a bio-functional food with a proliferative effect on rat
intestinal epithelial cells.
Acknowledgments
This study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Education (grant no.
2012R1A6A1028677).
References
|
1
|
Zhao Y, Wu J, Shang D, Ning J, Zhai Y,
Sheng X and Ding H: Subcellular distribution and chemical forms of
cadmium in the edible seaweed, Porphyra yezoensis. Food Chem.
168:48–54. 2015. View Article : Google Scholar
|
|
2
|
van Netten C, Hoption Cann SA, Morley DR
and van Netten JP: Elemental and radioactive analysis of
commercially available seaweed. Sci Total Environ. 255:169–175.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dousip A, Matanjun P, Sulaiman MR, Tan TS,
Ooi YBH and Lim TP: Effect of seaweed mixture intake on plasma
lipid and antioxidant profile of hyperholesterolaemic rats. J Appl
Phycol. 26:999–1008. 2014. View Article : Google Scholar
|
|
4
|
Jiang LF: The polysaccharides from
Porphyra yezoensis suppress the denaturation of bighead carp
myofibrillar protein. Int J Biol Macromol. 68:18–20. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shin ES, Hwang HJ, Kim IH and Nam TJ: A
glycoprotein from Porphyra yezoensis produces anti-inflammatory
effects in liposaccharide-stimulated macrophages via the TLR4
signaling pathway. Int J Mol Med. 28:809–815. 2011.PubMed/NCBI
|
|
6
|
Qian L, Zhou Y and Ma JX: Hypolipidemic
effect of the polysaccharides from Porphyra yezoensis. Int J Biol
Macromol. 68:48–49. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee MK, Kim IH, Choi YH and Nam TJ: A
peptide from Porphyra yezoensis stimulates the proliferation of
IEC-6 cells by activating the insulin-like growth factor I receptor
signaling pathway. Int J Mol Med. 35:533–538. 2015.
|
|
8
|
Song SH, Kim IH and Nam TJ: Effect of a
hot water extract of Chlorella vulgaris on proliferation of IEC-6
cells. Int J Mol Med. 29:741–746. 2012.PubMed/NCBI
|
|
9
|
Schlessinger J: Cell signaling by receptor
tyrosine kinases. Cell. 103:211–225. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hubbard SR and Till JH: Protein tyrosine
kinase structure and function. Annu Rev Biochem. 69:373–398. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ostman A and Böhmer FD: Regulation of
receptor tyrosine kinase signaling by protein tyrosine
phosphatases. Trends Cell Biol. 11:258–266. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sordella R, Bell DW, Haber DA and
Settleman J: Gefitinib-sensitizing EGFR mutations in lung cancer
activate anti-apoptotic pathways. Science. 305:1163–1167. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
DeYulia GJ Jr and Cárcamo JM: EGF
receptor-ligand interaction generates extracellular hydrogen
peroxide that inhibits EGFR-associated protein tyrosine
phosphatases. Biochem Biophys Res Commun. 334:38–42. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu C, Hale J, Ritchie K, Prasad NK and
Irudayaraj J: Receptor overexpression or inhibition alters cell
surface dynamics of EGF-EGFR interaction: New insights from
real-time single molecule analysis. Biochem Biophys Res Commun.
378:376–382. 2009. View Article : Google Scholar
|
|
15
|
Rajalingam K, Wunder C, Brinkmann V,
Churin Y, Hekman M, Sievers C, Rapp UR and Rudel T: Prohibitin is
required for Ras-induced Raf-MEK-ERK activation and epithelial cell
migration. Nat Cell Biol. 7:837–843. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Holt KH, Waters SB, Okada S, Yamauchi K,
Decker SJ, Saltiel AR, Motto DG, Koretzky GA and Pessin JE:
Epidermal growth factor receptor targeting prevents uncoupling of
the Grb2-SOS complex. J Biol Chem. 271:8300–8306. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Buday L, Egan SE, Rodriguez Viciana P,
Cantrell DA and Downward J: A complex of Grb2 adaptor protein, Sos
exchange factor, and a 36-kDa membrane-bound tyrosine
phosphoprotein is implicated in ras activation in T cells. J Biol
Chem. 269:9019–9023. 1994.PubMed/NCBI
|
|
18
|
Tidyman WE and Rauen KA: The RASopathies:
Developmental syndromes of Ras/MAPK pathway dysregulation. Curr
Opin Genet Dev. 19:230–236. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ashton-Beaucage D, Udell CM, Gendron P, et
al: A functional screen reveals an extensive layer of
transcriptional and splicing control underlying RAS/MAPK signaling
in Drosophila. PLoS Biol. 12:e10018092014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bonni A, Brunet A, West AE, Datta SR,
Takasu MA and Greenberg ME: Cell survival promoted by the Ras-MAPK
signaling pathway by transcription-dependent and -independent
mechanisms. Science. 286:1358–1362. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Choi YH, Yamaguchi K, Oda T and Nam TJ:
Chemical and mass spectrometry characterization of the red alga
Pyropia yezoensis chemoprotective protein (PYP): Protective
activity of the N-terminal fragment of PYP1 against
acetaminophen-induced cell death in Chang liver cells. Int J Mol
Med. 35:271–276. 2015.
|
|
22
|
Kim YM, Kim IH and Nam TJ: Capsosiphon
fulvescens glycoprotein reduces AGS gastric cancer cell migration
by downregulating transforming growth factor-β1 and integrin
expression. Int J Oncol. 43:1059–1065. 2013.PubMed/NCBI
|
|
23
|
Pardee AB: G1 events and regulation of
cell proliferation. Science. 246:603–608. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Go H, Hwang HJ and Nam TJ: Polysaccharides
from Capsosiphon fulvescens stimulate the growth of IEC-6 Cells by
activating the MAPK signaling pathway. Mar Biotechnol (NY).
13:433–440. 2011. View Article : Google Scholar
|
|
25
|
Hwang HJ, Kwon MJ, Kim IH and Nam TJ:
Chemoprotective effects of a protein from the red algae Porphyra
yezoensis on acetaminophen-induced liver injury in rats. Phytother
Res. 22:1149–1153. 2008. View Article : Google Scholar : PubMed/NCBI
|