Introduction
Intervertebral disc degeneration (IDD) is
characterized by damage of the disc structure, progressive loss of
water content and proteoglycan in the extracellular matrix (ECM)
(1). IDD is considered a
predominant source of spine-related diseases and chronic lower back
pain, which causes a major economic and social burden affecting
millions of people worldwide (2).
Current treatment options for IDD, including discectomy (3), intradiscal electrothermal therapy
(4) and arthroplasty (5), only address the clinical symptoms of
IDD and remain limited with unpredictable outcomes (6). Therefore, understanding the
pathophysiology and molecular mechanism underlying IDD appears to
be imperative for diagnosis and developing novel therapeutic
approaches.
The intervertebral disc consists of three
morphologically distinct regions, which are the nucleus pulposus
(NP), annulus fibrosis and cartilaginous endplates (7). The central NP is a gelatinous matrix
that is composed of large aggregating proteoglycans and a loose
network of collagen. The peripheral annulus fibrosis encases the
nucleus pulposus and is rich in type I collagen. The cartilaginous
endplates contain the peripheral vasculature, which can nourish the
disc (1). The main morphological
manifestations of IDD are vertebral instability, disc herniation
and spinal stenosis (8).
Significant changes in morphology, structure and composition are
accompanied by specific changes in the disc with aging and
degeneration, including alteration of the elastic modulus and
swelling pressure of the nucleus pulposus (9). Recently, histological grading
schemes for assessing human IDD have been developed (10,11). A 5-level grading system for lumbar
disc degeneration, proposed by Pfirrmann et al (11), was developed according to
T2-weighted magnetic resonance images. Additionally, the Thompson
scoring system scores disc degeneration over the spectrum from
healthier discs (grades I and II) to advanced degeneration (grade
V, the most advanced stage of degeneration) (10). However, there is a lack of studies
on the associations of the grades of disc degeneration.
IDD is a complex multi-factorial process (12). Several factors, such as
biological, mechanical and genetic factors, are widely considered
as significant contributors to the disc degenerative process
(1). For example, Bachmeier et
al (13) identified that
matrix metalloproteinase-3 (MMP-3), which was essential for matrix
degeneration, had an essential role in lumbar disc herniation and
degeneration. Additionally, Takahashi et al (14) revealed that the polymorphism 5A
allele, which often occurs in the promoter region of the gene that
regulates MMP-3 expression, was a possible risk factor for
accelerated IDD in the elderly. In addition, Pratsinis et al
(15) reported that
platelet-derived growth factor, insulin-like growth factor-I and
basic fibroblast growth factor could stimulate the proliferation of
intervertebral disc cells via the activation of the
extracellular-signal regulated kinase and Akt signaling pathways.
By contrast, microRNAs (miRNAs) that can regulate RNA degradation
or repression of translation have been identified as key regulators
in numerous biological processes (16). Recently, Yu et al (17) demonstrated that miR-10b could
promote nucleus pulposus cell proliferation by targeting homeobox
D10 through the ras homolog family member C-Akt pathway in IDD.
However, the molecular mechanism of IDD is not fully understood and
further investigations are required.
Recently, Chen et al (18) reported that mitogen-activated
protein (MAP) kinase kinase 6 (MAP2K6) and Ras homologous-related
BTB domain containing 2 may have significant roles in the
progression of grade III and IV disc degeneration, respectively.
Additionally, Tang et al (8) demonstrated that differentially
expressed genes (DEGs) identified in degenerative disc tissue
samples were mainly associated with transforming growth factor-β
and the ECM. Furthermore, Wang et al (19) reported that deregulated miR-155
could promote Fas-mediated apoptosis in human IDD by targeting
Fas-associated death domain-containing protein (FADD) and
caspase-3. In the present study, 3 microarray datasets were
integrated and micro-array analysis was used to identify the DEGs,
grade-specific genes and differentially expressed miRNAs in the IDD
samples compared with their corresponding controls. In addition,
comprehensive bioinformatics was used to analyze the significant
functions and pathways, and to analyze the miRNA-target
gene-pathway interaction associations. The study aimed to identify
the significant genes and miRNA alternations in the progression of
IDD and gain more insights into the molecular mechanisms of disc
degeneration. Understanding these mechanisms can aid in exploring
an appropriate molecular target and developing new therapeutic
methods for IDD.
Materials and methods
Microarray data
Microarray datasets of GSE19943 (19), GSE15227 (20) and GSE34095 (21) were downloaded from the National
Center of Biotechnology Information (NCBI), Gene Expression Omnibus
(GEO) database (http://www.ncbi.nlm.nih.gov/geo/). The miRNA
expression profile GSE19943 included 3 degenerative NP samples that
were collected from patients with IDD and 3 NP controls derived
from patients with scoliosis, which was based on the platform
GPL9946 Exiqon human miRCURY LNA™ microRNA Array V11.0. The
GSE15227 dataset included 5 grade II, 7 grade III, and 3 grade IV
discs from patients with herniated discs and IDD that were scored
using the Thompson scoring system, which was based on the platform
GPL1352 [U133_X3P] Affymetrix Human X3P array. The gene expression
profile GSE34095 included 3 degenerative NP samples that were
harvested from elderly patients with IDD and 3 non-degenerative
samples derived from younger patients with adolescent idiopathic
scoliosis as controls, which was based on the platform GPL96
[HG-U133A] Affymetrix Human Genome U133A array.
DEGs and differentially expressed miRNAs
screening
The t-test in the limma package (22) was used to identify DEGs in
degenerative samples compared with controls in GSE34095, DEGs in 7
grade III and 3 grade IV samples compared with 5 grade II samples
in GSE15227, and to identify differentially expressed miRNAs in 3
degenerated samples compared with controls in GSE19943. The false
discovery rate (FDR) was applied to perform multiple testing
corrections using the Benjamini and Hochberg method (23). The threshold for the DEGs and the
differentially expressed miRNAs were set as FDR <0.05.
miRNA-target gene and KEGG pathway data
collection
Human miRNAs and their associated targets were
downloaded from 3 high-quality online miRNA reference databases,
which store manual collections of experimentally supported miRNA
targets, miRecords (24),
miRTarBase (25) and TarBase 6.0
(26). In addition, all pathways
and associated genes were downloaded from the Kyoto Encyclopedia of
Genes and Genomes (KEGG) database (27).
Grade-specific network construction
DEGs identified in grade III and IV samples were
imported into Cytoscape software (28) to create protein-protein
interaction (PPI) network visualizations, respectively. The source
of the interaction network database was the Search Tool of the
Retrieval of Interacting Genes 9.0 database, which is a
comprehensive database containing functional links between proteins
that are experimentally derived, as well as links predicted by text
mining and comparative genomics (29). The threshold was set as confidence
score, 0.4. Furthermore, the specific genes that were only enriched
in one of the grade-specific networks and the common genes of the 2
grade-specific networks were extracted. The parameters describing
the network topology were calculated using the Cytoscape plugin
Network Analyzer (30). Logistic
regression analysis (31) was
conducted by SPSS 19.0 (IBM, Corp., Armonk, NY, USA) to select the
most noteworthy network features.
Cluster analysis
To test the availability of these 2 grade-specific
networks to reveal different stages of IDD progression, clustering
analysis was performed on all genes in the networks and the top 20%
nodes that had a higher degree in the network using Cytoscape
plugin Network Analyzer (30),
respectively. The results are represented by heat-maps.
GO and pathway enrichment analysis of
DEGs and the specific genes
GO and KEGG enrichment analysis were performed for
the specific genes using the Database for Annotation, Visualization
and Integrated Discovery (DAVID) online tool (32). The threshold was set as
P<0.05.
Furthermore, KEGG enrichment analyses were also
performed for DEGs that were identified based on GSE34095 using
DAVID. The threshold was set as P<0.05.
Measurement of miRNA-pathway
interactions
With the identified differentially expressed miRNAs,
as well as their target genes and the genes enriched in the KEGG
pathways, Fisher's exact test (33) was used to assess the corresponding
significance of the miRNA pathway.
Fisher's exact test was based on the hypergeometric
distribution to combine the results of the proportion of the miRNA
target gene in the functional gene set and the proportion of miRNA
target gene in the whole genome. A P-value for the null hypothesis
of the Fisher's exact test was examined as the genes that belong to
the target genes of miRNAs and the genes that belong to the
functional genes enriched.
The resulting P-value depicts the probability that
the examined pathway is significantly enriched with gene targets of
the selected miRNAs, the probability that at least x
functional genes are enriched in the K target genes of
miRNA. The P-value can be expressed as:
Where N is the total number of genes, M is the number
of genes in the functional genes set and K is the number of
target genes of miRNA.
Results
Screening of DEGs and differentially
expressed miRNAs
For GSE15227, 846 and 1,137 DEGs were identified in
grade III and IV discs, respectively. For GSE34095, a total of 961
DEGs were identified in the IDD samples compared with the controls.
Furthermore, for GSE19943, 77 differentially expressed miRNAs were
identified in the degenerative NP samples compared with the
controls.
Grade-specific network construction
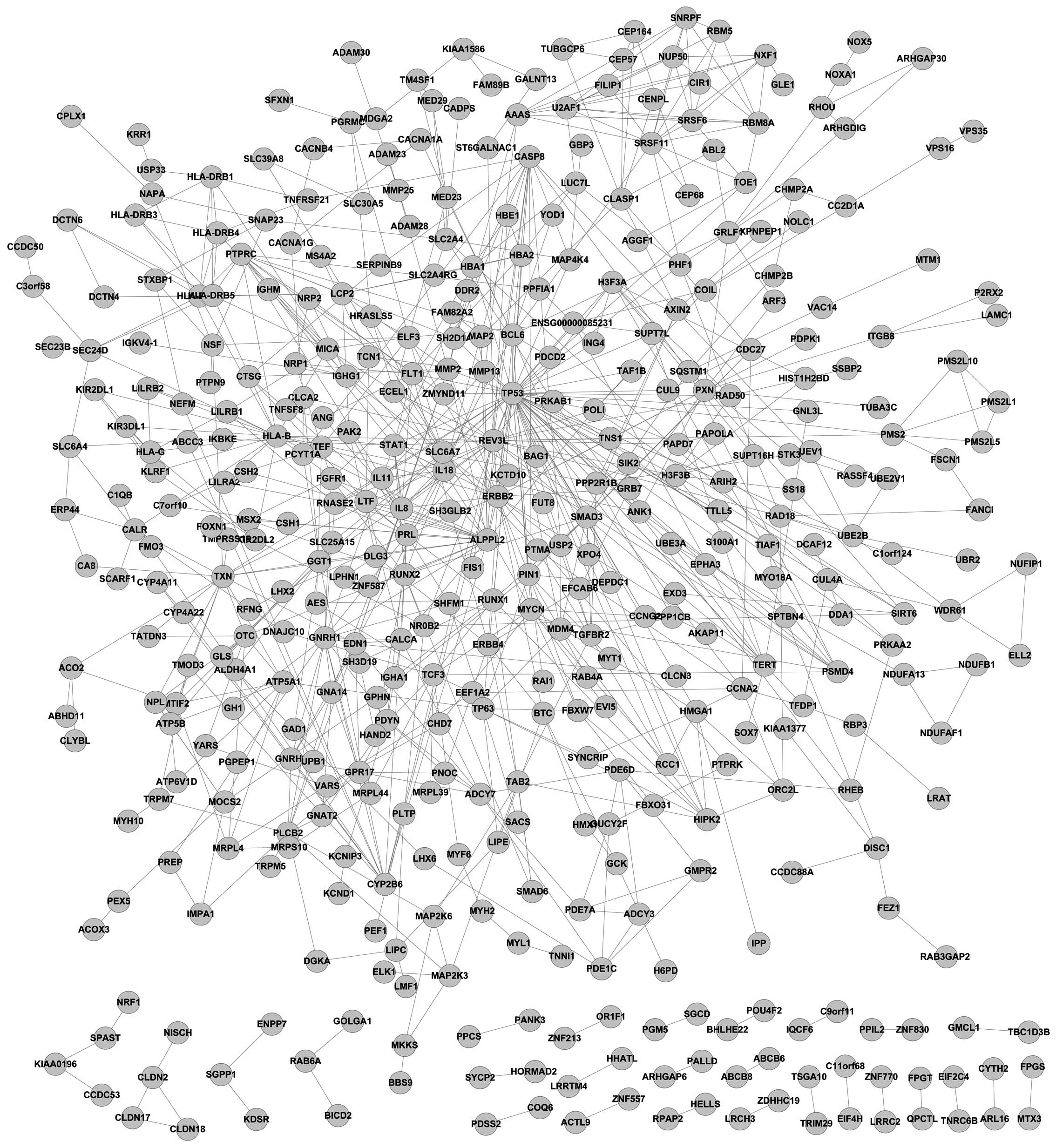
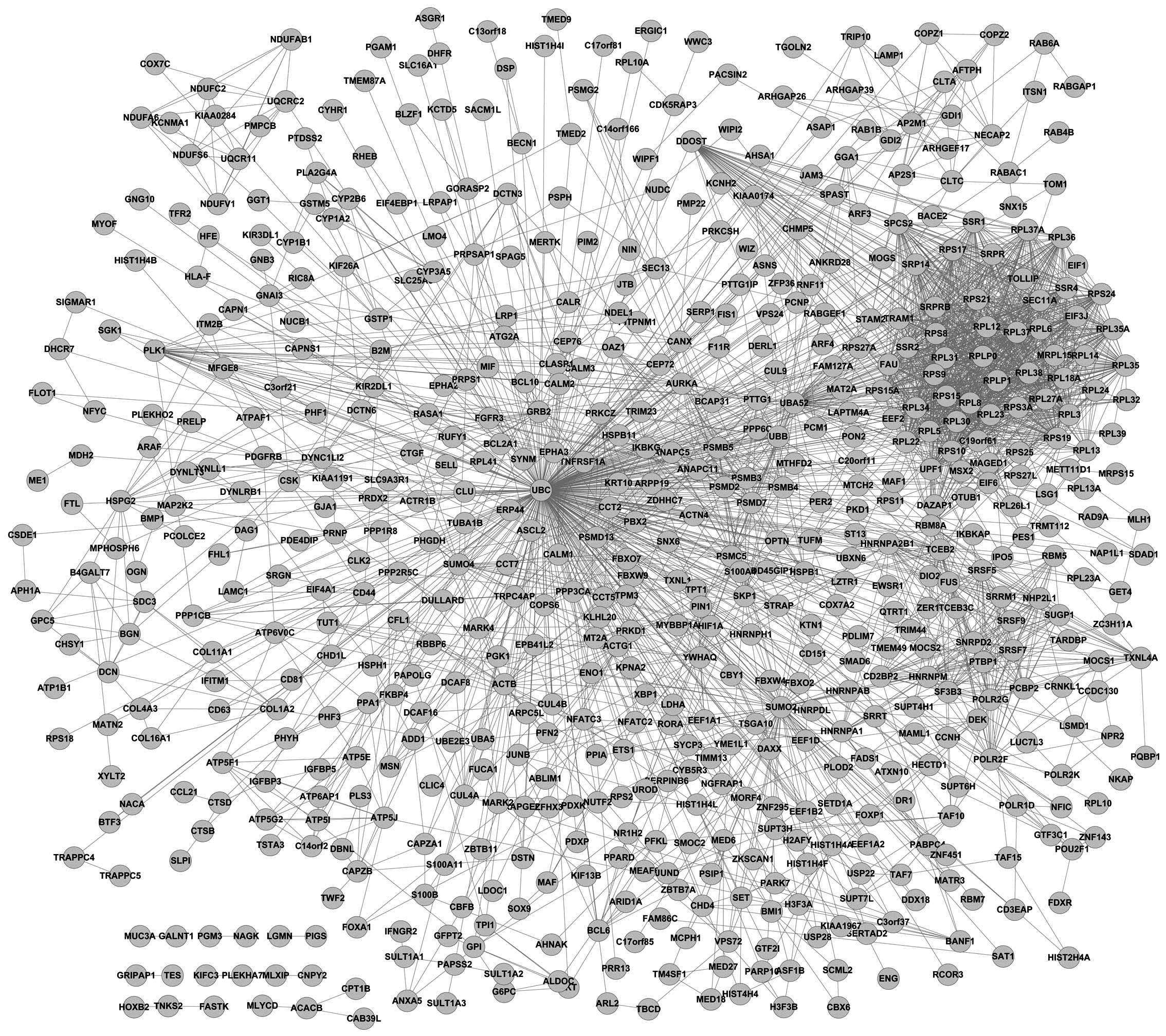
The grade-specific networks are shown in Figs. 1 and 2. In the present study, 746 grade
III-specific genes and 964 grade IV-specific genes were identified.
Tumor protein p53 (TP53) was a hub gene in the grade
III-specific network and ubiquitin C (UBC) was identified to
be a hub gene in the grade IV-specific network. Additionally, there
were 78 common genes in these 2 grade-specific networks. Network
topology analysis showed that a total of 16 network features were
identified. In addition, statistical regression analysis revealed
that 6 significant network features were obtained, as shown in
Table I, including average
shortest path length, betweenness centrality, closeness centrality,
neighborhood connectivity, radiality and stress.
 | Table ILogistic regression. |
Table I
Logistic regression.
| Feature | P-value | Exp (B) |
|---|
| Average shortest
path length | <0.0001 | 0.050 |
| Betweenness
centrality | <0.0001 | <0.0001 |
| Closeness
centrality | <0.0001 | <0.0001 |
| Neighborhood
connectivity | 0.030 | 1.015 |
| Radiality | 0.036 | <0.0001 |
| Stress | <0.0001 | 1.010 |
| Constant | 0.001 |
1.08×1011 |
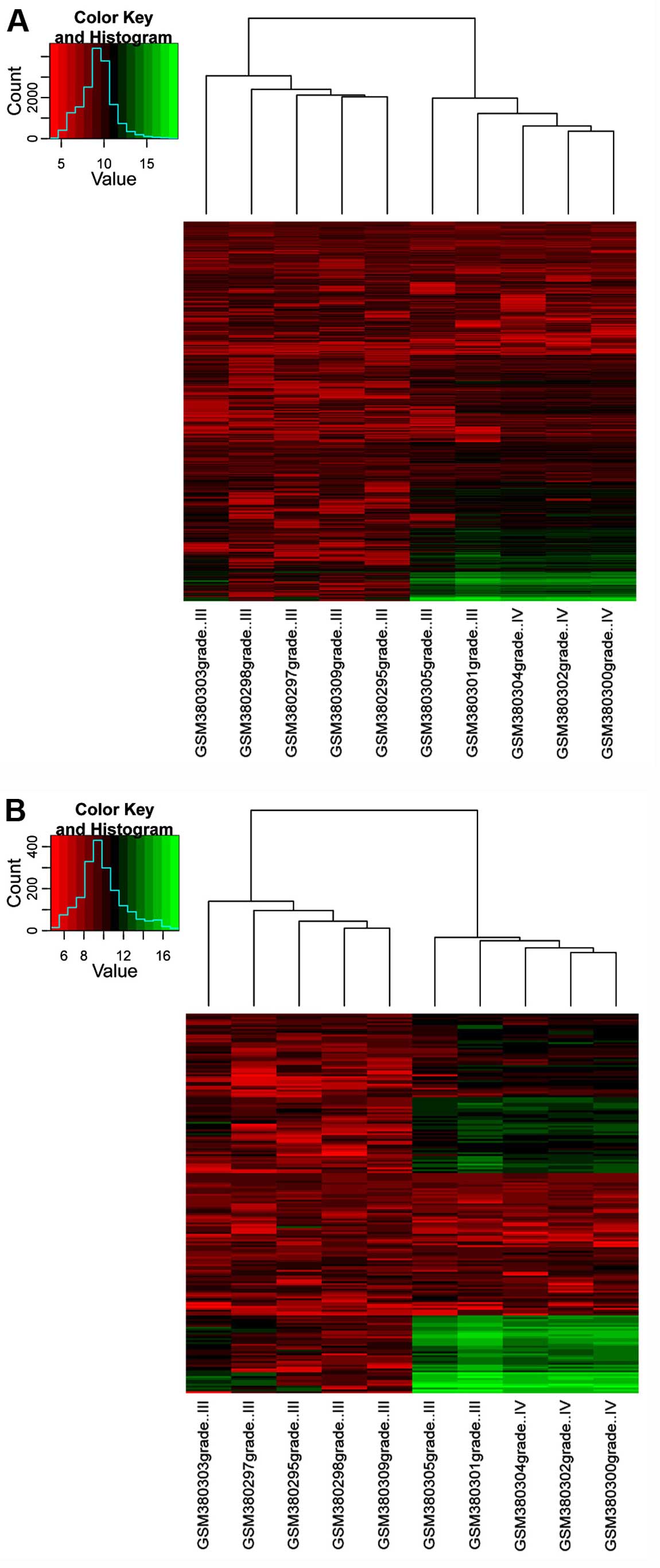
Clustering analysis
The clustering analysis was used to group the genes
and samples on the basis of similarities of gene expression. The
results of clustering analysis using all the specific genes in
these 2 networks are shown in Fig.
3A. The result showed that 2 grade III samples (GSM380301 and
GSM380305) were grouped into the region of grade IV samples;
however, the correlation was lower compared to the correlation of
the grade IV samples. By contrast, the results of clustering
analysis using the top 20% genes with a higher degree in these 2
networks are shown in Fig. 3B.
Consistently, the 2 grade III samples, GSM380301 and GSM380305,
were grouped into the region of grade IV samples.
GO and pathway enrichment analysis of
specific genes
GO and pathway analysis indicated that grade III-
and IV-specific genes were significantly enriched in different GO
terms and KEGG pathways. A total of 23 GO terms for grade
III-specific genes were enriched, including angiogenesis, adult
walking behavior and positive regulation of kinase activity
(Table II). GO terms for grade
IV-specific genes were mainly associated with the regulation of
ubiquitin-protein ligase activity, such as positive regulation of
ubiquitin-protein ligase activity, and regulation of protein
ubiquitination (Table III). In
addition, 4 KEGG pathways for grade III-specific genes were
significantly enriched, including Alzheimer's disease, oxidative
phosphorylation, Huntington's disease and Parkinson's disease. A
total of 5 KEGG pathways for grade IV-specific genes were
significantly enriched, including viral myocarditis,
graft-versus-host disease, type I diabetes mellitus, allograft
rejection and cell adhesion molecules (Table IV).
| Table IIGO terms of grade III-specific
genes. |
Table II
GO terms of grade III-specific
genes.
| GO term | P-value |
|---|
| Angiogenesis | 0.007 |
| Adult walking
behavior | 0.007 |
| Retina development
in camera-type eye | 0.008 |
| Activation of
protein kinase activity | 0.008 |
| Response to DNA
damage stimulus | 0.008 |
| Negative regulation
of biosynthetic process | 0.009 |
| Negative regulation
of cellular biosynthetic process | 0.011 |
| Negative regulation
of macromolecule biosynthetic process | 0.014 |
| Positive regulation
of protein kinase activity | 0.015 |
| Positive regulation
of kinase activity | 0.020 |
| Cytokine
secretion | 0.024 |
| Negative regulation
of gene expression | 0.024 |
| Negative regulation
of transcription | 0.025 |
| Positive regulation
of transferase activity | 0.026 |
| Negative regulation
of nucleobase, nucleoside, nucleotide and nucleic acid metabolic
process | 0.029 |
| Negative regulation
of nitrogen compound metabolic process | 0.034 |
| Vasculature
development | 0.037 |
| Patterning of blood
vessels | 0.038 |
| Tube
morphogenesis | 0.039 |
| Regulation of
protein kinase activity | 0.043 |
| Adult locomotory
behavior | 0.044 |
| Actin filament
organization | 0.044 |
| Cell-substrate
junction assembly | 0.048 |
| Negative regulation
of macromolecule metabolic process | 0.050 |
| Table IIIGO terms of grade IV-specific
genes. |
Table III
GO terms of grade IV-specific
genes.
| GO terms | P-value |
|---|
| Cellular
macromolecule localization | 0.036 |
| Cellular protein
localization | 0.030 |
| Establishment of
protein localization | 0.026 |
| Intracellular
protein transport | 0.038 |
| Positive regulation
of ligase activity | 0.031 |
| Positive regulation
of protein ubiquitination | 0.039 |
| Positive regulation
of ubiquitin-protein ligase activity | 0.019 |
| Positive regulation
of ubiquitin-protein ligase activity during mitotic cell cycle | 0.013 |
| Protein
transport | 0.036 |
| Regulation of
ligase activity | 0.025 |
| Regulation of
protein ubiquitination | 0.022 |
| Regulation of
ubiquitin-protein ligase activity | 0.015 |
| Regulation of
ubiquitin-protein ligase activity during mitotic cell cycle | 0.022 |
| Table IVPathway enrichment of grade III- and
IV-specific genes. |
Table IV
Pathway enrichment of grade III- and
IV-specific genes.
| Pathway | P-value |
|---|
| Grade III |
| hsa05010:
Alzheimer's disease | 0.009 |
| hsa00190:
Oxidative phosphorylation | 0.011 |
| hsa05016:
Huntington's disease | 0.012 |
| hsa05012:
Parkinson's disease | 0.021 |
| Grade IV |
| hsa05416: Viral
myocarditis |
9.37×10−4 |
| hsa05332:
Graft-versus-host disease | 0.012 |
| hsa04940: Type I
diabetes mellitus | 0.017 |
| hsa05330:
Allograft rejection | 0.040 |
| hsa04514: Cell
adhesion molecules | 0.049 |
miRNA-pathway-genes complex regulatory
associations analysis
Subsequent to combining miRecords, miRTarBase and
TarBase 6.0 databases, 5,489 miRNA-target pairs were collected in
the study, including 482 miRNAs and 2,331 target genes.
miRNA-pathway interactions analysis showed that there were 217
interaction pairs involved in 35 significant pathways. Furthermore,
the pathway enrichment analysis for DEGs in the degenerative NP
samples compared with the controls showed that 21 KEGG pathways
were significantly enriched. There were 6 pathways also regulated
by miRNAs, as shown in Table V, including focal adhesion, the ErbB
signaling pathway, the calcium signaling pathway, the MAPK
signaling pathway, apoptosis and pathways in cancer. Additionally,
only miR-129-5p was involved in the calcium signaling pathway,
apoptosis and pathways in cancer.
Discussion
In the present study, microarray analysis showed
that a total of 846 and 1,137 DEGs were identified in grade III and
IV discs, respectively. In addition, 961 DEGs were identified in
the IDD samples in GSE34095 and 77 differentially expressed miRNAs
were identified in the IDD samples. TP53 was a hub gene in
the grade III-specific network and UBC was identified as a
hub gene in grade IV-specific network. Six significant features
were identified by grade-specific network topology analysis.
Differentially expressed miRNAs were identified to participate in
35 pathways, among which 6 pathways were significantly enriched by
DEGs, including apoptosis.
The general concepts, such as centrality,
communicability and betweenness, quantify the important features in
a network (34). Estrada
(35) demonstrated that subgraph
centrality could be applied to the identification of essential
proteins in PPI networks. Additionally, the threshold for inclusion
was P<0.05 (significant) in the logistic regression model
(36). In line with the previous
study, the present study showed that 6 network features (average
shortest path length, betweenness centrality, closeness centrality,
neighborhood connectivity, radiality and stress) had significant
differences in grade III- and IV-specific networks (Table I). On this basis, these 6 features
may have a significant association with the grade of discs
degeneration and could be useful for assessing the IDD grade.
TP53 is a tumor-suppressor protein containing DNA
binding, transcriptional activation and oligomerization domains
(37). Vaghefi et al
(38) revealed that deacetylation
of p53 was a nerve growth factor (NGF)-dependent post-translational
mechanism of p53 activation. Additionally, Richardson et al
(39) reported that NGF had
increased expression in the painful degenerate intervertebral disc.
In addition, Liu et al (40) demonstrated that VEGF and
p53 were working simultaneously in neovascularization and
infiltration, and accelerating rat IDD progression. In line with
the previous study, TP53 was identified as a hub gene in the grade
III-specific network, which was derived from the microarray data of
GSE15227. Furthermore, specimens of GSE15227 are annulus fibrosus
cells in cultures from patients with herniated discs and IDD. Thus,
we hypothesized that TP53 may participate in neovascularization and
infiltration in annulus fibrosus in IDD progression, which could
also help the IDD grading system.
Additionally, the present study identified that the
GO terms for grade IV-specific genes were mainly associated with
the regulation of ubiquitin-protein ligase activity and UBC
was a hub gene in the grade IV-specific network derived from the
data of GSE15227. UBC encodes a poly-ubiquitin precursor that can
conjugate with different residues and lead to various effects
within a cell (41). Yew
(42) demonstrated that G1- and
S-phase events in vertebrates, which were essential for cell
proliferation, were specifically mediated by a multitude of
regulators associated with ubiquitination, of which UBC was
required. Furthermore, Gruber et al (43) reported that cells in IDD lost
their ability to proliferate and were subject to senescence. Thus,
we hypothesized that UBC may have a crucial role in
inhibiting cell proliferation of annulus fibrosus in IDD
progression and could be used for grading disc degeneration.
Furthermore, the present study showed that
miR-129-5p could participate in different pathways, including
apoptosis. The study by Kohyama et al (44) reported that induction of the
apoptosis of disc cells had an important role in the pathogenesis
of disc degeneration. In addition, Li et al (45) revealed that downregulation of
miR-129-5p could inhibit cell growth and induce apoptosis in
laryngeal squamous cell carcinoma by targeting adenomatous
polyposis coli. In the present study, the miRNA-pathway interaction
analysis was based on the information of differentially expressed
miRNAs derived from GSE19943 and the pathway enrichment analysis
for DEGs in degenerative NP samples was derived from GSE34095.
Additionally, tissues of GSE19943 and GSE34095 are NP from patients
with IDD and scoliosis. Furthermore, 6 overlapping pathways were
identified according to the results of miRNA-pathway interactions
analysis and pathway enrichment analysis for DEGs, including
apoptosis. Therefore, we hypothesized that miR-129-5p may have a
crucial role in modulating cell apoptosis in NP in IDD progression
and miR-129-5p could be a potential therapeutic target for IDD.
However, more experiments and further investigations are required
to verify this finding.
In conclusion, the present study identified that key
genes (TP53 and UBC) and miR-129-5p may participate
in the mechanism of IDD progression. Significant network features
identified in the study may aid in assessing the grade of disc
degeneration. TP53 may have an essential role in neovascularization
and infiltration in annulus fibrosus in grade III of IDD
progression and could help the IDD grading system. Additionally,
UBC may have a crucial role in inhibiting cell proliferation
of annulus fibrosus in IDD progression and could be used to grade
disc degeneration. Furthermore, miR-129-5p may have a crucial role
in modulating cell apoptosis in NP in IDD progression and
miR-129-5p could be a potential therapeutic target for IDD.
However, further experiments and studies are required to confirm
these results.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China: The effect and mechanism of
cytoskeletal elements in the mechanotransduction pathway within
intervertebral disc cells and its role in intervertebral disc
degeneration (grant no. 81471131) and the National Natural Science
Foundation of China: The effect and mechanism of chondroitin
sulphate proteoglycans in stem cell niche during stem cell
differentiation (grant no. 31300675).
References
|
1
|
Hadjipavlou AG, Tzermiadianos MN, Bogduk N
and Zindrick MR: The pathophysiology of disc degeneration: A
critical review. J Bone Joint Surg Br. 90:1261–1270. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang SZ, Rui YF, Tan Q and Wang C:
Enhancing intervertebral disc repair and regeneration through
biology: Platelet-rich plasma as an alternative strategy. Arthritis
Res Ther. 15:2202013. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yorimitsu E, Chiba K, Toyama Y and
Hirabayashi K: Long-term outcomes of standard discectomy for lumbar
disc herniation: A follow-up study of more than 10 years. Spine.
26:652–657. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Karasek M and Bogduk N: Twelve-month
follow-up of a controlled trial of intradiscal thermal anuloplasty
for back pain due to internal disc disruption. Spine. 25:2601–2607.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Anderson PA and Rouleau JP: Intervertebral
disc arthroplasty. Spine. 29:2779–2786. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shimer AL, Chadderdon RC, Gilbertson LG
and Kang JD: Gene therapy approaches for intervertebral disc
degeneration. Spine. 29:2770–2778. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Choi YS: Pathophysiology of degenerative
disc disease. Asian Spine J. 3:39–44. 2009. View Article : Google Scholar
|
|
8
|
Tang Y, Wang S, Liu Y and Wang X:
Microarray analysis of genes and gene functions in disc
degeneration. Exp Ther Med. 7:343–348. 2014.PubMed/NCBI
|
|
9
|
Antoniou J, Epure LM, Michalek AJ, Grant
MP, Iatridis JC and Mwale F: Analysis of quantitative magnetic
resonance imaging and biomechanical parameters on human discs with
different grades of degeneration. J Magn Reson Imaging.
38:1402–1414. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Thompson JP, Pearce RH, Schechter MT,
Adams ME, Tsang IK and Bishop PB: Preliminary evaluation of a
scheme for grading the gross morphology of the human intervertebral
disc. Spine. 15:411–415. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pfirrmann CW, Metzdorf A, Zanetti M,
Hodler J and Boos N: Magnetic resonance classification of lumbar
intervertebral disc degeneration. Spine. 26:1873–1878. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Adams MA and Roughley PJ: What is
intervertebral disc degeneration, and what causes it? Spine.
31:2151–2161. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bachmeier BE, Nerlich A, Mittermaier N,
Weiler C, Lumenta C, Wuertz K and Boos N: Matrix metalloproteinase
expression levels suggest distinct enzyme roles during lumbar disc
herniation and degeneration. Eur Spine J. 18:1573–1586. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Takahashi M, Haro H, Wakabayashi Y,
Kawauchi T, Komori H and Shinomiya K: The association of
degeneration of the inter-vertebral disc with 5a/6a polymorphism in
the promoter of the human matrix metalloproteinase-3 gene. J Bone
Joint Surg Br. 83:491–495. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pratsinis H and Kletsas D: PDGF, bFGF and
IGF-I stimulate the proliferation of intervertebral disc cells in
vitro via the activation of the ERK and Akt signaling pathways. Eur
Spine J. 16:1858–1866. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
van Rooij E and Kauppinen S: Development
of microRNA therapeutics is coming of age. EMBO Mol Med. 6:851–864.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu X, Li Z, Shen J, Wu WK, Liang J, Weng X
and Qiu G: MicroRNA-10b promotes nucleus pulposus cell
proliferation through RhoC-Akt pathway by targeting HOXD10 in
intervetebral disc degeneration. PLoS One. 8:e830802013. View Article : Google Scholar :
|
|
18
|
Chen Y, Chen K, Li M, Li C, Ma H, Bai YS,
Zhu XD and Fu Q: Genes associated with disc degeneration identified
using microarray gene expression profiling and bioinformatics
analysis. Genet Mol Res. 12:1431–1439. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang HQ, Yu XD, Liu ZH, Cheng X, Samartzis
D, Jia LT, Wu SX, Huang J, Chen J and Luo ZJ: Deregulated miR-155
promotes Fas-mediated apoptosis in human intervertebral disc
degeneration by targeting FADD and caspase-3. J Pathol.
225:232–242. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gruber HE, Hoelscher G, Loeffler B, Chow
Y, Ingram JA, Halligan W and Hanley EN Jr: Prostaglandin E1 and
misoprostol increase epidermal growth factor production in
3D-cultured human annulus cells. Spine J. 9:760–766. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tsai TT, Lai PL, Liao JC, Fu TS, Niu CC,
Chen LH, Lee MS, Chen WJ, Fang HC, Ho NY, et al: Increased
periostin gene expression in degenerative intervertebral disc
cells. Spine J. 13:289–298. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gentleman R, Carey VJ, Huber W, Irizarry
RA and Dudoit S: Bioinformatics and computational biology solutions
using R and Bioconductor. Springer; 2005, View Article : Google Scholar
|
|
23
|
Benjamini Y and Hochberg Y: Controlling
the false discovery rate: A practical and powerful approach to
multiple testing. J R Stat Soc B. 57:289–300. 1995.
|
|
24
|
Xiao F, Zuo Z, Cai G, Kang S, Gao X and Li
T: miRecords: An integrated resource for microRNA-target
interactions. Nucleic Acids Res. 37:D105–D110. 2009. View Article : Google Scholar
|
|
25
|
Hsu SD, Lin FM, Wu WY, Liang C, Huang WC,
Chan WL, Tsai WT, Chen GZ, Lee CJ, Chiu CM, et al: miRTarBase: A
database curates experimentally validated microRNA-target
interactions. Nucleic Acids Res. 39:D163–D169. 2011. View Article : Google Scholar
|
|
26
|
Vergoulis T, Vlachos IS, Alexiou P,
Georgakilas G, Maragkakis M, Reczko M, Gerangelos S, Koziris N,
Dalamagas T and Hatzigeorgiou AG: TarBase 6.0: Capturing the
exponential growth of miRNA targets with experimental support.
Nucleic Acids Res. 40:D222–D229. 2012. View Article : Google Scholar :
|
|
27
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar
|
|
28
|
Smoot ME, Ono K, Ruscheinski J, Wang PL
and Ideker T: Cytoscape 2.8: New features for data integration and
network visualization. Bioinformatics. 27:431–432. 2011. View Article : Google Scholar :
|
|
29
|
Franceschini A, Szklarczyk D, Frankild S,
Kuhn M, Simonovic M, Roth A, Lin J, Minguez P, Bork P, von Mering
C, et al: STRING v9.1: Protein-protein interaction networks, with
increased coverage and integration. Nucleic Acids Res.
41:D808–D815. 2013. View Article : Google Scholar :
|
|
30
|
Assenov Y, Ramírez F, Schelhorn S-E,
Lengauer T and Albrecht M: Computing topological parameters of
biological networks. Bioinformatics. 24:282–284. 2008. View Article : Google Scholar
|
|
31
|
Huang Y, Pepe MS and Feng Z: Logistic
regression analysis with standardized markers. Ann Appl Stat.
7:72013. View Article : Google Scholar
|
|
32
|
Sherman BT, Huang da W, Tan Q, Guo Y, Bour
S, Liu D, Stephens R, Baseler MW, Lane HC and Lempicki RA: DAVID
Knowledgebase: A gene-centered database integrating heterogeneous
gene annotation resources to facilitate high-throughput gene
functional analysis. BMC Bioinformatics. 8:4262007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Blevins L and McDonald C: Fisher's Exact
Test: an easy-to-use statistical test for comparing outcomes. MD
Comput. 2:151985.PubMed/NCBI
|
|
34
|
Estrada E and Higham DJ: Network
properties revealed through matrix functions. SIAM Rev. 52:696–714.
2010. View Article : Google Scholar
|
|
35
|
Estrada E: Virtual identification of
essential proteins within the protein interaction network of yeast.
Proteomics. 6:35–40. 2006. View Article : Google Scholar
|
|
36
|
Tailor A, Jurkovic D, Bourne TH, Collins
WP and Campbell S: Sonographic prediction of malignancy in adnexal
masses using multivariate logistic regression analysis. Ultrasound
Obstet Gynecol. 10:41–47. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dumont P, Leu JI, Della Pietra AC III,
George DL and Murphy M: The codon 72 polymorphic variants of p53
have markedly different apoptotic potential. Nat Genet. 33:357–365.
2003. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Vaghefi H and Neet KE: Deacetylation of
p53 after nerve growth factor treatment in PC12 cells as a
post-translational modification mechanism of neurotrophin-induced
tumor suppressor activation. Oncogene. 23:8078–8087. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Richardson SM, Doyle P, Minogue BM,
Gnanalingham K and Hoyland JA: Increased expression of matrix
metallopro-teinase-10, nerve growth factor and substance P in the
painful degenerate intervertebral disc. Arthritis Res Ther.
11:R1262009. View
Article : Google Scholar
|
|
40
|
Liu XW, Kang J, Fan XD and Sun LF:
Expression and significance of VEGF and p53 in rat degenerated
intervertebral disc tissues. Asian Pac J Trop Med. 6:404–406. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Marinovic AC, Zheng B, Mitch WE and Price
SR: Ubiquitin (UbC) expression in muscle cells is increased by
glucocorticoids through a mechanism involving Sp1 and MEK1. J Biol
Chem. 277:16673–16681. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yew PR: Ubiquitin-mediated proteolysis of
vertebrate G1- and S-phase regulators. J Cell Physiol. 187:1–10.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gruber HE, Ingram JA, Norton HJ and Hanley
EN Jr: Senescence in cells of the aging and degenerating
intervertebral disc: Immunolocalization of senescence-associated
β-galactosidase in human and sand rat discs. Spine. 32:321–327.
2007. View Article : Google Scholar
|
|
44
|
Kohyama K, Saura R, Doita M and Mizuno K:
Intervertebral disc cell apoptosis by nitric oxide: Biological
understanding of inter-vertebral disc degeneration. Kobe J Med Sci.
46:283–295. 2000.
|
|
45
|
Li M, Tian L, Wang L, Yao H, Zhang J, Lu
J, Sun Y, Gao X, Xiao H and Liu M: Down-regulation of miR-129-5p
inhibits growth and induces apoptosis in laryngeal squamous cell
carcinoma by targeting APC. PLoS One. 8:e778292013. View Article : Google Scholar : PubMed/NCBI
|