Introduction
Cutaneous wound healing is a dynamic and
well-ordered biological process that involves three critical steps:
inflammation, re-epithelialization and tissue remodeling (1). The re-epithelialization process
plays the most significant role and requires the proper migration
and proliferation of keratinocytes at the periphery of the wound
(2). Keratinocyte migration is a
complex process that involves the serial activation of kinases and
cytokines. Src, one of the proto-oncogenes encoding a
membrane-associated, non-receptor protein tyrosine kinase, has been
implicated in the regulation of a wide range of cellular functions,
including cell proliferation and cell migration (3). In normal and oncogenically
transformed cells, it has ben proposed that Src family kinases
regulate cell migration (4–6).
Additionally, it has been demonstrated that Src is essential for
wound healing in Drosophila as well as wound responses and
tissue regeneration in zebrafish (7,8).
Another study has reported that the activation of Src promotes
wound healing, whereas the inactivation of Src inhibits wound
closure in mouse corneal epithelial cells (9). However, it remains unclear whether
the role of Src in cutaneous wound healing is related to the
regulation of keratinocyte migration.
Keratinocyte migration also depends on the loss of
cell-matrix and cell-cell adhesion; the ability of keratinocytes to
detach from the underlying basal lamina and migrate through the
fibrin and extracellular matrix (ECM) meshwork of the wound is
important to the re-epithelialization process. Matrix
metalloproteinases (MMPs), which degrade different components of
the ECM, are essential for keratinocyte migration. Following
cutaneous injury, MMP expression is temporally and spatially
regulated in the wound; this helps to initiate and maintain
keratinocyte migration and is necessary for wound
re-epithelialization (10–12).
Human keratinocytes synthesize and secrete mainly MMP-1, MMP-2,
MMP-9 and MMP-10 (2). Several
studies have linked the gelatinases, MMP-2 and MMP-9, contribute to
cancer, infectious diseases, inflammation, vascular diseases and
wound healing (10,12,13). The mitogen-activated protein
kinase (MAPK) signaling pathway has been implicated in MMP-2
expression in oral cancer cells (14). Moreover, E-cadherin is a protein
which mediates cell-cell adhesion by forming homodimers on adjacent
cells. Accordingly, these data prompted us to investigate the
function of MAPK, MMPs and E-cadherin in the regulation of
Src-mediated keratinocyte migration in wound healing.
We hypothesized that Src accelerates keratinocyte
migration, at least in part, through MAPK, MMPs and E-cadherin. To
test this hypothesis, keratinocytes and in vivo wounds were
pre-treated with vector pcDNA3.1(+)-Src for overexpressing Src and
Src-specific small interfering RNA (siRNA) for the silencing of
Src, and then the effects on MAPK activation, cell migration,
E-cadherin, MMPs and wound healing were determined. Our in
vitro study found that Src promoted keratinocyte migration
through the upregulation of MMP-2 and the downregulation of
E-cadherin, and that the extracellular signal-regulated kinase
(ERK) pathway was involved in the Src-induced increase of MMP-2.
Our in vivo experiments showed that Src accelerated wound
healing. Thus, the present study offers valuable insights into the
molecular mechanisms responsible for keratinocyte migration and
wound healing, and it provides a rationale for the therapeutic
effect of Src on cutaneous wound healing.
Materials and methods
Animals and antibodies
Adult male Sprague-Dawley (SD) rats (n=50) weighing
220–250 g were purchased from the Center of Experimental Animals at
the Fourth Military Medical University (FMMU; Xi'an, China). The
experiments were conducted in accordance with the Guide for the
Care and Use of Laboratory Animals of the FMMU, and all
experimental protocols used in this study were approved by the
Animal Care Committee of the FMMU.
Cell culture
All human tissues were obtained from 4 patients
(mean age, 30 years) at Xijing Hospital (Xi'an, China).
Hypertrophic scar and surrounding normal skin tissues were obtained
from the same patients. Prior to the experiment, all patients were
informed about the purpose of the study as well as the procedures,
and voluntarily agreed to provide tissue. Written consent was
obtained from all participants, and all protocols were approved by
the Ethics Committee of Xijing Hospital, which is affiliated with
the FMMU. Briefly, the epidermal layer of human keratinocytes was
separated from the dermis and placed into a sterile 15-ml conical
tube containing 2 ml 0.05% trypsin-EDTA. The cells were incubated
at 37°C for approximately 15 min, during which time the cells were
triturated using a 2-ml pipette every 2–3 min to aid cell
dissociation. The cells were centrifuged at 180 × g for 7 min at
room temperature. After resuspension, the primary keratinocytes
were seeded in 75 cm2 tissue culture flasks at a density
of approximately 3×106 cells/flask in 15 ml keratinocyte
serum-free medium (KSFM) with the addition of bovine pituitary
extract (both from Gibco, Grand Island, NY, USA), and 1%
antibiotics (penicillin-streptomycin, both 100 U/ml). The medium
was changed every 2 days until the cells reached confluence, and
the keratinocytes were cultured continuously until use.
Western blot analysis
The keratinocytes were washed with ice-cold
phosphate-buffered saline (PBS) and lysed using RIPA buffer
supplemented with protease and phosphatase inhibitor mixtures
(Heart Biological Technology Co., Ltd., Xi'an, China) on ice. The
lysates were separated by centrifugation at 4°C and 14,000 × g for
10 min. Subsequently the protein concentration was determined using
a bicinchoninic acid (BCA) assay (Pierce, Rockford, IL, USA). Fifty
micrograms of total protein was subjected to sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred to PVDF membranes (Millipore Corp., Bedford, MA, USA).
After blocking with 5% non-fat milk, the membranes were incubated
with mouse anti-Src (1:200, sc-8056; Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA), rabbit anti-ERK1/2 (4695),
anti-phosphorylated (p-)ERK1/2 (4370), anti-c-Jun N-terminal kinase
(JNK) (9258), anti-p-JNK (4668), anti-p38 (8690), anti-p-p38 (4511)
(all at 1:1,000 dilution; Cell Signaling Technology, Inc., Danvers,
MA, USA), mouse anti-MMP-2 (ZS-135950), goat anti-MMP-9 (ZS-68400,
both at 1:200 dilution; Zhongshan Jinqiao Biological Technology,
Co. Ltd., Beijing, China), mouse anti-E-cadherin (1:1000, 610181;
BD Biosciences, Franklin Lakes, NJ, USA), goat anti-actin (1:200,
sc-1616; Santa Cruz Biotechnology, Inc.) overnight at 4°C. The
following day, the membranes were incubated with horseradish
peroxidase-conjugated secondary antibodies (1:3,000) at 37°C for 1
h. The immunoreactive proteins were then visualied using ECL
western blotting detection reagent (Millipore Corp., Billerica, MA,
USA) and detected using the MultiImage Light Cabinet filter
positions (Alpha Innotech Corp., San Leandro, CA, USA).
Plasmid vector for Src overexpression and
siRNA for silen cing of Src expression
Full-length human Src cDNA was amplified by reverse
transcription-polymerase chain reaction (RT-PCR) from normal skin
RNA. The PCR products were digested with NheI and
KpnI, and subcloned into the eukaryotic expression vector,
pcDNA3.1(+) (GeneCopoeia, Rockville, MD, USA). The recombinant
pcDNA3.1(+)-Src plasmid (GeneCopoeia) was confirmed by sequencing.
The keratinocytes were plated in 60 cm2 wells for 24 h
prior to transfection. Thereafter, the cells were transfected with
either 5 µg pcDNA3.1(+)-Src plasmid or the empty vector
(GeneCopoeia) using Lipofectamine 2000 transfection reagent
(Invitrogen, Carlsbad, CA, USA). The siRNA (80 nM; Thermo Fisher
Scientific, Waltham, MA, USA) duplex sequence was as follows:
sense, 5′CGAGUGCCUUAUCCAAGAATT-3′. The cells transfected with
scrambled siRNA (Thermo Fisher Scientific) (sense,
5′UUCUUGGAUAAGGCACUCGTT-3′), which were the mock group, were also
transfected using Lipofectamine 2000 transfection reagent. Western
blot analysis was performed after an additional 48 h.
Flow cytometric analysis
Cell cycle distribution was analyzed by flow
cytometry (FACSAria; BD Biosciences). Primary keratinocytes were
divided into the following groups: control, pcDNA3.1(+),
pcDNA3.1(+)-Src, mock and also the Src-siRNA transfected groups.
Twenty-four hours after transfection, the cells were harvested,
rinsed with PBS, fixed with 95% (v/v) ice-cold ethanol and
resuspended in staining buffer containing FITC-Annexin V and
propidium iodide (PI). The mixture was then incubated in the dark
at room temperature for 15 min. The DNA contents of the stained
nuclei were analyzed, and the number of cells in each cycle phase
was calculated.
In vitro cell migration assays
In the present study, the keratinocytes transfected
with pcDNA3.1(+)-Src plasmid, Src-siRNA, pcDNA3.1(+), scrambled
siRNA (mock group) and untransfected cells (as control), were grown
to confluence in 12-well plates in KSFM containing 10 µg/ml
mitomycin C (Invitrogen) for 1 h to completely inhibit cell
proliferation. A straight scratch was made on the keratinocytes
using a P200 pipette tip. The cells were then washed with PBS three
times, and further cultured in KSFM. After incubating for 0, 12 and
24 h, the gap width of scratch re-population was measured and
recorded, and then compared with the initial gap size at 0 h. Using
the ImageJ image processing program, the size of the denuded area
was determined at each time point from the digital images. Where
indicated, the cells were treated with JNK inhibitor (SP-600125 at
30 µM; Sigma-Aldrich, St. Louis, MO, USA) or the ERK
inhibitor (PD-98059 at 30 µM; Beyotime, Haimen, China) 1 h
prior to experiments.
Immunocytofluorescence analysis
For immunocytofluorescence analysis, keratinocytes
were fixed with 4% paraformaldehyde. Cells were permeabilized with
Triton X-100 and then blocked with BSA. Cells were incubated with
anti-E-cadherin (1:100, 610181; BD Biosciences) at 4°C overnight
and with CY3-555 IgG antibody (A28180; Invitrogen) at 37°C for 1 h
on the following day. DAPI was used for nuclear staining. Images
were captured by an FSX100 microscope (Olympus, Tokyo, Japan).
In vivo wound closure assay
Following anesthetization, a 1.0×1.0-cm2
full-thickness wound was punched onto the back of male SD rats. To
evaluate the effect of Src on wound healing, 20 µg
pcDNA3.1(+)-Src plasmids and 400 nM Src-siRNA duplexes were
topically transfected into the epidermis surrounding the wounds at
four sites on days 1, 5, 9, 13 and 17 for a total of five doses.
The same quantity of empty plasmid and scrambled siRNA were used as
controls. The process of wound healing was digitally photographed
and recorded. Wound area measurement was performed by digital
planimetry using ImageJ software. The relative residual wound area
was calculated as the ratio between the residual wound area at a
given time point and the original wound area × 100%. At least 10
rats were used for each time point examined. The average unhealed
wound gap was defined as the distance between the advancing edges
of epidermal keratinocyte migration. All images were captured with
the FSX100 microscope; multiple overlapping pictures were used to
artificially reconstitute the entire wounds.
Statistical analysis
Each experiment was repeated at least three times,
and the data are presented as the means ± SEM. Statistical
differences between groups were analyzed by the Student's t-test or
the Mann-Whitney U test as appropriate using SPSS 13.0. A P-value
<0.05 was considered to indicate a statistically significant
difference.
Results
Src promotes keratinocyte migration and
does not markedly affect keratinocyte proliferation in vitro
Keratinocyte migration and proliferation determine
the rate of wound re-epithelialization (1). In order to examine the role of Src
in keratinocyte migration in vitro, we first constructed
vector for overexpressing Src [pcDNA3.1(+)-Src] and siRNA for
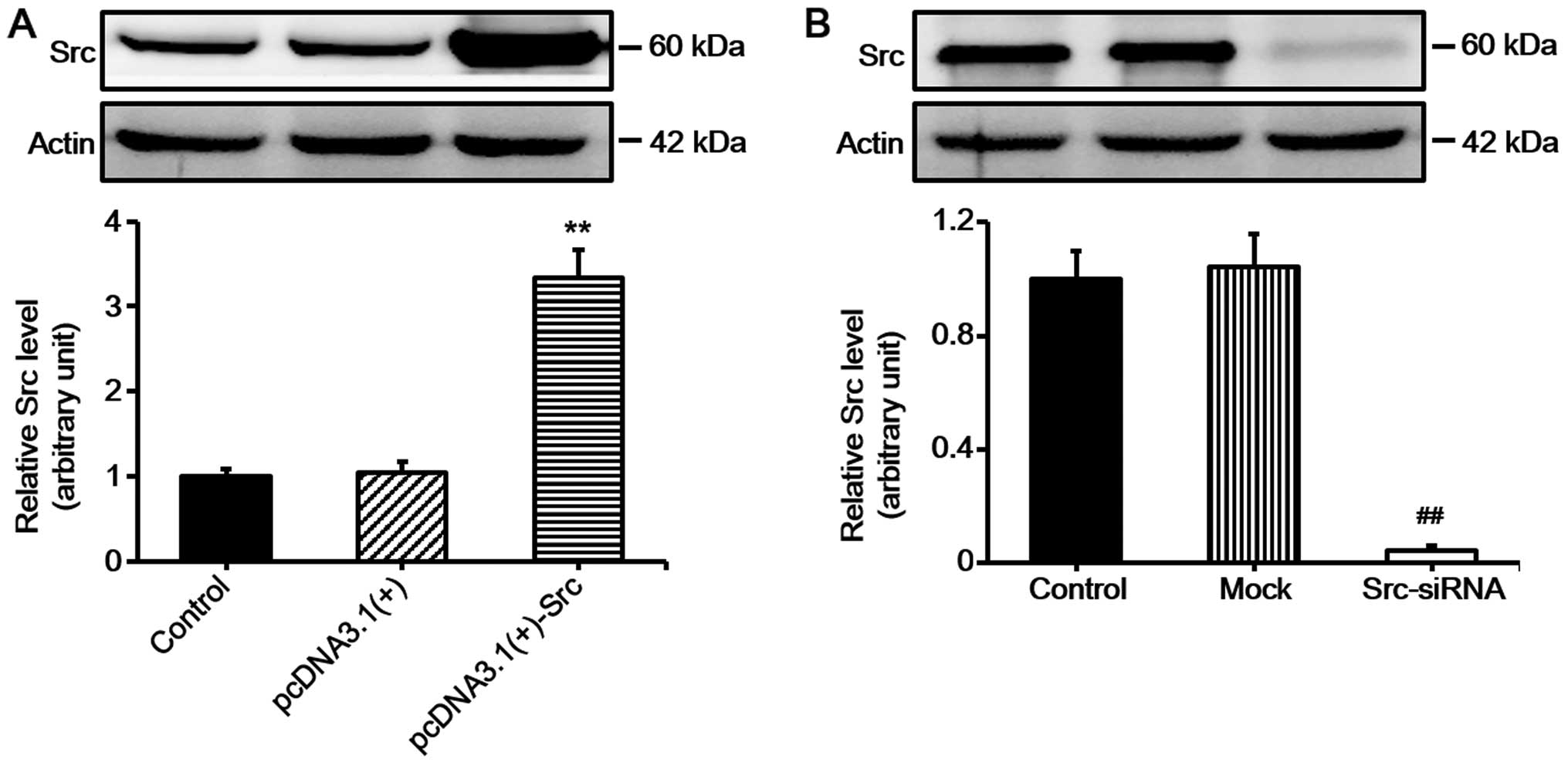
silencing Src (Src-siRNA). Western blot analysis confirmed the
effective overexpression of Src and silencing of Src (Fig. 1). Subsequently, scratch wound
healing assays were performed in order to evaluate the effects of
overexpressing Src and silencing of Src on keratinocyte migration.
The cells were pre-treated with mitomycin C for 1 h to inhibit
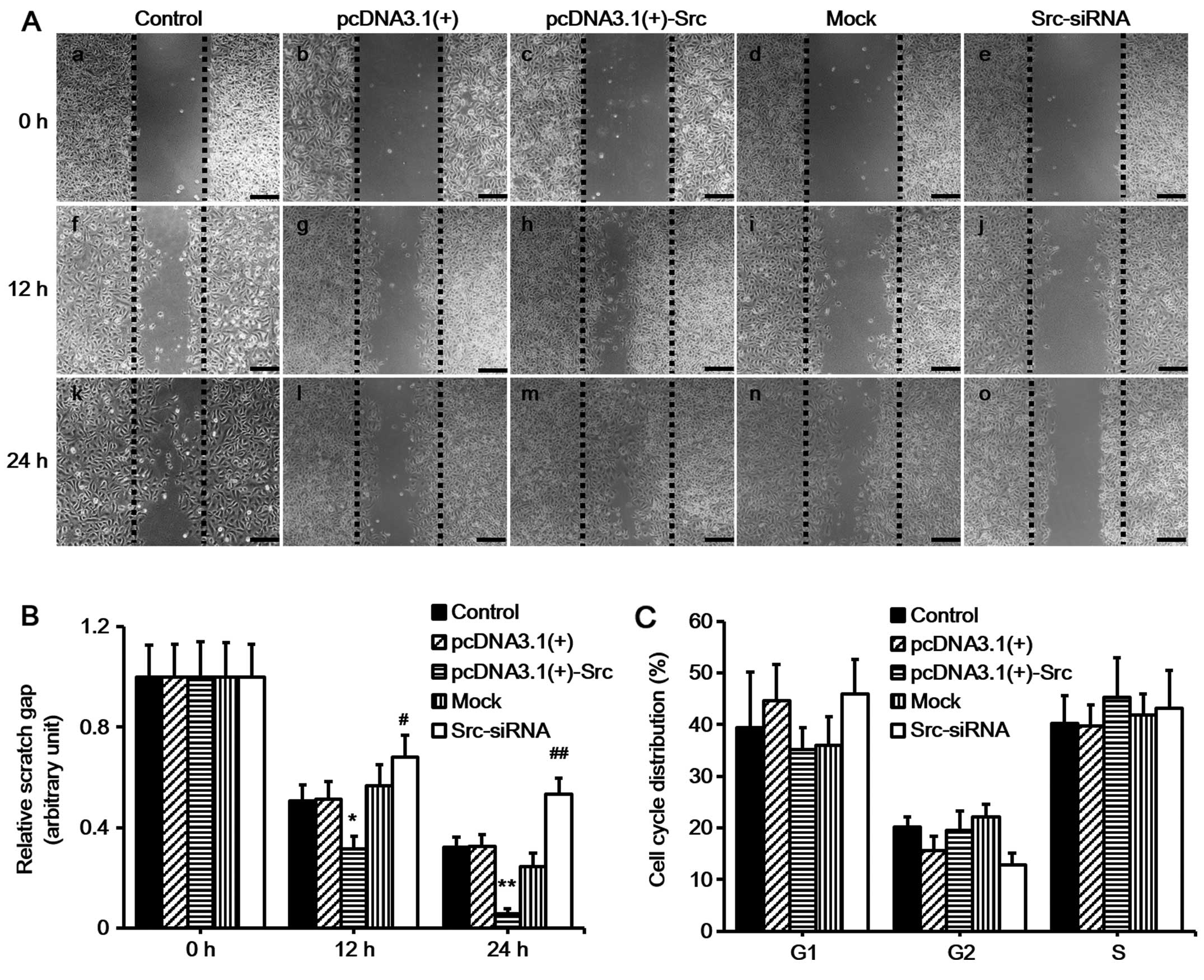
proliferation prior to the scratch assay. The results demonstrated
that keratinocytes over-expressing Src migrated into over 90% of
the scratched area, pcDNA3.1(+)-Src group, while the Src-silenced
keratinocytes migrated into only 30% of the area
(Src-siRNA-transfected group), compared with the untransfected
cells (as control) or negative vectors [mock and pcDNA3.1(+) group]
at 24 h post-scratching (Fig. 2A and
B). Flow cytometric analysis showed that Src did not
significantly affect the percentage of cells in the G1, G2, or S
phase, indicating that Src did not exert a marked effect on cell
proliferation (Fig. 2C). Taken
together, these results suggest that Src promotes keratinocyte
migration while exerting no marked effect on cell
proliferation.
Src promotes cutaneous wound healing in
vivo
Our in vitro experiment indicated that Src
promotes keratinocyte migration. As keratinocyte migration plays a
very important role in wound healing, we hypothesized that Src
promotes wound healing in vivo. As in the previous
experiment, the Src-overexpression vector, pcDNA3.1(+)-Src, and
Src-siRNA were also applied to wounds of SD rats (data not shown).
To test our hypothesis, we first established a rat model of
full-thickness wound healing. The pcDNA3.1(+), pcDNA3.1(+)-Src,
mock (scrambled siRNA) and Src-siRNA were injected at the wound
edges on days 1, 5, 9, 13 and 17 post-wounding. Wound healing was
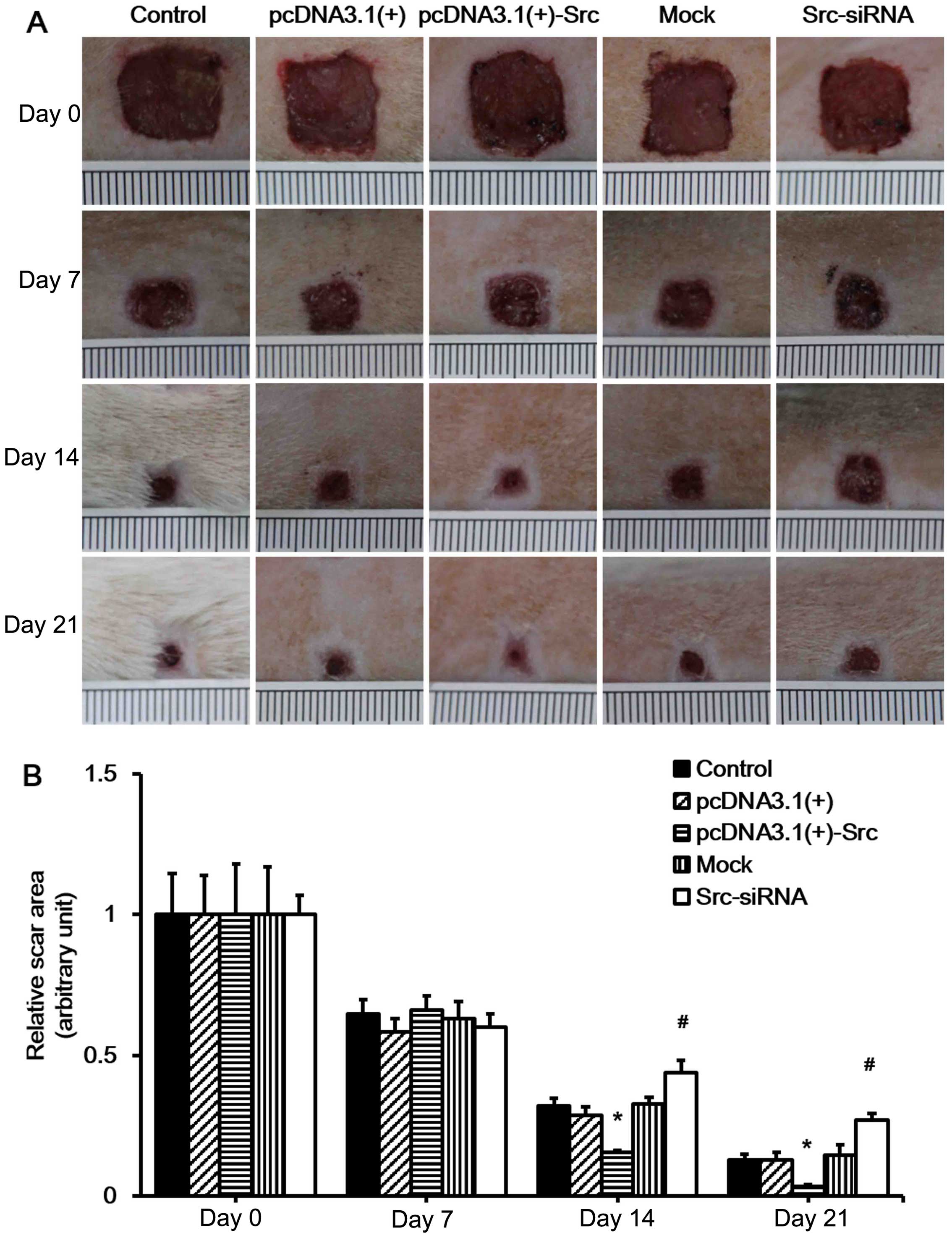
analyzed daily and selected wound images are shown (Fig. 3A). The results showed that
pcDNA3.1(+)-Src transfection significantly accelerated wound
closure on 14 and 21 days, while it was observed that the wounds of
the Src-siRNA-transfected rats healed at a slower rate on days 14
and 21, indicating that the initial wound healing in the Src group
happened as early as day 7 post-wounding (Fig. 3). Taken together, these results
suggest that overexpression of Src at the wound edge accelerates
the process of wound healing.
ERK and JNK signaling are involved in
Src-promoted keratinocyte migration
In the present study, in order to explore the role
of the MAPK pathways in Src-regulated keratinocyte migration, we
investigated the activation of MAPK pathways in keratinocytes
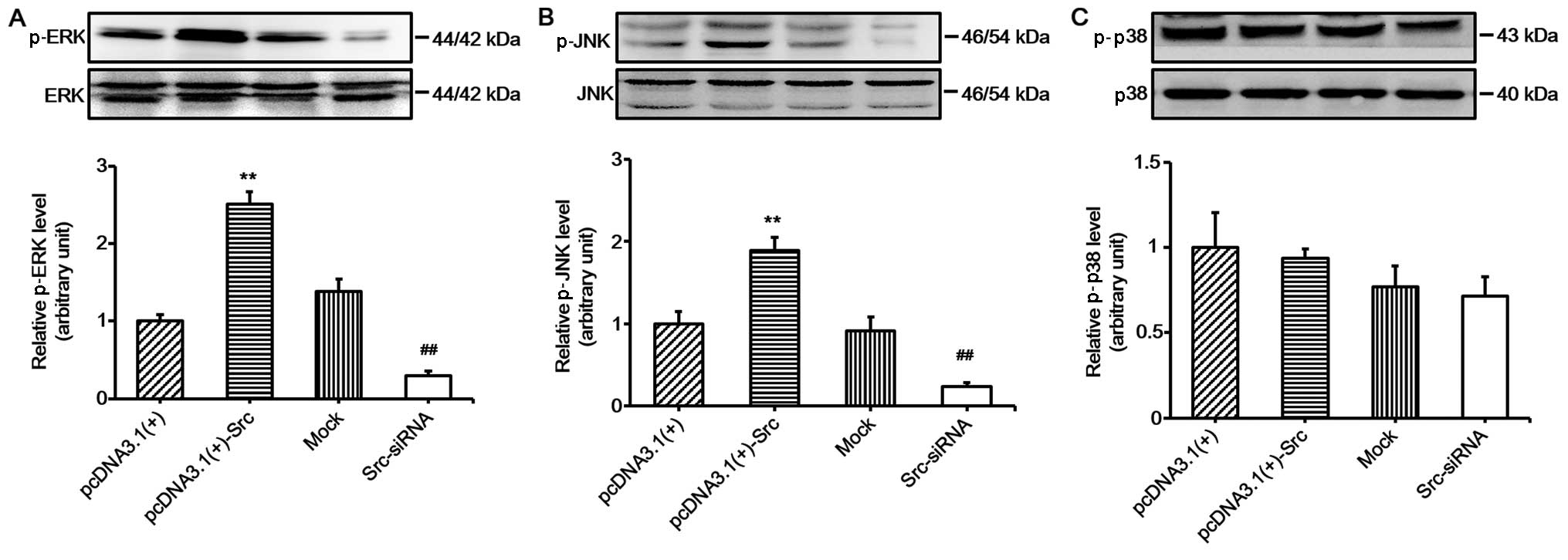
overexpressing Src and Src-silenced keratinocytes. The results
showed that the phosphorylation levels of JNK and ERK were
significantly increased in keratinocytes overexpressing Src and
decreased in Src-silenced keratino-cytes, while the phosphorylation
levels of p38 were relatively unchanged (Fig. 4). These results suggest that the
JNK and ERK pathways participate in Src-promoted keratinocyte
migration. To further confirm the important role of the JNK and ERK
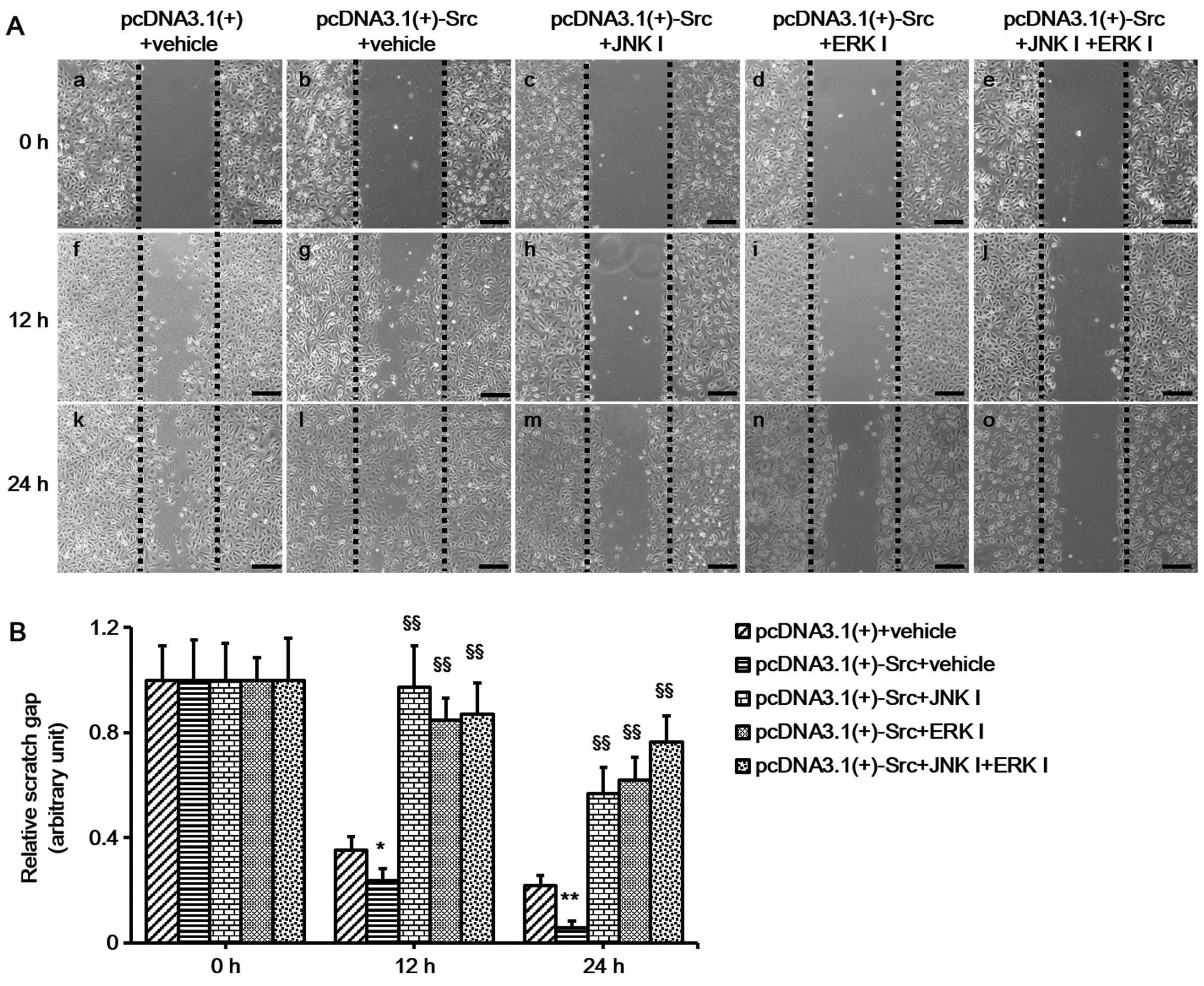
pathways, keratinocytes overexpressing Src were treated with ERK
inhibitor (PD-98059 at 30 µM) or JNK inhibitor (SP-600125 at
30 µM) or both. The cells were pretreated with mitomycin C
to inhibit cell proliferation prior to the scratch wound-healing
assays. The results showed that the ERK inhibitor, PD-98059, and
the JNK inhibitor, SP-600125, slowed down the course of scratch gap
closure when administered separately, while the combination of the
two further slowed the progression (Fig. 5). Thus, taken together, the
results of these experiments indicate that JNK and ERK signaling
play a crucial role in the process of Src-promoted keratinocyte
migration.
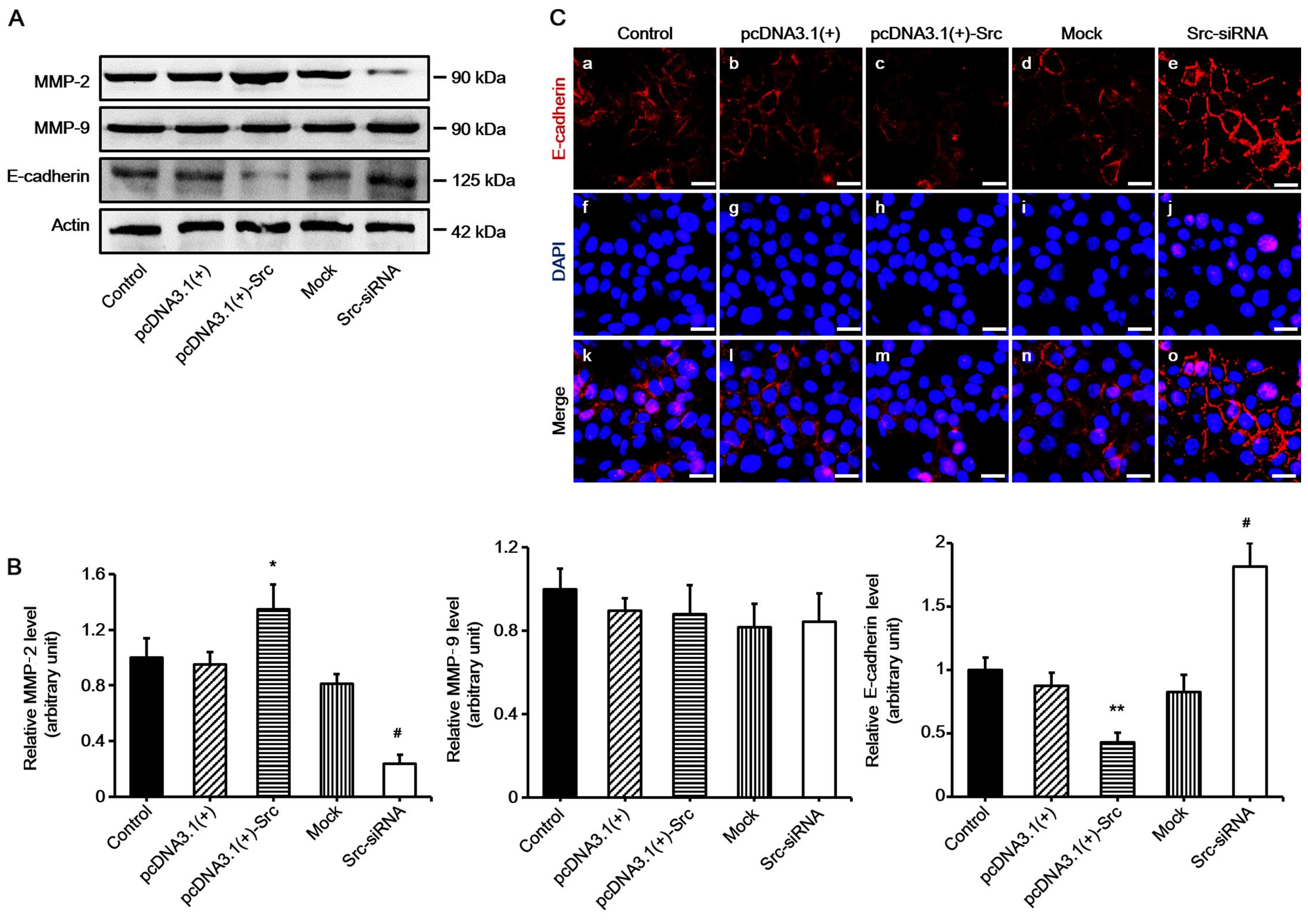
MMP-2 and E-cadherin are involved in
Src-promoted keratinocyte migration
Many factors play an important role in keratinocyte
migration, particularly MMPs and E-cadherin (15,16). In order to examine the mechanism
of Src-promoted keratinocyte migration, the expression levels of
MMP-2, MMP-9 and E-cadherin were determined. The results
demonstrated that the expression of MMP-2 was significantly
increased in keratinocytes overexpressing Src, and decreased in
Src-silenced keratinocytes compared with the untransfected cells
(acting as control) or negative vectors, which were the mock and
pcDNA3.1(+) groups. The expression of MMP-9 was not changed
significantly. By contrast to the results noted for MMP-2, the
expression levels of E-cadherin were markedly decreased in
keratinocytes overexpressing Src and increased in Src-silenced
keratinocytes (Fig. 6). Thus, our
findings suggest that MMP-2 and E-cadherin participate in
Src-promoted keratinocytes migration.
ERK signaling is involved in Src-induced
changes in MMP-2 expression
Our results indicated that the ERK and JNK pathways
played important roles in Src-promoted keratinocyte migration, and
the expression of MMP-2 and E-cadherin changed during this process.
In order to further elucidate whether ERK and JNK signaling played
an important role in Src-induced expression changes in MMP-2 and
E-cadherin, the keratinocytes were transfected with pcDNA3.1(+)-Src
vector and then treated with the ERK inhibitor, PD-98059, or the
JNK inhibitor, SP-600125, or both. The results demonstrated that
PD-98059 alone significantly prevented Src-induced changes in MMP-2
protein expression. The combination of the two inhibitors did not
abolish MMP-2 expression. Moreover, no significant change was
detected in the protein expression of MMP-9. The protein expression
of E-cadherin was almost unchanged when the inhibitors were used on
their own or in combination, indicating that a different mechanism
was involved in the regulation of E-cadherin (Fig. 7). Collectively, these results
indicate that ERK signaling plays an important role in Src-induced
changes in MMP-2 protein expression.
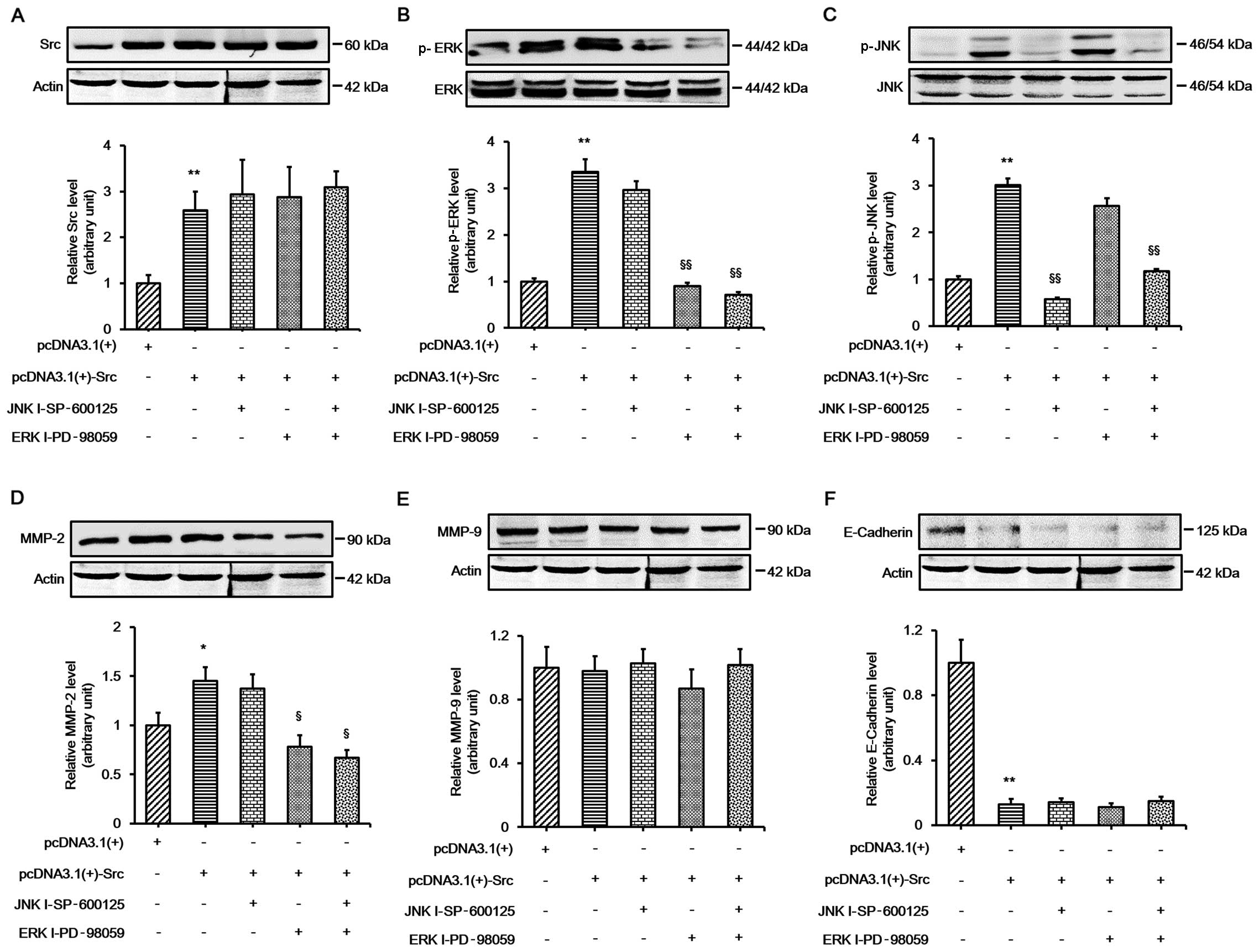 | Figure 7Involvement of extracellular
signal-regulated kinase (ERK) signaling in Src-induced changes in
MMP-2 expression. Western blot analysis showing the expression of
(A) Src, (B) ERK, (C) c-Jun N-terminal kinase (JNK), (D) MMP-2, (E)
MMP-9 and (F) E-cadherin in keratinocytes overexpressing Src
treated with ERK inhibitor (ERK I; PD-98059 at 30 µM), JNK
inhibitor (JNK I; SP-600125 at 30 µM) or both. Actin was
used as a loading control. Bars represent the means ± SEM of n=4
tissue samples. **P<0.01, pcDNA3.1(+)-Src group vs.
pcDNA 3.1(+) group, *P<0.05, pcDNA3.1(+)-Src group
vs. pcDNA 3.1(+) group; §§P<0.01, JNK I - SP-600125
vs. pcDNA3.1(+)-Src group, §§P<0.01, ERK I-PD-98059
vs. pcDNA3.1(+)-Src group, §§P<0.01, JNK I -
SP-600125 + ERK I-PD-98059 vs. pcDNA3.1(+)-Src group,
§P<0.05, ERK I-PD-98059 vs. pcDNA3.1(+)-Src group,
§P<0.05, JNK I - SP-600125 + ERK I-PD-98059 vs.
pcDNA3.1(+)-Src group. |
Discussion
Wound healing is a complex process which requires
re-epithelialization (17).
Keratinocyte migration is an essential process in wound healing,
and involves the serial activation of kinases and cytokines.
Studies have demonstrated that Src family members participate in
the regulation of the cell cycle, cell proliferation, migration,
adhesion, and differentiation (18). In corneal epithelial cells, Src
has been found to be activated along the leading edge of
scratch-wounded cultures (9);
however, the role of Src in keratinocytes has not previously been
reported in depth. In the present study, we found that Src
overexpression promoted keratinocyte migration and Src silencing
inhibited keratinocyte migration without exerting a marked effect
on keratinocyte proliferation in vitro (Figs. 1 and 2). The positive effects of Src on the
migration of human keratinocytes suggest that Src plays a
significant role in wound re-epithelialization. We also found that
Src promoted wound healing in vivo in SD rats (Fig. 3). Taken together, these results
demonstrate that the effect of Src on wound healing was mainly due
to its role in keratinocyte migration.
MMPs are an important family of cell
migration-related proteins which degrade different components of
the ECM. Previous research has shown that MMPs also contribute to
cell migration during wound healing (19). Several studies have demonstrated
that following skin wounding, keratinocytes bind with type I
collagen to stimulate an epidermal growth factor (EGF)
receptor-dependent cascade resulting in the expression of MMPs
(20–23). Previous research has revealed that
exogenous treatment with MMP inhibitors delayed wound healing and
re-epithelialization (24,25).
By contrast, the overexpression of MMPs was found to be detrimental
to the wound healing process, and the expression level of MMPs was
increased in chronic wounds (26,27). These findings suggest that MMPs
act positively and also negatively regulate wound healing
processes, and collectively they indicate the importance of MMPs in
wound healing. In addition, a previous study has shown that Src
promotes cancer cell migration by increasing the levels of MMP-2
and MMP-9 (28). Consistent with
this, in the present study we found that MMP-2 expression was
increased during Src-induced accelerated keratinocyte migration,
while the expression of MMP-9 was not changed significantly
(Fig. 6). The inconsistent
results may be related to the use of different cell lines in this
previous study and our present study.
An essential condition for cell migration is the
loosening of adherent junctions which facilitate cell-cell adhesion
through homotypic binding between E-cadherin molecules on adjacent
cells (29–31). Thus, E-cadherin is clearly another
important protein in cell migration. A previous study has shown
that Src kinase plays a crucial role in the maintenance of
epithelial integrity by upregulating E-cadherin during epithelial
morphogenesis (32). Similarly,
it has previously been suggested that the regulation of E-cadherin
expression in lung cancer is also linked to c-Src activity
(33). Furthermore, several
studies have demonstrated that the main role of Src in
re-epithelialization is to control the adhesion of leading cells by
regulating E-cadherin expression (34,35). In the present study, we found that
E-cadherin expression is partly regulated by Src (Fig. 6), and we suggest that a reduction
of E-cadherin expression plays an important role in Src-promoted
keratinocyte migration and accelerated wound healing. Our findings
concur with previous research in cancer cell migration (36).
Src has been implicated in the regulation of
signaling pathways associated with cell migration and
proliferation, such as Akt, STAT3 phosphorylation and Ras
activation (37–39). Previous research has indicated
that Src is involved in the activation of MAPK in cancer cells and
that it is considered a downstream target of Src signaling
(40–42). MAPKs, including ERK1/2, JNK and
p38, have all been implicated in the regulation of cell migration
and proliferation (43–46). Downregulation of the ERK1/2/MAPK
pathway leads to the decreased migration and proliferation of the
cell (47–49). Notably, the inhibition of Src
family kinases suppresses the activation of ERK1/2 (50). Additionally, it has been
demonstrated that inhibition of the JNK/MAPK pathway decreases the
migration of endothelial cells (51), and a correlation has been noted
between the p38/MAPK pathway and cell migration into the wound
along the wound edge (52). Thus,
in the present study the phosphorylation levels of JNK, ERK1/2 and
p38 were examined in keratinocytes overexpressing Src and
Src-silenced keratinocytes. We found that the phosphorylation
levels of JNK and ERK1/2 were significantly increased in
keratinocytes overexpressing Src and decreased in Src-silenced
keratinocytes, while the phosphorylation levels of p38 were
relatively unchanged (Fig. 4).
These findings suggest that the ERK1/2 and JNK pathways play an
integral role in Src-regulated wound healing and also that the p38
pathway is not involved in this process.
To further understand the mechanisms through which
Src regulates wound healing through the ERK1/2 and JNK pathways,
keratinocyte migration in vitro was evaluated after treating
keratinocytes overexpressing Src with the ERK1/2 inhibitor PD-98059
or the JNK inhibitor SP-600125, or both. The protein expression
levels of MMP-2, MMP-9 and E-cadherin were then assessed. We
demonstrated that administration of the ERK1/2 inhibitor, the JNK
inhibitor, or both, inhibited keratinocyte migration, and the
inhibition of ERK1/2 signaling decreased MMP-2 expression; the
inhibition of JNK activity did not markedly decrease MMP-2
expression (Figs. 5 and 7). No significant change was detected in
the protein expression of MMP-9 and E-cadherin when inhibitors were
used alone or in combination (Fig.
7). These findings suggest that the ERK1/2 pathway is involved
in the process of Src-mediated upregulation of MMP-2. Activation of
the MAPK pathway has been previously implicated in the regulation
of MMP gene expression (14).
Moreover, it has previously been reported that kaempferol reduces
MMP-2 expression by down-regulating ERK1/2 signaling pathways in
oral cancer cells (14). The
results of the present study reveal that ERK1/2 is involved in the
Src-mediated regulation of MMP-2 expression, whereas the protein
expression of E-cadherin is regulated through a different
mechanism. In the future, in order to elucidate the mechanisms
through which Src regulates wound healing via the ERK1/2 and JNK
pathways, further investigation is clearly warranted.
In conclusion, in the present study we discovered
that the overexpression of Src accelerates wound healing by
promoting keratinocyte migration, and that the mechanism appears to
involve the upregulation of MMP-2 via the ERK1/2 pathway. These
results provide us with a better understanding of the mechanisms
responsible for wound healing and provide a novel therapeutic
approach with which to improve wound healing.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (no. 81171811), the Special
Scientific Research Projects of National Health and Family Planning
Commission (no. 2015SQ00060), and the Natural Science Foundation of
Shaanxi Province (no. 2014JM4180).
References
|
1
|
Singer AJ and Clark RA: Cutaneous wound
healing. N Engl J Med. 341:738–746. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Santoro MM and Gaudino G: Cellular and
molecular facets of keratinocyte reepithelization during wound
healing. Exp Cell Res. 304:274–286. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cary LA, Klinghoffer RA, Sachsenmaier C
and Cooper JA: SRC catalytic but not scaffolding function is needed
for integrin-regulated tyrosine phosphorylation, cell migration,
and cell spreading. Mol Cell Biol. 22:2427–2440. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Avizienyte E, Fincham VJ, Brunton VG and
Frame MC: Src SH3/2 domain-mediated peripheral accumulation of Src
and phosphomyosin is linked to deregulation of E-cadherin and the
epithelial-mesenchymal transition. Mol Biol Cell. 15:2794–2803.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
González L, Agulló-Ortuño MT,
García-Martínez JM, Calcabrini A, Gamallo C, Palacios J, Aranda A
and Martín-Pérez J: Role of c-Src in human MCF7 breast cancer cell
tumorigenesis. J Biol Chem. 281:20851–20864. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ishizawar R and Parsons SJ: c-Src and
cooperating partners in human cancer. Cancer Cell. 6:209–214. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yoo SK, Freisinger CM, LeBert DC and
Huttenlocher A: Early redox, Src family kinase, and calcium
signaling integrate wound responses and tissue regeneration in
zebrafish. J Cell Biol. 199:225–234. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tsarouhas V, Yao L and Samakovlis C: Src
kinases and ERK activate distinct responses to Stitcher receptor
tyrosine kinase signaling during wound healing in Drosophila. J
Cell Sci. 127:1829–1839. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gao CY, Stepp MA, Fariss R and Zelenka P:
Cdk5 regulates activation and localization of Src during corneal
epithelial wound closure. J Cell Sci. 117:4089–4098. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kähäri VM and Saarialho-Kere U: Matrix
metalloproteinases in skin. Exp Dermatol. 6:199–213. 1997.
View Article : Google Scholar
|
|
11
|
Madlener M, Parks WC and Werner S: Matrix
metalloproteinases (MMPs) and their physiological inhibitors
(TIMPs) are differentially expressed during excisional skin wound
repair. Exp Cell Res. 242:201–210. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ravanti L and Kähäri VM: Matrix
metalloproteinases in wound repair (Review). Int J Mol Med.
6:391–407. 2000.PubMed/NCBI
|
|
13
|
Coussens LM, Tinkle CL, Hanahan D and Werb
Z: MMP-9 supplied by bone marrow-derived cells contributes to skin
carcinogenesis. Cell. 103:481–490. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lin CW, Chen PN, Chen MK, Yang WE, Tang
CH, Yang SF and Hsieh YS: Kaempferol reduces matrix
metalloproteinase-2 expression by down-regulating ERK1/2 and the
activator protein-1 signaling pathways in oral cancer cells. PLoS
One. 8:e808832013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xue M, Le NT and Jackson CJ: Targeting
matrix metalloproteases to improve cutaneous wound healing. Expert
Opin Ther Targets. 10:143–155. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Asai J, Hirakawa S, Sakabe J, Kishida T,
Wada M, Nakamura N, Takenaka H, Mazda O, Urano T, Suzuki-Inoue K,
et al: Platelets regulate the migration of keratinocytes via
podoplanin/CLEC-2 signaling during cutaneous wound healing in mice.
Am J Pathol. 186:101–108. 2016. View Article : Google Scholar
|
|
17
|
Martin P: Wound healing - aiming for
perfect skin regeneration. Science. 276:75–81. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Thomas SM and Brugge JS: Cellular
functions regulated by Src family kinases. Annu. Rev Cell Dev Biol.
13:513–609. 1997. View Article : Google Scholar
|
|
19
|
Raja, Sivamani K, Garcia MS and Isseroff
RR: Wound re-epithelialization: modulating keratinocyte migration
in wound healing. Front Biosci. 12:2849–2868. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pilcher BK, Wang M, Qin XJ, Parks WC,
Senior RM and Welgus HG: Role of matrix metalloproteinases and
their inhibition in cutaneous wound healing and allergic contact
hypersensitivity. Ann N Y Acad Sci. 878:12–24. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pilcher BK, Dumin JA, Sudbeck BD, Krane
SM, Welgus HG and Parks WC: The activity of collagenase-1 is
required for keratinocyte migration on a type I collagen matrix. J
Cell Biol. 137:1445–1457. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sudbeck BD, Pilcher BK, Welgus HG and
Parks WC: Induction and repression of collagenase-1 by
keratinocytes is controlled by distinct components of different
extracellular matrix compartments. J Biol Chem. 272:22103–22110.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pilcher BK, Dumin J, Schwartz MJ, Mast BA,
Schultz GS, Parks WC and Welgus HG: Keratinocyte collagenase-1
expression requires an epidermal growth factor receptor autocrine
mechanism. J Biol Chem. 274:10372–10381. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mirastschijski U, Haaksma CJ, Tomasek JJ
and Agren MS: Matrix metalloproteinase inhibitor GM 6001 attenuates
keratinocyte migration, contraction and myofibroblast formation in
skin wounds. Exp Cell Res. 299:465–475. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Agren MS: Matrix metalloproteinases (MMPs)
are required for re-epithelialization of cutaneous wounds. Arch
Dermatol Res. 291:583–590. 1999. View Article : Google Scholar
|
|
26
|
Saarialho-Kere UK: Patterns of matrix
metalloproteinase and TIMP expression in chronic ulcers. Arch
Dermatol Res. 290(Suppl): S47–S54. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fray MJ, Dickinson RP, Huggins JP and
Occleston NL: A potent, selective inhibitor of matrix
metalloproteinase-3 for the topical treatment of chronic dermal
ulcers. J Med Chem. 46:3514–3525. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang Y, Bai ZG, Yin J, Wu GC and Zhang ZT:
Role of c-Src activity in the regulation of gastric cancer cell
migration. Oncol Rep. 32:45–49. 2014.PubMed/NCBI
|
|
29
|
Perez-Moreno M, Jamora C and Fuchs E:
Sticky business: orchestrating cellular signals at adherens
junctions. Cell. 112:535–548. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Takeichi M: Cadherin cell adhesion
receptors as a morphogenetic regulator. Science. 251:1451–1455.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
van Roy F and Berx G: The cell-cell
adhesion molecule E-cadherin. Cell Mol Life Sci. 65:3756–3788.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shindo M, Wada H, Kaido M, Tateno M,
Aigaki T, Tsuda L and Hayashi S: Dual function of Src in the
maintenance of adherens junctions during tracheal epithelial
morphogenesis. Development. 135:1355–1364. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dong S, Khoo A, Wei J, Bowser RK,
Weathington NM, Xiao S, Zhang L, Ma H, Zhao Y and Zhao J: Serum
starvation regulates E-cadherin upregulation via activation of
c-Src in non-small-cell lung cancer A549 cells. Am J Physiol Cell
Physiol. 307:C893–C899. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Abreu-Blanco MT, Verboon JM, Liu R, Watts
JJ and Park-hurst SM: Drosophila embryos close epithelial wounds
using a combination of cellular protrusions and an actomyosin purse
string. J Cell Sci. 125:5984–5997. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Förster D and Luschnig S: Src42A-dependent
polarized cell shape changes mediate epithelial tube elongation in
Drosophila. Nat Cell Biol. 14:526–534. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Veracini L, Grall D, Schaub S, Beghelli-de
la Forest Divonne S, Etienne-Grimaldi MC, Milano G, Bozec A, Babin
E, Sudaka A, Thariat J and Van Obberghen-Schilling E: Elevated Src
family kinase activity stabilizes E-cadherin-based junctions and
collective movement of head and neck squamous cell carcinomas.
Oncotarget. 6:7570–7583. 2014. View Article : Google Scholar
|
|
37
|
Deo DD, Axelrad TW, Robert EG, Marcheselli
V, Bazan NG and Hunt JD: Phosphorylation of STAT-3 in response to
basic fibroblast growth factor occurs through a mechanism involving
platelet-activating factor, JAK-2, and Src in human umbilical vein
endothelial cells. Evidence for a dual kinase mechanism. J Biol
Chem. 277:21237–21245. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Goel R, Phillips-Mason PJ, Raben DM and
Baldassare JJ: alpha-Thrombin induces rapid and sustained Akt
phosphorylation by beta-arrestin1-dependent and -independent
mechanisms, and only the sustained Akt phosphorylation is essential
for G1 phase progression. J Biol Chem. 277:18640–18648. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wu W, Graves LM, Gill GN, Parsons SJ and
Samet JM: Src-dependent phosphorylation of the epidermal growth
factor receptor on tyrosine 845 is required for zinc-induced Ras
activation. J Biol Chem. 277:24252–24257. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Callera GE, Touyz RM, Tostes RC, Yogi A,
He Y, Malkinson S and Schiffrin EL: Aldosterone activates vascular
p38MAP kinase and NADPH oxidase via c-Src. Hypertension.
45:773–779. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Khadaroo RG, He R, Parodo J, Powers KA,
Marshall JC, Kapus A and Rotstein OD: The role of the Src family of
tyrosine kinases after oxidant-induced lung injury in vivo.
Surgery. 136:483–488. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Khadaroo RG, Parodo J, Powers KA, Papia G,
Marshall JC, Kapus A and Rotstein OD: Oxidant-induced priming of
the macrophage involves activation of p38 mitogen-activated protein
kinase through an Src-dependent pathway. Surgery. 134:242–246.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chen Y, Ramakrishnan DP and Ren B:
Regulation of angiogenesis by phospholipid lysophosphatidic acid.
Front Biosci (Landmark Ed). 18:852–861. 2013. View Article : Google Scholar
|
|
44
|
He M, Xue ZM, Li J and Zhou BQ:
Breviscapine inhibits high glucose-induced proliferation and
migration of cultured vascular smooth muscle cells of rats via
suppressing the ERK1/2 MAPK signaling pathway. Acta Pharmacol Sin.
33:606–614. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Makino T, Jinnin M, Muchemwa FC, Fukushima
S, Kogushi-Nishi H, Moriya C, Igata T, Fujisawa A, Johno T and Ihn
H: Basic fibroblast growth factor stimulates the proliferation of
human dermal fibroblasts via the ERK1/2 and JNK pathways. Br J
Dermatol. 162:717–723. 2010. View Article : Google Scholar
|
|
46
|
Yue GG, Fan JT, Lee JK, Zeng GZ, Ho TW,
Fung KP, Leung PC, Tan NH and Lau CB: Cyclopeptide RA-V inhibits
angiogenesis by down-regulating ERK1/2 phosphorylation in HUVEC and
HMEC-1 endothelial cells. Br J Pharmacol. 164:1883–1898. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gazel A, Nijhawan RI, Walsh R and
Blumenberg M: Transcriptional profiling defines the roles of ERK
and p38 kinases in epidermal keratinocytes. J Cell Physiol.
215:292–308. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Boulton TG, Nye SH, Robbins DJ, Ip NY,
Radziejewska E, Morgenbesser SD, DePinho RA, Panayotatos N, Cobb MH
and Yancopoulos GD: ERKs: a family of protein-serine/threonine
kinases that are activated and tyrosine phosphorylated in response
to insulin and NGF. Cell. 65:663–675. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kim MS, Kim YK, Eun HC, Cho KH and Chung
JH: All-trans retinoic acid antagonizes UV-induced VEGF production
and angiogenesis via the inhibition of ERK activation in human skin
keratinocytes. J Invest Dermatol. 126:2697–2706. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Matsubayashi Y, Ebisuya M, Honjoh S and
Nishida E: ERK activation propagates in epithelial cell sheets and
regulates their migration during wound healing. Curr Biol.
14:731–735. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ennis BW, Fultz KE, Smith KA, Westwick JK,
Zhu D, Boluro-Ajayi M, Bilter GK and Stein B: Inhibition of tumor
growth, angiogenesis, and tumor cell proliferation by a small
molecule inhibitor of c-Jun N-terminal kinase. J Pharmacol Exp
Ther. 313:325–332. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Harper EG, Alvares SM and Carter WG:
Wounding activates p38 map kinase and activation transcription
factor 3 in leading keratinocytes. J Cell Sci. 118:3471–3485. 2005.
View Article : Google Scholar : PubMed/NCBI
|