Introduction
A glioblastoma is a tumor of the central nervous
system and the most malignant of all astrocytic tumors, and it is
associated with a poor prognosis (1). Glioblastomas represent 25% of all
malignant nervous system tumors, with an annual incidence of
3/100,000 individuals in the United States, and of <2/100,000
individuals in Europe (1).
Exposure to radiation, second-hand tobacco smoke and rare
hereditary disorders are associated with an increased risk of
glioblastoma (1,2). The tumor consists of poorly
differentiated, neoplastic astrocytes with a diffuse infiltration
pattern, making surgery difficult (3). Deletion of annexin A7 (ANXA7) is
found in 75% of cases (4).
Genetic changes and loss of heterozygosity, epidermal growth factor
receptor (EGFR), p16, tumor protein 53 (TP53) and phosphatase and
tensin homolog (PTEN) are often observed (1). Survival rates are very low, with
<30% at 1 year and 3% at 5 years (1,5).
Currently available treatments have a limited efficacy: a clinical
trial using radiation therapy and temozolomide reported a 5-year
survival rate of 9.8% (6).
It has previously been shown that phosphoglucose
isomerase/autocrine motility factor (PGI/AMF) is important for cell
migration in high-grade glioma (7). AMF/neuroleukin (NLK)/maturation
factor (MF) is an extracellular phosphohexose isomerase (PHI) that
is secreted by malignant cells (8–10).
AMF acts in a cytokine-like manner through the AMF receptor (AMFR),
also known as 78 kDa glycoprotein (gp78), which is a
seven-transmembrane domain glycoprotein (11). AMF stimulates tumor angiogenesis
(12,13), ascites formation (14), apoptotic resistance (15–17), cell proliferation (17,18), matrix metalloproteinase-3 (MMP-3)
secretion (19) and cell motility
(12–14,20). All of these factors are associated
with tumor aggressiveness, invasion and metastatic spread. It has
been demonstrated that the mRNA expression of AMF is higher in
glioblastomas compared with that in astrocytomas, and the overall
survival of patients with AMF-positive glioblastoma is poorer than
in patients with AMF-negative glioblastoma (7).
The aim of the present study was to elucidate the
role that PGI/AMF plays in glioblastoma U87 cells by evaluating the
effect of silencing PGI/AMF expression on migration and sphere
forming ability of the cells, as well as on the side population
cell (SPC) proportion. The results of the present study provide
potential new targets for the treatment of glioblastoma.
Materials and methods
Cell culture
Human glioblastoma U87 cells were purchased from the
Cell Bank of the Chinese Academy of Sciences (Shanghai, China) and
were cultured in Dulbecco's modified Eagle's medium (DMEM) with 10%
heat-inactivated fetal bovine serum (FBS) (both from Gibco, Grand
Island, NY, USA) and 1% penicillin (100 U/ml)/streptomycin (100
µg/ml) at 37°C in an atmosphere with 5% CO2.
RNA interference
Sequences of specific small interfering RNA (siRNA)
targeting PGI/AMF were described previously (20). The target sequence for the PGI/AMF
siRNA was 5′-UGGUACCGCGAGCACCGCUTT-3′ modified with a 5′-FAM
(carboxyfluorescein). The scrambled siRNA sequence used was
5′-ACGUGACACGUUCGGAGAATT-3′ (both from GenePharma Co., Ltd.,
Shanghai, China). Following 24 h of incubation, 50 nM siRNA duplex
transfection was undertaken, into U87 cells using Lipofectamine
2000 (BD Biosciences, San Jose, CA, USA), according to the
manufacturer's instructions. The results were observed directly
under the fluorescence microscope (DM2500 LED; Leica, Wetzlar,
Germany) The efficiency of PGI/AMF silencing was analyzed after 24
h of transfection using reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) and western blot analysis.
Wound healing assay
The U87 cells transfected with PGI/AMF siRNA were
seeded onto 24-well plates 3 days after transfection and were
cultured to at least 95% confluence. Monolayer cells were washed
with serum-free Dulbecco's modified Eagle's medium (DMEM) (Gibco)
and then scraped with a plastic 200 µl pipette tip and
placed back in the incubator at 37°C. Images of the wounded areas
were captured using a CKX41 phase contrast microscope (Olympus,
Tokyo, Japan) at two time points (0 and 8 h after scraping). Images
were analyzed using Adobe Photoshop 7.0 software (Adobe Systems,
Inc., San Jose, CA, USA). The relative migration distance was
calculated using the following formula: relative migration distance
(%) = (A–B)/A × 100%, where A is the width of the wound 0 h after
scraping, and B is the width of the wound 8 h after scraping.
Migration assay
An in vitro migration assay was performed 3
days after transfection using Transwell cell culture chambers (no.
3422; Corning Inc., Corning, NY, USA). The cells were re-suspended
in DMEM supplemented with 0.1% FBS. The cell suspensions
(1×105 cells) were added to the upper compartment of the
chamber. The bottom chamber was filled with DMEM supplemented with
10% FBS. After 8 h of incubation, the topside of the insert
membrane was scrubbed with a cotton swab to remove the non-invasive
cells and the bottom side was fixed with methanol and stained with
1% hematoxylin and eosin (H&E). Migrating cells were counted
under a CKX41 phase contrast microscope at a magnification of ×400
within 10 randomly selected microscope fields.
SPC assay
The SPCs were assessed using previously published
methods (21,22). Briefly, three days after
transfection, the U87 cell suspensions were labeled with Hoechst
33342 dye (Sigma-Aldrich, St. Louis, MO, USA). The U87 cells were
resuspended at 1×106/ml in pre-warmed high-glucose DMEM
with 2% FBS. Hoechst 33342 dye was added at a final concentration
of 5 µg/ml in the presence or absence of reserpine (50
µmol/l) (Aladdin Chemicals Co., Ltd., Shanghai, China) as
controls for the SPC gating, and the cells were incubated at 37°C
for 90 min with intermittent shaking. At the end of the incubation,
the cells were washed with ice-cold PBS with 2% FBS, centrifuged
(200 × g for 5 min) at 4°C, and resuspended in ice-cold Hank's
Balanced Salt Solution (HBSS) containing 2% FBS, in the dark.
Analysis was undertaken using a FACSAria II flow cytometer (BD
Biosciences). The Hoechst 33342 dye was excited at 357 nm and its
fluorescence was analyzed by dual-wavelength measurements (blue,
402–446 nm; red, 650–670 nm).
Sphere formation assay
Three days after transfection, the U87 cells were
diluted in serum-free growth medium (1,000 cells/ml) and plated
(100 µl) in 96-well plates. The cells were cultured with
serum-free growth medium for 14 days. The culture medium consisted
of serum-free DMEM/F12 (Gibco, Invitrogen, Carlsbad, CA, USA)
supplemented with 2% B27 (Invitrogen), human recombinant fibroblast
growth factor 2 (FGF-2, 20 ng/ml) and epidermal growth factor (EGF,
20 ng/ml) (both from PeproTech, Rocky Hill, NJ, USA). After 14
days, the spheres were measured and those >100 µm were
counted as tumorsphere-forming units. The data calculated for the
number and size of the tumorspheres was the average of three
independent experiments. The spheres were counted and measured from
32 different wells/experiment.
RT-qPCR
Twenty-four hours after transfection, total RNA was
isolated from the U87 cells using TRIzol (Takara Bio, Otsu, Japan),
according to the manufacturer's instructions. RNA purity was
assessed by spectrophotometry (OD260/OD280
nm). RNA was transcribed into cDNA using TIANScript M-MLV
reverse transcriptase and the TIANScript RT kit (Tiangen Biotech
(Beijing) Co., Ltd., Beijing, China). cDNA (5 µl) was added
to the 2X Taq PCR Master Mix system (Tiangen Biotech Co., Ltd.).
For quantitative evaluation of the amplified product, 20–40 cycles
of PCR were performed preliminarily to determine the most suitable
number of amplifications for each reaction. PCR cycling conditions
consisted of 30 sec at 95°C, 30 sec at 55°C, and 1 min at 72°C for
PGI/AMF; and 30 sec at 95°C, 30 sec at 60°C, and 1 min at 72°C for
β-actin. PGI/AMF and β-actin were amplified using the following
primers: PGI/AMF forward, 5′-AATGCAGAGACGGCGAAGGAG-3′ and reverse,
5′-ACGAGAAGAGAAAGGGGAGTC-3′; β-actin forward,
5′-TGGCACCCAGCACAATGAA-3′ and reverse,
5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′. Amplification of β-actin was used
to estimate the efficiency of cDNA synthesis and to normalize
PGI/AMF expression. The amplified products were separated on 1%
agarose gel electrophoresis. The gel was stained with ethidium
bromide and images were captured using a camera (Panasonic, Kadoma,
Japan). The products were quantified by analyzing the band density
using Quantity One V4.6.2 software (Bio-Rad, Hercules, CA,
USA).
Protein extraction and western blot
analysis
Twenty-four hours after transfection, the U87 cells
were washed twice with PBS and lysed with 500 µl cell lysis
buffer for western blot analysis, supplemented with PMSF (both from
Beyotime, Jiangsu, China). The supernatant from the lysate was
collected after centrifugation (12,500 × g for 20 min at 4°C). The
cell supernatants were supplemented with PMSF and concentrated at
over 100-fold with a centrifugal evaporator (CVE-200D; (Eyela,
Tokyo, Japan), and the protein concentrations in the supernatant
were determined using a BCA assay. Equal amounts of protein (40 or
50 µg) were subjected to 10% SDS-PAGE and transferred onto
PVDF membranes (pore size, 0.45 µm) (Millipore, Billerica,
MA, USA). The membranes were blocked with 5% non-fat skimmed milk
in PBS for 1 h at room temperature and incubated with primary
antibodies. The following antibodies were used for western blot
analysis: rabbit anti-Akt (#9272; 1:1,000) and rabbit anti-p-Akt
(#9271; 1:1,000; both from Cell Signaling Technology, Danvers, MA,
USA), goat anti-PGI/AMF (sc-30392; 1:2,000; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA), mouse anti-SRY (sex
determining region Y)-box 2 (SOX2) (S1451; 1:1,000; Sigma-Aldrich)
or mouse anti-β-actin (TA-09; 1:1,000; ZSGB-Bio, Beijing, China)
overnight at 4°C. The membranes were processed with horseradish
peroxidase (HRP)-conjugated secondary antibodies [goat anti-mouse
IgG (ZB-5305), ZSGB-Bio; goat anti-rabbit IgG (SA00001-2),
Proteintech Group, Inc., Chicago, IL, USA; or donkey anti-goat IgG
(sc-2020), Santa Cruz Biotechnology, Inc.]. Immunoreactive bands
were visualized using enhanced chemiluminescence (ECL) western
blotting detection reagents (P0018; Beyotime, Beijing, China). The
blots were stripped and re-developed with β-actin antibody to
normalize data. Immunodetection was followed by visualization using
ECL, and densitometry using Quantity One V4.6.2 software
(Bio-Rad).
Statistical analysis
Data for each group are represented as the means ±
standard deviation (SD) from three independent experiments. Data
were evaluated using the unpaired t test and GraphPad Prism 5.0
(GraphPad Software Inc., San Diego, CA, USA). A two-sided P-value
<0.05 was considered to indicate a statistically significant
difference.
Results
Silencing of PGI/AMF in U87 cells
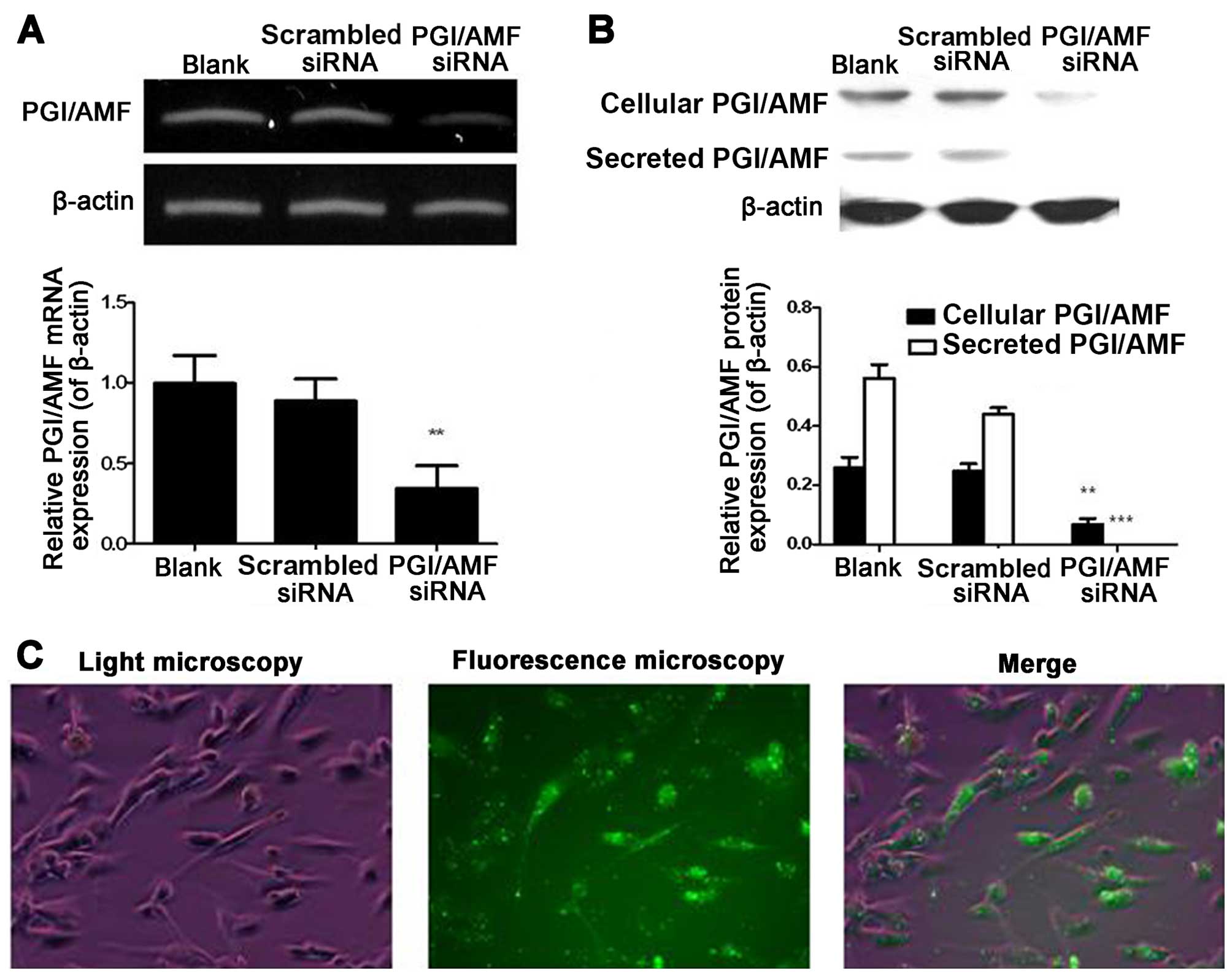
RT-qPCR and western blot analysis showed that
PGI/AMF mRNA and protein expression levels in U87 cells were
decreased by PGI/AMF siRNA (both P<0.01) (Fig. 1). Of note, the silencing of
endogenous PGI/AMF by siRNA led to the complete inhibition of
PGI/AMF secretion (Fig. 1B).
There was no statistical difference in mRNA and protein expression
between the untreated cells and the cells transfected with
scrambled siRNA (both P>0.05). Transfection efficiency, shown as
GFP-positive cells, was observed under a fluorescence microscope
(Fig. 1C).
Inhibition of wound healing capacity and
migration ability of U87 cells by PGI/AMF siRNA
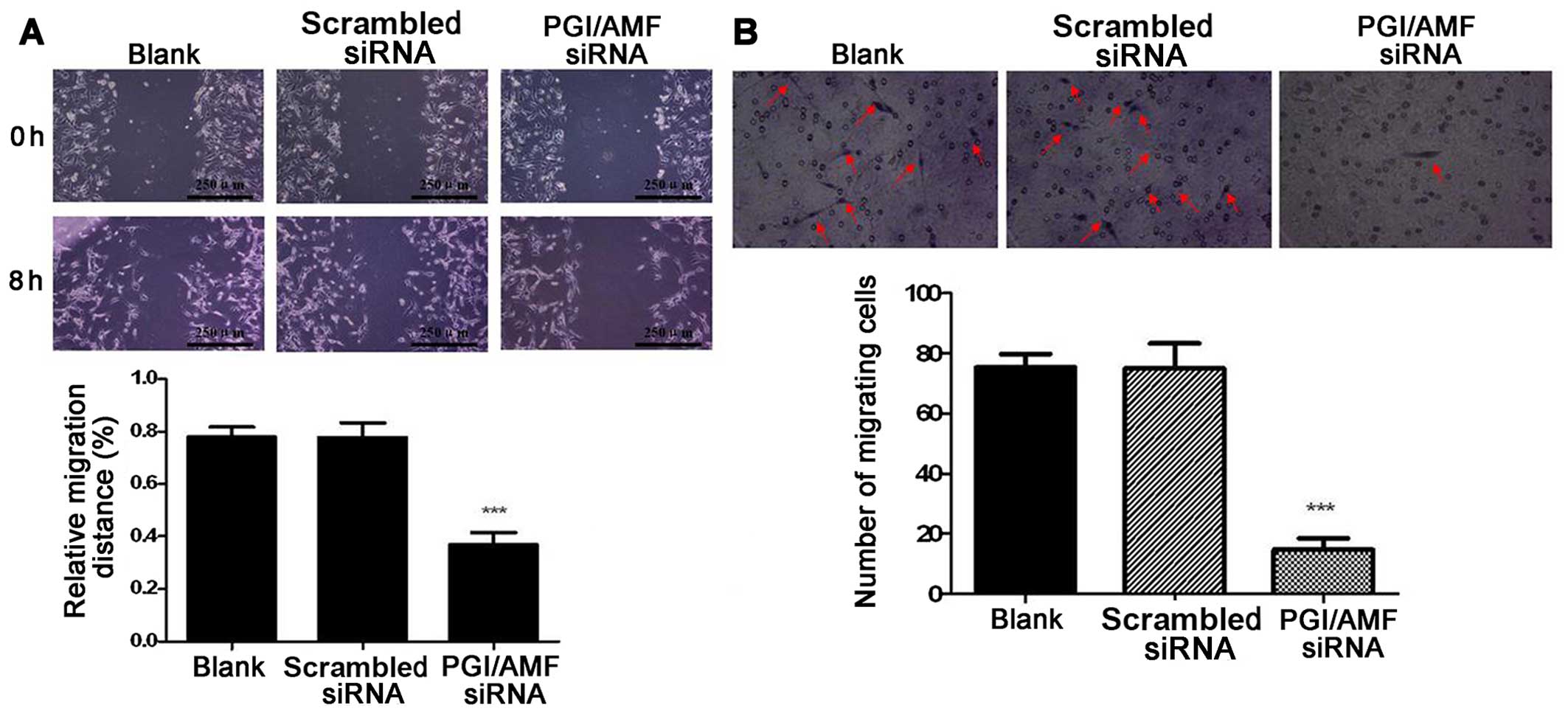
Wound healing capacity was significantly inhibited
by 52.6% in the cells transfected with PGI/AMF siRNA compared with
those transfected with scrambled siRNA (P<0.001) (Fig. 2A). The Transwell assay showed that
the silencing of PGI/AMF with siRNA significantly decreased the
number of migrating cells (−80.4% vs. scrambled siRNA, P<0.001)
(Fig. 2B).
Silencing of PGI/AMF reduces the ratio of
SPCs
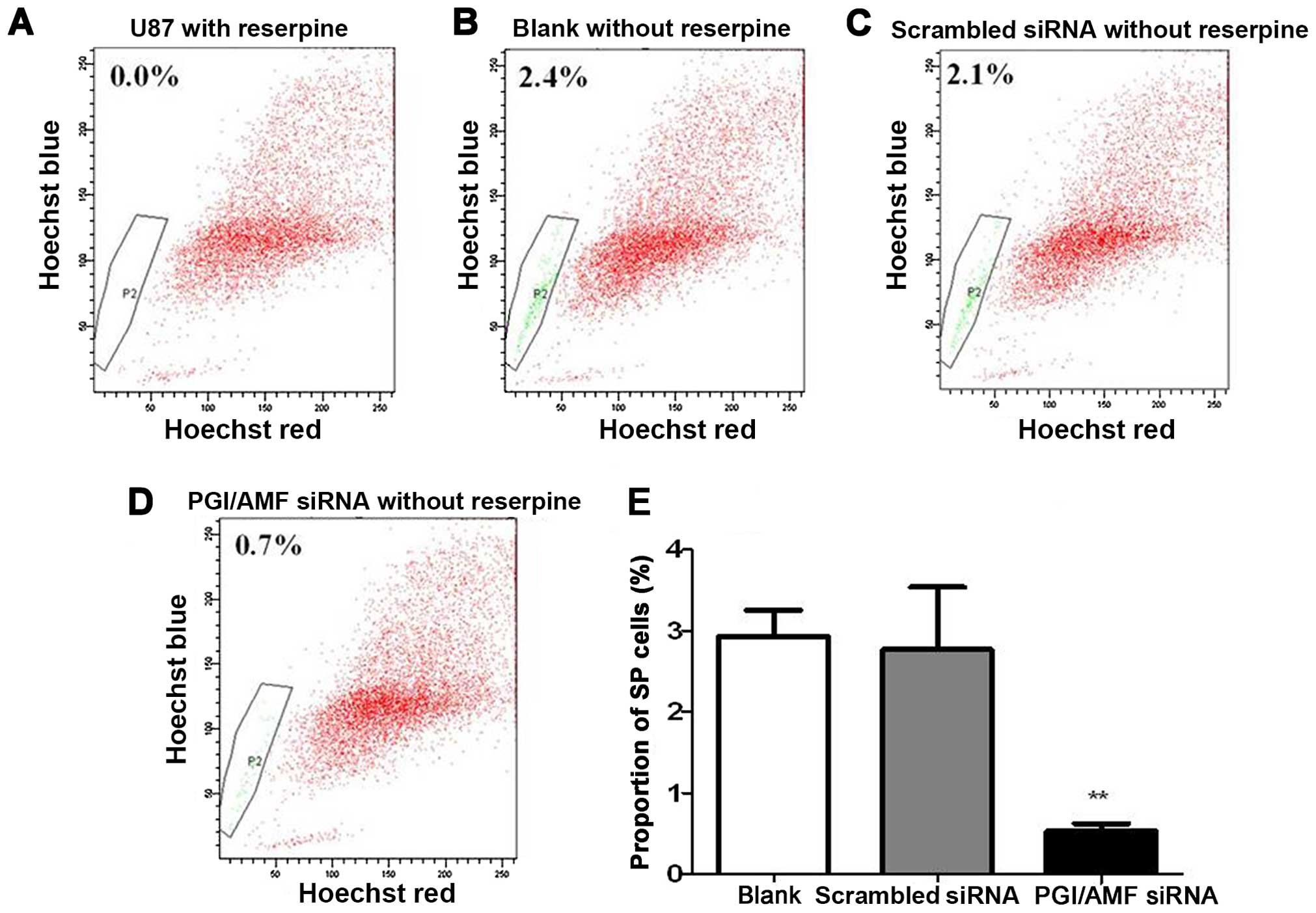
As shown in Figure
3, flow cytometric analysis revealed that the proportion of
SPCs was decreased by 80.7% or decreased to 19.3% in cells
transfected with PGI/AMF siRNA compared with those which had been
transfected with scrambled siRNA, 72 h after silencing of PGI/AMF
(P<0.01) (Fig. 3).
Silencing of PGI/AMF reduces tumorsphere
formation
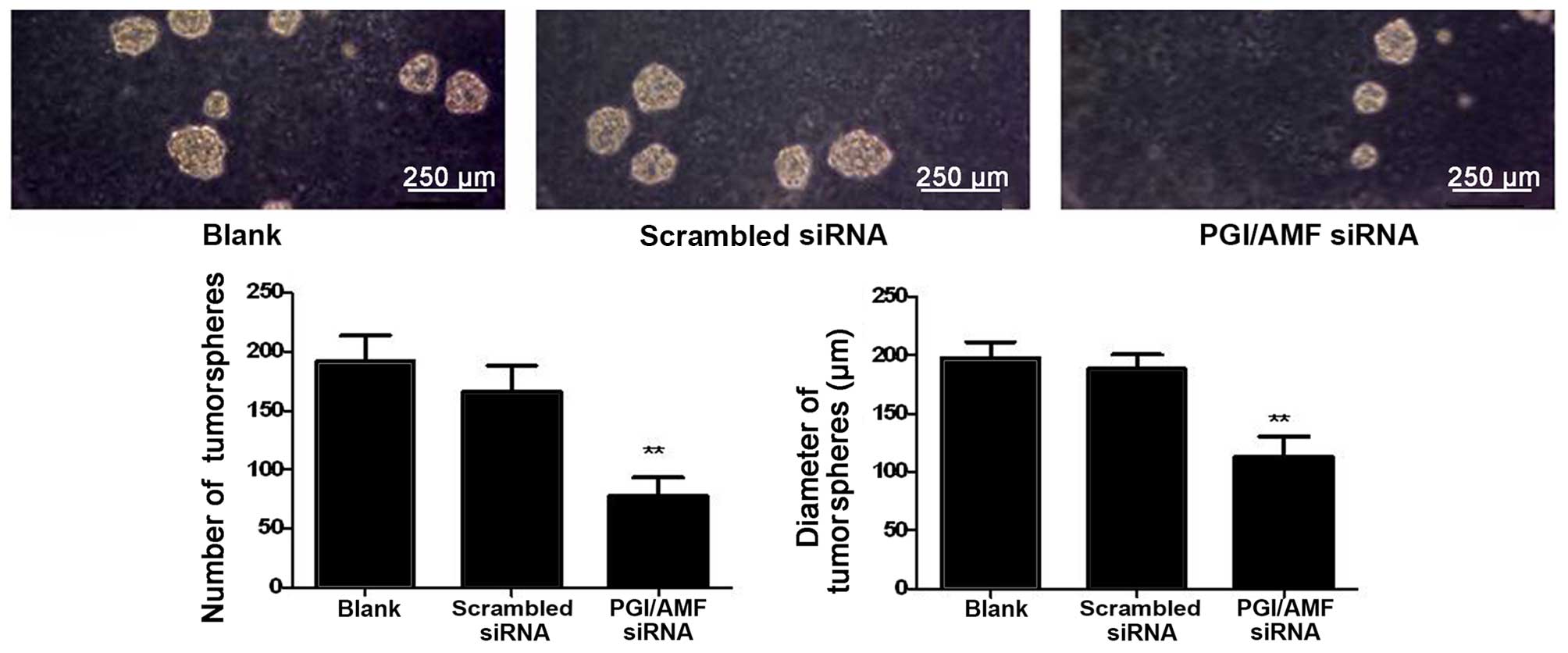
The U87 cells transfected with PGI/AMF siRNA formed
53.1% fewer tumorspheres compared with the control cells
transfected with scrambled siRNA (P<0.01) (Fig. 4). In addition, the size of the
tumorspheres was significantly reduced in the cells transfected
with PGI/AMF siRNA compared with the control cells (113.0±17.3 vs.
197.3±14.0 and 188.0±12.3 µm, P<0.01) (Fig. 4).
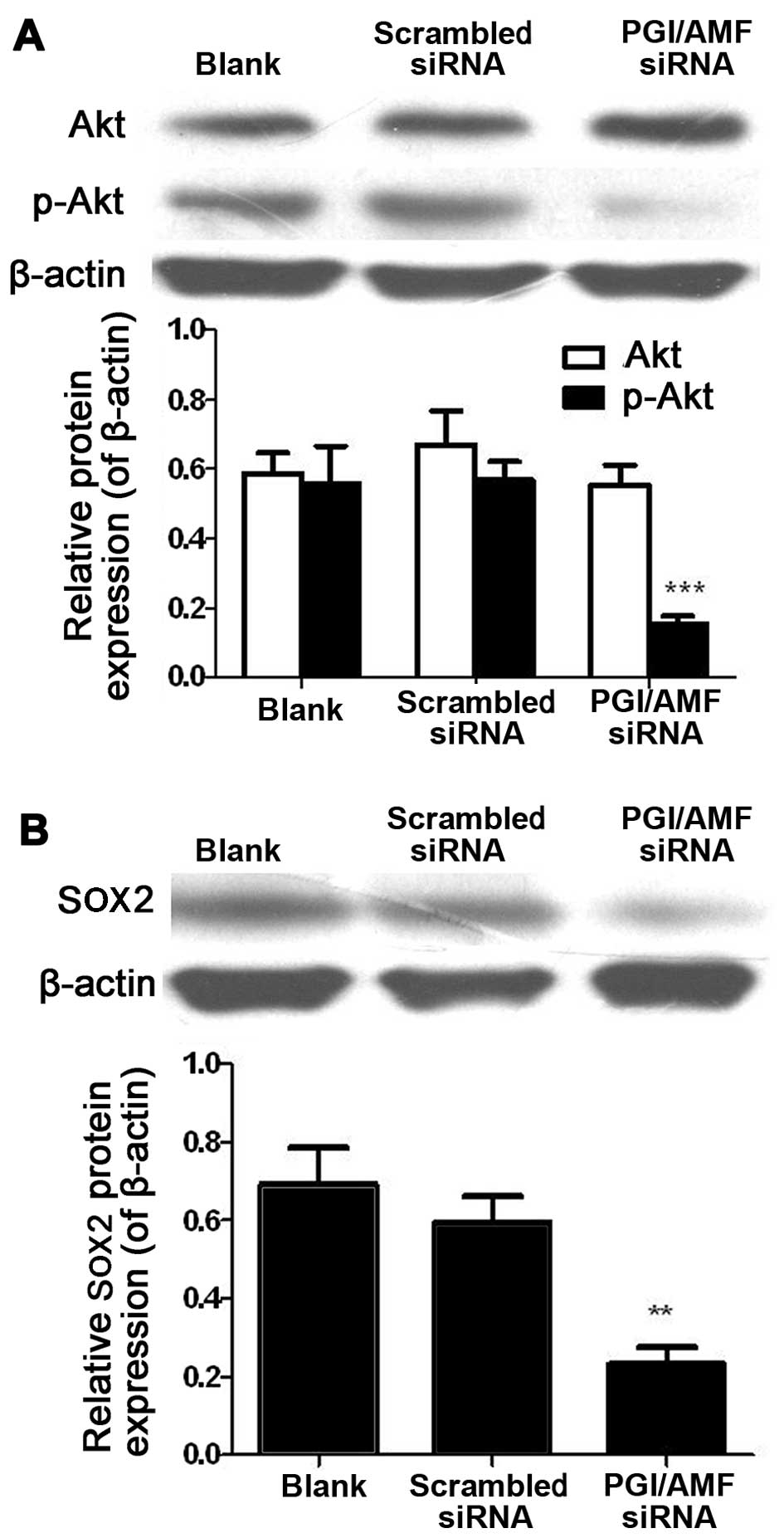
Silencing of PGI/AMF decreases the
phosphorylation level of Akt and the expression of SOX2 in U87
cells
Akt expression levels were similar in the U87 cells
transfected with PGI/AMF siRNA and in the control cells (both
P>0.05). However, the levels of p-Akt were markedly decreased
(−71.9%, P<0.001) (Fig. 5A).
SOX2 levels were also significantly decreased in the cells
transfected with PGI/AMF siRNA compared with scrambled siRNA
(−61.7%, P<0.01) (Fig.
5B).
Discussion
PGI/AMF has been reported to regulate the
proliferation and the survival of cells, and to prevent
stress-induced apoptosis of tumor cells (15,17,20). In the present study, silencing of
PGI/AMF inhibited the migration ability of the glioblastoma U87
cells, decreased the proportion of SPCs, decreased sphere formation
ability, and decreased the levels of p-Akt and the stemness marker,
SOX2.
Previous research has shown that adult stem cells
can be identified by an SPC phenotype. The SPCs, first described by
Goodell et al (23), are a
small subpopulation of cells presenting with enriched stem cell
activity and a distinctively low Hoechst 33342 dye-staining
pattern. Subsquent studies attributed this phenotype to the
expression of ATP-binding cassette sub-family G member 2 (ABCG2),
an ATP-binding cassette (ABC) transporter (24). Other studies have demonstrated the
presence of SPCs in human cancers of different origins including
acute myelogenous leukemia, neuroblastoma and glioma (21,25–27). SPCs are of great clinical
significance. Indeed, previous research has shown that these cells
are resistant to many drugs (25), contributing to the resistance of
tumors to chemotherapy. Thus, decreasing the proportion of SPCs
within tumors is a viable option for improving the effectiveness of
chemotherapy, particularly in aggressive tumors such as
glioblastomas. The present study suggests that silencing PGI/AMF is
a way to decrease the proportion of SPCs in glioblastomas.
It is now widely accepted that tumors contain a
mixed population of cells at various stages of differentiation, of
which only a fraction can perpetuate the tumor; these cells are
named cancer stem cells (28,29). However, given the impossibility of
directly identifying them due to a lack of specific markers, these
cells are operationally defined as the small fraction of cancer
cells that have the ability to propagate the tumor upon
transplantation into immunodeficient mice (28,29). The xenotransplanted tumor may be
serially transplanted into new recipient mice, highlighting the
capacity for perpetual self-renewal of this subset of cancer cells.
These cells share genetic and phenotypic features with normal
neural stem cells such as the expression of the transcription
factor, SOX2 (30,31). SOX2 plays an essential role in
maintaining the proliferative potential of neural stem/precursor
cells and in ensuring the production of sufficient cell numbers of
the appropriate type (32–34).
The results of the present study demonstrated that silencing of
PGI/AMF decreased the expression of SOX2, suggesting that PGI/AMF
signaling is involved in cancer stem cell proliferation and
malignant functions.
The serine/threonine kinase, Akt/protein kinase B
pathway is a nodal point regulating a number of tumor-associated
processes, including cell growth, cell cycle progression, survival,
migration and angiogenesis, and has been shown to be important in
many malignancies including glioblastoma (17,35–37). More specifically, the Akt pathway
has been shown to be activated in the majority of glioblastoma
multiforme tumors (35,36). In other research, activation of
the Akt pathway in a human astrocytic model of glioma resulted in
the conversion of anaplastic astrocytoma to glioblastoma multiforme
(37). In the present study,
although the total expression level of Akt was almost unchanged in
the silenced PGI/AMF cells compared with the control cells, the
p-Akt level was decreased. These results are supported by a
previous study showing that the overexpression of PGI/AMF in
NIH-3T3 fibroblasts led to an accumulation of cellular p-Akt
protein (17). These changes may
have led to the reduced aggressiveness of the cells treated with
silenced PGI/AMF. However, the present study did not explore the
causal relationship between PGI/AMF silencing and the Akt pathway.
More studies are necessary to correctly establish the chain of
events.
In conclusion, the silencing of PGI/AMF decreases
migration, tumorsphere formation and the proportion of side
population cells in glioblastoma U87 cells. We suggest that the Akt
pathway is involved. These results provide a potential new target
for the treatment of glioblastoma.
References
|
1
|
Brandes AA, Tosoni A, Franceschi E, Reni
M, Gatta G and Vecht C: Glioblastoma in adults. Crit Rev. Oncol
Hematol. 67:139–152. 2008.
|
|
2
|
Stupp R, Brada M, van den Bent MJ, Tonn JC
and Pentheroudakis G: High-grade glioma: ESMO Clinical Practice
Guidelines for diagnosis, treatment and follow-up. Ann Oncol.
25(Suppl 3): iii93–iii101. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sanai N, Alvarez-Buylla A and Berger MS:
Neural stem cells and the origin of gliomas. N Engl J Med.
353:811–822. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yadav AK, Renfrow JJ, Scholtens DM, Xie H,
Duran GE, Bredel C, Vogel H, Chandler JP, Chakravarti A, Robe PA,
et al: Monosomy of chromosome 10 associated with dysregulation of
epidermal growth factor signaling in glioblastomas. JAMA.
302:276–289. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stupp R, Hegi ME, Mason WP, van den Bent
MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B,
Belanger K, et al European Organisation for Research and Treatment
of Cancer Brain Tumour and Radiation Oncology Groups: National
Cancer Institute of Canada Clinical Trials Group: Effects of
radiotherapy with concomitant and adjuvant temozolomide versus
radiotherapy alone on survival in glioblastoma in a randomised
phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet
Oncol. 10:459–466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tanizaki Y, Sato Y, Oka H, Utsuki S, Kondo
K, Miyajima Y, Nagashio R and Fujii K: Expression of autocrine
motility factor mRNA is a poor prognostic factor in high-grade
astrocytoma. Pathol Int. 56:510–515. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Watanabe H, Takehana K, Date M, Shinozaki
T and Raz A: Tumor cell autocrine motility factor is the
neuroleukin/phosphohexose isomerase polypeptide. Cancer Res.
56:2960–2963. 1996.PubMed/NCBI
|
|
9
|
Niinaka Y, Paku S, Haga A, Watanabe H and
Raz A: Expression and secretion of neuroleukin/phosphohexose
isomerase/maturation factor as autocrine motility factor by tumor
cells. Cancer Res. 58:2667–2674. 1998.PubMed/NCBI
|
|
10
|
Haga A, Niinaka Y and Raz A: Phosphohexose
isomerase/autocrine motility factor/neuroleukin/maturation factor
is a multifunctional phosphoprotein. Biochim Biophys Acta.
1480:235–244. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shimizu K, Tani M, Watanabe H, Nagamachi
Y, Niinaka Y, Shiroishi T, Ohwada S, Raz A and Yokota J: The
autocrine motility factor receptor gene encodes a novel type of
seven transmembrane protein. FEBS Lett. 456:295–300. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Funasaka T, Haga A, Raz A and Nagase H:
Tumor autocrine motility factor is an angiogenic factor that
stimulates endothelial cell motility. Biochem Biophys Res Commun.
285:118–128. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Funasaka T, Haga A, Raz A and Nagase H:
Autocrine motility factor secreted by tumor cells upregulates
vascular endothelial growth factor receptor (Flt-1) expression in
endothelial cells. Int J Cancer. 101:217–223. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Funasaka T, Haga A, Raz A and Nagase H:
Tumor autocrine motility factor induces hyperpermeability of
endothelial and mesothelial cells leading to accumulation of
ascites fluid. Biochem Biophys Res Commun. 293:192–200. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Haga A, Funasaka T, Niinaka Y, Raz A and
Nagase H: Autocrine motility factor signaling induces tumor
apoptotic resistance by regulations Apaf-1 and Caspase-9 apoptosome
expression. Int J Cancer. 107:707–714. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Romagnoli A, Oliverio S, Evangelisti C,
Iannicola C, Ippolito G and Piacentini M: Neuroleukin inhibition
sensitises neuronal cells to caspase-dependent apoptosis. Biochem
Biophys Res Commun. 302:448–453. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsutsumi S, Hogan V, Nabi IR and Raz A:
Overexpression of the autocrine motility factor/phosphoglucose
isomerase induces transformation and survival of NIH-3T3
fibroblasts. Cancer Res. 63:242–249. 2003.PubMed/NCBI
|
|
18
|
Tsutsumi S, Yanagawa T, Shimura T,
Fukumori T, Hogan V, Kuwano H and Raz A: Regulation of cell
proliferation by autocrine motility factor/phosphoglucose isomerase
signaling. J Biol Chem. 278:32165–32172. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu FL, Liao MH, Lee JW and Shih WL:
Induction of hepatoma cells migration by phosphoglucose
isomerase/autocrine motility factor through the upregulation of
matrix metalloproteinase-3. Biochem Biophys Res Commun. 314:76–82.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Funasaka T, Hu H, Yanagawa T, Hogan V and
Raz A: Downregulation of phosphoglucose isomerase/autocrine
motility factor results in mesenchymal-to-epithelial transition of
human lung fibrosarcoma cells. Cancer Res. 67:4236–4243. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Feuring-Buske M and Hogge DE: Hoechst
33342 efflux identifies a subpopulation of cytogenetically normal
CD34+CD38− progenitor cells from patients
with acute myeloid leukemia. Blood. 97:3882–3889. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim MC, D'Costa S, Suter S and Kim Y:
Evaluation of a side population of canine lymphoma cells using
Hoechst 33342 dye. J Vet Sci. 14:481–486. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Goodell MA, Brose K, Paradis G, Conner AS
and Mulligan RC: Isolation and functional properties of murine
hematopoietic stem cells that are replicating in vivo. J Exp Med.
183:1797–1806. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou S, Schuetz JD, Bunting KD, Colapietro
AM, Sampath J, Morris JJ, Lagutina I, Grosveld GC, Osawa M,
Nakauchi H and Sorrentino BP: The ABC transporter Bcrp1/ABCG2 is
expressed in a wide variety of stem cells and is a molecular
determinant of the side-population phenotype. Nat Med. 7:1028–1034.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hirschmann-Jax C, Foster AE, Wulf GG,
Nuchtern JG, Jax TW, Gobel U, Goodell MA and Brenner MK: A distinct
'side population' of cells with high drug efflux capacity in human
tumor cells. Proc Natl Acad Sci USA. 101:14228–14233. 2004.
View Article : Google Scholar
|
|
26
|
Kondo T, Setoguchi T and Taga T:
Persistence of a small subpopulation of cancer stem-like cells in
the C6 glioma cell line. Proc Natl Acad Sci USA. 101:781–786. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wulf GG, Wang RY, Kuehnle I, Weidner D,
Marini F, Brenner MK, Andreeff M and Goodell MA: A leukemic stem
cell with intrinsic drug efflux capacity in acute myeloid leukemia.
Blood. 98:1166–1173. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vermeulen L, Sprick MR, Kemper K, Stassi G
and Medema JP: Cancer stem cells - old concepts, new insights. Cell
Death Differ. 15:947–958. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Singh SK, Clarke ID, Terasaki M, Bonn VE,
Hawkins C, Squire J and Dirks PB: Identification of a cancer stem
cell in human brain tumors. Cancer Res. 63:5821–5828.
2003.PubMed/NCBI
|
|
30
|
Boumahdi S, Driessens G, Lapouge G, Rorive
S, Nassar D, Le Mercier M, Delatte B, Caauwe A, Lenglez S, Nkusi E,
et al: SOX2 controls tumour initiation and cancer stem-cell
functions in squamous-cell carcinoma. Nature. 511:246–250. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Brafman DA, Moya N, Allen-Soltero S,
Fellner T, Robinson M, McMillen ZL, Gaasterland T and Willert K:
Analysis of SOX2-expressing cell populations derived from human
pluripotent stem cells. Stem Cell Rep. 1:464–478. 2013. View Article : Google Scholar
|
|
32
|
Bani-Yaghoub M, Tremblay RG, Lei JX, Zhang
D, Zurakowski B, Sandhu JK, Smith B, Ribecco-Lutkiewicz M, Kennedy
J, Walker PR, et al: Role of Sox2 in the development of the mouse
neocortex. Dev Biol. 295:52–66. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ferri AL, Cavallaro M, Braida D, Di
Cristofano A, Canta A, Vezzani A, Ottolenghi S, Pandolfi PP, Sala
M, DeBiasi S and Nicolis SK: Sox2 deficiency causes
neurodegeneration and impaired neurogenesis in the adult mouse
brain. Development. 131:3805–3819. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Graham V, Khudyakov J, Ellis P and Pevny
L: SOX2 functions to maintain neural progenitor identity. Neuron.
39:749–765. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Holland EC, Celestino J, Dai C, Schaefer
L, Sawaya RE and Fuller GN: Combined activation of Ras and Akt in
neural progenitors induces glioblastoma formation in mice. Nat
Genet. 25:55–57. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rajasekhar VK, Viale A, Socci ND, Wiedmann
M, Hu X and Holland EC: Oncogenic Ras and Akt signaling contribute
to glioblastoma formation by differential recruitment of existing
mRNAs to polysomes. Mol Cell. 12:889–901. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sonoda Y, Ozawa T, Aldape KD, Deen DF,
Berger MS and Pieper RO: Akt pathway activation converts anaplastic
astrocytoma to glioblastoma multiforme in a human astrocyte model
of glioma. Cancer Res. 61:6674–6678. 2001.PubMed/NCBI
|