Introduction
Obesity is associated with metabolic syndrome, a
group of risk factors for cardiovascular disease, type 2 diabetes
mellitus (T2DM) and other pathologies, including hypertension,
hyperlipidemia, coronary artery disease and cancers (1,2). A
worldwide survey conducted in 2013, found that the body mass index
(BMI) of adults had increased by 36.9% in men and by 38.0% in women
(3). Many factors play a role in
increasing the prevalence of obesity, including technological
developments in the food industry and the prevalence of fast food
(4). Moreover, obesity,
specifically abdominal obesity, causes serum adipokine imbalance,
insulin resistance and endothelial dysfunction (5). It is associated with dyslipidemia as
well as an increase in the ratio of total cholesterol (TC) to
high-density lipoprotein cholesterol (HDL-C) (6). A study performed on mice determined
that the consumption of a high-fat diet (HFD) resulted in elevated
levels of serum cholesterol and insulin, specifically fasting
insulin and pro-insulin (7).
Likewise, HFD-fed mice have been shown to have lower superoxide
dismutase (SOD) levels compared with normal mice (8), and obese mice have been shown to
have increased triglyceride (TG) and glucose levels, and
hypertension after 8 weeks on a HFD (9). HFDs are linked with both leptin and
insulin resistance (10), and
importantly, are associated with hepatic steatosis. Moreover,
HFD-fed mice had increased levels of serum glutamic oxaloacetic
transaminase (GOT) and glutamic-pyruvate transaminase (GPT)
compared with mice fed a normal diet (ND) and those fed HFD
supplemented with α-lipoic acid for 24 weeks (11). In addition, it has been shown that
HFD-fed mice have greater adipocyte hypertrophy and increased
levels of inflammatory cytokines [monocyte chemoat-tractant protein
(MCP)-1 and tumor necrosis factor (TNF)-α], as well as hepatic
steatosis (12). Catalase (CAT)
activity is significantly upregulated after the consumption of a
HFD; an increased CAT protein content translates to increased
enzyme activity in the mouse heart (13).
Tuna contains bioactive peptides formed from
enzymatically hydrolyzed byproducts (14). Hydrolyzed byproducts from tuna
dark muscle have been shown to exert anticancer effects on MCF-7
cells, with peptide fractions ranging from 390 to 1400 Da, having
the highest anti-proliferative activity (15).
Previous studies using western blot analysis have
revealed the anti-obesity effects of peptides derived from tuna
proteins, which have been shown to significantly reduce the
expression levels of the adipocyte marker genes,
CCAAT/enhancer-binding protein (C/EBP)α and peroxisome
proliferator-activated receptor-γ (PPAR-γ) (16). Moreover, these results were
supported by the upregulation of Wnt-10b mRNA expression and the
activation of the Wnt signaling pathway in 3T3-L1 adipocyte cells
(17). The Wnt signaling pathway
plays a key role in the differentiation of adipocytes and
downregulates the expression of adipocyte-specific genes (18). A previous study demonstrated that
the peptides present in boiled tuna extract inhibit the
differentiation of 3T3-L1 cells in vitro, and highlighted
the effectiveness of this model for metabolism and obesity research
(17). Moreover, these peptides
can decrease the expression levels of adipogenic genes (17,18). Based on these data, we were
interested in exploring the anti-obesity effects of boiled tuna
extract in vivo in C57BL/6N mice. We hypothesized that tuna
may improve body weight and affect metabolic systems. In this
study, using serum and western blot analyses, we identified the
genes responsible for the effects of tuna extract on obesity. Blood
and liver tissues were assessed in order to measure the levels of
TC, HDL-C, low-density lipoprotein cholesterol (LDL-C), insulin,
leptin, TG, glucose, CAT, SOD, GOT, GPT, aspartate transaminase
(AST) and alanine transaminase (ALT).
Materials and methods
Preparation of boiled tuna extract
The desalinated boiled tuna extract used was
prepared in Korea in 2015. First, boiled tuna extract was
centrifuged (10,000 rpm, 30 min, 22°C) to remove any suspended
solids that may interfere with the desalting step. This process
involves a change from 55 Brix, 13% salinity to 45 Brix, 12%
salinity. We performed membrane filtration (membrane 2319/size 200
Da) on the desalinated boiled tuna extract. We finally obtained a
tuna extract of 30 Brix, 1% salinity, which was subjected to heat
exchanger-type momentary sterilization (conditions: 110°C, 10 sec).
The tuna extract sample was then transferred to 1.5 ml tubes and
stored at −70°C until use.
Animals and diets
Male 5-week-old C57BL/6N mice were purchased from
Samtako Bio Korea Co. (Gyeonggi-do, Korea). The animal care and use
protocol in this study was approved by the Institutional Animal
Care and Use Committee (IACUC) of Pukyong National University,
Busan, Korea (Approval no. 2015-04). They were maintained on a 12-h
light-dark cycle for 1 week prior to the experiments, and were
housed in cages under controlled room temperature (22±2°C) and
humidity (55±5%). Mice had ad libitum access to a
commercially available diet (Samtako Bio Korea Co.) and water.
After 1 week, the feed was changed to the AIN-76 semi-purified diet
(MP0290545220; MP Biomedicals, LLC, Solon, OH, USA) with added lard
and corn oil to induce obesity for 10 weeks. The mice were divided
into 5 groups as follows: i) the ND group (n=10); the ii) HFD group
(n=10); iii) the group fed a HFD and 100 mg/kg boiled tuna extract
(HFD + T100) (n=10); iv) mice fed a HFD and 200 mg/kg boiled tuna
extract (HFD + T200) (n=10); and v) the group fed a HFD and 400
mg/kg boiled tuna extract group (HFD + T400) (n=10). The diet
composition is shown in Table
I.
 | Table IComposition of the experimental diets
(mg/kg/day). |
Table I
Composition of the experimental diets
(mg/kg/day).
| Component | Control | HFD | HFD + T100 | HFD + T200 | HFD + T400 |
|---|
| Casein | 875.0 | 742.0 | 741.1 | 741.1 | 741.1 |
| DL-methionine | 13.1 | 11.1 | 11.1 | 11.1 | 11.1 |
| Sucrose | 2187.5 | 1855 | 1852.8 | 1852.8 | 1852.8 |
| Corn starch | 656.3 | 556.5 | 555.8 | 555.8 | 555.8 |
| Cellulose | 218.8 | 185.5 | 185.3 | 185.3 | 185.3 |
| Corn oil | 218.8 | 185.5 | 185.3 | 185.3 | 185.3 |
| Mineral mix | 153.1 | 129.9 | 129.7 | 129.7 | 129.7 |
| Vitamin mix | 43.8 | 37.1 | 37.1 | 37.1 | 37.1 |
| Choline
bitartate | 8.8 | 7.4 | 7.4 | 7.4 | 7.4 |
| Lard | 0 | 630.7 | 630.0 | 630.0 | 630.0 |
| Cholesterol | 0 | 37.1 | 37.1 | 37.1 | 37.1 |
| Boiled tuna
powder | 0 | 0 | 4.375 | 8.75 | 17.5 |
| Total | 4375.0 | 4377.8 | 4377.0 | 4381.4 | 4390.1 |
| Calories | 16843.8 | 19959.8 | 19936.3 | 19936.3 | 19936.3 |
| Fat | 1968.8 | 7345.8 | 7337.1 | 7337.1 | 7337.1 |
| Fat (%
calorie) | 11.7 | 36.8 | 36.8 | 36.8 | 36.8 |
Analysis of blood
Mice were anesthetized with ether, and blood was
obtained by the intraorbital vein collection method. Blood samples
were centrifuged at 2,500 × g for 15 min at 4°C, and subsequently
stored at −70°C. Enzyme kits were used to measure the serum levels
of TC, HDL-C, TG, glucose (Asan Pharmaceutical Co., Ltd., Gyeonggi,
Korea), LDL-C (Cusabio Biotech Co., Ltd., Wuhan, China), leptin
(Enzo Life Sciences, Inc., Farmingdale, NY, USA), insulin (Alpco
Diagnostics, Windham, NH, USA), ALT and AST (both from
Sigma-Aldrich, St. Louis, MO, USA), according to the manufacturer's
instructions at an absorbance of 490 nm using a Benchmark
enzyme-linked immunosorbent assay (ELISA) plate reader (Bio-Rad
Laboratories, Hercules, CA, USA).
Analysis of liver tissue
Mice were anesthetized with ether, and liver tissues
were obtained. Liver tissue samples (0.2 g) were homogenized in 1
ml of phosphate-buffered saline (PBS), and the samples were
centrifuged at 2,500 × g for 15 min at 4°C and stored at −70°C. The
levels of CAT, SOD (Arbor Assays, Ann Arbor, MI, USA), GOT and GPT
(Asan Pharmaceutical Co., Ltd.) were measured using enzyme kits,
according to the manufacturer's instructions at an absorbance at
490 nm using a Benchmark ELISA plate reader (Bio-Rad
Laboratories).
Western blot analysis
The liver tissues were washed with PBS and lysis
buffer [20 mM Tris base (pH 8.0), 150 mM NaCl, 100 μM sodium
vanadate, 100 μM ammonium molybdate, 10% (v/v) glycerol,
0.1% (v/v) Nonidet P-40, 0.1% (w/v) SDS, 1 mM glycerophosphate, 1
μg/ml aprotinin, 1 μg/ml leupeptin, 1 μg/ml
pepstatin A, and 1 mM phenylmethanesulfonyl fluoride (PMSF)] was
added. Proteins were separated by 7–15% (w/v) SDS-PAGE and
transferred onto polyvinylidene fluoride membranes (Millipore,
Billerica, MA, USA). The membranes were blocked at room temperature
with 1% (w/v) bovine serum albumin in TBS-T [10 mM Tris-HCl (pH
7.5), 150 mM NaCl, 0.1% (v/v) Tween-20] and incubated on a shaker
with the following antibodies: anti-C/EBPα (sc-9314, anti-rabbit;
1:1,000), anti-C/EBPβ (sc-150, anti-rabbit; 1:1,000), anti-C/EBPδ
(sc-151, anti-rabbit; 1:1,000), anti-PPAR-γ (sc-1984, anti-goat;
1:1,000), anti-CD36 (sc-7641, anti-goat; 1:1,000), anti-lipoprotein
lipase (LPL) (sc-32382, anti-goat; 1:1,000), anti-sterol regulatory
element-binding protein-1 (SREBP-1) (sc-366, anti-rabbit; 1:1,000),
anti-fatty acid synthase (FAS) (sc-7886, anti-mouse; 1:1,000),
anti-acetyl-CoA carboxy-acetyl-CoA carboxylase (ACC) (sc-271965,
anti-mouse; 1:1,000), anti-fatty acid binding protein (FABP)
(sc-18661, anti-goat; 1:1,000), anti-glucose transporter type 4
(Glut4) (sc-1606, anti-goat; 1:1,000), anti-glyceraldehyde
3-phosphate dehydrogenase (GAPDH) (sc-25778, anti-rabbit; 1:1,000)
(all from Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). The
secondary antibodies used were peroxidase-conjugated goat
(sc-2741), mouse (sc-2032), or rabbit (sc-2031) antibodies
(1:10,000; all from GE Healthcare Bio-Sciences, Piscataway, NJ,
USA). Proteins were visualized using the SuperSignal West Pico
Stable Peroxide solution and the SuperSignal West Pico
Luminol/Enhancer solution (both from Thermo Fisher Scientific,
Inc., Rockford, IL, USA) and Kodak X-ray film.
Statistical analysis
The data are expressed as the means ± standard
deviation (SD). The SPSS software (version 10.0; SPSS, Inc.,
Chicago, IL, USA) was used to perform all statistical analyses.
Comparisons were made using analysis of variance (ANOVA) and
Duncan's multiple range test. The level of significance was set at
P<0.05.
Results
Mouse body, adipose tissue and liver
weight
At the end of the experimental period, the body
weight, liver weight and epididymal and abdominal adipose tissue
weights of the mice were measured.
After 10 weeks of consuming the HFD, the body
weights of the mice in the groups fed the HFD and tuna extract
significantly decreased compared with those of the mice fed the HFD
and not fed the tuna extract. Moreover, the epididymal adipose
tissue weight of the mice fed the HFD and 100 mg/kg boiled tuna
extract group was slightly decreased, and that of the HFD-fed mice
given 200 and 400 mg/kg of tuna extract was decreased to levels
similar to those of the mice in the ND group. The abdominal adipose
tissue weight significantly decreased in the mice fed the HFD and
200 mg/kg and 400 mg/kg of boiled tuna extract, while no
significant changes were observed in that of mice fed the HFD and
100 mg/kg of tuna extract. The liver weight of all mice fed the
boiled tuna extract was significantly decreased compared with the
mice fed the HFD and not given the tuna extract (Table II).
| Table IIBody, liver, and epididymal and
abdominal adipose tissue weights in mice fed a HFD and boiled tuna
extract for 10 weeks. |
Table II
Body, liver, and epididymal and
abdominal adipose tissue weights in mice fed a HFD and boiled tuna
extract for 10 weeks.
| Group | Weight (g)
|
|---|
| Body | Liver | Epididymal fat | Abdominal
subcutaneous fat |
|---|
| Control |
31.51±2.41a |
1.09±0.11a |
0.83±0.23a |
0.43±0.15a |
| HFD |
34.51±2.43b |
1.50±1.15c |
1.45±0.28c |
0.88±0.27b |
| HFD + T100
(mg/kg) |
32.10±0.31a |
1.30±0.22b |
1.13±0.31b |
0.74±0.31b |
| HFD + T200
(mg/kg) |
31.30±2.04a |
1.20±0.12a,b |
0.86±0.24a |
0.48±0.20a |
| HFD + T400
(mg/kg) |
30.96±1.72a |
1.08±0.87a |
0.79±0.27a |
0.43±0.23a |
Effects of boiled tuna extract on serum
enzyme levels in mice fed a HFD
Leptin is secreted by adipocytes, and its functions
are to regulate appetite and energy metabolism, resulting in
satiety. It is also directly related to insulin resistance and body
fat content (19). An increase in
serum TG levels is associated with a risk of heart disease
(20). In general, high
concentrations of TG result in the accumulation of TC, and LDL-C
can build up on artery walls (21). However, increased HDL-C levels
attenuate the accumulation of LDL-C and protect against heart
disease by transporting LDL-C, TC and TG from the arteries
(22). Generally, glucose levels
are increased in obesity and result in increased TC and TG levels
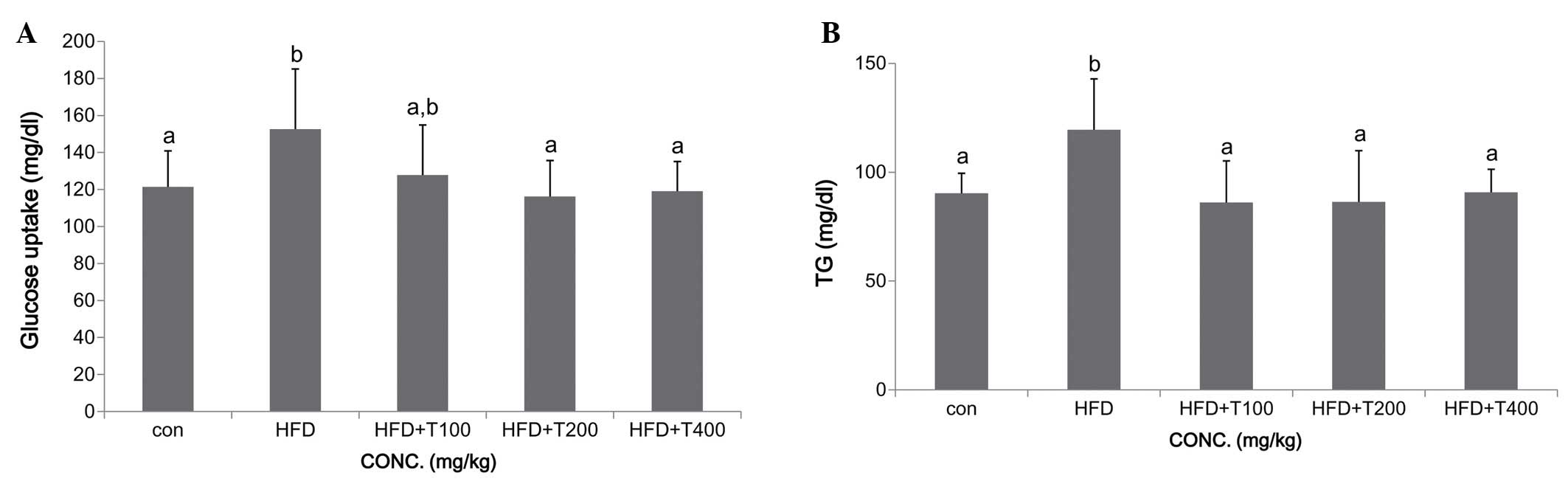
(17). In this study, we observed
increased glucose and TG serum levels in mice fed a HFD. However,
the glucose and TG levels were significantly decreased in all the
groups fed the HFD and the tuna extract [glucose: ND (control,
con), 121.4±19.4 mg/dl; HFD, 152.6±32.5 mg/dl; HFD + T100,
127.8±27.0 mg/dl; HFD + T200, 116.2±19.5 mg/dl; HFD + T400,
119.1±16.0 mg/dl; TG: ND, 90.4±9.1 mg/dl; HFD, 119.5±23.4 mg/dl;
HFD + T100, 127.86.1±19.1 mg/dl; HFD + T200, 86.3±23.5 mg/dl; HFD +
T400, 90.7±10.6 mg/dl] (Fig. 1).
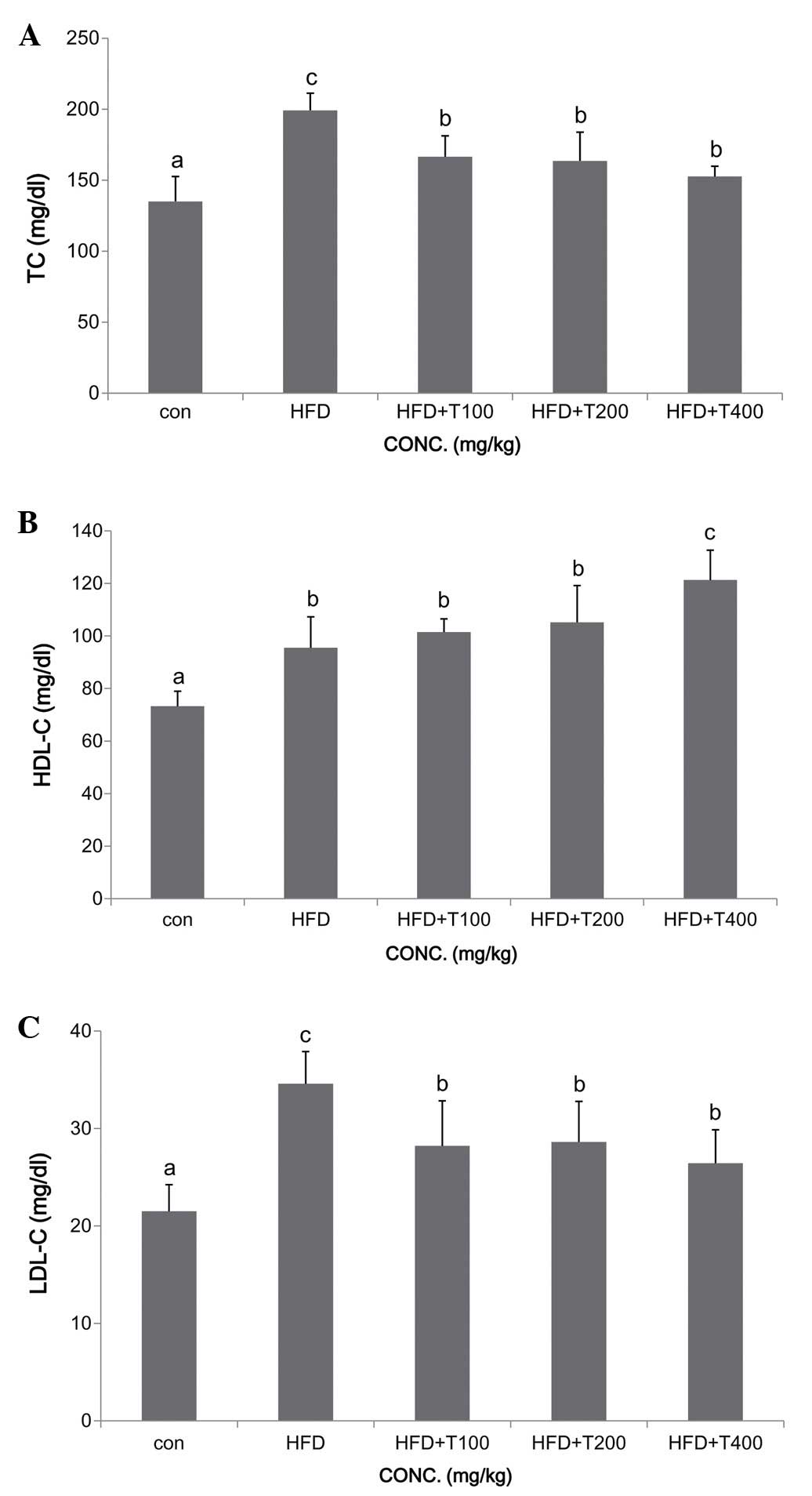
Additionally, the TC and LDL-C levels were significantly decreased
in all the groups fed the HFD and the tuna extract (TC: ND,
135.0±17.5 mg/dl; HFD, 199.2±12.1 mg/dl; HFD + T100, 166.5±14.8
mg/dl; HFD + T200, 163.6±20.2 mg/dl; HFD + T400, 152.6±7.2 mg/dl;
LDL-C: ND, 21.5±2.7 mg/dl; HFD, 34.6±3.3 mg/dl; HFD + T100,
28.2±4.6 mg/dl; HFD + T200, 28.6±4.1 mg/dl; HFD + T400, 26.4±3.4
mg/dl). However, the HDL-C levels were only increased in the group
fed the HFD and 400 mg/kg of the boiled tuna extract (HDL-C: ND,
73.2±5.7 mg/dl; HFD, 95.5±11.8 mg/dl; HFD + T100, 101.5±5.1 mg/dl;
HFD + T200, 105.2±14.0 mg/dl; HFD + T400, 121.3±11.3 mg/dl)
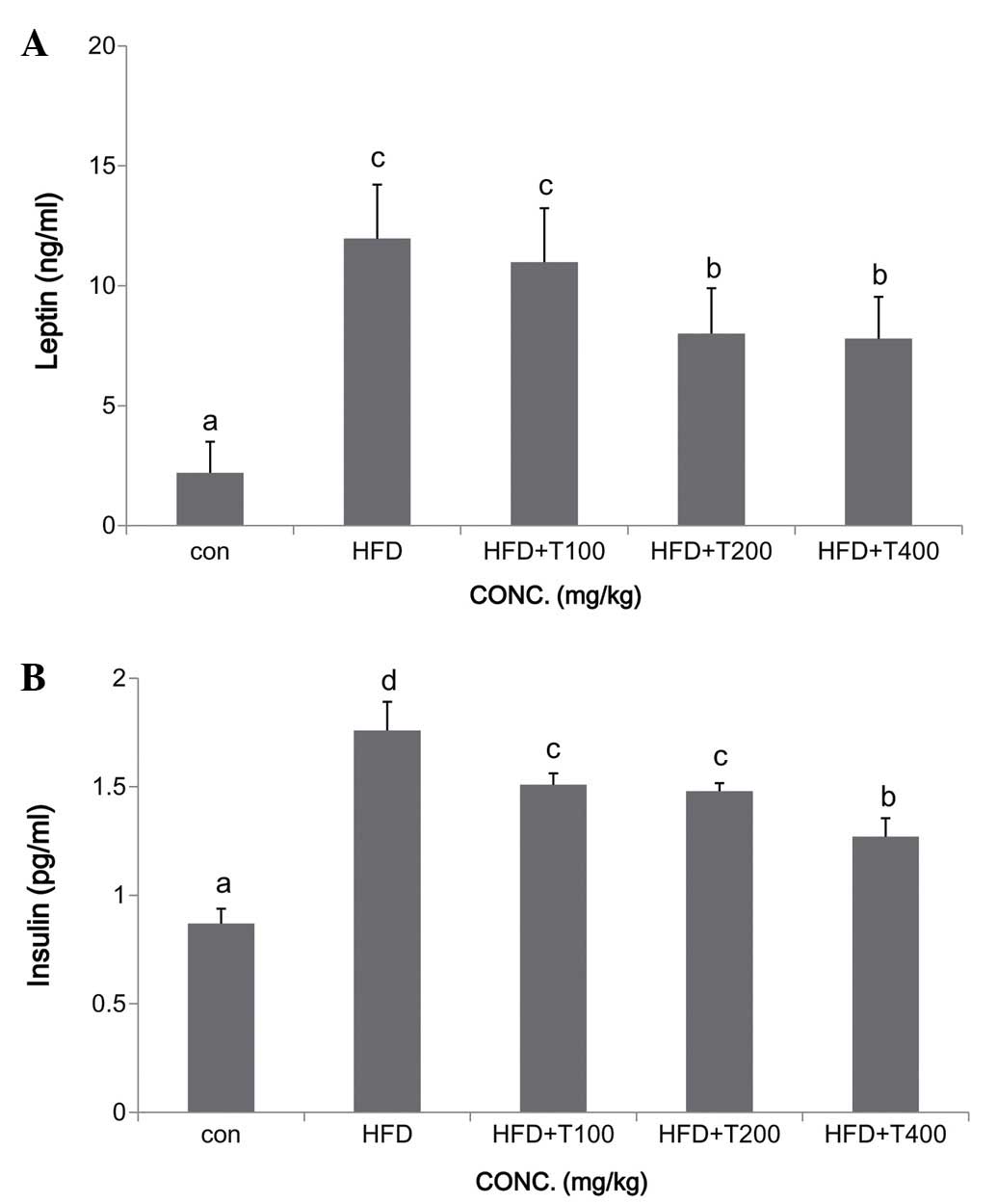
(Fig. 2A). The serum insulin
levels deceased significantly in a dose-dependent manner in the
HFD-fed mice given 100, 200, and 400 mg/kg of the boiled tuna
extract, and the leptin levels were significantly decreased in the
mice fed HFD and 200 and 400 mg/kg of the boiled tuna extract
(insulin: ND, 0.87±0.17 ng/ml; HFD, 1.76±0.26 ng/ml; HFD + T100,
1.51±0.32 ng/ml; HFD + T200, 1.48±0.32 ng/ml; HFD + T400, 1.27±0.39
ng/ml; leptin: ND, 2.2±1.3 ng/ml; HFD, 12.0±2.3 ng/ml; HFD + T100,
11.0±2.3 ng/ml; HFD + T200, 8.0±1.9 ng/ml; HFD + T400, 7.8±1.7
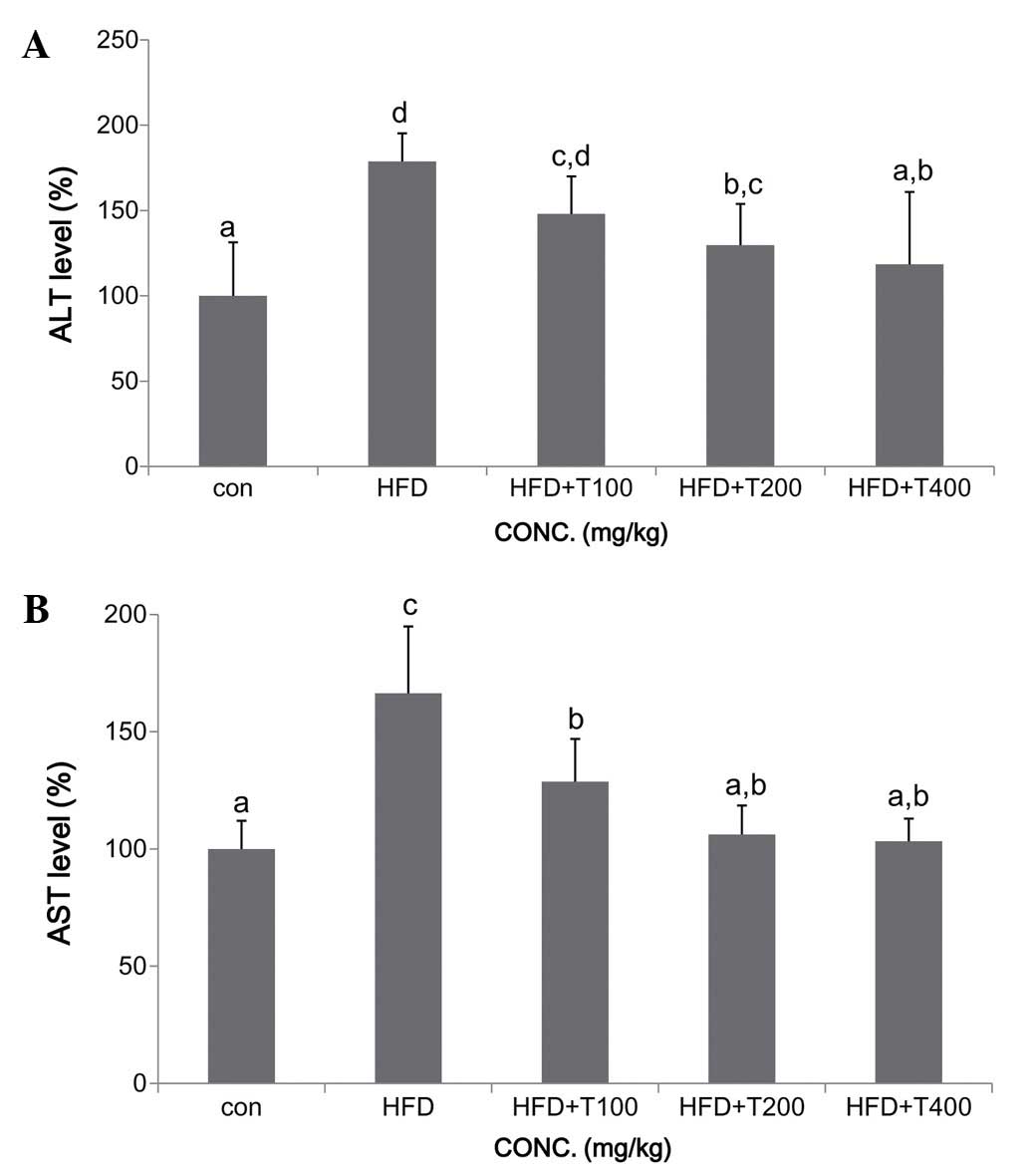
ng/ml) (Fig. 3). To determine the
damaging effects of HFD on the liver, the levels of ALT and AST
were measured. The levels of ALT and AST were significantly
elevated in the HFD-fed mice compared with the mice fed the ND.
However, in all the mice fed the boiled tuna extract, the levels of
ALT and AST decreased, and in particular, in the mice fed 400 mg/kg
of the tuna extract, the levels of ALT and AST reached levels
similar to those of the control ND-fed mice (Fig. 4).
Effects of boiled tuna extract on liver
tissue enzyme levels
After blood collection, the livers of all the mice
were removed and weighed immediately. CAT, an enzyme that plays a
role in the reduction of hydrogen peroxide with water, forms an
antioxidant enzyme system with SOD. In general, increases in
various metabolic processes induce oxidative stress and activate
SOD and CAT (23); in obesity,
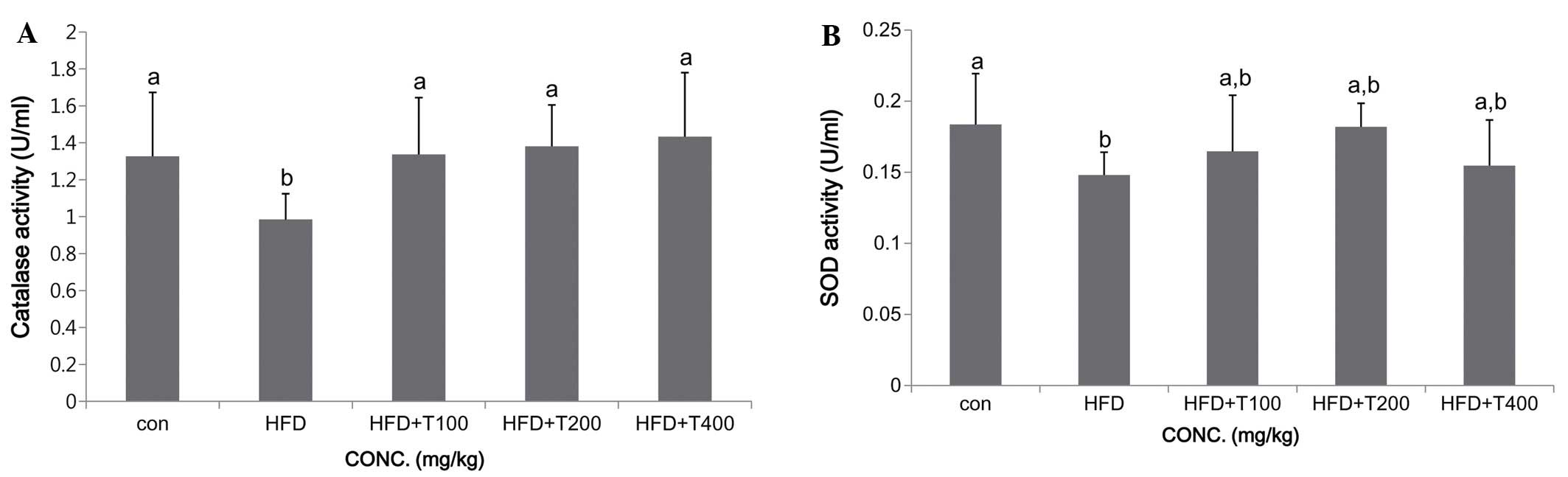
the levels of these enzymes are decreased (24). In this study, the SOD and CAT
levels in the livers of the mice in all groups are shown in
Fig. 5. Consumption of the HFD
resulted in impaired protection of the liver and decreased SOD and
CAT levels, whereas the consumption of the tuna extract increased
the SOD and CAT levels (SOD: ND, 0.18±0.04 U/ml; HFD, 0.15±0.02
U/ml; HFD + T100, 0.16±0.04 U/ml; HFD + T200, 0.18±0.02 U/ml; HFD +
T400, 0.15±0.03 U/ml; CAT: ND, 1.33±0.35 U/ml; HFD, 0.98±0.14 U/ml;
HFD + T100, 1.34±0.30 U/ml; HFD + T200, 1.38±0.22 U/ml; HFD + T400,
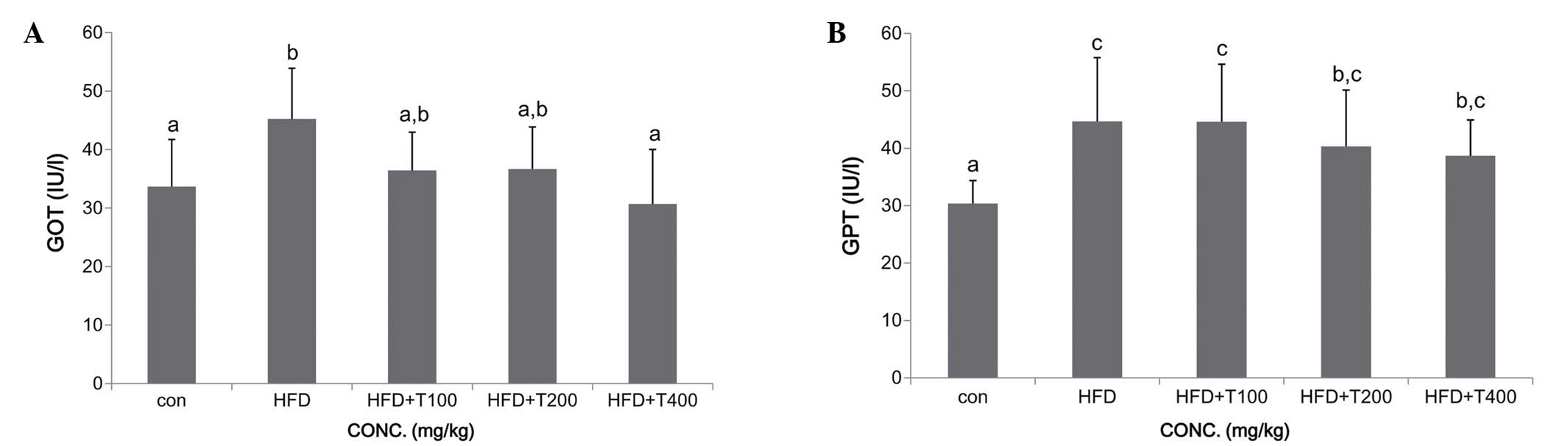
1.43±0.35 U/ml). Moreover, the consumption of a HFD can result in
hepatic steatosis (25). In our
study, the mice fed the HFD exhibited increased serum levels of GOT
and GPT compared with the ND-fed group, whereas the GOT and GPT
levels in the HFD-fed mice given the boiled tuna extract were
decreased compared with the HFD-fed group. However, the decrease in
the GTP levels was not significant in the mice fed 100 mg/kg of the
tuna extract (GOT: ND, 33.7±8.1 IU/l; HFD, 45.2±8.7 IU/l; HFD +
T100, 36.4±6.6 IU/l; HFD + T200, 36.7±7.2 IU/l; HFD + T400,
30.7±9.3 IU/l; GPT: ND, 30.4±4.0 IU/l; HFD, 44.7±11.1 IU/l; HFD +
T100, 44.6±10.0 IU/l; HFD + T200, 40.3±9.8 IU/l; HFD + T400,
38.7±6.2 IU/l) (Fig. 6).
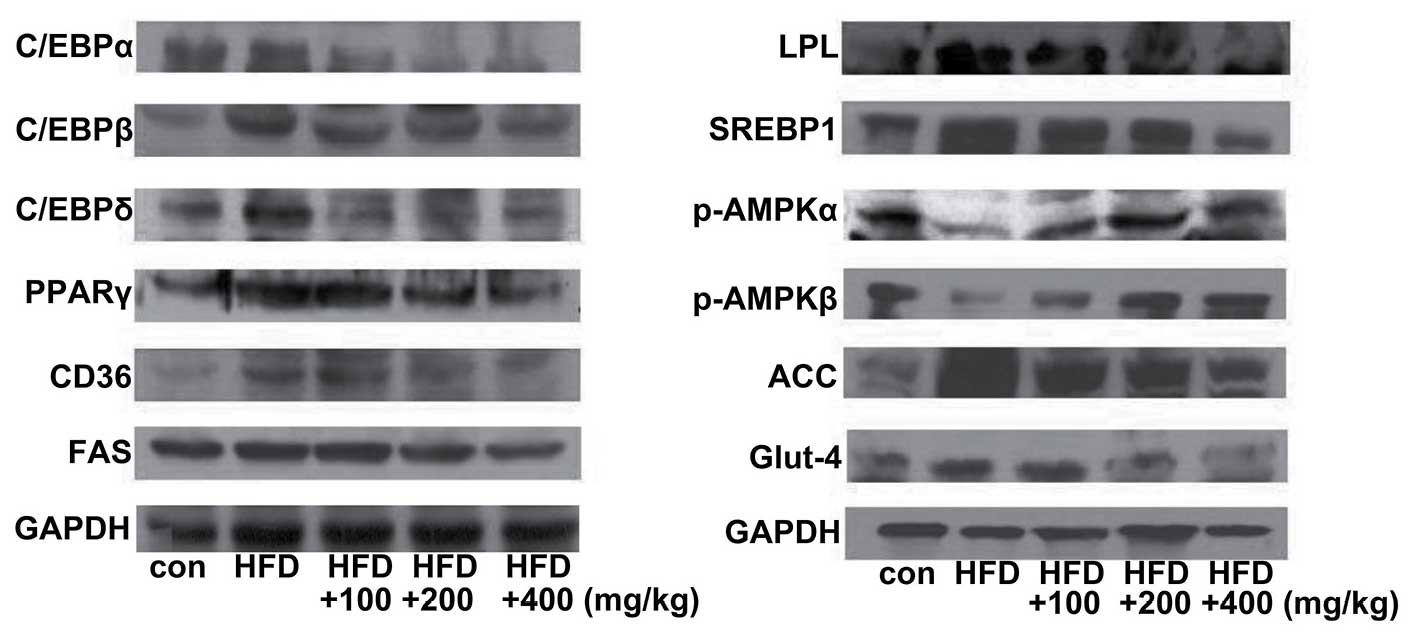
Expression levels of lipogenic- and
adipogenic-related genes in the liver tissue of mice fed a HFD
To elucidate the mechanisms underlying the effects
of boiled tuna extract on lipid metabolism, the levels of
lipogenic- and adipogenic-related genes in liver tissue were
measured. Compared with the ND-fed control group, the HFD group
exhibited increased expression levels of adipocyte markers, such as
C/EBPα, C/EBPβ, C/EBPδ, PPAR-γ, CD36, SREBP-1, LPL and FAS.
However, the expression levels of these genes were significantly
decreased in the mice fed the HFD and boiled tuna extract. The
activation of adenosine monophosphate-activated protein kinase α
and β (AMPKα, β) inhibits the expression of adipogenic-related
genes, including ACC and SREBP (26). In this study, AMPK expression was
downregulated in the HFD-fed mice, whereas it was upregulated in
the mice fed the HFD and boiled tuna extract. Glut4 is involved in
the active transport of glucose and is upregulated by C/EBPs and is
inhibited in conditions of insulin resistance via the inhibition of
C/EBPs and PPAR-γ. In this study, we confirmed the downregulation
of Glut4 by the inhibition of C/EBPs by treatment with tuna
extract. Our data clearly demonstrated the downregulation of C/EBPs
and Glut4, and the inhibition of lipogenic- and adipogenic-related
genes in the mice fed the HFD and various concentrations of the
tuna extract (Fig. 7).
Discussion
Obesity is associated with many pathologies,
including metabolic disease, hypertension, atherosclerosis and an
increased risk of death from environmental pollution, irregular
eating habits and the globalization of fast food (27). Many studies have focused on
strategies with which to exert anti-obesity effects by inhibiting
the lipogenic and adipogenic pathways. In previous studies, we
demonstrated the anti-obesity effects of peptides from boiled tuna
extract in 3T3-L1 mouse adipocytes (16,17). In this study, we examined these
effects in vivo by administering boiled tuna extract (100,
200, or 400 mg/kg) to C57BL/6N mice with obesity induced by being
fed a HFD for 10 weeks. Our data clearly demonstrated that when the
HFD-fed mice were fed various concentrations of tuna extract, the
body, liver, and epidydimal and abdominal fat weights decreased
(Table I). Moreover, tuna is a
high source of protein and contains nutritional components that
affect atherosclerosis by decreasing serum cholesterol levels. The
TC, HDL-C, and LDL-C levels in the mice fed the tuna extract are
shown in Fig. 1. Both the TC and
LDL-C levels decreased in a dose-dependent manner in the mice fed
the HFD and boiled tuna extract compared with the HFD-fed group.
However, the HDL-C levels only increased significantly in the mice
fed the a tuna extract at a dose of 400 mg/kg (Fig. 2).
Leptin is a key adipose-derived regulator of energy
expenditure and food intake, and its secretion levels are
positively associated with the extent of TG stores in adipose
tissue (28). Additionally,
changes in insulin secretion occur with obesity, and are associated
with several diseases, including diabetes. Several risk factors in
subjects with hypertension are associated with insulin resistance,
including low HDL-C and high TG levels and glucose intolerance
(29). In this study, we
confirmed significant and dose-dependent decreases in TG levels and
glucose uptake in mice fed a HFD and boiled tuna extract compared
with the mice fed the HFD with no tuna extract (Fig. 1). However, the leptin levels were
not affected by the consumption of 100 mg/kg of tuna extract
(Fig. 3A). ALT and AST play
important roles in the formation of amino acids in the liver, and
their activity increases with HFD-induced liver damage (30,31). Increased ALT and AST activity is
also associated with elevated serum cholesterol levels due to
complications with bile acid production in liver disorders
(32). Therefore, the activity of
serum ALT and AST is a useful indicator for determining liver
deterioration due to obesity. Our results demonstrated a
significant recovery of serum (ALT and AST) and liver (GOT and GPT)
enzyme function in the mice fed the tuna extract (Figs. 5 and 6).
C/EBPα and PPAR-γ induce the expression of each
other in a feedback loop, thereby maintaining cell differentiation
(33). In vivo studies
using mice have demonstrated that the disruption of PPAR-γ and
C/EBP impairs the formation of white adipose tissue and brown
adipose tissue and the differentiation of adipocytes (34). This raises the possibility that
PPAR-γ and C/EBPs may also directly promote adipogenesis- and
lipogenesis-related gene expression (16). In this study, the consumption of a
HFD upregulated C/EBP and PPAR-γ and lipogenic and adipogenic gene
expression, whereas the consumption of various concentrations of
boiled tuna extract attenuated these effects (Fig. 7). In conclusion, the consumption
of a HFD and boiled tuna extract resulted in the restoration of
body, liver, and epididymal and abdominal fat weight, as well as
serum glucose, TG, TC, HDL-C, LDL-C, insulin, leptin, AST and ALT
levels, and in the hepatic SOD and CAT levels, and the levels of
various genes involved in adipogenesis and lipogenesis, to levels
similar to those of the controls, highlighting its potential
anti-obesity effects.
Acknowledgments
This study was a part of the project titled
'Functional materials and foods using fisheries by-products',
funded by the Ministry of Oceans and Fisheries, Korea
(20130279).
References
|
1
|
Esser N, Legrand-Poels S, Piette J, Scheen
AJ and Paquot N: Inflammation as a link between obesity, metabolic
syndrome and type 2 diabetes. Diabetes Res Clin Pract. 105:141–150.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lei F, Zhang XN, Wang W, Xing DM, Xie WD,
Su H and Du LJ: Evidence of anti-obesity effects of the pomegranate
leaf extract in high-fat diet induced obese mice. Int J Obes.
31:1023–1029. 2007. View Article : Google Scholar
|
|
3
|
Ng M, Fleming T, Robinson M, Thomson B,
Graetz N, Margono C, Mullany EC, Biryukov S, Abbafati C, Abera SF,
et al: Global, regional, and national prevalence of overweight and
obesity in children and adults during 1980–2013: a systematic
analysis for the Global Burden of Disease Study 2013. Lancet.
384:766–781. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
McPherson K: Reducing the global
prevalence of overweight and obesity. Lancet. 384:728–730. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ritchie SA and Connell JM: The link
between abdominal obesity, metabolic syndrome and cardiovascular
disease. Nutr Metab Cardiovasc Dis. 17:319–326. 2007. View Article : Google Scholar
|
|
6
|
Paccaud F, Schlüter-Fasmeyer V,
Wietlisbach V and Bovet P: Dyslipidemia and abdominal obesity: an
assessment in three general populations. J Clin Epidemiol.
53:393–400. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Eisinger K, Liebisch G, Schmitz G,
Aslanidis C, Krautbauer S and Buechler C: Lipidomic analysis of
serum from high fat diet induced obese mice. Int J Mol Sci.
15:2991–3002. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang J and Ryu HK: The effects of
Momordica charantia on obesity and lipid profiles of mice fed a
high-fat diet. Nutr Res Pract. 9:489–495. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vargas-Robles H, Rios A, Arellano-Mendoza
M, Escalante BA and Schnoor M: Antioxidative diet supplementation
reverses high-fat diet-induced increases of cardiovascular risk
factors in mice. Oxid Med Cell Longev. 2015:4674712015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Koch CE, Lowe C, Pretz D, Steger J,
Williams LM and Tups A: High-fat diet induces leptin resistance in
leptin-deficient mice. J Neuroendocrinol. 26:58–67. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang Y, Li W, Liu Y, Sun Y, Li Y, Yao Q,
Li J, Zhang Q, Gao Y, Gao L and Zhao J: Alpha-lipoic acid improves
high-fat diet-induced hepatic steatosis by modulating the
transcription factors SREBP-1, FoxO1 and Nrf2 via the
SIRT1/LKB1/AMPK pathway. J Nutr Biochem. 25:1207–1217. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gao M, Ma Y and Liu D: High-fat
diet-induced adiposity, adipose inflammation, hepatic steatosis and
hyperinsulinemia in outbred CD-1 mice. PLoS One. 10:e01197842015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rindler PM, Plafker SM, Szweda LI and
Kinter M: High dietary fat selectively increases catalase
expression within cardiac mitochondria. J Biol Chem. 288:1979–1990.
2013. View Article : Google Scholar :
|
|
14
|
Suarez-Jimenez GM, Burgos-Hernandez A and
Ezquerra-Brauer JM: Bioactive peptides and depsipeptides with
anticancer potential: sources from marine animals. Mar Drugs.
10:963–986. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hsu KC, Li-Chan ECY and Jao CL:
Antiproliverative activity of peptides prepared from enzymatic
hydrolisate of tuna dark muscle on human breast cancer cell line
MCF-7. J Food Chem. 126:617–622. 2011. View Article : Google Scholar
|
|
16
|
Kim YM, Kim EY, Kim IH and Nam TJ: Peptide
derived from desalinated boiled tuna extract inhibits adipogenesis
through the downregulation of C/EBP-α and PPAR-γ in 3T3-L1
adipocytes. Int J Mol Med. 35:1362–1368. 2015.PubMed/NCBI
|
|
17
|
Kim YM, Kim IH, Choi JW, Lee MK and Nam
TJ: The ant-obesity effects of a tuna peptide on 3T3-L1 adipocytes
are mediated by the inhibition of the expression of lipogenic and
adipogenic genes and by the activation of the Wnt/β-catenin
signaling pathway. Int J Mol Med. 36:327–334. 2015.PubMed/NCBI
|
|
18
|
Fasshauer M and Paschke R: Regulation of
adipocytokines and insulin resistance. Diabetologia. 46:1594–1603.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kopelman PG: Obesity as a medical problem.
Nature. 404:635–643. 2000.PubMed/NCBI
|
|
20
|
Chi MS, Koh ET and Stewart TJ: Effects of
garlic on lipid metabolism in rats fed cholesterol or lard. J Nutr.
112:241–248. 1982.PubMed/NCBI
|
|
21
|
Kwon OJ, Kim MY and Roh SS: Improving
effect of extract of Ganoderma lucidum in atherosclerosis from LDL
receptor knockout mouse. Korea J Herbol. 31:17–23. 2016. View Article : Google Scholar
|
|
22
|
Sack FM, Tonkin AM, Craven T, Pfeffer MA,
Shepherd J, Keech A, Furberg CD and Braunwald E: Coronary heart
disease in patients with low LDL-cholesterol: benefit of
pravastatin in diabetics and enhanced role for HDL-cholesterol and
triglycerides as risk factors. Circulation. 105:1424–1428. 2002.
View Article : Google Scholar
|
|
23
|
Scandalios JG: Oxygen stress and
superoxide dismutases. Plant Physiol. 101:7–12. 1993.PubMed/NCBI
|
|
24
|
Kim JY, Shin M and Heo YR: Effect of
stabilized rice bran on obesity and antioxidative enzyme activity
in high fat diet-induced obese C57BL/6 mice. J Korean Soc Food Sci
Nutr. 43:1148–1157. 2014. View Article : Google Scholar
|
|
25
|
Park PJ, Kim CW, Cho SY, Rha CS, Seo DB
and Lee SJ: Aqueous spray-dried green tea extract regulates body
weight and epididymal fat accumulation in mice. Korean J Food Sci
Technol. 42:103–108. 2010.
|
|
26
|
Jeong HJ, Park JH and Kim MJ: Ethanol
extract of Hippophae rhamnides L. leaves inhibits adipogenesis
through AMP-activated protein kinase (AMPK) activation in 3T3-L1
preadipocytes. Hangug Jaweon Sigmul Haghoeji. 28:582–590. 2015.
|
|
27
|
Friedman JM and Halaas JL: Leptin and the
regulation of body weight in mammals. Nature. 395:763–770. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Reaven GM: Banting lecture 1988. Role of
insulin resistance in human disease. Diabetes. 37:1595–1607. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Modan M, Halkin H, Almog S, Lusky A,
Eshkol A, Shefi M, Shitrit A and Fuchs Z: Hyperinsulinemia. A link
between hypertension obesity and glucose intolerance. J Clin
Invest. 75:809–817. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kang YH, Park YK, Ha TY and Moon KD:
Effects of pine needle extracts on serum and liver lipid contents
in rats fed high fat diet. J Korean Soc Food Nutr. 25:367–373.
1996.
|
|
31
|
Lim AK, Jung MJ, Kim DW, Hong JH, Jung HK,
Kim KS, Kim YH and Kim DI: An extrapolation concentration decision
effect antihyperlipidemic of aglycone isoflavone from
biotransformation soybean on the fed high-fat diet rats. J Korean
Soc Food Sci Nutr. 38:1167–1173. 2009. View Article : Google Scholar
|
|
32
|
Rosen ED, Hsu CH, Wang X, Sakai S, Freeman
MW, Gonzalez FJ and Spiegelman BM: C/EBPalpha induces adipogenesis
through PPARgamma: a unified pathway. Genes Dev. 16:22–26. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gray SL, Dalla Nora E and Vidal-Puig AJ:
Mouse models of PPAR-gamma deficiency: dissecting PPAR-gamma's role
in metabolic homoeostasis. Biochem Soc Trans. 33:1053–1058. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang ND, Finegold MJ, Bradley A, Ou CN,
Abdelsayed SV, Wilde MD, Taylor LR, Wilson DR and Darlington GJ:
Impaired energy homeostasis in C/EBP alpha knockout mice. Science.
269:1108–1112. 1995. View Article : Google Scholar : PubMed/NCBI
|