Introduction
Breast cancer is the most prevalent type of cancer
with the highest incidence of all cancer types among women in
China, accounting for 23% of diagnosed cancer cases and for 14% of
cancer mortalities each year (1–3).
Despite advances being made in the screening and adjuvant treatment
of breast cancer, the mortality rates associated breast cancer
remain high and metastasis is the leading cause of mortality
(4). Human epidermal growth
factor receptor-2 (HER-2) is a member of the epidermal growth
factor receptor membrane tyrosine kinase, which plays an important
role in the proliferation, invasion and migration of cells
(5). An increased expression of
HER-2 is found in 20–30% of patients with breast cancer and is
associated with a poor prognosis and metastasis (6).
Tumor metastasis, an aggressive process in the
development of tumor cells, is a multiple complex process involving
invasion and migration. Epithelial-mesenchymal transition (EMT), an
important process originally described in embryonic development,
has been reported to play a vital role in the metastasis of some
carcinomas (7). During the
process of EMT, focal adhesion kinase (FAK) promotes the
phosphorylation of β-catenin to increase the expression of
transcription factors associated with EMT.
FAK is a non-receptor tyrosine kinase that has been
shown to be upregulated in various human cancer cells, including
lung, colon, thyroid, hepatocellular, prostate, pancreatic and
breast cancers (8 and refs therein). It has also been reported that
FAK plays a vital role in various biological processes, including
adhesion, angiogenesis, motility, EMT and metastasis (9). It has also been reported that HER-2
influences the metastasis of breast cancer cells through the
cSrc/FAK pathway (10). The
activation of cSrc/FAK is critical for the phosphorylation of
p130Cas. The interaction of p130Cas with FAK
promotes cell migration by activating c-Jun N-terminal kinases
(JNKs) (11).
Recently, researches have studied phytochemicals
extracted from fruits, vegetables and other herbs. It has been
defined that these nutrients, including anthocyanins, play an
important role on oncotherapy (12). A previous study suggested that a
phytochemical named oleuropein reduced tumor growth in xenografted
mice and enhanced the efficacy of trastuzumab against breast cancer
cells (13). Citrus fruits and
their bioactive ingredients have also been shown to suppress cancer
growth by inducing apoptsis (14). The overexpression of oncogenes,
the inactivation of tumor-related suppressor genes and the loss of
apoptosis are some of the mechanisms that have been widely
researched in molecular oncology. Phytochemicals, such as fungal
substances has been shown to control the expression of oncogenes
and tumor suppressor in vivo and in vitro (15 and
refs therein). Rutin modulates intracellular signaling pathways to
control cancer cell growth and apoptosis in vivo and in
vitro (16 and refs therein). Viscum album has also
emerged as a natural source with substantial biological activities,
such as modulating endoplasmic reticulum stress and targeting
cytoskeletal machinery in cancer cells (17).
Black rice anthocyanins (BRACs) are a category of
anthocyanins extracted from the aleurone layer of black rice, which
is regarded as a healthy food due to its beneficial effects on the
liver and gastrointestinal tract (18). It has been demonstrated that
anthocyanins exert an inhibitory effect on carcinogenesis, and
inhibit cancer progression and metastasis through cell signal
transduction (19). Hui et
al (20) reported that BRACs
induced the apoptosis and inhibited the angiogenesis of MDA-MB-453
cells through an intrinsic pathway. In their study, BALB/c nude
mice bearing MDA-MB-453 tumor xenografts orally ingested BRACs (100
mg/kg/day), which suppressed tumor growth and angiogenesis by
decreasing the expression of angiogenic factors. Previous studies
have also indicated that BRACs inhibited lung nodule formation and
pulmonary metastasis in ErbB2-positive MDA-MB-453 subcutaneous
xenografts, and that the RAS/RAF/MAPK pathway played a vital role
in this inhibitory effect (21,22). However, limited information is
available on the inhibitory effects of BRACs on EMT and FAK
signaling associated with this anti-metastatic effect.
The present study thus investigated whether FAK
signaling plays a role in the anti-metastatic effects of BRACs on
HER-2-positive breast cancer in vitro. It was found that
BRACs exert a suppressive effect on the process of EMT.
Materials and methods
Reagents
The BRACs used in this study were purchased from the
New Star Natural Plant Development Company (Jilin, China). The FAK
inhibitor, 1,2,4,5-benzenetetramine tetrahydrochloride (Y15), was
obtained from Sigma (St. Louis, MO, USA). Fetal bovine serum (FBS)
and horse serum were purchased from Gibco-BRL (Grand Island, NY,
USA). Dulbecco's modified Eagle's medium (DMEM)/high glucose and
DMEM/F12 were purchased from HyClone (Beijing, China). L15 medium
was obtained from Keygen Biotech (Nanjing, China). Primary
antibodies against FAK (ab40794), phospho-FAK (ab38458, ab81298),
Src (ab16885), phospho-Src (ab24789), and p130Cas
(ab136514) were purchased from Abcam (Cambridge, MA, USA).
Phospho-p130Cas (#4011) antibody was purchased from Cell
Signaling Technology (Danvers, MA, USA). Primary antibodies against
E-cadherin (610181), vimentin (550513), and fibronectin (610077)
were purchased from BD Biosciences (Bedford, MA, USA).
Cell culture
The human breast cancer cell lines, MCF-7 (HER-2
negative), MDA-MB-453 (HER-2 positive), and the normal breast cell
line, MCF-10A, were purchased from the Chinese Academy of Science.
The MCF-7 cells were cultured in DMEM/high glucose with 10% FBS,
and the MCF-10A cells were cultured in DMEM/F12 supplemented with
6% horse serum, 10 µg/ml insulin (Wako, Osaka, Japan), 20
ng/ml epidermal growth factor (PeproTech, Rocky Hill, NJ, USA), 100
ng/ml cholera toxin and 0.5 µg/ml hydrocortisone in
humidified 5% CO2 at 37°C. The MDA-MB-453 cells were
cultured in L15 medium with 10% FBS in 100% O2 at
37°C.
Cell counting kit-8 (CCK-8) assay
The MCF-10A, MCF-7 and MDA-MB-453 cells
(1×103 cells/well in a 96-well plate) were incubated
overnight at 4°C. Following incubation, the cells were treated with
or without BRACs at various concentrations (0, 50, 100, 200 and 400
µg/ml) for 24 h. CCK-8 assays (C0037; Beyotime, Shanghai,
China) were performed according to the instructions provided with
the kits, and the absorbance was detected at 450 nm using a
microplate reader (Biotek, Shanghai, China).
Cell adhesion assay
The cell adhesion assay was performed as described
in a previous study (23).
Briefly, the cells were pre-treated with various concentrations of
BRACs (0, 50, 100, 200 and 400 µg/ml) or Y15 (0, 2, 4, 8 and
16 µM) for 24 h. A 96-well plate was pre-coated with
fibronectin (10 µg/ml) for 1 h at 37°C. The plate was then
incubated with 3% bovine serum albumin (BSA) in phosphate-buffered
saline (PBS) for 30 min to block non-specific binding sites; the
cells (5×104/well) were then seeded on a fibronectin
pre-coated plate at 37°C with 5% CO2, and the plate was
then washed with PBS 3 times to remove the unattached cells after 1
h. The attached cells were fixed with 3.7% paraformaldehyde for 10
min and stained with 0.1% crystal violet (C0121, Beyotime) in 2%
ethanol for 10 min followed by washing 3 times with PBS. The plate
with attached the cells was rinsed with water and dried. The cells
with crystal violet were eluted in 10% acetic acid, and the
absorbance was measured at 595 nm using a microplate reader (Thermo
Fisher Scientific, Inc., Waltham, MA, USA).
Wound healing assay
The MCF-10A, MCF-7, and MDA-MB-453 cells were grown
in 24-well plates to form a confluent monolayer. A cross-scratch
wound was created using 200-µl sterile pipette tips. The
cells were then treated with BRACs (200 µg/ml) or Y15 (10
µM) for 24 h. The images were observed under an inverted
light microscope (Olympus, Tokyo, Japan). Migration distances were
measured using ImageJ software.
Cell invasion assay
The in vitro invasion features of the cells
were measured by a cell invasion assay (ECM550; Millipore,
Billerica, MA, USA), according to the manufacturer's instructions.
The cells (3×105/chamber) were seeded in the ECM-coated
upper compartment, and media including BRACs (200 µg/ml) or
Y15 (10 µM) were then added to each upper insert. The lower
compartment was filled with a medium containing serum. Following 48
h of incubation at 37°C, non-invading cells were removed using
cotton swabs, and the Transwell membrane was then fixed and stained
with crystal violet. The number of invading cells was counted in
random fields under a light microscope (DM4000B; Leica, Wetzlar,
Germany) followed by several times rinses in water.
Western blot analysis
The cells pre-treated with BRACs (200 µg/ml)
or Y15 (10 µM) were lysed in a radioimmunoprecipitation
assay (RIPA) buffer [50 mM Tris-HCL (pH 7.4), 150 mM NaCl, 1%
Triton X-100, 1% sodium deoxycholate, 0.1% sodium dodecyl sulfate
(SDS) and 1 mM phenylmethanesulfonyl fluoride] supplemented with
protease inhibitor cocktail tablets for 30 min at 4°C. Lysates were
centrifuged at 13,000 rpm for 30 min at 4°C, and the undissolved
debris was discarded. The protein concentration was detected using
biocinchoninic acid assay kits (P0010; Beyotime). Equal amounts of
protein (40 µg) were separated on sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred onto
polyvinylidene fluoride membranes (Millipore). The membranes were
blocked with TBST [20 mM Tris-HCL (pH 7.4), 150 mM NaCl, 0.1%
Tween-20] containing 5% non-fat milk or 3% BSA. After blocking, the
membranes were incubated with primary antibodies (1:1,000)
overnight at 4°C. The membranes were washed 3 times with TBST
followed by incubation with horseradish peroxidase-conjugated
secondary antibodies (1:10,000; Beyotime) for 1 h at room
temperature. The proteins were detected using an enhanced
chemiluminescence detection kit (Thermo Fisher Scientific,
Inc.).
Immunoprecipitation analysis
The cells were rinsed twice with cold PBS, followed
by lysing with RIPA buffer containing phosphatase inhibitors.
Anti-ErbB2, FAK, cSrc, or p130Cas antibodies were added
to protein A/G agarose beads (Millipore) and incubated for 1 h at
37°C with gentle mixing. The mixture was centrifuged at 4°C, the
supernatant was discarded, and the beads were washed 3 times with
PBST. Equal amounts of proteins were added to the beads for
incubation overnight at 4°C with gentle shaking. The beads were
suspended by boiling with sample buffer after washing with PBST and
analyzed by western blot analysis.
Statistical analysis
Data are presented as the means ± standard deviation
(SD). One-way analysis of variance and Dunnett's t-test were used
to determine the statistical significance between values of the
various experimental and control groups. A value of P<0.05 was
considered to indicate a statistically significant difference.
Results
BRACs inhibit the metastasis of
Her-2-positive breast cancer cells
The effects of BRACs on the proliferation of
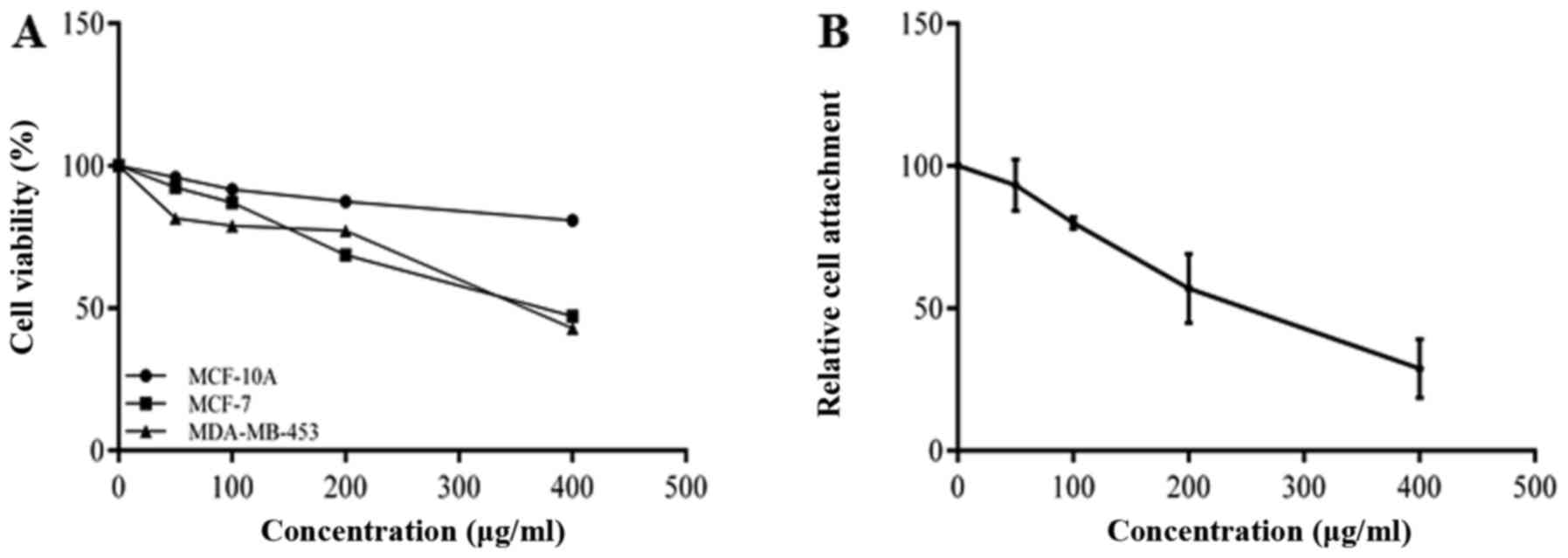
MDA-MB-453 cells were examined by CCK-8 assay. The results shown in
Fig. 1A indicate that BRACs
inhibited the viability of the MDA-MB-453 cells in a dose-dependent
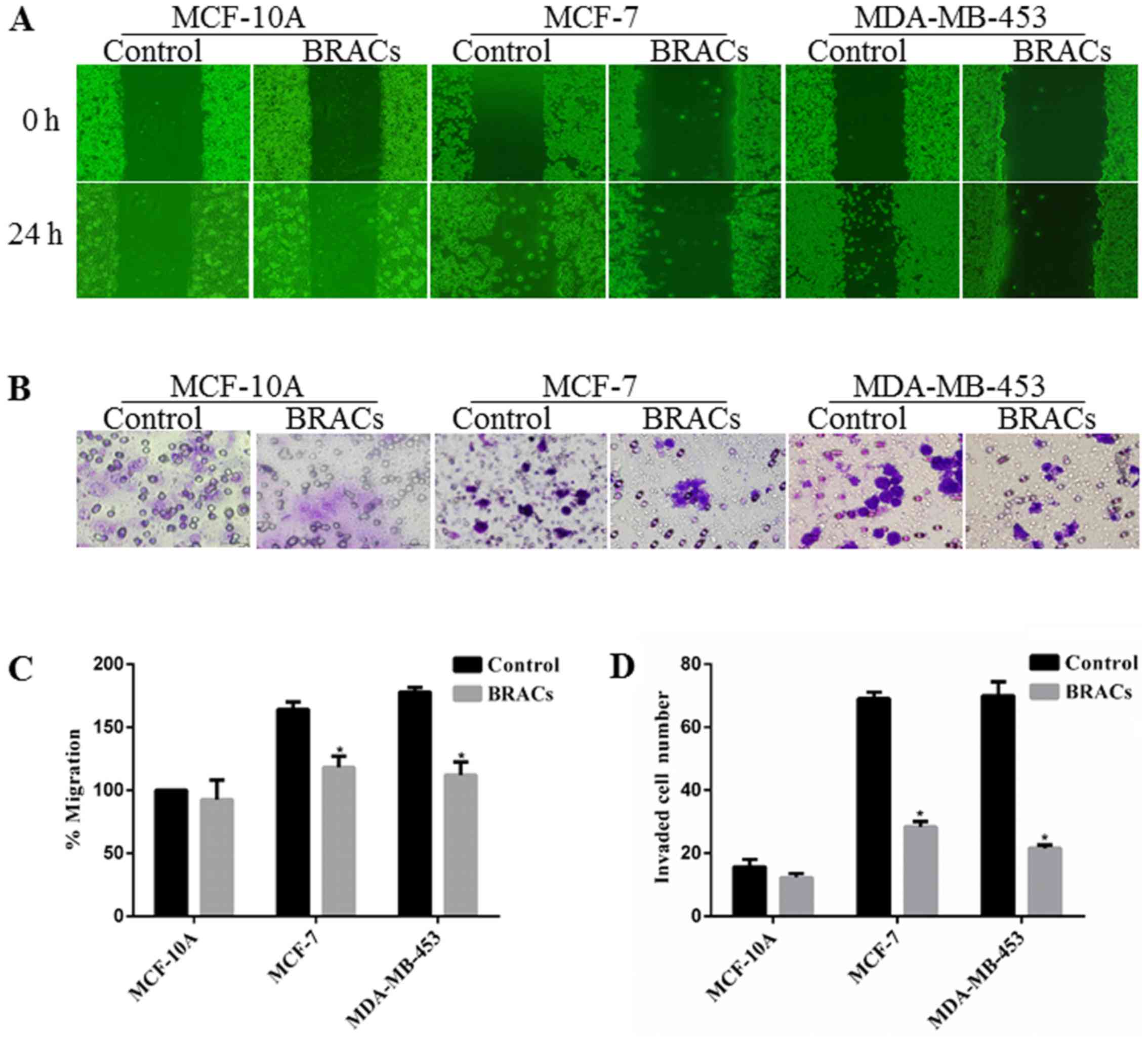
manner. A wound healing assay was also used to examine the effect
of BRACs on the migration of the breast cancer cells. BRACs
significantly decreased the migration distance of MDA-MB-453 cells
by 37% compared with the untreated cells after 24 h (Fig. 2A and C). The results also revealed
that BRACs did not affect the migration ability of the normal
mammary epithelial MCF-10A cells. In addition, a Matrigel-coated
membrane was used to examine the invasive ability of the MDA-MB-453
cells. The results indicated that BRACs decreased the invasive
ability of the MDA-MB-453 cells, the number of invaded cells has
been reduced by 68% (Fig. 2B and
D). The adhesion of the cancer cells to extracellular matrix
(ECM) and cell-ECM interaction are important processes of
metastasis; therefore, this study examined the adhesion of the
MDA-MB-453 cells to fibronectin, which is an essential protein in
the ECM (24). BRACs inhibited
the adhesion of the MDA-MB-453 cells to fibronectin in a
concentration-dependent manner (Fig.
1B). These results suggest that BRACs exerts anti-metastatic
effects on HER-2-positive breast cancer cells.
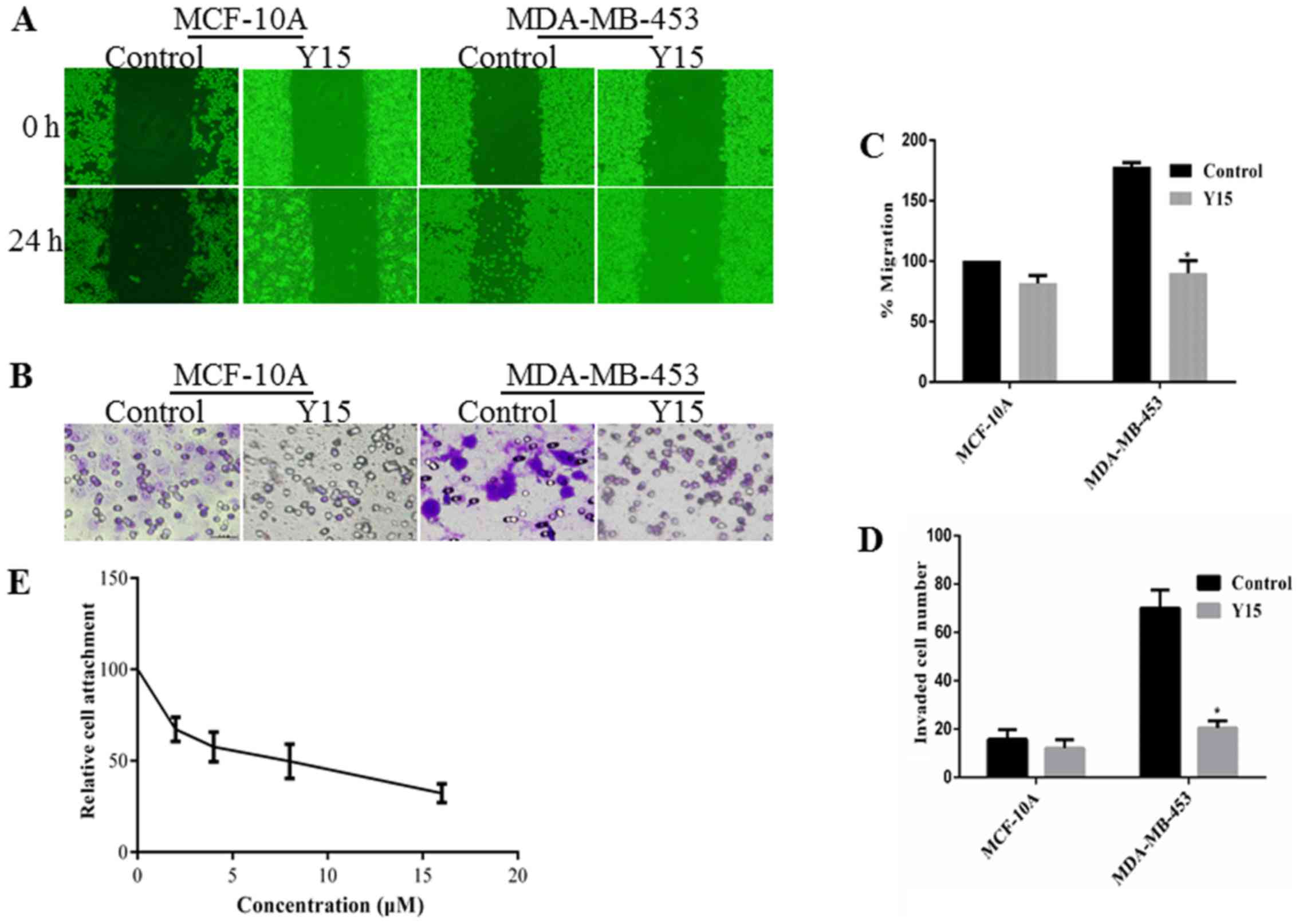
Y15, a small molecule inhibitor that inhibits the
growth of breast cancers, can inhibit the phosphorylation of FAK by
binding with the Y397 site. The Y397 site is an
autophosphory-lation site of FAK that provides a binding for cSrc
to lead to downstream signaling by mediating the phosphorylation of
FAK. Y15 significantly decreased the migration distance and the
invasive ability of the MDA-MB-453 cells (Fig. 3). The adhesion between fibronectin
and MDA-MB-453 cells was also reduced by Y15. Thus, Y15 exerted
anti-metastatic effects similar to those of BRACs.
BRACs reduces EMT in Her-2-positive
breast cancer cells
EMT plays an important role in the metastasis of
breast carcinomas (7). The
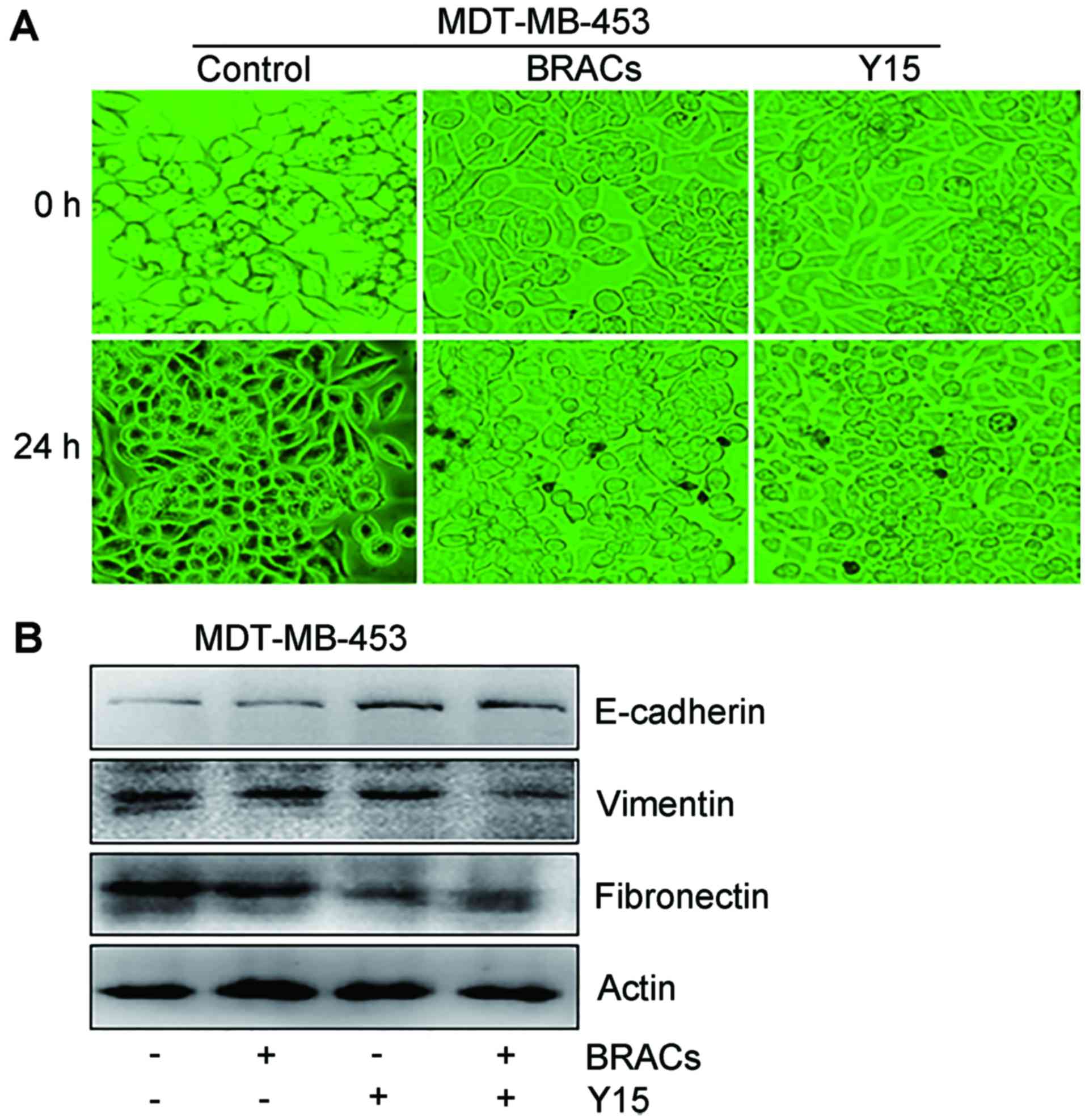
present study examined whether BRACs influence EMT in MDA-MB-453
cells. The morphology of the MDA-MB-453 cells significantly changed
from a fibroblast-like to a cobblestone-like appearance following
treatment with BRACs or Y15, suggesting that BRACs reduced EMT in
the MDA-MB-453 cells (Fig. 4A).
The results of western blot analysis also revealed that BRACs
increased the expression of the epithelial marker, E-cadherin, and
decreased the expression of the mesenchymal markers, fibronectin
and vimentin, in the MDA-MB-453 cells (Fig. 4B). BRACs exerted an anti-EMT
effect that was similar to the effect of Y15 (Fig. 4B). BRACs together with Y15 further
augmented the expression of E-cadherin and exerted further
suppressive eftects on the expression of fibronectin and
vimentin.
BRACs reduce EMT by inhibiting
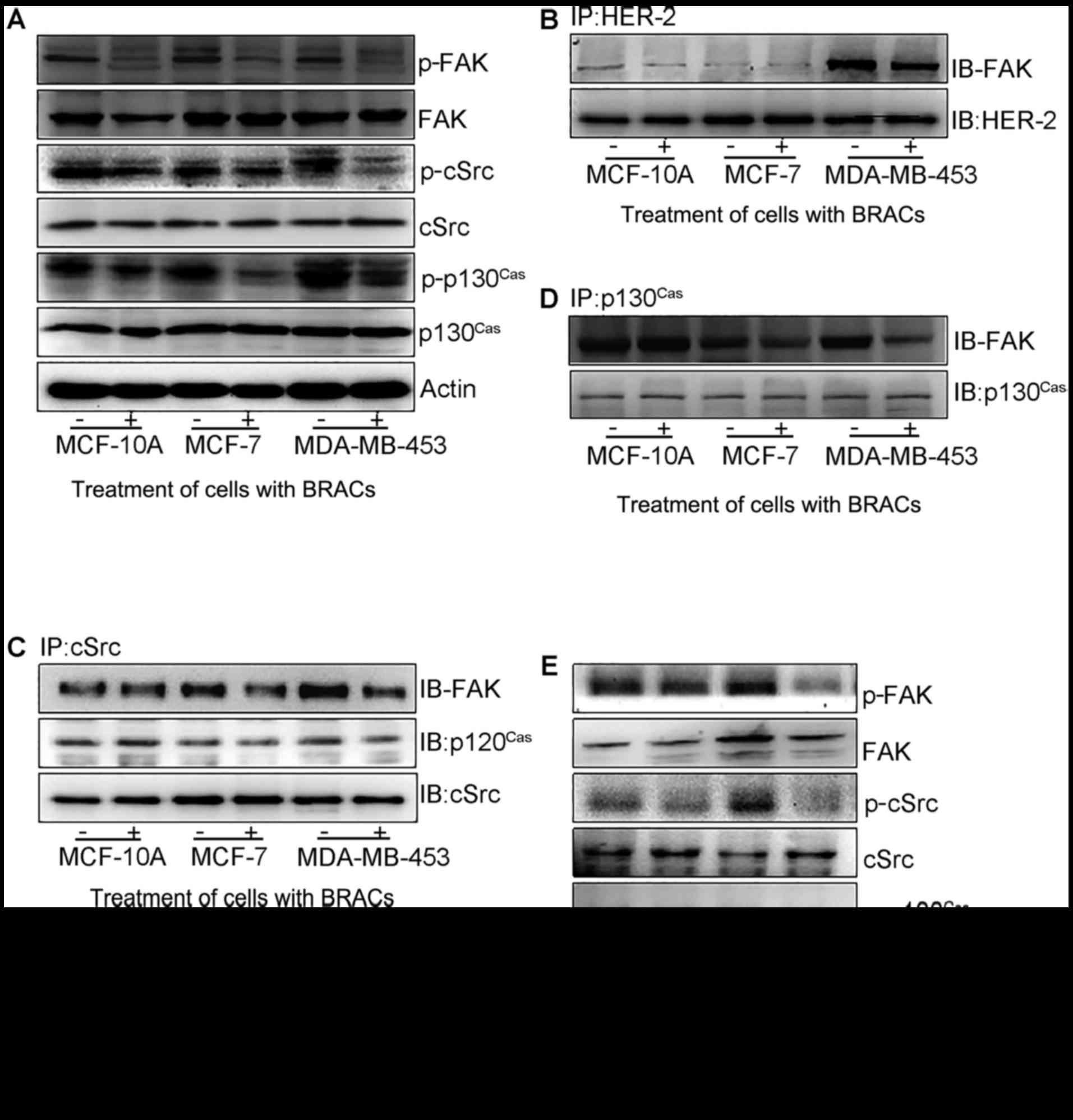
cSrc/FAK/p130Cas signaling
cSrc/FAK/p130Cas signaling plays a key
role in favoring migration, invasion and metastasis in cancer cells
regulated by EMT (10). To
elucidate the molecular mechanisms underlying the anti-metastatic
effects of BRACs, the effects of BRACs treatment on the expression
and phosphorylation of cSrc/FAK/p130Cas were evaluated
by western blot analysis (Fig.
5A). BRACs significantly decreased the phosphorylation of cSrc,
FAK and p130Cas in the MDA-MB-453 cells. Additional
experiments were performed to determine the effects of BRACs on the
association between HER-2 and FAK, FAK and cSrc, cSrc and
p130Cas, and between FAK and p130Cas by
western blot analysis. BRACs treatment inhibited the interactions
among these proteins (Fig. 5B–D).
BRACs weakened the association between HER-2 and FAK, attenuated
the activation of FAK by HER-2, and inhibited the contact among
FAK, cSrc and p130Cas proteins. It also reduced the
interaction among these proteins further by suppressing the
transduction of FAK signaling. The results of western blot analysis
revealed that Y15 also suppressed the phosphorylation of cSrc, FAK
and p130Cas in the MDA-MB-453 cells (Fig. 5E).
Discussion
Anthocyanins, a species of natural flavonoids
existing widely in food such as cabbages, berries, grapes, cereal
and other dark-colored fruits and vegetables, appear to have
potential benefits such as antioxidant, anti-inflammatory,
anticancerous and anti-metastatic activities (25). Anthocyanins exert their effects in
a native or metabolized form by entering the circulation in a
sufficient quantity. However, these beneficial effects are
dependent on the bioavailability and absorbed substances of
anthocyanins (26). As hydrolyzed
by the colon microflora or intestinal enzymes, anthocyanins are
usually absorbed slowly in the intestine due to the eliminated
hydrophilic moiety which is usually necessary for anthocyanins to
cross the intestine membrane via passive diffusion (27). Anthocyanins are then further
metabolized in the liver and small intestine followed by entering
the circulation and targeting different tissues in a conjugated
structure by sulfation, glucuronidation or methylation. Ultimately,
anthocyanins are secreted in bile or eliminated in urine (28). In biotransprotation, anthocyanins
can be transported through one human intestinal epithelial cell
model in intact glycone forms (29), then stored in tissues rather than
circulated in the blood circulation. Anthocyanins can be restored
in the liver, cortex, eye and cerebellum after consuming blueberry
anthocyanins, as shown in an in vivo study (30 and refs
therein).
During the complex processes of metastasis, cancer
cell invasion of the basement membrane is one of the earliest and
critical steps (31). The most
important step is the invasion of the ECM, known as the degradation
of basement membrane collagen, by some proteolytic enzymes such as
matrix metal-loproteinases (MMP) and urokinase-type plasminogen
activator (u-PA). BRACs have been found to inhibit cancer cell
invasion by decreasing the expression of MMP and u-PA (32). The expression of MMP-2 and MMP-9
is regulated by FAK in the cell migration and invasion of
hepatocellular carcinoma (33).
The present study indicated that BRACs reduced the
adhesion, migration and invasion of human HER-2-positive breast
cancer MDA-MB-453 cells. It also significantly altered the
morphology of the MDA-MB-453 cells from a mesenchymal to an
epithelial phenotype. The results of western blot analysis revealed
that BRACs increased the expression of the epithelial marker,
E-cadherin, and decreased the expression of the mesenchymal
markers, fibronectin and vimentin. These results indicate that the
inhibitory effect of BRACs on EMT plays an important role in their
anti-metastatic effects.
To delineate whether FAK signaling plays a
significant role in the metastasis of MDA-MB-453 cells, the present
study used adhesion, wound healing and Transwell assays to analyze
the adhesion, migration, invasion and EMT status of cells
pretreated with Y15. Y15 inhibited the phosphorylation of FAK by
binding with the Y397 site. The Y397 site is an autophosphorylation
site of FAK that provides a binding for cSrc to lead to downstream
signaling by mediating the phosphorylation of FAK. A previous study
demonstrated that Y15 inhibits the growth of breast cancers
(34). The present study
demosntrated that the adhesion, migration, invasion of the
MDA-MB-453 cells and EMT were reduced by Y15. The aforementioned
results indicated that BRACs exerted similar anti-metastatic
effects to those of Y15, and taht FAK played a vital role in the
anti-metastastic effects of BRACs.
In the present study, the expression and
phosphorylation of FAK, cSrc and p130Cas were
investigated. The total level of FAK, cSrc and p130Cas
was not affected, but the phosphorylation levels were reduced by
BRACs and Y15. BRACs decreased the interaction between HER-2 and
FAK, FAK and cSrc, cSrc and p130Cas, and between FAK and
p130Cas. It also decreased the activation and
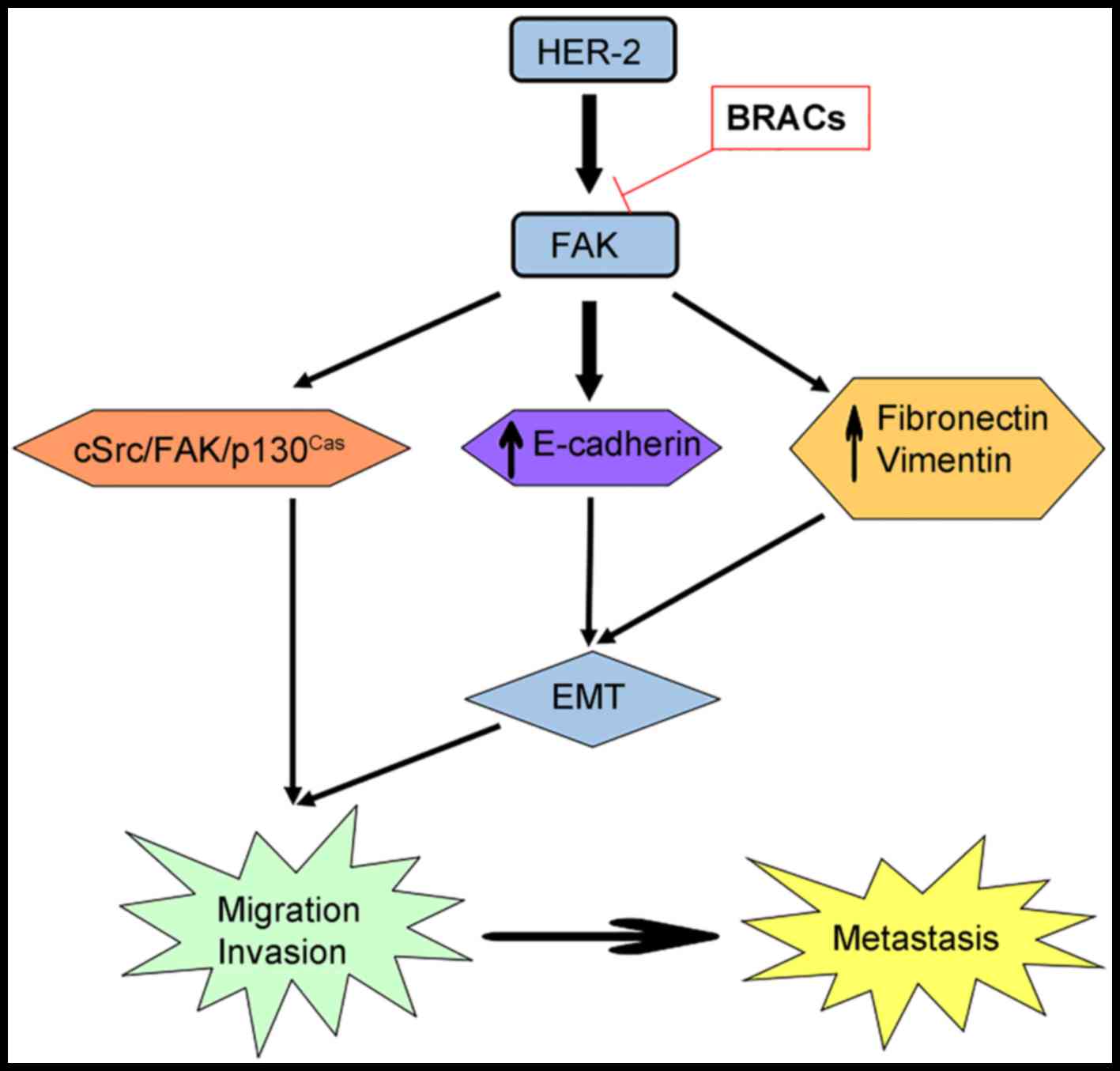
transduction of FAK signaling. Taken together, this study suggests
that BRACs suppress the metastasis of HER-2-positive breast cancer
in vitro, and that the cSrc/FAK/p130Cas pathway
plays a vital role in this inhibitory effect (Fig. 6). It has been demonstrated
anthocyanins from raspberry extract (RES) enhance the effects of
radiotherapy and chemotherapy in the treatment of pancreatic cancer
by controlling apoptosis and signaling pathways (35). Another study also indicated that
RES enhanced the susceptibility of centchroman in breast cancer
cells (36). These studies
suggest anthocyanins combined with anticancer drugs may be used to
enhance the anticancer effects. In conclusion, our results may
probe usefule to future studies on BRACs using animals and humans.
BRACs may have potential for use as supplementary treatments with
anticancer drugs.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (nos. 81273074 and 81573154), the
Innovation Research Team Project of Sichuan Province Youth Science
and Technology Foundation (no. 2014TD0021), and the Scientific
Research Innovation Team Project of Sichuan Provincial University
Foundation (no. 14TD00234). The authors would like to thank Mrs.
Weiwei Zhang for assisting with the revision of the manuscript.
References
|
1
|
Friedenreich CM: Physical activity and
breast cancer: Review of the epidemiologic evidence and biologic
mechanisms. Recent Results Cancer Res. 188:125–139. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zheng L, Zhou B, Meng X, Zhu W, Zuo A,
Wang X, Jiang R and Yu S: A model of spontaneous mouse mammary
tumor for human estrogen receptor- and progesterone
receptor-negative breast cancer. Int J Oncol. 45:2241–2249. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mendoza G, Portillo A and Olmos-Soto J:
Accurate breast cancer diagnosis through real-time PCR her-2 gene
quantification using immunohistochemically-identified biopsies.
Oncol Lett. 5:295–298. 2013.
|
|
5
|
Gutierrez C and Schiff R: HER2: Biology,
detection, and clinical implications. Arch Pathol Lab Med.
135:55–62. 2011.PubMed/NCBI
|
|
6
|
Slamon DJ, Godolphin W, Jones LA, Holt JA,
Wong SG, Keith DE, Levin WJ, Stuart SG, Udove J, Ullrich A, et al:
Studies of the HER-2/neu proto-oncogene in human breast and ovarian
cancer. Science. 244:707–712. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Heerboth S, Housman G, Leary M, Longacre
M, Byler S, Lapinska K, Willbanks A and Sarkar S: EMT and tumor
metastasis. Clin Transl Med. 4:62015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Heffler M, Golubovskaya VM, Dunn KM and
Cance W: Focal adhesion kinase autophosphorylation inhibition
decreases colon cancer cell growth and enhances the efficacy of
chemotherapy. Cancer Biol Ther. 14:761–772. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Golubovskaya VM: Targeting FAK in human
cancer: From finding to first clinical trials. Front Biosci
(Landmark Ed). 19:687–706. 2014. View
Article : Google Scholar
|
|
10
|
Vadlamudi RK, Sahin AA, Adam L, Wang RA
and Kumar R: Heregulin and HER2 signaling selectively activates
c-Src phosphorylation at tyrosine 215. FEBS Lett. 543:76–80. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dolfi F, Garcia-Guzman M, Ojaniemi M,
Nakamura H, Matsuda M and Vuori K: The adaptor protein Crk connects
multiple cellular stimuli to the JNK signaling pathway. Proc Natl
Acad Sci USA. 95:15394–15399. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Han C, Ding H, Casto B, Stoner GD and
D'Ambrosio SM: Inhibition of the growth of premalignant and
malignant human oral cell lines by extracts and components of black
raspberries. Nutr Cancer. 51:207–217. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fayyaz S, Aydin T, Cakir A, Gasparri ML,
Panici PB and Farooqi AA: Oleuropein mediated targeting of
signaling network in cancer. Curr Top Med Chem. 16:2477–2483. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Farooqi AA, Wang Z, Hasnain S, Attar R,
Aslam A, Mansoor Q and Ismail M: Citrus fruits and their bioactive
ingredients: Leading four horsemen from front. Asian Pac J Cancer
Prev. 16:2575–2580. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin X, Farooqi AA and Ismail M: Recent
progress in fungus-derived bioactive agents for targeting of
signaling machinery in cancer cells. Drug Des Devel Ther.
9:1797–1804. 2015.PubMed/NCBI
|
|
16
|
Perk AA, Shatynska-Mytsyk I, Gerçek YC,
Boztaş K, Yazgan M, Fayyaz S and Farooqi AA: Rutin mediated
targeting of signaling machinery in cancer cells. Cancer Cell Int.
14:1242014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Attar R, Tabassum S, Fayyaz S, Ahmad MS,
Nogueira DR, Yaylim I, Timirci-Kahraman O, Kucukhuseyin O, Cacina
C, Farooqi AA, et al: Natural products are the future of anticancer
therapy: Preclinical and clinical advancements of Viscum album
phytometabolites. Cell Mol Biol (Noisy-le-grand). 61:62–68.
2015.
|
|
18
|
Kong S, Kim DJ, Oh SK, Choi IS, Jeong HS
and Lee J: Black rice bran as an ingredient in noodles: Chemical
and functional evaluation. J Food Sci. 77:C303–C307. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sehitoglu MH, Farooqi AA, Qureshi MZ, Butt
G and Aras A: Anthocyanins: Targeting of signaling networks in
cancer cells. Asian Pac J Cancer Prev. 15:2379–2381. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hui C, Bin Y, Xiaoping Y, Long Y, Chunye
C, Mantian M and Wenhua L: Anticancer activities of an
anthocyanin-rich extract from black rice against breast cancer
cells in vitro and in vivo. Nutr Cancer. 62:1128–1136. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Luo LP, Han B, Yu XP, Chen XY, Zhou J,
Chen W, Zhu YF, Peng XL, Zou Q and Li SY: Anti-metastasis activity
of black rice anthocyanins against breast cancer: Analyses using an
ErbB2 positive breast cancer cell line and tumoral xenograft model.
Asian Pac J Cancer Prev. 15:6219–6225. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen XY, Zhou J, Luo LP, Han B, Li F, Chen
JY, Zhu YF, Chen W and Yu XP: Black Rice Anthocyanins Suppress
Metastasis of Breast Cancer Cells by Targeting RAS/RAF/MAPK
Pathway. Biomed Res Int. 2015:4142502015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu M, Bower KA, Wang S, Frank JA, Chen G,
Ding M, Wang S, Shi X, Ke Z and Luo J: Cyanidin-3-glucoside
inhibits ethanol-induced invasion of breast cancer cells
overexpressing ErbB2. Mol Cancer. 9:2852010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu M, Bower KA, Chen G, Shi X, Dong Z, Ke
Z and Luo J: Ethanol enhances the interaction of breast cancer
cells over-expressing ErbB2 with fibronectin. Alcohol Clin Exp Res.
34:751–760. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hou DX: Potential mechanisms of cancer
chemoprevention by anthocyanins. Curr Mol Med. 3:149–159. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Crozier A, Jaganath IB and Clifford MN:
Dietary phenolics: Chemistry, bioavailability and effects on
health. Nat Prod Rep. 26:1001–1043. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Scalbert A and Williamson G: Dietary
intake and bioavailability of polyphenols. J Nutr. 130(Suppl 8):
2073S–2085S. 2000.PubMed/NCBI
|
|
28
|
Manach C, Scalbert A, Morand C, Rémésy C
and Jiménez L: Polyphenols: Food sources and bioavailability. Am J
Clin Nutr. 79:727–747. 2004.PubMed/NCBI
|
|
29
|
Kamiloglu S, Capanoglu E, Grootaert C and
Van Camp J: Anthocyanin absorption and metabolism by human
intestinal caco-2 cells - A review. Int J Mol Sci. 16:21555–21574.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fang J: Bioavailability of anthocyanins.
Drug Metab Rev. 46:508–520. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen PN, Kuo WH, Chiang CL, Chiou HL,
Hsieh YS and Chu SC: Black rice anthocyanins inhibit cancer cells
invasion via repressions of MMPs and u-PA expression. Chem Biol
Interact. 163:218–229. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang LS and Stoner GD: Anthocyanins and
their role in cancer prevention. Cancer Lett. 269:281–290. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen JS, Huang XH, Wang Q, Huang JQ, Zhang
LJ, Chen XL, Lei J and Cheng ZX: Sonic hedgehog signaling pathway
induces cell migration and invasion through focal adhesion
kinase/AKT signaling-mediated activation of matrix
metalloproteinase (MMP)-2 and MMP-9 in liver cancer.
Carcinogenesis. 34:10–19. 2013. View Article : Google Scholar
|
|
34
|
Golubovskaya VM, Nyberg C, Zheng M, Kweh
F, Magis A, Ostrov D and Cance WG: A small molecule inhibitor,
1,2,4,5-benzenetetraamine tetrahydrochloride, targeting the y397
site of focal adhesion kinase decreases tumor growth. J Med Chem.
51:7405–7416. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Veeraraghavan J, Natarajan M, Lagisetty P,
Awasthi V, Herman TS and Aravindan N: Impact of curcumin, raspberry
extract, and neem leaf extract on rel protein-regulated cell
death/radiosensitization in pancreatic cancer cells. Pancreas.
40:1107–1119. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Singh N, Zaidi D, Shyam H, Sharma R and
Balapure AK: Polyphenols sensitization potentiates susceptibility
of MCF-7 and MDA MB-231 cells to Centchroman. PLoS One.
7:e377362012. View Article : Google Scholar : PubMed/NCBI
|