Introduction
Intestinal ischemia/reperfusion (I/R) can be a fatal
syndrome that occurs in a variety of important clinical scenarios,
including major trauma, septic shock, and small bowel or liver
transplantation (1). Acute
intestinal I/R results in the disruption of intestinal barrier
function, initiating the systemic inflammatory response syndrome,
and finally leading to the multiple organ dysfunction syndrome,
with high mortality rate (2,3).
Intestinal epithelial cells (IECs) are the most dominant sectors of
the intestinal epithelial barrier (IEB). Tight junctions (TJs),
which are composed of a cluster of transmembrane proteins
[including tight junction protein 1 (ZO-1), occludin and claudins]
and firmly connect adjacent epithelial cells, regulate the
permeability of ions, water and nutrients via the paracellular
pathway, establish cell polarity and are a primary determinant of
IEB function (4). Recent studies
have demonstrated that disruption of the expression and
distribution of TJs caused by intestinal I/R are responsible for
hyperpermeability and intestinal IEB dysfunction in several animal
models (5,6). The precise molecular mechanisms of
dynamic regulation of TJs under this pathophysiology remain
unclear.
Aryl hydrocarbon receptor (AhR), a ligand-dependent
transcription factor widely expressed in the cytosol of various
cells and tissues, is involved in numerous biological processes,
including cell proliferation, apoptosis, differentiation and the
inflammatory response (7–10). Upon binding with ligands, such as
6-formylindolo(3,2-b)carbazole (Ficz), activated AhR translocates
from the cytosol to the nucleus and dimerizes with aryl hydrocarbon
receptor nuclear translocator (ARNT) to initiate the transcription
of target genes, such as cytochrome P450 (CYP1A1), which contains
functional AhR responsive elements (AhREs). Recently, studies have
focused on the protective role of AhR activation in immune cells in
the gut. Increasing evidence has demonstrated that AhR activation
ameliorated mucosal injury in several animal models of experimental
colitis (11,12), whereas AhR-deficient mice
developed more severe colitis than their wild-type counterparts
(13–15). Although it has been previously
reported that AhR activation by MG132 alleviated liver injury
caused by intestinal I/R (16),
to the best of our knowledge, no study has investigated the effects
of AhR activation by Ficz on intestinal structure and barrier
function following intestinal I/R in vivo and under hypoxia
in vitro.
Previous studies have demonstrated that deficiency
of Notch1 signaling is involved in the disrupted distribution of
TJs and increased permeability, while restoration of epithelial
Notch1 activation is associated with normal barrier function in a
recombination activating 1 (RAG1)-adoptive transfer model of
colitis in vivo, and in the Caco-2 cell line in vitro
(17,18). Additionally, Notch1 signaling is
also involved in the regeneration and adaptation of intestinal
epithelium in acute and chronic pathological conditions, including
I/R (19,20). All these results indicate that
Notch1 signaling may be important for the maintenance of intestinal
renewal and barrier function.
Studies also confirmed that Notch1 has several AhREs
in its promoters and is induced by
2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD; an exogenous ligand of
AhR) and Ficz in hepatocytes, intestinal innate lymphocytes (ILCs)
and bone marrow-derived dendritic cells (BMDCs) (21–23). Notably, activated Notch signaling
promotes the production of certain unknown endogenous stimulators
of AhR in a concanavalin A-induced hepatitis model (24). As AhR and Notch1 are expressed in
the intestinal epithelium, and Notch1 signaling has been confirmed
to have a crucial role in maintaining the intestinal homeostasis
under different pathological conditions including I/R (17–20), the positive feedback loop of
AhR-Notch1 signaling may be a underlying mechanism in regulation of
the intestinal homeostasis under pathological conditions. In the
present study, we hypothesized that AhR activation by Ficz may
maintain the epithelial architecture and barrier function via
upregulation of Notch1 signaling in a mouse model of acute
intestinal I/R and cell model of hypoxia.
Materials and methods
Animals
The 6–8 week-old male C57BL/6 mice weighing 20–23 g
were purchased from the Laboratory Animal Center, the Third
Military Medical University (Chongqing, China). All animals were
housed in plastic boxes with ad libitum access to standard
rodent chow and water at 20–22°C with a constant 12-h light/dark
cycle. An initial adaptation period of 1 week was allowed prior to
the experiments. Mice were randomly divided into three groups: Sham
operation (sham; n=7), I/R (n=7), I/R + Ficz (Enzo Life Sciences,
Inc., Farmingdale, NY, USA) (n=7). All animal experiments and
protocols were approved by the Animal Care and Use Committee of the
Third Military Medical University. All efforts were made to
minimize the suffering of the mice.
Intestinal I/R and treatment
All animals were fasted for 12 h and were free to
drink water prior to surgery. All experimental protocols were
administrated under aseptic conditions. All mice were
intraperitoneally injected with 40 mg/kg pentobarbital as
anesthesia and then subjected to aseptic laparotomy at medioventral
line. For I/R and I/R + Ficz groups, the superior mesenteric artery
(SMA) of each mouse was occluded with a non-traumatic microvascular
clamp for 20 min. The mice in sham group received the same
operation except occlusion of the SMA. Then, all the clamps were
removed and the incisions were sutured layer-by-layer. With normal
saline (NS) vehicle, Ficz (1 µg/mouse) were administered
intraperitoneally to the mice in I/R + Ficz group to activate AhR.
Equal volumes of NS were administered by the same route to the mice
in other groups as control. Following the surgery, all mice were
given free access to food and water and sacrificed at 6 h after
reperfusion. The small bowel of each mouse was quickly excised and
processed for histological assessment and IEC isolation.
IECs isolation
IECs isolation was performed according to our
previous study (25). Briefly,
the whole small intestine was removed and placed in tissue culture
media [RPMI-1640, with 10% fetal calf serum (both from Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA)]. The section was
cut into 5-mm pieces, washed in an isolation buffer [190 mg
ethylenediaminetetraacetic acid (EDTA) and 80 mg
dichlorodiphenyltrichloroethane dissolved in 500 ml
phosphate-buffered saline (PBS)], and incubated in the same buffer
with continuous brisk stirring at 37°C for 30 min. The supernatant
was filtered rapidly through a glass wool column. Following
centrifugation at 400 × g at 4°C for 5 min, the pellets were
purified in 40% isotonic Percoll (GE Healthcare Life Sciences,
Logan, UT, USA) and cells recovered in the suspension were
collected for RNA or protein extraction.
Histological examination
Segments of treated jejunum were harvested, fixed in
4% paraformaldehyde at 4°C overnight for histological evaluation.
Tissues were embedded in paraffin, cut into 5-µm sections
and dehydrated successively in 70, 80, 95 and 100% gradient ethanol
at room temperature for 5 min for hematoxylin and eosin (H&E)
staining. The sections were stained with hematoxylin (1 g/l) for 5
min and eosin (5 g/l) for 3 min both at room temperature. The
intestinal mucosal injury was assessed under a light microscope
(Leica Microsystems GmbH, Wetzlar, Germany) at ×200 magnification
in accordance with the Chiu's intestinal mucosa injury criteria
(26) and graded from 0 to 5.
Immunohistochemistry staining
Tissues were fixed with 4% paraformaldehyde at 4°C
overnight, embedded in the paraffin, sliced into 5-µm
sections, dehydrated successively in 70, 80, 95 and 100% in the
gradient ethanol at room temperature for 5 min, treated with 0.3%
hydrogen peroxide in methanol at room temperature for 20 min.
Subsequently, the sections were soaked in citrate buffer (pH 6.0)
and heated by microwave for 20 min for antigen retrieval. The
sections were incubated with anti-AhR antibody (ab2770; 1:200;
Abcam, Cambridge, UK), anti-Notch1 antibody (D1E11; 3608; 1:200;
Cell Signaling Technology, Inc., Danvers, MA, USA), anti-ZO-1
(21773-1-AP; 1:200) and anti-occludin (13409-1-AP; 1:200)
antibodies (ProteinTech Group, Inc., Chicago, IL, USA) at 4°C
overnight and then blocking with 5% bovine serum albumin (BSA;
SA1020; ready to use; Wuhan Boster Biological Technology Ltd.,
Wuhan, China) at room temperature for 20 min. These sections were
successively incubated with biotinylated goat anti-mouse and rabbit
secondary antibody (SA1020, ready to use) and avidin-biotin complex
(SA1020; ready to use) (both from Wuhan Boster Biological
Technology Ltd.) at 37°C for 20 min. The peroxidase activities were
detected by diaminobenzidine (0.5 mg/ml) staining at room
temperature for 20 sec. Following counterstaining with hematoxylin
(1 g/l) at room temperature for 1 min, histological evaluation was
performed under a light microscope at ×400 magnification.
Detection of intestinal permeability
The transepithelium electrical resistance (TER;
Ω/cm2), which indicates tissue viability and IEB
function (27), was determined
according to Ohm's law. The permeability of the small bowel was
assessed by TER. Tissues were placed in modified Ussing chambers
(Physiologic Instruments, Inc., San Diego, CA, USA) and bathed with
5 ml Krebs buffer (110.0 mM NaCl, 3.0 mM CaCl2, 5.5 mM
KCl, 1.4 mM KH2PO4, 29.0 mM
NaHCO3, 1.2 mM MgCl2, pH 7.4) on mucosal and
serosal sides. The buffer was continuously oxygenated with 5%
CO2 in O2 and maintained at 37°C by
water-jacketed reservoirs. After a 20 min period of equilibration,
the detections were performed for up to 90 min in Ussing chambers.
TER was calculated by the injected short-circuit current and
spontaneous potential difference.
Cell culture and treatment
The human Caco-2 colon carcinoma cell line was
obtained from American Type Culture Collection (Manassas, VA, USA)
and maintained in a culture media composed of minimum essential
medium (MEM; Gibco; Thermo Fisher Scientific, Inc.) supplemented
with 15% fetal bovine serum (Gibco; Thermo Fisher Scientific,
Inc.), 1% non-essential amino acids (HyClone; GE Healthcare Life
Sciences, Logan, UT, USA), 100 U/ml penicillin, and 100
µg/ml streptomycin (Beyotime Institute of Biotechnology,
Haimen, China) at 37°C in a 5% CO2 humidified incubator
and subcultured by partial digestion with 0.25% trypsin and 0.53 mM
EDTA in PBS without Ca2+ or Mg2+. The culture
medium was changed every other day.
The in vitro hypoxic environment was used to
simulate in vivo I/R condition. Caco-2 cells were seeded on
6-well plates at a density of 1×106 cells/well. Once the
monolayers reached 70–80% confluence, they were cultured with
serum-free MEM basic media overnight and then exposed to hypoxic
(Hx) environments (1% O2, 5% CO2 and 94%
N2, and 90% humidity) at 37°C for 0, 6, 12 or 24 h, with
or without Ficz (Enzo Life Sciences, Inc.) or
N-[N-(3,5-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl
ester (DAPT; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) at
appropriate concentrations. The controls were incubated under
normoxic (Nx) conditions.
Immunofluorescence analysis
The confluent Caco-2 mono-layers cultured on glass
bottom cell culture dish were treated as described above and then
rinsed three times in PBS, fixed with 4% paraformaldehyde at 4°C
for 20 min, and then permeabilized using 0.2% Triton X-100 in PBS
at room temperature for 30 min. Following blocking in 5% BSA at
room temperature for 2 h, monolayers were incubated with mouse
anti-AhR antibody (ab2770; 1:50; Abcam), rabbit anti-Notch1
intracellular domain (NICD1) antibody (D1E11; 3608; 1:50; Cell
Signaling Technology, Inc.), rabbit anti-ZO-1 (21773-1-AP; 1:50)
and rabbit anti-occludin (13409-1-AP; 1:50) antibodies (both from
ProteinTech) at 4°C overnight. Monolayers were then washed three
times and incubated with fluorescein isothiocyanate-conjugated goat
anti-mouse (A0568; 1:300) or Cy3-conjugated goat anti-rabbit
(A0516; 1:300) secondary antibodies (Beyotime Institute of
Biotechnology) for 1 h. DAPI (1 mg/ml; Invitrogen; Thermo Fisher
Scientific, Inc.) was used for nuclear staining at room temperature
for 10 min. Following extensive rinsing, the images were captured
using a laser scanning fluorescence microscopy (TCS SP5; Leica
Microsystems GmbH).
Measurement of TER
The integrity of the confluent Caco-2 cells was
assessed by measuring the TER. Caco-2 cells were seeded on
Millicell filters (0.33 cm2 area, 0.4-µm pore
diameter and 6.5 mm diameter) at a density of 0.5×105
cells/cm2 as monolayers and experiments were performed
when confluent. The TER of the treated monolayers was detected
using a Millicell ERS-2 voltohmmeter (EMD Millipore, Billerica, MA,
USA) according to Ohm's law. TER (Ω/cm2) = (total
resistance − blank resistance) (Ω)/area (cm2).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA extracted by isothiocyanate/chloroform
extraction using TRIzol (Life Technologies; Thermo Fisher
Scientific, Inc.) was reversed into cDNA using a PrimeScript RT
reagent kit (Takara Bio, Inc., Otsu, Japan). The amplified cDNA was
utilized for the template DNA and performed with specific primers
for PCR assay. The RT reaction was performed in thermocycler
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) at 42°C for 2 min,
37°C for 15 min, and 85°C for 5 sec. Subsequently, The average
quantification cycle threshold (Ct) of the triplicate samples was
calculated as the quantity of gene pruduct and the relative mRNA
expression levels were analyzed by 2−ΔΔCt method
(28) by Rotor-Gene Q series
software (Qiagen GmbH, Hilden, Germany) in accordance with the
standard PCR conditions (94°C for 10 min, and 40 cycles of 45 sec
at 94°C, 30 sec at 59°C, 45 sec at 72°C). In the present study, the
following primers were used: β-actin (mouse) forward
5′-CTTCTTTGCAGCTCCTTCGTT-3′ and reverse
5′-AGGAGTCCTTCTGACCCATTC-3′; CYP1A1 (mouse) forward
5′-CCAAGAGCTGCTCAGCATAG-3′ and reverse 5′-GGCATCCAGGGAAGAGTTAG-3′;
Notch1 (mouse) forward 5′-ATGTCAATGTTCGAGGACCAG-3′ and reverse
5′-TCACTGTTGCCTGTCTCAAG-3′; hes family bHLH transcription factor 1
(HES-1; mouse) forward 5′-CCAGCCAGTGTCAACACGA-3′ and reverse
5′-AATGCCGGGAGCTATCTTTCT-3′. Data were collected and analyzed with
Rotor-Gene Q series software (Qiagen GmbH).
Western blotting
Proteins were extracted from the murine IECs and
Caco-2 monolayers in cold radioimmunoprecipitation assay buffer
[PBS, 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS, 1 µg/ml
phenylmethanesulfonyl fluoride, 1.0 mM sodium orthovanadate and 1X
mammalian protease inhibitor cocktail (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany)]. Following centrifugation at 14,462 × g at 4°C
for 15 min, proteins were quantified by the Bradford method
utilizing the bicinchoninic acid assay reagent (Beyotime Institute
of Biotechnology). Equal volumes of proteins were loaded into 10%
SDS-polyacrylamide gels and transferred onto 0.2-µm
polyvinylidene difluoride-Plus membranes. Following blocking with
5% BSA at room temperature for 1 h, the membranes were incubated at
4°C overnight with the follo wing primary antibodies: anti-GAPDH
antibody (sc-47724; 1:1,000; Santa Cruz Biotechnology, Inc.),
anti-AhR antibody (ab2770; 1:500; Abcam), anti-CYP1A1 antibody
(13241-1-AP; 1:500; ProteinTech), anti-NICD1 antibody (D1E11; 3608;
1:500; Cell Signaling Technology, Inc.), anti-HES-1 antibody
(sc-166378; 1:500; Santa Cruz Biotechnology, Inc.), anti-ZO-1
(21773-1-AP; 1:500) and anti-occludin (13409-1-AP; 1:500)
antibodies (both from ProteinTech). Following washing, the
membranes were incubated with horseradish peroxidase-conjugated
goat anti-mouse secondary antibody (BA1051; 1:5,000; Wuhan Boster
Biological Technology Ltd.) or horseradish peroxidase-conjugated
goat anti-rabbit secondary antibody (BA1055; 1:5,000; Wuhan Boster
Biological Technology Ltd.) for 1 h at room temperature and
visualized with an enhanced chemiluminescence system (EMD
Millipore) and imaging system (Kodak Gel Logic 4000R Imaging
System; Carestream Health, Inc., Rochester, NY, USA). Protein
expression was normalized to that of GAPDH and the densitometry of
the western blot bandings was analyzed using ImageJ software
(Version 1.50i; National institutes of Health, Bethesda, MD,
USA).
Statistical analysis
Data are presented as the mean ± standard deviation.
Differ ences among groups were evaluated by analysis of one-way
analysis of variance with Student-Newman-Keuls test using SPSS
statistical software package (version 13.0; SPSS, Inc., Chicago,
IL, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Ficz ameliorates the mucosal histological
lesions and loss of intestinal barrier function following I/R in
mice
It was demonstrated by our previous study that
intestinal I/R resulted in the intestinal barrier dysfunction in
mice, as indicated by the decreased TER and the increased
paracellular permeability (6).
Since AhR activation by MG132 has been reported to alleviate the
liver injury caused by intestinal I/R in rats (16), to investigate into the effect of
AhR activation by Ficz on I/R-induced intestinal barrier
dysfunction, the morphological structure of intestinal mucosa and
epithelial permeability were detected in the current study. The
representative images of the small intestine demonstrated that
edema and hyperemia were clear in the I/R group, but rarely
observed in the sham or IR + Ficz group (Fig. 1A). The histological examination
was performed on the basis of criteria proposed by Chiu et
al (26). The typical
histologic features exhibited in the I/R and I/R + Ficz groups were
characterized by shortening of the villi, shedding of the
epithelium, and prominent mucosal and mucodermal immune cell
infiltration (Fig. 1B). The
results indicated that Ficz significantly improved the intestinal
morphological lesions (scored 2.33±0.52), compared with those in
the I/R group (scored 4.50±0.55). Additionally, there was no damage
in the jejunal mucosa in the sham group (scored 0; Fig. 1C). To further confirm the effects
of Ficz on the maintenance of IEB function, TER was detected by
Ussing chambers as the permeability of intestinal mucosa (27). Intestinal I/R caused a markedly
decrease in TER (96.07±7.53 Ω/cm2) compared with the
sham group (145.4±10.79 Ω/cm2). Ficz partially rescued
the reduction in TER following intestinal I/R (123.8±5.56
Ω/cm2), though the TER remained lower than that in the
sham group (Fig. 1D). These
findings revealed that Ficz significantly attenuated the
I/R-induced epithelial barrier dysfunction.
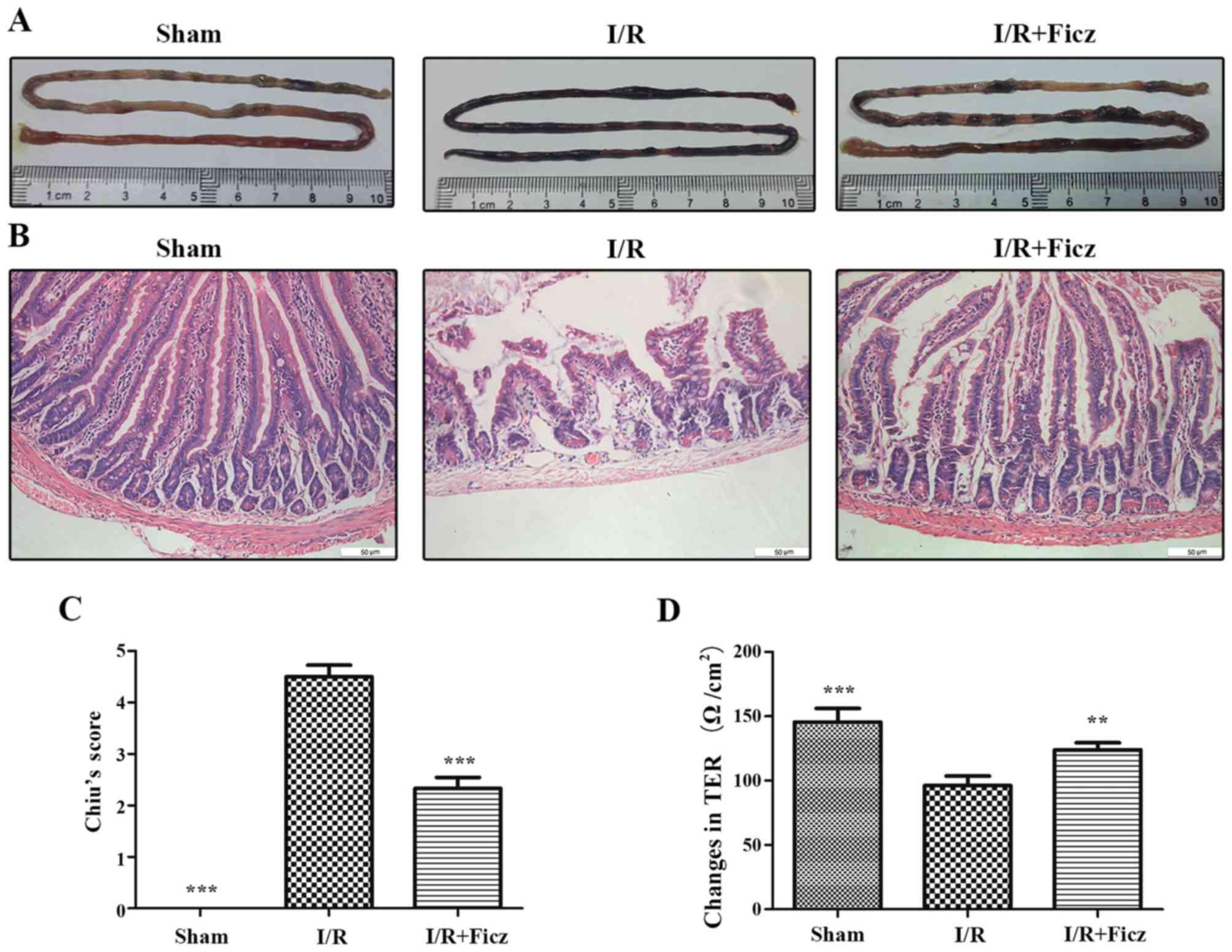 | Figure 1Ficz alleviates the mucosal damage
and hyperpermeability induced by murine I/R. (A) Representative
images of small intestine harvested from three groups (sham, I/R
and I/R + Ficz). (B) Hematoxylin and eosin staining of the gut
tissues captured by inverted microscope (original magnification,
×200; scale bar, 50 µm; n=5). (C) Graphic representation of
Chiu's score according to (B). (D) Epithelial permeability was
assessed by TER, which was detected using Ussing chambers. Data are
presented as the mean ± standard deviation. **P<0.01,
***P<0.001 vs. I/R group, n=3. Ficz,
6-formylindolo(3,2-b)carbazole; I/R, ischemia/reperfusion; TER,
transepithelium electrical resistance. |
Ficz maintains the expression and
distribution of TJ proteins following intestinal I/R
Our previous study confirmed that intestinal I/R
lead to the disrupted distribution of TJs (5). To determine the effects of AhR
activation on the development of TJs following I/R in mice,
immunohistochemistry staining and western blotting were performed
to detect the distribution and expression of TJ proteins, including
ZO-1 and occludin. The results demonstrated that, in the sham
group, ZO-1 and occludin were located at the apex of the villous
enterocytes in a chicken wire-like pattern. The distribution of
ZO-1 and occludin was clearly disrupted following intestinal I/R
(Fig. 2A). The ZO-1 and occludin
protein expression in the small bowel following intestinal I/R was
also significantly decreased compared with the sham group (Fig. 2B and C). However, Ficz ameliorated
the altered distribution of ZO-1 and occludin following intestinal
I/R, and normal structures reappeared and were maintained (Fig. 2A). Ficz also markedly increased
the protein expression of ZO-1 and occludin following intestinal
I/R compared with the I/R group (Fig.
2B and C). These results suggested that AhR activation by Ficz
has an important effect on in the development of TJs following
intestinal I/R.
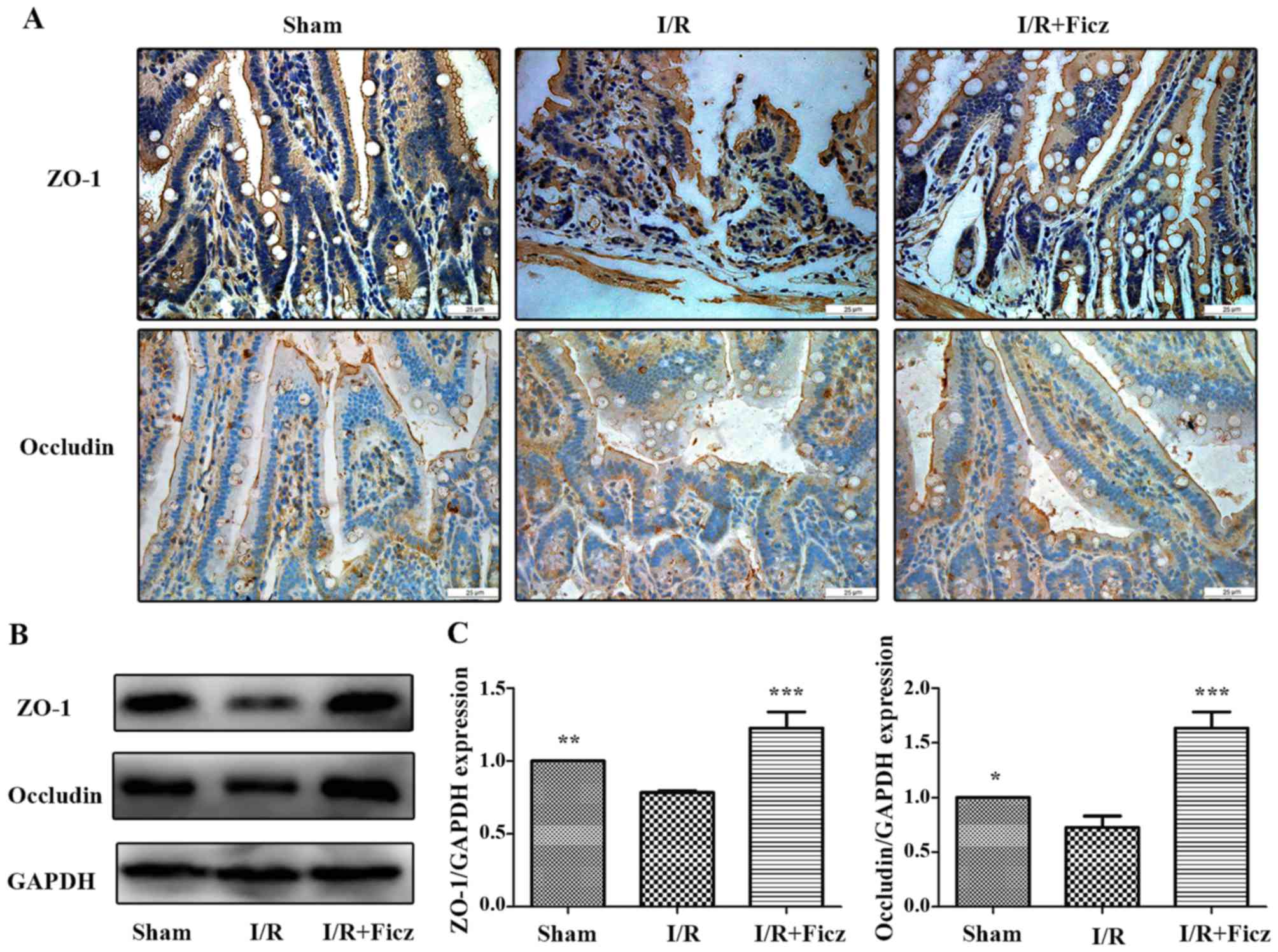 | Figure 2Ficz maintains the integrity and
sufficient quantity of tight junctions (ZO-1 and occludin)
following murine I/R. (A) Expression and distribution of ZO-1 and
occludin in three groups (sham, I/R and I/R + Ficz) were detected
by immunohistochemistry staining (original magnification, ×400;
scale bar, 25 µm; n=3). (B) Western blotting of ZO-1,
occludin and GAPDH protein levels. (C) Relative expression of
proteins in (B), GAPDH was used to verify equivalent loading. Data
are presented as the mean ± standard deviation.
*P<0.05, **P<0.01,
***P<0.001 vs. I/R group; n=3. I/R,
ischemia/reperfusion; Ficz, 6-formylindolo(3,2-b)carbazole; ZO-1,
tight junction protein 1. |
Activation of AhR by Ficz upregulates
epithelial Notch1 signaling following intestinal I/R in mice
Previous studies have reported that AhR activation
by endogenous and exogenous ligands can induce the expression of
Notch1 gene expression (21–23), and AhR and Notch1 signaling have
important roles in maintaining intestinal epithelial homeostasis;
thus, the effects of I/R and Ficz on the expression and activity of
AhR-Notch1 signaling pathway were investigated in the current
study. As CYP1A1 is part of the AhR gene battery (29) and is induced through AhR
signaling, CYP1A1 induction was used as an index to assess the
activity of the AhR signaling pathway (30). NICD1 and HES-1 expression were
used to determine the activity of Notch1 signaling. The results
demonstrated that AhR and NICD1 were expressed in the nucleus and
cytosol of intestinal epithelium and in the lamina propria
(Fig. 3A). I/R in mice reduced
the expression and activity of AhR-Notch1 signaling, while Ficz
administration (1 µg/mouse) significantly upregulated the
expression and activity of these signaling pathways, as indicated
by the elevated AhR, CYP1A1, NICD1 and HES-1 mRNA and protein
expression levels (Fig. 3B–D),
which suggested that activation of AhR by Ficz upregulated
epithelial Notch1 signaling following intestinal I/R in mice.
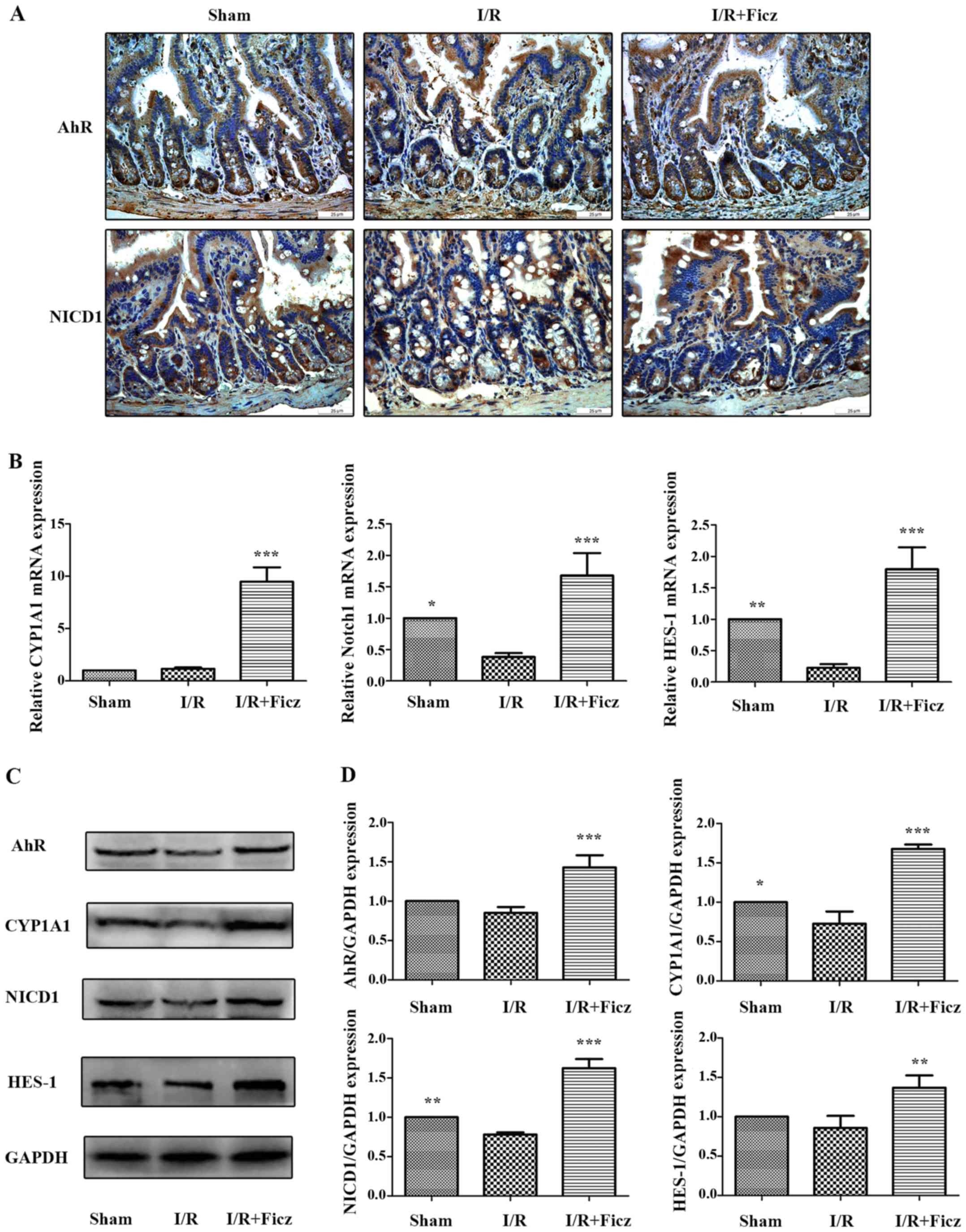 | Figure 3Ficz activates the epithelial
AhR-Notch1 signaling pathway following intestinal I/R in mice. (A)
AhR and Notch1 expression in sham, I/R and I/R + Ficz groups
detected by immunohistochemistry staining (original magnification,
×400; scale bar, 25 µm; n=3). (B) Reverse
transcription-quantitative polymerase chain reaction analysis of
the mRNA levels of the CYP1A1, Notch1 and HES-1 in mice treated as
in (A). (C) Western blotting of AhR, CYP1A1, NICD1, HES-1 and GAPDH
protein levels. (D) Relative expressions of proteins determined by
densitometry, GAPDH was used to verify equivalent loading. Data are
presented as the mean ± standard deviation. *P<0.05,
**P<0.01, ***P<0.001 vs. I/R group;
n=3. I/R, ischemia/reperfusion; Ficz, 6-formylindolo(3,2-b)
carbazole; AhR, aryl hydrocarbon receptor; NICD1, Notch1
intracellular domain; CYP1A1, cytochrome P450; HES-1, hes family
bHLH transcription factor 1. |
AhR activation protects TJs against
hypoxia-induced morphological destruction via upregulation of
Notch1 signaling in Caco-2 monolayers
To further elucidate the underlying mechanism of the
protective effects of AhR activation following I/R, Caco-2 cell
lines were subjected to hypoxic conditions in vitro. As is
demonstrated in Figs. 4 and
5, 12 h of hypoxia was optimal to
mimic the in vivo I/R and the optimum concentration of Ficz
to activate AhR-Notch1 signaling was 100 nM. Immunofluorescence
demonstrated that AhR and NICD1 were expressed in the cytosol of
the monolayers, and following treatment with Ficz, the AhR and
NICD1 appeared to have increased expression in the nuclei of the
monolayers (Fig. 6A). Western
blotting demonstrated that hypoxia downregulated the expression of
AhR, NICD1, CYP1A1 and HES-1, while administration of Ficz
significantly increased the expression of the proteins, and thus,
activated AhR-Notch1 signaling under normoxic and hypoxic
conditions (Fig. 6B and C).
Immunofluorescence and western blotting were also performed to
detect the formation and distribution of epithelial TJs (ZO-1 and
occludin). ZO-1 and occludin were predominantly expressed on the
membrane of the monolayers, however occludin was also detected in
the cytoplasm (Fig. 7A). The data
demonstrated that hypoxia reduced expression and disrupted the
distribution of ZO-1 and occludin, whereas Ficz maintained their
expression and distribution in Caco-2 monolayers under hypoxic
conditions (Fig. 7B and C), in
accordance with the in vivo experiments. TER was used to
assess the endothelial barrier function. Compared with the normoxic
condition (62.67±4.41 Ω/cm2), the TER of Caco-2
monolayers decreased markedly when exposed to hypoxic conditions
(38.20±2.39 Ω/cm2), while administration of Ficz under
hypoxic conditions restored the TER (51.4±03.15 Ω/cm2;
Fig. 7D). Overall, activation of
AhR by Ficz was confirmed to maintain the epithelial barrier
function under hypoxic conditions and maintain ZO-1 and occludin
expression/distribution by activation of Notch1 signaling in
vitro.
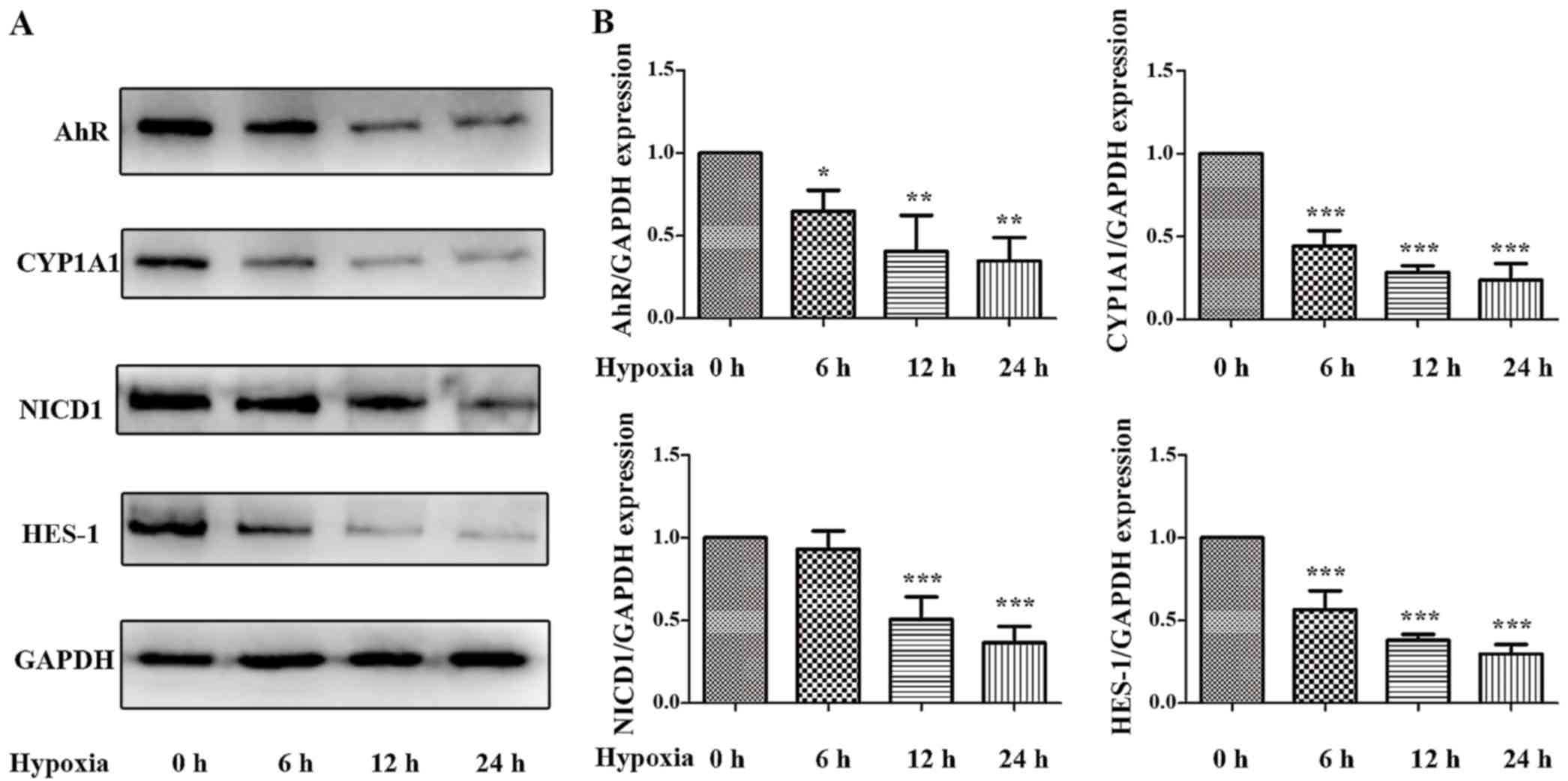 | Figure 4Effects of hypoxia on the expression
and activity of AhR-Notch1 signaling pathway in Caco-2 monolayers.
Caco-2 cells were subjected to hypoxic condition for 0, 6, 12 and
24 h. (A) Western blotting of AhR, CYP1A1, NICD1, HES-1 and GAPDH
protein levels. (B) Relative expressions of proteins determined
using desittometry. GAPDH was used to verify equivalent loading.
Data are presented as mean ± standard deviation.
*P<0.05, **P<0.01,
***P<0.001 vs. hypoxia 0 h group; n=3. AhR, aryl
hydrocarbon receptor; CYP1A1, cytochrome P450; NICD1, Notch1
intracellular domain; HES-1, hes family bHLH transcription factor
1 |
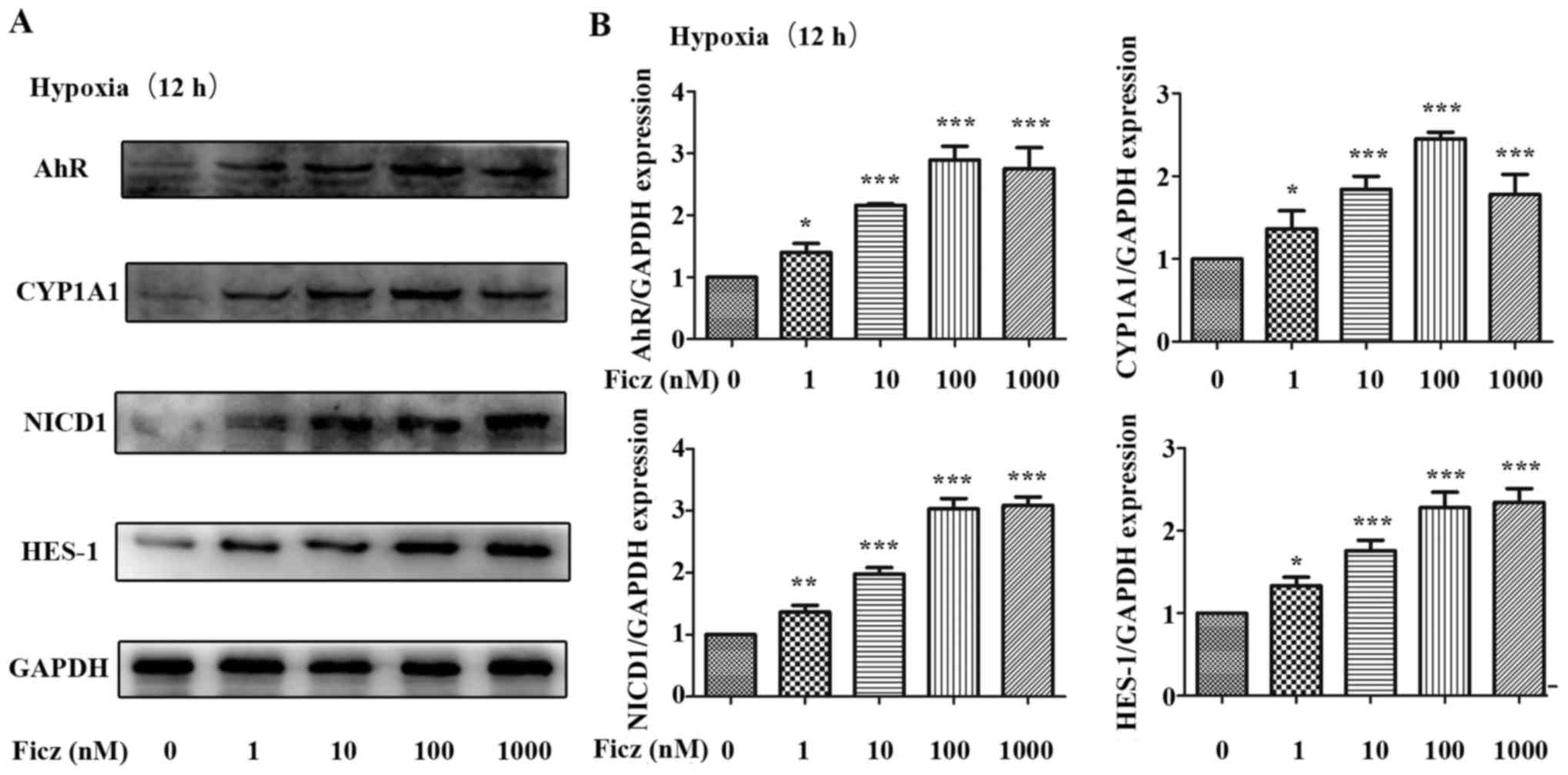 | Figure 5Effects of Ficz on the expression and
activity of AhR-Notch1 signaling pathway in hypoxic Caco-2 cells.
Caco-2 monolayers were administered with Ficz at five different
concentrations (0, 1, 10, 100, 1,000 nM) respectively and then
exposed to hypoxia for 12 h. (A) Western blotting of AhR, CYP1A1,
NICD1, HES-1 and GAPDH protein levels. (B) Relative protein
expressions determined by densitometry, GAPDH was used to verify
equivalent loading. Data are presented as the mean ± standard
deviation. *P<0.05, **P<0.01,
***P<0.001 vs. Ficz 0 nM group; n=3. Ficz,
6-formylindolo(3,2-b)carbazole; AhR, aryl hydrocarbon receptor;
CYP1A1, cytochrome P450; NICD1, Notch1 intracellular domain; HES-1,
hes family bHLH transcription factor 1. |
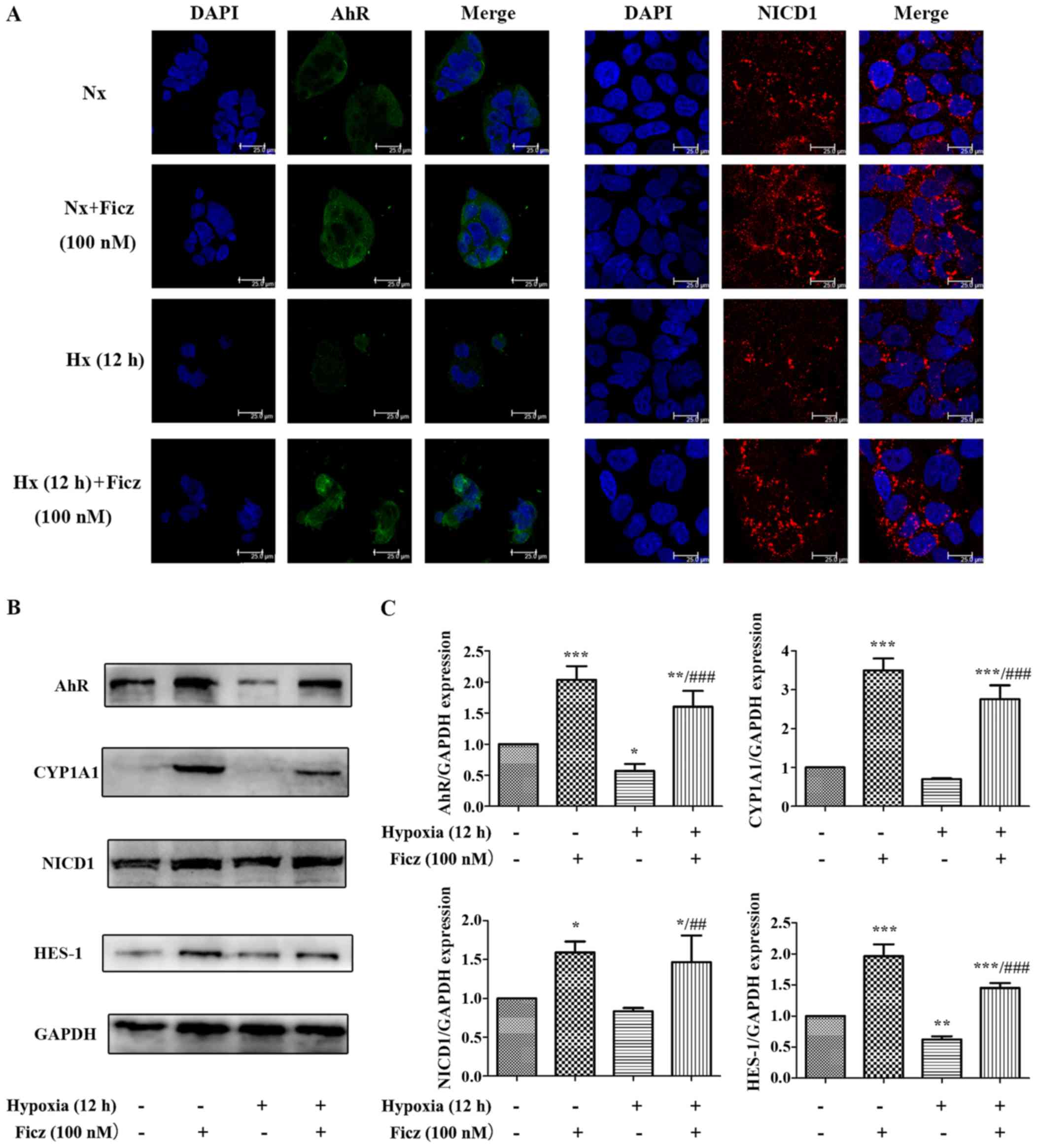 | Figure 6Ficz activates AhR-Notch1 signaling
in Caco-2 cells under normoxic and hypoxic conditions. (A)
Immunofluorescence staining was performed to evaluate the
expression of AhR (green) and NICD1 (red) in four groups (Nx, Nx +
Ficz, Hx and Hx + Ficz). Nuclei were counterstained with DAPI
(blue; original magnification, ×1,200; scale bar, 25 µm;
n=3). (B) Western blotting was used to measure the protein levels
of AhR, CYP1A1, NICD1, HES-1 and GAPDH. (C) Relative protein
expressions determined by densitometry, GAPDH was used to verify
equivalent loading. Data are presented as the mean ± standard
deviation. *P<0.05, **P<0.01,
***P<0.001 vs. Nx group; ##P<0.01,
###P<0.001 vs. Hx group; n=3. AhR, aryl hydrocarbon
receptor; NICD1, Notch1 intracellular domain; Nx, normoxia; Ficz,
6-formylindolo(3,2-b)carbazole; Hx, hypoxia; CYP1A1, cytochrome
P450; HES-1, hes family bHLH transcription factor 1. |
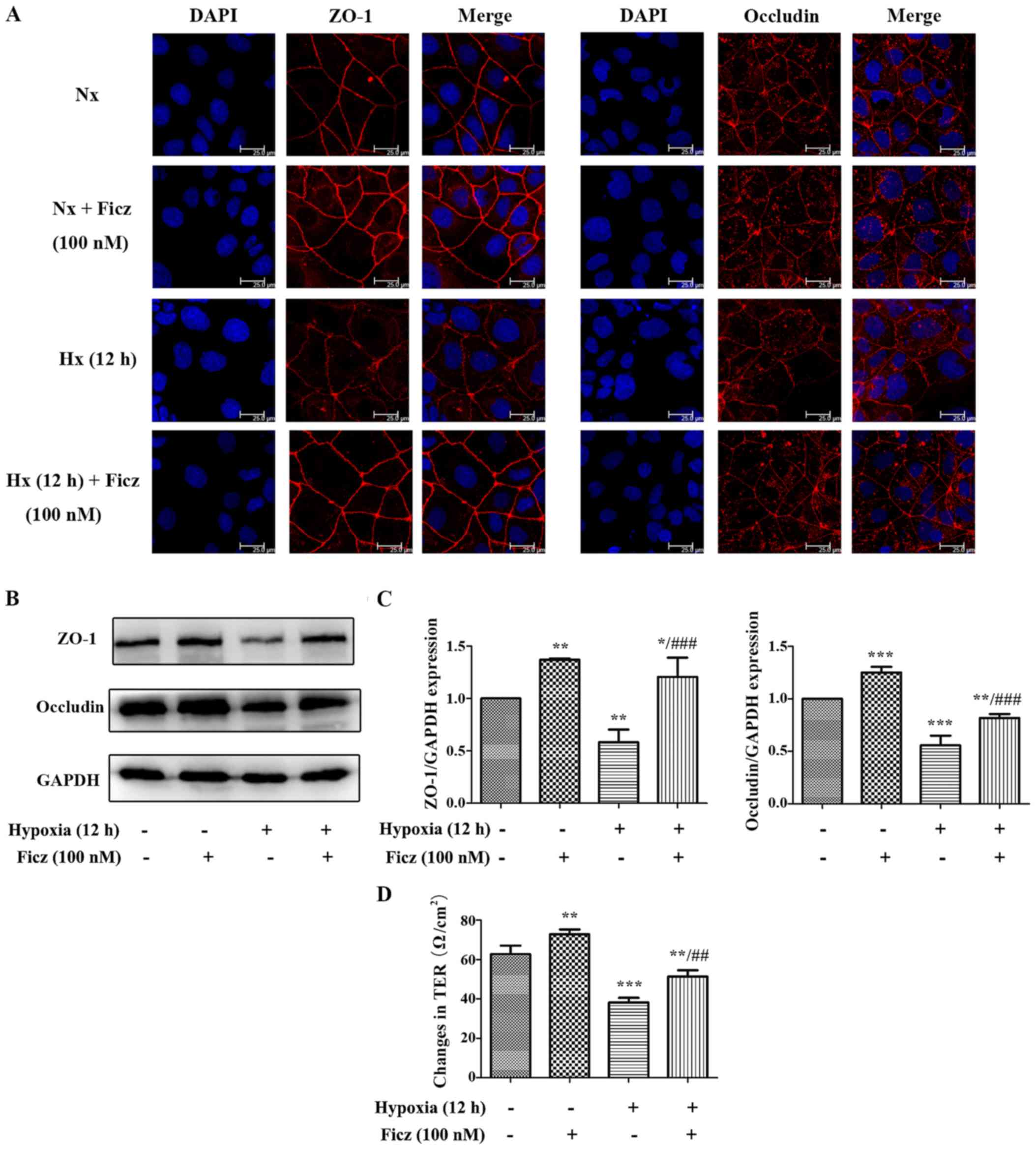 | Figure 7Ficz protects the tight junctions
(ZO-1 and occludin) from hypoxia-induced morphological damage and
maintains the barrier function in Caco-2 monolayers. (A) The
expression and distribution of ZO-1 and occluldin (red) were
detected by immunofluorescence staining in four groups (Nx, Nx +
Ficz, Hx and Hx + Ficz). Nuclei were counterstained with DAPI
(blue) (original magnification, ×1,200; scale bar, 25 µm;
n=3). (B) Western blotting was employed to investigate the protein
expression of ZO-1, occludin and GAPDH. (C) Relative protein
expressions determined by densitometry, GAPDH was used to verify
equivalent loading. (D) The TER was detected by the Millipore
electric resistance system to evaluate the epithelial permeability
of Caco-2 cells in four groups. Data are presented as the mean ±
standard deviation. *P<0.05, **P<0.01,
***P<0.001 vs. Nx group; ##P<0.01,
###P<0.001 vs. Hx group; n=3. ZO-1, tight junction
protein 1; Nx, normoxia; Ficz, 6-formylindolo(3,2-b)carbazole; Hx,
hypoxia; TER, transepithelium electrical resistance. |
Blocking Notch1 signaling with DAPT
counteracts the effects of Ficz on the development of TJs in
hypoxic Caco-2 monolayers
To determine whether the upregulation of Notch1
signaling was the underlying mechanism of AhR activation in
maintaining the endothelial barrier function in Caco-2 monolayers,
20 µM DAPT, a γ-secretase inhibitor, was administrated to
block the Notch1 signaling (31).
The results demonstrated that, under hypoxic conditions, the AhR
and CYP1A1 expression in Caco-2 monolayers were similar in the Hx +
Ficz and Hx + Ficz + DAPT groups (Fig. 8A and B), suggesting that DAPT had
no effect on the upstream AhR signaling. Additionally, the
expression levels of NICD1 and HES-1 in the Hx + DAPT group were
significantly decreased compared with the Hx + Ficz group, while
the levels in the Hx + Ficz + DAPT group remained similar to the Hx
+ DAPT group, suggesting that the inhibitory effect of DAPT on
Notch1 signaling was strong and cannot be altered by Ficz (Fig. 8A and B). As expected, the ZO-1 and
occludin expression were not elevated by Ficz when combined with
DAPT under hypoxic conditions (Fig.
8A and B), suggesting that the potential mechanism of AhR
activation in the development of TJs in hypoxic Caco-2 cells is
mediated by upregulation of Nocth1 signaling.
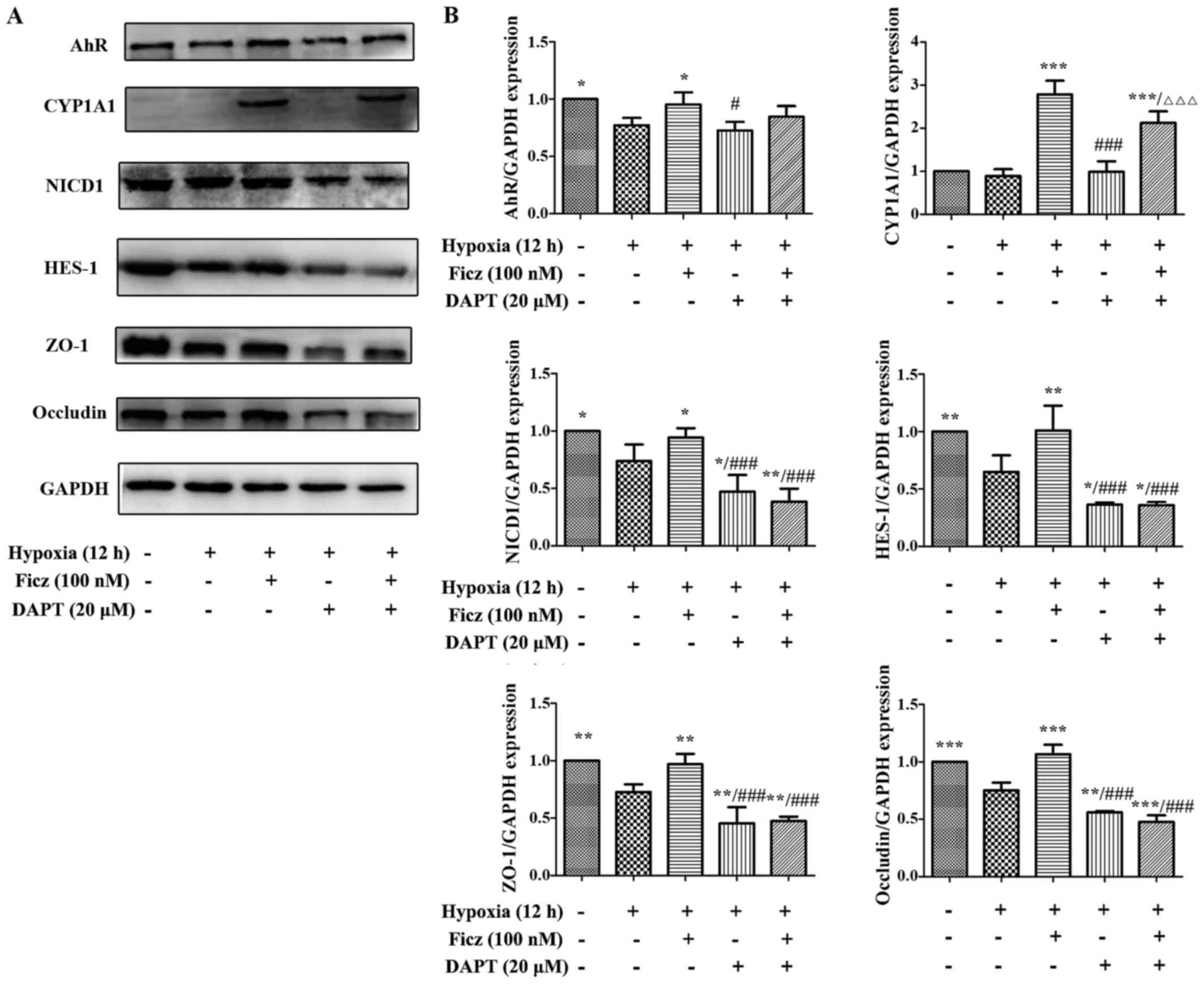 | Figure 8Blocking Notch1 signaling with DAPT
obstructed the effects of Ficz on the development of tight
junctions in hypoxic Caco-2 cells. (A) Western blottting was
performed to assess the protein levels of aryl hydrocarbon receptor
(AhR)-Notch1 signaling components and TJs (ZO-1 and occludin) in 5
groups (Nx, Hx, Hx + Ficz, Hx + DAPT and Hx + Ficz + DAPT). (B)
Relative protein expressions determined by densitometry, GAPDH was
used to verify equivalent loading. Data are presented as the mean ±
standard deviation. *P<0.05, **P<0.01,
***P<0.001 vs. Hx group; ###P<0.001 vs.
Hx + Ficz group; ∆∆∆P<0.001 vs. Hx+DAPT group; n=3.
AhR, aryl hydrocarbon receptor; NICD1, Notch1 intracellular domain;
CYP1A1, cytochrome P450; HES-1, hes family bHLH transcription
factor 1; ZO-1, tight junction protein 1; Ficz,
6-formylindolo(3,2-b)carbazole; DAPT,
N-[N-(3,5-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl
ester; Nx, normoxia; Hx, hypoxia. |
Discussion
Intestinal I/R injury is a major clinical challenge
that occurs in patients with severe trauma, midgut volvulus,
mesenteric artery embolism, or following other vascular and
abdominal surgeries (1,32). The interruption of intestinal
blood flow rapidly reduces tissue oxygenation, damages intestinal
epithelium and increases mucosal permeability. Although the
pathophysiological progression of intestinal I/R is well
understood, currently, there is no efficient solution to solve this
problem. In the present study, the potential effect of AhR
activation by Ficz in modulating intestinal barrier function in a
mouse model of intestinal I/R was investigated. The results
demonstrated that activation of AhR by Ficz upregulated the Notch1
signaling pathway, maintained the intestinal histomorphology and
osmotic balance and reversed the disrupted distribution of TJs
following intestinal I/R in mice or hypoxia in Caco-2 monolayers.
Furthermore, inhibition of Notch1 signaling by DAPT reversed the
effects of Ficz on TJs in hypoxic Caco-2 cells. In this way, it is
clear that upregulation of Notch1 signaling is the underlying
mechanism by which AhR activation alleviates intestinal barrier
dysfunction following I/R.
Ficz is an endogenous AhR ligand with higher
affinity and lower toxicity than TCDD (33). To explore the underlying molecular
mechanism, intraperitoneal administration of Ficz to activate AhR
signaling in the mouse I/R model. Previous studies have
demonstrated that AhR is downregulated in intestinal tissues of
patients with inflammatory bowel disease. Activation of AhR by Ficz
maintained the survival rate, body weight and development of
histopathological lesions, downregulated proinflammatory cytokines
and altered the intraepithelial lymphocyte phenotype in animal
models of colitis induced by dextran sodium sulfate or
trinitrobenzene sulfonic acid (11,12). Jing et al (16) also reported that the expression
and activity of AhR were downregulated in rat liver tissues
following intestinal I/R, while MG132 administration maintained the
expression and activity of AhR and alleviated the liver injury
under intestinal I/R conditions. These findings are in accordance
with the results of the current study indicating that intestinal
I/R reduced the expression and activity of AhR, while Ficz
increased the expression and activity of AhR following intestinal
I/R, suggesting that AhR signaling may be involved in the
intestinal mucosal injury induced by I/R.
Encouraged by these findings, whether activation of
AhR by Ficz had protective effects against intestinal I/R was also
investigated. Histological examination was performed to estimate
the effects of Ficz on the morphological changes following
intestinal I/R. The results suggested that the intestinal
morphological features arising from intestinal I/R were
characterized by shortening of the villi, shedding of the
epithelium, multiple erosions, immune cell infiltration, hemorrhage
and necrosis. The results were consistent with previous studies on
animal models of intestinal I/R (6,34).
The intestinal mucosal lesions induced by I/R were more severe than
those of the sham group, while Ficz markedly alleviated the
intestinal mucosal lesions following intestinal I/R, which
indicated that activation of AhR by Ficz could maintain the
intestinal mucosal morphology in a mouse model of acute intestinal
I/R.
AhR regulation of Notch signaling has been confirmed
by numerous studies. Accumulating evidence has indicated that
several AhREs are present in Notch1 gene promoters, and AhR ligands
(TCDD, Ficz) upregulated their transcription in hepatocytes, ILCs
and BMDCs (21–23). Alam et al (24) reported that Notch signaling
induced endogenous stimulation of AhR in activated CD4+
T cells. Additionally, Notch signaling downregulated microRNA-223,
which was identified as a negative regulator of the AhR/ARNT
pathway (35). Considering these
previous findings, the effects of AhR activation by Ficz on the
expression and activity of epithelial Notch1 signaling following
intestinal I/R in mice were investigated in the current study. Our
previous study demonstrated that Notch1 signaling was activated at
2 h after intestinal I/R in rats, whereas after 6 h of reperfusion,
the Notch1 mRNA level and NICD1 protein level were decreased
(20). This temporary activation
of Notch1 signaling may due to the stress response induced by the
early stage of I/R. Consistent with these findings, the results of
the present study demonstrated that the expression of NICD1 and
HES-1 decreased following intestinal I/R while AhR activation by
Ficz significantly increased the expression of NICD1 and HES-1
following intestinal I/R, which confirmed that AhR activation by
Ficz upregulated Notch1 signaling in intestinal epithelial cells
following intestinal I/R.
TJs maintain IEB function by regulating the
permeability to water, ions and nutrients (4). The alteration of the TJ function is
dynamically regulated by various signaling pathways, including
mitogen-activated protein kinases, myosin light-chain kinase and
phosphoinositide 3-kinase/protein kinase B (36). However, Dahan et al
(17) reported that activation of
the epithelial Notch1 signaling pathway by CD4+ T cells
adoptive transfer maintained claudin-5 expression in colonic
tissues of recombination activating gene 1-null mice, and
interference of Notch1 signaling in Caco-2 cells increased the
expression of claudin-2, while occludin expression decreased.
Mathern et al (18) also
repo rted that in Notch1 small interfering RNA-treated mice, the
expression of claudin-5 in colonic tissues was also significantly
decreased. All these findings supported that Notch1 signaling may
have a crucial role in maintaining the intestinal barrier function
by regulating the development of TJ proteins. Additionally, Pope
et al (37) demonstrated
that upregulation of claudin-1 in colonic epithelium activated
Notch1 signaling by increasing matrix metalloproteinase-9 and
phospho-ERK signaling, this suggested that TJs and Notch1 signaling
may have an interaction that maintains the intestinal barrier
function. Movahedan et al (38) also reported that the ZO-1
formation is delayed in Notch1-null corneal epithelial cells in
vitro. Thus, it was speculated whether activation of AhR-Notch1
signaling by Ficz alleviates the barrier dysfunction by maintaining
the expression of TJ proteins and TJ architecture following
intestinal I/R. The results of the current study demonstrated that
the intestinal I/R caused disrupted expression and distribution of
TJs (ZO-1 and occludin) and increased intestinal mucosal
permeability, which was in accordance with our previous studies
(5,6). Ficz ameliorated the disruption of
TJs and maintained the mucosal permeability following intestinal
I/R. These results indicated that Ficz may have a vital role in the
maintenance of the intestinal barrier function under I/R
conditions, potentially mediated via the activation of AhR-Notch1
signaling.
A Caco-2 cell model of hypoxia (12 h) was used to
mimic intestinal I/R and to explore the role of AhR activation in
the regulation of endothelial barrier function in vitro. As
with murine I/R, activated AhR maintained the epithelial barrier
function and paracellular permeability under hypoxic conditions in
Caco-2 cells through the recovery of TJs (ZO-1 and occludin). The
results also demonstrated that activation of AhR by Ficz repressed
the downregulation of Notch1 signaling induced by hypoxia. The
activity of AhR-Notch1 signaling and TJ protein expression were
detected in Caco-2 monolayers following treatment with DAPT, a
blocker of Notch signaling. The data confirmed that, in hypoxic
condition, blocking of Notch1 signaling with DAPT had marked effect
on the expression and activity of AhR with or without Ficz. By
contrast, loss of Notch1 counteracted the development of TJs (ZO-1
and occludin) induced by Ficz. Activated AhR was demonstrated to
have an important role in epithelial barrier defense under hypoxic
conditions, and the upregulation of the Notch1 signaling pathway
has been demonstrated to be the potential molecular mechanism
involved.
In conclusion, in the present study has provided
novel insights into the protective role of AhR in regulation of
intestinal barrier dysfunction induced by I/R in vivo and
in vitro. Activation of AhR may be a novel approach for the
treatment of acute intestinal I/R in the clinic.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (grant no. NSFC 81330013 to
H.Y., grant no. NSFC 81300275 to L.H.S, grant no. NSFC 81270451 to
W.D.X. and grant no. NSFC 81501661 to M.Y), and the Program of
Changjiang Scholars and Innovative Research (grant no. IRT 13050 to
HY).
References
|
1
|
Arda-Pirincci P and Bolkent S: The role of
epidermal growth factor in prevention of oxidative injury and
apoptosis induced by intestinal ischemia/reperfusion in rats. Acta
Histochem. 116:167–175. 2014. View Article : Google Scholar
|
|
2
|
Granger DN and Korthuis RJ: Physiologic
mechanisms of post-ischemic tissue injury. Annu Rev Physiol.
57:311–332. 1995. View Article : Google Scholar
|
|
3
|
Matthijsen RA, Derikx JP, Kuipers D, van
Dam RM, Dejong CH and Buurman WA: Enterocyte shedding and
epithelial lining repair following ischemia of the human small
intestine attenuate inflammation. PLoS One. 4:e70452009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee SH: Intestinal permeability regulation
by tight junction: implication on inflammatory bowel diseases.
Intest Res. 13:11–18. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang Y, Qiu Y, Wang W, Xiao W, Liang H,
Zhang C and Yang H, Teitelbaum DH, Sun LH and Yang H: Adenosine A2B
receptor modulates intestinal barrier function under hypoxic and
ischemia/reperfusion conditions. Int J Clin Exp Pathol.
7:2006–2018. 2014.PubMed/NCBI
|
|
6
|
Cai Y, Wang W, Liang H, Sun L, Teitelbaum
DH and Yang H: Keratinocyte growth factor improves epithelial
structure and function in a mouse model of intestinal
ischemia/reperfusion. PLoS One. 7:e447722012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xie G, Peng Z and Raufman JP: Src-mediated
aryl hydrocarbon and epidermal growth factor receptor cross talk
stimulates colon cancer cell proliferation. Am J Physiol
Gastrointest Liver Physiol. 302:G1006–G1015. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Marlowe JL, Fan Y, Chang X, Peng L,
Knudsen ES, Xia Y and Puga A: The aryl hydrocarbon receptor binds
to E2F1 and inhibits E2F1-induced apoptosis. Mol Biol Cell.
19:3263–3271. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mandavia C: TCDD-induced activation of
aryl hydrocarbon receptor regulates the skin stem cell population.
Med Hypotheses. 84:204–208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qiu J, Heller JJ, Guo X, Chen ZM, Fish K,
Fu YX and Zhou L: The aryl hydrocarbon receptor regulates gut
immunity through modulation of innate lymphoid cells. Immunity.
36:92–104. 2012. View Article : Google Scholar :
|
|
11
|
Monteleone I, Rizzo A, Sarra M, Sica G,
Sileri P, Biancone L, MacDonald TT, Pallone F and Monteleone G:
Aryl hydrocarbon receptor-induced signals up-regulate IL-22
production and inhibit inflammation in the gastrointestinal tract.
Gastroenterology. 141:237–248. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ji T, Xu C, Sun L, Yu M, Peng K, Qiu Y,
Xiao W and Yang H: Aryl hydrocarbon receptor activation
down-regulates IL-7 and reduces inflammation in a mouse model of
DSS-induced colitis. Dig Dis Sci. 60:1958–1966. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qiu J, Guo X, Chen ZM, He L, Sonnenberg
GF, Artis D, Fu YX and Zhou L: Group 3 innate lymphoid cells
inhibit T-cell-mediated intestinal inflammation through aryl
hydrocarbon receptor signaling and regulation of microflora.
Immunity. 39:386–399. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Arsenescu R, Arsenescu V, Zhong J, Nasser
M, Melinte R, Dingle RW, Swanson H and de Villiers WJ: Role of the
xenobiotic receptor in inflammatory bowel disease. Inflamm Bowel
Dis. 17:1149–1162. 2011. View Article : Google Scholar :
|
|
15
|
Li Y, Innocentin S, Withers DR, Roberts
NA, Gallagher AR, Grigorieva EF, Wilhelm C and Veldhoen M:
Exogenous stimuli maintain intraepithelial lymphocytes via aryl
hydrocarbon receptor activation. Cell. 147:629–640. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jing H, Shen G, Wang G, Zhang F, Li Y, Luo
F, Yao J and Tian XF: MG132 alleviates liver injury induced by
intestinal ischemia/reperfusion in rats: involvement of the AhR and
NFκB pathways. J Surg Res. 176:63–73. 2012. View Article : Google Scholar
|
|
17
|
Dahan S, Rabinowitz KM, Martin AP, Berin
MC, Unkeless JC and Mayer L: Notch-1 signaling regulates intestinal
epithelial barrier function, through interaction with
CD4+ T cells, in mice and humans. Gastroenterology.
140:550–559. 2011. View Article : Google Scholar
|
|
18
|
Mathern DR, Laitman LE, Hovhannisyan Z,
Dunkin D, Farsio S, Malik TJ, Roda G, Chitre A, Iuga AC, Yeretssian
G, et al: Mouse and human Notch-1 regulate mucosal immune
responses. Mucosal Immunol. 7:995–1005. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen G, Sun L, Yu M, Meng D, Wang W, Yang
Y and Yang H: The Jagged-1/Notch-1/Hes-1 pathway is involved in
intestinal adaptation in a massive small bowel resection rat model.
Dig Dis Sci. 58:2478–2486. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen G, Qiu Y, Sun L, Yu M, Wang W, Xiao
W, Yang Y, Liu Y, Yang S, Teitelbaum DH, et al: The
jagged-2/notch-1/hes-1 pathway is involved in intestinal epithelium
regeneration after intestinal ischemia-reperfusion injury. PLoS
One. 8:e762742013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee JS, Cella M, McDonald KG, Garlanda C,
Kennedy GD, Nukaya M, Mantovani A, Kopan R, Bradfield CA, Newberry
RD, et al: AHR drives the development of gut ILC22 cells and
postnatal lymphoid tissues via pathways dependent on and
independent of Notch. Nat Immunol. 13:144–151. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Stevens EA, Mezrich JD and Bradfield CA:
The aryl hydrocarbon receptor: a perspective on potential roles in
the immune system. Immunology. 127:299–311. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xia M, Viera-Hutchins L, Garcia-Lloret M,
Noval Rivas M, Wise P, McGhee SA, Chatila ZK, Daher N, Sioutas C
and Chatila TA: Vehicular exhaust particles promote allergic airway
inflammation through an aryl hydrocarbon receptor-Notch signaling
cascade. J Allergy Clin Immunol. 136:441–453. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Alam MS, Maekawa Y, Kitamura A, Tanigaki
K, Yoshimoto T, Kishihara K and Yasutomo K: Notch signaling drives
IL-22 secretion in CD4+ T cells by stimulating the aryl
hydrocarbon receptor. Proc Natl Acad Sci USA. 107:5943–5948. 2010.
View Article : Google Scholar
|
|
25
|
Qiu Y, Yu M, Yang Y, Sheng H, Wang W, Sun
L, Chen G, Liu Y, Xiao W and Yang H: Disturbance of intraepithelial
lymphocytes in a murine model of acute intestinal
ischemia/reperfusion. J Mol Histol. 45:217–227. 2014. View Article : Google Scholar
|
|
26
|
Chiu CJ, McArdle AH, Brown R, Scott HJ and
Gurd FN: Intestinal mucosal lesion in low-flow states. I. A
morphological, hemodynamic, and metabolic reappraisal. Arch Surg.
101:478–483. 1970. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Polentarutti BI, Peterson AL, Sjöberg AK,
Anderberg EK, Utter LM and Ungell AL: Evaluation of viability of
excised rat intestinal segments in the Ussing chamber:
investigation of morphology, electrical parameters, and
permeability characteristics. Pharm Res. 16:446–454. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
29
|
Nebert DW and Roe AL: Role of the aromatic
hydrocarbon receptor and [Ah] gene battery in the oxidative stress
response, cell cycle control, and apoptosis. Biochem Pharmacol.
59:65–85. 2000. View Article : Google Scholar
|
|
30
|
Delescluse C, Lemaire G, de Sousa G and
Rahmani R: Is CYP1A1 induction always related to AHR signaling
pathway. Toxicology. 153:73–82. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen G, Zhang Z, Cheng Y, Xiao W, Qiu Y,
Yu M, Sun L, Wang W, Du G, Gu Y, et al: The canonical Notch
signaling was involved in the regulation of intestinal epithelial
cells apoptosis after intestinal ischemia/reperfusion injury. Int J
Mol Sci. 15:7883–7896. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huang CY, Hsiao JK, Lu YZ, Lee TC and Yu
LC: Anti-apoptotic PI3K/Akt signaling by sodium/glucose transporter
1 reduces epithelial barrier damage and bacterial translocation in
intestinal ischemia. Lab Invest. 91:294–309. 2011. View Article : Google Scholar
|
|
33
|
Jönsson ME, Franks DG, Woodin BR, Jenny
MJ, Garrick RA, Behrendt L, Hahn ME and Stegeman JJ: The tryptophan
photo-product 6-formylindolo[3,2-b]carbazole (FICZ) binds multiple
AHRs and induces multiple CYP1 genes via AHR2 in zebrafish. Chem
Biol Interact. 181:447–454. 2009. View Article : Google Scholar
|
|
34
|
Daniel RA, Cardoso VK, Góis E Jr, Parra
RS, Garcia SB, Rocha JJ and Féres O: Effect of hyperbaric oxygen
therapy on the intestinal ischemia reperfusion injury. Acta Cir
Bras. 26:463–469. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ogando J, Tardáguila M, Díaz-Alderete A,
Usategui A, Miranda-Ramos V, Martínez-Herrera DJ, de la Fuente L,
García-León MJ, Moreno MC, Escudero S, et al: Notch-regulated
miR-223 targets the aryl hydrocarbon receptor pathway and increases
cytokine production in macrophages from rheumatoid arthritis
patients. Sci Rep. 6:202232016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
González-Mariscal L, Tapia R and Chamorro
D: Crosstalk of tight junction components with signaling pathways.
Biochim Biophys Acta. 1778:729–756. 2008. View Article : Google Scholar
|
|
37
|
Pope JL, Bhat AA, Sharma A, Ahmad R,
Krishnan M, Washington MK, Beauchamp RD, Singh AB and Dhawan P:
Claudin-1 regulates intestinal epithelial homeostasis through the
modulation of Notch-signalling. Gut. 63:622–634. 2014. View Article : Google Scholar
|
|
38
|
Movahedan A, Afsharkhamseh N, Sagha HM,
Shah JR, Milani BY, Milani FY, Logothetis HD, Chan CC and Djalilian
AR: Loss of Notch1 disrupts the barrier repair in the corneal
epithelium. PLoS One. 8:e691132013. View Article : Google Scholar : PubMed/NCBI
|














