Introduction
Among all skin tumors, melanoma is the most
aggressive form because of rapid metastasis and resistance to
conventional radio- and chemotherapy (1–3).
The advances in understanding the microenvironment of melanoma and
cell biology make it obvious that the treatment needs to be
multi-directional. Melanoma is a highly immunogenic tumor (4) and numerous immunotherapeutic
strategies have been tested (5–8).
Although it has been demonstrated that the various immune-based
therapies induce an increase in circulating tumor antigen-specific
T cells, these approaches have produced a poor therapy response,
due to tumor-induced immune suppression and tumor evasion of
anti-tumor immune responses (9).
Regulatory T cells (Tregs) are CD4+
CD25+ cells characterized by the forkhead box protein 3
(FOXP3) transcription factor expression, which is the most
specific marker for Tregs (10,11). These cells exert an
immunosuppressive function and FOXP3 is a prerequisite for
this suppressive activity, ultimately leading to tumor immune
evasion/escape (12,13). Additionally, patients with an
altered expression or function of FOXP3 can develop serious
autoimmune diseases and cancers (14,15). FOXP3, a member of the
forkhead/winged-helix family of transcription factors,
constitutively translocate into the nucleus where it binds to
specific sequences of DNA to regulate the transcription of its
target genes (16,17). Although FOXP3 protein expression
was considered to be restricted to lymphocytes, recently it has
been reported to be expressed in various human malignancies, such
as pancreatic, lung, colon, breast, ovarian, prostate cancers,
hepatocellular carcinoma, and melanoma (18-28). FOXP3 has been also
associated with an unfavorable disease course (24,25,27) and identified as an independent
prognostic factor and a marker of tumor progression and metastasis
(29–33). Indeed, numerous studies have
demonstrated that metastases and poor clinical response of melanoma
are closely related to the large number of Tregs and high
FOXP3 expression (27,34–36).
Multiple signaling pathways, including NOTCH and
transforming growth factor-β (TGF-β/Smad), are closely associated
with FOXP3 transcription (37–41). NOTCH signaling regulates essential
cell processes, such as stem cell self-renewal, proliferation,
differentiation and apoptosis (42–44). Previous experimental data have
shown that aberrant NOTCH signaling may lead to cancer, although
its effect greatly depends on tissue type and interaction with
other signaling pathways (45,46). Activation of the NOTCH receptor is
triggered by its interaction with NOTCH ligands (Delta-like 1, 3,
4; Jagged-1, 2) present on adjacent cells (47). Upon ligand binding, the NOTCH
intracellular domain (NICD) is proteolytically cleaved and
translocated into the nucleus where it interacts with its
corresponding co-activators to promote the transcription of
downstream target genes (48,49). Dysregulated NOTCH signaling has
been involved in the development and progression of many types of
cancer (50–56). Findings have shown that the
upregulation of NOTCH signaling may play a role in melanoma cells
transformation and progression (50–62,33).
In addition to NOTCH, TGF-β is known as a
double-edged sword during cancer progression, being a tumor
suppressor or a tumor promoter, depending on the context of signal
activation (63–65).
TGF-β is a pleiotropic cytokine that negatively
regulates the activity of immune cells, playing an important role
in the control of T-cell functions, growth and differentiation
(66). Moreover, TGF-β signaling
is involved in Tregs differentiation being required for their in
vivo expansion and immuno suppressive capacity (67). In vitro studies have shown
that TGF-β may trigger FOXP3 expression in CD4+
CD25- naive T cells, switching them towards a
CD4+CD25+ regulatory phenotype, probably
through activation of Smads, which results in a positive
autoregulatory loop (68,69). Furthermore, all human tumors
overproduce TGF-β, whose autocrine and paracrine actions promote
tumor cell invasiveness and metastasis (70–74). TGF-β signaling can synergize with
NOTCH in many processes (75–77). Previous findings have identified
the bidirectional regulation of NOTCH and TGF-β,
through different context-dependent mechanisms and a functional
synergism in the regulation of hairy and enhancer of split 1
(HES1), a direct target of the NOTCH signal, has been
demonstrated (78–80). It has been previously shown that
the induction of FOXP3-Tregs is cooperatively regulated by NOTCH
signaling and TGF-β (76,79,81–83).
Few reports have shown the association between
FOXP3 and NOTCH in cancers (84,85) and the cross-talk between them is
unexplored in melanoma. Since TGF-β and NOTCH are involved in the
regulation of the FOXP3 gene transcription, we investigated,
in melanoma in vitro models, the mechanisms of
TGF-β1-induced FOXP3 gene expression in relation to NOTCH
signaling inactivation. For this reason, we have used a synthetic
tripeptide aldehyde containing γ-secretase inhibitor (GSI), a
pharmacological agent known to block NOTCH processing and
activation through the inhibition of proteolysis and translocation
of NIDC to the nucleus (86).
Materials and methods
Human melanoma cell lines and culture
conditions
Human epithelial melanocytes (NHEM) were purchased
from Lonza (Lonza Group, Ltd., Basel, Switzerland), cultured in
Melanocyte Growth Medium (Lonza Group, Ltd.) and used as controls.
WM35 (from primary lesion), A375 and A2058 (from metastatic lesion)
melanoma cell lines, a kind gift from V. Russo (Tumor Targeting
Research Unit, San Raffaele Scientific Institute, Milano, Italy)
were cultured in RPMI-1640 medium (Gibco; Life Technologies, Inc.,
Monza, Italy), supplemented with 2 mmol/l L-glutamine, 100 IU
penicillin, 100 μg/ml streptomycin, 10% of heat-inactivated
fetal calf serum (Gibco; Life Technologies, Inc.) and maintained
under an atmosphere of 5% CO2 at 37°C. For the western
blot analysis, reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) and immunocytochemical analysis (ICC), 70–80%
confluent cultures were used.
Immunocytochemical analysis
A total of 1×105 cells (WM35, A375 and
A2058) were grown on glass slides. Cells were washed in
phosphate-buffered saline (PBS) and fixed with 4% paraformaldehyde,
pH 7.4 for 20 min at room temperature. The cells were permeabilized
with 0.5% Triton X-100 for 4 min and after washing in PBS were
treated with 1% BSA to block non-specific binding sites. FOXP3
immunodetection was performed using a primary antibody anti-FOXP3
(1:100 dilution; ThermoFisher Scientific, eBioscience, Inc., San
Diego, USA) for 2 h at room temperature, revealed using the Immuno
Cruz Staining System (Santa Cruz Biotechnology, Inc, Santa Cruz,
CA, USA). The cells were counterstained with hematoxylin for 30
sec. Appropriate positive and negative controls were carried
out.
RNA extraction and RT-qPCR
Total RNA was extracted from WM35, A375 and A2058
cell lines stimulate or not with TGF-β1 using the Micro-to-Midi
total RNA purification system (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) according to the manufacturer's
instructions. RNA was reverse transcribed into cDNA using the
SuperScript III First-Strand Synthesis system (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions.
mRNA expression was measured using SYBR-Green
RT-qPCR using the Rotor-Gene Q thermal cycler (Qiagen, Inc.,
Valencia, CA, USA).
Amplification reactions were performed using primers
specific for FOXP3 (forward, 5′-CACAACATGCGACCCC CTTTCACC-3′ and
reverse, 5′-AGGTTGTGGCGGAT GGCGTTCTTC-3′), NOTCH1 and HES1
(QuantiTect® Primer Assay; Qiagen, Inc.). The PCR
reaction was carried out in 25 μl buffer, containing 50 ng
cDNA, 1 μM of each primer and 12.5 μl 2X RotorGene
SYBR-Green PCR Master Mix (Qiagen, Inc.). The thermal cycling
conditions were as follows: denaturation at 95°C for 5 min,
followed by 40 cycles of denaturation for 10 sec at 95°C and
annealing and extension for 15 sec at 60°C. As housekeeping gene,
glyceraldehyde 3-phosphate dehydrogenase (GAPDH; QuantiTect
Primer assay; Qiagen, Inc.) was used. Transcripts quantification
was carried out utilizing the software supplied with Rotor-Gene Q.
The experiments were repeated three times.
Protein extraction and western blot
analysis
Cells were lysed in RIPA buffer (Thermo Fisher
Scientific, Inc.) in the presence of 1 mM of the protease inhibitor
phenylmethylsulfonyl fluoride (PMSF; Thermo Fisher Scientific,
Inc.) and incubated on ice for 30 min. Protein concentration was
determined by Bradford assay, using the Quick Start Bradford kit
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). Total proteins
were fractionated using SDS-PAGE (Invitrogen; Thermo Fisher
Scientific, Inc.) and transferred onto nitrocellulose membranes
(Trans-Blot Transfer Medium Pure Nitrocellulose Membrane 0.45
μm; Bio-Rad Laboratories, Inc.). Membranes were blocked with
5% non-fat dried milk in TBS buffer containing (20 mM Tris, 500 mM
NaCl, and 0.05% Tween-20) and incubated overnight with the
appropriate primary monoclonal antibody: Anti-NOTCH1-NICD (dilution
1:1,000; cat. no. 14-5785-81; ThermoFisher Scientific, eBioscience,
Inc.), anti-FOXP3 (dilution 1:1,000; cat. no. 14-5773-80;
ThermoFisher Scientific, eBioscience, Inc.), anti-HES1 (dilution
1:500; cat. no. AB5702 ; EMD Millipore, Billerica, MA, USA),
anti-Smad3 and phospho-Smad3 (dilution 1:1,000; cat. nos. 9513S and
9520S, respectively; Cell Signaling Technology, Inc., Danvers, MA,
USA). Horseradish peroxidase-conjugated anti-rabbit or anti-goat
IgG was used as the secondary antibody and the protein bands were
detected using the enhanced chemiluminescence detection system (ECL
detection system; Bio-Rad Laboratories, Inc.). Protein levels were
determined using laser densitometry and normalized to GAPDH
(Calbiochem; Merck KGaA, Darmstadt, Germany) levels in each
sample.
GSI and TGF-β treatment
A synthetic tripeptide aldehyde inhibitor, GSI
(Calbiochem; Merck KGaA), a potent GSI, was used to block
NOTCH1-mediated signal transduction in melanoma cell lines. Cells
in the logarithmic growth phase were seeded at densities of
4×105 cells/ml (WM35) and 2×105 cells/ml
(A375 and A2058) and treated or not with GSI, at different
concentrations (5, 10 and 20 μM) for the desired period of
time. The cells were then stimulated with recombinant human
(rh)TGF-β (5 ng/ml; Gibco; Life Technologies, Inc.) for 48 h.
Control cells were treated with an equal volume of dimethyl
sulfoxide (DMSO). Expression of FOXP3 and NOTCH signaling, cell
growth proliferation and inhibition of melanoma cell lines were
analyzed.
Cell proliferation-cytotoxicity
assay
GSI effects on cell proliferation was measured using
the [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide
(MTT); Sigma-Aldrich, St. Louis, MO, USA] colorimetric assay as
described by Cardile et al (87). In brief, melanoma cell lines
(1–2×104 cells/well) were grown overnight in 24-well
plates and then treated with vehicle alone or different
concentrations of GSI. After 24, 48 and 72 h treatment, the cells
were incubated with 20 μl of 0.5% MTT in PBS for 4 h at 37°C
in a humidified 95% air/5% CO2; supernatant was removed
and 100 μl of DMSO was added to each well. Optical density
was measured at 550 nm (Titertek Multiskan; DAS). Cell viability
was expressed as a percentage of treated cells with respect to
appropriate controls. Trypan blue dye exclusion assay was used to
evaluate the percentage of dead cells with respect to the total
number of cells.
Statistical analysis
Differences between TGF-β1 stimulated or not were
compared by Student's t-test. Differences among multiple groups
were compared by analysis of variance (ANOVA test) and a post hoc
test for multiple comparisons (Tukey's test). Data are presented as
mean ± SD. P-values <0.05 were considered statistically
significant.
Results
Upregulation of FOXP3 expression on human
melanoma cell lines by TGF-β1
We used different approaches to examine the baseline
gene expression of FOXP3 as well as the protein levels, in
human melanoma cell lines at different stages, primary (WM35)
versus metastatic (A375 and A2058) cells. Normal human epidermal
melanocytes (NHEM) were used as a control.
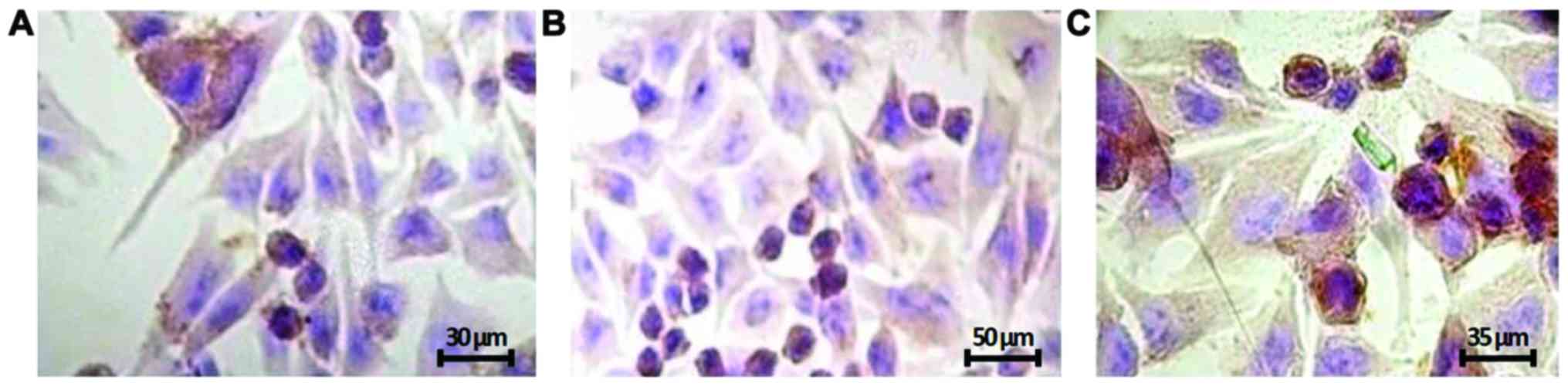
First, we observed by immunocytochemistry that
FOXP3 was mainly localized in the nucleus and, was less
evident, in the perinuclear region and cytoplasm. Moreover, the
three melanoma cell lines showed different staining intensity,
being the A2058 cell lines that were mostly expressed (Fig. 1), while NHEM FOXP3 staining
was undetectable (data not shown).
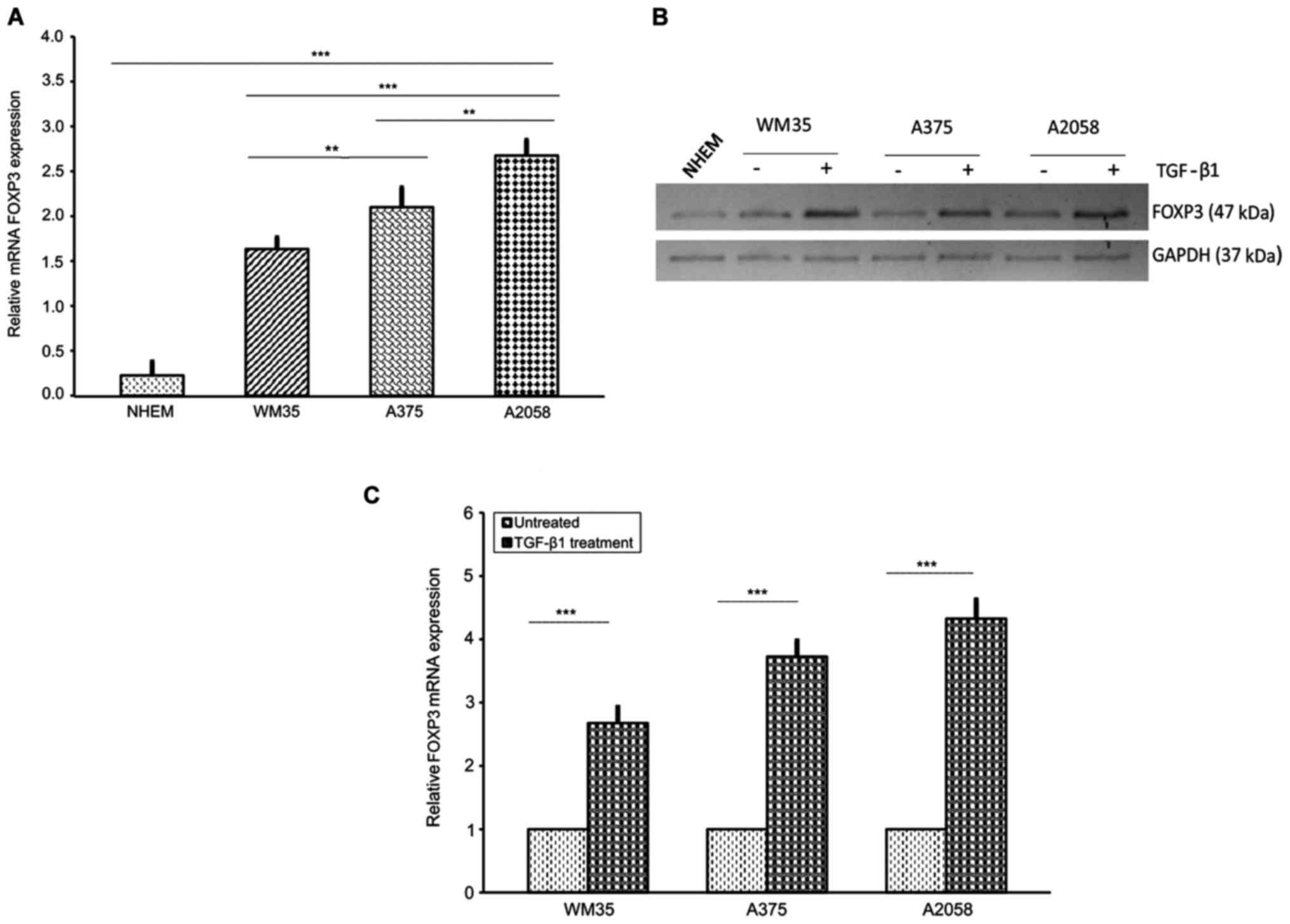
Next, we examined the relative mRNA level of
FOXP3 in WM35, A375 and A2058 melanoma cells. As expected,
FOXP3 transcriptional levels, assessed by RT-qPCR, were
higher in A2058 cells compared to A375 and WM35 cells, and very low
in NHEM (Fig. 2A). In accordance
with the RT-qPCR data, western blot analysis revealed that FOXP3
protein level expression was higher in A2058 compared to A375 and
WM35 cells, whereas NHEM cells had a very weak expression (Fig. 2B). Moreover, since TGF-β1
regulates T-cell function through FOXP3 (69), we examined the effect of this
cytokine on FOXP3 expression in melanoma cells. Stimulation
with rhTGF-β1 (5 ng/ml) for 48 h significantly increased the mRNA
expression of FOXP3, approximately of 1.68-, 2.74- and
3.3-fold in WM35, A375 and A2058 cells, respectively, compared to
untreated cells (Fig. 2C).
Shorter treatments did not induce any appreciable change in FOXP3
expression. Western blot analysis confirmed the upregulation of
TGF-β1-induced FOXP3 protein levels (Fig. 2B). Altogether, our results show a
very high expression of the transcription factor FOXP3 in human
metastatic melanoma cells, suggesting that FOXP3 could be
considered a biological marker of melanoma progression, probably
contributing to metastasis, as described by other authors (29,34–36).
Modulation of FOXP3 expression through
the NOTCH signaling pathway
It has been shown that NOTCH signaling may be
involved in the activation of FOXP3 promoter (39,85). To analyze the potential role of
NOTCH in modulating the FOXP3 expression in melanoma cells,
we used the GSI, a GSI, responsible for inhibition of NOTCH
cleavage into the active NICD (47,86).
First, we tested, by RT-qPCR, the efficacy of GSI in
FOXP3 expression reduction. WM35, A375 and A2058 cells were
treated with DMSO, as a control, or increasing concentrations of
GSI (5, 10 and 20 μM) for 24, 48 and 72 h. FOXP3 mRNA
expression decreased in a concentration- and time-dependent manner
in all cell lines (Fig. 3A and
B). GSI in low doses (5, 10 μM) for 24 and 48 h did not
show any significant modification of FOXP3 gene expression
(data not shown). At 72 h (Fig.
3A), we found a FOXP3 mRNA reduction of 40.7, 52.9%;
40.1, 62.2 and 53%, 62.6% in WM35, A375 and A2058 cells treated
with 20 μM GSI compared to low doses (5, 10 μM),
respectively. Furthermore, at 20 μM GSI, a modest reduction
of FOXP3 mRNA was observed in the three melanoma cell lines
at 24 and 48 h, while a strong and significant reduction was
observed at 72 h (Fig. 3B). Thus,
20 μM GSI for 72 h was the proper concentration and time for
treating melanoma cells. We also verified whether FOXP3 was
regulated at the translation level. In agreement with the RT-qPCR
data, we observed a decrease of FOXP3 protein levels, in a
dose-dependent manner, in the three melanoma cell lines after 72 h
of GSI treatment. Fig. 3C shows
the protein levels in the WM35 cell line treated with 5, 10 and 20
μM GSI at 72 h.
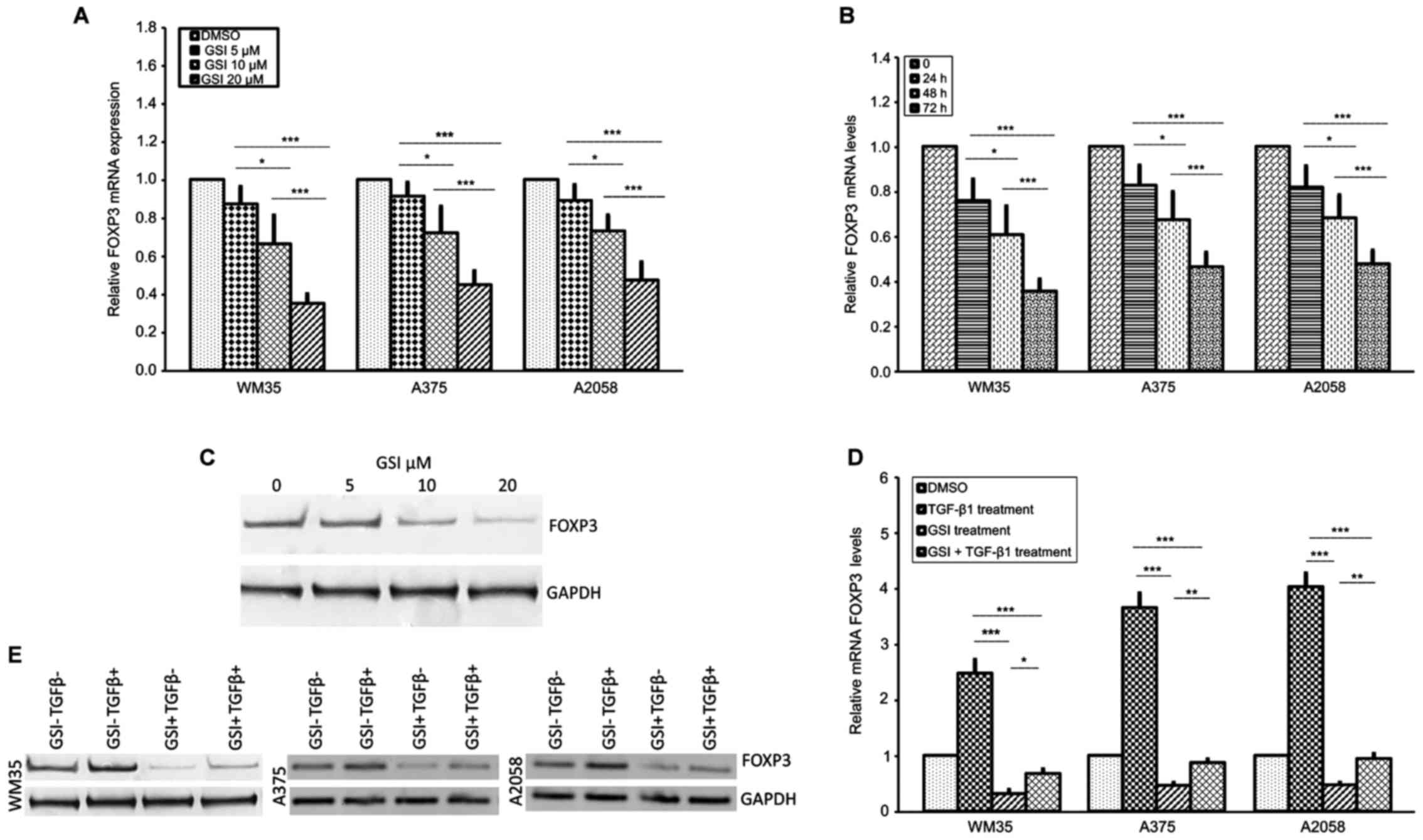 | Figure 3Effect of GSI on FOXP3 expression in
melanoma cell lines. (A) Inhibition of FOXP3 mRNA is shown after 72
h of 5-, 10- and 20 μM GSI treatment in melanoma cells.
RT-qPCR shows that FOXP3 mRNA levels were downregulated in
GSI-treated WM35, A375 and A2058 melanoma cells in a dose-dependent
manner. Maximum inhibition of FOXP3 was observed at 20 μM of
GSI. (B) Inhibition of FOXP3 mRNA after 24, 48 and 72 h with 20
μM/GSI treatment in WM35, A375 and A2058 melanoma cell
lines. A statistically significant time-dependent decrease in FOXP3
mRNA level was observed in each melanoma cell line. (C) Inhibition
of FOXP3 protein expression after 72 h of 5-, 10- and 20 μM
GSI treatment in WM35 melanoma cells. Western blot analysis showed
that the protein levels of FOXP3 were downregulated in GSI-treated
WM35 cells in a dose-dependent manner. GAPDH expression was used as
a loading control. (D and E) Effect of GSI/TGF-β1 treatment on
FOXP3 mRNA and protein expression in melanoma cell lines.
Inhibition of FOXP3 mRNA and protein levels are shown after 72 h of
GSI treatment in WM35, A375 and A2058 melanoma cells. In
vitro GSI treatment downregulated TGF-β1-induced FOXP3 mRNA and
protein levels in all the melanoma cell lines. As an internal
control, GAPDH was used for normalization. Data are shown as
mean ± SD of three independent experiments. The comparison of mRNA
FOXP3 expression in multiple groups was performed by ANOVA and
Tukey's test. GSI, γ-secretase inhibitor; FOXP3, forkhead box
protein 3; TGF-β, transforming growth factor-β; GAPDH,
glyceraldehyde 3-phosphate dehydrogenase. *P<0.01;
**P<0.001; ***P<0.0001. |
To validate the possible relationship between
NOTCH1/TGF-β1 in regulating FOXP3 expression, we also
examined whether GSI, a NOTCH signaling inhibitor, influenced the
TGF-β1-dependent FOXP3 upregulation. WM35, A375 and A2058
cell lines were pre-treated overnight with 20 μM GSI, to
block NOTCH activation, using an equal volume of DMSO as control,
and the cells were subsequently stimulated with rhTGF-β1 for 48 h.
Fig. 3D shows that 20 μM GSI
significantly decreased FOXP3 mRNA levels in WM35, A375 and A2058
cell lines treated with TGF-β1, although the downregulation did not
reach the levels observed in cells treated with GSI alone. Data
were confirmed by western blot analysis (Fig. 3E).
Taken together, these data indicate that
upregulation of FOXP3 by TGF-β1 may require input from the
NOTCH signaling pathway.
Activation of NOTCH1 pathway by TGF-β1 in
human melanoma cell lines
Since TGF-β1 upregulates FOXP3 levels and in
considering that NOTCH/TGF-β signaling is involved in the
tumorigenic process of cancers (50,65,88), including melanoma (62,89,90), we investigated whether TGF-β1 was
able to affect the NOTCH signaling in melanoma cell lines. We
tested NOTCH1NICD and the NOTCH-specific target gene
HES1 expression in WM35, A375, A2058 and NHEM cells by
RT-qPCR and western blot analysis. Generally, NICD levels reflect
the activation status of NOTCH signaling.
Our results showed that NOTCH1NICD and
HES1 expression were significantly increased in the three
melanoma cell lines compared to NHEM cells at both mRNA and protein
levels (Fig. 4A and D). This
increase was more evident in WM35 compared to A375 and A2058 cells.
Next, to confirm the association between TGF-β1 and NOTCH1, we
stimulated WM35, A375 and A2058 cells with rhTGF-β1 (5 ng/ml) for
48 h. We found higher mRNA and protein levels of
NOTCH1NICD and HES1, in WM35, A375 and A2058 cells,
after stimulation with rhTGF-β1, compared to untreated cells
(Fig. 4B–D), confirming earlier
studies carried out in other cancer types (78,79,88,91).
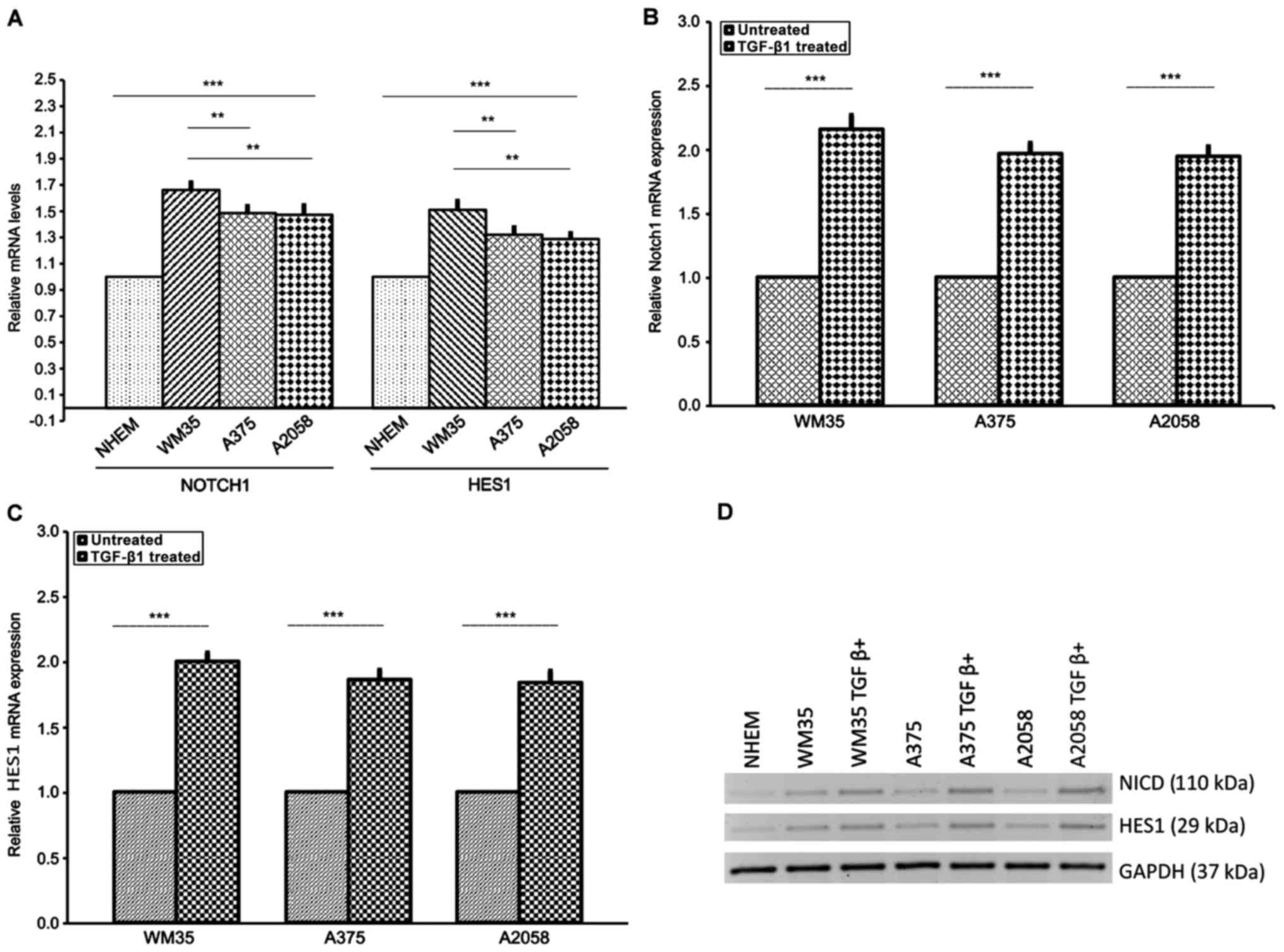 | Figure 4Expression of NOTCH1NICD
and NOTCH-specific target gene HES1 in melanoma cell lines.
(A) mRNA of NOTCH1NICD and HES1 was measured by RT-qPCR
in NHEM, WM35 and A375 and A2058 melanoma cells. Melanocytes served
as the control. WM35 showed a higher level of NOTCH1NICD
mRNA and HES1 mRNA than A375 and A2058 cells. (B) Protein level of
NOTCH1NICD and HES1 was measured by western blot
analysis in WM35, A375 and A2058 melanoma cell lines. All of the
melanoma cell lines positively expressed NOTCH1NICD and
HES1. (B-D) Effect of TGFβ-1 treatment on NOTCH1NICD,
HES1 mRNA and protein levels in melanoma cell lines. Treatment with
rhTGF-β1 (5 ng/ml) for 48 h induced a higher increase of
NOTCH1NICD and HES1 mRNA and their own protein levels in
WM35, A375 and A2058 melanoma cells. As an internal control, GAPDH
was used for normalization. Data are shown as mean ± SD of three
independent experiments. The comparison of mRNA NOTCH-1 and HES1
expression in multiple groups was performed by ANOVA and Tukey's
test. HES1, hairy and enhancer of split 1; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; TGF-β,
transforming growth factor-β; rh, recombinant human; GAPDH,
glyceraldehyde 3-phosphate dehydrogenase. **P<0.001;
***P<0.0001. |
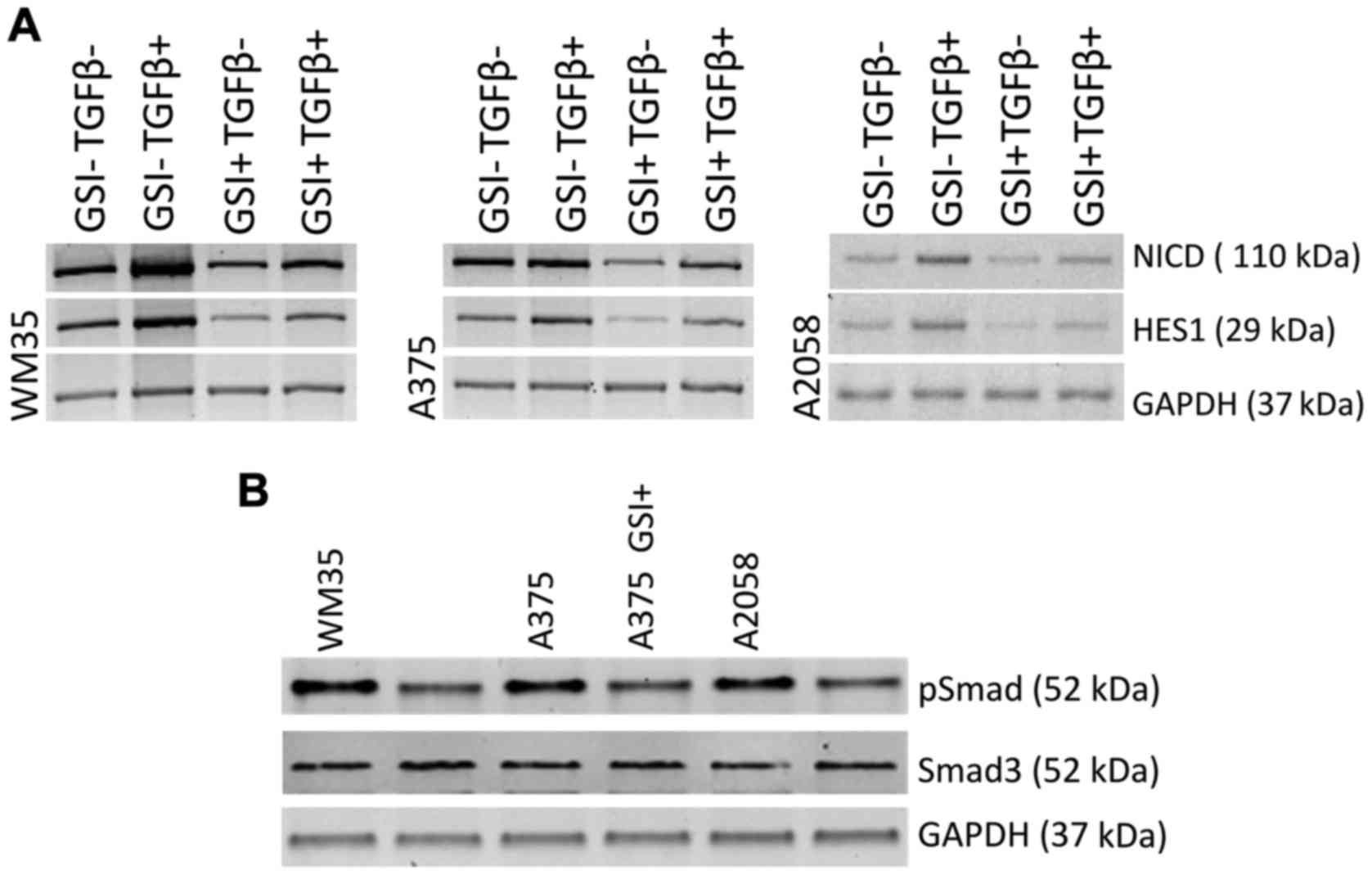
GSI prevents TGF-β1-dependent NOTCH1
activation pathway
In parallel, we explored whether NOTCH signaling
inhibition could prejudice NOTCH/TGF-β axis. Western blot analysis
revealed that NOTCH-1 protein and its downstream effector
HES1 were downregulated in GSI-treated melanoma cells
compared with vehicle control (Fig.
5A), confirming the ability of the drug to affect the NOTCH
signaling pathway (61).
Furthermore, to corroborate the effects of the NOTCH inhibition on
TGF-β1-induced NOTCH1NICD activation, WM35, A375 and
A2058 cell lines were treated with TGF-β1 alone or combined with
GSI. Notably, we found that the upregulation of TGF-β1-induced of
the NOTCH1NICD protein level was strongly decreased but
not completely abolished by concomitant GSI treatment in WM35, A375
and A2058 cells. The same trend was evident for downstream target
genes HES1 (Fig. 5A).
Finally, to confirm the effect of NOTCH on
TGF-β/Smad signaling, we treated WM35, A375 and A2058 cells with
and without GSI for 72 h. Phosphorylated Smad3 (pSMAD3), a marker
of constitutive TGF-β1 receptor activity, was detected in WM35,
A375 and A2058 cells; GSI treatment consistently decreased pSMAD3
levels in all melanoma cell lines (Fig. 5B), without interfere with the
unphosphorylated Smad3 protein levels. These data highlighted the
effect of NOTCH on TGFβ/Smad signaling in melanoma cell lines.
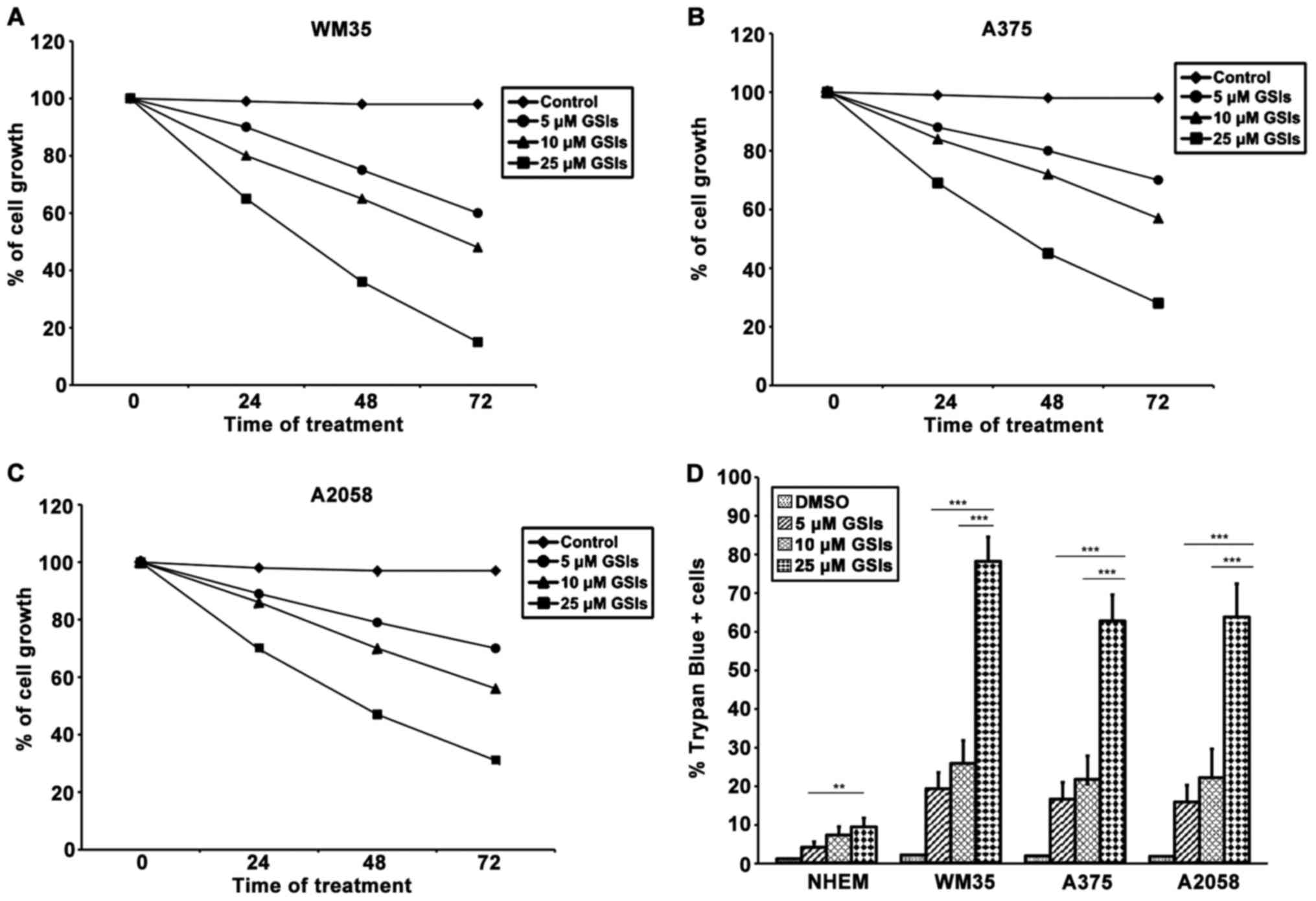
Cytotoxic and anti-proliferative effects
of GSI on melanoma cells
To further investigate whether GSI could be an
effective therapeutic target for melanoma, we first tested the
effect of the drug on cell proliferation in WM35, A375 and A2058
after treatment to 0, 5, 10 and 20 μM of GSI for 24, 48 and
72 h. MTT analyses showed that melanoma cell lines, without GSI
treatment, exhibited a linear growth up to confluence. In these
experiments, DMSO, which was used as a vehicle control, did not
affect cell growth, while GSI treatment induced a marked cell
growth inhibition, in a dose- and time-dependent manner. The
concentrations of 5 and 10 μM showed a weak growth
inhibition, but at 20 μM, GSI significantly inhibited
proliferation at each incubation time for all the cell lines, the
maximum inhibition was attained at 72 h (Fig. 6A–C). However, GSI effects were
significantly greater in WM35 cells rather than in A375 and A2058;
no difference was noted between A375 and A2058 cells. MTT assay did
not discriminate if the decrease of proliferation rate was
attributed to growth arrest or cell death, since both mechanisms
induce a decrease in cell numbers and an apparent loss of
viability. To establish this, a Trypan blue exclusion test was
performed on melanoma cell lines. As shown in Fig. 6D, GSI exerted a significantly more
toxic effect on the melanoma cells compared to DMSO. The
cytotoxicity of GSI on WM35 cells was significantly higher than
that of A375 and A2058 cells. GSI showed very low toxicity in
normal melanocytes.
Discussion
Many studies have shown that FOXP3 is
expressed not only by Tregs, but also in a variety of tumor cells,
including melanoma (18–20,25,27,28,33). Expression of FOXP3 by
cancer cells may cause the inhibition of tumor directed T-cell
responses and may favor tumor cells immune-evasion mechanisms
(22,23,27,30,31,33). By contrast, other studies have
suggested that FOXP3 plays a critical role in suppressing the
development of several types of tumors, such as ovarian, prostate
and breast cancer, through the inhibition of cell proliferation,
migration and invasion or by modulating the expression of oncogenes
or tumor suppressor genes (29,91–93). Thus, the FOXP3 gene or
protein exerts different functions in different types of tumors. In
any case, the role of FOXP3 in carcinogenesis is intriguing and
remain controversial.
Multiple signaling pathways, including NOTCH and
TGF-β/Smad pathways, are involved in FOXP3 transcription regulation
(17,37,38,41,76,83,95). Few studies have shown the
association between NOTCH/FOXP3 in cancers (84,85) and to the best of our knowledge
there are no reports investigating directly the relationship
between NOTCH/TGF-β signaling and FOXP3 transcription factor in
melanoma.
In the present study, we investigated the
involvement of NOTCH/TGF-β1 signaling pathways in regulating the
FOXP3 transcription factor and demonstrated, for the first time,
that FOXP3 expression was modulated by NOTCH/TGF-β1 pathways in
primary and metastatic melanoma cell lines. The subcellular
localization of FOXP3 in human melanoma cell lines at
different stages of cancer progression was studied by
immunocytochemistry. Our results showed that the intensity of
FOXP3 expression in melanoma metastatic cells lines (A375
and A2058) was higher than that in primary melanoma cells (WM35),
while FOXP3 staining was undetectable in melanocytes. These results
underline that FOXP3 staining gradually increase from the primary
to the metastatic melanoma cell lines.
This result suggested that FOXP3 expression may be
associated with metastatic spread. These data partially confirm the
study by Quaglino et al (32), which demonstrated a significant
association between FOXP3 expression in primary melanomas and
development of visceral metastases. Our study also showed an
heterogeneous subcellular localization of FOXP3 mainly in the
nucleus, less in cytoplasm and in perinuclear region. Metastatic
melanoma cell lines exhibited strong FOXP3 positive staining in the
nucleus and weak staining in the cytoplasm. Similar to our results,
Brody et al (96) reported
a nuclear FOXP3 expression in Tregs localized at the primary
melanomas and at the interface of metastasis with the lymph node
parenchyma. Chen et al (97) showed that activation of
CD4+ CD25+ Treg induced a shift in the
subcellular localization of FOXP3 from a primarily
cytoplasmic/perinuclear pattern, in most cells, to a nuclear
pattern, suggesting that the change in the FOXP3 expression pattern
may be a result of post-translational modifications. Similar
results were obtained by Niu et al (27) and subcellular staining of FOXP3
was demonstrated in other types of cancer (19–21,24) due to post-translational
modification and types of cancer (97). However, the exact involvement of
this variable expression of FOXP3 remains unclear.
Subsequently, we confirmed in vitro, by
RT-qPCR and western blot analysis, FOXP3 expression in
melanoma cell lines. Our study has revealed that FOXP3 was
strongly expressed in metastatic melanoma cell lines. A higher
FOXP3 at mRNA and protein levels was more evident in the
metastatic melanoma cell line A2058, compared to A375, cell lines
derived from the dermis of a malignant melanoma. In addition,
FOXP3 expression was slightly lower in the primary melanoma
cells (WM35), but was still significantly higher than that in the
melanocytes.
This result suggests that FOXP3 is a
biological marker of melanoma progression and may contribute to
metastasis. These results are particularly noteworthy and confirm
those reported by other studies that associated high FOXP3
levels with metastasis in several tumors, including melanoma
(22,31,33,98,99).
Since the potential role of FOXP3 has been
demonstrated in various cancer types on immune surveillance
(20), we examined the effects of
TGF-β1 on the induction of FOXP3 in melanoma cell lines. We showed
that TGF-β1 treatment upregulated FOXP3 expression at the
transcriptional and post-translational level, more in A2058 and
A373 cells than in WM35 cells.
Overall, our data strongly emphasize the role of
TGF-β1 and FOXP3 in promoting melanoma progression.
Melanoma is a type of highly immunogenic cancer and
is a rich source of TGF-β (65).
It is possible that TGF-β1, one of the many factors present in the
tumor microenvironment, can induce FOXP3 and the regulatory
activity in Treg cells (27,28,68,69,100). FOXP3-expressing melanoma cells
may have Treg-like activity, thus suppressing effector T-cell
activity (28,34). It is possible that FOXP3
immunosuppressive function in the FOXP3-driven metastatic process
requires a crosstalk between tumor cells and the micro-environment
(34).
It has been demonstrated that, for the majority of
the Treg cells, FOXP3 expression is transient and its
persistence is highly dependent on the TGF-β exposure present in
tumoral microenvironment (101).
The identification of regulatory mechanisms that
potentially lead to a decreased expression of FOXP3 may
offer insight into the control of tumor cell proliferation and
progression in melanoma and provide new perspectives to develop
potential therapeutic targets. Although numerous cell surface
molecules could mediate this condition, we focused on NOTCH
signaling because it was shown that NOTCH may be involved in the
activation of FOXP3 promoter through RBP-J- and
HES1-dependent mechanisms (39).
In addition, emerging evidence indicated that TGF-β1 and NOTCH act
in concert to regulate the transcription of target genes (102–104). For example, TGF-β1, through
effector Smad3, and NOTCHNICD physically interact to
coordinately regulate the transcription of Hes1 and
FOXP3 (76–79). Further findings have shown that
the NOTCH ligand Jagged2 promotes Treg cell proliferation, leading
to an increase in TGF-β production (105).
NOTCH/TGF-β1 pathways are important regulators of
many fundamental processes of cancer cell biology, such as tumor
growth, angiogenesis, invasion and tumor progression (72,78,79,80,88,91,106,107).
Aberrant expression of TGF-β1 and NOTCH pathway has
been demonstrated previously in melanoma (57,61,74,90,108).
In accordance with the above reported studies, we
found that both the NOTCHNICD and Hes1 mRNA and protein
levels were higher in primary melanoma (WM35) compared to
metastatic melanoma cells (A375 and A2058). Moreover, TGF-β1
treatment induced upregulation of NOTCHNICD and Hes1 in
all the melanoma cell lines. The increase of Hes1 by TGF-β1 is
consistent with earlier studies, one of which demonstrated a
cooperative interaction between Smad3 and NOTCH on CSL binding
elements (79,91,109). Hes1 is the most well
characterized target gene of NOTCH, and its upregulated expression
symbolizes the activated NOTCH signaling.
Consistent with our data, other reports have found
that NOTCH signaling are upregulated in primary lesions of human
malignant melanoma (61,62,89).
This suggests that NOTCH signaling may be involved
in melanomagenesis, by activating MAPK/PI3K/AKT signaling pathways,
and corroborate the idea that it has less effect on metastatic cell
lines, suggesting its primary function in early transformation
events (57,60,110). In contrast to oncogenetic role
in melanoma, NOTCH pathway is downregulated in other cancers,
including skin carcinoma (111–113). A recent study of Banerjee et
al (114) showed that
inhibition of NOTCH causes pathologic activation of liver stromal
cells, promoting angiogenesis and growth of hepatic metastases. In
a later study, Talora et al (115) showed that in HPV-positive
cervical cancer cells, activated NOTCH causes growth suppression.
The involvement of NOTCH in cancer development is complex, since
NOTCH can function as an oncogene or a tumor suppressor depending
on the tissue type, different cell context and on crosstalk with
other signaling pathways (45,46). Previous findings have shown that
NOTCH signaling is largely regulated by γ-secretase activity,
responsible for cleavage of NOTCH into its active intracellular
transactivator NICD and for its translocation to the nucleus, where
it induces the transcription of target genes (47,86). Recently, there has been an
increased interest in targeting the NOTCH pathway using GSIs as a
new target therapy for those tumors with NOTCH activation (86,116,117). Thus, when we treated the
melanoma cell lines with the GSI, a strong effect on both growth
inhibition and cellular death in WM35, A375 and A2058 cells was
evident.
We have observed that low GSI doses did not induce
cell growth inhibition or cell death, as shown by MTT assay and dye
test. Instead, a high dose of GSI, led to inhibition of the
proliferation rate, with concomitant induction of death of WM35,
A375 and A2058 cells due to a strong NOTCH inhibition. Thus, the
level of treatment operating on NOTCH signaling appears to be
critical for the proliferation outcome.
The role of GSI in inhibiting the growth of
melanoma cell lines is consistent with the previous findings in
other cancers (110,118–120). Notably, some research has shown
an opposite outcome; the overexpression of NOTCH signaling can
inhibit the growth of cancer cells through induction of cell cycle
arrest (121,122).
Our results underline the role of NOTCH as an
oncogene in melanoma because its downregulation causes inhibition
of cell growth and induction of cellular death in all the melanoma
cell lines.
Based on the data presented herein and in
consideration that FOXP3 signaling may function as a potential
oncogenic factor in melanoma (28,32,33,99,123), we hypothesized that the
pharmacological inhibition of NOTCH by GSI, could reduce the
tumorigenic activity that NOTCH exerts through several signaling
pathways in melanoma cells, such as FOXP3 pathways and TGF-β/Smad3
signaling.
Of note, we have found that GSI treatment strongly
decreased FOXP3 expression at the transcriptional and
translational level in WM35, A375 and A2058 cells in a dose- and
time-dependent manner. Consistent with our results, prior studies
have shown that blockade of the NOTCH1 inhibited FOXP3
expression and Treg suppressor function (76,82,85,124). This finding emphasizes the role
of NOTCH signaling in Treg differentiation and FOXP3
transcription. In addition, we found that GSI reduced the
upregulation of TGF-β1-mediated FOXP3 gene and protein in
primary and metastatic melanoma cells. In this regard, it is
interesting to note that various reports have shown cell-type
specific effect of TGF-β1, as a mediator of FOXP3 and NOTCH
signaling pathways (76–78). In other cases, however, the
NOTCHNICD signaling blocks TGF-β1 signaling by mutually
interfering with the Smad3 (125–127). This emphasizes the complexity of
the interaction between FOXP3 and NOTCH/TGF-β signaling that may
produce different signaling outcomes depending on other signaling
pathways.
This study further confirms the functional
integration between NOTCH and TGF-β1 signaling pathways and
underlines the synergistic effect of NOTCH on a subset of
Smad3-inducible genes. We found that GSI markedly induced both a
decreased level of NOTCHNICD protein, that of its
downstream gene Hes1, and attenuated strongly the levels of
TGF-β1-induced NOTCHNICD and Hes1 protein in WM35, A375
and A2058 melanoma cell lines. In addition, we detected that the
downregulation of NOTCHNICD by GSI decreased the pSmad3
protein, a downstream transcription factor of TGF-β1.
NOTCHNICD, not only interacts with pSmad3, facilitating
its nuclear translocation (91,128), but also remains bound with
pSmad3 in the nucleus where they cooperatively upregulate the
transcription factor FOXP3 (76,
81,129). Tone et al (130), have demonstrated that the
mechanism underlying TGF-β-driven of FOXP3 expression
involves the induction of activated Smad3 (pSmad3), which acts as a
powerful transcription factor for the FOXP3 gene.
Taken together, our results sustain the role of
NOTCH signaling in mediating the FOXP3 expression in
melanoma cells by a dual mechanism: direct modulation of
FOXP3 transcription and cooperative interaction with the
TGF-β1 pathway in the modulation of FOXP3 expression. Our
data suggest a possible crosstalk between NOTCH1/TGF-β1 and
FOXP3 pathways in melanoma cells.
Moreover, our data show that NOTCHNICD
activation has an effect on TGF-β/Smad signaling and confirm that
the NOTCH and TGF-β1 pathways are intertwined to regulate FOXP3
transcription factor in melanoma cell lines. Nevertheless, future
studies are needed to validate our data. In vivo experiments
are required to explore the role of NOTCH/TGF-β pathway in the
regulation of FOXP3 transcription factor in melanoma.
Finally, this study may provide a double additional
rationale for targeting the NOTCH and FOXP3 signaling pathways for
treatment of melanoma.
Despite various advances on the comprehension of
the signal transduction pathways that modulate FOXP3
transcriptional activity, there are still many uncertainties.
A growing body of evidence suggests a connection
between FOXP3 and NOTCH/TGF-β signaling pathways and their link
with cancer recurrence, metastasis, and patient prognosis (76,85). Taken together these studies
highlight the need of a more detailed understanding of how the
NOTCH/TGF-β/FOXP3 signals interact with other pathways in order to
design rationally oriented targeted therapy experiments and
trials.
The problem of secondary resistance to targeted
therapy is a common problem in oncology. Thus, the use of drugs
specifically targeting NOTCH, such as GSI, combined with other
drugs, either standard chemotherapeutic agents or selective
pathway-specific inhibitors, such as TGF-β1 or Braf inhibitors,
could offer a potential strategy for therapeutic investigations in
melanoma. Therefore, our study not only corroborated some of these
findings, but also identified a novel interaction between NOTCH and
TGF-β1 in modulating FOXP3 expression in melanoma cells.
Although further studies are needed to clarify the role and
molecular mechanisms that govern the association between FOXP3 and
NOTCH/TGF-β signaling in the progression of melanoma, the current
study provides new insight into the carcinogenesis of melanoma.
Acknowledgments
Not applicable.
References
|
1
|
Buzaid AC: Management of metastatic
cutaneous melanoma. Oncology (Williston Park). 18:1443–1450;
discussion 1457–1459. 2004.
|
|
2
|
La Porta CA: Mechanism of drug sensitivity
and resistance in melanoma. Curr Cancer Drug Targets. 9:391–397.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fava P, Astrua C, Chiarugi A, Crocetti E,
Pimpinelli N, Fargnoli MC, Maurichi A, Rubegni P, Manganoni AM,
Bottoni U, et al: Differences in clinicopathological features and
distribution of risk factors in Italian melanoma patients.
Dermatology. 230:256–262. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Maio M: Melanoma as a model tumour for
immune-oncology. Ann Oncol. 23(Suppl 8): viii10–14. 2012.
View Article : Google Scholar
|
|
5
|
Shrayer DP, Bogaars H, Wolf SF, Hearing VJ
and Wanebo HJ: A new mouse model of experimental melanoma for
vaccine and lymphokine therapy. Int J Oncol. 13:361–374.
1998.PubMed/NCBI
|
|
6
|
Nakai N, Katoh N, Kitagawa T, Ueda E,
Takenaka H and Kishimoto S: Immunoregulatory T cells in the
peripheral blood of melanoma patients treated with melanoma
antigen-pulsed mature monocyte-derived dendritic cell vaccination.
J Dermatol Sci. 54:31–37. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Russo A, Ficili B, Candido S, Pezzino FM,
Guarneri C, Biondi A, Travali S, McCubrey JA, Spandidos DA and
Libra M: Emerging targeted therapies for melanoma treatment
(Review). Int J Oncol. 45:516–524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nakamura K and Okuyama R: Immunotherapy
for advanced melanoma: Current knowledge and future directions. J
Dermatol Sci. 83:87–94. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Slingluff CL Jr, Chianese-Bullock KA,
Bullock TN, Grosh WW, Mullins DW, Nichols L, Olson W, Petroni G,
Smolkin M and Engelhard VH: Immunity to melanoma antigens: From
self-tolerance to immunotherapy. Adv Immunol. 90:243–295. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hori S, Nomura T and Sakaguchi S: Control
of regulatory T cell development by the transcription factor Foxp3.
Science. 299:1057–1061. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ramsdell F: Foxp3 and natural regulatory T
cells: Key to a cell lineage. Immunity. 19:165–168. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sakaguchi S: Naturally arising
CD4+ regulatory t cells for immunologic self-tolerance
and negative control of immune responses. Annu Rev Immunol.
22:531–562. 2004. View Article : Google Scholar
|
|
13
|
Takeuchi Y and Nishikawa H: Roles of
regulatory T cells in cancer immunity. Int Immunol. 28:401–409.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu R, Li S, Yang WH and Wang L: IPEX
syndrome, FOXP3 and cancer. J Syndr. 1:72013.PubMed/NCBI
|
|
15
|
Martin F, Ladoire S, Mignot G, Apetoh L
and Ghiringhelli F: Human FOXP3 and cancer. Oncogene. 29:4121–4129.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Coffer PJ and Burgering BM: Forkhead-box
transcription factors and their role in the immune system. Nat Rev
Immunol. 4:889–899. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shen Z, Chen L, Hao F and Wu J:
Transcriptional regulation of Foxp3 gene: Multiple signal pathways
on the road. Med Res Rev. 29:742–766. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu H: FOXP3 expression and prognosis: Role
of both the tumor and T cells. J Clin Oncol. 27:1735–1736. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hinz S, Pagerols-Raluy L, Oberg HH,
Ammerpohl O, Grüssel S, Sipos B, Grützmann R, Pilarsky C,
Ungefroren H, Saeger HD, et al: Foxp3 expression in pancreatic
carcinoma cells as a novel mechanism of immune evasion in cancer.
Cancer Res. 67:8344–8350. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Karanikas V, Speletas M, Zamanakou M,
Kalala F, Loules G, Kerenidi T, Barda AK, Gourgoulianis KI and
Germenis AE: Foxp3 expression in human cancer cells. J Transl Med.
6:192008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang WH, Jiang CL, Yan W, Zhang YH, Yang
JT, Zhang C, Yan B, Zhang W, Han W, Wang JZ and Zhang YQ: FOXP3
expression and clinical characteristics of hepatocellular
carcinoma. World J Gastroenterol. 16:5502–5509. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fu HY, Li C, Yang W, Gai XD, Jia T, Lei YM
and Li Y: FOXP3 and TLR4 protein expression are correlated in
non-small cell lung cancer: Implications for tumor progression and
escape. Acta Histochem. 115:151–157. 2013. View Article : Google Scholar
|
|
23
|
Kim M, Grimmig T, Grimm M, Lazariotou M,
Meier E, Rosenwald A, Tsaur I, Blaheta R, Heemann U, Germer CT, et
al: Expression of Foxp3 in colorectal cancer but not in Treg cells
correlates with disease progression in patients with colorectal
cancer. PLoS One. 8:e536302013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Merlo A, Casalini P, Carcangiu ML,
Malventano C, Triulzi T, Mènard S, Tagliabue E and Balsari A: FOXP3
expression and overall survival in breast cancer. J Clin Oncol.
27:1746–1752. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wolf D, Wolf AM, Rumpold H, Fiegl H,
Zeimet AG, Muller-Holzner E, Deibl M, Gastl G, Gunsilius E and
Marth C: The expression of the regulatory T cell-specific forkhead
box transcription factor FoxP3 is associated with poor prognosis in
ovarian cancer. Clin Cancer Res. 11:8326–8331. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kiniwa Y, Miyahara Y, Wang HY, Peng W,
Peng G, Wheeler TM, Thompson TC, Old LJ and Wang RF:
CD8+ Foxp3+ regulatory T cells mediate
immunosuppression in prostate cancer. Clin Cancer Res.
13:6947–6958. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Niu J, Jiang C, Li C, Liu L, Li K, Jian Z
and Gao T: Foxp3 expression in melanoma cells as a possible
mechanism of resistance to immune destruction. Cancer Immunol
Immunother. 60:1109–1118. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ebert LM, Tan BS, Browning J, Svobodova S,
Russell SE, Kirkpatrick N, Gedye C, Moss D, Ng SP, MacGregor D, et
al: The regulatory T cell-associated transcription factor FoxP3 is
expressed by tumor cells. Cancer Res. 68:3001–3009. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Douglass S, Ali S, Meeson AP, Browell D
and Kirby JA: The role of FOXP3 in the development and metastatic
spread of breast cancer. Cancer Metastasis Rev. 31:843–854. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zeng C, Yao Y, Jie W, Zhang M, Hu X, Zhao
Y, Wang S, Yin J and Song Y: Up-regulation of Foxp3 participates in
progression of cervical cancer. Cancer Immunol Immunother.
62:481–487. 2013. View Article : Google Scholar
|
|
31
|
Triulzi T, Tagliabue E, Balsari A and
Casalini P: FOXP3 expression in tumor cells and implications for
cancer progression. J Cell Physiol. 228:30–35. 2013. View Article : Google Scholar
|
|
32
|
Quaglino P, Osella-Abate S, Marenco F,
Nardò T, Gado C, Novelli M, Savoia P and Bernengo MG: FoxP3
expression on melanoma cells is related to early visceral spreading
in melanoma patients treated by electrochemotherapy. Pigment Cell
Melanoma Res. 24:734–736. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gerber AL, Münst A, Schlapbach C, Shafighi
M, Kiermeir D, Hüsler R and Hunger RE: High expression of FOXP3 in
primary melanoma is associated with tumour progression. Br J
Dermatol. 170:103–109. 2014. View Article : Google Scholar
|
|
34
|
Viguier M, Lemaître F, Verola O, Cho MS,
Gorochov G, Dubertret L, Bachelez H, Kourilsky P and Ferradini L:
Foxp3 expressing CD4+CD25(high) regulatory T cells are
overrepresented in human metastatic melanoma lymph nodes and
inhibit the function of infiltrating T cells. J Immunol.
173:1444–1453. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Knol AC, Nguyen JM, Quéreux G, Brocard A,
Khammari A and Dréno B: Prognostic value of tumor-infiltrating
Foxp3+ T-cell subpopulations in metastatic melanoma. Exp
Dermatol. 20:430–434. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jandus C, Bioley G, Speiser DE and Romero
P: Selective accumulation of differentiated FOXP3(+) CD4 (+) T
cells in metastatic tumor lesions from melanoma patients compared
to peripheral blood. Cancer Immunol Immunother. 57:1795–1805. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang L and Zhao Y: The regulation of
Foxp3 expression in regulatory CD4(+)CD25(+)T cells: Multiple
pathways on the road. J Cell Physiol. 211:590–597. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang X, Liu Y, Dai L, Liu Q, Jia L, Wang
H, An L, Jing X, Liu M, Li P and Cheng Z: Foxp3 downregulation in
NSCLC mediates epithelial-mesenchymal transition via NF-κB
signaling. Oncol Rep. 36:2282–2288. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ou-Yang HF, Zhang HW, Wu CG, Zhang P,
Zhang J, Li JC, Hou LH, He F, Ti XY, Song LQ, et al: Notch
signaling regulates the FOXP3 promoter through RBP-J- and
Hes1-dependent mechanisms. Mol Cell Biochem. 320:109–114. 2009.
View Article : Google Scholar
|
|
40
|
Maruyama T, Konkel JE, Zamarron BF and
Chen W: The molecular mechanisms of Foxp3 gene regulation. Semin
Immunol. 23:418–423. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Uzdensky AB, Demyanenko SV and Bibov MY:
Signal transduction in human cutaneous melanoma and target drugs.
Curr Cancer Drug Targets. 13:843–866. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu J, Sato C, Cerletti M and Wagers A:
Notch signaling in the regulation of stem cell self-renewal and
differentiation. Curr Top Dev Biol. 92:367–409. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Artavanis-Tsakonas S, Rand MD and Lake RJ:
Notch signaling: Cell fate control and signal integration in
development. Science. 284:770–776. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bray SJ: Notch signalling: A simple
pathway becomes complex. Nat Rev Mol Cell Biol. 7:678–689. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Radtke F and Raj K: The role of Notch in
tumorigenesis: Oncogene or tumour suppressor. Nat Rev Cancer.
3:756–767. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Roy M, Pear WS and Aster JC: The
multifaceted role of Notch in cancer. Curr Opin Genet Dev.
17:52–59. 2007. View Article : Google Scholar
|
|
47
|
Kopan R and Ilagan MX: The canonical Notch
signaling pathway: Unfolding the activation mechanism. Cell.
137:216–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Fortini ME: Notch signaling: The core
pathway and its posttranslational regulation. Dev Cell. 16:633–647.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Schroeter EH, Kisslinger JA and Kopan R:
Notch-1 signalling requires ligand-induced proteolytic release of
intracellular domain. Nature. 393:382–386. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Koch U and Radtke F: Notch signaling in
solid tumors. Curr Top Dev Biol. 92:411–455. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Gao J, Dong Y, Zhang B, Xiong Y, Xu W,
Cheng Y, Dai M, Yu Z, Xu H and Zheng G: Notch1 activation
contributes to tumor cell growth and proliferation in human
hepatocellular carcinoma HepG2 and SMMC7721 cells. Int J Oncol.
41:1773–1781. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Bolós V, Mira E, Martínez-Poveda B, Luxán
G, Cañamero M, Martínez-A C, Mañes S and de la Pompa JL: Notch
activation stimulates migration of breast cancer cells and promotes
tumor growth. Breast Cancer Res. 15:R542013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Reedijk M, Odorcic S, Zhang H, Chetty R,
Tennert C, Dickson BC, Lockwood G, Gallinger S and Egan SE:
Activation of Notch signaling in human colon adenocarcinoma. Int J
Oncol. 33:1223–1229. 2008.PubMed/NCBI
|
|
54
|
Yuan X, Wu H, Xu H, Han N, Chu Q, Yu S,
Chen Y and Wu K: Meta-analysis reveals the correlation of Notch
signaling with non-small cell lung cancer progression and
prognosis. Sci Rep. 5:103382015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hijioka H, Setoguchi T, Miyawaki A, Gao H,
Ishida T, Komiya S and Nakamura N: Upregulation of Notch pathway
molecules in oral squamous cell carcinoma. Int J Oncol. 36:817–822.
2010.PubMed/NCBI
|
|
56
|
Ai Q, Ma X, Huang Q, Liu S, Shi T, Zhang
C, Zhu M, Zhang Y, Wang B, Ni D, et al: High-level expression of
Notch1 increased the risk of metastasis in T1 stage clear cell
renal cell carcinoma. PLoS One. 7:e350222012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Pinnix CC, Lee JT, Liu ZJ, McDaid R,
Balint K, Beverly LJ, Brafford PA, Xiao M, Himes B, Zabierowski SE,
et al: Active Notch1 confers a transformed phenotype to primary
human melanocytes. Cancer Res. 69:5312–5320. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Howard JD, Moriarty WF, Park J, Riedy K,
Panova IP, Chung CH, Suh KY, Levchenko A and Alani RM: Notch
signaling mediates melanoma-endothelial cell communication and
melanoma cell migration. Pigment Cell Melanoma Res. 26:697–707.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Müller CS: Notch signaling and malignant
melanoma. Adv Exp Med Biol. 727:258–264. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Liu ZJ, Xiao M, Balint K, Smalley KS,
Brafford P, Qiu R, Pinnix CC, Li X and Herlyn M: Notch1 signaling
promotes primary melanoma progression by activating
mitogen-activated protein kinase/phosphatidylinositol 3-kinase-Akt
pathways and up-regulating N-cadherin expression. Cancer Res.
66:4182–4190. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Balint K, Xiao M, Pinnix CC, Soma A, Veres
I, Juhasz I, Brown EJ, Capobianco AJ, Herlyn M and Liu ZJ:
Activation of Notch1 signaling is required for
beta-catenin-mediated human primary melanoma progression. J Clin
Invest. 115:3166–3176. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Massi D, Tarantini F, Franchi A,
Paglierani M, Di Serio C, Pellerito S, Leoncini G, Cirino G,
Geppetti P and Santucci M: Evidence for differential expression of
Notch receptors and their ligands in melanocytic nevi and cutaneous
malignant melanoma. Mod Pathol. 19:246–254. 2006. View Article : Google Scholar
|
|
63
|
Akhurst RJ and Derynck R: TGF-beta
signaling in cancer - a double-edged sword. Trends Cell Biol.
11:S44–S51. 2001.PubMed/NCBI
|
|
64
|
Trapani JA: The dual adverse effects of
TGF-beta secretion on tumor progression. Cancer Cell. 8:349–350.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Massagué J: TGFbeta in cancer. Cell.
134:215–230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Li MO, Wan YY, Sanjabi S, Robertson AK and
Flavell RA: Transforming growth factor-beta regulation of immune
responses. Annu Rev Immunol. 24:99–146. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Huber S, Schramm C, Lehr HA, Mann A,
Schmitt S, Becker C, Protschka M, Galle PR, Neurath MF and Blessing
M: Cutting edge: TGF-beta signaling is required for the in vivo
expansion and immunosuppressive capacity of regulatory
CD4+CD25+ T cells. J Immunol. 173:6526–6531.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Chen W, Jin W, Hardegen N, Lei KJ, Li L,
Marinos N, McGrady G and Wahl SM: Conversion of peripheral
CD4+CD25− naive T cells to
CD4+CD25+ regulatory T cells by TGF-beta
induction of transcription factor Foxp3. J Exp Med. 198:1875–1886.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Pyzik M and Piccirillo CA: TGF-beta1
modulates Foxp3 expression and regulatory activity in distinct
CD4+ T cell subsets. J Leukoc Biol. 82:335–346. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Oft M, Heider KH and Beug H: TGFbeta
signaling is necessary for carcinoma cell invasiveness and
metastasis. Curr Biol. 8:1243–1252. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zhang HJ, Wang HY, Zhang HT, Su JM, Zhu J,
Wang HB, Zhou WY, Zhang H, Zhao MC, Zhang L and Chen XF:
Transforming growth factor-β1 promotes lung adenocarcinoma invasion
and metastasis by epithelial-to-mesenchymal transition. Mol Cell
Biochem. 355:309–314. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Lee D, Chung YH, Kim JA and Lee YS, Lee D,
Jang MK, Kim KM, Lim YS, Lee HC and Lee YS: Transforming growth
factor beta 1 overexpression is closely related to invasiveness of
hepatocellular carcinoma. Oncology. 82:11–18. 2012. View Article : Google Scholar
|
|
73
|
Teraoka H, Sawada T, Yamashita Y, Nakata
B, Ohira M, Ishikawa T, Nishino H and Hirakawa K: TGF-β1 promotes
liver metastasis of pancreatic cancer by modulating the capacity of
cellular invasion. Int J Oncol. 19:709–715. 2001.PubMed/NCBI
|
|
74
|
Malaponte G, Zacchia A, Bevelacqua Y,
Marconi A, Perrotta R, Mazzarino MC, Cardile V and Stivala F:
Co-regulated expression of matrix metalloproteinase-2 and
transforming growth factor-β in melanoma development and
progression. Oncol Rep. 24:81–87. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Ostroukhova M, Qi Z, Oriss TB,
Dixon-McCarthy B, Ray P and Ray A: Treg-mediated immunosuppression
involves activation of the Notch-HES1 axis by membrane-bound
TGF-beta. J Clin Invest. 116:996–1004. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Samon JB, Champhekar A, Minter LM, Telfer
JC, Miele L, Fauq A, Das P, Golde TE and Osborne BA: Notch1 and
TGFbeta1 cooperatively regulate Foxp3 expression and the
maintenance of peripheral regulatory T cells. Blood. 112:1813–1821.
2008. View Article : Google Scholar
|
|
77
|
Zhou J, Jain S, Azad AK, Xu X, Yu HC, Xu
Z, Godbout R and Fu Y: Notch and TGFβ form a positive regulatory
loop and regulate EMT in epithelial ovarian cancer cells. Cell
Signal. 28:838–849. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zavadil J, Cermak L, Soto-Nieves N and
Böttinger EP: Integration of TGF-beta/Smad and Jagged1/Notch
signalling in epithelial-to-mesenchymal transition. EMBO J.
23:1155–1165. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Blokzijl A, Dahlqvist C, Reissmann E, Falk
A, Moliner A, Lendahl U and Ibáñez CF: Cross-talk between the Notch
and TGF-beta signaling pathways mediated by interaction of the
Notch intracellular domain with Smad3. J Cell Biol. 163:723–728.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Klüppel M and Wrana JL: Turning it up a
Notch: Cross-talk between TGF beta and Notch signaling. BioEssays.
27:115–118. 2005. View Article : Google Scholar
|
|
81
|
Barbarulo A, Grazioli P, Campese AF,
Bellavia D, Di Mario G, Pelullo M, Ciuffetta A, Colantoni S, Vacca
A, Frati L, et al: Notch3 and canonical NF-kappaB signaling
pathways cooperatively regulate Foxp3 transcription. J Immunol.
186:6199–6206. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Burghardt S, Claass B, Erhardt A, Karimi K
and Tiegs G: Hepatocytes induce Foxp3+ regulatory T
cells by Notch signaling. J Leukoc Biol. 96:571–577. 2014.
View Article : Google Scholar
|
|
83
|
Mota C, Nunes-Silva V, Pires AR, Matoso P,
Victorino RM, Sousa AE and Caramalho I: Delta-like 1-mediated Notch
signaling enhances the in vitro conversion of human memory CD4 T
cells into FOXP3-expressing regulatory T cells. J Immunol.
193:5854–5862. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Trehanpati N, Shrivastav S, Shivakumar B,
Khosla R, Bhardwaj S, Chaturvedi J, Sukriti, Kumar B, Bose S, Mani
Tripathi D, et al: Analysis of Notch and TGF-β signaling expression
in different stages of disease progression during hepatitis B virus
infection. Clin Transl Gastroenterol. 3:e232012. View Article : Google Scholar
|
|
85
|
Luo X, Tan H, Zhou Y, Xiao T, Wang C and
Li Y: Notch1 signaling is involved in regulating Foxp3 expression
in T-ALL. Cancer Cell Int. 13:342013. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Josien H: Recent advances in the
development of gamma-secretase inhibitors. Curr Opin Drug Discov
Devel. 5:513–525. 2002.PubMed/NCBI
|
|
87
|
Cardile V, Frasca G, Libra M, Caggia S,
Umezawa K, Panico A and Malaponte G: Dehydroxymethylepoxyquinomicin
inhibits expression and production of inflammatory mediators in
interleukin-1beta-induced human chondrocytes. Cell Physiol Biochem.
25:543–550. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Ohnuki H and Tosato G: Notch and TGFβ:
Functional partners facilitating tumor progression. OncoImmunology.
3:e290292014. View Article : Google Scholar
|
|
89
|
Hoek K, Rimm DL, Williams KR, Zhao H,
Ariyan S, Lin A, Kluger HM, Berger AJ, Cheng E, Trombetta ES, et
al: Expression profiling reveals novel pathways in the
transformation of melanocytes to melanomas. Cancer Res.
64:5270–5282. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Perrot CY, Javelaud D and Mauviel A:
Insights into the transforming growth factor-β signaling pathway in
cutaneous melanoma. Ann Dermatol. 25:135–144. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Zhang J, Wang Y, Li D and Jing S: Notch
and TGF-β/Smad3 pathways are involved in the interaction between
cancer cells and cancer-associated fibroblasts in papillary thyroid
carcinoma. Tumour Biol. 35:379–385. 2014. View Article : Google Scholar
|
|
92
|
Zhang HY and Sun H: Up-regulation of Foxp3
inhibits cell proliferation, migration and invasion in epithelial
ovarian cancer. Cancer Lett. 287:91–97. 2010. View Article : Google Scholar
|
|
93
|
Wang L, Liu R, Li W, Chen C, Katoh H, Chen
GY, McNally B, Lin L, Zhou P, Zuo T, et al: Somatic single hits
inactivate the X-linked tumor suppressor FOXP3 in the prostate.
Cancer Cell. 16:336–346. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zuo T, Liu R, Zhang H, Chang X and Liu Y,
Wang L, Zheng P and Liu Y: FOXP3 is a novel transcriptional
repressor for the breast cancer oncogene SKP2. J Clin Invest.
117:3765–3773. 2007.PubMed/NCBI
|
|
95
|
Liu Y, Zhang P, Li J, Kulkarni AB,
Perruche S and Chen W: A critical function for TGF-beta signaling
in the development of natural
CD4+CD25+Foxp3+ regulatory T
cells. Nat Immunol. 9:632–640. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Brody JR, Costantino CL, Berger AC, Sato
T, Lisanti MP, Yeo CJ, Emmons RV and Witkiewicz AK: Expression of
indoleamine 2,3-dioxygenase in metastatic malignant melanoma
recruits regulatory T cells to avoid immune detection and affects
survival. Cell Cycle. 8:1930–1934. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Chen C, Rowell EA, Thomas RM, Hancock WW
and Wells AD: Transcriptional regulation by Foxp3 is associated
with direct promoter occupancy and modulation of histone
acetylation. J Biol Chem. 281:36828–36834. 2006. View Article : Google Scholar
|
|
98
|
Dimitrakopoulos FI, Papadaki H,
Antonacopoulou AG, Kottorou A, Gotsis AD, Scopa C, Kalofonos HP and
Mouzaki A: Association of FOXP3 expression with non-small cell lung
cancer. Anticancer Res. 31:1677–1683. 2011.PubMed/NCBI
|
|
99
|
Franco-Molina MA, Miranda-Hernández DF,
Mendoza- Gamboa E, Zapata-Benavides P, Coronado-Cerda EE, Sierra-
Rivera CA, Saavedra-Alonso S, Taméz-Guerra RS and Rodríguez-Padilla
C: Silencing of Foxp3 delays the growth of murine melanomas and
modifies the tumor immunosuppressive environment. OncoTargets Ther.
9:243–253. 2016. View Article : Google Scholar
|
|
100
|
Fantini MC, Becker C, Monteleone G,
Pallone F, Galle PR and Neurath MF: Cutting edge: TGF-beta induces
a regulatory phenotype in CD4+CD25− T cells
through Foxp3 induction and down-regulation of Smad7. J Immunol.
172:5149–5153. 2004. View Article : Google Scholar
|
|
101
|
Selvaraj RK and Geiger TL: A kinetic and
dynamic analysis of Foxp3 induced in T cells by TGF-beta. J
Immunol. 178:7667–7677. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Guo X and Wang XF: Signaling cross-talk
between TGF-beta/BMP and other pathways. Cell Res. 19:71–88. 2009.
View Article : Google Scholar
|
|
103
|
Wang Y, Shen RW, Han B, Li Z, Xiong L,
Zhang FY, Cong BB and Zhang B: Notch signaling mediated by
TGF-β/Smad pathway in concanavalin A-induced liver fibrosis in
rats. World J Gastroenterol. 23:2330–2336. 2017. View Article : Google Scholar :
|
|
104
|
Yan XC, Cao J, Liang L, Wang L, Gao F,
Yang ZY, Duan JL, Chang TF, Deng SM, Liu Y, et al: miR-342-5p is a
notch downstream molecule and regulates multiple angiogenic
pathways including notch, vascular endothelial growth factor and
transforming growth factor β signaling. J Am Heart Assoc.
5:e0030422016. View Article : Google Scholar
|
|
105
|
Kared H, Adle-Biassette H, Foïs E, Masson
A, Bach JF, Chatenoud L, Schneider E and Zavala F:
Jagged2-expressing hematopoietic progenitors promote regulatory T
cell expansion in the periphery through notch signaling. Immunity.
25:823–834. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Stockhausen MT, Sjö J and Axelson H:
Regulation of the Notch target gene Hes-1 by TGFalpha induced
Ras/MAPK signaling in human neuroblastoma cells. Exp Cell Res.
310:218–228. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Pisklakova A, Grigson E, Ozerova M, Chen
F, Sullivan DM and Nefedova Y: Anti-myeloma effect of
pharmacological inhibition of Notch/gamma-secretase with RO4929097
is mediated by modulation of tumor microenvironment. Cancer Biol
Ther. 17:477–485. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Tas F, Karabulut S, Yasasever CT and
Duranyildiz D: Serum transforming growth factor-beta 1 (TGF-β1)
levels have diagnostic, predictive, and possible prognostic roles
in patients with melanoma. Tumour Biol. 35:7233–7237. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Takizawa T, Ochiai W, Nakashima K and Taga
T: Enhanced gene activation by Notch and BMP signaling cross-talk.
Nucleic Acids Res. 31:5723–5731. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Asnaghi L, Ebrahimi KB, Schreck KC, Bar
EE, Coonfield ML, Bell WR, Handa J, Merbs SL, Harbour JW and
Eberhart CG: Notch signaling promotes growth and invasion in uveal
melanoma. Clin Cancer Res. 18:654–665. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Sriuranpong V, Borges MW, Ravi RK, Arnold
DR, Nelkin BD, Baylin SB and Ball DW: Notch signaling induces cell
cycle arrest in small cell lung cancer cells. Cancer Res.
61:3200–3205. 2001.PubMed/NCBI
|
|
112
|
Thélu J, Rossio P and Favier B: Notch
signalling is linked to epidermal cell differentiation level in
basal cell carcinoma, psoriasis and wound healing. BMC Dermatol.
2:72002. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Panelos J, Tarantini F, Paglierani M, Di
Serio C, Maio V, Pellerito S, Pimpinelli N, Santucci M and Massi D:
Photoexposition discriminates Notch 1 expression in human cutaneous
squamous cell carcinoma. Mod Pathol. 21:316–325. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Banerjee D, Hernandez SL, Garcia A,
Kangsamaksin T, Sbiroli E, Andrews J, Forrester LA, Wei N,
Kadenhe-Chiweshe A, Shawber CJ, et al: Notch suppresses
angiogenesis and progression of hepatic metastases. Cancer Res.
75:1592–1602. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Talora C, Cialfi S, Segatto O, Morrone S,
Kim Choi J, Frati L, Paolo Dotto G, Gulino A and Screpanti I:
Constitutively active Notch1 induces growth arrest of HPV-positive
cervical cancer cells via separate signaling pathways. Exp Cell
Res. 305:343–354. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Takebe N, Nguyen D and Yang SX: Targeting
notch signaling pathway in cancer: Clinical development advances
and challenges. Pharmacol Ther. 141:140–149. 2014. View Article : Google Scholar :
|
|
117
|
Olsauskas-Kuprys R, Zlobin A and Osipo C:
Gamma secretase inhibitors of Notch signaling. Onco Targets Ther.
6:943–955. 2013.PubMed/NCBI
|
|
118
|
Ji X, Wang Z, Geamanu A, Sarkar FH and
Gupta SV: Inhibition of cell growth and induction of apoptosis in
non-small cell lung cancer cells by delta-tocotrienol is associated
with notch-1 down-regulation. J Cell Biochem. 112:2773–2783. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Wang M, Wu L, Wang L and Xin X:
Down-regulation of Notch1 by gamma-secretase inhibition contributes
to cell growth inhibition and apoptosis in ovarian cancer cells
A2780. Biochem Biophys Res Commun. 393:144–149. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Hu J, Zhu X and Lu Q: Antiproliferative
effects of γ-secretase inhibitor, a Notch signalling inhibitor, in
multiple myeloma cells and its molecular mechanism of action. J Int
Med Res. 41:1017–1026. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Qi R, An H, Yu Y, Zhang M, Liu S, Xu H,
Guo Z, Cheng T and Cao X: Notch1 signaling inhibits growth of human
hepatocellular carcinoma through induction of cell cycle arrest and
apoptosis. Cancer Res. 63:8323–8329. 2003.PubMed/NCBI
|
|
122
|
Wang L, Qin H, Chen B, Xin X, Li J and Han
H: Overexpressed active Notch1 induces cell growth arrest of HeLa
cervical carcinoma cells. Int J Gynecol Cancer. 17:1283–1292. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Miranda-Hernández DF, Franco-Molina MA,
Mendoza-Gamboa E, Zapata-Benavides P, Sierra-Rivera CA,
Coronado-Cerda EE, Rosas-Taraco AG, Taméz-Guerra RS and
Rodríguez-Padilla C: Expression of Foxp3, CD25 and IL-2 in the
B16F10 cancer cell line and melanoma is correlated with tumor
growth in mice. Oncol Lett. 6:1195–1200. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Del Papa B, Sportoletti P, Cecchini D,
Rosati E, Balucani C, Baldoni S, Fettucciari K, Marconi P, Martelli
MF, Falzetti F and Di Ianni M: Notch1 modulates mesenchymal stem
cells mediated regulatory T-cell induction. Eur J Immunol.
43:182–187. 2013. View Article : Google Scholar
|
|
125
|
Rao P and Kadesch T: The intracellular
form of notch blocks transforming growth factor beta-mediated
growth arrest in Mv1Lu epithelial cells. Mol Cell Biol.
23:6694–6701. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Sun XF, Sun XH, Cheng SF, Wang JJ, Feng
YN, Zhao Y, Yin S, Hou ZM, Shen W and Zhang XF: Interaction of the
transforming growth factor-β and Notch signaling pathways in the
regulation of granulosa cell proliferation. Reprod Fertil Dev.
28:1873–1881. 2016. View Article : Google Scholar
|
|
127
|
Masuda S, Kumano K, Shimizu K, Imai Y,
Kurokawa M, Ogawa S, Miyagishi M, Taira K, Hirai H and Chiba S:
Notch1 oncoprotein antagonizes TGF-beta/Smad-mediated cell growth
suppression via sequestration of coactivator p300. Cancer Sci.
96:274–282. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Asano N, Watanabe T, Kitani A, Fuss IJ and
Strober W: Notch1 signaling and regulatory T cell function. J
Immunol. 180:2796–2804. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Xu L, Kitani A, Stuelten C, McGrady G,
Fuss I and Strober W: Positive and negative transcriptional
regulation of the Foxp3 gene is mediated by access and binding of
the Smad3 protein to enhancer I. Immunity. 33:313–325. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Tone Y, Furuuchi K, Kojima Y, Tykocinski
ML, Greene MI and Tone M: Smad3 and NFAT cooperate to induce Foxp3
expression through its enhancer. Nat Immunol. 9:194–202. 2008.
View Article : Google Scholar
|