Introduction
Intestinal ischemia and reperfusion (II/R) is
encountered under various clinical conditions, and contributes to
multi-organ failure and high levels of mortality (60–80%) (1–3).
II/R not only induces intestinal damage, but also affects remote
organs, including the lungs, leading to acute lung injury (ALI) and
acute respiratory distress syndrome in patients (4,5).
II/R-induced ALI is caused by an excessive systemic inflammatory
response, which is triggered by the release of proinflammatory
cytokines and bacteria-derived endotoxins from the reperfused
ischemic gut tissue (6–8). In addition, animal models and
clinical data support the concept that excessive elevation of
proinflammatory cytokines is a major contributor in remote organ
injury following II/R (9–11).
Among the numerous proinflammatory cytokines, tumor
necrosis factor-α (TNF-α) has a critical role in the occurrence and
development of ALI caused by II/R (12,13). Preclinical and clinical studies
have reported that suppressing the expression of TNF-α may reduce
the progression of inflammation in numerous diseases, including
Crohn's disease (14–16). Using an anti-TNF antibody, Caty
et al revealed that blocking TNF ameliorates pulmonary
microvascular permeability (13).
In addition, Sorkine et al demonstrated that soluble TNF-α
receptors have the ability to reduce bowel ischemia-induced lung
permeability and neutrophil sequestration (17). These results suggest that TNF-α
may serve major roles in the lung injury induced by II/R. However,
pharmacological treatment options for ALI following II/R are
limited, with most targeting proinflammatory cytokines and
oxidative stress pathways (18).
Therefore, there is an urgent requirement to identify an effective
approach for ALI treatment.
Downregulation of mRNA transcripts by RNA
interference (RNAi) and small interfering (si)RNA (19) has been adopted as an invaluable
research tool, which holds promise as a novel strategy for drug
development (14), via the
suppression of targeted gene expression (20). As a therapeutic method, the RNAi
approach is safe and has incomparable long-term effects.
It has been hypothesized that blocking expression of
the proinflammatory cytokine TNF-α may protect the lungs from
remote organ injury following II/R. Therefore, the present study
employed a rat model of II/R injury and used short hairpin (sh)RNA
technology to examine the efficacy of TNF-α knockdown on
II/R-induced ALI, and to investigate its association with
interleukin-10 (IL-10) expression in lung tissues.
Materials and methods
Animals and grouping
Adult male Sprague-Dawley rats (8–12 weeks old),
weighing 230–280 g, were obtained from the Experimental Animal
Center of Sichuan University (Chengdu, China). Guide lines for
Laboratory Animal Care and Safety from the National Institutes of
Health (Bethesda, MD, USA) were followed. The rats were maintained
in plastic cages (2 rats/cage) with soft bedding, and were given
free access to food and water. Rats were maintained under the
following conditions: Controlled room temperature, 22–25°C
humidity, 45–50%; 12-h light/dark cycle. Animal care and all
experimental protocols were approved by the Institutional Medical
Experimental Animal Care Committee of Kunming Medical University
(Kunming, China).
A total of 152 rats were randomly divided into the
following two groups, as described in Table I: i) Sham and ii) II/R (8, 16 and
24 h). In the II/R group, rats underwent II/R injury via occlusion
of the superior mesenteric artery (SMA) and coeliac artery (CA) for
40 min, and subsequent reperfusion for 8, 16 or 24 h. In addition,
the II/R group was subdivided into the negative control (II/R +
Lv-NC vector) and TNF-α shRNA groups (II/R + RSH054951-5-HIVmU6),
as described in Table II. Rats
in the negative control group were injected with the Lv-NC vector,
whereas rats in the TNF-α shRNA group were injected with the TNF-α
shRNA lentivirus.
 | Table IAnimal model and number of rats
distribution. |
Table I
Animal model and number of rats
distribution.
| Group | Model | Lung edema | H&E | IF | qPCR/WB |
|---|
| Sham | Sham | 8 | 6 | 6 | 8 |
| 8 h | II/R | 8 | 6 | 6 | 8 |
| 16 h | II/R | 8 | 6 | 6 | 8 |
| 24 h | II/R | 8 | 6 | 6 | 8 |
| Table IIAnimal grouping and and number of
rats distribution. |
Table II
Animal grouping and and number of
rats distribution.
| Group | Model (24 h) | H&E | qPCR/WB | IF |
|---|
| Control | II/R+Lv-NC
vector | 6 | 8 | 6 |
| TNF-α shRNA |
II/R+RSH054951-5-HIVmU6 | 6 | 8 | 6 |
Production of lentivirus TNF-α shRNA
plasmid
To investigate the function of TNF-α in rat lungs
following II/R, human immunodeficiency virus (HIV)-based vectors
were used. TNF-α gene information was gathered from the National
Center for Biotechnology Information (https://www.ncbi.nlm.nih.gov/nuccore/NM_012675.3).
One potential shRNA sequence targeting TNF-α mRNA
(gcccgtagcccacgtcgta) was used to silence TNF-α expression (called
RSH054951-5-HIVmU6; provided by GeneCopoeia, Inc., Rockville, MD,
USA), whereas one non-sense shRNA sequence was used as a negative
control; sequences were designed and purchased from GeneCopoeia,
Inc.
RNA knockdown efficiency screening
In order to screen the efficiency of the potential
TNF-α shRNA sequence, PC12 cells, purchased from the Animal
Research Institute of the Chinese Academy of Medical Sciences
(Beijing, China), were seeded in 6-well plates and incubated at
37°C in an atmosphere containing 5% CO2, prior to
transfection with shRNA sequences. Brie fly, cultured PC12 cells
(1×105/ml), were transfected at 4°C with a mixture
including 1 μg shRNA fragment and 3 μl transfection
reagents (SuperFectin™ II; Shanghai Pufei Biological Technology
Co., Ltd., Shanghai, China) according to the manufacturer's
protocol. A total of 12 h post-transfection, basal culture medium
was replaced with Dulbecco's modified Eagle's medium (DMEM;
HyClone; GE Healthcare, Logan, UT, USA) containing 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) and penicillin-streptomycin solution (50 U/ml; HyClone; GE
Healthcare). Subsequently, total RNA was extracted from PC12 cells
48 h post-transfection and the effects of shRNA on TNF-α mRNA
expression were examined using quantitative polymerase chain
reaction (qPCR) (21). The
protein expression levels were evaluated by western blotting.
Recombinant lentiviral vector
production
Lentiviral vector production was conducted according
to the manufacturer's protocol of the Lenti-Pac™ HIV Expression
Packaging kit (GeneCopoeia, Inc.). Briefly, 293Tα lentiviral
packaging cells (GeneCopoeia, Inc.) were cultured in DMEM
supplemented with 10% heat-inactivated FBS at 37°C in an atmosphere
containing 5% CO2. Cell confluence between 70 and 80%
was optimal for transfection. Lentiviral expression plasmid (1.25
μg) was mixed with 2.5 μl (0.5 μg/μl)
packing mix (Lenti-Pac HIV) and the mixture was added to 75
μl Opti-MEM® I (Gibco; Thermo Fisher Scientific,
Inc.). Diluted EndoFectin Lenti was also added to the vector
mixture. Following incubation at room temperature for 25 min, the
mixtures were added to the culture medium of 293Tα cells.
Subsequently, 8 h post-transfection, the culture medium was
replaced with heated DMEM containing 10% FBS. Furthermore, Titer
Boost reagent (10 μl; GeneCopoeia, Inc.) was added to
improve virus generation. Transfection efficiency was confirmed by
the detection of mCherryFP, which was fused to the plasmid vector,
under a fluorescence microscope. A total of 72 h post-transfection,
cells were stimulated, the culture medium was collected and
centrifuged at 3,600 × g for 10 min at 4°C, and the supernatant was
filtered. Lentiviral stocks were aliquoted and stored at −80°C
until further use (21).
II/R model and lentivirus injection
Rats were fasted with no restriction of water access
for 24 h prior to surgery. II/R was induced by SMA and CA
occlusion, as described previously (22). Briefly, the rats were anesthetized
intraperitoneally (i.p.) with ketamine-xylazine (100 and 20 mg/kg
i.p., respectively) and placed in a supine position. The SMA and CA
were exposed and isolated through a midline laparotomy and clamped
with an atraumatic microvascular clip for 40 min. After 40 min
ischemia, the artery clamps were removed and intestinal perfusion
was re-established. Sham animals underwent the same surgical
procedure without artery clamping.
For TNF-α interference, the previously described
lentiviruses were injected through the diaphragm into the right
lung tissue (5 μl/2×108/ml). After 3 min, the
needle tip was pulled out and the diaphragm was sutured. The
arteries were clamped following lentiviral injection. After 40 min
of ischemia, the SMA and CA were loosened and the skin was sutured,
after which 0.5 ml normal saline was injected into the
enterocoelia. Rats in the negative control group were injected with
Lv-NC.
Tissue harvest
At the end of reperfusion (8, 16 and 24 h),
experimental and sham rats were anaesthetized with
ketamine-xylazine (100 and 20 mg/kg i.p., respectively) and
euthanized with pentobarbital sodium (200mg/kg i.p.). Subsequently,
lung tissues from rats in each group were collected and
analyzed.
Lung edema determination
Lung edema was estimated by comparing the lung
wet/dry weight ratio. At the end of the experiments, lungs were
immediately removed and weighed to obtain the wet weight. The
tissues were then dried in an oven at 90°Cfor 24 h and were weighed
again to obtain the dry weight. Lung wet/dry weight ratio was
calculated as previously described (23).
Histological analysis
Histological analysis of the lungs was performed by
hematoxylin and eosin (H&E) staining. Briefly, tissue samples
were fixed in 10% (v/v) formalin in neutral-buffered solution for
72 h at room temprature, and the fixed tissues were embedded in
paraffin. Subsequently, tissue blocks were cut into 5-μm
sections, transferred to glass slides and stained with hematoxylin
for 7 min, and eosin for 15 sec respectively, at room temprature.
Finally, these sections were observed under a light microscope to
detect morphological alterations. Lung injury was scored as
previously described (24).
Reverse transcription (RT)-qPCR
The mRNA expression levels of TNF-α and IL-10 were
detected using RT-qPCR. Briefly, total RNA was isolated from the
lung tissues using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
RNA was reverse transcribed to cDNA using the RevertAid™ First
Strand cDNA Synthesis kit (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. qPCR was then performed
to determine the expression levels of target genes. The primers and
TaqMan probes were designed with Primer Premier 5.0 (Premier
Biosoft International, Palo Alto, CA, USA). The primer sequences
were as follows (5′-3′): TNF-α, forward GCCCACGTCGTAGCAA, reverse,
GTCTTTGAGATCCATGCCAT (annealing temperature, 52°C); IL-10, forward
CAGAAATCAAGGAGCATTTG, reverse CTGCTCCACTGCCTTGCTTT (annealing
temperature, 50°C); and β-actin, forward GAAGATCAAGATCATTGCTCCT and
reverse TACTCCTGCTTGCTGATCCA (annealing temperature, 52°C). The rat
β-actin housekeeping gene was used as an internal control. The qPCR
reactive system was established as follows: 2X PCR, Master Mix
(12.5 μl), PCR water nuclear-free (10.5 μl), forward
primer (0.5 μl), reverse prime (0.5 μl), cDNA
template (1 μl); total, 25 μl; 1 μl water was
added as a negative control instead of cDNA. Amplification was
conducted using an ABI 7300 PCR system (Applied Biosystems; Thermo
Fisher Scientific, Inc.), under the following conditions: Initial
denaturation for 1 cycle at 95°C for 2 min, followed by
denaturation at 95°C for 15 sec, and amplification at 53°C for 20
sec and followed by extension at 60°C for 30 sec for a total of 40
cycles. The quantification cycle (Cq) of each sample was recorded
as a quantitative measure of the amount of PCR product in the
sample. Finally, the relative mRNA expression levels of the target
genes were calculated following normalization to β-actin mRNA using
the 2−ΔΔCq method (25).
Immunofluorescence (IF) staining
A total of 8, 16 and 24 h following reperfusion,
paraffin-embedded lung sections underwent IF staining of TNF-α and
IL-10. Following routine de-paraffinization and rehydration, tissue
sections were incubated with PBS containing 3% goat serum (Sigma,
St. Louis, MO, USA) for 30 min at 37°C, and were then incubated
overnight at 4°C with TNF-α (1:500, rabbit) and IL-10 (1:100,
rabbit; catalog no. Ab9969) primary antibodies (both from Abcam,
Cambridge, UK) which were diluted in PBS containing 2% normal goat
serum. A negative control was performed by adding PBS instead of
the primary antibody. Subsequently, sections were washed three
times with PBS and were incubated with Cy3 fluorescence-labeled
secondary antibody (1:200, anti-rabbit; catalog no. 111-165-003;
Jackson Laboratory, Bar Harbor, ME, USA), in the dark for 30 min at
37°C. Sections were then washed three times with PBS, mounted onto
gelatin-coated glass microscope slides, air dried and cover-slipped
in a glycerol-based mounting medium. Cell nuclei were visualized by
DAPI-Fluoromount (Beyotime Institute of Biotechnology, Shanghai,
China). Photomicrographs were captured under a fluorescence
microscope (Leica Microsystems GmbH, Wetzlar, Germany).
Western blotting
Tissue samples were lysed and homogenized in 50 ml
radioimmunoprecipitation acid lysis buffer (Beyotime Institute of
Biotechnology) containing a 2% protease inhibitor cocktail tablet
(Roche Diagnostics GmbH, Mannheim, Germany). Subsequently, a
Bicinchoninic Acid protein assay kit (Beyotime Institute of
Biotechnology) was used to detect the protein concentration and,
100 μg total protein was then resolved by 15% SDS-PAGE in
electrophoresis buffer (24.8 mM Tris, 192 mM glycine and 0.1% SDS)
at 60 V for 30 min, and 100 V for 1.5 h. The precipitated proteins
were then transferred to polyvinylidene fluoride membranes (EMD
Millipore, Billerica, MA, USA) at 350 mA for 4 h, using transfer
buffer: 24.8 mM Tris, 192 mM glycine and 10% methanol. The
membranes were blocked with Tris-buffered saline with 1/1,000 Tween
(TBST) containing 5% non-fat milk for 1 h at room temperature.
Subsequently, the membranes were incubated with TNF-α primary
antibody (1:5,000, rabbit anti-rat; ab9755; Abcam) in TBS overnight
at 4°C; β-actin (1:1,000; catalog no. ABM40028; Abcam) was used as
an internal control. Following incubation with the primary
antibodies, membranes were repeatedly rinsed in TBST four times
prior to incubation for 1.5 h with a secondary antibody at room
temprature (goat anti-rabbit immunoglobulin G; 1:5,000; catalog no.
ab6721; Abcam). Finally, the membranes were rinsed four times in
TBST and detected using ChemiDoc XRS System with Image Lab Software
2.0 (Bio-Rad Laboratories, Inc., Hercules, CA, USA) using an
Enhanced Chemiluminescence reagent (catalog no. BL520A; Bio-Rad
Laboratories, Inc.).
Statistical analysis
Statistical analysis was conducted using SPSS 18.0
software (SPSS, Inc., Chicago, IL, USA) and the experiments were
repeated 3 times. Data are presented as the means ± standard
deviation and were subjected to statistical analysis using one-way
analysis of variance (ANOVA) or Student's t-test. For multiple
group comparisons, ANOVA with Tukey's post hoc multiple comparisons
test was applied. P<0.05 was considered to indicate a
statistically significant difference.
Results
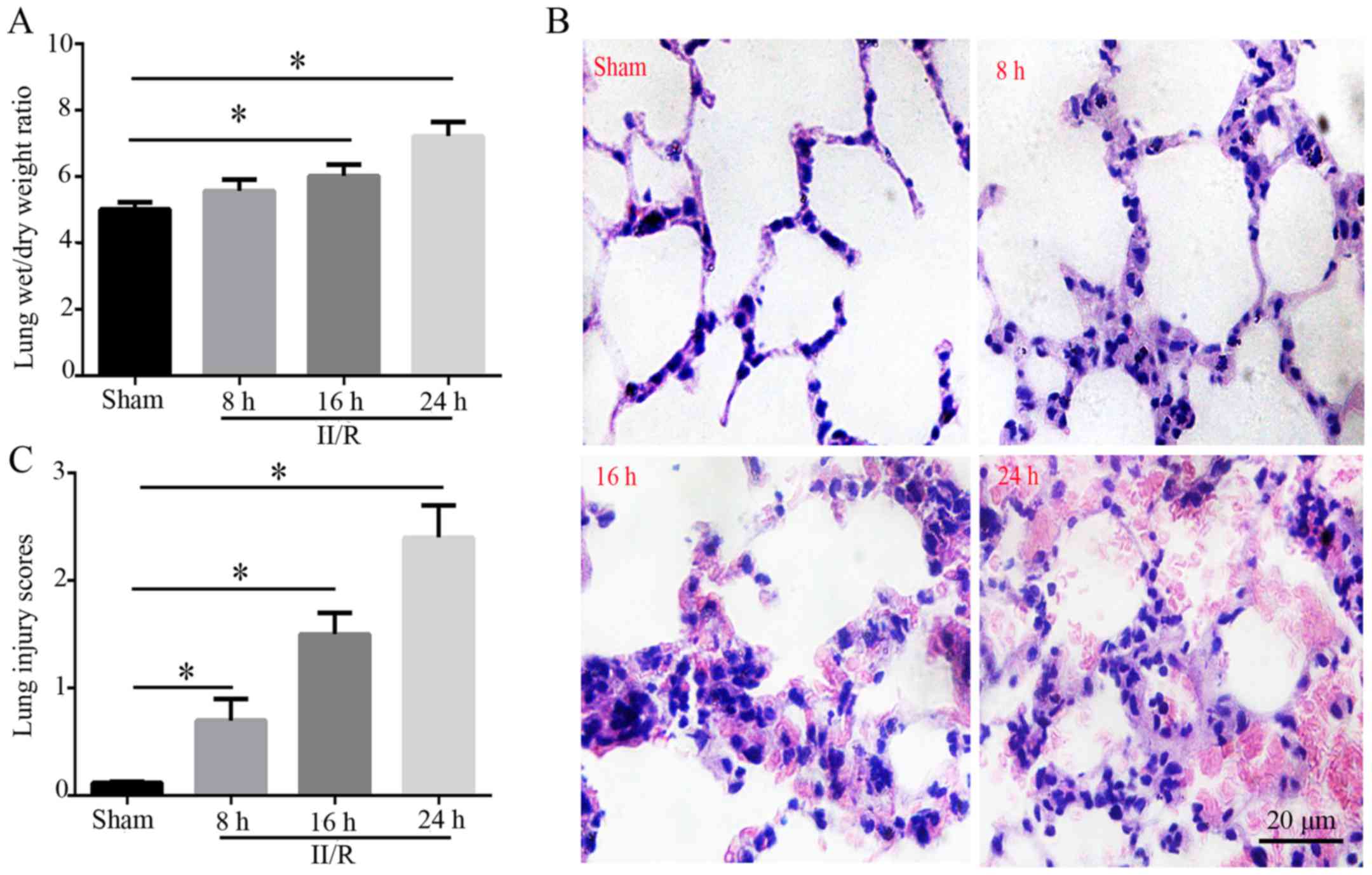
Lung edema and morphology
Compared with in the sham group, the lung wet/dry
weight ratio of the II/R group was significantly increased 8, 16
and 24 h after reperfusion; the results indicated that pulmonary
edema was aggravated as the time interval lengthened (P<0.05;
Fig. 1A). As shown in Fig. 1B, lung tissue sections were
stained with H&E and observed under a light microscope;
alveolar structure in the sham group was ordered, and no
congestion, neutrophil invasion or interstitial edema was detected.
However, in the II/R group, the alveolar structure was disordered
and integrity of the alveolar wall was damaged, which was
accompanied by a thickened alveolar wall and edema. In addition,
neutrophils accumulated in the alveolar space, alveolar capillary
congestion and exudation occurred, part of the alveolus pulmonis
collapsed, and there was hemorrhaging in the alveolar space. Based
on these observations, lung injury scores were significantly
increased in the II/R group compared with in the sham group 8, 16
and 24 h post-reperfusion (P<0.05; Fig. 1C).
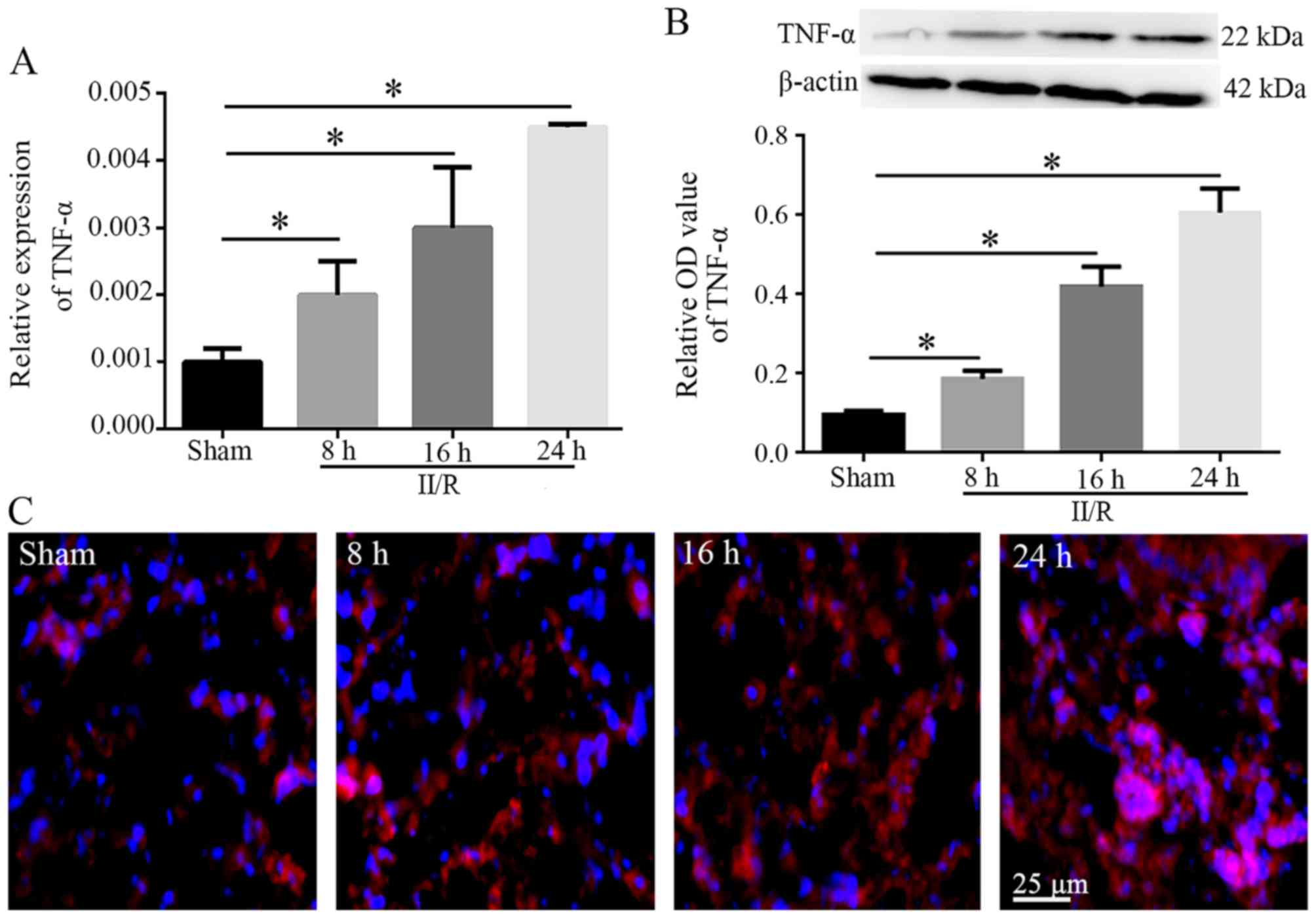
Expression levels of TNF-α in lung
tissues
Between 8 and 24 h post-reperfusion, the mRNA and
protein expression levels of TNF-α in the II/R group exhibited an
increasing trend compared with in the sham group (P<0.05;
Fig. 2A and B). In addition, IF
labeling of TNF-α, which exhibited red fluorescence, indicated that
weak TNF-α immunostaining was detected in the lung tissues of the
sham group, whereas strong immunostaining was detected in the lung
tissues of the II/R group 8, 16 and 24 h post-reperfusion. In
particular, TNF-α immunostaining was diffuse and intense 24 h
following reperfusion (Fig.
2C).
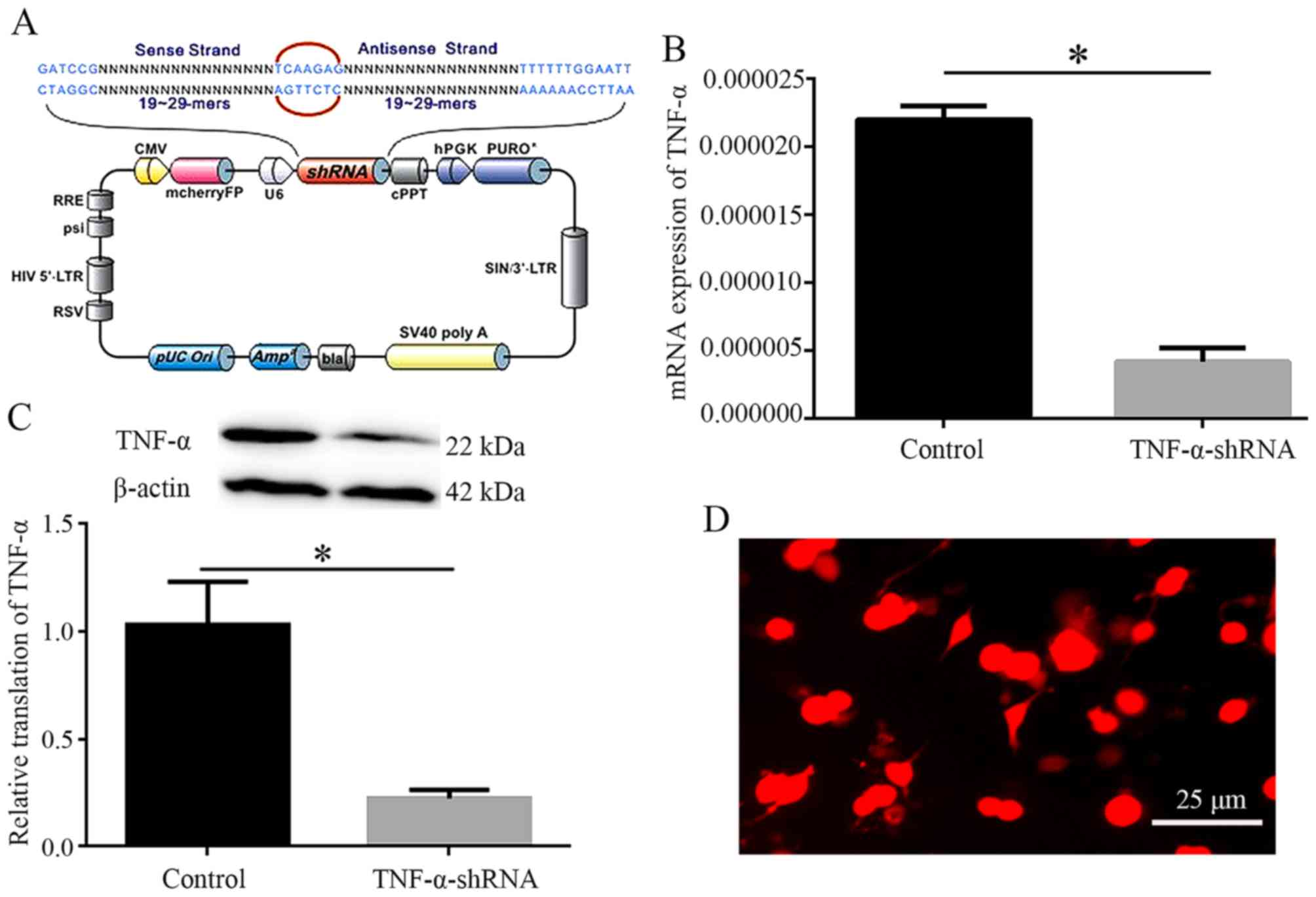
Efficiency of TNF-α knockdown
Lentivirus-mediated TNF-α interference was used to
knockdown the expression of TNF-α. The lentivirus-carried TNF-α
shRNA sequence is presented in Fig.
3A. PC12 cells were subjected to shRNA transfection, in order
to determine the interfering efficiency. Compared with in the
control group, the mRNA and protein expression levels of TNF-α were
reduced in the shRNA plasmid transfection groups (P<0.05), as
determined by qPCR and western blotting (Fig. 3B and C). Subsequently, the shRNA
sequence was inserted into a plasmid to produce a recombinant
vector, after which recombinant vectors containing TNF-α shRNA and
mCherryFP were transfected into 293Tα cells. IF detection confirmed
that 293Tα cell emitted red fluorescence, indicating successful
transfection (Fig. 3D). These
findings confirmed that lentivirus-mediated TNF-α interference was
successful, and with the addition of lentivirus, TNF-α was
significantly inhibited.
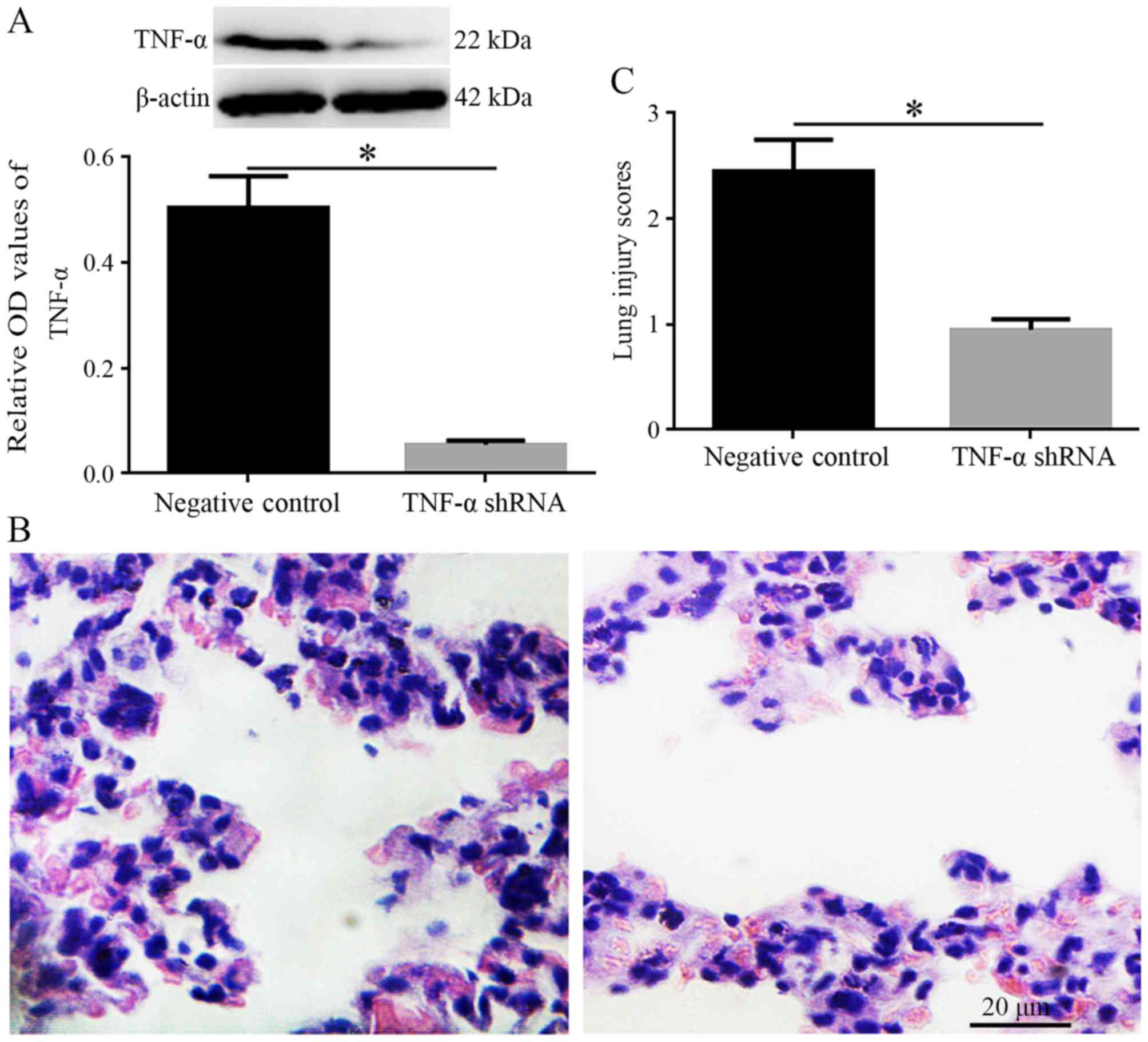
TNF-α knockdown ameliorates ALI following
II/R
Following TNF-α inhibition, western blotting
confirmed that the protein expression levels of TNF-α were
significantly decreased in lung tissues (P<0.05; Fig. 4A). Furthermore, H&E staining
indicated that inflammatory responses were improved, and lung
injury scores were reduced 24 h after reperfusion in the TNF-α
group compared with in the negative control group (P<0.05;
Fig. 4B and C).
Knockdown of TNF-α upregulates the
expression of IL-10
To determine the effects of TNF-α interference on
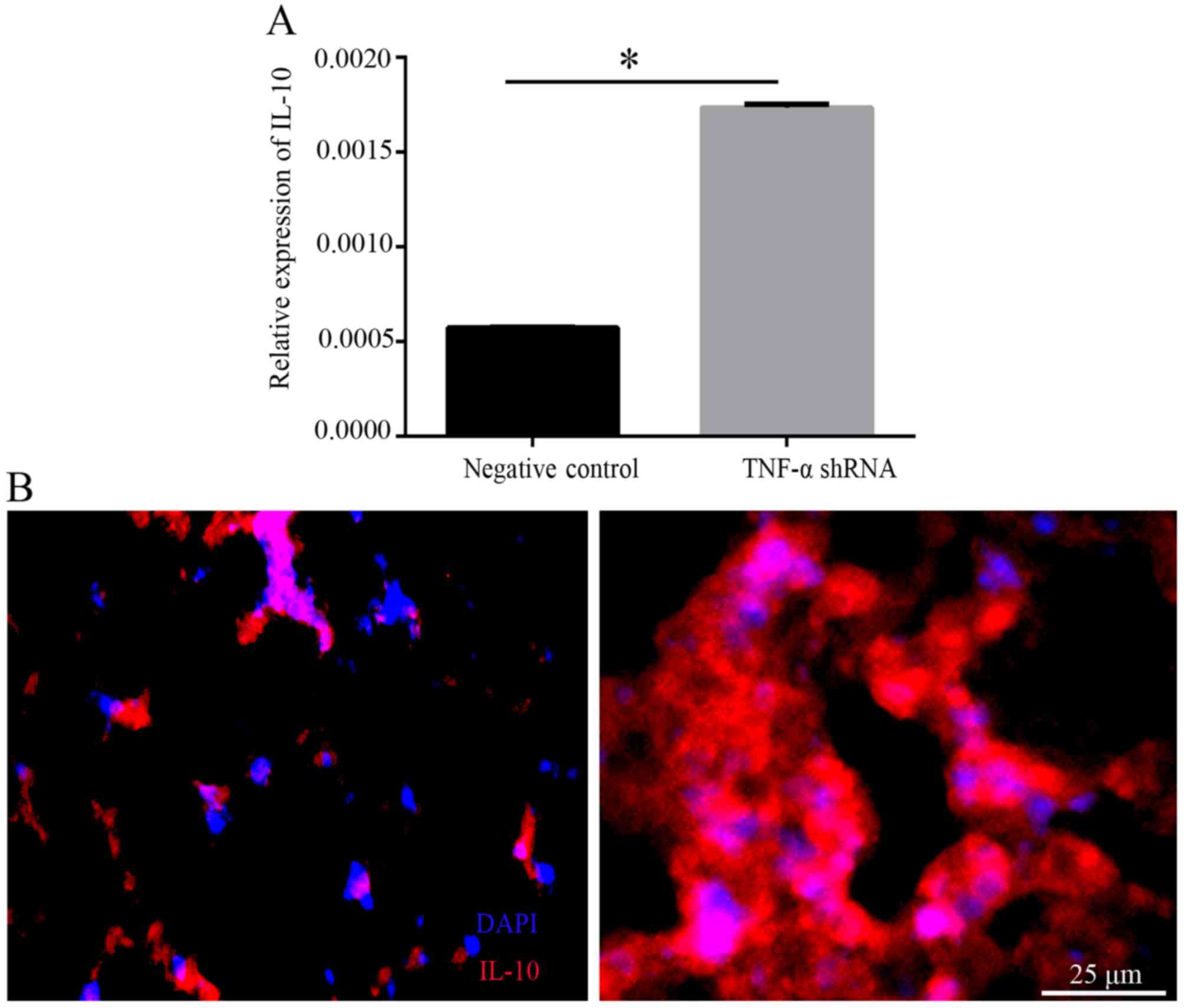
the production of IL-10, the expression levels of IL-10 were
detected in lung tissues. qPCR demonstrated that the mRNA
expression levels of IL-10 were significantly increased in the
TNF-α shRNA group compared with in the negative control group
(P<0.05; Fig. 5A).
Furthermore, immunostaining of IL-10 was stronger in the TNF-α
shRNA group than in the negative control group (Fig. 5B).
Discussion
The present study demonstrated that TNF-α knockdown
may alleviate the inflammatory response associated with
II/R-induced ALI by interfering with TNF-α expression and
upregulating IL-10 in the lung tissues. These findings suggested
that TNF-α RNA interference may be used as a strategy for the
prevention or treatment of II/R-induced ALI in future clinical
trials.
The present results indicated that II/R induced an
acute inflammatory response in the lungs, in which adherence and
infiltration of neutrophils was increased, and interstitial edema
occurred; these observations were associated with worsened lung
injury scores. These observations may significantly contribute to
II/R-induced lung injury. Previous studies have reported that
activated neutrophils are an important factor in tissue injury and
serve a significant role in the progression of ALI (26–28). Therefore, the results of the
present study demonstrated that lung injury was caused by II/R,
thus confirming the model reliability.
The present study suggested that TNF-α, either
locally produced at the site of ischemia or generated directly from
the lung tissue affected by II/R, had important effects on the
lungs. The results indicated that TNF-α expression was upregulated
in lung tissues, alongside ultrastructural alterations and lung
injuries. In addition, the alterations in TNF-α expression levels
differed with the time post-reperfusion. In the present study, the
expression levels of TNF-α were increased in the lung tissues of
the II/R group after 8 h reperfusion and peaked 24 h
post-reperfusion. A previous study provided evidence to suggest
that in I/R injury, excessive elevation of proinflammatory
cytokines is a major contributor in remote organ injury (12). The proinflammatory molecule,
TNF-α, can induce direct tissue damage and is also a potent
activator of neutrophils (29,30). In a previous study, Sorkine et
al revealed that serum TNF concentration peaked 30 min after
reperfusion, but returned to baseline values within 180 min
(17). In addition, Narita et
al reported that plasma TNF-α levels in an II/R group were
increased 30 min after the start of ischemia, and fluctuated during
I/R (30 min after intestinal reperfusion); however, 180 min after
reperfusion, the plasma TNF levels were not significantly increased
in the intestinal I/R rats compared with in the sham rats (12). Conversely, the present results
indicated that the longer the duration of reperfusion, the higher
the levels of TNF-α, this may underlie the reason why more severe
lung injury was detected 24 h post-reperfusion. These findings
indicated that the mRNA expression levels of TNF-α in lung tissue
may be more associated with the degree of lung injury compared with
plasma levels prior to 24 h post-reperfusion.
The present study demonstrated that TNF-α lentiviral
interference decreased the acute inflammatory response, lung injury
and lung edema induced by II/R in rats, and it was indicated that
its protective role may involve upregulation of IL-10. Until
recently, siRNAs have been considered particularly specific
(31,32). However, there is growing evidence
to suggest that the technique has limitations with regards to siRNA
specificity and long-term effects (14,20), whereas shRNAs maintain longer
expression and were therefore used in the present study.
In previous studies, blockade of TNF-α has been
reported to improve or prevent inflammation in animal models and in
humans for the treatment of disease (33–36). In addition, it has been
demonstrated that nanoparticle-mediated TNF-α knockdown in
peritoneal macrophages may be used to reduce local and systemic
inflammation, thereby presenting a novel therapeutic strategy for
arthritis, psoriasis and other skin disorders (22,37). Furthermore, potent RNAi against
systemic TNF-α production provides a promising approach for the
treatment of hepatic injury and other inflammatory diseases
(38).
The treatment for II/R-induced ALI is currently
limited. It has previously been reported that remote intestinal
ischemic preconditioning may confer cytoprotection in critical
organs, including the lungs, by attenuating the release of the
proinflammatory cytokines TNF-α and IL-1 (12,39). Furthermore, it has been revealed
that applying hydrogen-rich saline, ELR-CXC chemokine inhibitors or
heparin-binding EGF-like growth factor attenuates II/R-induced lung
injury by inhibiting II/R injury-associated inflammatory events in
clinical situations (40–42). However, few studies have focused
on the protective effects of TNF-α knockdown, the possible
mechanisms underlying II/R-induced ALI and the potential clinical
application of TNF-α RNAi in patients with II/R-induced ALI. To the
best of our knowledge, the present study is the first to
demonstrate the protective effects of TNF-α RNAi on II/R-induced
ALI via the upregulation of IL-10 expression (Fig. 6).
 | Figure 6Diagram of the underlying mechanism
by which TNF-α shRNA serves a protective role in a rat model of
II/R-induced ALI. At various time points post-reperfusion, evidence
of ALI was detected by hematoxylin and eosin staining. qPCR, WB and
IF indicated that the expression levels of TNF-α were significantly
increased in the lung tissues. TNF-α shRNA protected lungs from
II/R-induced acute injury by upregulating IL-10 expression. The
circles above the arrows indicate inhibition. ALI, acute lung
injury; IF, immunofluorescence; II/R, intestinal ischemia and
reperfusion; IL-10, interleukin-10; qPCR, quantitative polymerase
chain reaction; shRNA, short hairpin RNA; TNF-α, tumor necrosis
factor-α; WB, western blotting. |
In conclusion, the present study demonstrated that
TNF-α may be a major contributor in II/R-induced ALI, and TNF-α
RNAi may alleviate the severity of ALI. Notably, TNF-α RNAi exerted
a protective effect on II/R-induced ALI via upregulation of the
anti-inflammatory cytokine IL-10. Based on these findings, TNF-α
knockdown may be considered a novel therapeutic strategy for the
treatment of II/R-induced ALI.
Acknowledgments
The authors would like to thank Dr Qing-Jie Xia
(West China Hospital, Sichuan University, Chengdu, China) for
suggestions on this study.
Funding
The present study was supported by a grant from the
Key Natural Science Foundation of Yunnan (grant no. 2013FZ264).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
ZY, XRZ, THW and YHZ designed the project and were
major contributors in writing and revising the manuscript. QZ and
SLW participated in the production of recombinant lentiviral
vector. LLX and PZ carried out the II/R model and lentivirus
injection. QZ, BY and ZBZ carried out the histological analysis.
QZ, SYF, ZY and XRZ carried out the qPCR, IF staining, and western
blot analysis. ZY, XRZ, THW and ZYH analyzed the data. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Animal care and all experimental protocols were
approved by the Institutional Medical Experimental Animal Care
Committee of Kunming Medical University (Kunming, China).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tadros T, Traber DL, Heggers JP and
Herndon DN: Effects of interleukin-1alpha administration on
intestinal ischemia and reperfusion injury, mucosal permeability,
and bacterial translocation in burn and sepsis. Ann Surg.
237:101–109. 2003. View Article : Google Scholar
|
|
2
|
Tendler DA: Acute intestinal ischemia and
infarction. Semin Gastrointest Dis. 14:66–76. 2003.
|
|
3
|
Higuchi S, Wu R, Zhou M, Marini CP,
Ravikumar TS and Wang P: Gut hyperpermiability after ischemia and
reperfusion: attenuation with adrenomedullin and its binding
protein treatment. Int J Clin Exp Pathol. 1:409–418. 2008.
|
|
4
|
Harward TR, Brooks DL, Flynn TC and Seeger
JM: Multiple organ dysfunction after mesenteric artery
revascularization. J Vasc Surg. 18:459–467. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cui T, Miksa M, Wu R, Komura H, Zhou M,
Dong W, Wang Z, Higuchi S, Chaung W, Blau SA, et al: Milk fat
globule epidermal growth factor 8 attenuates acute lung injury in
mice after intestinal ischemia and reperfusion. Am J Respir Crit
Care Med. 181:238–246. 2010. View Article : Google Scholar :
|
|
6
|
Bellingan GJ: The pulmonary physician in
critical care * 6: the pathogenesis of ALI/ARDS. Thorax.
57:540–546. 2002. View Article : Google Scholar
|
|
7
|
Souza DG, Vieira AT, Soares AC, Pinho V,
Nicoli JR, Vieira LQ and Teixeira MM: The essential role of the
intestinal microbiota in facilitating acute inflammatory responses.
J Immunol. 173:4137–4146. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tamion F, Richard V, Lyoumi S, Daveau M,
Bonmarchand G, Leroy J, Thuillez C and Lebreton JP: Gut ischemia
and mesenteric synthesis of inflammatory cytokines after
hemorrhagic or endotoxic shock. Am J Physiol. 273:G314–G321.
1997.PubMed/NCBI
|
|
9
|
Roten R, Markert M, Feihl F, Schaller MD,
Tagan MC and Perret C: Plasma levels of tumor necrosis factor in
the adult respiratory distress syndrome. Am Rev Respir Dis.
143:590–592. 1991. View Article : Google Scholar
|
|
10
|
Ito K, Ozasa H and Horikawa S: Edaravone
protects against lung injury induced by intestinal
ischemia/reperfusion in rat. Free Radic Biol Med. 38:369–374. 2005.
View Article : Google Scholar
|
|
11
|
Guzel A, Kanter M, Guzel A, Pergel A and
Erboga M: Anti-inflammatory and antioxidant effects of infliximab
on acute lung injury in a rat model of intestinal
ischemia/reperfusion. J Mol Histol. 43:361–369. 2012. View Article : Google Scholar
|
|
12
|
Narita K, Kuwabara Y and Fujii Y: Lung
injury after intestinal ischemia-reperfusion may be avoided by the
reduced absorption of locally produced cytokines. Surg Today.
34:937–942. 2004. View Article : Google Scholar
|
|
13
|
Caty MG, Guice KS, Oldham KT, Remick DG
and Kunkel SI: Evidence for tumor necrosis factor-induced pulmonary
micro-vascular injury after intestinal ischemia-reperfusion injury.
Ann Surg. 212:694–700. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim DH and Rossi JJ: Strategies for
silencing human disease using RNA interference. Nat Rev Genet.
8:173–184. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Elbashir SM, Harborth J, Lendeckel W,
Yalcin A, Weber K and Tuschl T: Duplexes of 21-nucleotide RNAs
mediate RNA interference in cultured mammalian cells. Nature.
411:494–498. 2001. View
Article : Google Scholar
|
|
16
|
Hall J: Unravelling the general properties
of siRNAs: strength in numbers and lessons from the past. Nat Rev
Genet. 5:552–557. 2004. View
Article : Google Scholar
|
|
17
|
Sorkine P, Setton A, Halpern P, Miller A,
Rudick V, Marmor S, Klausner JM and Goldman G: Soluble tumor
necrosis factor receptors reduce bowel ischemia-induced lung
permeability and neutrophil sequestration. Crit Care Med.
23:1377–1381. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Artigas A, Bernard GR, Carlet J, Dreyfuss
D, Gattinoni L, Hudson L, Lamy M, Marini JJ, Matthay MA, Pinsky MR,
et al: The American-European Consensus Conference on ARDS, part 2:
ventilatory, pharmacologic, supportive therapy, study design
strategies, and issues related to recovery and remodeling. Acute
respiratory distress syndrome. Am J Respir Crit Care Med.
157:1332–1347. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bosher JM and Labouesse M: RNA
interference: genetic wand and genetic watchdog. Nat Cell Biol.
2:E31–E36. 2000. View
Article : Google Scholar
|
|
20
|
Younis A, Siddique MI, Kim CK and Lim KB:
RNA interference (RNAi) induced gene silencing: a promising
approach of Hi-Tech plant breeding. Int J Biol Sci. 10:1150–1158.
2014. View Article : Google Scholar :
|
|
21
|
Liu R, Zhao W, Zhao Q, Liu SJ, Liu J, He
M, Xu Y, Wang W, Liu W, Xia QJ, et al: Endoplasmic reticulum
protein 29 protects cortical neurons from apoptosis and promoting
corticospinal tract regeneration to improve neural behavior via
caspase and Erk signal in rats with spinal cord transection. Mol
Neurobiol. 50:1035–1048. 2014. View Article : Google Scholar
|
|
22
|
Howard KA, Paludan SR, Behlke MA,
Besenbacher F, Deleuran B and Kjems J: Chitosan/siRNA
nanoparticle-mediated TNF-alpha knockdown in peritoneal macrophages
for anti-infla mmatory treatment in a murine arthritis model. Mol
Ther. 17:162–168. 2009. View Article : Google Scholar
|
|
23
|
Pei YH, Cai XM, Chen J, Sun BD, Sun ZR,
Wang X and Qian XM: The role of p38 MAPK in acute paraquat-induced
lung injury in rats. Inhal Toxicol. 26:880–884. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim K, Li Y, Jin G, Chong W, Liu B, Lu J,
Lee K, Demoya M, Velmahos GC and Alam HB: Effect of valproic acid
on acute lung injury in a rodent model of intestinal ischemia
reperfusion. Resuscitation. 83:243–248. 2012. View Article : Google Scholar
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
26
|
Sayan H, Ozacmak VH, Sen F, Cabuk M, Atik
DY, Igdem AA and Ozacmak ID: Pharmacological preconditioning with
erythropoietin reduces ischemia-reperfusion injury in the small
intestine of rats. Life Sci. 84:364–371. 2009. View Article : Google Scholar
|
|
27
|
Sukhotnik I, Slijper N, Pollak Y,
Chemodanov E, Shaoul R, Coran AG and Mogilner JG: Parenteral
omega-3 fatty acids (Omegaven) modulate intestinal recovery after
intestinal ischemia-reperfusion in a rat model. J Pediatr Surg.
46:1353–1360. 2011. View Article : Google Scholar
|
|
28
|
Grommes J and Soehnlein O: Contribution of
neutrophils to acute lung injury. Mol Med. 17:293–307. 2011.
View Article : Google Scholar :
|
|
29
|
Zhao C, Sun J, Fang C and Tang F:
1,8-Cineol attenuates LPS-induced acute pulmonary inflammation in
mice. Inflammation. 37:566–572. 2014. View Article : Google Scholar
|
|
30
|
Rinkema LE, Bemis KG and Fleisch JH:
Production and antagonism of cutaneous vascular permeability in the
guinea pig in response to histamine, leukotrienes and A23187. J
Pharmacol Exp Ther. 230:550–557. 1984.PubMed/NCBI
|
|
31
|
Fattal E and Bochot A: Ocular delivery of
nucleic acids: antisense oligonucleotides, aptamers and siRNA. Adv
Drug Deliv Rev. 58:1203–1223. 2006. View Article : Google Scholar
|
|
32
|
Oshitari T, Brown D and Roy S: siRNA
strategy against overexpression of extracellular matrix in diabetic
retinopathy. Exp Eye Res. 81:32–37. 2005. View Article : Google Scholar
|
|
33
|
Neurath MF, Fuss I, Pasparakis M,
Alexopoulou L, Haralambous S, Meyer zum Büschenfelde KH, Strober W
and Kollias G: Predominant pathogenic role of tumor necrosis factor
in experimental colitis in mice. Eur J Immunol. 27:1743–1750. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Myers KJ, Murthy S, Flanigan A, Witchell
DR, Butler M, Murray S, Siwkowski A, Goodfellow D, Madsen K and
Baker B: Antisense oligonucleotide blockade of tumor necrosis
factor-α in two murine models of colitis. J Pharmacol Exp Ther.
304:411–424. 2003. View Article : Google Scholar
|
|
35
|
Powrie F, Leach MW, Mauze S, Menon S,
Caddle LB and Coffman RL: Inhibition of Th1 responses prevents
inflammatory bowel disease in scid mice reconstituted with
CD45RBhi CD4+ T cells. Immunity. 1:553–562.
1994. View Article : Google Scholar
|
|
36
|
Hartmann G, Bidlingmaier C, Siegmund B,
Albrich S, Schulze J, Tschoep K, Eigler A, Lehr HA and Endres S:
Specific type IV phosphodiesterase inhibitor rolipram mitigates
experimental colitis in mice. J Pharmacol Exp Ther. 292:22–30.
2000.
|
|
37
|
Jakobsen M, Stenderup K, Rosada C, Moldt
B, Kamp S, Dam TN, Jensen TG and Mikkelsen JG: Amelioration of
psoriasis by anti-TNF-alpha RNAi in the xenograft transplantation
model. Mol Ther. 17:1743–1753. 2009. View Article : Google Scholar
|
|
38
|
Yin L, Song Z, Qu Q, Kim KH, Zheng N, Yao
C, Chaudhury I, Tang H, Gabrielson NP, Uckun FM, et al:
Supramolecular self-assembled nanoparticles mediate oral delivery
of therapeutic TNF-α siRNA against systemic inflammation. Angew
Chem Int Ed Engl. 52:5757–5761. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang Z, Ji Y, Wang S, Wang R, Li Z, Kang
A, Xu H, Shi M and Zhao M: Protective effect of intestinal ischemic
preconditioning on ischemia reperfusion-caused lung injury in rats.
Inflammation. 38:424–432. 2015. View Article : Google Scholar
|
|
40
|
Mao YF, Zheng XF, Cai JM, You XM, Deng XM,
Zhang JH, Jiang L and Sun XJ: Hydrogen-rich saline reduces lung
injury induced by intestinal ischemia/reperfusion in rats. Biochem
Biophys Res Commun. 381:602–605. 2009. View Article : Google Scholar
|
|
41
|
Zhao X, Town JR, Yang A, Zhang X, Paur N,
Sawicki G and Gordon JR: A novel ELR-CXC chemokine antagonist
reduces intestinal ischemia reperfusion-induced mortality, and
local and remote organ injury. J Surg Res. 162:264–273. 2010.
View Article : Google Scholar
|
|
42
|
James IA, Chen CL, Huang G, Zhang HY,
Velten M and Besner GE: HB-EGF protects the lungs after intestinal
ischemia/reperfusion injury. J Surg Res. 163:86–95. 2010.
View Article : Google Scholar
|