Introduction
The blood-brain barrier (BBB), as a membrane barrier
in the central nervous system (CNS) that separates the brain from
circulating blood, is vital to maintain neural microenvironment
homeostasis and low permeability, which is closely associated with
CNS disease (1). Endothelial
cells are not only the primary components of the BBB, but they are
also important in the BBB. Angiogenesis is critical in the
formation of new blood vessels within this brain microvessel
network (2). The initiation of
angiogenesis and the formation of early vascular structures depend
on endothelial cells (3), which
are essential for maintaining the function and regulating the
proliferation of vessels. Endothelial cells are also important in
damage repair, ischemic adaptation and tumor formation (4,5).
However, changes in the function and structure of the vascular wall
occur with increasing age or through stimuli from other
environmental factors, including hypertension, smoking and
drinking. This leads to the compromised ability of endothelial
cells in proliferation, migration and damage repair (6,7),
which results in disrupted vascular hemostasis and vessel
aging.
It is accepted that the dynamic reorganization of
the actin cytoskeleton is important in cellular morphogenesis and
motility. The phosphatidylinositol 3-kinase (PI3K)-AKT pathway
exerts functions downstream of growth factors in facilitating
endothelial cell growth, and its downstream targets are involved in
vascular remodeling (8-10). Girders of actin filament (girdin),
as an actin-binding protein in mammalian tissues, is expressed in
limited cell types, including immature endothelial cells, which can
be phosphorylated by AKT at serine 1417 (11,12). It has been reported that girdin is
important in cell migration, which relies on extracellular signals
and the surrounding microenvironment. Therefore, it is of value to
delay the aging of endothelial cells and modulate the relevant
molecule signaling for the prevention of CNS disease.
Platelets (PLTs), as fragments of cytoplasm derived
from megakaryocytes, are components of the blood in mammals
(13). The importance of PTLs in
the inflammatory response has been reported. In addition, extensive
evidence has revealed that PLTs are essential in vascular biology,
including angiogenesis and tissue regeneration (14,15). Through various angiogenic
stimulators, PLTs are capable of promoting de novo vessel
growth. In the presence of vascular endothelial growth factor and
platelet-derived growth factor, the proliferation of endothelial
cells can be induced and their migration can be increased (16-18). Previously, attributable to
concentrated biologically active molecules, platelet-rich plasma
(PRP), as PLTs concentrate, has become more common as a clinical
treatment to induce healing and regeneration, and control
inflammation (19-21). Furthermore, exosome derived from
PLTs has attracted wide attention owing to its role in angiogenesis
and neovascularization (22,23). PLTs can be engulfed by endothelial
cells (24), however, the precise
effect of internalized PLTs on endothelial cells remains to be
elucidated.
Multiple BBB in vitro models have been
established, including monoculture, co-culture and triple culture
systems. These systems have their respective advantages and
disadvantages (25-27). In the present study, human brain
microvascular endothelial cells (HBMECs) were used to mimic the BBB
owing to its brain origin. Therefore, this model offers a more
representative BBB model compared to several other cell lines
currently used (26). In the
present study, the effects of internalized PLTs on HBMEC
senescence, proliferation, apoptosis, invasion and migration were
determined, and the associated mechanisms were established in
vitro. This information may provide a novel method for delaying
the aging of endothelial cells, inspiring a potential strategy for
angiogenesis in the treatment of CNS diseases.
Materials and methods
Cell culture and treatment
The HBMECs (ScienCell Research Laboratories,
Carlsbad, CA, USA) were cultured with endothelial cell medium
(ScienCell Research Laboratories) containing 10% fetal bovine serum
(FBS) and 1% endothelial cell growth supplement (ScienCell Research
Laboratories) in a 5% CO2 humidified incubator at 37°C.
The HBMECs (5×104 cells per cm2) were seeded
on a collagen/fibronectin-coated Transwell insert. Subsequently,
human astrocytes (HAs) (ScienCell Research Laboratories) were
seeded (5×104 cells per cm2) onto the carrier
plates in the Transwell and cultured with astrocyte medium
(ScienCell Research Laboratories). The medium was replaced with
fresh medium every other day. After 3 days, the insert coated with
HBMECs was added to dishes containing the HAs. Lipofectamine 3000
(cat. no. L3000008; Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) was used for cell transfection. Short hairpin
(sh)-girdin plasmid (cat. no. sc-94984-SH) and control shRNA
plasmid (cat. no. sc-108060) were purchased from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). Mutated girdin was mutated
at Ser-1417 to Ala, which was resistant to phosphorylation, using a
site-directed mutagenesis kit (Tiangen Biotech Co., Ltd., Beijing,
China). Treatment with triciribine (Merck Millipore, Darmstadt,
Germany) at 20 µM for 48 h was performed to inhibit AKT
signals (28).
Isolation and labeling of PLTs
Healthy volunteers (8 men and 8 women) with an
average age of 30.2±12.6 years, were recruited at Beijing Hospital
(Beijing, China) in August 2014, with the approval of the Ethics
Committee of Beijing Hospital. Informed consent was obtained from
the volunteers. The PLTs were isolated from the whole blood of
volunteers using a platelet preparation kit (cat. no. CS0257;
Leagene Biotech Co., Ltd., Beijing, China). Briefly, 30 ml venous
blood was collected and mixed with anticoagulant. The blood was
then transferred to a centrifuge for 15 min at 2,000 × g at 4°C.
The upper supernatant was carefully collected and the
centrifugation was repeated to acquire the PLTs. The PLTs were
washed with 100 nM PGE-1 (Sigma-Aldrich; Merck Millipore) and 1.9
mM theophylline. The PLTs were labeled with PKH-26 according to the
manufacturer's protocol (Mini 26; Sigma; Merck Millipore).
According to a previous study (29), the PLTs were added to the medium
at an HBMEC:PL ratio of 1:40, and were incubated for 20 h at
37°C.
β-galactosidase (β-gal) staining
The activity of senescence-associated (SA)-β-gal was
detected with a β-gal staining kit (cat. no. C0602; Beyotime
Institute of Biotechnology, Haimen, China). Briefly, the cells were
fixed in 0.2% glutaraldehyde solutions for 5 min, following which
the cells were stained with X-gal solution for 18 h at 37°C. The
SA-β-gal-positive cells were observed under a phase-contrast
microscope at ×100 or ×400 magnification (Olympus, Tokyo, Japan).
The proportions of cells positive for SA-β-gal activity were
determined as a percentage of the total number of cells counted in
each dish.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay
An MTT kit (Beyotime Institute of Biotechnology) was
used to detect cell viability. The cells (2×103 cells
per well) were seeded into a 96-well plate (Corning Costar,
Cambridge, MA, USA). After 24 h, the cells were treated with PLTs
alone or with PLTs plus cytochalasin D (1 µg/ml,) for 14 h
at 37°C, following which 100 µg MTT was added to the wells.
The incubation with cytochalasin D was 30 min only. The medium was
removed following incubation for 4 h. Subsequently, 150 µl
dimethyl sulfoxide (DMSO) was added to dissolve the purple formazan
formed by the reduction of MTT. The absorbance at a 490 nm was
measured with a microplate reader.
Flow cytometry for detection of the
engulfment of PLTs
The engulfment of fluorescent PKH26-labeled PLTs by
endothelial cells was determined using flow cytometry. Briefly, the
cells were incubated in HEPES-buffer, comprising 134 mM NaCl, 6 mM
KCl, 2 mM CaCl2, 1 mM MgCl2, 10 mM HEPES and
10 mM glucose (pH 7.40), and anti-PLT antibody (AIP21; cat. no.
ab112238; Abcam, Cambridge, MA, USA; 100 nM) for 30 min at 4°C,
followed by a 15-min wash with 1X PBS buffer. Flow cytometric
analysis (BD FACSCalibur; BD Biosciences, Franklin Lakes, NJ, USA)
was used to analyze the mean fluorescence intensity.
Flow cytometric analysis of
apoptosis
Apoptosis was determined using an Annexin V/PI
apoptosis kit (MultiSciences Biotech Co., Ltd., Hangzhou, China).
The cells were harvested and centrifuged at 2,000 × g at 4°C for 5
min. Following collection, the cells were resuspended in 0.5 ml 1X
annexin-binding buffer at a density of 5×105 cells/ml.
Annexin V-FITC and PI were added and the cells were incubated for
10 min at room temperature in the dark. The samples were
immediately analyzed by flow cytometry.
Transmission electron microscopic
observation
In brief, the cells were fixed in 4%
glutaraldehyde/1% paraformaldehyde for 2 h at room temperature, and
then post-fixed in 1% OsO4 for 1 h at 4°C. Following
dehydration in graded ethanol (70, 95 and 100%), the cells were
embedded in LX112. Ultrathin sections (50 nm) were stained with 1%
uranyl acetate and lead citrate, and were then examined for
mitochondrial morphology on a H7000 electron microscope operating
at 80 kV (Hitachi, Ltd., Tokyo, Japan).
Confocal microscopy
The cell sections were incubated with 3% hydrogen
peroxide for 10 min. Normal goat serum (Beijing Solarbio Science
& Technology Co., Ltd., Beijing, China) was added to the
sections and incubated at room temperature for 2 h. Subsequently,
the sections were incubated with anti-PLT antibody (100 nM; cat.
no. ab53304; Abcam), anti-girdin antibody (1:500; cat. no.
ab179481; Abcam), or anti-girdin (phosphorylated S1417) antibody (8
µg/ml; cat. no. 28067; Takara Bio, Inc., Otsu, Japan) at 4°C
overnight. The secondary antibodies, FITC-donkey anti-rabbit IgG
(1:200; JacksonImmuno Research Laboratories, Inc., West Grove, PA,
USA) and anti-rabbit IgG (H+L) F(ab')2 Fragment (1:500; Alexa
Fluor® 488 Conjugate; cat. no. 4412; Cell Signaling
Technology, Inc., Danvers, MA, USA) were added and incubated at 4°C
overnight. Finally, the sections were mounted with FlourSave
(Calbiochem; Merck Millipore) mounting reagent. Images of the
histological sections were captured using Northern Eclipse software
version 6.0 (Empix Imaging, Inc., Mississauga, ON, Canada) on a
Zeiss Axioplan 2 imaging microscope (Carl Zeiss, Toronto, ON,
Canada).
Transwell assay
A precoated cell invasion kit (pore size
8.0-µm; Corning Costar) and Matrigel (250 µg/ml; BD
Biosciences) in Transwell chambers were used in Transwell assays.
The cells were plated at a density of 5×104/ml in the
upper chambers without FBS. The cells migrated to the lower chamber
containing medium with 30% FBS. The invaded cells were fixed with
paraformaldehyde after 60 h incubation and stained with 0.1%
crystal violet. The numbers of cells that invaded through the
membrane were counted under a light microscope (Nikon Ni-E; Nikon
Corporation, Tokyo, Japan) in four randomly selected fields per
well.
Western blot analysis
The samples were incubated with an NP40 lysis buffer
(Beijing Solarbio Science & Technology Co., Ltd.) with protease
inhibitors (Promega Corporation, Madison, WI, USA). Then the
concentration of the protein samples was estimated using a
bicinchoninic acid protein quantitation kit (Thermo Fisher
Scientific, Inc.). Protein extract (35 µg per group) was
separated on 8% SDS-polyacrylamide gels and transferred onto a
nitrocellulose membrane. The membrane was blocked in 5% skim milk
powder solution with 0.1% Tris-buffered saline/Tween 20 at room
temperature for 2 h. The membrane was then incubated with
anti-active caspase-3 antibody (1 µg/ml; cat. no. ab2302;
Abcam), anti-AKT1/2/3 (phospho Y315+Y316+Y312) antibody (1:500;
cat. no. ab131443), anti-pan-AKT antibody (1 µg/ml; cat. no.
ab85683), anti-girdin antibody (1:40; cat. no. 18979; Takara Bio,
Inc.), anti-girdin (phospho S1417) antibody (1:50; cat. no. 28067;
Takara Bio, Inc.) or anti-GAPDH (6C5; 1:500; cat. no. ab8245;
Abcam) overnight at 4°C. The target bands were detected using a
horseradish peroxidase-conjugated secondary antibody (1:2,000; cat.
no. ab6721; Abcam) by incubation for 1 h at room temperature and
then visualized using a BeyoECL plus enhanced chemiluminescence
detection system (Beyotime Institute of Biotechnology). The band
intensity was normalized to that of GAPDH and expressed as a
relative ratio.
Statistical analysis
Statistical analysis was performed with SPSS 17.0
statistical software (SPSS, Inc., Chicago, IL, USA). For
comparisons between groups, Student's t-test or one-way analysis of
variance followed by Dunnet's multiple comparison test was used.
P<0.05 was considered to indicate a statistically significant
difference.
Results
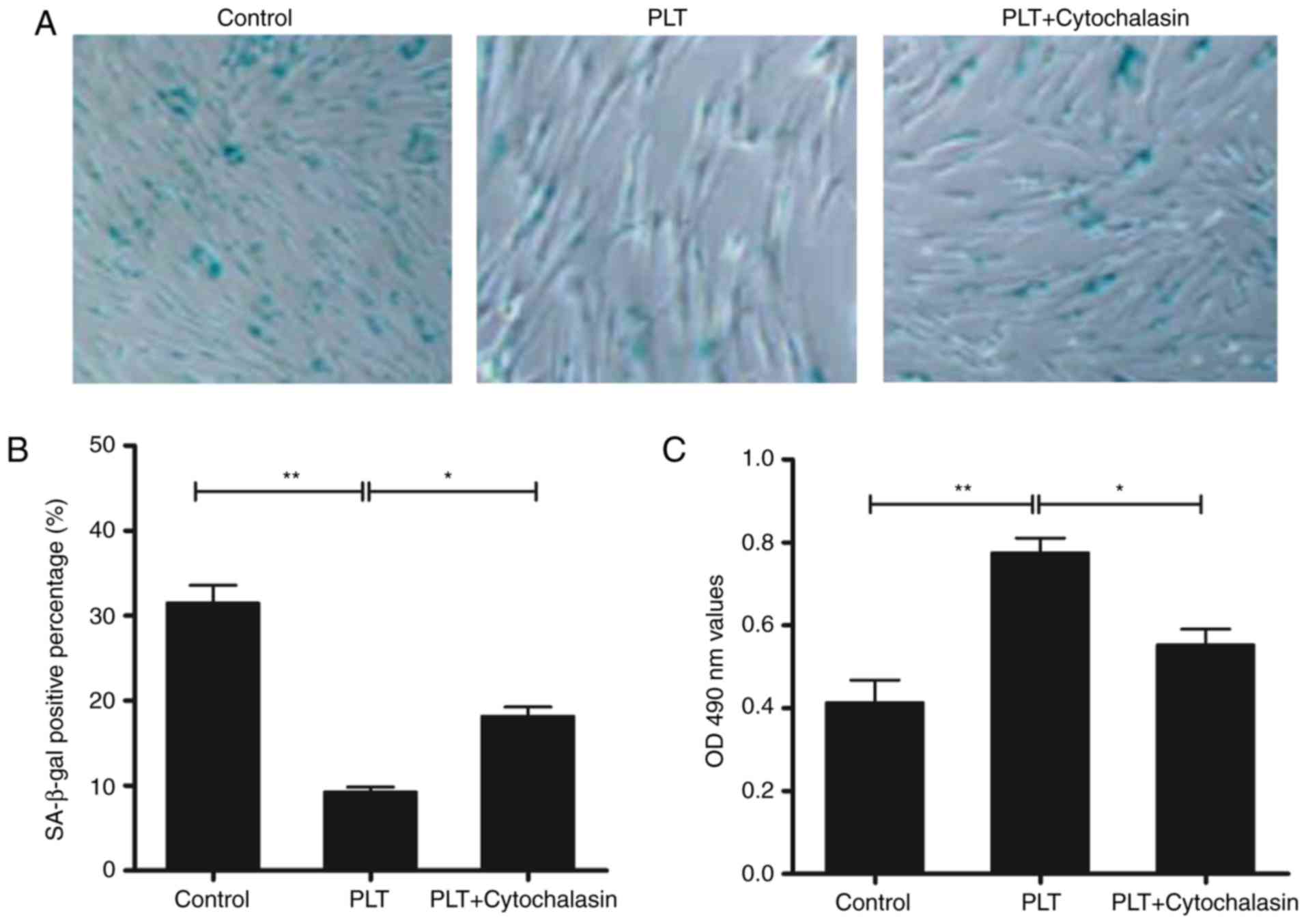
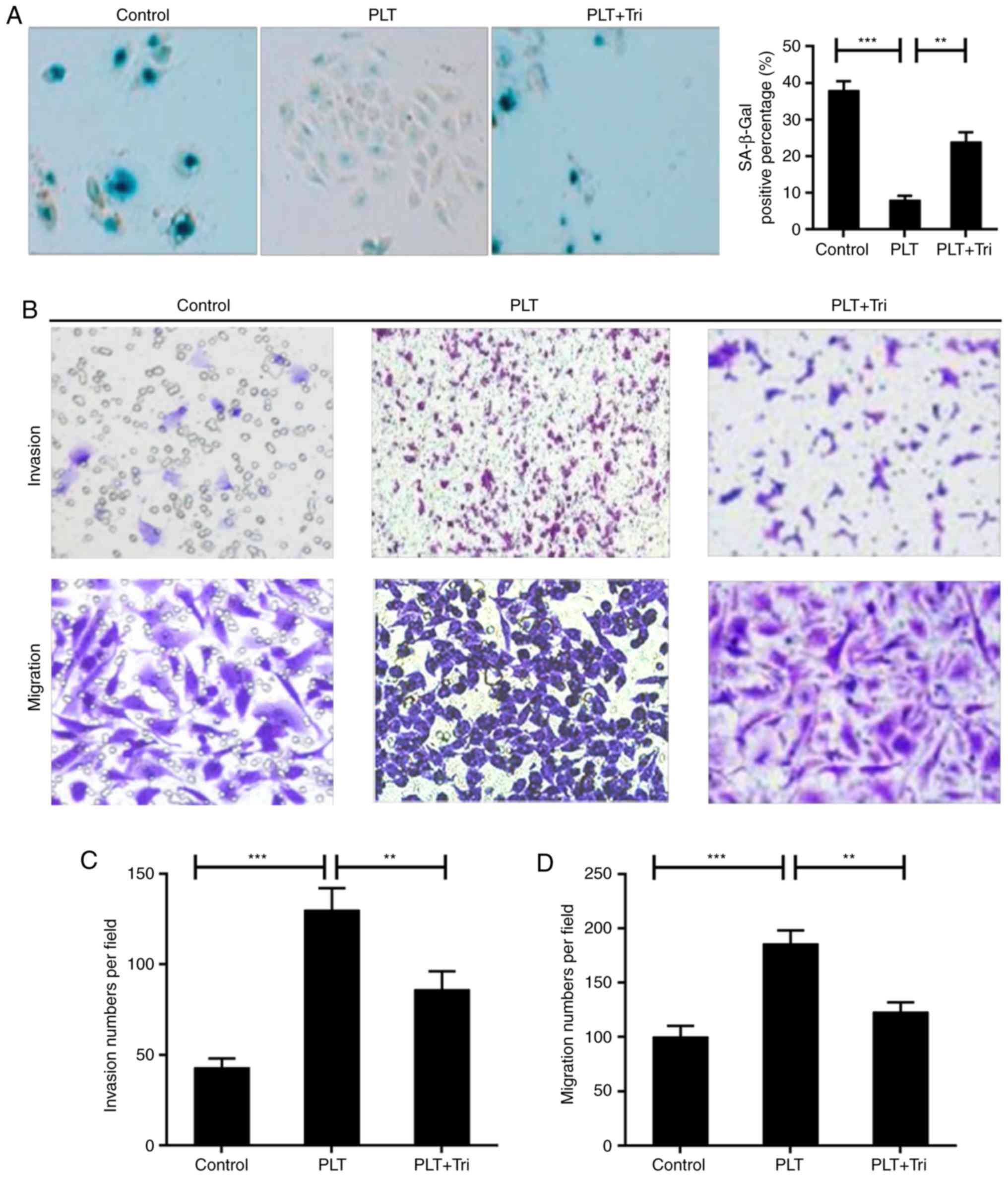
PLTs promotes resistance to cell
senescence
The expression of SA-β-gal can be induced in
senescent cells. As shown in Fig. 1A
and B, the activity of SA-β-gal was reduced by PLT treatment.
Cytochalasins can alter cell morphology by inhibiting actin
polymerization. Following treatment with cytochalasin D, the
decline in activity was recovered partially. In addition, the cell
viability was increased in the PLT group; and was decreased by
cytochalasin D (Fig. 1C).
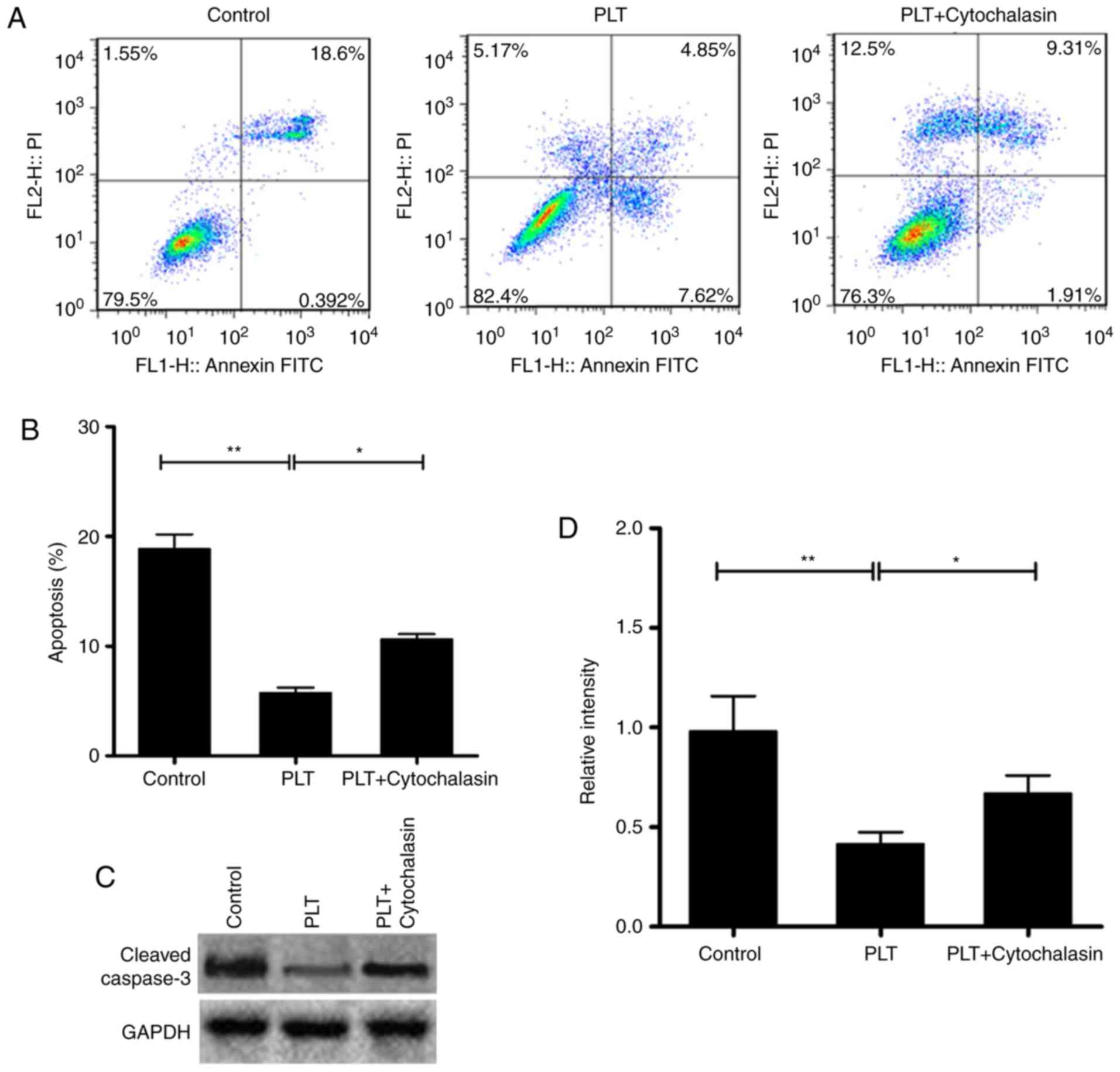
PLTs depress the apoptosis of HBMECs
The proliferation of cells can be inhibited by
senescence or apoptosis. The apoptosis of HBMECs was determined in
the present study. The results revealed that the level of apoptosis
was decreased significantly in the PLT group, compared with that in
the control group. By contrast, the number of apoptotic cells was
higher in the cytochalasin D+PLT group, compared with that in the
PLT group (Fig. 2A and B). In
addition, the expression of cleaved caspase-3 was suppressed in the
PLT group, compared with the control. Cytochalasin D recovered the
protein level of cleaved caspase-3 (Fig. 2C and D).
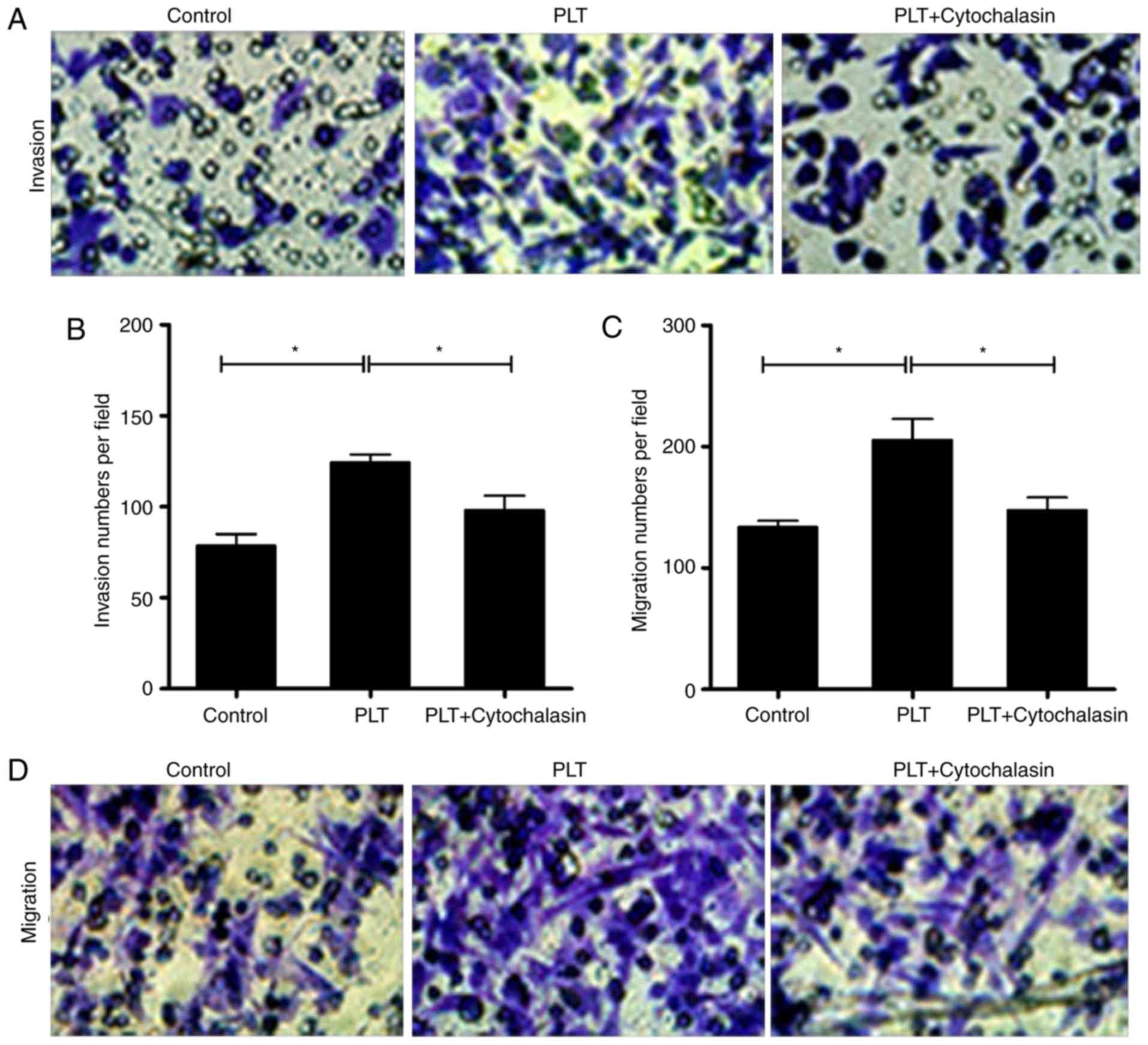
PLTs enhance the invasion and migration
capability of HBMECs
Invasion and migration are the essential motilities
of endothelial cells (30). The
Transwell assays revealed that the invasion and migration abilities
of the cells were increased by PLTs, whereas cytochalasin D
repressed the cell invasion and migration induced by PLTs (Fig. 3A–D).
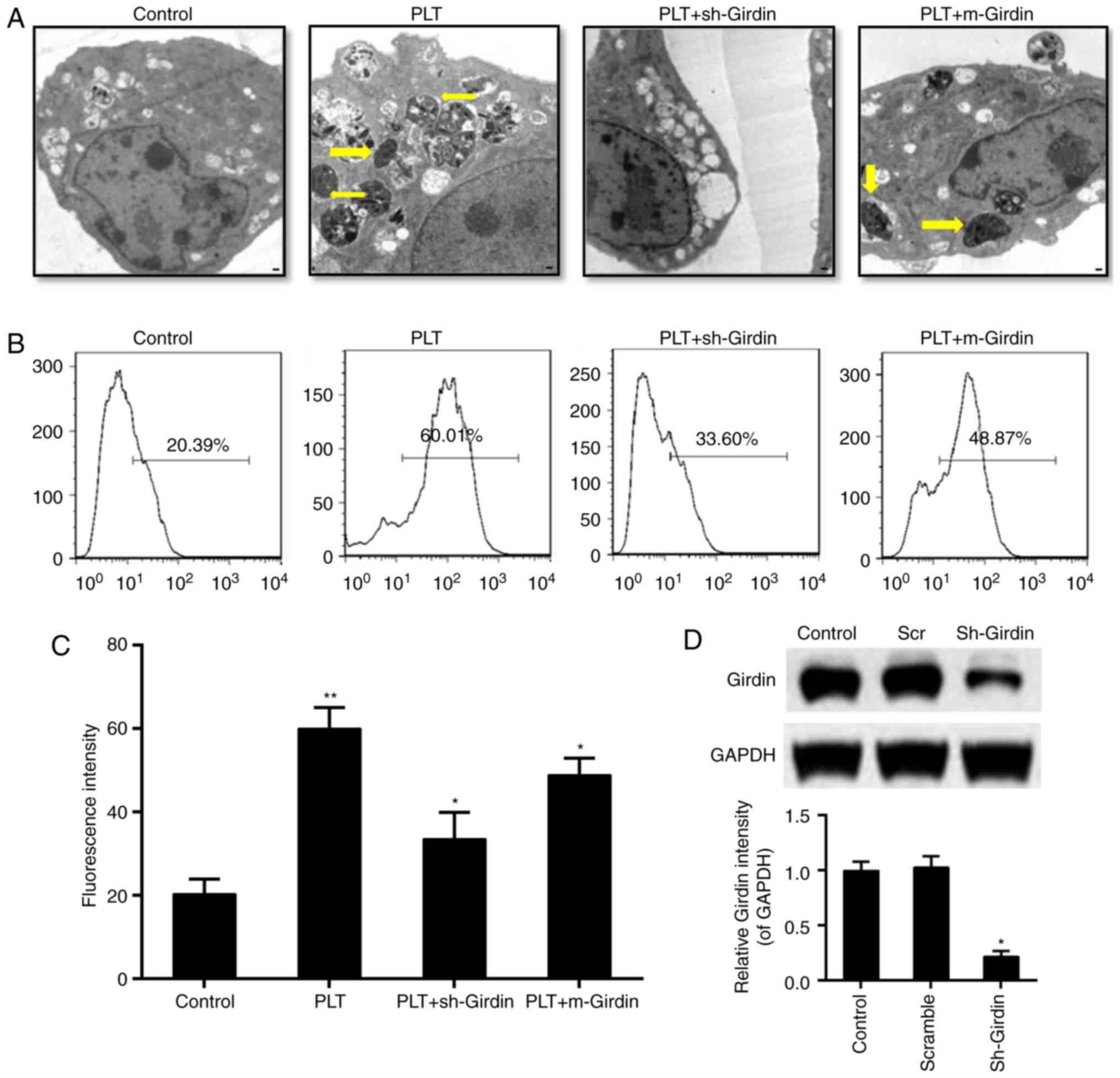
Effect of girdin on the engulfment of
HBMECs to PLTs
The results from the transmission electron
microscopy showed that, compared with the control group, knockdown
of the expression of girdin inhibited the engraftment of
endothelial cells to PLTs; and the mutated girdin at the 1417 locus
also inhibited the engraftment of PLTs (Fig. 4A). In addition, the fluorescent
intensity of PLT-labeling was reduced by the knockdown of girdin or
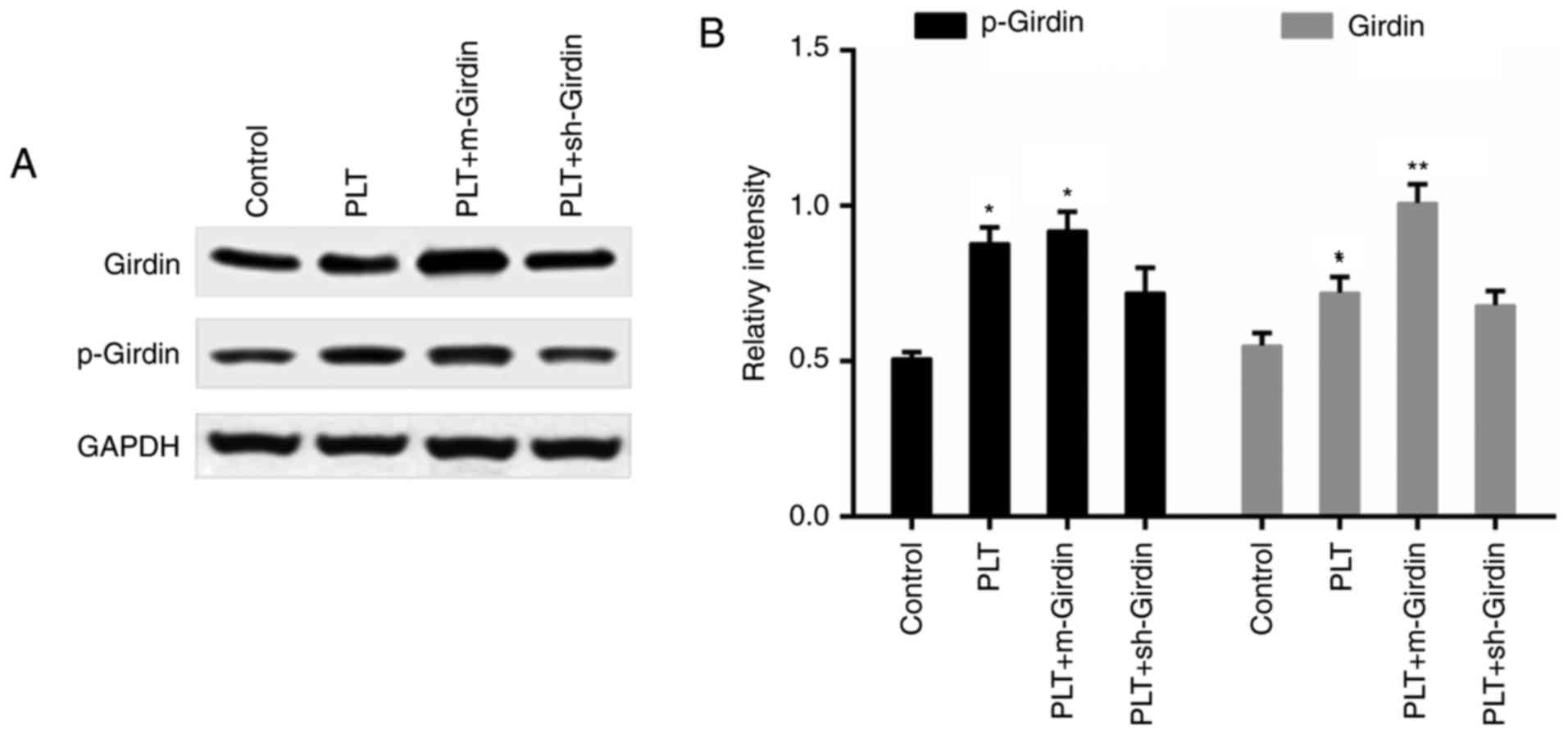
the mutated girdin, compared with that in the PLT group (Fig. 4B and C). The downregulation of
girdin was confirmed by western blot analysis (Fig. 4D). Furthermore, the engulfment of
PLTs promoted the expression of girdin and p-girdin; whereas the
expression of p-girdin was inhibited by the knockdown of girdin,
while the expression of p-girdin did not appear to be influenced by
m-girdin (Fig. 5A and B).
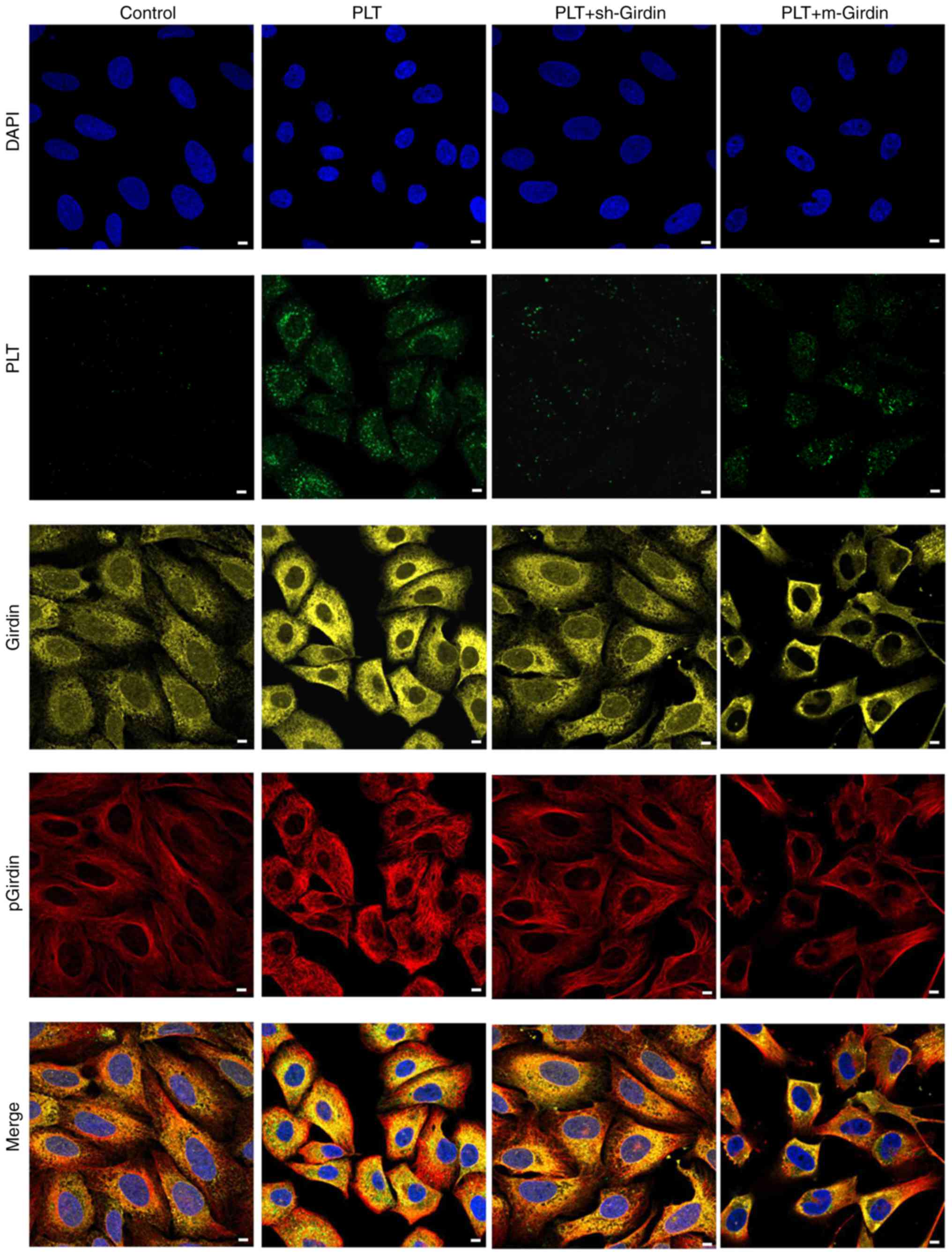
Girdin is co-located with PLTs in
HBMECs
An immunofluorescence assay was performed to confirm
the effect of girdin on the engulfment of PLTs. As shown in
Fig. 6, girdin and p-girdin were
located in the Golgi apparatus, and green fluorescence indicated
phagocytic PLTs. Compared with the PLT group, the knockdown of
girdin or the mutated girdin prevented the engulgment of PLTs.
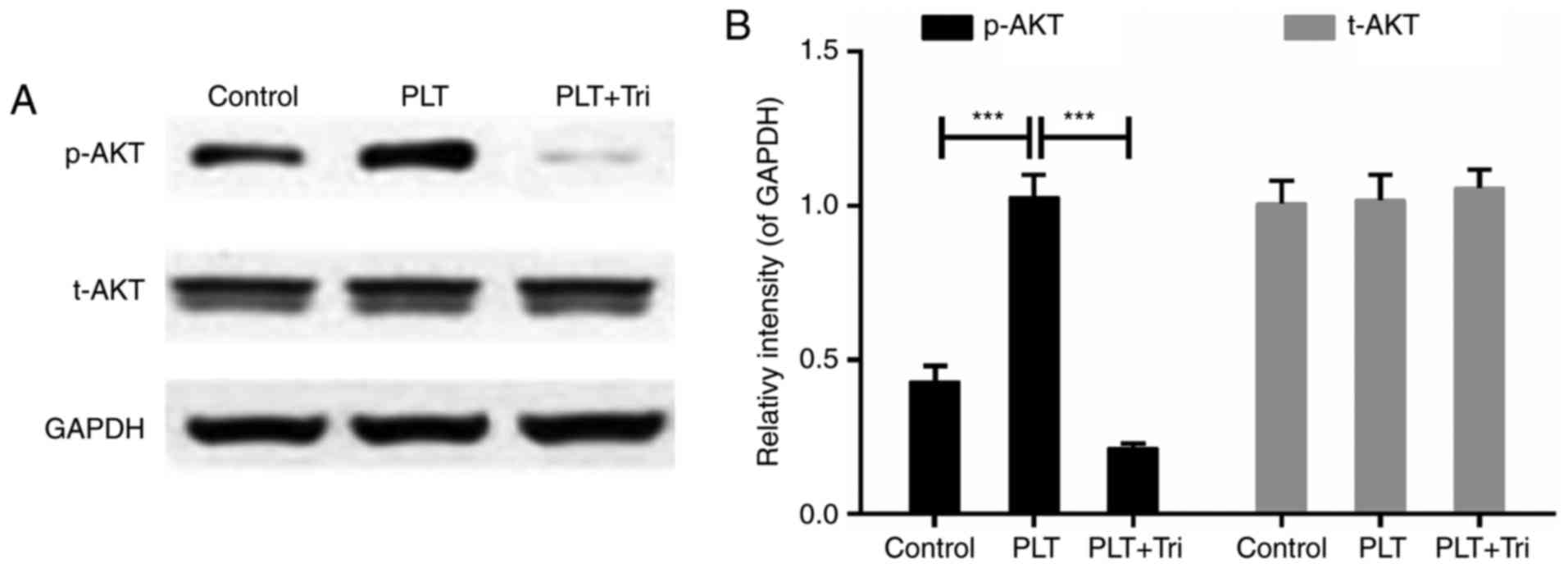
Akt signals are involved in the
engulfment of PLTs
It has been reported that girdin is a substrate of
AKT/PKB, which is reported to be positively associated with
phagocytosis (31). Triciribine,
a specific inhibitor of AKT phosphorylation (32), was used in the present study to
further determine the involvement of AKT. The inhibition of
senescence of HBMECs caused by PLTs was reversed by triciribine
(Fig. 7A). In addition, the cell
invasion and migration capabilities were decreased by triciribine,
compared with those in the PLT group (Fig. 7B–D). Mechanically, p-AKT was
increased in PLT group, whereas treatment with triciribine
significantly inhibited the activity of AKT (Fig. 8A and B). This indicated that the
activation of AKT signals was involved in the engulfment of
PLTs.
Discussion
BBB is a well-differentiated microvessel network in
the CNS and important to the hemostasis of brain tissues. Brain
angiogenesis is a critical step in the development of the BBB,
which is dependent on the invasion of endothelial cells into the
avascular neuroectoderm and formation of intraneural vessels
(33,34). It is promising for the protection
of endothelial cells from injury and aging. In the present study,
co-cultures of HAs and HBMECs in vitro were used to mimic
the BBB in living cells. It was shown that PLTs promoted the
resistance to senescence of HBMECs, whereas this effect was
mitigated by treatment with cytochalasin D. Although the effect of
extracellular PLTs cannot completely excluded, it was suggested
that the survival of HBMECa was partly promoted by the phagocytosis
of PLTs. The apoptosis of HBMECs was further examined, and the
results revealed that the level of apoptosis was significantly
decreased by PLTs. In addition, the expression of cleaved
caspase-3, an apoptosis-associated protein (35), was inhibited by PLTs. These
results indicated that treatment with PLTs increased proliferation
and depressed endothelial apoptosis. This was consistent with
previous reports that PLTs enhance the growth and integrity of
endothelial cells (36,37). The invasion and migration of
endothelial cells is an important event in angiogenesis (38). To further confirm the effect of
PLTs, a Transwell assay was performed. It was shown that the PLTs
augmented the invasion and migration abilities of the endothelial
cells. Taken together, treatment with PLTs was beneficial for the
healthy situation of endothelial cells. Consistently, several
studies have confirmed that PRP exerts protective effect via
anti-inflammatory and anti-apoptotic effects, and by promoting
angiogenesis (20,39). However, platelet aggregation and
abnormal angiogenesis are considered to be harmful to human health
(40-42). The concentration used in the
present study may be not the optimal concentration for delaying
cell aging, and further investigations are warranted for its
potential clinical use. It is important to regulate the
phagocytosis of PLTs in an appropriate manner.
PLTs can be engulfed by endothelial cells (24,29). The phagocytosis of PLTs was
observed under a transmission electron microscopic. It was shown
that the PLTs were engulfed by HBMECs. Girdin, as an actin-binding
AKT substrate, is able to regulate angiogenesis (12). Girdin is also important for the
migration of endothelial cells during postnatal angiogenesis
(11,12). Therefore, it is possible that
girdin may be associated with the protective effect of PLTs on
endothelial cells. As expected, the depletion of girdin and mutated
girdin (resistant to phosphorylation at 1417) suppressed the
phagocytosis of PLTs by HBMECs. In addition, the expression of
girdin and p-girdin were induced by PLTs. The co-localization of
PLTs, girdin and p-girdin further demonstrated the potential role
of girdin in the positive effect of PLTs on endothelial cells. As
the reduction of girdin and p-girdin reduced the phagocytosis of
PLTs, the depletion of girdin may have a negative feedback effect
on the phagocytosis in endothelial cells of PLTs.
In consideration of the regulatory association
between girdin and AKT, the involvement of AKT signals in the
present study model was investigated using its specific inhibitor.
The results confirmed that the repression of AKT signals reversed
the effect of PLTs through the aggravation of senescence and the
inhibition of invasion and migration of HBMECs. It was shown that
phagocytosis of the PLTs induced the activation of AKT signals and
thus contributed to cell growth and survival. This result was in
line with previous reports (36,43). In addition, girdin can be
phosphorylated by AKT at serine 1417. Therefore, the
phosphorylation of girdin mediated by AKT may be the mechanism by
which phagocytosis of PLTs promoted the survival of endothelial
cells. This result was compatible with a previous study that
highlighted the major effect of girdin and its Akt-mediated
phosphorylation on migration and proliferation following vascular
injury (19). Girdin is also
associated with Wnt signaling pathways (44), indicating the versatile function
of girdin in different signaling pathways. The intracellular
signaling pathways are complex and there may be cross-talk,
therefore, it is possible that other signals or pathways are
involved in the phagocytosis of PLTs. For example, forkhead box 3a
(Fox3a), another downstream target of the AKT signal, has been
shown to be essential in angiogenesis (45); additionally, the extracellular
signal-regulated kinase (ERK) pathway is another important
signaling pathway for cell survival, angiogenesis and phagocytosis
(46-48). Although the role of other signals,
including the Fox3a and ERK pathways, were not elucidated in the
present study, they may be modulated following the phagocytosis of
PLTs and may be potential targets of novel therapeutics for
delaying the aging of endothelial cells.
In conclusion, it was shown that the engulfment of
PLTs delayed endothelial cell aging via the promotion of cell
proliferation and inhibition of apoptosis, and the enhancement of
cell invasion and migration abilities. Girdin was essential for the
phagocytosis of PLTs, in which AKT may be involved in the
phosphorylation of girdin. The results of the present study
indicated the significance of understanding the effect of PLTs on
endothelial cells, which may assist in developing novel
therapeutics for CNS diseases associated with the aging of
endothelial cells.
Acknowledgments
Not applicable.
Funding
This study was supported by the Natural Science
Foundation of China (grant no. 31371160).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YLa, YoLi and DL wrote the manuscript. YLa, YoLi,
DL, PL, JW, YD and GY performed the experiments. YLa and YaLi
designed the study. YLa, YoLi, DL, PL and JW performed the data
analysis. YLa, YoLi, and DL revised the manuscript and all authors
reviewed the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Beijing Hospital (Beijing, China). Informed consent
was obtained from the volunteers.
Consent for publication
Informed consent was obtained from all participants
for the publication of their data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yang Y and Rosenberg GA: Blood-brain
barrier breakdown in acute and chronic cerebrovascular disease.
Stroke. 42:3323–3328. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee SW, Kim WJ, Choi YK, Song HS, Son MJ,
Gelman IH, Kim YJ and Kim KW: SSeCKS regulates angiogenesis and
tight junction formation in blood-brain barrier. Nat Med.
9:900–906. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lahteenvuo J and Rosenzweig A: Effects of
aging on angiogenesis. Circ Res. 110:1252–1264. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Capettini LS, Cortes SF, Silva JF,
Alvarez-Leite JI and Lemos VS: Decreased production of neuronal
NOS-derived hydrogen peroxide contributes to endothelial
dysfunction in atherosclerosis. Br J Pharmacol. 164:1738–1748.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pelham CJ, Keen HL, Lentz SR and Sigmund
CD: Dominant negative PPARgamma promotes atherosclerosis, vascular
dysfunction, and hypertension through distinct effects in
endothelium and vascular muscle. Am J Physiol Regul Integr Comp
Physiol. 304:R690–R701. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Erusalimsky JD and Skene C: Mechanisms of
endothelial senescence. Exp Physiol. 94:299–304. 2009. View Article : Google Scholar
|
|
7
|
Rivard A, Fabre JE, Silver M, Chen D,
Murohara T, Kearney M, Magner M, Asahara T and Isner JM:
Age-dependent impairment of angiogenesis. Circulation. 99:111–120.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fujio Y and Walsh K: Akt mediates
cytoprotection of endothelial cells by vascular endothelial growth
factor in an anchorage-dependent manner. J Biol Chem.
274:16349–16354. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shiojima I and Walsh K: Role of Akt
signaling in vascular homeostasis and angiogenesis. Circ Res.
90:1243–1250. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sata M, Maejima Y, Adachi F, Fukino K,
Saiura A, Sugiura S, Aoyagi T, Imai Y, Kurihara H, Kimura K, et al:
A mouse model of vascular injury that induces rapid onset of medial
cell apop-tosis followed by reproducible neointimal hyperplasia. J
Mol Cell Cardiol. 32:2097–2104. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Enomoto A, Murakami H, Asai N, Morone N,
Watanabe T, Kawai K, Murakumo Y, Usukura J, Kaibuchi K and
Takahashi M: Akt/PKB regulates actin organization and cell motility
via Girdin/APE. Dev Cell. 9:389–402. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kitamura T, Asai N, Enomoto A, Maeda K,
Kato T, Ishida M, Jiang P, Watanabe T, Usukura J, Kondo T, et al:
Regulation of VEGF-mediated angiogenesis by the Akt/PKB substrate
Girdin. Nat Cell Biol. 10:329–337. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Angiolillo DJ, Ferreiro JL, Price MJ,
Kirtane AJ and Stone GW: Platelet function and genetic testing. J
Am Coll Cardiol. 62(Suppl 17): S21–S31. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
de Groot PG, Urbanus RT and Roest M:
Platelet interaction with the vessel wall. Handb Exp Pharmacol.
210:87–110. 2012. View Article : Google Scholar
|
|
15
|
Sopova K, Tatsidou P and Stellos K:
Platelets and platelet interaction with progenitor cells in
vascular homeostasis and inflammation. Curr Vasc Pharmacol.
10:555–562. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sharma D, Brummel-Ziedins KE, Bouchard BA
and Holmes CE: Platelets in tumor progression: A host factor that
offers multiple potential targets in the treatment of cancer. J
Cell Physiol. 229:1005–1015. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Radziwon-Balicka A, Moncada de la Rosa C
and Jurasz P: Platelet-associated angiogenesis regulating factors:
A pharmacological perspective. Can J Physiol Pharmacol. 90:679–688.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Andrae J, Gallini R and Betsholtz C: Role
of platelet-derived growth factors in physiology and medicine.
Genes Dev. 22:1276–1312. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dohan Ehrenfest DM, Andia I, Zumstein MA,
Zhang CQ, Pinto NR and Bielecki T: Classification of platelet
concentrates (Platelet-Rich Plasma-PRP, Platelet-Rich Fibrin-PRF)
for topical and infiltrative use in orthopedic and sports medicine:
Current consensus, clinical implications and perspectives. Muscles
Ligaments Tendons J. 4:3–9. 2014.PubMed/NCBI
|
|
20
|
Kakudo N, Morimoto N, Kushida S, Ogawa T
and Kusumoto K: Platelet-rich plasma releasate promotes
angiogenesis in vitro and in vivo. Med Mol Morphol. 47:83–89. 2014.
View Article : Google Scholar
|
|
21
|
Marx RE: Platelet-rich plasma: Evidence to
support its use. J Oral Maxillofac Surg. 62:489–496. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hayon Y, Dashevsky O, Shai E, Brill A,
Varon D and Leker RR: Platelet microparticles induce angiogenesis
and neurogenesis after cerebral ischemia. Curr Neurovasc Res.
9:185–192. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ohtsuka M, Sasaki K, Ueno T, Seki R,
Nakayoshi T, Koiwaya H, Toyama Y, Yokoyama S, Mitsutake Y, Chibana
H, et al: Platelet-derived microparticles augment the adhesion and
neovascularization capacities of circulating angiogenic cells
obtained from atherosclerotic patients. Atherosclerosis.
227:275–282. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kuckleburg CJ, McClenahan DJ and
Czuprynski CJ: Platelet activation by histophilus somni and its
lipooligosaccharide induces endothelial cell proinflammatory
responses and platelet internalization. Shock. 29:189–196.
2008.PubMed/NCBI
|
|
25
|
Deli MA, Abraham CS, Kataoka Y and Niwa M:
Permeability studies on in vitro blood-brain barrier models:
Physiology, pathology, and pharmacology. Cell Mol Neurobiol.
25:59–127. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bachmeier C, Mullan M and Paris D:
Characterization and use of human brain microvascular endothelial
cells to examine β-amyloid exchange in the blood-brain barrier.
Cytotechnology. 62:519–529. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Helms HC, Abbott NJ, Burek M, Cecchelli R,
Couraud PO, Deli MA, Förster C, Galla HJ, Romero IA, Shusta EV, et
al: In vitro models of the blood-brain barrier: An overview of
commonly used brain endothelial cell culture models and guidelines
for their use. J Cereb Blood Flow Metab. 36:862–890. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gloesenkamp CR, Nitzsche B, Ocker M, Di
Fazio P, Quint K, Hoffmann B, Scherubl H and Hopfner M: AKT
inhibition by triciribine alone or as combination therapy for
growth control of gastroenteropancreatic neuroendocrine tumors. Int
J Oncol. 40:876–888. 2012.
|
|
29
|
Jiang P, Ren YL, Lan Y, Li JL, Luo J, Li J
and Cai JP: Phagocytosis of platelets enhances endothelial cell
survival under serum deprivation. Exp Biol Med (Maywood).
240:876–883. 2015. View Article : Google Scholar
|
|
30
|
van Moorselaar RJ and Voest EE:
Angiogenesis in prostate cancer: Its role in disease progression
and possible therapeutic approaches. Mol Cell Endocrinol.
197:239–250. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Luster AD, Alon R and von Andrian UH:
Immune cell migration in inflammation: Present and future
therapeutic targets. Nat Immunol. 6:1182–1190. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Garrett CR, Coppola D, Wenham RM, Cubitt
CL, Neuger AM, Frost TJ, Lush RM, Sullivan DM, Cheng JQ and Sebti
SM: Phase I pharmacokinetic and pharmacodynamic study of
triciri-bine phosphate monohydrate, a small-molecule inhibitor of
AKT phosphorylation, in adult subjects with solid tumors containing
activated AKT. Invest New Drugs. 29:1381–1389. 2011. View Article : Google Scholar
|
|
33
|
Risau W and Wolburg H: Development of the
blood-brain barrier. Trends Neurosci. 13:174–178. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Plate KH: Mechanisms of angiogenesis in
the brain. J Neuropathol Exp Neurol. 58:313–320. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Porter AG and Janicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Johnson SA, Balboa RS, Dessel BH, Monto
RW, Siegesmund KA and Greenwalt TJ: The mechanism of the
endothelial supporting function of intact platelets. Exp Mol
Pathol. 3:115–127. 1964. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lang D, Dohle F, Terstesse M, Bangen P,
August C, Pauels HG and Heidenreich S: Down-regulation of monocyte
apoptosis by phagocytosis of platelets: Involvement of a caspase-9,
caspase-3, and heat shock protein 70-dependent pathway. J Immunol.
168:6152–6158. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shin WS, Maeng YS, Jung JW, Min JK, Kwon
YG and Lee ST: Soluble PTK7 inhibits tube formation, migration, and
invasion of endothelial cells and angiogenesis. Biochem Biophys Res
Commun. 371:793–798. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Moussa M, Lajeunesse D, Hilal G, El Atat
O, Haykal G, Serhal R, Chalhoub A, Khalil C and Alaaeddine N:
Platelet rich plasma (PRP) induces chondroprotection via increasing
autophagy, anti-inflammatory markers, and decreasing apoptosis in
human osteoarthritic cartilage. Exp Cell Res. 352:146–156. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Luzak B, Golanski J, Rozalski M, Krajewska
U, Olas B and Watala C: Extract from Aronia melanocarpa fruits
potentiates the inhibition of platelet aggregation in the presence
of endothelial cells. Arch Med Sci. 6:141–144. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Metzig C, Grabowska E, Eckert K, Rehse K
and Maurer HR: Bromelain proteases reduce human platelet
aggregation in vitro, adhesion to bovine endothelial cells and
thrombus formation in rat vessels in vivo. In Vivo. 13:7–12.
1999.PubMed/NCBI
|
|
42
|
Wen H, Lu Y, Yao H and Buch S: Morphine
induces expression of platelet-derived growth factor in human brain
microvascular endothelial cells: Implication for vascular
permeability. PLoS One. 6:e217072011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Rauch BH, Millette E, Kenagy RD, Daum G,
Fischer JW and Clowes AW: Syndecan-4 is required for
thrombin-induced migration and proliferation in human vascular
smooth muscle cells. J Biol Chem. 280:17507–17511. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Enomoto A, Ping J and Takahashi M: Girdin,
a novel actin-binding protein, and its family of proteins possess
versatile functions in the Akt and Wnt signaling pathways. Ann N Y
Acad Sci. 1086:169–184. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Skurk C, Maatz H, Kim HS, Yang J, Abid MR,
Aird WC and Walsh K: The Akt-regulated forkhead transcription
factor FOXO3a controls endothelial cell viability through
modulation of the caspase-8 inhibitor FLIP. J Biol Chem.
279:1513–1525. 2004. View Article : Google Scholar
|
|
46
|
Yang JY, Michod D, Walicki J and Widmann
C: Surviving the kiss of death. Biochem Pharmacol. 68:1027–1031.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Curry JM, Eubank TD, Roberts RD, Wang Y,
Pore N, Maity A and Marsh CB: M-CSF signals through the MAPK/ERK
pathway via Sp1 to induce VEGF production and induces angiogenesis
in vivo. PLoS One. 3:e34052008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Jehle AW, Gardai SJ, Li S, Linsel-Nitschke
P, Morimoto K, Janssen WJ, Vandivier RW, Wang N, Greenberg S, Dale
BM, et al: ATP-binding cassette transporter A7 enhances
phagocytosis of apoptotic cells and associated ERK signaling in
macrophages. J Cell Biol. 174:547–556. 2006. View Article : Google Scholar : PubMed/NCBI
|