1. Introduction
Trafficking protein particle complex subunit 9
(TRAPPC9) is a protein subunit of the transport protein particle II
(TRAPPII), a conserved trafficking molecule in organisms ranging
from yeast to humans (1). TRAPPII
shares similarity with complexes TRAPPI and TRAPPIII, in that all
three complexes comprise six core proteins. However, TRAPPII is the
only complex that contains TRAPPC9 in addition to two other
TRAPPII-specific proteins (TRAPPC10 and TRAPPC2L) (2). The TRAPPII complex is a Rab1 guanine
exchange factor known to bind coat protein I (COPI) and to serve as
a tethering complex for COPI-coated vesicles to the early Golgi
membrane. During TRAPPII complex formation, TRAPPC9 has been shown
to interact with TRAPPC2 and TRAPPC10. In addition, TRAPPC9 has
been reported to bind and interact with p150(Glued) at the
trans-Golgi region (3). Studies
have also suggested that mutations in TRAPPC9 are linked with a
form of mental retardation (MR) associated with severe osseous
deformities, including short stature and polydactylism (4). Clinical phenotypes associated with
TRAPPC9 mutation have been linked with decreased activation of
nuclear factor-κB (NF-κB). NF-κB is critical in the activation of
genes associated with multiple pathways (5). In the present review, the possible
roles and functions of TRAPPC9 in normal and disease processes are
highlighted.
2. Structure
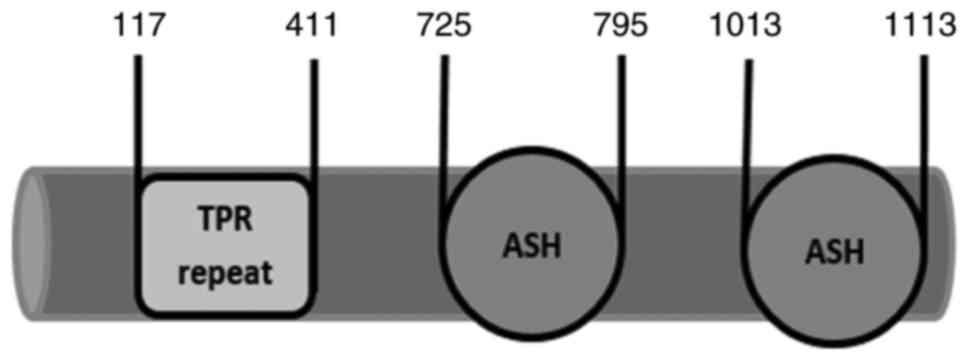
A survey of the human proteome by Schou et al
(6) identified two protein
domains in TRAPPC9: An ASPM, SPD-2, Hydin (ASH) domain at the
C-terminal, and predicted α-solenoid-bearing stretches of multiple
tetratricopeptide (TPR) repeats (Fig.
1). The TPR repeats have been identified in numerous proteins
and may serve as binding elements in multiprotein complexes
(7). The TPR repeats are also
considered to regulate diverse biological processes, including
organelle targeting and protein import, vesicle fusion and
biomineralization (7-9). By contrast, the ASH
domain is typically present in proteins associated with cilia,
flagella, the centrosome and the Golgi complex (10).
3. Expression
In humans, TRAPPC9 (MIM no. 611966) is encoded by
the gene located at locus 8q24.3 and contains 23 exons. TRAPPC9 is
a conserved protein with sequence similarity of the human gene
found in mouse (92%), chicken (87%) and zebrafish (85%) (9). TRAPPC9 has a conserved region
(Trs120), first identified in Saccharomyces cerevisiae
(12,13). Two variants have been identified
in humans, one encoding a 1,148-amino acid protein and the second
encoding a 1,246-amino acid protein. Compared to the human form,
the mouse TRAPPC9 gene is located on 15_NC_000081.6 and five
variants have been identified; the first variant encodes a
3,324-amino acid protein (NM_029640.2), the second encodes a
4,069-amino acid protein (NM_180662.2), the third encodes a
4,688-amino acid protein (NM_001164641.1), the fourth encodes a
3,127 amino acid protein (NM_001164642.1) and the fifth encodes a
2,833-amino acid protein (NM_001164643.1). TRAPPC9 is expressed at
high levels in the developing cortical plate of the human embryonic
brain (at 11.5 weeks gestation) and in the mouse brain during its
adult phase according to in situ hybridization data
(13). In mice, the expression of
TRAPPC9 has been localized in neurons of the cerebral cortex, the
hippo-campus and deep gray matter (13). Another study revealed that TRAPPC9
may also be expressed in mouse colon and small intestine tissues by
conventional reverse transcription-polymerase chain reaction
analysis (11). Further analysis
of human tissues by Zhang et al (11) using northern blot analysis
indicated a high expression of TRAPPC9 in muscle and the kidneys,
and low expression in the brain, heart and placenta. TRAPPC9 was
also shown to be weakly expressed in immune organs and cells,
including the thymus, spleen and peripheral blood leukocytes.
4. Function
TRAPPC9 is part of the TRAPPII complex, which is
important in intra-Golgi and endosomal trafficking in yeast
(14,16). In mammals, TRAPPC9 is
predominantly expressed at the endoplasmic reticulum (ER) exit
sites (17). All TRAPP complexes
share six core subunits, namely trafficking protein particle
complex subunits 20, 23, 31 and 33 (Trs20, 23, 31 and 33,
respectively) and two copies of Bet3, in addition to Bet5 and the
six core proteins. TRAPPII also contains four specific proteins,
Trs65, Trs120 (TRAPPC9), Trs130 (TRAPPC10) and Tcal17, specific to
TRAPPII (18,20). TRAPPC9 has been shown to be
involved in the NF-κB signaling pathway by physically interacting
and regulating NF-κB-inducing kinase (NIK) and inhibitor of NF-κB
(IκB) kinase subunit β (IKKβ; also known as IKK2) activation
(21). A previous study indicated
that TRAPPC9 may regulate enteric neuronal differentiation through
the NF-κB signaling pathway (11).
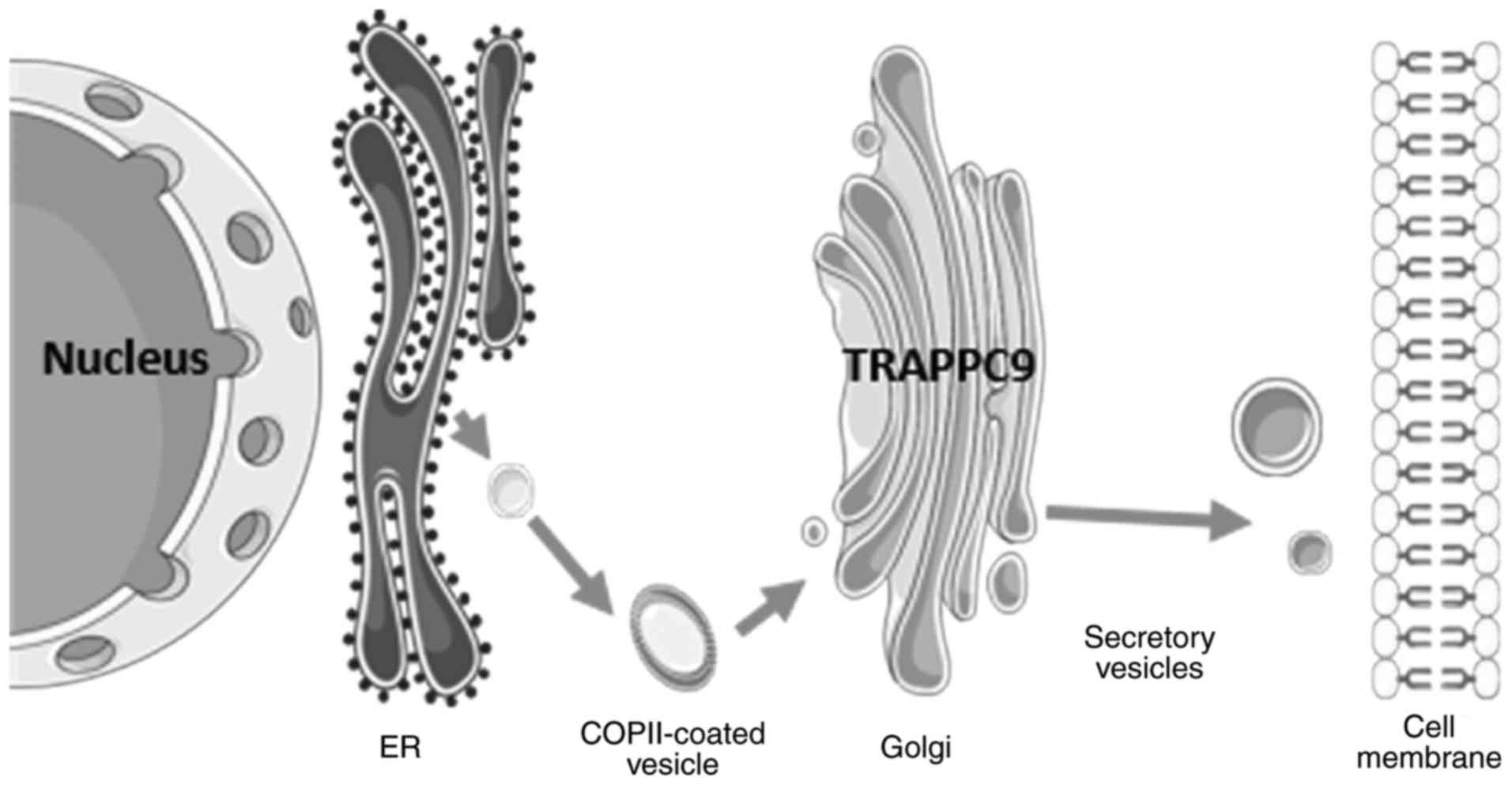
5. Trafficking
Newly synthesized proteins must be transported from
the ER to the Golgi complex via the secretory pathway (22). To ensure the directionality and
accuracy of the protein transport process, the process is completed
and regulated by intracellular membrane traffic complexes. In yeast
and mammals, several of the mechanisms that have been suggested to
underlie the secretory pathways within cells have been indicated
through decades of research (23).
TRAPPC9 is part of the TRAPPII complex, which is
expressed predominantly at the ER exit sites in mammals and has
been suggested to function in intra-Golgi and endosome-to-Golgi
transport (17) (Fig. 2). TRAPPC9 interacts with the
TRAPPC10 subunit of TRAPPII, which is considered to function as a
guanine exchange factor for Ypt/Rab GTPase by activating Rab1
(24). Site-directed mutagenesis
of TRAPPC10 has indicated that it may be implicated in autophagy,
and may be important in cytoplasmic-to-vacuole targeting and
starving-induced autophagy in Saccharomyces cerevisiae
(24). Compared to TRAPPC10,
mutation in TRAPPC9 reportedly causes the accumulation of aberrant
membrane structures that resemble Berkeley bodies, which is the
transport medium between the cytoplasm and vacuole within the
cytoplasm-to-vacuole targeting pathway in yeast; this results in
disruption to the trafficking of proteins that recycle through the
early endosome (15). TRAPPII is
enriched on coat protein I (COPI)-coated vesicles and interacts
directly with γ1COP, a COPI coat adaptor subunit and the heptameric
complex that forms the coat of COPI vesicles (23-25). Zong et al (3) demonstrated that TRAPPC9 may bind and
interact with p150(Glued) at the same carboxyl terminal domain of
p150(Glued) that binds Sec23 and Sec24. TRAPPC9 has also been shown
to co-localize with the late Golgi marker Sec7p (15). Previous site-directed mutagenesis
of TRAPPC9 resulted in defects in the localization of COPI.
Furthermore, it had been suggested that TRAPPC9 may serve to
uncouple p150(Glued) from the COPII coat and to relay the
vesicle-dynactin interaction at the target membrane (3). The functions associated with TRAPPC9
are summarized in Table I
(3,11,15,21,40-42).
 | Table ISummary of functions associated with
TRAPPC9. |
Table I
Summary of functions associated with
TRAPPC9.
| Author, date | Organism | Designation | Function | (Refs.) |
|---|
| Zong et al,
2012 | COS cells | TRAPPC9 | TRAPPC9 bound
directly to p150(Glued) via the same carboxyl terminal domain of
p150(Glued) that binds Sec23 and Sec24. TRAPPC9 reported to
uncouple p150 (Glued) from the COPII coat and relay the
vesicle-dynactin interaction at the target membrane. TRAPPC9
inhibits the interaction between p150 (Glued) and Sec23/Sec24 in
vivo and in vitro. | (3) |
| Zhang et al,
2014 | Bovine | NIBP | Expression of
TRAPPC9 is high in the mouse enteric nervous system. | (11) |
| Cai et al,
2005 | Yeast | Trs120 | TRAPPC9 is required
for vesicle trafficking from early endosome to late Golgi. TRAPPC9
co-localizes with the late Golgi marker Sec7p. TRAPPC9 mutation
causes accumulation of aberrant membrane structures that resemble
Saccharomyces cerevisiae organelle Berkeley bodies and
disrupt the traffic of proteins that recycle through the early
endosome. | (15) |
| Hu et al,
2005 | PC12 cells | NIBP | Novel NIK and IKKb
binding protein mainly expressed in brain, muscle, heart, and
kidney. Expression is low in immune tissues, including the spleen,
thymus, and peripheral blood leukocytes. TRAPPC9 shows physical
interaction with NIK and IKKb. Overexpression of TRAPPC9
potentiates the activation of NF-κB through increased IKK complex
phosphorylation. Downregulation of TRAPPC9 prevents TNFα-induced
activation of NF-κB and decreases gene expression of B-cell
lymphoma 2-extra large in pheochromocytoma PC12 cells. | (21) |
| Li et al,
2017 | 293T cells | TRAPPC9 | Regulates lipid
droplet size and the association of Rab18 on the lipid droplet
surface. | (40) |
| Salamat et
al, 2011 | Bovine | NIBP | TRAPPC9 interacts
with the N terminus of the bovine viral diarrhea virus NS5A. NS5A
co-localizes with TRAPPC9 on the endoplasmic reticulum in the
cytoplasm of infected cells. | (41) |
| Thellmann et
al, 2010 | Arabidopsis | AtTRS120 | TRAPPC9 is required
for cell plate biogenesis. Mutations in TRAPPC9 show canonical
cytokinesis-defective seedling-lethal phenotypes, including cell
wall stubs and incomplete cross walls. TRAPPC9 mutants show vesicle
accumulation at the equator of diving cells fails to assemble into
a cell plate. | (42) |
6. NF-κB
The NF-κB family comprises structurally related
transcription factors that regulate various biological processes,
including stress responses, immunity and inflammation (28). The activation of NF-κB is mediated
by ‘canonical’ and ‘non-canonical’ pathways (29). The canonical pathway is activated
by multiple stimuli, including receptor activator of NF-κB ligand
(RANKL), tumor necrosis factor (TNF)-α and other inflammatory
mediators (30). In turn, this
pathway is mediated by activation of the IKK complex, which
regulates phosphorylation and proteolysis of the IκBα inhibitors
and consequent nuclear translocation of the RelA and p50
transcriptional activators (30).
By contrast, the non-canonical NF-κB activation pathway involves
the activation of NIK to stimulate IKKα-induced phosphorylation,
the proteolytic processing of NF-κB2 into p52, and the nuclear
translocation of RelB (31). The
most well-characterized non-canonical NF-κB signaling is a subset
of TNF receptor superfamily members, including B-cell-activating
factor belonging to the TNF family receptor, cluster of
differentiation 40 and lymphotoxin β-receptor (32). TRAPPC9 has been shown to
physically interact with NIK and IKKβ, but not IKKα or IKKγ
(21). Therefore, TRAPPC9 is
implicated in the canonical and non-canonical NF-κB activation
pathways (Fig. 3). Of note,
TRAPPC9 may potentiate the activation of NF-κB through increased
phosphorylation of the IKK complex (21).
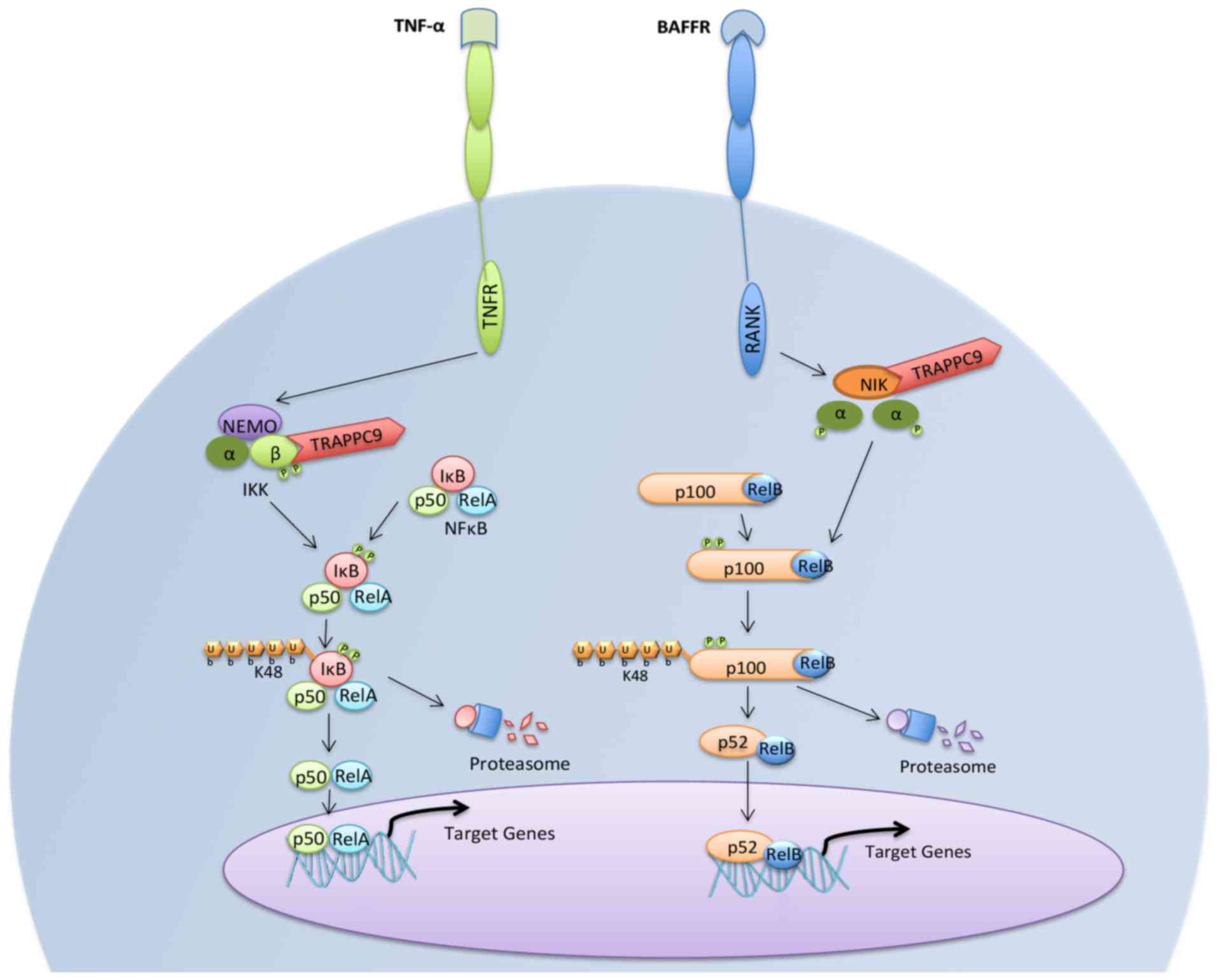 | Figure 3Activation of the canonical and
non-canonical pathways of NF-κB. TRAPPC9 mediates activation of the
NF-κB pathways via binding to IKKβ and NIK. Canonical and
non-canonical NF-κB pathway activation induces transcription
factors that are involved in various biological processes,
including the immune response, inflammation, cell growth and
survival, and development. TRAPPC9, trafficking protein particle
complex 9; NF-κB, nuclear factor-κB; NIK, nuclear
factor-κB-inducing kinase; IKKβ, inhibitor of NF-κB kinase subunit
β; TNF-α, tumor necrosis factor-α; TNFR, TNF receptor; BAFF, B-cell
activating factor; BAFFR; BAFF receptor; NEMO, NF-κB essential
modulator. |
7. TRAPPC9 in human diseases
Central nervous system
Mutations in TRAPPC9 have been identified in
patients with non-syndromic autosomal recessive MR (NS-ARMR)
(5). One such mutation was
identified in consanguineous family members and resulted in TRAPPC9
truncation; c.1422C>T. Mutation has also been identified in
three consanguineous Israeli-Arab female adolescents and in a large
consanguineous Pakistani family, where it was caused by the
nonsense mutation R475X in exon 7 (13,19). In addition, the novel homozygous
nonsense mutation c.2065G>T in exon 11 of the TRAPPC9 gene was
identified in a Pakistani family genome by whole exome sequencing
(33). Another mutation resulting
in a frameshift and premature truncation (p.Leu772TrpfsX7) is
caused by a homozygous 4-bp deletion, c.2311-2314 delTGTT,
identified in an Iranian family (34). A homozygous nonsense mutation
resulting in p.Arg570Ter (R570 X) due to c.1708C>T transcription
has been identified in three Tunisian brothers from a
consanguineous family (35). All
patients with NS-ARMR exhibit a similar clinical MR phenotype
ranging from moderate to severe. The clinical phenotype of patients
with mutation in the TRAPPC9 gene is moderate to severe MR,
including variable postnatal microcephaly. Mild facial dysmorphism
and truncal obesity have also been reported in Tunisian brothers,
whereas no facial dysmorphism has been observed in Pakistani,
Israeli-Arab or Iranian patients (13,5,35).
Phenotypes associated with TRAPPC9 mutations have been consistently
associated with postnatal microcephaly, speech delay,
neuroradiological abnormality of the cerebral white matter, corpus
callosum and cerebellum, peculiar facial appearance, obesity and
hypotonia (36,37). Magnetic resonance imaging in
affected individuals has revealed reduced cerebral white matter
volume with sulcal enlargement, thinning of the corpus callosum and
mild cerebellar volume loss. These phenotypes have been linked with
the downregulated activation of NF-κB, and it is possible that the
trafficking function of TRAPPC9 may also be affected. Human
mutations associated with TRAPPC9 have been summarized in Table II (13,33,37,36).
| Table IISummary of human mutations associated
with TRAPPC9. |
Table II
Summary of human mutations associated
with TRAPPC9.
| Author, year | Designation | Genotype | Phenotype
(Refs.) | |
|---|
| Mochida et
al, 2009 | TRAPPC9 | Nonsense nucleotide
mutation. | Defects in axonal
connectivity. Variable postnatal microcephaly of a consanguineous
Israeli Arab family. | (13) |
| Abbasi et
al, 2017 | TRAPPC9 | Homozygous nonsense
mutation c.2065G>T in exon 11 of the TRAPPC9 gene. | Severe intellectual
disability, motor delay, and absent speech. | (33) |
| Marangi et
al, 2013 | TRAPPC9 | Homozygous splice
sit mutation causing exon skipping with and premature termination.
frameshift | Peculiar facial
appearance, obesity, hypotonia, moderate-to-severe intellectual
disability, and brain abnormalities. Identified in two Italian
sisters born to healthy and apparently nonconsanguineous
parents. | (37) |
| Philippe et
al, 2009 | TRAPPC9 | Nonsense mutation
(c.1708C>T[p.R570X]) within exon 9. | Mild microcephaly
and white matter abnormalities. | (35) |
Liver disease
A previous study indicated that certain genetic loci
are associated with features of histological severity in
nonalcoholic fatty liver disease in a cohort of Hispanic boys
(38). In this study, 234
Hispanic boys (aged 2-17 years) with available clinical, laboratory
and histological data enrolled in the Nonalcoholic Steatohepatitis
Clinical Research Network were included in the analysis of 624,297
single nucleotide polymorphisms (SNPs). The median age and body
mass index z-score were 12.0 years [interquartile range (IQR),
11.0-14.0] and 2.4 (IQR, 2.1-2.6), respectively. Notably, the
nonalcoholic fatty liver disease activity score (1-4, vs. 5-8) was
associated with SNP rs11166927 on chromosome 8 in the TRAPPC9
region (P=8.7-07) (38).
Breast/colon cancer
Zhang et al (39) investigated the potential
regulatory mechanisms underlying the constitutive and inducible
activation of NF-κB in cancer as they remain to be fully
elucidated. The study investigated whether a novel NIK- and
IKK2-binding protein (NIBP/TRAPPC9) is required for maintaining the
malignancy of cancer cells in an NF-κB-dependent manner by
polymerase chain reaction analysis of a human cancer survey
tissue-scan cDNA array, immunostaining of a human frozen tumor
tissue array and immunoblotting of a high-density reverse-phase
cancer protein lysate array. The study indicated that TRAPPC9 was
extensively expressed in the majority of tumor tissues,
particularly in breast and colon cancer. More specifically, TRAPPC9
appeared to promote tumorigenesis via NF-κB signaling in breast
MDA-MB-231 and colon HCT116 cancer cells. The downregulation of
TRAPPC9 significantly inhibited the growth/proliferation,
invasion/migration, colony formation and xenograft tumorigenesis of
the breast and colon cancers cells (39).
8. Concluding remarks
TRAPPC9 is a conserved protein. The protein has been
found to be expressed in different human tissues and is implicated
in several intracellular protein trafficking processes. In
addition, TRAPPC9 has been observed to be important in NF-κB
signaling by directly binding and regulating NIK and IKKβ; via
these interactions, TRAPPC9 has been implicated in the canonical
and non-canonical NF-κB activation pathways. TRAPPC9 mutation may
also be essential in the pathogenesis of a number of human
diseases. Collectively these data highlight the importance of
understanding the normal physiological roles of TRAPPC9 in
NF-κB-mediated signaling and in trafficking, as this can assist in
improving our understanding of the role of TRAPPC9 in different
disease processes.
Acknowledgements
The authors would like to thank the School of
Biomedical Sciences, Kent State University for their support.
Funding
The present study was supported by the Cook
Philanthropy Foundation (grant no., 72264).
Availability of data and materials
Data sharing is not applicable to this article, as
no datasets were generated or analyzed during the current
study.
Authors' contributions
TM and NJH contributed equally to manuscript writing
and editing. AN assisted with preparing the figures. FFS
contributed to the manuscript editing and writing. All authors read
and approved the final manuscript for publication.
Ethics approval and consent to
participate
No applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kim JJ, Lipatova Z and Segev N: TRAPP
complexes in secretion and autophagy. Front Cell Dev Biol.
4:202016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yip CK, Berscheminski J and Walz T:
Molecular architecture of the TRAPPII complex and implications for
vesicle tethering. Nat Struct Mol Biol. 17:1298–1304. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zong M, Satoh A, Yu MK, Siu KY, Ng WY,
Chan HC, Tanner JA and Yu S: TRAPPC9 mediates the interaction
between p150 and COPII vesicles at the target membrane. PLoS One.
7:e299952012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mohamoud HS, Ahmed S, Jelani M, Alrayes N,
Childs K, Vadgama N, Almramhi MM, Al-Aama JY, Goodbourn S and Nasir
J: A missense mutation in TRAPPC6A leads to build-up of the
protein, in patients with a neurodevelopmental syndrome and
dysmorphic features. Sci Rep. 8:20532018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mir A, Kaufman L, Noor A, Motazacker MM,
Jamil T, Azam M, Kahrizi K, Rafiq MA, Weksberg R, Nasr T, et al:
Identification of mutations in TRAPPC9, which encodes the NIK- and
IKK-beta-binding protein, in nonsyndromic autosomal-recessive
mental retardation. Am J Hum Genet. 85:909–915. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schou KB, Morthorst SK, Christensen ST and
Pedersen LB: Identification of conserved, centrosome-targeting ASH
domains in TRAPPII complex subunits and TRAPPC8. Cilia. 3:62014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zeytuni N and Zarivach R: Structural and
functional discussion of the tetra-trico-peptide repeat, a protein
interaction module. Structure. 20:397–405. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Clairfeuille T, Norwood SJ, Qi X, Teasdale
RD and Collins BM: Structure and membrane binding properties of the
endosomal tetratricopeptide repeat (TPR) domain-containing sorting
nexins SNX20 and SNX21. J Biol Chem. 290:14504–14517. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hwang JY, Ahn SJ, Kwon MG, Seo JS, Hwang
SD and Son MH: Interferon-induced protein 56 (IFI56) is induced by
VHSV infection but not by bacterial infection in olive flounder
(paralichthys olivaceus). Fish Shellfish Immunol. 66:382–389. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ponting CP: A novel domain suggests a
ciliary function for ASPM, a brain size determining gene.
Bioinformatics. 22:1031–1035. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Y, Bitner D, Pontes Filho AA, Li F,
Liu S, Wang H, Yang F, Adhikari S, Gordon J, Srinivasan S and Hu W:
Expression and function of NIK- and IKK2-binding protein (NIBP) in
mouse enteric nervous system. Neurogastroenterol Motil. 26:77–97.
2014. View Article : Google Scholar :
|
|
12
|
Sacher M, Barrowman J, Schieltz D, Yates
JR III and Ferro-Novick S: Identification and characterization of
five new subunits of TRAPP. Eur J Cell Biol. 79:71–80. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mochida GH, Mahajnah M, Hill AD,
Basel-Vanagaite L, Gleason D, Hill RS, Bodell A, Crosier M,
Straussberg R and Walsh CA: A truncating mutation of TRAPPC9 is
associated with autosomal-recessive intellectual disability and
postnatal microcephaly. Am J Hum Genet. 85:897–902. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sacher M, Jiang Y, Barrowman J, Scarpa A,
Burston J, Zhang L, Schieltz D, Yates JR III, Abeliovich H and
Ferro-Novick S: TRAPP, a highly conserved novel complex on the
cis-Golgi that mediates vesicle docking and fusion. EMBO J.
17:2494–2503. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cai H, Zhang Y, Pypaert M, Walker L and
Ferro-Novick S: Mutants in trs120 disrupt traffic from the early
endosome to the late golgi. J Cell Biol. 171:823–833. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Barrowman J, Sacher M and Ferro-Novick S:
TRAPP stably associates with the golgi and is required for vesicle
docking. EMBO J. 19:862–869. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zahoor MA, Yamane D, Mohamed YM, Suda Y,
Kobayashi K, Kato K, Tohya Y and Akashi H: Bovine viral diarrhea
virus non-structural protein 5A interacts with NIK- and
IKKbeta-binding protein. J Gen Virol. 91:1939–1948. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu S and Liang Y: A trapper keeper for
TRAPP, its structures and functions. Cell Mol Life Sci.
69:3933–3944. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sacher M, Barrowman J, Wang W, Horecka J,
Zhang Y, Pypaert M and Ferro-Novick S: TRAPP I implicated in the
specificity of tethering in ER-to-golgi transport. Mol Cell.
7:433–442. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Choi C, Davey M, Schluter C, Pandher P,
Fang Y, Foster LJ and Conibear E: Organization and assembly of the
TRAPPII complex. Traffic. 12:715–725. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu WH, Pendergast JS, Mo XM, Brambilla R,
Bracchi-Ricard V, Li F, Walters WM, Blits B, He L, Schaal SM and
Bethea JR: NIBP, a novel NIK and IKK(beta)-binding protein that
enhances NF-(kappa)B activation. J Biol Chem. 280:29233–29241.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Barrowman J, Bhandari D, Reinisch K and
Ferro-Novick S: TRAPP complexes in membrane traffic: Convergence
through a common rab. Nat Rev Mol Cell Biol. 11:759–763. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Delic M, Valli M, Graf AB, Pfeffer M,
Mattanovich D and Gasser B: The secretory pathway: Exploring yeast
diversity. FEMS Microbiol Rev. 37:872–914. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zou S, Chen Y, Liu Y, Segev N, Yu S, Liu
Y, Min G, Ye M, Zeng Y, Zhu X, et al: Trs130 participates in
autophagy through GTPases Ypt31/32 in saccharomyces cerevisiae.
Traffic. 14:233–246. 2013. View Article : Google Scholar :
|
|
25
|
Montpetit B and Conibear E: Identification
of the novel TRAPP associated protein Tca17. Traffic. 10:713–723.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Scrivens PJ, Shahrzad N, Moores A, Morin
A, Brunet S and Sacher M: TRAPPC2L is a novel, highly conserved
TRAPP-interacting protein. Traffic. 10:724–736. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yamasaki A, Menon S, Yu S, Barrowman J,
Meerloo T, Oorschot V, Klumperman J, Satoh A and Ferro-Novick S:
mTrs130 is a component of a mammalian TRAPPII complex, a Rab1 GEF
that binds to COPI-coated vesicles. Mol Biol Cell. 20:4205–4215.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Israel LP, Benharoch D, Gopas J and
Goldbart AD: A pro-inflammatory role for nuclear factor kappa B in
childhood obstructive sleep apnea syndrome. Sleep. 36:1947–1955.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Brasier AR: The NF-kappaB regulatory
network. Cardiovasc Toxicol. 6:111–130. 2006. View Article : Google Scholar
|
|
30
|
Hayden MS and Ghosh S: Shared principles
in NF-kappaB signaling. Cell. 132:344–362. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sun SC: The noncanonical NF-κB pathway.
Immunol Rev. 246:125–140. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sun SC: Non-canonical NF-kappaB signaling
pathway. Cell Res. 21:71–85. 2011. View Article : Google Scholar
|
|
33
|
Abbasi AA, Blaesius K, Hu H, Latif Z,
Picker-Minh S, Khan MN, Farooq S, Khan MA and Kaindl AM:
Identification of a novel homozygous TRAPPC9 gene mutation causing
non-syndromic intellectual disability, speech disorder, and
secondary micro-cephaly. Am J Med Genet B Neuropsychiatr Genet.
174:839–845. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Najmabadi H, Motazacker MM, Garshasbi M,
Kahrizi K, Tzschach A, Chen W, Behjati F, Hadavi V, Nieh SE,
Abedini SS, et al: Homozygosity mapping in consanguineous families
reveals extreme heterogeneity of non-syndromic autosomal recessive
mental retardation and identifies 8 novel gene loci. Hum Genet.
121:43–48. 2007. View Article : Google Scholar
|
|
35
|
Philippe O, Rio M, Carioux A, Plaza JM,
Guigue P, Molinari F, Boddaert N, Bole-Feysot C, Nitschke P, Smahi
A, et al: Combination of linkage mapping and microarray-expression
analysis identifies NF-kappaB signaling defect as a cause of
autosomal-recessive mental retardation. Am J Hum Genet. 85:903–908.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kakar N, Goebel I, Daud S, Nürnberg G,
Agha N, Ahmad A, Nürnberg P, Kubisch C, Ahmad J and Borck G: A
homozygous splice site mutation in TRAPPC9 causes intellectual
disability and microcephaly. Eur J Med Genet. 55:727–731. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Marangi G, Leuzzi V, Manti F, Lattante S,
Orteschi D, Pecile V, Neri G and Zollino M: TRAPPC9-related
autosomal recessive intellectual disability: Report of a new
mutation and clinical phenotype. Eur J Hum Genet. 21:229–232. 2013.
View Article : Google Scholar :
|
|
38
|
Wattacheril J, Lavine JE, Chalasani NP,
Guo X, Kwon S, Schwimmer J, Molleston JP, Loomba R, Brunt EM, Chen
YI, et al: Genome-wide associations related to hepatic histology in
nonalcoholic fatty liver disease in hispanic boys. J Pediatr.
190:100–107.e102. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang Y, Liu S, Wang H, Yang W, Li F, Yang
F, Yu D, Ramsey FV, Tuszyski GP and Hu W: Elevated NIBP/TRAPPC9
mediates tumorigenesis of cancer cells through NFκB signaling.
Oncotarget. 6:6160–6178. 2015.PubMed/NCBI
|
|
40
|
Li C, Luo X, Zhao S, Siu GK, Liang Y, Chan
HC, Satoh A and Yu SS: COPI-TRAPPII activates Rab18 and regulates
its lipid droplet association. EMBO J. 36:441–457. 2017. View Article : Google Scholar
|
|
41
|
Salamat MK, Dron M, Chapuis J, Langevin C
and Laude H: Prion propagation in cells expressing PrP
glycosylation mutants. J Virol. 85:3077–3085. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Thellmann M, Rybak K, Thiele K, Wanner G
and Assaad FF: Tethering factors required for cytokinesis in
arabidopsis. Plant Physiol. 154:720–732. 2010. View Article : Google Scholar : PubMed/NCBI
|