Introduction
Osteoarthritis (OA) is a chronic joint disease
characterized by the progressive degradation of articular
cartilage, accompanied by remodeling of the subchondral bone and
consequent articular dysfunction. OA affects 20-25% of individuals
aged >45 years and 37.4% of individuals aged ≥60 years old
(1,2). Knee OA may cause severe pain,
physical disability and reduced quality of life. Furthermore, OA is
one of the major reasons for knee arthroplasty and leads to high
healthcare costs (3,4).
A variety of risk factors contribute to OA,
including age, gender, obesity, history of knee injury, genetic
factors and nutritional factors (5). Research in the field of molecular
biology has identified an imbalance in the homeostasis of the
cartilage matrix during OA, as well as inflammation caused by
various biomarkers, such as prostaglandins, adipokines and
cytokines among others (6).
Moderate physical exercise is recognized to be an
effective therapeutic treatment for knee OA (7). Research revealed that a moderate
level of exercise is beneficial in rat knee OA models, and affects
subchondral or trabecular bone metabolism (8-10).
Various cytokines, growth factors and proteins secreted by
osteoclasts and osteoblasts of OA subchondral bone cysts aggravate
the disease and are alleviated by moderate physical exercise
(11-14).
Autophagy, as an essential cellular homeostatic
mechanism in eukaryotic cells, involves continuous biosynthesis and
renewal of cellular organelles and macromolecules. It is recognized
that autophagy serves an important role in the knee OA process, in
which it has been demonstrated that multiple signal transduction
pathways may be involved (15).
Previous studies reported that autophagy in chondrocytes mainly
serves as a protective response of cells to negative stimulation,
such as hypoxia (16), glucose
starvation or nutrient insufficiency (17), aging (18) and certain inflammatory mediators
(19-21). Nevertheless, complex results have
been derived from in vivo studies (18,19,22). Autophagy may not only be a
protective or homeostatic mechanism in normal or OA cartilage, but
may also be an essential process for the differentiation and
maturation of chondrocytes, which occurs constantly during the
pathological process of OA (23).
A previous study reported that physical exercise
affects autophagy in numerous tissues and organs, including muscle,
adipose, heart and brain tissues (24). The present study was designed to
detect the alterations in autophagy in the cartilage following
treadmill exercise using a rat OA model that was induced by
monosodium iodoacetate (MIA) injection. The results may help to
develop a novel theory regarding the treatment effect of exercise
on OA.
Materials and methods
Experimental animals
The experimental protocol was approved by the Ethics
Committee of Shengjing Hospital, China Medical University
(Shenyang, China). A total of 30 male Sprague-Dawley rats (230±10
g; 8 weeks old; specific-pathogen-free) were purchased from HFK
Bioscience Co., Ltd. (Beijing, China) and kept in plastic cages
with sawdust bedding in a controlled environment (22±2°C; 70%
humidity) under a 12:12-h light/dark cycle with the lights turned
on at 06:00 a.m. Animals were able to move freely in the cages,
were fed a standard diet and had access to tap water ad
libitum. The body weight of rats was recorded weekly prior to
the treadmill exercise each Monday. Adaptive exercise was applied
to all animals at a speed of 10 m/min for 10 min/day prior to
regular treadmill exercise to eliminate stress.
OA model and treadmill running
protocols
The animals were randomly divided into three groups
(n=10 per group), as follows: Control (sterile saline injection
only), OA (MIA injection) and OA with treadmill exercise (OAE). An
intra-articular injection of MIA was used in both knees following
the protocol described in our previous study (25), in order to generate the OA model
according to the study of Guzman et al (26). All rats were kept sedentary in the
cages; however, the OAE group began treadmill exercise at 24 h
post-injection with a speed of 18 m/min for 30 min/day (moderate
exercise) for 5 days/week, which is considered to be beneficial for
cartilage damage caused by MIA injection. The treatment schedule is
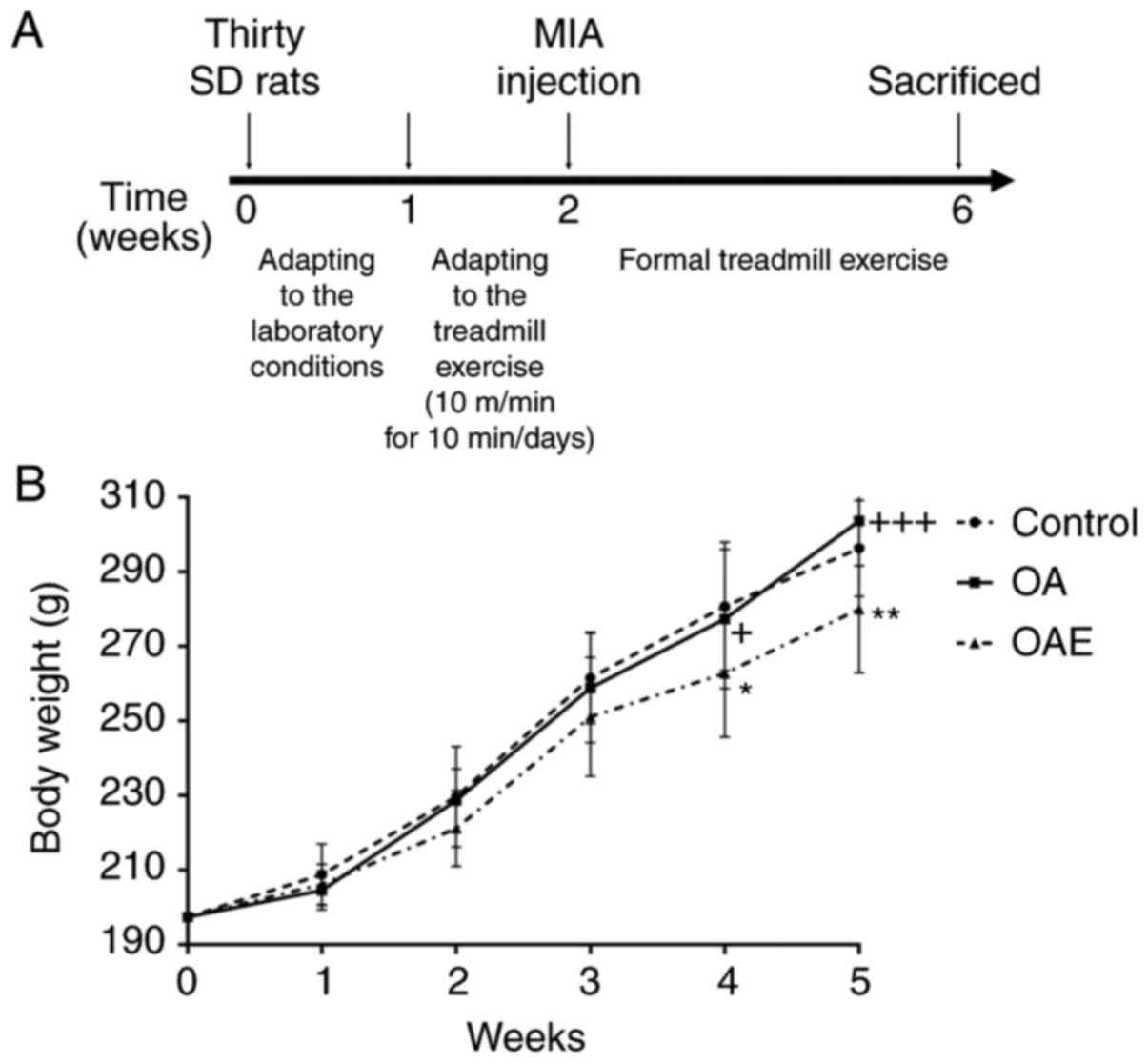
shown in Fig. 1A.
Sampling and tissue preparation
After 4 weeks of exercise, all animals were
anesthetized 2 h subsequent to the last exercise session. Blood
samples (3 ml) were obtained immediately and centrifuged at 3,000 ×
g for 10 min in 4°C to obtain the serum. All rats were euthanized,
and the joints of the left knee were separated and fixed in 4%
paraformaldehyde solution at room temperature. Intra-articular
lavage fluid (IALF) was obtained from the synovial cavity of the
right knee by injection and recovery with 0.2 ml PBS three times
using a 1-ml syringe. Cartilage was scraped off from the
weight-bearing area of the condyles of the right femur and tibia
using a scalpel, and was carefully collected. The left knee joints
were kept at room temperature, while other samples were kept at
−80°C.
Histological analysis
Following 2 weeks of soaking in 4% paraformaldehyde
solution, the left knee joints were washed with PBS five times (10
min/wash) and transferred to a 10% EDTA solution (EDTA-2Na,
dissolved in 0.1 M PBS) to decalcify the samples. Subsequently,
samples were kept at a constant temperature of 37°C for 1 month and
the solution was renewed every 3 days. Dehydration in an ethanol
series and embedding in paraffin was performed successively
following the decalcification, and serial 4.5-µm sagittal
sections were then cut for histological examination. Hematoxylin
and eosin (H&E) staining and toluidine blue staining were used
to obtain scores according to the Osteoarthritis Research Society
International (OARSI) and the modified Mankin scoring systems,
respectively, in order to grade and stage OA development in
cartilage (27). Synovitis
scoring, which reflects the inflammation state in the synovium
(28), was also performed by
H&E staining.
ELISA of IALF and serum levels
IL-1β and IL-4 levels in the knee IALF and serum
were detected using ELISA kits (cat. nos. ml037361 and ml102825;
Shanghai Enzyme-linked Biotechnology Co., Ltd., Shanghai, China),
following the manufacturer’s protocol. Additionally, the
concentration of protein in the IALF was measured using a
bicinchoninic acid (BCA) assay kit (cat. no. P0010S; Beyotime
Institute of Biotechnology, Shanghai, China) to correct the error
in harvesting.
Immunohistochemical assay
Left knee joint sections were deparaffinized in
xylene and rehydrated in graded ethanol and water. Enzymatic
antigen retrieval (cat. no. AR0026; Boster Biological Technology,
Pleasanton, CA, USA) was performed for 30 min at 37°C for antigen
retrieval, and left knee joint sections were then stained using a
two-step method, according to the protocol of the SPlink Detection
kit (cat. no. SP-9001; Zhongshan Golden Bridge Biotechnology Co.,
Ltd., Beijing, China). The sections were incubated overnight at 4°C
with rabbit polyclonal antibodies against type II collagen (1:500
dilution; cat. no. ab34712), light chain 3β (LC3B; 1:100 dilution;
cat. no. ab48394), BECLIN1 (1:800 dilution; cat. no. ab62557) (all
from Abcam, Cambridge, MA, USA) and sequestosome 1 (SQSTM1; 1:20
dilution; cat. no. 8420-1-AP; ProteinTech Group, Wuhan, China).
Sections were then incubated with biotinylated secondary antibody
and streptavidin/horseradish peroxidase were incubated for 30 min
at room temperature successively according to the instructions of
the SPlink Detection kit. Finally, the sections were visualized
with 3,3-diaminobenzidine tetrahydrochloride for 1 min and
counterstained with hematoxylin for 5 min. The negative control
group was assayed according to the abovementioned procedure, by
substituting the primary antibody with PBS. The type II collagen
content was evaluated based on the optical density of positive area
measured using the image analysis software Image-Pro Plus, version
6.0 (Media Cybernetics, Inc., Rockville, MD, USA). To evaluate the
positive cells of LC3B, BECLIN1 and SQSTM1 in the different zones
of the cartilage (29), three
images of each protein were captured under ×200 magnification,
representing the cartilages in the loaded parts. The total number
and positive number of chondrocytes, as well as the number of
different zones, were counted in each image, respectively. Finally,
the percentage of positive cells was calculated.
Western blotting
Following removal from storage at −80°C, the
cartilage was washed in PBS twice, and ultrasonication was
performed in radioimmunoprecipitation assay lysis buffer (cat. no.
P0013C) with 1 mM phenylmethylsulfonyl fluoride (cat. no. ST506)
(both from Beyotime Institute of Biotechnology). The lysates were
centrifuged at 14,000 × g for 5 min at 4°C, and the supernatants
were obtained to measure the protein concentration with the BCA
assay kit. Equal amounts of protein (50 µg) were added to an
SDS-PAGE gel [8% for type II collagen and a disintegrin and
metalloproteinase with thrombospondin motifs 5 (ADAMTS5); 12% for
LC3B and β-actin] following heating with loading buffer (cat. no.
NP0007; Thermo Fisher Scientific, Inc., Waltham, MA, USA) at 100°C
for 5 min, and then samples were transferred to polyvinylidene
difluoride membranes. Following blocking at room temperature for 2
h with 1% bovine serum albumin (cat. no. A8020; Solarbio Science
& Technology Co., Ltd., Beijing, China) in Tris-buffered saline
with 0.1% Tween-20 (TBST), the membranes were incubated with
primary antibodies overnight at 4°C, including the following:
Rabbit polyclonal anti-type II collagen (molecular weight, 142 kDa;
1:5,000 dilution; cat. no. ab34712; Abcam), rabbit polyclonal
anti-ADAMTS5 (molecular weight, 100 kDa; 1:200 dilution; cat. no.
BA3020; Boster Biological Technology), rabbit polyclonal anti-LC3B
(molecular weight, 15 kDa for LC3-II and 17 kDa for LC3-I; 1:1,000
dilution; cat. no. ab48394; Abcam) and mouse monoclonal
anti-β-actin (molecular weight, 42 kDa; cat. no. AA128; 1:1,000;
Beyotime Institute of Biotechnology) antibodies. Following washing
five times with TBST for 5 min each time, the peroxidase-conjugated
AffiniPure goat anti-rabbit and anti-mouse IgG (H+L) secondary
antibodies (cat. nos. ZB5301 and ZB5305; Zhongshan Golden Bridge
Biotechnology Co., Ltd.) were applied at a dilution of 1:10,000 in
TBST and incubated at room temperature for 1.5 h. The membrane was
washed with TBST again, detected with enhanced chemiluminescence
(cat. no. P0018; Beyotime Institute of Biotechnology) and
quantified using Image-Pro Plus software, version 6.0 (Media
Cybernetics, Inc.). β-actin served as the internal reference.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Following removal of the cartilage from the −80°C
freezer, a mortar and pestle were used to grind up the samples in a
low-temperature environment maintained by liquid nitrogen.
Approximately 50 mg cartilage samples was collected for total RNA
isolation with RNAiso Plus (cat. no. 9108; Takara Biotechnology
Co., Ltd., Dalian, China) and High-Salt Solution for Precipitation
(Plant) (cat. no. 9193; Takara Biotechnology Co., Ltd.). A total of
1 µg total RNA was then reverse-transcribed into
first-strand complementary DNA (cDNA) with a PrimeScript RT reagent
kit with gDNA Eraser (cat. no. RR047A; Takara Biotechnology Co.,
Ltd.). The cDNA samples were used to conduct qPCR with an ABI Prism
7500 Fast Real-Time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.) in a 20-µl system using SYBR Premix Ex Taq
II (cat. no. RR820A; Takara Biotechnology Co., Ltd.) with a
two-step method (95°C for 30 sec, 95°C 5 sec, 40 cycles of 60°C for
34 sec). All kits and reagents were used according to the
manufacturer’s protocol. The following primers were used: BECLIN1
forward, 5′-CTG GAC CGA GTG ACC ATT CA-3′ and reverse, 5′-AGA CAC
CAT CCT GGC GAG TT-3′; LC3B forward, 5′-TTC TTC CTC CTG GTG AAT
GG-3′ and reverse, 5′-CTG GGA GGC ATA GAC CAT GT-3′; SQSTM1
forward, 5′-CTG TGG TGG GAA CTC GCT AT-3′ and reverse, 5′-GAA AGA
TGA GCT TGC TGT GCT-3′; β-actin forward, 5′-CAC CCG CGA GTA CAA CCT
TC-3′ and reverse, 5′-CCC ATA CCC ACC ATC ACA CC-3′. The expression
levels of the PCR products were calculated using the
2−ΔΔCq method, with β-actin serving as the housekeeping
gene (30).
Statistical analysis
All data are expressed as the mean and 95%
confidence interval (CI), and were analyzed using IBM SPSS
statistical software version 20 (IBM Corp., Armonk, NY, USA). To
evaluate the normality of the results, the Kolmogorov-Smirnov and
Shapiro-Wilk tests (when sample size was ≤5) were performed.
Comparisons among groups were determined by one-way analysis of
variance with Tukey’s tests for pairwise comparisons, following the
Levene’s test to examine homogeneity of variance. If homogeneity of
variance was missing, Games-Howell method was used as a
replacement. P<0.05 was considered to indicate a statistically
significant difference.
Results
Weight change
There were no significant changes between the
control and the OA group throughout the procedure. At the same
time, there was a significant difference between the OAE and the
control, the OAE and the OA group at the fourth and fifth week
(Fig. 1B).
Histological analysis of knee
cartilage
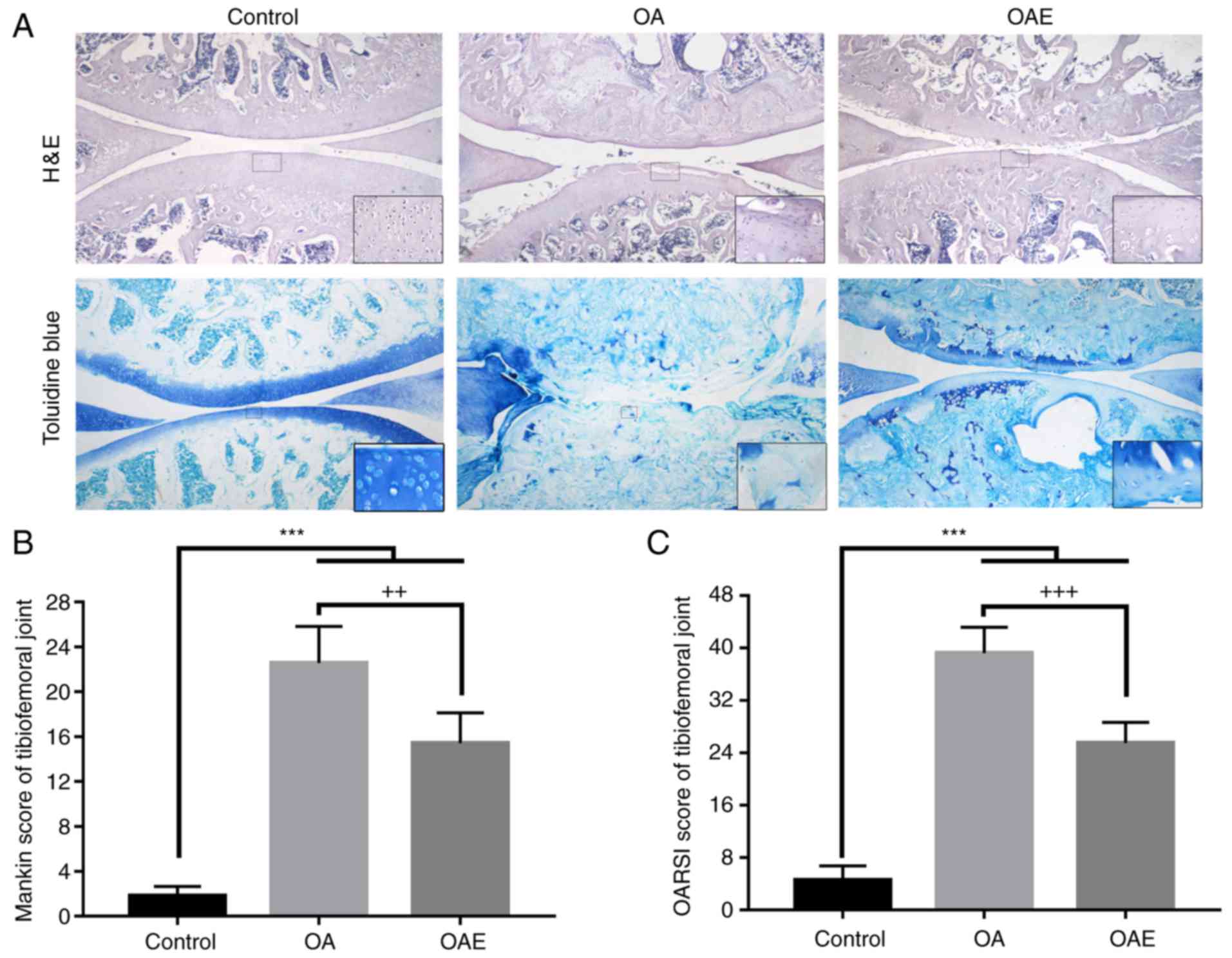
The joint cartilage of the control group exhibited a
slight degeneration in the superficial zone (SZ), including
cartilage matrix edema and chondrocyte proliferation (Fig. 2A). By contrast, in the OA group,
severe damage was observed in sections stained with H&E and
toluidine blue, such as denudation in the deep zone (DZ) and
microfractures in the fibrocartilage. In the OAE group, the
cartilage exhibited different alterations, from superficial
fibrillation to matrix loss in the mid zone. Furthermore, the
Mankin score in the control group was 1.80 (95% CI, 0.95-2.65),
while it was 22.55 (95% CI, 19.27-25.83) in the OA group and 15.40
(95% CI, 12.69-18.11) in the OAE group (Fig. 2B). The OARSI score was 4.55 (95%
CI, 2.36-6.74) in the control group, 39.20 (95% CI, 35.23-43.17) in
the OA group, and 25.50 (95% CI, 22.36-28.64) in the OAE group
(Fig. 2C). The OA and OAE group
scores were significantly increased compared with the control,
while there was a marked reduction in the OAE group in comparison
with the scores in the OA group.
Type II collagen and ADAMTS5
alterations
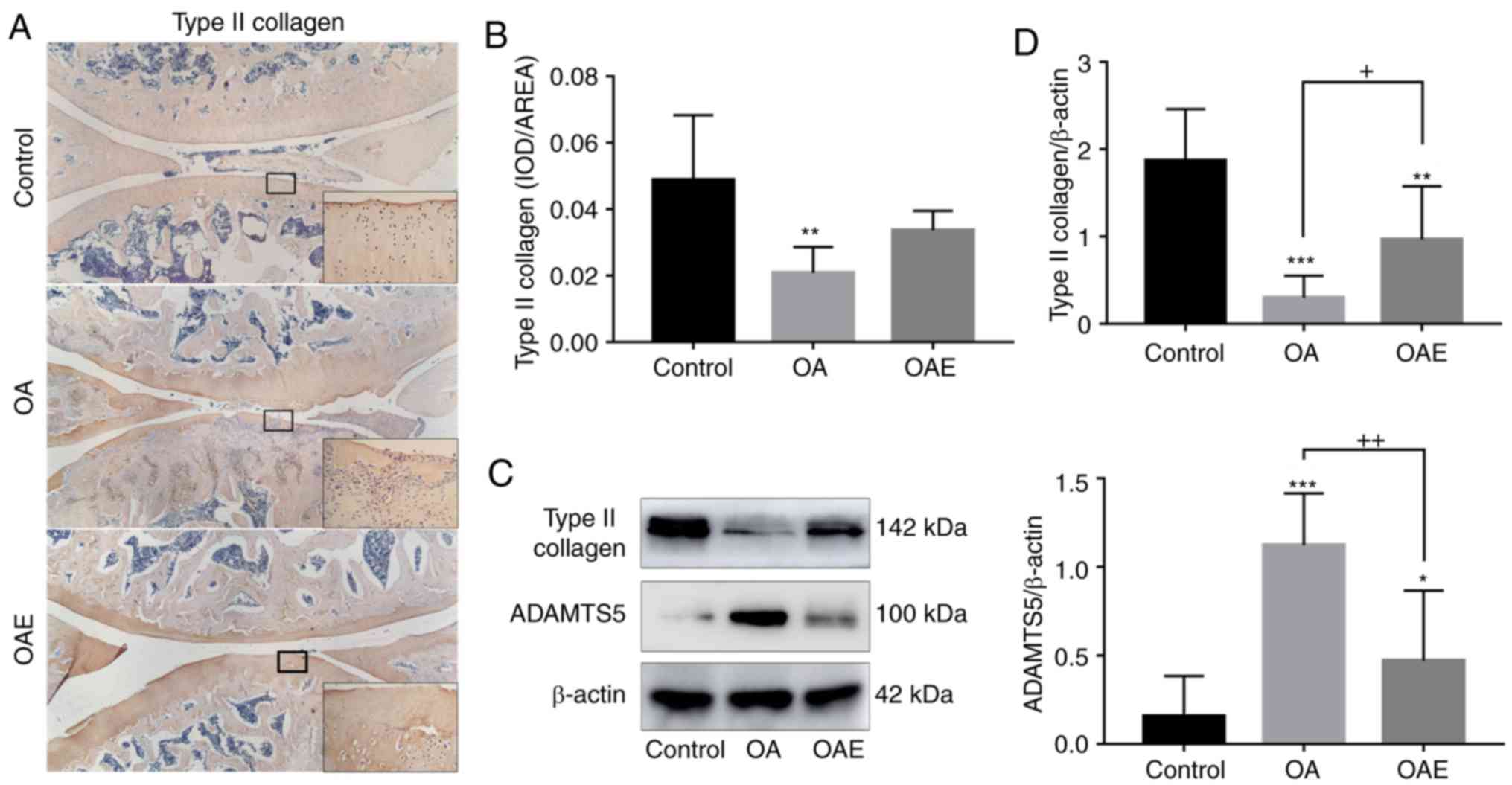
As demonstrated via immunohistochemical analysis,
type II collagen expression in the OA group was much weaker in
comparison with that in the control group, and this reduction was
notably attenuated by exercise in the OAE group (relative
percentages, 43 and 69% in the OA and OAE groups, respectively;
Fig. 3A and B). The expression of
type II collagen detected via western blotting exhibited a similar
trend to the immunohistochemistry results [control group, 1.87 (95%
CI, 1.28-2.46); OA group, 0.30 (95% CI, 0.05-0.55); and OAE group,
0.97 (95% CI, 0.35-1.58); Fig. 3C and
D]. On the contrary, the expression of ADAMTS5 was markedly
increased in the OA group and moderately increased in the OAE group
[control group, 0.16 (95% CI, −0.07-0.38); OA group, 1.12 (95% CI,
0.83-1.42), and OAE group, 0.47 (95% CI, 0.08-0.87; Fig. 3C and D].
Inflammation in synovium, and IL-1β and
IL-4 levels detected by ELISA
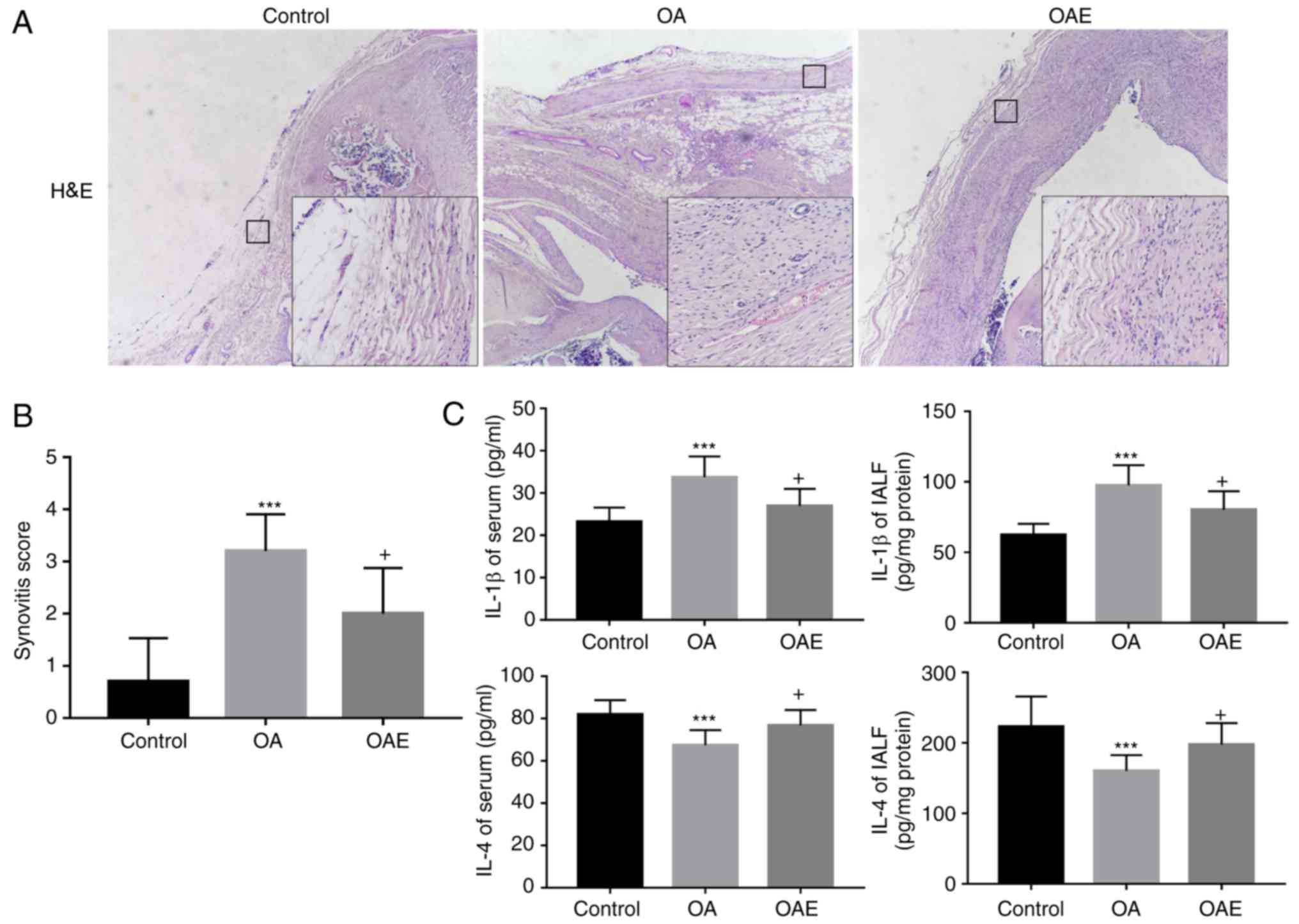
As compared with the control group, H&E staining
of the synovium of the OA group displayed an enlargement of the
synovial lining cell layer and inflammatory cell infiltration,
accompanied by an increase of the cellularity (Fig. 4A). However, the synovial stroma
was mostly normal in the OAE group. The synovitis score in the
control group was 0.90 (95% CI, 0.22-1.58), while the scores in the
OA and OAE groups were 3.20 (95% CI, 2.49-3.91) and 2.00 (95% CI,
1.12-2.88), respectively (Fig. 4A and
B).
ELISA revealed that, compared with the control
group, the IL-1β level was significantly increased in the serum and
IALF of the OA group, whereas the expression of IL-4 was notably
diminished. In the OAE group, the variation in IL-1β and IL-4
levels was alleviated compared with that in the OA group, although
the differences between the two groups were not significant
(Fig. 4C). More specifically, the
serum IL-1β levels were 23.18 (95% CI, 19.77-26.58 pg/ml), 33.75
(95% CI, 28.83-38.66 pg/ml) and 27.56 (95% CI, 23.10-32.01 pg/ml)
in the control, OA and OAE groups, respectively. The IL-1β levels
in IALF were 62.31 (95% CI, 56.71-67.91 pg/mg protein), 97.48 (95%
CI, 87.26-107.69 pg/mg protein) and 87.18 (95% CI, 78.05-96.31
pg/mg protein) in the three groups, respectively. Furthermore, the
IL-4 levels in the control, OA and OAE groups were 83.98 (95% CI,
76.33-91.63 pg/ml), 67.35 (95% CI, 62.21-72.49 pg/ml) and 74.75
(95% CI, 69.02-80.48 pg/ml) in the serum, respectively; these
levels in the IALF were 206.40 (95% CI, 174.82-237.98 pg/mg
protein), 160.34 (95% CI, 144.32-176.36 pg/mg protein) and 172.63
(95% CI, 144.76-200.50 pg/mg protein), respectively (Fig. 4C).
Autophagy markers in the articular
cartilage
As demonstrated by immunohistochemical assay, LC3B
was expressed widely in the SZ and DZ of normal cartilage, with a
positive detection rate of 41 and 57% of cells, respectively
(Fig. 5). However, LC3B level in
the OA group was markedly reduced to 18 and 26% in the SZ and DZ,
respectively. In the OAE group, these values were 30 and 50%,
respectively. In contrast to LC3B levels, the number of
SQSTM1-positive cells gradually increased in the control, OA and
OAE groups, with 20, 29 and 43% of cells detected respectively in
the SZ, and 31, 45 and 58% detected respectively in the DZ. In
terms of BECLIN1, it was notably increased in the different zones
in both the OA and OAE groups compared with the control group. The
positive detection rates of BECLIN1 were 41, 72 and 76% in the SZ,
and 47, 80 and 74% in the DZ in the control, OA and OAE groups,
respectively (Fig. 5).
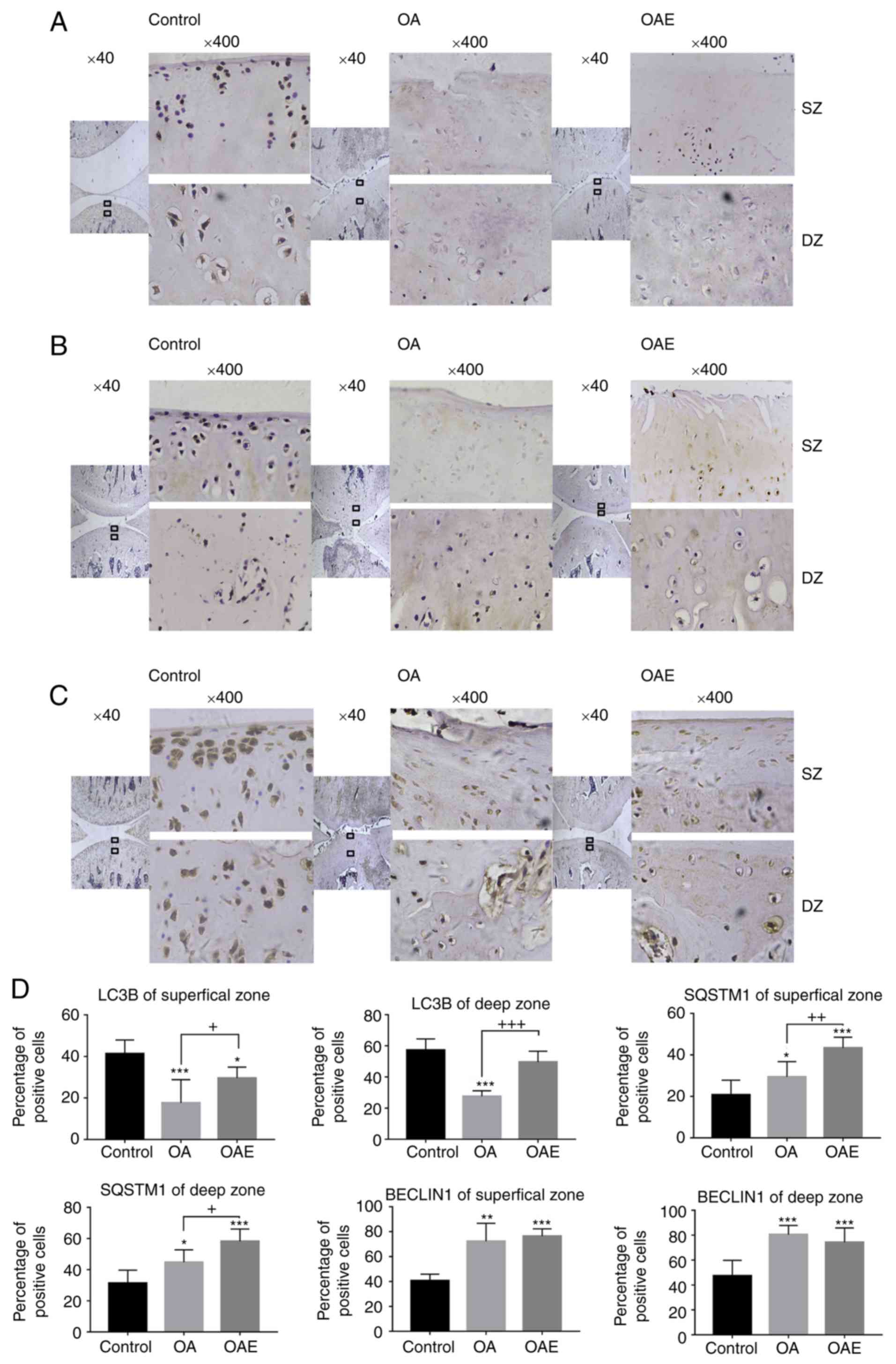 | Figure 5Immunohistochemical staining for
LC3B, SQSTM1 and BECLIN1. (A) LC3B exhibited a decrease in the SZ
and DZ of the cartilage in both the OA and OAE groups, while
treadmill exercise increased LC3B expression in OA rats. (B) SQSTM1
expression was increased in the OA and OAE groups, while treadmill
exercise evidently increased SQSTM1 level in the SZ and DZ of the
cartilage. (C) BECLIN1 expression was increased in the OA and OAE
groups in both the SZ and DZ of the cartilage, with no significant
difference between the OA and OAE groups. (D) Results are expressed
as the mean and 95% confidence interval (one-way analysis of
variance; n=5). *P<0.05, **P<0.01 and
***P<0.001 vs. control group;
+P<0.05,++P<0.01 and
+++P<0.001. OA, osteoarthritis; OAE, OA with
treadmill exercise; LC3B, light chain 3B; SQSTM1, sequestosome 1;
SZ, superficial zone; DZ, deep zone. |
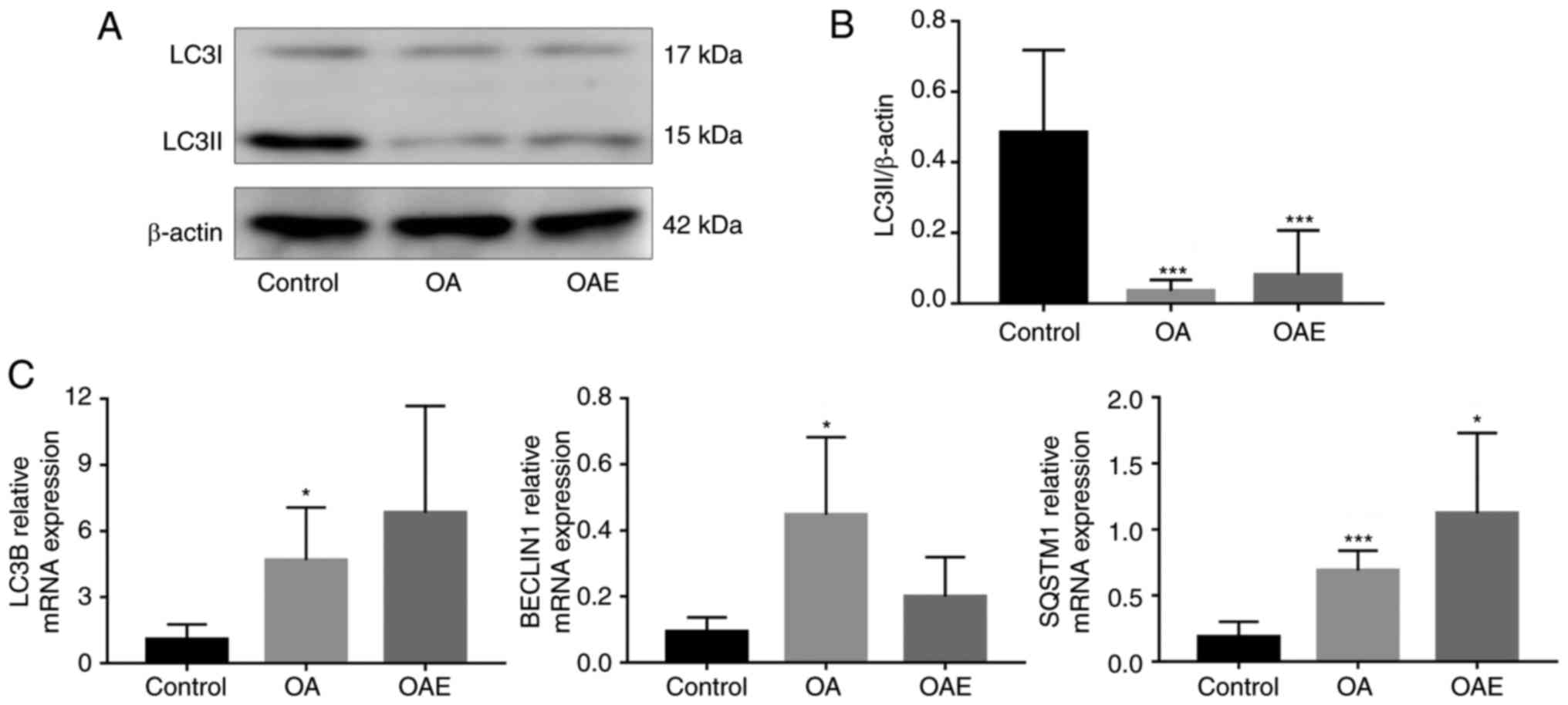
The western blot analyses of LC3B protein are
presented in Fig. 6A, while the
quantitative analysis of LC3II levels among the groups is presented
in Fig. 6B. A marked decrease in
LC3II levels was observed in both the OA and OAE groups, with a
ratio relative to the control group of 0.073 and 0.167,
respectively. RT-qPCR analysis also indicated that, compared with
the control group, the mRNA expression levels were significantly
increased in the OA group of LC3B and BECLIN1 and in the OA and OAE
groups of SQSTM1 (Fig. 6C).
Although the difference of LC3B, BECLIN1 and SQSTM1 between the OA
and OAE groups was not statistically significant, the mRNA
expression levels of LC3B and SQSTM1 exhibited an increasing
tendency in OAE. In contrast to LC3B and SQSTM1, the mRNA
expression of BECLIN1 was markedly decreased in the OAE group
compared with the OA group. The relative mRNA expression of LC3B in
the control, OA and OAE groups was 1.08 (95% CI, 0.40-1.76), 4.67
(95% CI, 2.27-7.07) and 6.83 (95% CI, 1.99-11.68), respectively.
The relative BECLIN1 levels in these groups were 0.093 (95% CI,
0.049-1.137), 0.447 (95% CI, 0.213-0.682) and 0.2 (95% CI,
0.081-0.319), respectively. Similarly, the relative SQSTM1 levels
were 0.188 (95% CI, 0.074-0.302), 0.689 (95% CI, 0.539-0.840) and
1.122 (95% CI, 0.515-1.729), respectively (Fig. 6C).
Discussion
A number of studies have illustrated the therapeutic
effect of moderate treadmill exercise in different models of OA
(9-13). It has also been reported that
systemic factors, such as body weight, and local factors, including
muscle strength, are associated with the occurrence and development
of OA (31). In the present
study, it was observed that the weight gain in the OAE group was
significantly lower compared with the other two groups (Fig. 1B). However, excessive exercise can
promote the occurrence of OA (32), suggesting that these prophylactic
treatments for OA are limited.
In the present study, an OA model induced by MIA
injection was established, with the typical pathological
alterations observed in histological analysis, while moderate
treadmill exercise was applied in the OAE group and the condition
has improved significantly. Alterations in cartilaginous damage
were represented by the degradation of type II collagen, the major
form of collagen in the matrix. Type II collagen, which is
synthesized by chondrocytes and secreted to the extracellular
matrix, may be degraded by various proteases, including ADAMTS5
(33). The synthesis of type II
collagen is inhibited by IL-1β and the expression of ADAMTS5 is
activated simultaneously. In the present study, the histological
appearance of the synovium exhibited severe inflammation in OA
group which was partially relieved in OAE group. IL-1β, a
traditional inflammatory factor, was significantly increased in the
OA group, while its expression was attenuated in the OAE group, in
both the serum and IALF. It was also observed that the level of
IL-4, an anti-inflammatory cytokine that inhibits the
proinflammatory effect of IL-1β, was inversely correlated with that
of IL-1β. These proteins have been reported to have an important
association with the occurrence of OA (34,35). Compared with our previous study
(25), the results of the present
study indicated a consistent trend in the expression levels of
IL-1β and type II collagen.
Autophagy, which is highly conserved in eukaryotes,
has been demonstrated to serve an important role in various systems
and relevant diseases, including OA. It has been reported that
increased expression of LC3B and BECLIN1 mRNA results in increased
autophagy in OA chondrocytes and cartilage at the outset, which
represents a compensatory response to stress (22). However, in cartilage with mild or
severe OA, these two proteins are strongly expressed in the OA cell
clusters, as well as the middle zone and DZ. Their expression is
however reduced in the SZ of the cartilage, and this is accompanied
by an increase in cell death and cartilage damage (36,37). To the best of our knowledge,
studies on the alteration in autophagy in an OA model following
treatment with exercise are lacking in the literature.
In the present study, the results appeared to be
consistent with the findings of a previous study demonstrating that
LC3B was decreased in different zones of the cartilage in OA
(18). However, BECLIN1 was
strongly expressed in the OA group in the SZ and DZ, and the
increase was statistically significant; these results were partly
in accordance with those obtained by Sasaki et al (22). This may be explained by the
complexity of autophagy in different periods and zones of OA, and
the distinction between a rat model induced via MIA injection and
spontaneous OA in the elderly. SQSTM1 is reported to be a link
between LC3 and ubiquitinated substrates (38), and the present study observed an
increase in its expression in OA cartilage. A possible explanation
for this effect is that the upregulated transcriptional level
supplemented the consumed protein during degradation in the
lysosome (39). Another important
finding of the current study was that exercise markedly promoted
the expression of LC3B and SQSTM1 in whole cartilage with
immunohistochemical assay compared with the OA group. Notably, no
differences were observed in BECLIN1 expression between the OA and
OAE groups, which may be partly explained by the excessive
expression in OA cartilage. In addition, western blot analysis
revealed that LC3II, which serves as a marker of autophagosome
formation, was markedly decreased in the OA group and this was
partially alleviated in the OAE group. It was also demonstrated
that, although the mRNA expression levels of the three genes of
interest was elevated following prolonged stimulation with MIA,
only BECLIN1 and SQSTM1 protein levels were increased in cartilage,
while LC3B protein was decreased. Furthermore, the decrease
detected in BECLIN1 mRNA expression in OA following treadmill
exercise may be due to the complex role of BECLIN1 in autophagy and
apoptosis (40). It can be
hypothesized that the excessive consumption of LC3B limited the
speed of autophagy and caused the accumulation of BECLIN1 and
SQSTM1 proteins in the OA group, which may explain the upregulated
mRNA expression of LC3B and simultaneous decrease of its protein
expression. Finally, the results demonstrated that the mRNA
expression levels of LC3B and SQSTM1 exhibited an increasing trend
in the OAE group in comparison with the OA group, although this
result was not found to be significant, likely due to the small
sample size.
Our previous study demonstrated that multiple
sessions of short-term exercise were associated with an improved
chondroprotective effect compared with one long session of exercise
for the same total time (25).
The important role of autophagy has been recently reported in
chondrogenesis in the regulation of the growth plate (41). In addition, a previous study
indicated that the proliferation of chondrocytes was associated
with mechanical stimulation (42). Therefore, it can be hypothesized
that the increased expression of autophagy-associated proteins in
cartilage caused by moderate treadmill exercise is associated with
chondrocyte differentiation and proliferation. The present study
may provide a basis for a new therapeutic strategy for OA via
moderate exercise. Future studies on this topic are recommended, as
several questions remain unanswered at present.
Although the effects of exercise on autophagy have
been examined in numerous organs and systems, there is little
evidence for bone and cartilage, particularly in OA. The findings
of the present study, while preliminary, suggest that moderate
exercise has a marked effect on autophagy in the cartilage in OA,
although the limitations of the detection method for autophagy
caused in animal models should be considered. Furthermore, the
limitations of small sample size and bias during observation cannot
be neglected.
In conclusion, the present study provided strong
evidence on the hypothesis that moderate treadmill exercise has a
protective effect on the articular cartilage of rats with OA
induced via MIA injection. In spite of several limitations, the
present study suggested that moderate treadmill exercise may
promote autophagy in all zones of the articular cartilage in rats
with MIA-induced OA.
Acknowledgments
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81772420).
Availability of data and material
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors’ contributions
XZ, HZ and LB conceived and designed the study. XZ,
YG, YY and XL conducted the experiments. HZ, YG and XL performed
the statistical analysis, and designed the figures. XZ wrote the
manuscript. YY and LB reviewed and edited the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Medical
Research and New Technology Ethics Committee of Shengjing Hospital,
China Medical University (Shenyang, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Vaughan MW, LaValley MP, Felson DT,
Orsmond GI, Niu J, Lewis CE, Segal NA, Nevitt MC and Keysor JJ:
Affect and incident participation restriction in adults with knee
osteoarthritis. Arthritis Care Res (Hoboken). 70:542–549. 2018.
View Article : Google Scholar
|
|
2
|
Gómez R, Villalvilla A, Largo R, Gualillo
O and Herrero- Beaumont G: TLR4 signalling in
osteoarthritis-finding targets for candidate DMOADs. Nat Rev
Rheumatol. 11:159–170. 2015. View Article : Google Scholar
|
|
3
|
Rahmati M, Mobasheri A and Mozafari M:
Inflammatory mediators in osteoarthritis: A critical review of the
state-of-the-art, current prospects, and future challenges. Bone.
85:81–90. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Barbour KE, Hootman JM, Helmick CG, Murphy
LB, Theis KA, Schwartz TA, Kalsbeek WD, Renner JB and Jordan JM:
Meeting physical activity guidelines and the risk of incident knee
osteoarthritis: A population-based prospective cohort study.
Arthritis Care Res (Hoboken). 66:139–146. 2014. View Article : Google Scholar
|
|
5
|
Blagojevic M, Jinks C, Jeffery A and
Jordan KP: Risk factors for onset of osteoarthritis of the knee in
older adults: A systematic review and meta-analysis. Osteoarthritis
Cartilage. 18:24–33. 2010. View Article : Google Scholar
|
|
6
|
Berenbaum F and van den Berg WB:
Inflammation in osteoarthritis: Changing views. Osteoarthritis
Cartilage. 23:1823–1824. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cifuentes DJ, Rocha LG, Silva LA, Brito
AC, Rueff-Barroso CR, Porto LC and Pinho RA: Decrease in oxidative
stress and histological changes induced by physical exercise
calibrated in rats with osteoarthritis induced by monosodium
iodoacetate. Osteoarthritis Cartilage. 18:1088–1095. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Boudenot A, Presle N, Uzbekov R, Toumi H,
Pallu S and Lespessailles E: Effect of interval-training exercise
on subchondral bone in a chemically-induced osteoarthritis model.
Osteoarthritis Cartilage. 22:1176–1185. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Siebelt M, Groen HC, Koelewijn SJ, de
Blois E, Sandker M, Waarsing JH, Müller C, van Osch GJ, de Jong M
and Weinans H: Increased physical activity severely induces
osteoarthritic changes in knee joints with papain induced
sulfate-glycosaminoglycan depleted cartilage. Arthritis Res Ther.
16:R322014. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nam J, Perera P, Liu J, Wu LC, Rath B,
Butterfield TA and Agarwal S: Transcriptome-wide gene regulation by
gentle treadmill walking during the progression of
monoiodoacetate-induced arthritis. Arthritis Rheum. 63:1613–1625.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Iijima H, Aoyama T, Ito A, Yamaguchi S,
Nagai M, Tajino J, Zhang X and Kuroki H: Effects of short-term
gentle treadmill walking on subchondral bone in a rat model of
instability-induced osteoarthritis. Osteoarthritis Cartilage.
23:1563–1574. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Iijima H, Ito A, Nagai M, Tajino J,
Yamaguchi S, Kiyan W, Nakahata A, Zhang J, Wang T, Aoyama T, et al:
Physiological exercise loading suppresses post-traumatic
osteoarthritis progression via an increase in bone morphogenetic
proteins expression in an experimental rat knee model.
Osteoarthritis Cartilage. 25:964–975. 2017. View Article : Google Scholar
|
|
13
|
Galois L, Etienne S, Grossin L,
Watrin-Pinzano A, Cournil-Henrionnet C, Loeuille D, Netter P,
Mainard D and Gillet P: Dose-response relationship for exercise on
severity of experimental osteoarthritis in rats: A pilot study.
Osteoarthritis Cartilage. 12:779–786. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hong Y, Kim H, Lee Y, Lee S, Kim K, Jin Y,
Lee SR, Chang KT and Hong Y: Salutary effects of melatonin combined
with treadmill exercise on cartilage damage. J Pineal Res.
57:53–66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mizushima N: Physiological functions of
autophagy. Curr Top Microbiol Immunol. 335:71–84. 2009.PubMed/NCBI
|
|
16
|
Milner PI, Fairfax TP, Browning JA,
Wilkins RJ and Gibson JS: The effect of O2 tension on pH
homeostasis in equine articular chondrocytes. Arthritis Rheum.
54:3523–3532. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim J, Kundu M, Viollet B and Guan KL:
AMPK and mTOR regulate autophagy through direct phosphorylation of
Ulk1. Nat Cell Biol. 13:132–141. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Caramés B, Taniguchi N, Otsuki S, Blanco
FJ and Lotz M: Autophagy is a protective mechanism in normal
cartilage, and its aging-related loss is linked with cell death and
osteoarthritis. Arthritis Rheum. 62:791–801. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Caramés B, Taniguchi N, Seino D, Blanco
FJ, D’Lima D and Lotz M: Mechanical injury suppresses autophagy
regulators and pharmacologic activation of autophagy results in
chondroprotection. Arthritis Rheum. 64:1182–1192. 2012. View Article : Google Scholar
|
|
20
|
López de Figueroa P, Lotz MK, Blanco FJ
and Caramés B: Autophagy activation and protection from
mitochondrial dysfunction in human chondrocytes. Arthritis
Rheumatol. 67:966–976. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cetrullo S, D’Adamo S, Guidotti S, Borzì
RM and Flamigni F: Hydroxytyrosol prevents chondrocyte death under
oxidative stress by inducing autophagy through sirtuin 1-dependent
and -independent mechanisms. Biochim Biophys Acta. 1860.1181–1191.
2016.
|
|
22
|
Sasaki H, Takayama K, Matsushita T, Ishida
K, Kubo S, Matsumoto T, Fujita N, Oka S, Kurosaka M and Kuroda R:
Autophagy modulates osteoarthritis-related gene expression in human
chondrocytes. Arthritis Rheum. 64:1920–1928. 2012. View Article : Google Scholar
|
|
23
|
Srinivas V, Bohensky J and Shapiro IM:
Autophagy: A new phase in the maturation of growth plate
chondrocytes is regulated by HIF, mTOR and AMP kinase. Cells
Tissues Organs. 189:88–92. 2009. View Article : Google Scholar :
|
|
24
|
Watson K and Baar K: mTOR and the health
benefits of exercise. Semin Cell Dev Biol. 36:130–139. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang Y, Wang Y, Kong Y, Zhang X and Bai L:
The effects of different frequency treadmill exercise on lipoxin A4
and articular cartilage degeneration in an experimental model of
monosodium iodoacetate-induced osteoarthritis in rats. PLoS One.
12:e01791622017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guzman RE, Evans MG, Bove S, Morenko B and
Kilgore K: Mono-iodoacetate-induced histologic changes in
subchondral bone and articular cartilage of rat femorotibial
joints: An animal model of osteoarthritis. Toxicol Pathol.
31:619–624. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pritzker KP, Gay S, Jimenez SA, Ostergaard
K, Pelletier JP, Revell PA, Salter D and van den Berg WB:
Osteoarthritis cartilage histopathology: Grading and staging.
Osteoarthritis Cartilage. 14:13–29. 2006. View Article : Google Scholar
|
|
28
|
Krenn V, Morawietz L, Burmester GR, Kinne
RW, Mueller-Ladner U, Muller B and Haupl T: Synovitis score:
Discrimination between chronic low-grade and high-grade synovitis.
Histopathology. 49:358–364. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guilak F, Alexopoulos LG, Upton ML, Youn
I, Choi JB, Cao L, Setton LA and Haider MA: The pericellular matrix
as a transducer of biomechanical and biochemical signals in
articular cartilage. Ann NY Acad Sci. 1068:498–512. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
31
|
Roos EM and Arden NK: Strategies for the
prevention of knee osteoarthritis. Nat Rev Rheumatol. 12:92–101.
2016. View Article : Google Scholar
|
|
32
|
Bomer N, Cornelis FM, Ramos YF, den
Hollander W, Storms L, van der Breggen R, Lakenberg N, Slagboom PE,
Meulenbelt I and Lories RJ: The effect of forced exercise on knee
joints in Dio2(−/−) mice: Type II iodothyronine
deiodinase-deficient mice are less prone to develop OA-like
cartilage damage upon excessive mechanical stress. Ann Rheum Dis.
75:571–577. 2016. View Article : Google Scholar
|
|
33
|
Rasheed Z, Rasheed N and Al-Shaya O:
Epigallocatechin-3- O-gallate modulates global microRNA expression
in interleukin-1β-stimulated human osteoarthritis chondrocytes:
Potential role of EGCG on negative co-regulation of microRNA-140-3p
and ADAMTS5. Eur J Nutr. 57:917–928. 2018. View Article : Google Scholar
|
|
34
|
Jeon JE, Schrobback K, Meinert C, Sramek
V, Hutmacher DW and Klein TJ: Effect of preculture and loading on
expression of matrix molecules, matrix metalloproteinases, and
cytokines by expanded osteoarthritic chondrocytes. Arthritis Rheum.
65:2356–2367. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wojdasiewicz P, Poniatowski ŁA and
Szukiewicz D: The role of inflammatory and anti-inflammatory
cytokines in the pathogenesis of osteoarthritis. Mediators Inflamm.
2014.561459:2014.
|
|
36
|
Barranco C: Osteoarthritis: Activate
autophagy to prevent cartilage degeneration. Nat Rev Rheumatol.
11:1272015. View Article : Google Scholar
|
|
37
|
Caramés B, Olmer M, Kiosses WB and Lotz
MK: The relationship of autophagy defects to cartilage damage
during joint aging in a mouse model. Arthritis Rheumatol.
67:1568–1576. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bjørkøy G, Lamark T, Brech A, Outzen H,
Perander M, Overvatn A, Stenmark H and Johansen T: p62/SQSTM1 forms
protein aggregates degraded by autophagy and has a protective
effect on huntingtin-induced cell death. J Cell Biol. 171:603–614.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sanchez AM, Bernardi H, Py G and Candau
RB: Autophagy is essential to support skeletal muscle plasticity in
response to endurance exercise. Am J Physiol Regul Integr Comp
Physiol. 307:R956–R969. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kang R, Zeh HJ, Lotze MT and Tang D: The
Beclin 1 network regulates autophagy and apoptosis. Cell Death
Differ. 18:571–580. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kang X, Yang W, Feng D, Jin X, Ma Z, Qian
Z, Xie T, Li H, Liu J, Wang R, et al: Cartilage-specific autophagy
deficiency promotes ER stress and impairs chondrogenesis in
PERK-ATF4-CHOP-dependent manner. J Bone Miner Res. 32:2128–2141.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Khozoee B, Mafi P, Mafi R and Khan WS:
Mechanical stimulation protocols of human derived cells in
articular cartilage tissue engineering-a systematic review. Curr
Stem Cell Res Ther. 12:260–270. 2017. View Article : Google Scholar
|