Introduction
Brain ischemic stroke, a serious disease in the
central nervous system (CNS), is becoming a prominent public health
risk (1-3). Generally, vascular recanalization to
obtain timely reperfusion is the preferred treatment in clinical
practice. However, reperfusion can induce further unexpected brain
injury, termed cerebral ischemia reperfusion injury (CIR) (4). Previous studies have demonstrated
that the inflammatory response has an important role in the outcome
of stroke (5,6). Thus, anti-inflammatory therapy may
lessen neurological deficits by ischemic stroke and it may serve as
a potential therapeutic strategy following ischemic stroke
(3).
Biliverdin (BV) is a metabolite of heme catabolism
that has a protective role in lung graft injury, hemorrhagic shock
and resuscitation-induced lung injury through anti-inflammatory and
antioxidant mechanisms (7,8). A
previous study has suggested that exogenously administered carbonic
oxide (CO) and BV have potent cytoprotective effects on intestinal
ischemia reperfusion injury (9).
In addition, BV administration can ameliorate CIR in rats, and the
mechanism may be related to the downregulation of proinflammatory
factors. To date, two pathways are known to be involved in the
anti-inflammatory mechanism of BV. By activating the nitric
oxide-dependent BV reductase, BV reduces the expression of toll
like receptor-4 (TLR-4) in murine macrophages. BV regulates the
lipopolysaccharide (LPS)-mediated expression of complement C5a
receptor 1 via the mammalian target of rapamycin (mTOR) pathway
(10,11). While the anti-inflammatory
mechanism of BV has been the focus of previous studies, the
molecular network upstream and downstream of BV is largely
unknown.
MicroRNAs (miRNAs), small non-coding RNAs of 21-23
bp in length, serve crucial roles in several biological processes.
By binding to the 3'-untranslated region (UTR) of target mRNA,
miRNAs induce mRNA cleavage or translation inhibition (12). Each miRNA has multiple potential
mRNA targets, and therefore, a broad-spectrum gene expression can
be affected by an specific miRNA (13,14). Recent studies have demonstrated
that miRNA inhibitory activity can be quantified by examining their
target mRNA expression levels (15). However, the relation between miRNA
and mRNA during CIR pathogenesis following BV administration
remains to be determined.
In the present study, it was hypothesized that miRNA
and mRNA expression may be regulated by the BV anti-inflammatory
mechanism in CIR rats. To explore the potential miRNA-mRNA
regulatory mechanism and relevant signaling pathways, a rat middle
cerebral artery occlusion (MCAO) model was first established and
then BV treatment was performed. Subsequently, the expressional
network of miRNAs and mRNAs was examined by microarray,
bioinformatics, and integrated genomics analyses. Reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) was
performed to validate the reliability of microarray data. Lastly,
miRNA204-5p was confirmed to directly target ETS proto-oncogene 1
(Ets1) following BV administration in CIR rats. To the best of our
knowledge, this is the first report of the integrated molecular
network of miRNA with gene microarray expression following BV
treatment in CIR rats. The present data suggested that miRNAs, such
as miRNA204-5p, can act as key regulators to target Ets1 in
endogenous responses to stroke. Therefore, manipulation of these
miRNAs might serve as a novel diagnosis or therapeutic indication
for acute stroke patients.
Materials and methods
Animals and experimental groups
Animal care and all experimental protocols involving
animals were approved by the Animal Care and Welfare Committee of
Kunming Medical University (Kunming, China). A total of 36 adult
male Sprague-Dawley (SD) rats (specific pathogen-free; age, 8-12
weeks; weight, 220±10 g; Kunming Medical University Laboratory
Animal Research Center, Kunming, China) were used in the study. All
rats were housed at room temperature (21-25°C) with 45-50% humidity
and a 12-h light/dark cycle, with free access to soft food and tap
water. Rats were randomly divided into three groups (Table I): Sham group (S, sham operation);
Brain ischemic and vehicle control group (C, CIR + normal saline);
and brain ischemic with BV treatment group (BV, CIR+BV).
 | Table IExperimental groups and methods. |
Table I
Experimental groups and methods.
| Number of rats
|
|---|
| Group | TTC stain/NSS | Microarray | qPCR |
|---|
| S | 3 | 3 | 6 |
| C | 3 | 3 | 6 |
| BV | 3 | 3 | 6 |
Animal model of MCAO and BV
administration
The right middle cerebral artery was occluded
according to the standard operation procedures for MCAO in rats
(16). Briefly, animals were
anaesthetized by an intraperitoneal (ip) injection of 5 mg/kg
xylazine HCl and 40 mg/kg ketamine HCl, and then a midline neck
incision was made. The right common carotid artery (RCCA), right
external carotid artery, and right internal carotid artery were
isolated for inserting the nylon monofilament (diameter 0.24 mm;
Johnson & Johnson, New Brunswick, NJ, USA) through a small
incision in the RCCA. The monofilament was fixed in position
tightly and then the incision was sutured. Rat body temperature was
maintained at 36.5±0.5°C using a heating lamp. A laser Doppler
system (Peri-Flux System 5000; Perimed, Jarfalla, Sweden) was used
to supervise regional cerebral blood flow (rCBF). Rats in which
rCBF did not drop <20% of the baseline levels following MCAO
were excluded from analysis. Sham group rats underwent the same
procedures without inserting a nylon thread. After 2 h of tMCAO,
CBF was recovered by removing the nylon thread, and the incision
was closed.
BV HCl (Frontier Scientific, Inc., Logan, UT, USA)
was dissolved in 0.2 N NaOH and adjusted to final pH 7.4 with HCl.
As previously published, BV was diluted in saline and injected (35
mg/kg ip) to rats 15 min prior to reperfusion, then once again 4 h
after reperfusion, and twice per day thereafter (17). In the vehicle control group, the
same volume of saline alone was injected in the same way.
Stroke-onset was assessed by circadian rhythm disturbances in heart
rate (HR) and mean arterial blood pressure (MABP).
Neural behavioral test
Rats from the three groups were scored by an
evaluator with neurological severity scores (NSS), as previously
reported (17), at day 1 and 2
post-reperfusion. The NSS includes four physiological function
evaluation scores: Feeling, movement, reflection and balance.
Scores 1-6 indicate mild injury, 7-12, moderate injury, and 13-18,
severe injury. The neural behavioral test was conducted by an
evaluator that was blinded to the treatments.
Evaluation of cerebral infarct volume
indicated by 2,3,5-triphenyltetrazolium chloride (TTC)
staining
Whole brain tissue was harvested after 2 h of
ischemia, followed by 48 h of reperfusion, and the integrity of the
brain was maintained upon removal (n=3 for each group). Brain
tissue from bregma was cut into 2 mm thick coronal slices. Brain
slices were then incubated in 2% TTC solution at 37°C for 30 min in
the dark, as previously described (18). After staining, the sections were
washed with PBS (3 times, 1 min each) and fixed in 4%
paraformaldehyde for 24 h. Color images of these sections were
directly obtained with a stereomicroscope by an evaluator that was
blinded to the treatments, and the infarct areas of each section
were measured with ImageJ 1.4 software (National Institutes of
Health, Bethesda, MD, USA). To compensate for the effect of brain
edema following cerebral infarction, the corrected infarct volume
was calculated as the sum of the infarct areas multiplied by the
section thickness (2 mm), and expressed as a % of the contralateral
(non-occluded) hemisphere.
RNA isolation
Expression levels of inflammatory factors were the
most obvious at 2 h ischemia and 6 h following reperfusion,
according to our previous study (17); therefore, the ipsilateral ischemic
cortex at 6 h following reperfusion was selected to perform miRNA
and mRNA assays. In the C and BV groups, the lesion tissue from the
ipsilateral ischemic cortex was obtained from rats at 2 h ischemia
and 6 h post-reperfusion, with the corresponding cortex harvested
from the S group as well. Three samples of each group were pooled
into one S pool, one C pool and one BV pool for microarray
analysis, and each pool was on a different microarray chip.
Briefly, after rats were anaesthetized by an ip injection of 5
mg/kg xylazine HCl and 40 mg/kg ketamine HCl, the brain cortex of
ischemia regions was harvested and fresh-frozen in liquid nitrogen.
Total RNA was isolated using TRIzol (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and purified with RNeasy mini
kit (Qiagen GmbH, Hilden, Germany), according to the manufacturer's
instructions. RNA quality and quantity was measured with a NanoDrop
spectrophotometer (ND-1000; NanoDrop Technologies; Thermo Fisher
Scientific, Inc.) and RNA integrity was determined by gel
electrophoresis.
miRNA microarray analysis
After total RNA samples were labelled using the
miRCURY Hy3/Hy5 Power labeling kit (Exiqon; Qiagen), they were
hybridized to a 12-Bay Hybridization System (Nimblegen Systems,
Inc., Madison, WI, USA) to detect miRNA expressional profiles.
Then, the positive signal was recorded using the Axon GenePix 4000B
micro-array scanner (Axon Instruments; Molecular Devices, LLC,
Sunnyvale, CA, USA). Subsequently, the scanned images were imported
into GenePix Pro 6.0 software (Axon Instruments; Molecular Devices,
LLC) for grid alignment and data extraction. Replicated miRNAs were
averaged and miRNAs with intensities ≥30 in all samples were
selected for calculating the normalization factor. Expressed data
were normalized using the Median normalization. Following
normalization, differentially expressed miRNAs between two samples
were filtered through fold change. The miRNA array results
following normalization and fold changes are listed in Table II. Finally, hierarchical
clustering was performed to display distinguishable miRNA
expression profiles among samples.
| Table IImiRNA array ratios following
normalization and fold change. |
Table II
miRNA array ratios following
normalization and fold change.
| microRNA array
ratio after normalization
| Fold change
| |
|---|
| microRNA | S | C | BV | S vs. C | C vs. BV | Change trend |
|---|
| rno-miR-324-5p | 0.09 | 0.62 | 0.04 | 6.59 | 17.59 | Up/down |
| rno-miR-3072 | 0.29 | 1.15 | 0.23 | 3.91 | 5.03 | Up/down |
| rno-miR-124-3p | 26.93 | 126.82 | 37.02 | 4.71 | 3.43 | Up/down |
| rno-miR-375-5p | 0.06 | 0.264 | 0.07 | 4.51 | 3.62 | Up/down |
|
rno-miR-3573-3p | 6.33 | 28.02 | 10.82 | 4.43 | 2.59 | Up/down |
| rno-miR-150-5p | 2.11 | 8.58 | 2.35 | 4.07 | 3.65 | Up/down |
| rno-miR-935 | 0.20 | 0.73 | 0.23 | 3.60 | 3.21 | Up/down |
|
rno-miR-133a-3p | 0.16 | 0.57 | 0.11 | 3.53 | 4.94 | Up/down |
| rno-miR-539-3p | 1.37 | 4.40 | 0.75 | 3.20 | 5.88 | Up/down |
| rno-miR-370-3p | 0.14 | 0.44 | 0.16 | 3.14 | 2.77 | Up/down |
|
rno-miR-181b-5p | 3.17 | 0.03 | 1.51 | 116.9 | 55.69 | Down/up |
| rno-miR-204-5p | 1.55 | 0.02 | 0.82 | 68.77 | 36.55 | Down/up |
|
rno-miR-664-1-5p | 0.81 | 0.02 | 0.60 | 32.67 | 24.00 | Down/up |
| rno-miR-136-5p | 19.47 | 0.64 | 14.36 | 30.41 | 22.42 | Down/up |
|
rno-miR-126a-5p | 4.24 | 0.11 | 2.24 | 37.64 | 19.86 | Down/up |
| rno-miR-124-5p | 0.75 | 0.02 | 0.30 | 47.74 | 19.25 | Down/up |
| rno-miR-363-5p | 2.09 | 0.06 | 0.78 | 35.69 | 13.28 | Down/up |
| rno-miR-27a-3p | 4.85 | 0.23 | 2.88 | 21.51 | 12.76 | Down/up |
| rno-miR-29b-3p | 24.68 | 1.41 | 15.96 | 17.51 | 11.32 | Down/up |
|
rno-miR-376a-5p | 0.84 | 0.02 | 0.34 | 41.32 | 16.92 | Down/up |
Gene microarray analysis
Sample labeling and array hybridization were
performed according to the Agilent One-Color Microarray-Based Gene
Expression Analysis protocol (Agilent Technologies, Inc., Santa
Clara, CA, USA). Briefly, mRNA was purified from total RNA after
removal of rRNA (mRNA-ONLY Eukaryotic mRNA Isolation kit;
Epicentre; Illumina, Inc., San Diego, CA, USA). Then, each sample
was amplified and transcribed into fluorescent cRNA along the
entire length of the transcripts without 3' bias utilizing a random
priming method (Arraystar Flash RNA Labeling kit; Arraystar, Inc.
Rockville, MD, USA). The labeled cRNAs were purified with RNeasy
Mini kit (Qiagen GmbH). Labeled samples hybridized for 17 h at 65°C
in an Agilent hybridization oven. Then, the hybridized arrays were
washed, fixed and scanned with the Agilent DNA Microarray Scanner
(part no. G2505C). Agilent Feature Extraction software (version
11.0.1.1; Agilent Technologies, Inc.) was used to analyze acquired
array images. Quantile normalization and subsequent data processing
were performed with GeneSpring GX v12.1 software package (Agilent
Technologies, Inc.). Differentially expressed mRNAs between the two
samples were identified through fold change filtering.
qPCR verification for miRNA microarray
data
To validate the microarray data, first 20
differentially expressed miRNAs were selected for qPCR. Total RNA
from the brain cortex of ischemia region was extracted with TRIzol
(Thermo Fisher Scientific, Inc.). Poly(A) Tailing and reverse
transcription were performed with the All-in-one miRNA qRNA miRNA
qPCR Start kit (Yijing, Guangzhou, China) in a DNA thermal cycler
(T100 TM; Bio-Rad Laboratories, Inc., Hercules, CA, USA) at 85°C
for 5 min. Then, qPCR was performed using a SYBR-Green RT-PCR
Master Mix kit (Takara Biotechnology Co., Ltd., Dalian, China). The
annealing temperature was 53°C. The reaction was performed at 95°C
for 2 min and 40 cycles of 95°C for 20 sec, 53°C for 30 sec and
60°C for 40 sec. The primers used (Sangon Biotech Co., Ltd.,
Shanghai, China) are listed in Table III. The U6 gene was used as the
internal control. Relative levels of miRNA were calculated using
the formula 2−ΔΔCq (19). At least three independent
biological replicates were used for miRNA.
| Table IIIPrimer sequence cat. nos for
microRNAs. |
Table III
Primer sequence cat. nos for
microRNAs.
| microRNA | Catalog number |
|---|
| rno-miR-29c-3p | RmiRQP0375 |
|
rno-miR-181b-5p | RmiRQP0234 |
| rno-miR-204-5p | RmiRQP0306 |
|
rno-miR-664-1-5p | RmiRQP3276 |
| rno-miR-136-5p | RmiRQP0173 |
|
rno-miR-126a-5p | RmiRQP0098 |
| rno-miR-124-5p | RmiRQP0073 |
|
rno-miR-376a-5p | RmiRQP1253 |
| rno-miR-363-5p | RmiRQP0450 |
| rno-miR-27a-3p | RmiRQP0359 |
| rno-miR-29b-3p | RmiRQP0373 |
| rno-miR-324-5p | RmiRQP0412 |
| rno-miR-3072 | RmiRQP2830 |
| rno-miR-124-3p | RmiRQP0074 |
| rno-miR-375-5p | RmiRQP3367 |
|
rno-miR-3573-3p | RmiRQP1852 |
| rno-miR-150-5p | RmiRQP0210 |
| rno-miR-935 | RmiRQP0840 |
|
rno-miR-133a-3p | RmiRQP0166 |
| rno-miR-539-3p | RmiRQP3241 |
| rno-miR-370-3p | RmiRQP0456 |
| U6 | RmiRQP9003 |
Bioinformatics prediction
Predicted target genes of candidate miRNAs were
determined using three bioinformatics prediction tools:
TargetScanv6.2 (http://www.targetscan.org/mamm_31/), miRmap
(http://mirmap.ezlab.org/), and miRDB (http://www.mirdb.org/miRDB/). The selection criteria
were correlation >0.99 or correlation <-0.99, and P-value
<0.05. The genes that overlapped in all three databases were
selected for further functional analyses. The miRNAs and predicted
mRNA target genes were then subjected to Gene Ontology (GO) and
Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses using David
v6.7 (http://david.abcc.ncifcrf.gov/)
online. Cytoscape version 3.4.0 was used to visualize the
connections between the miRNAs and their predicted gene targets.
Additionally, the sequences of the target genes were obtained from
the TargetScanv6.2 (http://www.targetscan.org/mamm_31/), miRmap
(http://mirmap.ezlab.org/) and miRDB (http://www.mirdb.org/miRDB/) databases, and the
complementarity of the miRNAs with the 3'UTR regions of target
genes was examined by manual method.
Integration of miRNA and mRNA
First, significant differentially expressed mRNAs
were identified by fold change ≥2 of each data set for further data
analysis. Further testing involved determination of the overlap
between predicted gene targets of candidate miRNAs and mRNAs array.
In order to select relatively novel genes, those overlapped mRNAs
were searched in PubMed web source (https://www.ncbi.nlm.nih.gov/pubmed/), and candidate
genes were verified by qPCR.
qPCR verification for candidate
genes
A total of 33 candidate genes were verified using
qPCR. Total RNA from the brain cortex of ischemia region was
extracted with TRIzol (Thermo Fisher Scientific, Inc.). After RNA
samples were reverse transcribed into cDNA using the RevertAidTM
First Strand cDNA Synthesis kit (cat. no. K1622; Thermo Fisher
Scientific, Inc.), qPCR was performed using a SYBR Green RT-PCR
Master Mix kit (Takara Biotechnology Co., Ltd.). The annealing
temperature was 53°C. The reaction was performed at 95°C for 2 min
and 40 cycles of 95°C for 20 sec, 53°C for 30 sec and 60°C for 40
sec. The primers are listed in Table
IV. β-actin was used as the internal control. Relative
expression levels of mRNA were calculated using the formula
2−ΔΔCq (19). At least
three independent biological replicates were used for mRNA.
| Table IVPrimer sequences and annealing
temperatures. |
Table IV
Primer sequences and annealing
temperatures.
| Gene | Primer | Sequence | Annealing
temperature |
|---|
| Sox7 | Forward |
CCTCTGCAGTCACCTTAGCC | 51°C |
| Reverse |
GTCCATGTCGCCAAGAAGTT | |
| Fbxo33 | Forward |
TCCAGCACATACCTCAGCAG | 50°C |
| Reverse |
TGGTGTTGGCAATGGAGTTA | |
| Plekha8 | Forward |
TGCTGAGATCAACCTGCAAC | 50°C |
| Reverse |
GCAATCTGCCTTTGTGGATT | |
| Ets1 | Forward |
ACTGTGTGCCCTGGGTAAAG | 53°C |
| Reverse |
GAGTTCTTCCGAGCTGATGG | |
| Gcnt2 | Forward |
AGCGAAAGGACGTCTGTCAT | 51°C |
| Reverse |
GTAGACGTTCTGGGGCATGT | |
| Csrnp1 | Forward |
GCTTTCAGTGTCCGGAGTTC | 53°C |
| Reverse |
GCCATCACAGTGACAACCAC | |
| En2 | Forward |
CCAGGTCTCGAAAACCAAAG | 50°C |
| Reverse |
ACCGCCAAAGTGTTCTTGTT | |
| Rgs1 | Forward |
AACTCCTTGCCAACCAGATG | 50°C |
| Reverse |
TGTGGGAGTTGGTGTTTTGA | |
| Litaf | Forward |
TCCAGGACCTTACCAAGCAG | 50°C |
| Reverse |
AGGAAGGACAGCACATCTGG | |
| Rnd3 | Forward |
CGGACAGATGTCAGCACACT | 51°C |
| Reverse |
GTGGCCCTCTGTGATTTGTT | |
| Zc3h12d | Forward |
GAAGGATGGCGTCATTGTCT | 51°C |
| Reverse |
TCTCCGGGTGGTAGAATCTG | |
| P4hb | Forward |
CAATTTTGCCACCACTTCCT | 50°C |
| Reverse |
CTTCCACCTCATTGGCTGTT | |
| Mthfd2 | Forward |
TCCTTGCGGACATTGTGATA | 50°C |
| Reverse |
TCTGGCCTGAGCACTTTCTT | |
| Mafk | Forward |
GACTAATCCCAAGCCCAACA | 51°C |
| Reverse |
TCCAGCTCCTCCTTCTGTGT | |
| B4galt1 | Forward |
GGCCTGAAGAGCAACTTGAC | 50°C |
| Reverse |
AATGATGGCCACCTTGTGA | |
| Slc25a25 | Forward |
CTTGATGGGCAACTGGACTT | 51°C |
| Reverse |
ACTCGTTCCAGTCGATGGTC | |
| Esm1 | Forward |
TGATTTCGGTGACGAGTTTG | 49°C |
| Reverse |
TCTCTCACAGCATTGCCATC | |
| Vps37c | Forward |
GCATGAAGATCGAGGAGGAG | 54°C |
| Reverse |
TAGGGCAAAGGGTAGGGAGT | |
| Zmiz1 | Forward |
ACTCTGTCGCACAGTGATGG | 51°C |
| Reverse |
TGGAGAACTGTTGCTGTTGC | |
| Rhoq | Forward |
TTCGACCACTACGCAGTCAG | 50°C |
| Reverse |
AAGGGGACATTTGGTGCATA | |
| Plp2 | Forward |
CTACTCCTCCCTGTCGGTGA | 51°C |
| Reverse |
TCGTAGCCAAAGAGCAAGGT | |
| Zkscan1 | Forward |
GGTTCAGGCGCTTCTGTTAC | 53°C |
| Reverse |
GCCTCTTCTCCACTGTCAGG | |
| Dgkg | Forward |
TGTCCACCAACGCTGTGTAT | 51°C |
| Reverse |
GGTAGGCAGCAAAATGTGGT | |
| Megf9 | Forward |
TACTCCTCCTCCCCCAGAGT | 51°C |
| Reverse |
GTGACACCCAGTTTGCATTG | |
| Zbtb3 | Forward |
CAGGTCCAGGGCAGTATCAT | 53°C |
| Reverse |
GAGGTCCCCTGACTGTGTGT | |
| Guca1b | Forward |
TGGAACACAAGCTCAAGTGG | 52°C |
| Reverse |
AGACAGCTGACCGTCTCCAT | |
| Pnrc1 | Forward |
GAAAATTTCCCCTCCCCATA | 50°C |
| Reverse |
TCTTCCCTCGGTTTTCCTTT | |
| Ankrd12 | Forward |
CAGCAGTGGGCACAGAGATA | 51°C |
| Reverse |
GGCTTCCACAGACGAAAGAG | |
| Kcnab3 | Forward |
ATTTGCCAATCGTTCAGACC | 51°C |
| Reverse |
TCTCCACCTTCTCCCTCTGA | |
| Map2k6 | Forward |
AGTGGACTGTCCGTTTACCG | 50°C |
| Reverse |
TGAGCACATTTGAAGGCTTG | |
| Scn4b | Forward |
GCCACCACCATCTACGCTAT | 53°C |
| Reverse |
CTCTAGGGTGATGCGGTCAT | |
| Cmklr1 | Forward |
GTGCCCCTACCACACACTCT | 53°C |
| Reverse |
TGGTGAAGCTCCTGTGACTG | |
| β-actin | Forward |
TACTCCTGCTTGCTGATCCA | 53°C |
| Reverse |
CTGTCCACCTTCCAGCAGA | |
Luciferase assay
The miRNA mimics and the 3'UTR plasmids of Ets1,
glucosaminyl N-acetyl transferase 2 (GcnT2), Rho family GTPase 3
(Rnd3), prolyl 4-hydroxylase subunit β (P4hb), MAF bZIP
transcription factor B (mafb), and mitogen-activated protein kinase
kinase 6 (map2K6) were supplied by RiboBio Co., Ltd. (Guangzhou,
China). 293T cells (5×104 per well) were seeded in a
96-well plate 24 h prior to transfection. Each well was transfected
with 1 ng/µl of the 3'UTR luciferase vector and 50 nM of the
miRNA mimic using a FECT transfection kit (RiboBio Co., Ltd.). The
assay was performed using the Dual-Luciferase Reporter Assay System
(Promega Corporation, Madison, WI, USA) 48 h after transfection,
using Renilla luciferase as the reporter and firefly luciferase as
the control. Luminescence was measured with a Synergy 2 microplate
reader (BioTek Instruments, Inc., Winooski, VT, USA).
Statistical analysis
All data were represented as mean ± standard
deviation. For multiple group comparisons, one-way analysis of
variance with Tukey's post hoc test was applied. All statistical
analyses were performed with SPSS 19.0 software (IBM Corp., Armonk,
NY, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
BV treatment decreases cerebral
infarction volume and may improve functional recovery
To examine whether BV treatment following MCAO could
reduce the functional deficits caused by ischemic brain injury, TTC
staining was used to detect the infarct volume at 48 h
post-reperfusion. The borders of the TTC stain enclosing the white
infarct area were readily distinguishable in contrast to the red
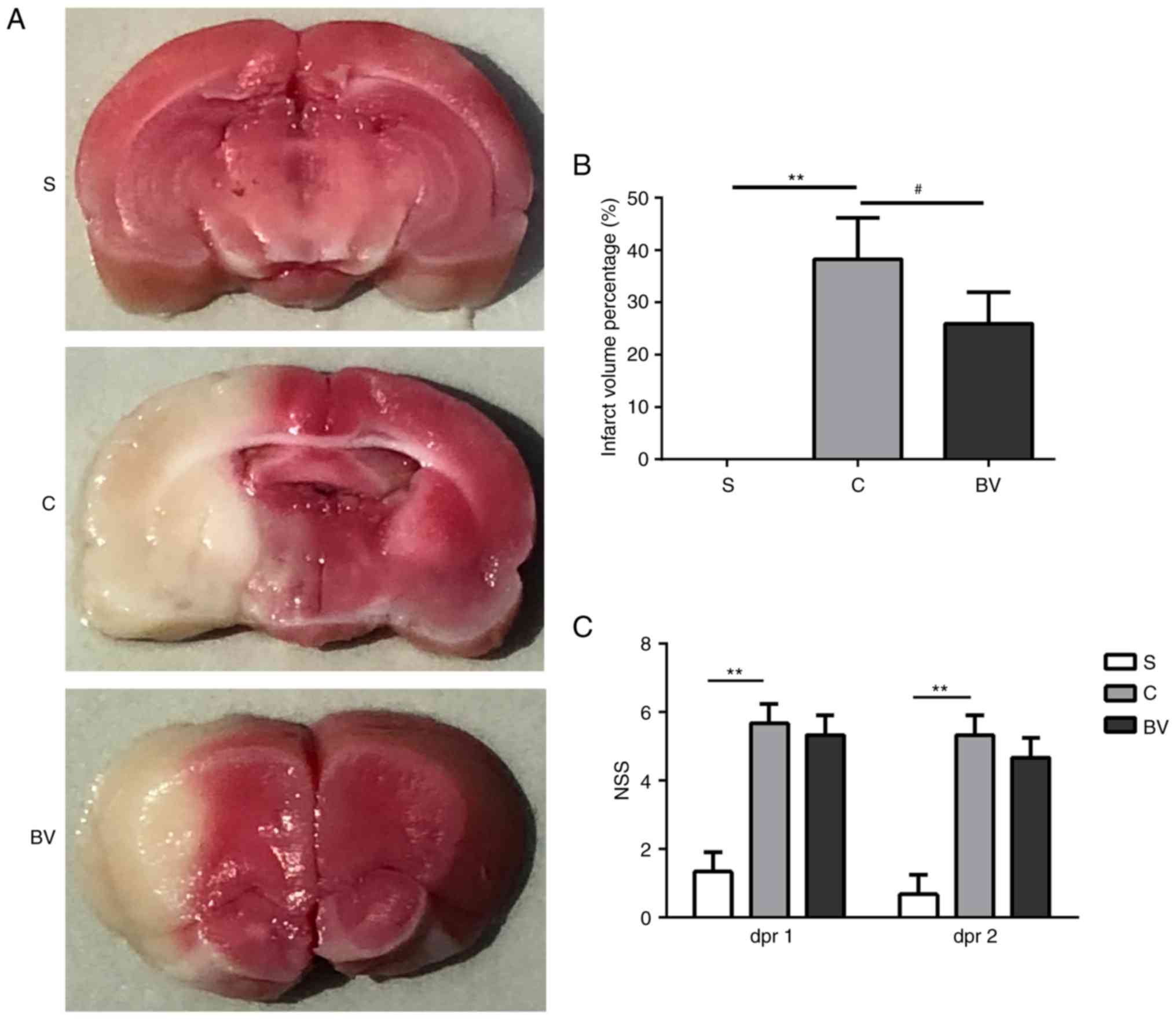
color which is the normal tissue (Fig. 1A). The infarct volumes % observed
in the C group was significantly larger compared with the S group
(38.25±7.87 vs. 00.00±0.00; P<0.01; Fig. 1A and B). The infarct volumes
observed in the BV group were obviously decreased compared with the
C group (38.25±7.87 vs. 25.93±6.02; P<0.05; Fig. 1A and B).
To determine the effect of BV treatment on the
neurological function following MCAO, NSS was used to assess the
functional recovery. The NSS test was performed at day 1 and 2
post-reperfusion. Compared with the S group, functional deficits
were impaired by ischemic insult in the C group (P<0.01;
Fig. 1C). However, a slight
recovery of neurological functions was observed in the BV group
(Fig. 1C). Therefore, it can be
concluded that BV treatment effectively decreased cerebral
infarction volume and may improve functional recovery.
In addition, the effects of BV administration on
MABP and heart rate (HR) were evaluated in the S, C and BV groups
(Table V). Compared with the S
group, MABP was decreased in the C group, while BV treatment
reversed this decline. Compared with the S group, HR was increased
in the C group, and BV treatment significantly alleviated this
increase.
| Table VMABP and HR in the different
experimental groups. |
Table V
MABP and HR in the different
experimental groups.
| Group | MABP (mmHg)
| HR (bpm)
|
|---|
| Pre-operation | 48 h
post-operation | Pre-operation | 48 h
post-operation |
|---|
| S | 76.6+7.5 | 76.4+4.3 | 329.2+9.1 | 330.2+15.3 |
| C | 75.6+3.9 | 63.2+4.3a | 327.2+13.1 | 351.2+24.1b |
| BV | 75.8+4.5 | 68.8+3.6b, c | 328.8+8.8 | 343.4+10.3b, c |
BV administration alters miRNA expression
in the rat MCAO model
miRNA microarray analysis was used to detect the
expression profiles of miRNAs in brain tissues isolated from the
three groups (n=3 rats per group), in order to examine whether BV
treatment can alter the expression profile of miRNAs following
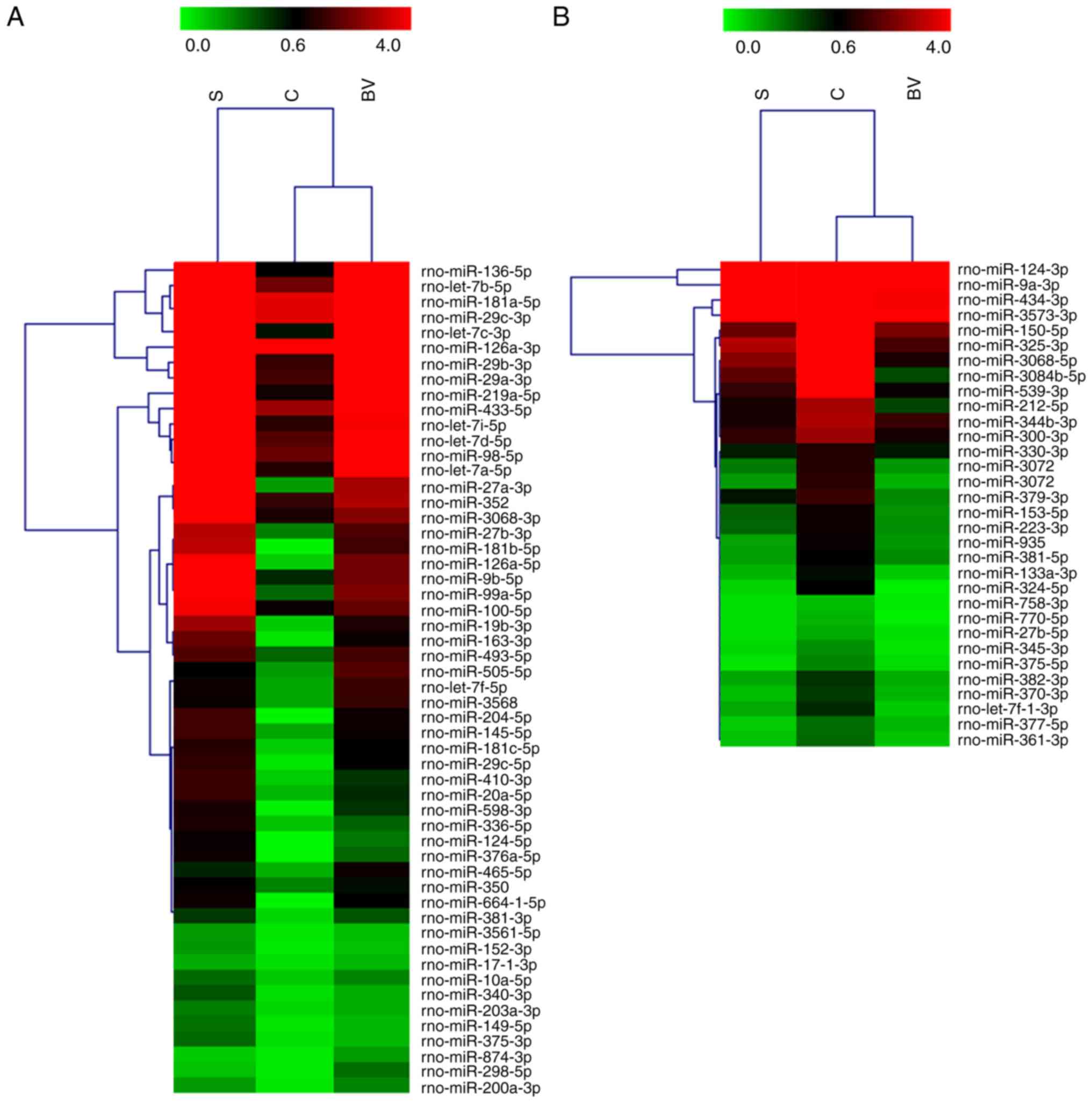
cerebral ischemia. miRNA microarray analysis revealed that 86
miRNAs were differentially expressed (fold change ≥2) in the BV
group compared with the other groups (Fig. 2). The dramatic expression change
in the three groups indicates that the BV administration altered
the miRNA response to stroke.
qPCR verification of miRNA array
results
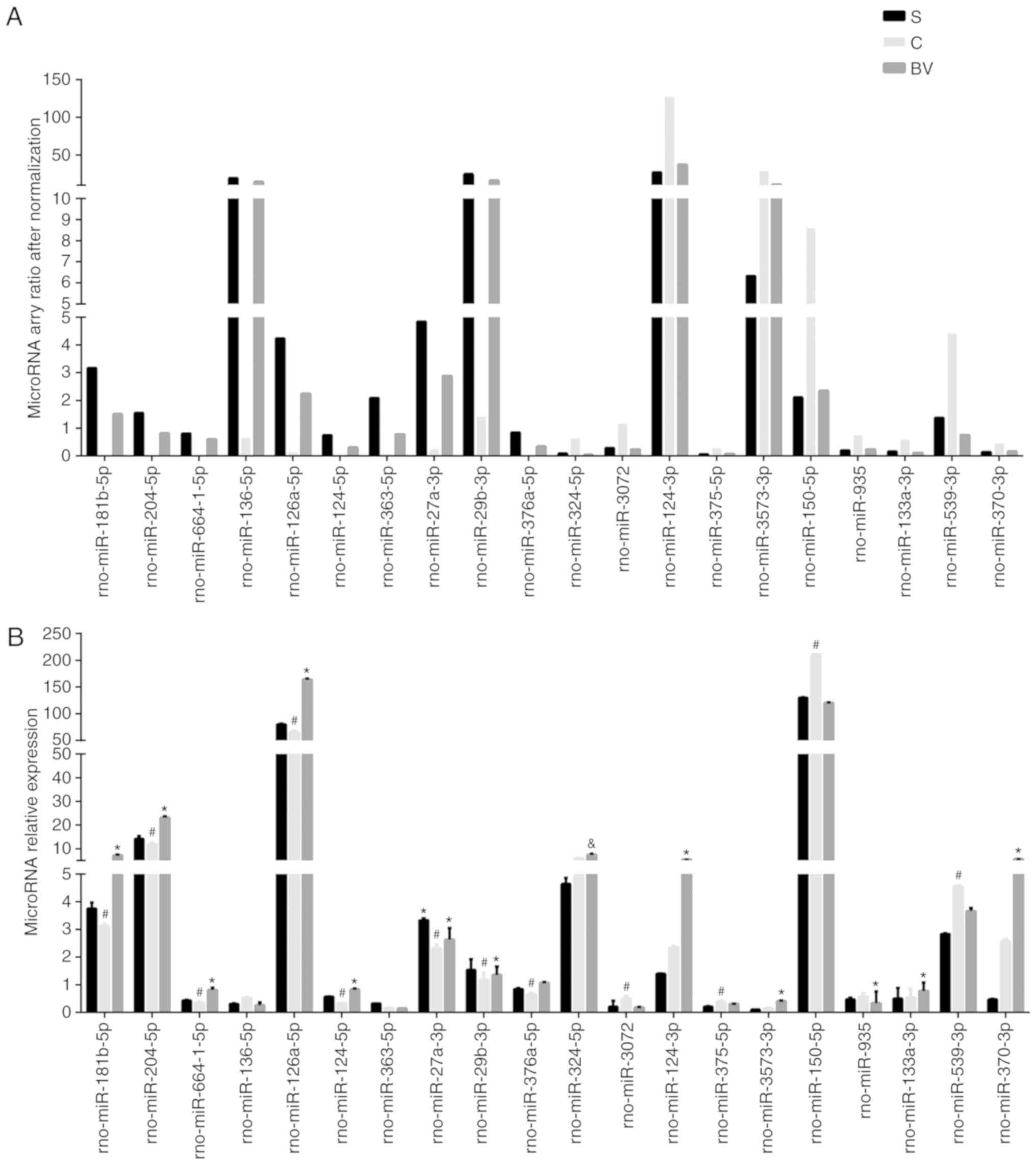
A total of 20 miRNAs were selected from the miRNA
array with the top 10 fold change of 'Up/Down' or 'Down/Up' groups.
These included rno-miR-181b-5p, -204-5p, -664-1-5p, -126a-5p,
-124-5p, -376a-5p, -27a-3p, -29b-3p, -136-5p, -363-5p, -324-5p,
-3072, -124-3p, -375-5p, -3573-3p, -150-5p, -935, -133a-3p,
-539-3p, -370-3p (Fig. 3A).
Compared with the S group, miRNAs were upregulated in the C group,
but downregulated in the BV group, from the group referred as
'Up/Down', which contains 32 miRNAs. By contrast, the other 54
microRNAs changed trends were on the contrary referred to as
'Down/Up'. Except for rno-miR-136-5p, -363-5p, -324-5p, -124-3p,
3573-3p, 133a-3p, -370-3p, all other miRNAs verified by qPCR
exhibited similar alterations as detected by microarray (Fig. 3B).
GO and KEGG analysis of miRNAs
To determine the potential influence of the
differentially expressed miRNAs following BV treatment in MCAO, the
potential target genes of these miRNAs were predicted by Targetscan
v6.2, miRmap and miRDB analyses. Some miRNAs had a large number of
target genes, including rno-miR-181b-5p, -124-5p, -376a-5p,
-664-1-5p, -126a-5p; -27a-3p, -3072, -935, 539-3p, while others had
just one target, such as rno-miR-375-5p.
A total of 1,052 predicted target genes were
identified ('Up/Down' miRNAs group, 225 genes; 'Down/Up' miRNAs
group, 727 genes). Among them, 970 predicted genes had David IDs
and were subjected to GO and KEGG analysis in DAVID v6.7.
('Up/Down' miRNAs group, 222 genes; 'Down/Up' miRNAs group, 718
genes).
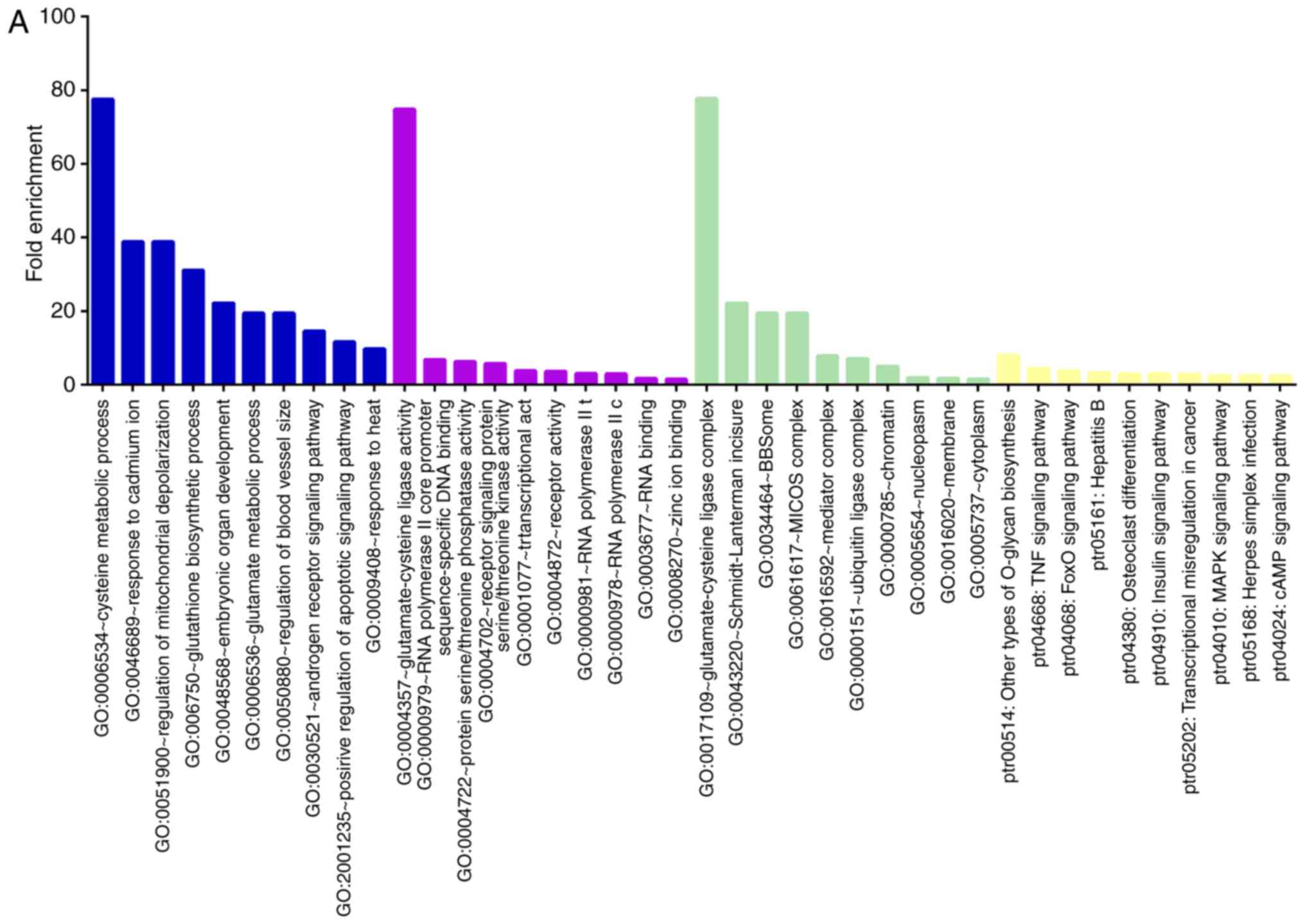
Analyzing the target genes of the 'Up/Down' group
revealed that GO processes associated with metabolism were
significantly overrepresented in these genes, such as regulation of
mitochondrial depolarization, glutamate metabolic process,
glutathione biosynthetic process, and protein serine/threonine
phosphatase activity. In addition, overrepresented pathways
included the TNF, MAPK, cAMP, FoxO and insulin signaling pathways,
as well as herpes simplex infection and transcriptional
misregulation in cancer (Fig.
4A).
Analyzing the target genes of the 'Down/Up' group
revealed overrepresentation of GO processes associated with
ubiquitination, such as ubiquitin conjugating enzyme binding,
thiol-dependent ubiquitin-specific protease activity,
thiol-dependent ubiquitin-specific protease activity, SCF and
Cul3-RING ubiquitin ligase complex, ubiquitin ligase complex, the
pathway including proximal tubule bicarbonate reclamation, inositol
phosphate metabolism, phosphatidylinositol signaling system,
GABAergic synapse, and the NOD-like receptor signaling pathway. The
inflammation-related TNF pathway and the neuronal function-related
biological functions, including positive regulation of apoptotic
signaling pathway, postsynaptic density, synapse, GABAergic synapse
and regulation of blood vessel size, may explain how BV exerts its
neuroprotective effects in MCAO (Fig.
4B).
Integration analysis of miRNA and
mRNA
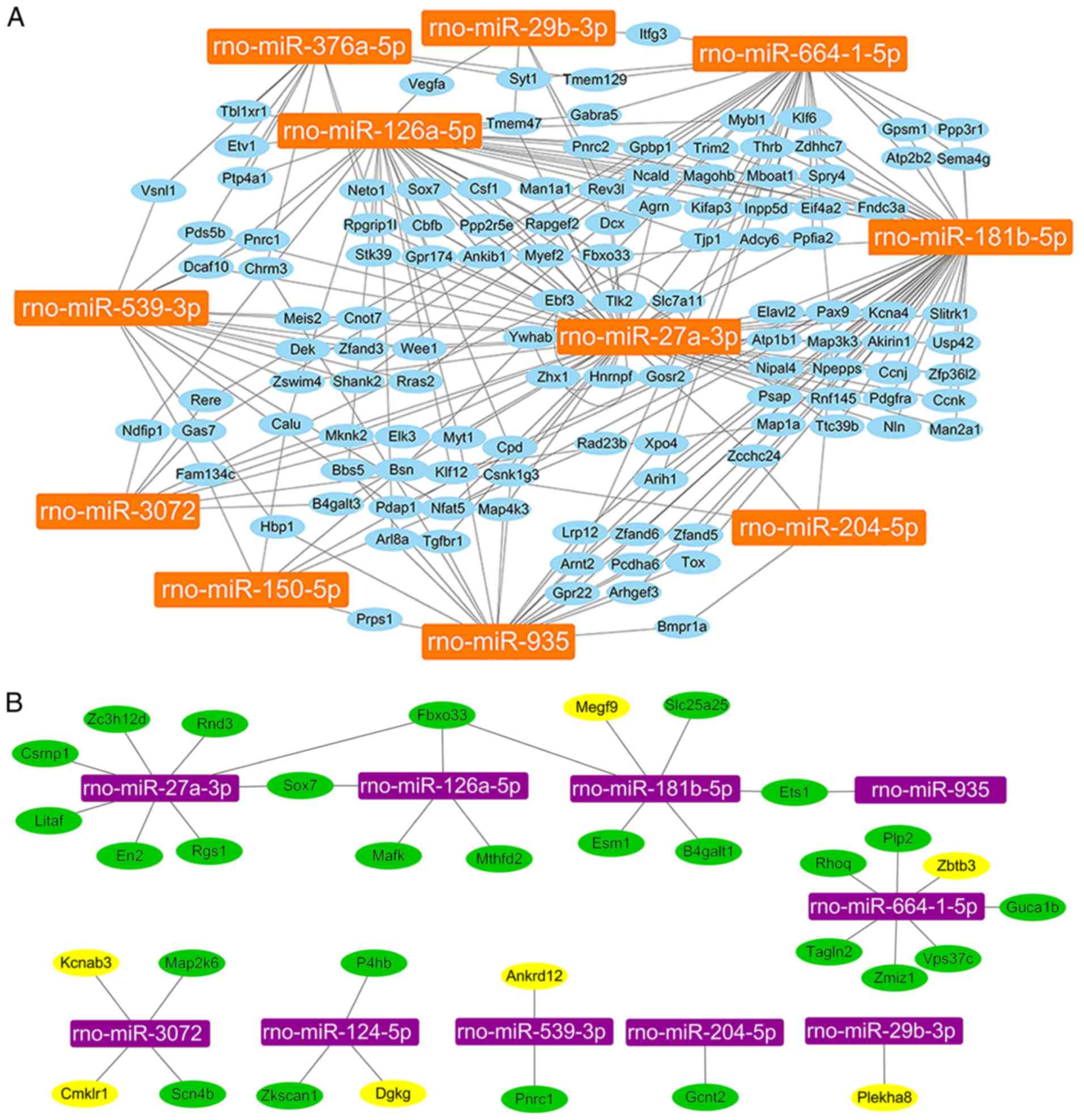
Because each miRNA has multiple potential mRNA
targets, one mRNA can regulated by multiple miRNAs. The 13 miRNAs
that were confirmed by qPCR were further analyzed and their 1,052
predicted target genes were screened. The results demonstrated that
except for rno-miR-124-5p and rno-miR-375-5p, 126 genes had at
least two miRNAs co-regulation (Fig.
5A). In order to screen more accurate target genes of miRNAs,
the results from the miRNA microarray were compared to the results
from the mRNA microarray. The mRNA microarray assay revealed a
total of 1,718 genes with fold change ≥2 (group C vs. group S,
group BV vs. group C). Of the 1,718 genes, 49 genes overlapped with
miRNA target genes (Sox7, Fbxo33, Spry4, Plekha8, Ets1, Cd4, Gcnt2,
Dll4, Nfe2l2, Pparg, Csrnp1, En2, Rgs1, Litaf, Rnd3, Zc3h12d,
Hbegf, Ier3, P4hb, Mthfd2, Mafk, Nr4a3, Tnf, Il1a, Adamts1, Dusp5,
B4galt1, Slc25a25, Cyr61, Esm1, Pgf, Vps37c, Zmiz1, Rhoq, Emp1,
Tagln2, Plp2, Zkscan1, Dgkg, Megf9, Zbtb3, Guca1b, Pnrc1, Ankrd12,
Kcnab3, Map2k6, Scn4b, Tlr5, Cmklr1). Among the 49 genes, 33
candidate genes were selected for qPCR verification, including
Sox7, Fbxo33, Plekha8, Ets1, Gcnt2, Csrnp1, En2, Rgs1, Litaf, Rnd3,
Zc3h12d, P4hb, Mthfd2, Mafk, B4galt1, Slc25a25, Esm1, Vps37c,
Zmiz1, Rhoq, Tagln2, Plp2, Zkscan1, Dgkg, Megf9, Zbtb3, Guca1b,
Pnrc1, Ankrd12, Kcnab3, Map2k6, Scn4b, Cmklr1 (Fig. 5B).
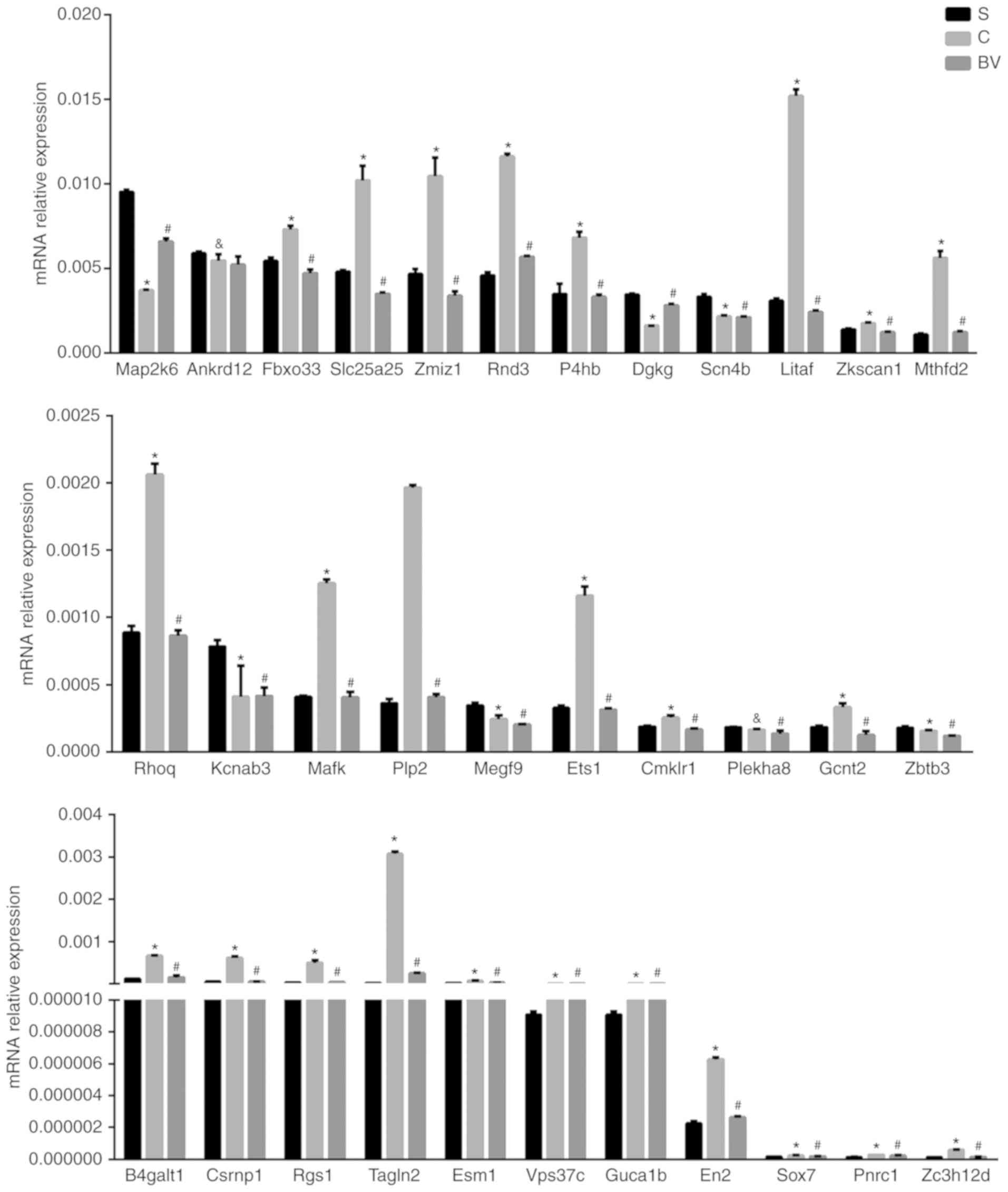
qPCR verification of target mRNAs
qPCR was performed to identify target mRNAs of
miRNAs, which may be related with BV treatment in MCAO. Based on
the criteria that the target mRNA should display an inverse
expression correlation with the miRNA, 33 mRNAs were tested. These
included the genes Sox7, Fbxo33, Plekha8, Ets1, Gcnt2, Csrnp1, En2,
Rgs1, Litaf, Rnd3, Zc3h12d, P4hb, Mthfd2, Mafk, B4galt1, Slc25a25,
Esm1, Vps37c, Zmiz1, Rhoq, Tagln2, Plp2, Zkscan1, Dgkg, Megf9,
Zbtb3, Guca1b, Pnrc1, Ankrd12, Kcnab3, Map2k6, Scn4b, Cmklr1.
Excepting Plekha8, Zbtb3, Megf9, Dgkg, Ankrd12, Kcnab3 and Cmklr1.
The results demonstrated that out of these 33 genes, 26 were
changed in accordance to the 'microRNA-mRNA' regulatory mechanism
identified by the microarray analyses (Figs. 5B and 6).
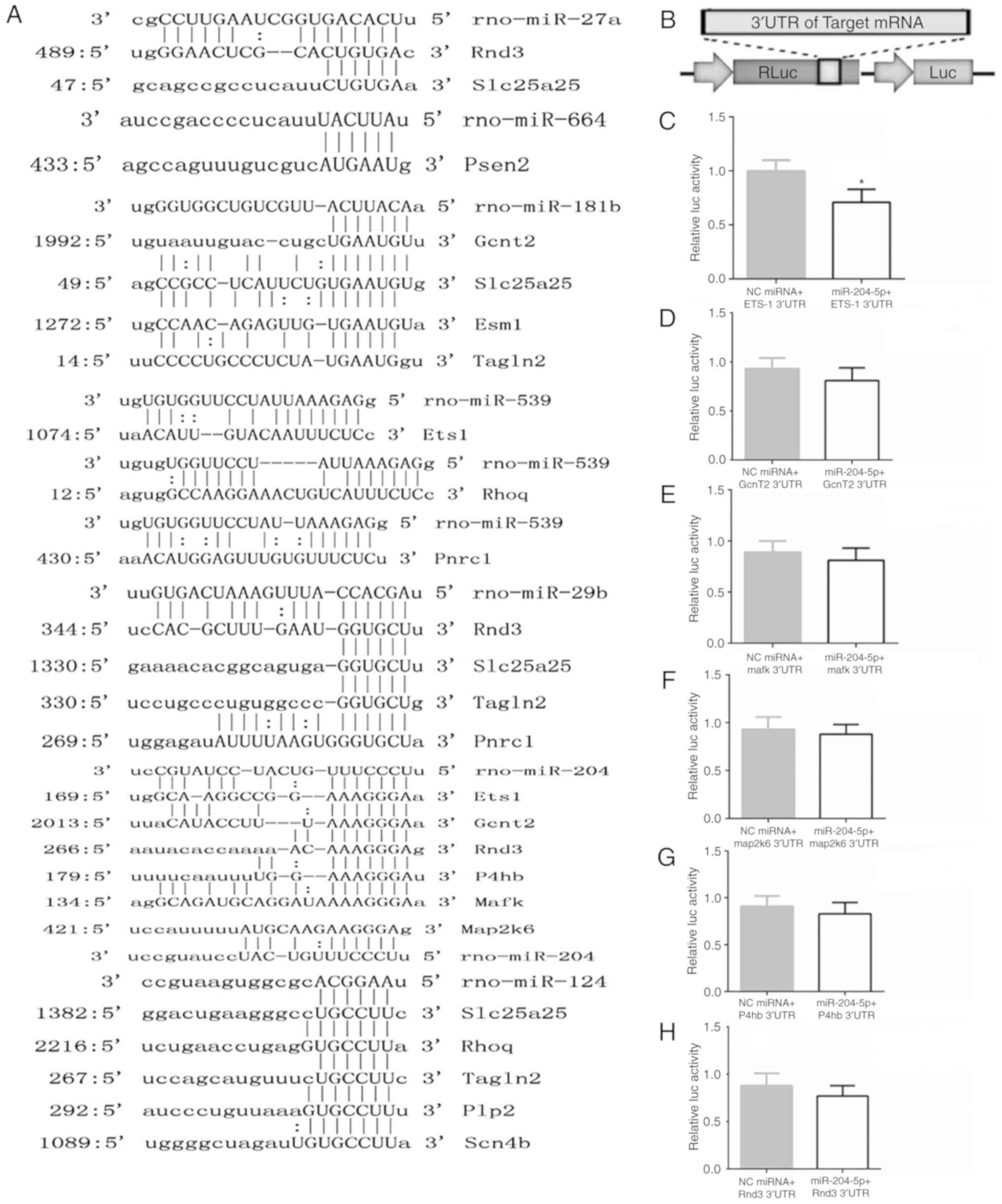
Additionally, using luciferase assay, the regulation
relationship of Ets1 and miR-204-5p was examined. Firstly, the
conserved binding site of the target gene was compared with the
miRNAs, and it was hypothesized that an effect of miR-204-5p on the
Ets1 3'UTR was mediated via a single, highly conserved binding site
(Fig. 7A). Then, the full-length
3'UTR of the transcript was cloned downstream of a luciferase
reporter vector (Fig. 7B). The
reporter vector, in combination with miRNA mimics, was
co-transfected into 293T cells, and luciferase activity was
monitored 48 h later. A robust decrease in luciferase activity was
observed in the miR-204-5p group, while the negative control (NC)
miRNA had no effect on luciferase activity (Fig. 7C).
Discussion
The present study investigated the molecular network
of miRNA and mRNA expression to modulate critical processes in CIR
rats following BV administration. To clarify the influence of miRNA
and mRNA on CIR rats, a model of BV treatment in MCAO rats was
established and the expression change of mRNA and miRNA were
evaluated by microarray analysis. The microarray data were then
verified using qPCR. Furthermore, combined with bioinformatics
prediction, BV treatment was demonstrated to alter the expression
profile of multiple miRNAs and mRNAs in the cortex of rats. Among
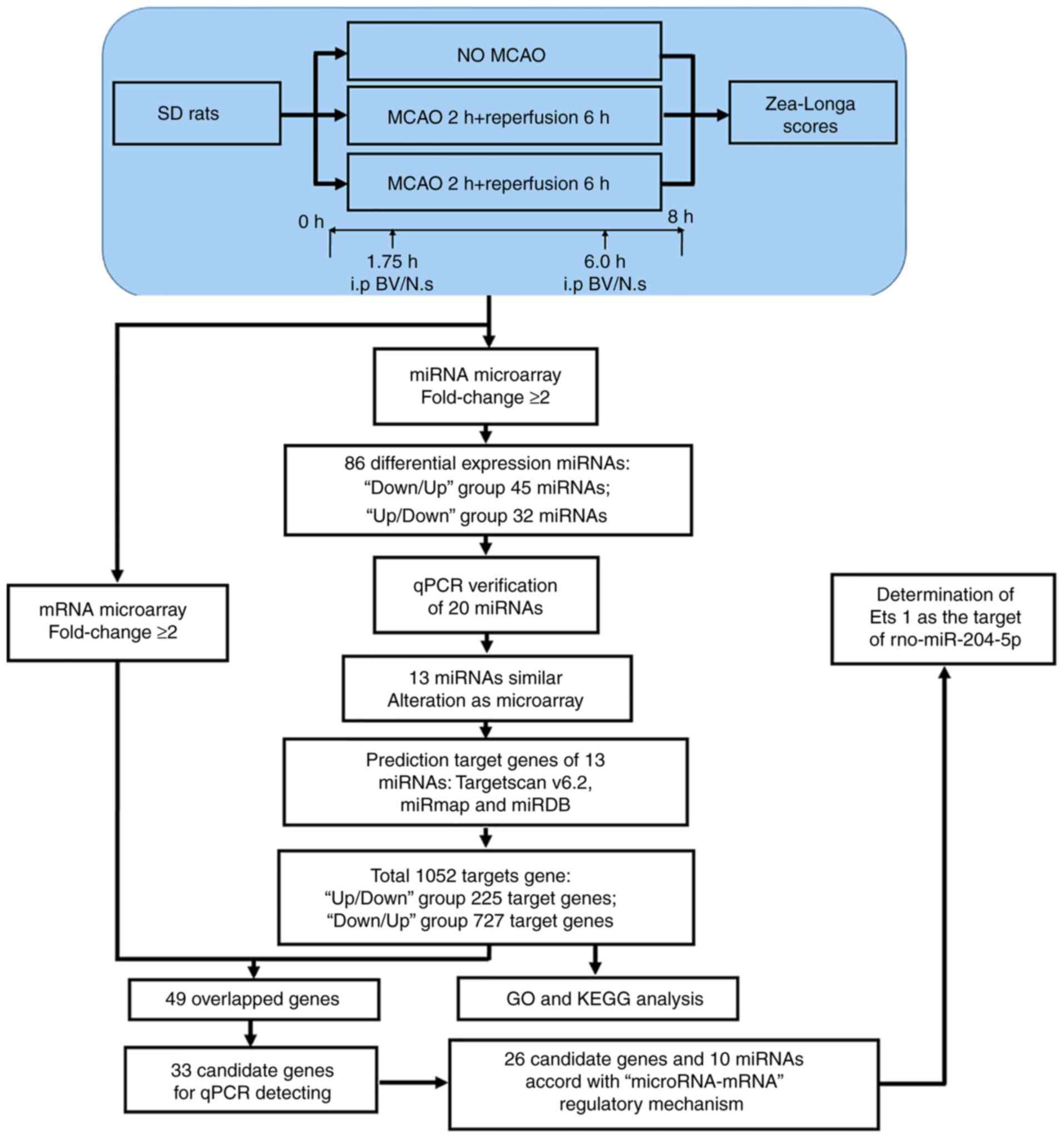
them, there were 10 miRNAs and 26 candidate genes that comprised a
core 'microRNA-mRNA' regulatory mechanism (Fig. 8).
First, the present study successfully established
the MCAOs model, as confirmed by TTC staining. At 48 h
post-reperfusion, BV treatment markedly reduced the infarct size
and slightly decreased the NSS score, compared with the vehicle
control group. Generally, the changes of morphology and function
were followed by changes in gene expression. A previous study has
indicated that administration of BV resulted in rapid appearance of
bilirubin in the serum and significantly suppressed IRI-induced
liver dysfunction in swine (20).
Exogenously administered CO and BV have been demonstrated to
exhibit potent cytoprotective effects on intestinal IRI (9). Recent evidence has demonstrated that
BV has a protective role against lung graft injury, hemorrhagic
shock and resuscitation-induced lung injury through
anti-inflammatory and antioxidant mechanisms (8,21).
The present findings suggested that the molecular network of
miRNA-mRNA expression may regulate critical processes to attenuate
the release of inflammatory factors in CIR rats following BV
administration.
Next, through GO analysis, the function of 26
candidate genes was divided into six categories. The first category
was related to cell proliferation, and included the genes En2,
Rnd3, Sox7, Mthfd2, β4GalT, MaP2K6, Gcnt2, and Zmiz1. Engrailed 2
(En2) is expressed in all cerebellar cell types, and is critical
for regulating the formation of specific fissures and general
growth of the cerebellum (22).
Downregulation of Rnd3 promotes glioma cell proliferation (23,24). Overexpression of sex determining
region Y-box 7 (Sox7) antagonizes cell growth and promotes
apoptosis in a number of cancer cell lines (25-27). Furthermore, the overexpression of
Mthfd2 promotes tumor cell proliferation (28,29). β4GalT functions in several types
of cancer, such as renal cancer, leukemia and neural crest-derived
tumors, and is correlated with cancer cell proliferation,
metastasis, invasiveness and drug resistance (30-32), Map2k6 can effectively inhibit cell
proliferation and induce apoptosis in ovarian cancer HO8910 cells
(33). In addition,
downregulation of Gcnt2 can promote DNA hyper-methylation (34), and enhance cell migration and
invasion in breast cancer and prostate cancer (35,36). Lastly, ectopic expression of zinc
finger MIZ-type containing 1 (Zmiz1) induces cutaneous squamous
cell malignancies in a mouse model of cancer (37). In the present study, the
expression levels of En2, Rnd3, Sox7, Mthfd2, β4GalT, Gcnt2 and
Zmiz1 were downregulated, while MaP2K6 was upregulated following BV
administration, which suggested that these genes were involved in
the process of CIR following BV treatment.
The second category was related to the development
and function of the nervous system, and included the genes Csrnp1
and En2. Cystein-serine-rich nuclear protein 1 (Csrnp1) is
essential for cephalic neural progenitor proliferation and survival
in zebrafish (38). EN2 has key
roles in developing mesencephalic dopa-minergic (mDA) neurons and
also impacts on the adult mDA neuronal biological function
(39). In the present study, the
expression change of Csrnp1 and En2 may regulate the functional
remodeling in our experiment condition.
The third category related to the anti-inflammatory
response, and included the genes Litaf, Zc3h12d and Rgs1.
LPS-induced TNF factor is known to activate the transcription of
multiple cytokines, such as TNF-α, IL-8 and IL-β, in a variety of
cellular processes and many inflammatory diseases (21,40-42). Zc3h12a and Zc3h12d recognize a set
of common target mRNAs encoding proteins that serve important roles
in the course of the inflammation (43). By reducing the stability of mRNAs
encoding proinflammatory factors, Zc3h12d attenuates inflammatory
responses (44,45). Regulator of G-protein Signaling
(Rgs1) can inhibit chemokine receptor signaling as potential
therapeutic targets in leukocyte trafficking and vascular
inflammation (46). Endothelial
cell-specific molecule-1 (Esm1) is secreted by endothelial cells
and upregulated by inflammatory cytokines, such as TNF (47). In the present study, the increase
of Litaf, Zc3h12d and Rgs1 may be associated with the
anti-inflammation activity in CIR brains following BV
treatment.
The fourth category was related to angiogenesis, and
included the genes Rnd3, Sox7, Ets1, Esm1 and Esm1. Rnd3 acts as a
novel proangiogenic factor involved in cardiac responsive
angiogenesis through HIF1α-VEGFA signaling promotion (24). Sox7 regulates angiogenesis and
vasculogenesis through mechanisms that are redundant with those of
Sox18 (48,49). TF-bearing mEMPs increase
angiogenesis operating via paracrine regulation of neighboring
endothelial cells, signaling through the β1-integrin pathway
Rac1-ERK1/2-Ets1 and triggering CCL2 production to form new and
competent mature neovessels (50). Esm1 promotes the angiogenesis and
tumor cell growth of prolactinomas (51). Together, the expression of those
molecules may regulate the angiogenesis process that is beneficial
to the vessel development in ischemia brains following BV
administration.
The fifth category was related to cell apoptosis,
and included the genes Mafk, Ets1 and Plp2. Protein K (avian)
(Mafk) enhances oxidative stress-induced apoptosis (52). Knockdown of erythroblastosis virus
E26 oncogene homolog 1 (Ets1) promotes apoptosis in myocardial
ischemic reperfusion injury (53). Downregulation of Plp2 increases
endoplasmic reticulum-stress-induced neuronal apoptosis and risk
for adverse neurological outcomes after hypoxia ischemia injury
(54). Inhibiting Tagln2
expression could mediate the hypoxia-induced apoptosis in cardiac
myocytes (55). Therefore, these
gene expressions may influence function by regulating cell
apoptosis in the present model.
Finally, the sixth category includes genes such as
Rhoq, slc25a25, vps37c, P4hb, Scn4b and Zkscan1. Overexpression of
member Q (Rhoq) may increase invasion potential in colorectal
cancer (56). Solute carrier
family 25 (slc25a25) is localized in the mitochondrial inner
membrane and maintains ATP homeostasis and endurance performance
(57). Derepression of the
slc25a25 can boost mitochondrial respiration and promote the
product of ATP (58). Hyperoxia
reduces the survival benefit of U87 and U251 cells with prolyl
4-hydroxylase β polypeptide (P4hb) over-expression through the
unfolded protein response (UPR) (59). When cultured mouse cerebral
cortical neurons were exposed to hypoxia for 24 h, P4hb was
upregulated, and it may participate protein-ubiquitination pathways
in the neuronal response to hypoxia (59). Vacuolar protein sorting 37 homolog
C (vps37c) is a component of endosomal sorting complex required for
transport-I important for viral budding (60). Guca1b is upregulated in dog heart
failure (61). Sodium channel
voltage-gated type IV β (Scn4b) function may relate with
Na+ channels (62). In
adenocarcinomas of the gastroesophageal junction, Zkscan1 mRNA is
overexpressed (63). The present
study is the first to report that the aforementioned genes are
involved in the neuroprotection activity of BV in treated CIR
rats.
Furthermore, the present study screened a series of
miRNAs and several crucial findings were observed in CIR rats
following BV administration. By analysis of bionetwork, these
miRNAS were predicted to function with their target mRNAs. Compared
with the S group, miR-27a, miR-126a-3p, miR-181b-5p, miR-664,
miR-124-5p and miR-204-5p were downregulated in the C group and
upregulated in the BV group, while miR-935 was upregulated in the C
group and downregulated in the BV group. A previous study indicated
that miR-27 might unravel unknown pathways in virus-associated CNS
dysfunction (64). The present
results revealed that BV administration on CIR rats induced
downregulation of En2, Rnd3 and Sox7 (the target genes of miR-27a)
which may promote cell proliferation; downregulation of Litaf,
Zc3h12d, Rgs1 (the target genes of miR-27a) may promote
anti-inflammatory response; downregulation of Rnd3 and Sox7 (the
target genes of miR-27a) may inhibit angiogenesis; and
downregulation of Csrnp1and En2 (the target genes of miR-27a) are
poorly expressed to CNS. In sepsis, the downregulation of
miR-126a-3p in endothelial cells resulted in increased apoptosis,
and decreased proliferation and migration (65). The present results suggested that
BV administration in CIR rats induced downregulation of Mafk (the
target gene of miR-126a-3p) and may promote cell apoptosis; while
downregulation of Mthfd2 (the target gene of miR-126a-3p) may
inhibit cell proliferation; and downregulation of Sox7 (the target
gene of miR-126a-3p) may promote cell proliferation. miR-181b-5p
participates in eosinophilic airway inflammation by regulating
IL-13, IL-1β and CCL11 expression via targeting SPP1 (66). The present results indicated that
BV administration in CIR rats induced the change of miR-181b-5p,
supporting its role in the current model. miR-664 has been known to
negatively regulate proteolipid protein 2 (Plp2), promote cell
proliferation and invasion in T-cell acute lymphoblastic leukaemia
and melanoma (67,68). Combining knowledge from the PubMed
database with the current results suggests that BV administration
in CIR rats induced downregulation of Zmiz1 (the target gene of
miR-664) to inhibit cell proliferation. miR-124-5p functions are as
a tumor suppressor and serve as a molecular marker for glioma
diagnosis (69). In addition, BV
administration in CIR rats induced downregulation of P4hb (the
target gene of miR-124-5p), which may participate in
protein-ubiquitination pathways in the neuronal response to
hypoxia; moreover, downregulation of Zkscan1 (the target gene of
miR-124-5p) mRNA may relate to the gastroesophageal junction.
Overexpression of miR-935 promotes cell proliferation of gastric
cancer and lung cancer (70,71), where the target gene Ets1 is not
consistant with the miRNA-mRNA regulatory mechanism following qPCR
testing.
In addition, for miR-204-5p, several studies have
demonstrated that miR-204 is significantly increased at 24 h
post-oxygen-glucose deprivation (OGD) by using RT-qPCR to determine
the expression of 16 miRNAs of interest at 1 and 24-h post-OGD
(72). Overexpression of
miR-204-5p inhibits cell migration and proliferation in glioma
cancer (73). Through the
luciferase reporter assay, the present results confirmed the
regulation of miR-204-5p on the target gene Ets1, and Ets1 was
downregulated in CIR rats following BV administration. It has been
reported that Ets1 mRNA is increased following MCAO-induced stroke
(74). Ets1 was required for
activation of RAS-regulated cell migration genes, but also
identified a surprising role for Ets1 in the repression of genes
such as DUSP4, DUSP6 and SPRY4 that provide negative feedback to
the RAS/ERK pathway (75). The
present results suggested that miR-204-5p directly regulates Ets-1,
which may indicate a potential mechanism by which BV improves
neural behavior in CIR rats.
The present study provides a global view of the
influence of miRNA/mRNA expression on the genomic response in CIR
rats following BV treatment. miR-27a, miR-181b-5p, Litaf, Zc3h12d
and Rgs1 have been demonstrated to participate in inflammatory
response. BV treatment upregulated miR-27a, miR-181b-5p and miR-204
5p expression and induced their target genes downregulation to
influence the anti-inflammatory effect of BV in CIR. Therefore, BV
may influence crucial biological functions, such as cell
proliferation, apoptosis, maintaining ATP homeostasis and
angiogenesis, by miRNAs regulating target genes, such as miRNA
204-5p directly targeting Ets1.
Funding
This research was supported by the Natural Science
Foundation of China (grant no. 81760248), the Foundation of Yunnan
Provincial Science and Technology Department (grant no.
2017FE468-034), the Foundation of Yunnan Health Department (grant
no. 2016NS043), Weifeng Yu Expert Workstation (grant no. 2017IC067)
and Key Applied and Basic Research Program in Yunnan Province
(grant no. 2018FA042).
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
ZYZ and JLS conceived and designed the project. ZYZ,
CC and JJL analyzed and interpreted the data. YJ, JL and ZM
provided technical support. ZYZ, THW and JLS wrote and revised the
manuscript. THW and JLS supervised the study. All authors have read
and approved the final manuscript.
Ethics approval and consent to
participate
All experimental protocols involving animals were
approved by the Animal Care and Welfare Committee of Kunming
Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
CIR
|
cerebral ischemia reperfusion
injury
|
|
BV
|
biliverdin
|
|
CO
|
carbonic oxide
|
|
miRNA
|
microRNA
|
|
MCAO
|
middle cerebral artery occlusion
|
|
SPF
|
specific pathogen free
|
|
ip
|
intraperitoneally
|
|
RCCA
|
right common carotid artery
|
|
CBF
|
cerebral blood flow
|
|
TTC
|
2,3,5-triphenyltetrazolium
chloride
|
Acknowledgments
Not applicable.
References
|
1
|
Sarady-Andrews JK, Liu F, Gallo D, Nakao
A, Overhaus M, Ollinger R, Choi AM and Otterbein LE: Biliverdin
administration protects against endotoxin-induced acute lung injury
in rats. Am J Physiol Lung Cell Mol Physiol. 289:L1131–L1137. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhou XQ, Zeng XN, Kong H and Sun XL:
Neuroprotective effects of berberine on stroke models in vitro and
in vivo. Neurosci Lett. 447:31–36. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen GY and Nuñez G: Sterile inflammation:
Sensing and reacting to damage. Nat Rev Immunol. 10:826–837. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Deng H, Zuo X, Zhang J, Liu X, Liu L, Xu
Q, Wu Z and Ji A: Alphalipoic acid protects against cerebral
ischemia/reperfusion-induced injury in rats. Mol Med Rep.
11:3659–3665. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yamasaki Y, Matsuura N, Shozuhara H,
Onodera H, Itoyama Y and Kogure K: Interleukin-1 as a pathogenetic
mediator of ischemic brain damage in rats. Stroke. 26:676–681.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lavine SD, Hofman FM and Zlokovic BV:
Circulating antibody against tumor necrosis factor-alpha protects
rat brain from reperfusion injury. J Cereb Blood Flow Metab.
18:52–58. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fondevila C, Shen XD, Tsuchiyashi S,
Yamashita K, Csizmadia E, Lassman C, Busuttil RW, Kupiec-Weglinski
JW and Bach FH: Biliverdin therapy protects rat livers from
ischemia and reperfusion injury. Hepatology. 40:1333–1341. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kosaka J, Morimatsu H, Takahashi T,
Shimizu H, Kawanishi S, Omori E, Endo Y, Tamaki N, Morita M and
Morita K: Effects of biliverdin administration on acute lung injury
induced by hemorrhagic shock and resuscitation in rats. PLoS One.
8:e636062013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nakao A, Kaczorowski DJ, Sugimoto R,
Billiar TR and McCurry KR: Application of heme oxygenase-1, carbon
monoxide and biliverdin for the prevention of intestinal
ischemia/reperfusion injury. J Clin Biochem Nutr. 42:78–88. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wegiel B, Gallo D, Csizmadia E, Roger T,
Kaczmarek E, Harris C, Zuckerbraun BS and Otterbein LE: Biliverdin
inhibits Toll-like receptor-4 (TLR4) expression through nitric
oxide-dependent nuclear translocation of biliverdin reductase. Proc
Natl Acad Sci USA. 108:18849–18854. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Holst B, Raby AC, Hall JE and Labéta MO:
Complement takes its Toll: An inflammatory crosstalk between
Toll-like receptors and the receptors for the complement
anaphylatoxin C5a. Anaesthesia. 67:60–64. 2012. View Article : Google Scholar
|
|
12
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yu J, Liu F, Yin P, Zhu X, Cheng G, Wang
N, Lu A, Luan W, Zhang N, Li J, et al: 2.2 Integrating miRNA and
mRNA expression profiles in response to heat stress-induced injury
in rat small intestine. Funct Integr Genomics. 11:203–213. 2011.
View Article : Google Scholar
|
|
14
|
Nassar FJ, El Eit R and Nasr R: An
integrative analysis of microRNA and mRNA profiling in CML stem
cells. Methods Mol Biol. 1465:219–241. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen JH, He HC, Jiang FN, Militar J, Ran
PY, Qin GQ, Cai C, Chen XB, Zhao J, Mo ZY, et al: Analysis of the
specific pathways and networks of prostate cancer for gene
expression profiles in the Chinese population. Med Oncol.
29:1972–1984. 2012. View Article : Google Scholar
|
|
16
|
Chiang T, Messing RO and Chou WH: Mouse
model of middle cerebral artery occlusion. J Vis Exp.
2761:2011.
|
|
17
|
Li JJ, Zou ZY, Liu J, Xiong LL, Jiang HY,
Wang TH and Shao JL: Biliverdin administration ameliorates cerebral
ischemia reperfusion injury in rats and is associated with
proinflammatory factor downregulation. Exp Ther Med. 14:671–679.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kam KY, Yu SJ, Jeong N, Hong JH, Jalin AM,
Lee S, Choi YW, Lee CK and Kang SG: p-Hydroxybenzyl alcohol
prevents brain injury and behavioral impairment by activating Nrf2,
PDI, and neurotrophic factor genes in a rat model of brain
ischemia. Mol Cells. 31:209–215. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
20
|
Andria B, Bracco A, Attanasio C, Castaldo
S, Cerrito MG, Cozzolino S, Di Napoli D, Giovannoni R, Mancini A,
Musumeci A, et al: Biliverdin protects against liver ischemia
reperfusion injury in swine. PLoS One. 8:e699722013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou J, Yang Z, Tsuji T, Gong J, Xie J,
Chen C, Li W, Amar S and Luo Z: LITAF and TNFSF15, two downstream
targets of AMPK, exert inhibitory effects on tumor growth.
Oncogene. 30:1892–1900. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Orvis GD, Hartzell AL, Smith JB, Barraza
LH, Wilson SL, Szulc KU, Turnbull DH and Joyner AL: The engrailed
homeobox genes are required in multiple cell lineages to coordinate
sequential formation of fissures and growth of the cerebellum. Dev
Biol. 367:25–39. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu BH, Lin X, Yang X, Dong H, Yue X,
Andrade KC, Guo Z, Yang J, Wu L, Zhu X, et al: Downregulation of
RND3/RhoE in glioblastoma patients promotes tumorigenesis through
augmentation of notch transcriptional complex activity. Cancer Med.
4:1404–1416. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yue XJ, Lin X, Yang T, Yang X, Yi X, Jiang
X, Li X, Li T, Guo J, Dai Y, et al: Rnd3/RhoE modulates
hypoxia-inducible factor 1α/vascular endothelial growth factor
signaling by stabilizing hypoxia-inducible factor 1α and regulates
responsive cardiac angiogenesis. Hypertension. 67:597–605. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guo L, Zhong D, Lau S, Liu X, Dong XY, Sun
X, Yang VW, Vertino PM, Moreno CS, Varma V, et al: Sox7 is an
independent checkpoint for beta-catenin function in prostate and
colon epithelial cells. Mol Cancer Res. 6:1421–1430. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang Y, Huang SY, Dong W, Li L, Feng Y,
Pan L, Han Z, Wang X, Ren G, Su D, et al: SOX7, down-regulated in
colorectal cancer, induces apoptosis and inhibits proliferation of
colorectal cancer cells. Cancer Lett. 277:29–37. 2009. View Article : Google Scholar
|
|
27
|
Wang C, Qin L, Min Z, Zhao Y, Zhu L, Zhu J
and Yu S: SOX7 interferes with β-catenin activity to promote
neuronal apoptosis. Eur J Neurosci. 41:1430–1437. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gustafsson Sheppard N, Jarl L, Mahadessian
D, Strittmatter L, Schmidt A, Madhusudan N, Tegnér J, Lundberg EK,
Asplund A, Jain M and Nilsson R: The folate-coupled enzyme MTHFD2
is a nuclear protein and promotes cell proliferation. Sci Rep.
5:150292015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tedeschi PM, Vazquez A, Kerrigan JE and
Bertino JR: Mitochondrial methylenetetrahydrofolate dehydrogenase
(MTHFD2) overexpression is associated with tumor cell proliferation
and is a novel target for drug development. Mol Cancer Res.
13:1361–1366. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
la Torre A, Muscarella LA, Parrella P,
Balsamo T, Bisceglia M, Valori VM, la Torre A, Barbano R, Perrella
E, Poeta ML, et al: Aberrant genes promoter methylation in neural
crest-derived tumors. Int J Biol Marker. 27:E389–E394. 2012.
View Article : Google Scholar
|
|
31
|
Zhou H, Zhang Z, Liu C, Jin C, Zhang J,
Miao X and Jia L: B4GALT1 gene knockdown inhibits the hedgehog
pathway and reverses multidrug resistance in the human leukemia
K562/adriamycin-resistant cell line. IUBMB Life. 64:889–900. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xie H, Zhu Y, An H, Wang H, Zhu Y, Fu H,
Wang Z, Fu Q, Xu J and Ye D: Increased B4GALT1 expression
associates with adverse outcome in patients with non-metastatic
clear cell renal cell carcinoma. Oncotarget. 7:32723–32730.
2016.PubMed/NCBI
|
|
33
|
Yuan J, Kang JL, Liao H, Wang XX, Nie ML,
Shuai R and Deng C: Mitogen-activated protein kinase kinase
6-fusion protein (MAP2K6-FP) potentiates the anti-tumor effects of
paclitaxel in ovarian cancer. Anticancer Agent Med Chem.
15:1308–1316. 2015. View Article : Google Scholar
|
|
34
|
Nakamura K, Yamashita K, Sawaki H, Waraya
M, Katoh H, Nakayama N, Kawamata H, Nishimiya H, Ema A, Narimatsu H
and Watanabe M: Aberrant methylation of GCNT2 Is tightly related to
lymph node metastasis of primary CRC. Anticancer Res. 35:1411–1421.
2015.PubMed/NCBI
|
|
35
|
Zhang H, Meng F, Wu S, Kreike B, Sethi S,
Chen W, Miller FR and Wu G: Engagement of I-branching {beta}-1,
6-N-acetylglucosaminyltransferase 2 in breast cancer metastasis and
TGF-{beta} signaling. Cancer Res. 71:4846–4856. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mikami J, Tobisawa Y, Yoneyama T,
Hatakeyama S, Mori K, Hashimoto Y, Koie T, Ohyama C and Fukuda M:
I-branching N-acetylglucosaminyltransferase regulates prostate
cancer invasiveness by enhancing α5β1 integrin signaling. Cancer
Sci. 107:359–368. 2016. View Article : Google Scholar :
|
|
37
|
Rogers LM, Riordan JD, Swick BL, Meyerholz
DK and Dupuy AJ: Ectopic expression of Zmiz1 induces cutaneous
squamous cell malignancies in a mouse model of cancer. J Invest
Dermatol. 133:1863–1869. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Feijóo CG, Sarrazin AF, Allende ML and
Glavic A: Cystein-serine-rich nuclear protein 1, Axud1/Csrnp1, is
essential for cephalic neural progenitor proliferation and survival
in zebrafish. Dev Dyn. 238:2034–2043. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rekaik H, Blaudin de Thé FX, Prochiantz A,
Fuchs J and Joshi RL: Dissecting the role of Engrailed in adult
dopaminergic neurons-insights into Parkinson disease pathogenesis.
FEBS Lett. 589:3786–3794. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shiomi N, Myokai F, Naruishi K, Oyaizu K,
Senoo K, Yamaguchi T, Amar S and Takashiba S: Cloning and
characterization of lipopolysaccharide-induced tumor necrosis
factor alpha factor promoter. Fems Immunol Med Mic. 47:360–368.
2006. View Article : Google Scholar
|
|
41
|
Bushell KN, Leeman SE, Gillespie E, Gower
AC, Reed KL, Stucchi AF, Becker JM and Amar S: LITAF mediation of
increased TNF-α secretion from inflamed colonic lamina propria
macrophages. PLoS One. 6:e258492011. View Article : Google Scholar
|
|
42
|
Tang XR and Amar S: Kavain involvement in
LPS-induced signaling pathways. J Cell Biochem. 117:2272–2280.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wawro M, Kochan J, Krzanik S, Jura J and
Kasza A: Intact NYN/PIN-like domain is crucial for the degradation
of inflammation-related transcripts by ZC3H12D. J Cell Biochem.
118:487–498. 2017. View Article : Google Scholar
|
|
44
|
Minagawa K, Wakahashi K, Kawano H,
Nishikawa S, Fukui C, Kawano Y, Asada N, Sato M, Sada A, Katayama Y
and Matsui T: Posttranscriptional modulation of cytokine production
in T cells for the regulation of excessive inflammation by TFL. J
Immunol. 192:1512–1524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang H, Wang WC, Chen JK, Zhou L, Wang M,
Wang ZD, Yang B, Xia YM, Lei S, Fu EQ and Jiang T: ZC3H12D
attenuated inflammation responses by reducing mRNA stability of
proinflammatory genes. Mol Immunol. 67:206–212. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Patel J, McNeill E, Douglas G, Hale AB, de
Bono J, Lee R, Iqbal AJ, Regan-Komito D, Stylianou E, Greaves DR
and Channon KM: RGS1 regulates myeloid cell accumulation in
atherosclerosis and aortic aneurysm rupture through altered
chemokine signalling. Nat Commun. 6:66142015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Grigoriu BD, Depontieu F, Scherpereel A,
Gourcerol D, Devos P, Ouatas T, Lafitte JJ, Copin MC, Tonnel AB and
Lassalle P: Endocan expression and relationship with survival in
human non-small cell lung cancer. Clin Cancer Res. 12:4575–4582.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang C, Basta T and Klymkowsky MW: SOX7
and SOX18 are essential for cardiogenesis in Xenopus. Dev Dyn.
234:878–891. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Herpers R, van de Kamp E, Duckers HJ and
Schulte-Merker S: Redundant roles for sox7 and sox18 in
arteriovenous specification in zebrafish. Circ Res. 102:12–15.
2008. View Article : Google Scholar
|
|
50
|
Arderiu G, Peña E and Badimon L:
Angiogenic microvascular endothelial cells release microparticles
rich in tissue factor that promotes postischemic collateral vessel
formation. Arterioscl Throm Vasc Biol. 35:348–357. 2015. View Article : Google Scholar
|
|
51
|
Cai L, Leng ZG, Guo YH, Lin SJ, Wu ZR, Su
ZP, Lu JL, Wei LF, Zhuge QC, Jin K and Wu ZB: Dopamine agonist
resistance-related endocan promotes angiogenesis and cells
viability of prolactinomas. Endocrine. 52:641–651. 2016. View Article : Google Scholar
|
|
52
|
Bensellam M, Montgomery MK, Luzuriaga J,
Chan JY and Laybutt DR: Inhibitor of differentiation proteins
protect against oxidative stress by regulating the
antioxidant-mitochondrial response in mouse beta cells.
Diabetologia. 58:758–770. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Bian C, Xu TD, Zhu H, Pan D, Liu Y, Luo Y,
Wu P and Li D: Luteolin inhibits ischemia/reperfusion-induced
myocardial injury in rats via downregulation of microRNA-208b-3p.
PLoS One. 10:e01448772015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhang LL, Wang T and Valle D: Reduced PLP2
expression increases ER-stress-induced neuronal apoptosis and risk
for adverse neurological outcomes after hypoxia ischemia injury.
Hum Mol Genet. 24:7221–7226. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Li AY, Yang Q and Yang K: miR-133a
mediates the hypoxia-induced apoptosis by inhibiting TAGLN2
expression in cardiac myocytes. Mol Cell Biochem. 400:173–181.
2015. View Article : Google Scholar
|
|
56
|
Han SW, Kim HP, Shin JY, Jeong EG, Lee WC,
Kim KY, Park SY, Lee DW, Won JK, Jeong SY, et al: RNA editing in
RHOQ promotes invasion potential in colorectal cancer. J Exp Med.
211:613–621. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Sako H, Yada K and Suzuki K: Genome-wide
analysis of acute endurance exercise-induced translational
regulation in mouse skeletal muscle. PLoS One. 11:e01483112016.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Karunakaran D, Thrush B, Nguyen MA,
Richards L, Geoffrion M, Singaravelu R, Ramphos E, Shangari P,
Ouimet M, Pezacki JP, et al: Macrophage mitochondrial energy status
regulates cholesterol efflux and is enhanced by anti-miR33 in
atherosclerosis. Circ Res. 117:266–278. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lee D, Sun S, Ho AS, Kiang KM, Zhang XQ,
Xu FF and Leung GK: Hyperoxia resensitizes chemoresistant
glioblastoma cells to temozolomide through unfolded protein
response. Anticancer Res. 34:2957–2966. 2014.PubMed/NCBI
|
|
60
|
Eastman SW, Martin-Serrano J, Chung W,
Zang T and Bieniasz PD: Identification of human VPS37C, a component
of endosomal sorting complex required for transport-I important for
viral budding. J Biol Chem. 280:628–636. 2005. View Article : Google Scholar
|
|
61
|
Lanfear DE, Yang JJ, Mishra S and Sabbah
HN: Genome-wide approach to identify novel candidate genes for beta
blocker response in heart failure using an experimental model.
Discov Med. 11:359–366. 2011.PubMed/NCBI
|
|
62
|
Lewis AH and Raman IM: Resurgent current
of voltage-gated Na(+) channels. J Physiol. 592:4825–4838. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Yeh I, Botton T, Talevich E, Shain AH,
Sparatta AJ, de la Fouchardiere A, Mully TW, North JP, Garrido MC,
Gagnon A, et al: Activating MET kinase rearrangements in melanoma
and Spitz tumours. Nat Commun. 6:71742015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Ebrahimie E, Nurollah Z, Ebrahimi M,
Hemmatzadeh F and Ignjatovic J: Unique ability of pandemic
influenza to downregulate the genes involved in neuronal disorders.
Mol Biol Rep. 42:1377–1390. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Chu M, Qin S, Wu R, Zhou X, Tang X, Zhang
S, Zhao Q, Wang H, Liu Y, Han X, et al: Role of MiR-126a-3p in
endothelial injury in endotoxic mice. Criti Care Med. 44:e639–e650.
2016. View Article : Google Scholar
|
|
66
|
Huo X, Zhang K, Yi L, Mo Y, Liang Y, Zhao
J, Zhang Z, Xu Y and Zhen G: Decreased epithelial and plasma
miR-181b-5p expression associates with airway eosinophilic
inflammation in asthma. Clin Exp Allergy. 46:1281–1290. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Ding Z, Jian S, Peng X, Liu Y, Wang J,
Zheng L, Ou C, Wang Y, Zeng W and Zhou M: Loss of MiR-664
expression enhances cutaneous malignant melanoma proliferation by
upregulating PLP2. Medicine (Baltimore). 94:e13272015. View Article : Google Scholar
|
|
68
|
Zhu H, Miao MH, Ji XQ, Xue J and Shao XJ:
miR-664 negatively regulates PLP2 and promotes cell proliferation
and invasion in T-cell acute lymphoblastic. leukaemia Biochem
Biophys Res Commun. 459:340–345. 2015. View Article : Google Scholar
|
|
69
|
Chen Q, Lu G, Cai Y, Li Y, Xu R, Ke Y and
Zhang S: MiR-124-5p inhibits the growth of high-grade gliomas
through posttranscriptional regulation of LAMB1. Neuro Oncol.
16:637–651. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Yang M, Cui G, Ding M, Yang W, Liu Y, Dai
D and Chen L: miR-935 promotes gastric cancer cell proliferation by
targeting SOX7. Biomed Pharmacother. 79:153–158. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zhang L, Zeng D, Chen Y, Li N, Lv Y, Li Y,
Xu X and Xu G: miR-937 contributes to the lung cancer cell
proliferation by targeting INPP4B. Life Sci. 155:110–115. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Keasey MP, Scott HL, Bantounas I, Uney JB
and Kelly S: MiR-132 is upregulated by ischemic preconditioning of
cultured hippocampal neurons and protects them from subsequent OGD
toxicity. J Mol Neurosci. 59:404–410. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Stylli SS, Adamides AA, Koldej RM, Luwor
RB, Ritchie DS, Ziogas J and Kaye AH: miRNA expression profiling of
cerebrospinal fluid in patients with aneurysmal subarachnoid
hemorrhage. J Neurosurg. 126:1131–1139. 2017. View Article : Google Scholar
|
|
74
|
Pulliam JV, Xu Z, Ford GD, Liu C, Li Y,
Stovall KC, Cannon VS, Tewolde T, Moreno CS and Ford BD:
Computational identification of conserved transcription factor
binding sites upstream of genes induced in rat brain by transient
focal ischemic stroke. Brain Res. 1495:76–85. 2013. View Article : Google Scholar
|
|
75
|
Plotnik JP, Budka JA, Ferris MW and
Hollenhorst PC: ETS1 is a genome-wide effector of RAS/ERK signaling
in epithelial cells. Nucleic Acids Res. 42:11928–11940. 2014.
View Article : Google Scholar : PubMed/NCBI
|