Introduction
Oral squamous cell carcinoma (OSCC) is the sixth
most common malignancy worldwide (1,2).
According to statistics, >300,000 new cases are reported
annually in the US (3). Although
progress has been made in OSCC therapy, including surgery, and
chemo- or radiotherapy, the 5-year-survival rate of OSCC remains
<60% due to local recurrence and nodal metastasis (4). Therefore, there is an urgent
requirement to identify potential molecular therapeutic targets for
the treatment of OSCC.
Inhibitor of nuclear factor (NF)-κB (IκB) kinase β
(IKKβ), a key catalytic subunit of the IKK complex, is crucial in
the activation of NF-κB, which activates cellular programs critical
for cell survival and proliferation (5,6).
IKKβ can activate the NF-κB dimers by means of IKK-mediated,
phosphorylation-induced degradation of the IκB inhibitor, which
enables the NF-κB dimers to enter the nucleus and activate specific
target gene expression (7). In
OSCC, the activation of NF-κB induces epithelial-mesenchymal
transition, and the elevated expression of NF-κB is correlated with
enhanced invasion and metastasis (8-11).
The constitutive activation of NF-κB in OSCC has also been reported
(12). It has been reported that
inhibiting IKKβ can markedly reduce the cellular invasiveness of
SCC-25 cells and may be useful for OSCC therapy (13). However, the molecular regulatory
mechanism of the IKKβ/NF-κB signaling pathway in OSCC remains to be
fully elucidated.
MicroRNAs (miRNAs) are a class of small non-coding
RNA, which negatively regulate gene expression at the
post-transcriptional level by binding to the 3′-untranslated region
(3′-UTR) of the mRNAs of target genes (14,15). Alterations in miRNA expression
have been implicated in the pathogenesis of a variety of human
diseases, notably cancer (16-18). A number of miRNAs have been
identified to contribute to the development of OSCC. For example,
Zhang et al demonstrated that miRNA (miR)-375 was
significantly lower in OSCC tissues, and investigated the
prognostic value of miR-375 in patients with OSCC (19). Feng et al showed that
miR-22 suppressed cell proliferation, migration and invasion in
OSCC by targeting NLR family pyrin domain containing 3 (20). However, whether there are other
miRNAs involved, and the specific mechanisms require further
investigation.
In the present study, an miRNA microarray was
performed to investigate the expression of miRNAs in OSCC tissues
and the most downregulated of these, miR-199a-5p, was selected for
further analysis. In vitro experiments were performed to
investigate the functional role of miR-199a-5p in OSCC cells and to
examine the underlying mechanisms. The findings of these
experiments suggested that miR-199a-5p may be a potential target
for OSCC treatment and may be important in the development of
OSCC.
Materials and methods
Clinical specimens
Samples of 60 pairs of tumor tissues and matched
tumor-adjacent tissues were obtained from patients with OSCC with
pathologically diagnostic criteria between January 2014 and July
2016 in the Department of Oral and Maxillofacial Surgery, the First
Affiliated Hospital of Xinxiang Medical University (Weihui, China).
The clinicopathological data are shown in Table I. Written consent for tissue
donation for research purposes was obtained from each patient prior
to tissue collection. The protocol was approved by the Ethics
Committee of the First Affiliated Hospital of Xinxiang Medical
University. All the tissue samples were collected, immediately
snap-frozen in liquid nitrogen and stored at −80°C until RNA was
extracted.
 | Table ICorrelation between miR-199a-5p and
clinicopathological features in patients with oral squamous cell
carcinoma. |
Table I
Correlation between miR-199a-5p and
clinicopathological features in patients with oral squamous cell
carcinoma.
| Clinical
parameter | Cases (n=60) | miR-199-5p
expression
| P-value |
|---|
| High (22) | Low (38) |
|---|
| Sex | | | | 0.512 |
| Male | 36 | 12 | 24 | |
| Female | 24 | 10 | 14 | |
| Age (years) | | | | 0.242 |
| ≥50 | 41 | 13 | 28 | |
| <50 | 19 | 9 | 10 | |
| Site | | | | 0.381 |
| Buccal mucosa | 31 | 13 | 18 | |
| Non-buccal
mucosa | 29 | 9 | 20 | |
| Alcohol use | | | | 0.694 |
| Yes | 28 | 11 | 17 | |
| No | 32 | 11 | 21 | |
| Smoking habit | | | | 0.801 |
| Yes | 26 | 10 | 16 | |
| No | 34 | 12 | 22 | |
| Tumor size
(cm) | | | |
0.024a |
| ≥2 | 22 | 4 | 18 | |
| <2 | 38 | 18 | 20 | |
|
Differentiation | | | |
0.015a |
| Well and
moderate | 37 | 18 | 19 | |
| Poor | 23 | 4 | 19 | |
| Lymph node
metastasis | | | |
0.009b |
| Present | 35 | 8 | 27 | |
| Absent | 25 | 14 | 11 | |
| cTNM stage | | | |
0.030a |
| I + II | 15 | 9 | 6 | |
| III + IV | 45 | 13 | 32 | |
Microarray analysis
Total RNA was extracted from the OSCC tissues using
an miRNeasy Mini kit (Qiagen, Inc., Valencia, CA, USA). The purity
and quantity of the total RNA were evaluated via NanoDrop ND-1000
spectrophotometry (Thermo Fisher Scientific, Inc., Waltham, MA,
USA) and the Agilent 2100 Bioanalyzer. Total RNA (200 ng) was
labeled with fluorescence dye hy3 or hy5 using the miRCURY Hy3/Hy5
Power Labeling kit and hybridized on the miRCURY™ LNA Array
(v.16.0), both obtained from Exiqon; Qiagen, Inc., according to the
manufacturer’s protocol. Following washing with PBS, the Axon
GenePix 4000B microarray scanner (Axon Instruments; Molecular
Devices, LLC, Sunnyvale, CA, USA) was used to scan the fluorescence
intensity of the microarray. The scanned images were then imported
into the GenePix Pro 6.0 program (Axon Instruments; Molecular
Devices, LLC) for grid alignment and data extraction. Finally, the
heat map of the 57 miRNAs with the most marked differences was
created using a method of hierarchical clustering in GeneSpring GX
software, version 7.3 (Agilent Technologies, Inc., Santa Clara, CA,
USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
miRNA was prepared using the miRNeasy Mini kit
(Qiagen, Inc.) according to the manufacturer’s protocol. The
concentration and quality of total RNA was determined using a
NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Inc.).
For miRNA reverse transcription, cDNA was synthesized using the
PrimeScript RT reagent kit (Takara Bio, Inc., Tokyo, Japan). For
mRNA, total RNA was isolated using TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer’s
protocol and reverse transcribed with the SuperScript III
First-Strand Synthesis System (Thermo Fisher Scientific, Inc.).
qPCR analyses for miRNA and mRNA were performed on an ABI PRISM
7300 sequence detection system in an SYBR-Green I Real-Time PCR kit
(Applied Biosystems; Thermo Fisher Scientific, Inc.). First,
20-µl PCRs included 2 µl cDNA, 10 µl of 2X
qPCR mix, 250 nM concentrations of forward (1 µl) and
reverse primers (1 µl), and 6 µl ddH2O.
The reaction mixtures were denatured at 95°C for 3 min, followed by
40 two-step cycles of 95°C for 10 sec and 60°C for 30 sec. The
primers for RT-qPCR analysis were as follows: miR-199a-5p forward,
5′-TCCCAGTGTTCAGACTACC-3′ and miR-199a-5p reverse,
5′-TTTGGCACTAGCACATT-3′; IKKβ forward, 5′-ACTTGGCGCCCAATGACCT-3′
and IKKβ reverse, 5′-CTCTGTTCTCCTTGCTGCA-3′; GAPDH forward,
5′-GAAGATGGTGATGGGATTTC-3′ and GAPDH reverse,
5′-GAAGGTGAAGGTCGGAGT-3′; U6 forward, 5′-TGCGGGTGCTCGCTTCGCAGC-3′
and U6 reverse, 5′-CCAGTGCAGGGTCCGAGGT-3′. The expression of
miR-199a-5p and IKKβ in the tissues was normalized to the
expression of U6 and GAPDH, respectively. The RT-qPCR assays were
performed in triplicate and the change in expression level was
calculated using the 2−ΔΔCq method (21).
Cell lines and culture
The SCC-25, CAL-27, Tca8113, SCC-4 OSCC and 293 cell
lines were obtained from the American Type Culture Collection
(Manassas, VA, USA) and cultured in Dulbecco's modified Eagle's
medium supplemented with 10% fetal bovine serum (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany), 100 U/ml penicillin and 100
µg/ml streptomycin at 37°C in a 5% CO2
atmosphere. The normal oral mucosa cell line (Human Oral
Keratinocyte; HOK, Invitrogen; Thermo Fisher Scientific, Inc.) was
used as control and was maintained in oral keratinocyte media,
supplemented with 1% keratinocyte growth factor plus epithelial
growth factor mixture (Invitrogen; Thermo Fisher Scientific, Inc.)
at 37°C in a 5% CO2 atmosphere.
Cell transfection
The miR-199a-5p mimics, mimics negative control
(mimics NC), miR199a-5p inhibitor, and inhibitor NC were purchased
from Shanghai GenePharma Co., Ltd. (Shanghai, China). The detailed
information regarding miR-199a-5p mimics, miR-199a-5p inhibitor and
their controls are as follows: i) miR-199a-5p mimic sense,
5′-CCCAGUGUUCAGACUACCUGUUC-3′; mimics NC sense,
5′-CGGTGUGUUCAGACUACCUGUUC-3′; ii) miR-199a-5p inhibitor,
5′-AACAGGTAGTCTGAACACT-3′; inhibitor NC, 5′-TAACACGTCTATACGCCCA-3′.
To induce the overexpression of IKKβ, the coding domain sequences
of IKKβ mRNA were amplified by PCR, and inserted into the pcDNA 3.0
vector to enhance its expression (Invitrogen; Thermo Fisher
Scientific, Inc.), named pcDNA-IKKβ. The empty pcDNA3.1 was used as
a negative control (NC). The Tca8113 and SCC-4 cells
(1.0×106/well) were seeded and grown overnight in
six-well plates. The following day, Lipofectamine 2000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) was used for transient
transfection of the cells with miR-199a-5p mimics (50 nM),
miR-199a-5p inhibitor (100 nM) or NC (100 nM), whereas
Oligofectamine transfection reagent (Thermo Fisher Scientific,
Inc.) was used for transfection with miR-199a-5p mimics (50 nM) + 2
µg pcDNA-IKKβ for 48 h, following the manufacturer's
protocol. The transfection efficiency was confirmed by analyzing
the expression levels of miR-199a-5p using RT-qPCR at 24
post-transfection. At 48 h post-transfection, the cells were
harvested for western blot or RT-qPCR analyses, or other
experiments at the indicated times.
Cell proliferation
The Tca8113 and SCC-4 cells (5×103/well)
were seeded in 96-well plates overnight. At 24, 48 and 72 h
post-transfection, Cell Counting Kit-8 solution (CCK-8; Dojindo
Molecular Technologies, Inc., Kumamoto, Japan) was added to cells
and incubated at 37°C for an additional 2 h. The absorbance rates
were then measured at 450 nm using a micro-plate reader (Infinite
M200; Tecan Group, Ltd., Mannedorf, Austria). All experiments were
performed in triplicate.
Cell apoptosis
Apoptosis was measured using an Annexin V-FITC
Apoptosis Detection kit (Abcam, Cambridge, UK) according to the
manufacturer's protocol. At 48 h post-transfection, the Tca8113 and
SCC-4 cells were harvested and washed twice with PBS, and were
stained with Annexin V and propidium iodide (PI). Following
incubation at room temperature in the dark for 15 min, cell
apoptosis was analyzed on a FACScan flow cytometer (Beckman
Coulter, Inc., Brea, CA, USA).
Cell cycle analysis
Cell cycle distribution was determined using flow
cytometry (22). Briefly, at 48 h
post-transfection, the Tca8113 and SCC-4 cells were harvested by
trypsinization and plated in 6-cm dishes at a density of
1.0×105 cells/dish. The cells were then washed with PBS
and fixed in 70% ethanol overnight at 4°C, following which they
were then stained with 40 µg/ml PI, and incubated at 4°C for
30 min in the dark. The cells were analyzed by flow cytometry using
a FACSCalibur flow cytometer (BD Biosciences, San Jose, CA,
USA).
Immunohistochemistry (IHC)
Two paired OSCC tissues and matched tumor-adjacent
tissues were embedded in paraffin and sliced. The thickness of the
tissue sections were 4-5 mm. Immunohistochemical staining were
performed as described previously (23), IKKβ was detected using anti-IKKβ
antibody (cat. no. sc8014; 1:200; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA).
Bioinformatics analysis and luciferase
reporter assay
miRNA target prediction tools, including PicTar
version 2007 (https://pictar.mdc-berlin.de/) and TargetScan Release
7.0 (http://targetscan.org/) were used to
search for the putative targets of miR-199a-5p. The 3′-UTR of IKKβ
and the mutated sequence were inserted into the pGL3 control vector
(Promega Corporation, Madison, WI, USA) to construct the wild-type
(wt) IKKβ-3′-UTR vector and mutant IKKβ-3′-UTR vector,
respectively. For the luciferase reporter assay, 293 cells were
transfected with the corresponding vectors; at 48 h
post-transfection, the dual-luciferase reporter assay system
(Promega Corporation) was used to measure the luciferase activity.
All experiments were performed in triplicate.
Western blot analysis
Total protein was extracted using
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Shanghai, China) supplemented with protease
inhibitors (Roche Diagnostics, Guangzhou, China). The
concentrations of total cellular protein were determined using a
BCA assay kit (Pierce; Thermo Fisher Scientific, Inc.). The
extraction and isolation of nuclear and cytoplasmic proteins were
performed according to the Nuclear and Cytoplasmic Protein
Extraction kit (Beyotime Institute of Biotechnology). Briefly, the
cells were collected following washing with 1 ml ice-cold PBS and
then centrifuged for 5 min at 400 x g at 4°C, and the pellet was
dissolved with cytoplasmic protein extraction agent A supplemented
with PMSF. The tubes were vortexed for 4 sec and incubated for
10-15 min on ice to promote lysis. Cytoplasmic protein extraction
agent B was then added, vortexed for 5 sec and incubated on ice for
5 sec. The samples were then centrifuged for 5 min at 14,000 x g at
4°C and the supernatant, containing the cytoplasmic fraction, was
immediately frozen for further analysis. The pellet was
re-suspended in nuclear protein extraction agent supplemented with
PMSF. Following vortexing 15-20 times for 30 min and centrifuging
for 10 min at 14,000 x g at 4°C, the supernatants containing the
nuclear extracts were obtained. The nuclear and cytoplasmic
proteins were quantified with the BCA kit according to the
manufacturer's protocol. The protein samples (40 µg/lane)
were analyzed on an 8% SDS-PAGE gel and transferred onto
polyvinylidene difluoride membranes (GE Healthcare, Freiburg,
Germany) by electroblotting. The membranes were blocked for 1 h
with 5% non-fat milk at room temperature and then incubated with
primary antibodies against IKKβ (cat no. 8943; 1:1,000 dilution;
Cell Signaling Technology, Inc., Danvers, MA, USA), total p65 (cat.
no. 8242; 1:1,000 dilution; Cell Signaling Technology, Inc.),
nuclear phosphorylated (p-)p65 (cat. no. 3033; 1:1,000 dilution;
Cell Signaling Technology, Inc.), inhibitor of NF-κB (IκB)-α (cat.
no. sc-52900; 1:1,000 dilution; Santa Cruz Biotechnology, Inc.),
p-IκB-α (cat. no. 2859; 1:1,000 dilution; Cell Signaling
Technology, Inc.), Histone H3 (cat. no. 9728; 1:1,000 dilution;
Cell Signaling Technology, Inc.), β-actin (cat. no. 4970; 1:2,000
dilution; Cell Signaling Technology, Inc.) and α-tubulin (cat. no.
ab7291; 1:2,000; Abcam) at 4°C overnight. Following incubation with
the corresponding horseradish peroxidase-conjugated goat
anti-rabbit or goat anti-rat secondary antibodies (cat. nos. ab6721
and 6785; 1:2,000; Abcam) for 1 h at room temperature, the bands
were detected using an enhanced chemiluminescence kit (GE
Healthcare). The intensities of the bands of interest were analyzed
using ImageJ software (version 1.46; National Institutes of Health,
Bethesda, MD, USA). β-actin and α-tubulin proteins were used as the
inner controls of the cytoplasmic proteins; Histone H3 protein was
used as the inner control of the nuclear proteins. Each experiment
was run in triplicate.
NF-κB activity assay
The Tca8113 and SCC-4 cells were plated in 6-well
plates at a concentration of 5×104 cells/well. The cells
were allowed to attach overnight and then co-transfected with 20 ng
of the pGL4.32 [luc2P/NF-κB-RE/Hygro] vector and 5 ng of the pRL-TK
vector in each well (Promega Corporation). After 6 h, the cells
were washed with PBS and then transfected with miR-199a-5p mimics
and pcDNA-IKKβ for 24 h. The cells were washed in PBS and harvested
in 500 µl 1X passive lysis buffer. Luciferase activity was
quantified using the Promega luciferase assay kit on a
luminometer.
Statistical analysis
Data are presented as the mean ± standard deviation.
SPSS 19.0 statistical software (IBM Corp., Armonk, NY, USA) was
used to perform all statistical analyses. When only two groups were
compared, Student's t-test was conducted. One-way analysis of
variance followed by Tukey's post hoc test was applied to compare
differences between multiple groups. Pearson's or Spearman's
analyses were used for correlation analysis. P≤0.05 was considered
to indicate a statistically significant difference.
Results
Expression of miR-199a-5p is
downregulated in OSCC and correlates with clinicopathologic
parameters
To examine the potential involvement of miRNAs in
the development of OSCC, miRNA microarray profiling was performed
in three pairs of OSCC tissues and matched tumor-adjacent tissues.
The miRNA microarray identified 39 miRNAs that were upregulated in
OSCC and 18 miRNAs that were downregulated in OSCC tissues
(Fig. 1A). Of the downregulated
miRNAs, miR-199a-5p was identified as one of the most markedly
downregulated, which is consistent with a previous study (24). Of note, miR-199a-5p has previously
been reported to function as a tumor suppressor in a several types
of human cancers (25-29). However, its role in the
tumorigenesis of OSCC remains to be fully elucidated. Therefore,
the present study focused on miR-199a-5p in OSCC for molecular and
clinical analyses, to clarify the previously unknown role of
miR-199a-5p.
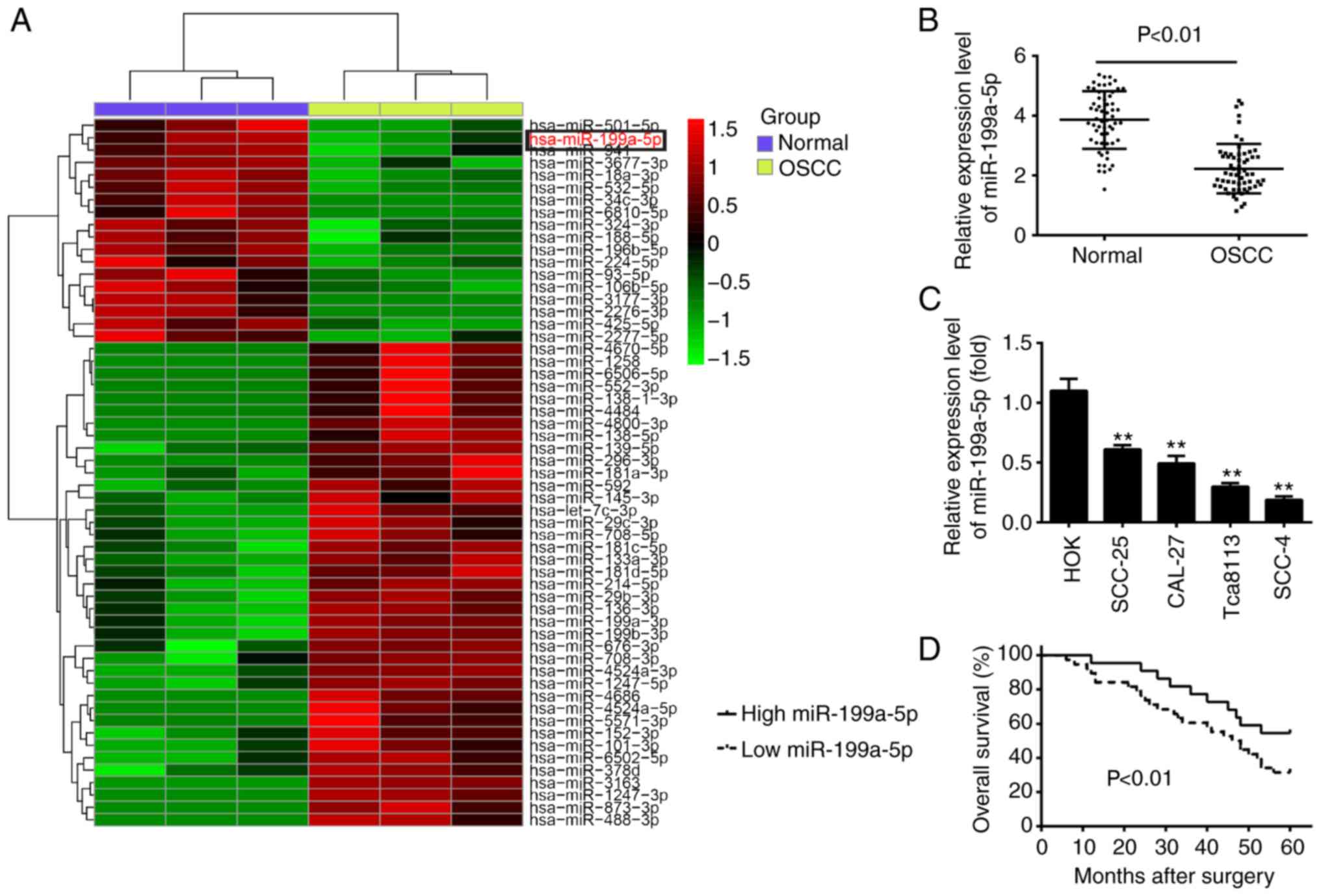 | Figure 1miR-199a-5p is downregulated in OSCC
tissues and cell lines. (A) Heat map of miRNA profiles representing
the significantly up/downregulated miRNAs. The color code in the
heat maps is linear, with green as the lowest and red as the
highest. The miRNAs that were upregulated are shown in green to
red, whereas the miRNAs that were downregulated are shown from red
to green. (B) Expression of miR-199a-5p was validated by RT-qPCR
analysis in OSCC tissues and matched tumor-adjacent tissues (n=60).
(C) Expression of miR-199a-5p was measured in four OSCC cell lines
(SCC-25, CAL-27, Tca8113 and SCC-4), and the HOK normal oral mucosa
cell line, used as a control, by RT-qPCR analysis. Data are
presented as the mean ± standard deviation of three independent
experiments. **P<0.01 vs. HOK cells. (D) Kaplan-Meier
survival curves of patients with OSCC according to the expression
of miR-199a-5p. Patients with a low tumor expression of miR-199a-5p
showed significantly shorter survival rates than those with a high
tumor expression of miR-199a-5p (P<0.01; log-rank test). OSCC,
oral squamous cell carcinoma; miR, microRNA; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; HOK, Human
Oral Keratinocyte. |
To further verify the dysregulation of miR-199a-5p,
RT-qPCR analysis was performed based on 60 paired tumor tissues and
matched tumor-adjacent tissues. The results showed that miR-199a-5p
was downregulated in the OSCC tissues (Fig. 1B). In addition, to further
elucidate whether the altered expression of miR-199a-5p occurred in
OSCC cells, RT-qPCR was performed to detect miR-199a-5p in four
OSCC cell lines (SCC-25, CAL-27, Tca8113 and SCC-4) and a normal
oral mucosa cell line (HOK) used as a control. As shown in Fig. 1C, consistent with the results in
the OSCC tissues, miR-199a-5p was significantly decreased in the
four OSCC cell lines compared with the HOK cells, suggesting that
alteration in the expression of miR-199a-5p may contribute, at
least in part, to the carcinogenesis of OSCC.
To determine the clinical values of miR-199a-5p, the
mean expression level of miR-199a-5p was used as a cut-off value to
divide 60 patients with OSCC into two groups: miR-199a-5p high
expression group and miR-199a-5p low expression group. The
associations between the expression of miR-199a-5p and
clinicopathological features are summarized in Table I. It was found that a low
expression of miR-199a-5p was associated with tumor size, tumor
differentiation, lymph node metastasis and tumor-node-metastasis
(TNM) stage. In this cohort, compared with the patients in the high
miR-199a-5p expression group, patients in the low miR-199a-5p
expression group had a lower 5-year overall survival (OS) rate
(Fig. 1D). These result indicated
that miR-199a-5p may serve as an effective biomarker for the
prognosis of patients with OSCC.
Overexpression of miR-199a-5p suppresses
cell viability, inhibits cell cycle and promotes cell
apoptosis
Previous evidence shows that miR-199a-5p has a tumor
suppressive function in several types of cancer, including ovarian
cancer and hepatocellular carcinoma (26,28). As the findings in the present
study showed that this miRNA was downregulated in OSCC, it was
hypothesized that miR-199a-5p may serve as a tumor-suppressive
miRNA in OSCC. To confirm this hypothesis, Tca8113 and SCC-4 cells,
which showed the lowest level of miR-199a-5p among the four OSCC
cell lines, were transfected with the miR-199a-5p mimics or
mimics-NC to investigate the biological function of miR-199a-5p in
OSCC. The RT-qPCR assays revealed that the expression of
miR-199a-5p was significantly upregulated following transfection
with the miR-199a-5p mimics, compared with that in the NC control
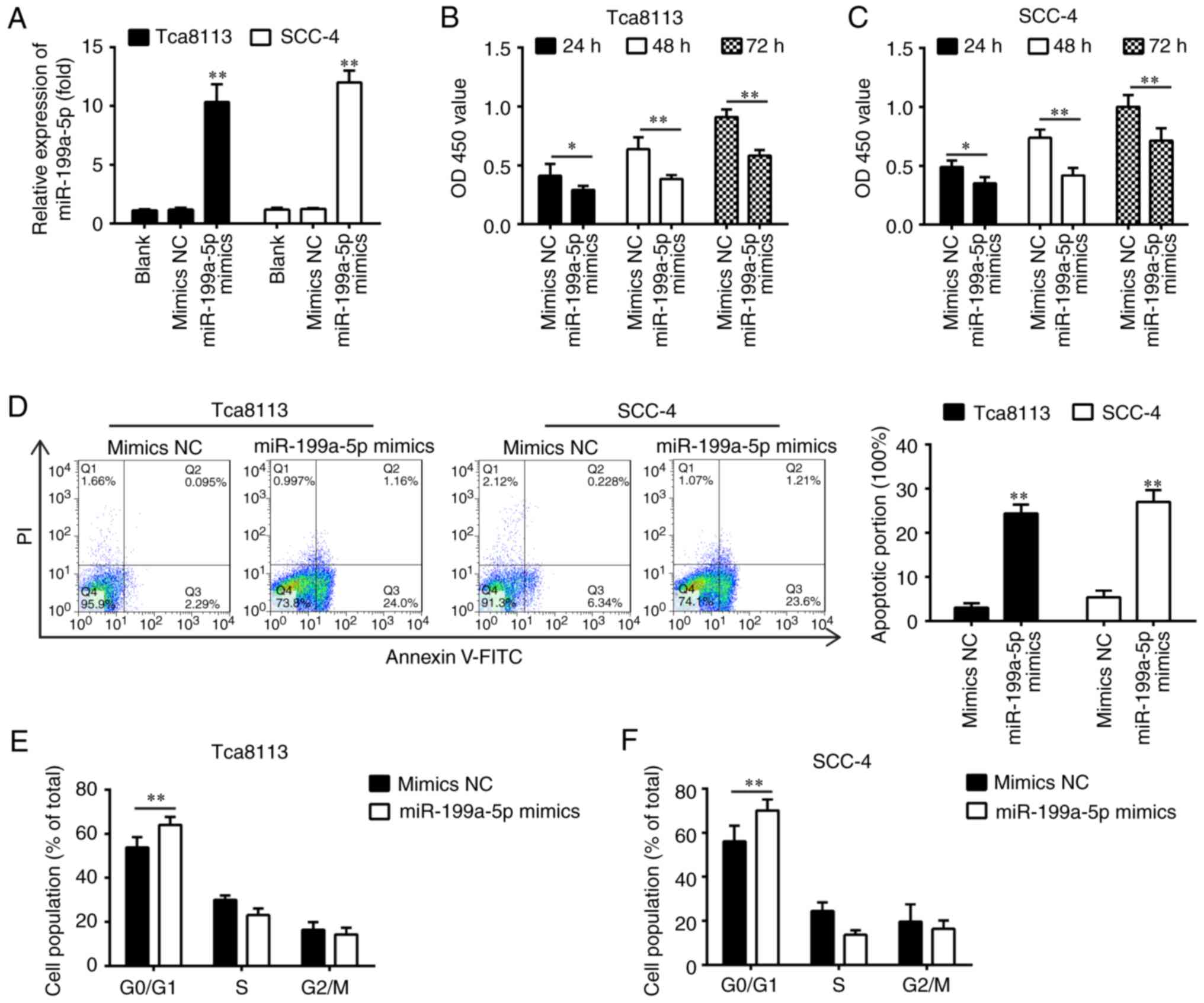
group in the Tca8113 and SCC-4 cells (Fig. 2A). The CCK-8 assay revealed that
transfection with miR-199a-5p mimics significantly inhibited cell
viability compared with that in the mimics NC-transfected cells
(Fig. 2B and C), and a
significant induction of apoptosis in the Tca8113 and SCC-4 cells
was observed (Fig. 2D). The
results of the flow cytometric analysis revealed that the
overexpression of miR-199a-5p led to G0/G1 phase arrest by
elevating the percentage of cells at the G0/G1 phase in Tca8113 and
SCC-4 cells (Fig. 2E and F),
suggesting that miR-199a-5p decreased viability partially through
inducing cell apoptosis and cell cycle arrest. These results
support the hypothesis that miR-199a-5p functions as a tumor
suppressor in the development of OSCC.
IKKβ is a direct target of
miR-199a-5p
To examine the molecular mechanism by which
miR-199a-5p functions in OSCC, candidate target genes of
miR-199a-5p were computationally screened using the TargetScan and
PicTar algorithms. Among several predicted target genes, IKKβ was
noted for its high scores in both algorithms. The bioinformatics
analysis showed that miR-199a-5p directly targeted the IKKβ gene
and that the target sequences were highly conserved among species
(Fig. 3A). In our previous study,
it was shown that IKKβ was a direct target of miR-199a-5p in
ovarian cancer cells (30).
However, the association between miR-199a-5p and IKKβ in OSCC has
not been clarified. To verify whether IKKβ is a direct target of
miR-199a-5p, a dual-luciferase reporter assay was performed by
integrating sequences of the IKKβ 3′-UTR containing the binding
sites for miR-199a-5p or corresponding mutated sequences into 293T
cells (Fig. 3B). The luciferase
reporter gene assay showed that the overexpression of miR-199a-5p
markedly repressed the luciferase activity, whereas the knockdown
of miR-199a-5p increased the relative luciferase activity of
constructs containing the wt IKKβ 3′-UTR. However, the luciferase
activity of the reporter containing the mutant binding site showed
no significant change (Fig. 3C).
Subsequently, the effect of miR-199a-5p on the expression of IKKβ
was measured at the protein level in OSCC cells by western blot
analysis. As shown in Fig. 3D,
the expression of IKKβ at the protein level was significantly
downregulated following the overexpression of miR-199a-5p, but was
upregulated following knockdown of miR-199a-5p in the Tca8113 and
SCC-4 cells. As miR-199a-5p was down-regulated in the OSCC tumor
samples, the expression levels of IKKβ were also detected in OSCC
tumor tissues and adjacent tissues by IHC. As shown in Fig. 3E, the protein expression level of
IKKβ was markedly upregulated in OSCC tissues compared with matched
tumor-adjacent tissues. In addition, the expression of IKKβ was
examined in cell lines and clinical tissue samples. As shown in
Fig. 3F and G, the expression of
IKKβ was also upregulated in cell lines and clinical tissue
samples. In addition, there was an inverse correlation between the
miR-199a-5p and IKKβ in tumor tissues (Fig. 3H). Taken together, these data
indicate that miR-199a-5p may function as a tumor suppressor by
targeting IKKβ.
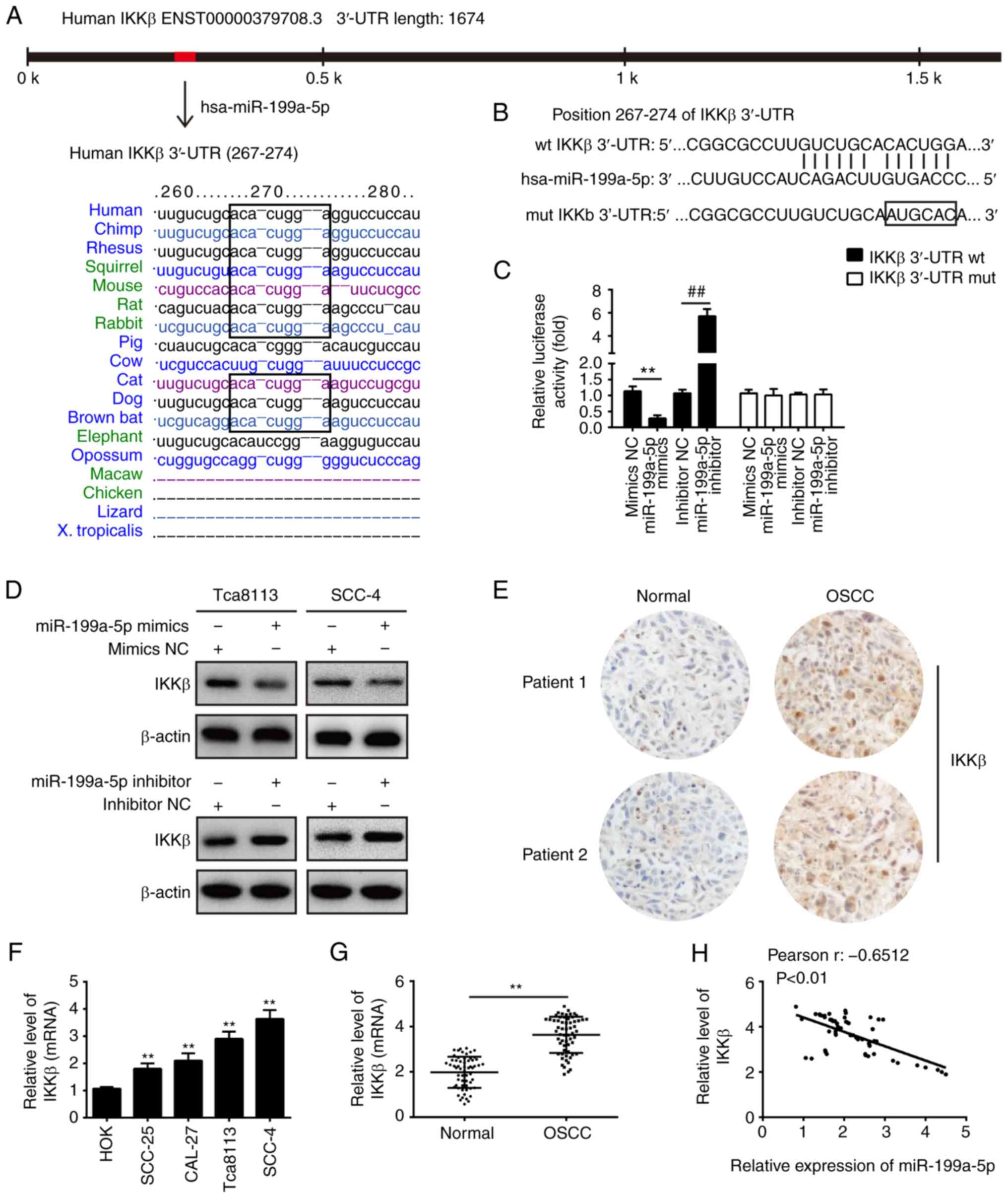 | Figure 3IKKβ is a direct target of
miR-199a-5p. (A) miR-199a-5p sequence is shown to be highly
conserved among species. (B) Putative binding sites of miR-199a-5p
and IKKβ. (C) Luciferase assay of 293T cells co-transfected with
firefly luciferase constructs containing the IKKβ wt or mut 3′-UTRs
and miR-199a-5p mimics, mimics NC, miR-199a-5p inhibitor or
inhibitor NC, as indicated (n=3). Data are presented as the mean ±
standard deviation of three independent experiments.
**P<0.01 vs. mimics NC, ##P<0.01 vs.
inhibitor NC. (D) Protein expression of IKKβ following transfection
with miR-199a-5p mimics or miR-199a-5p inhibitor was measured by
western blot analysis. **P<0.01 vs. mimics NC,
##P<0.01 vs. inhibitor NC. (E) Expression of IKKβ was
assessed by immunohistochemistry in OSCC tissues and matched
tumor-adjacent tissues (magnification, x200). (F) Expression was of
IKKβ examined in four OSCC cell lines (SCC-25, CAL-27, Tca8113 and
SCC-4) and HOK cells, used as a control, by RT-qPCR analysis. Data
are presented as the mean ± standard deviation of three independent
experiments. **P<0.01 vs. HOK cells. (G) Expression
of IKKβ was measured by RT-qPCR analysis in OSCC tissues and
matched tumor-adjacent tissues (n=60). **P<0.01 vs.
Normal group. (H) Spearman's rank correlation analysis revealed a
negative correlation between the expression of IKKβ and miR-199a-5p
(r=-0.6512, P<0.01). miR, microRNA; 3′-UTR, 3′-untranslated
region; wt, wild-type; mut, mutant; NC, negative control; IKKβ,
inhibitor of nuclear factor-κB kinase β; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; HOK, Human
Oral Keratinocyte. |
miR-199a-5p inhibits cell viability and
induces cell apoptosis by targeting IKKβ
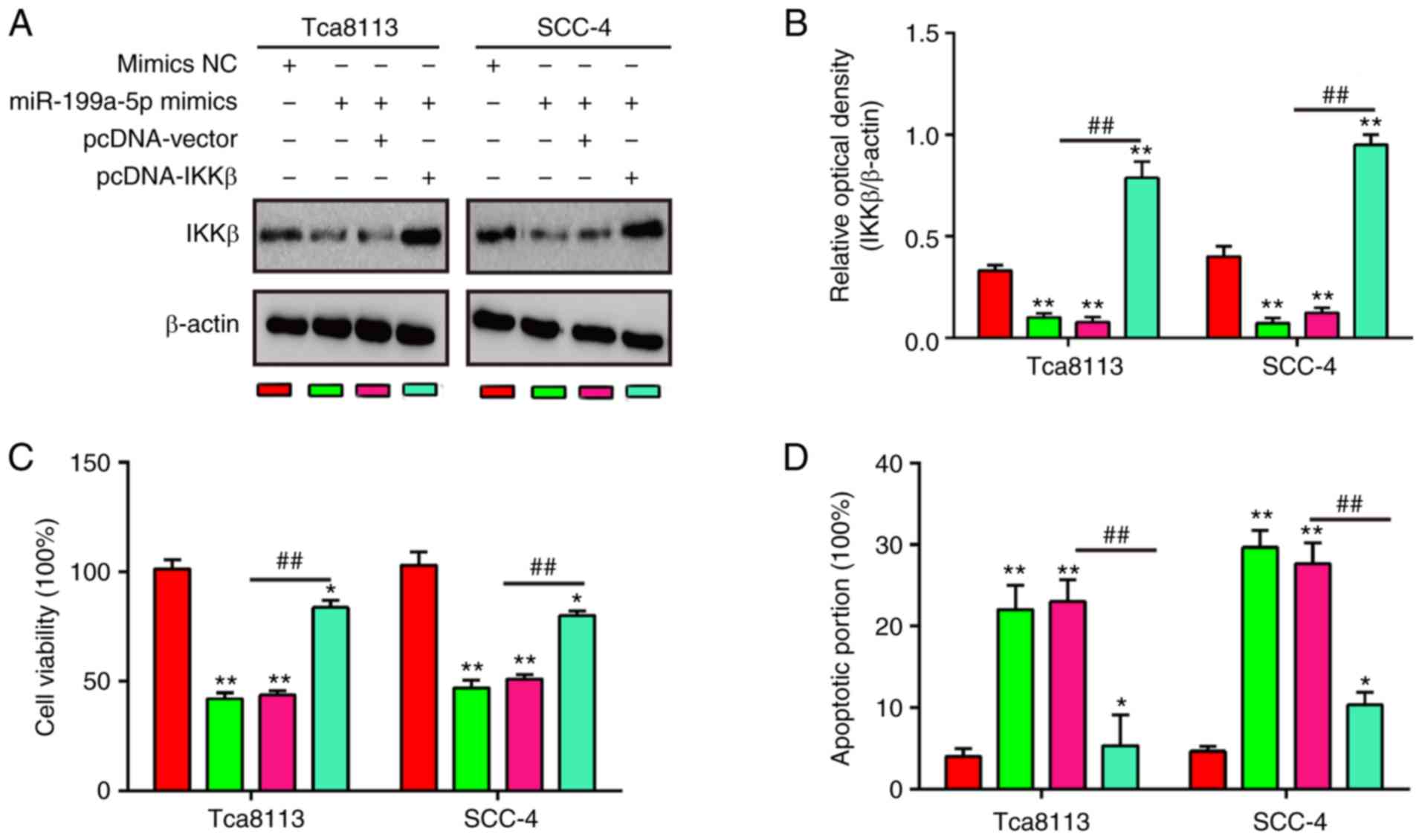
To investigate whether IKKβ mediated the inhibitory
effects of miR-199a-5p on cell viability and cell apoptosis, rescue
experiments were performed by trans-fecting pcDNA-IKKβ into Tca8113
and SCC-4 cells with higher expression of miR-199a-5p. It was found
that the protein expression of IKKβ was significantly increased in
the miR-199a-5p-overexpressing Tca8113 and SCC-4 cells when the
pcDNA-IKKβ plasmid was transfected (Fig. 4A and B). Cell viability and cell
apoptosis were then examined using a CCK-8 assay and flow
cytometry. As shown in Fig. 4C,
the decreased cell viability induced by miR-199a-5p mimics was
rescued by the overexpression of IKKβ. Furthermore, the flow
cytometry data indicated that the promotion of apoptosis caused by
miR-199a-5p mimics was attenuated when IKKβ was overexpressed
(Fig. 4D). These findings
demonstrated that IKKβ is involved in the tumor-suppressive
functions of miR-199a-5p in OSCC cells.
miR-199a-5p inhibits the IKKβ-mediated
activation of the NF-κB pathway
IKKβ has been shown to activate the NF-κB pathway
(31-33), and IKKβ/NF-κB is a classic
signaling pathway that is important in tumorigenesis (34,35). To investigate whether miR-199a-5p
influences the activity of the NF-κB signaling pathway, NF-κB
reporter luciferase activity and the expression levels of
downstream proteins in the NF-κB signaling pathway, namely
total-p65, cytoplasm-p-p65, nuclear p-p65, p-IκB-α and IκB-α, were
evaluated. In addition, the phosphorylation of IκB-α (levels of
p-IκB-α) were quantified in Tca8113 and SCC-4 cells. The results
showed that the miR-199a-5p mimics suppressed the levels of
p-IκB-α, whereas the overexpression of IKKβ reversed these
inhibitory effects of the miR-199a-5p mimics in Tca8113 and SCC-4
cells (Fig. 5A and B). To confirm
the possibility that miR-199a-5p can suppress the NF-κB signaling
pathway, the effect of miR-199a-5p on the expression of cytoplasmic
p-p65 and nuclear p-p65 was examined. As shown in Fig. 5A, C and D, the expression of
cytoplasmic p-p65 was significantly increased in the
miR-199a-5p-overexpressing Tca8113 and SCC-4 cells, whereas the
reverse changes were observed in the expression level of nuclear
p-p65, compared with the mimics NC group. By contrast, the
overexpression of IKKβ reversed the effects of miR-199a-5p on the
expression levels of cytoplasmic p-p65 and nuclear p-p65.
Furthermore, the overexpression of miR-199a-5p led to a significant
decrease in NF-κB reporter luciferase activity, and this reduction
was reversed by the overexpression of IKKβ (Fig. 5E). Overall, these results
demonstrate that miR-199a-5p inhibited the activation of the NF-κB
pathway through the downregulation of IKKβ.
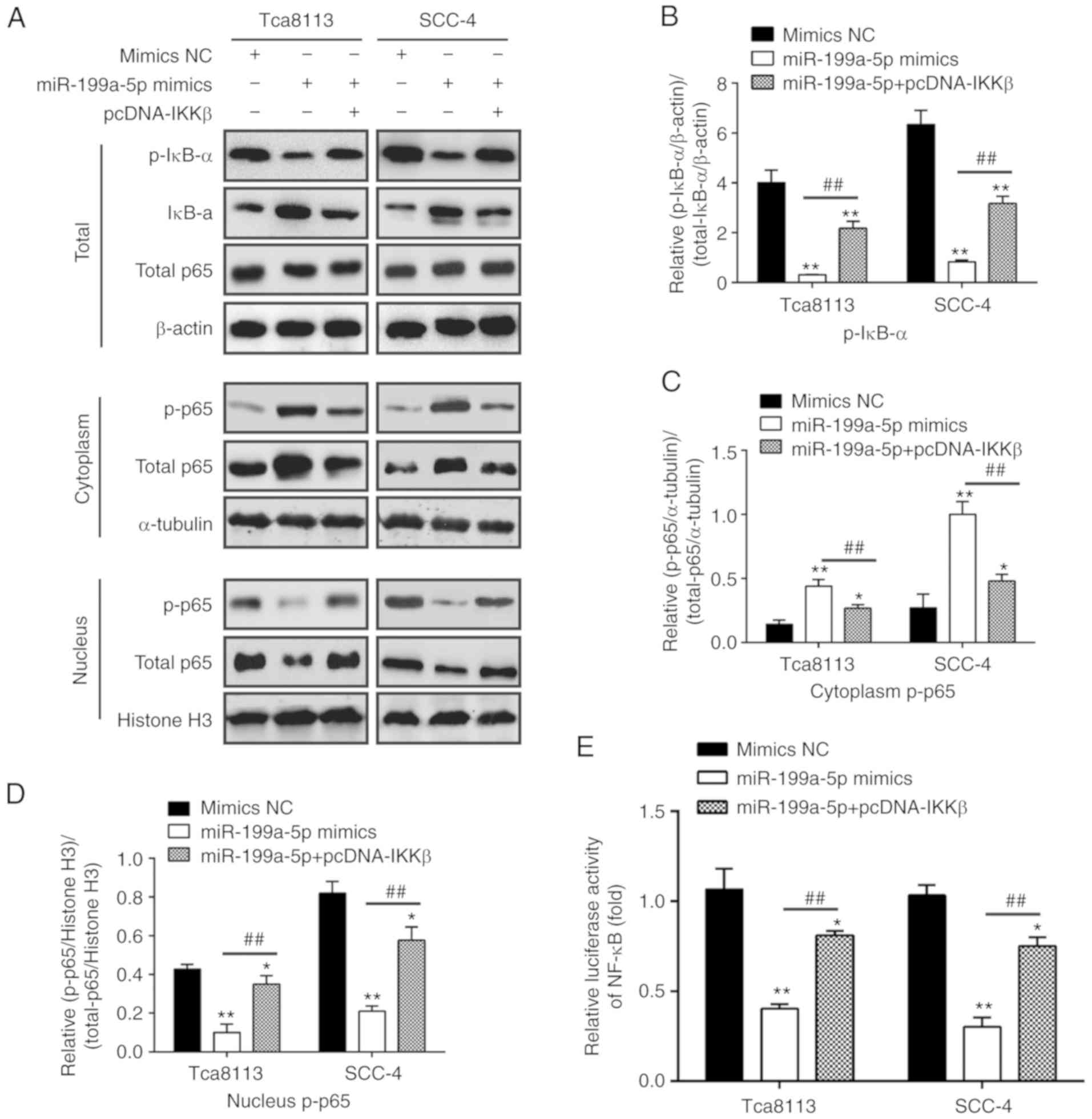 | Figure 5miR-199a-5p inhibits the
IKKβ-mediated activation of the NF-κB pathway. Tca8113 and SCC-4
cells were transfected with the miR-199a-5p mimics or mimics-NC for
48 h, and were used for western blot and NF-κB activity assays. (A)
Levels of nuclear p-p65, cytoplasm-p-p65, total p65, p-IκB-α and
IκB-α were measured by western blot analysis in the whole cell
lysate (upper), cytoplasm (middle) and nuclei (lower). β-actin
protein was used as the inner control of total proteins; α-tubulin
and Histone H3 protein was used as the inner control of the
cytoplasmic and nuclear proteins, respectively. (B) Phosphorylation
levels of IκB-α were quantified as (p-IκB-α/control)/(total
IκB-α/control). Expression levels of p-p65 in the (C) cytoplasm and
(D) nucleus were quantified. α-tubulin protein was used as the
inner control of the cytoplasmic proteins; Histone H3 protein was
used as the inner control of the nuclear proteins. (E) NF-κB
activity was quantified using a Promega luciferase assay kit. Data
are presented as the mean ± standard deviation of three independent
experiments. *P<0.05 and **P<0.01 vs.
mimics NC group; ##P< 0.01 vs. miR-199a-5p mimics
group. miR, microRNA; NC, negative control; NF-κB, nuclear
factor-κB; IκB-α, inhibitor of NF-κB-α; IKKβ, inhibitor of NF-κB
kinase β; p-, phosphorylated. |
Discussion
In the present study, miR-199a-5p was found to be
down-regulated in OSCC tissues and cell lines, and a low expression
of miR-199a-5p was associated with tumor differentiation, lymph
node metastasis and TNM stage. The overexpression of miR-199a-5p
also suppressed cell viability, caused cell cycle arrest and
promoted cell apoptosis. Additionally, IKKβ was confirmed as a
functional target of miR-199a-5p and miR-199a-5p inhibited the
IKKβ-mediated activation of the NF-κB pathway, and thus inhibited
malignancy in OSCC cells. These findings indicate that miR-199a-5p,
as a potential biomarker for the clinical diagnosis and prognosis
of OSCC, may be an effective anticancer target for the treatment of
OSCC.
Increasing evidence has demonstrated that miRNAs
function as either oncogenic or tumor-suppressing genes in OSCC.
For example, Lu et al found that miR-654-5p was upregulated
in late-stage OSCC tissues, and promoted the proliferation and
metastasis of OSCC in vitro and in vivo (36). Shiah et al demonstrated
that miR-329 and miR-410 promoted the proliferation and
invasiveness of OSCC cells by targeting Wnt-7b (37). Wang et al showed that
miR-139-5p was downregulated in OSCC tissues, and that the
overexpression of miR-139-5p inhibited the proliferation, invasion
and migration ability of OSCC cells by targeting homeobox A9
(38). Understanding the role of
miRNAs that are aberrantly expressed in OSCC can assist in
understanding the underlying mechanisms of OSCC and improve
therapeutic approaches for OSCC. In the present study, a large set
of miRNAs were found to be significantly deregulated in OSCC
tissues using an miRNA microarray, and miR-199a-5p was one of the
most markedly downregulated miRNAs. Its lower expression was
further confirmed by RT-qPCR analysis. It was also observed that a
low expression of miR-199a-5p was closely associated with tumor
differentiation, lymph node metastasis, TNM stage, and a poor OS
rate. Taken together, these findings suggest that miR-199a-5p may
be important in OSCC carcinogenesis.
A large number of studies have investigated the
expression of miR-199a-5p in human cancer and have reported it to
be downregulated in several types of cancer (39,40). Several studies have identified the
tumor suppressor functions of miR-199a-5p (41-43). For example, Cheng et al
showed that miR-199a suppressed the proliferation of ovarian
cancer-initiating cells in vitro and in vivo by
targeting targets cluster of differentiation-44 (26). In addition, it was shown that the
re-expression of miR-199a suppressed renal cancer cell
proliferation and survival by targeting glycogen synthase
kinase-3-β (GSK-β) (27).
However, whether miR-199a-5p was involved in OSCC remained to be
elucidated. In the present study, the experiments showed that the
enforced expression of miR-199a-5p inhibited cell proliferation,
inhibited cell cycle and induced the apoptosis of Tca8113 and SCC-4
cells, indicating that miR-199a-5p also serves as a tumor
suppressor in OSCC.
miR-199a-5p has been reported to downregulate the
expression of several target genes in different types of tumor,
including CD44 (25), GSK-3β
(27) and connective tissue
growth factor (44). IKKβ, one of
the catalytic subunits of the IKK complex, is an inhibitor of the
NF-κB signaling pathway (5).
Several studies have shown that IKKβ functions as an oncogene in
different types of carcinoma (45-47). For example, Greten et al
found that IKKβ contributed to tumor promotion by suppressing
apoptosis through the mitochondrial pathway in epithelial cells
(48). Zhang et al
demonstrated that IKKβ promoted proliferation and migration, and
inhibited apoptosis in prostate cancer cells (49). In addition IKKβ acts as downstream
molecule of certain miRNAs to mediate the role of the miRNAs in
different tumor types, including miR-429 (50) and miR-497 (51). In the present study, IKKβ was
predicted to be a target of miR-199a-5p using online informatics
tools, and this hypothesis was tested and confirmed by a luciferase
reporter assay. In addition, it was found that the levels of IKKβ
in cell lines and OSCC tissues were significantly higher than those
in HOK cells and matched adjacent tissues, respectively. It was
also found that there was an inverse correlation between
miR-199a-5p and IKKβ in tumor tissues. Furthermore, the
overexpression of IKKβ reversed the suppressive effects induced by
the enhanced expression of miR-199a-5p in OSCC cells. Taken
together, these data indicate that miR-199a-5p exerts its antitumor
effects by targeting IKKβ.
Activation of canonical NF-κB has been found to be
important in the pathogenesis of several types of tumor in humans
(45,52). In OSCC, NF-κB is constitutively
active and is involved in promoting the invasion of OSCC cells,
which suggests that inhibiting the activity of NF-κB may constitute
a promising therapeutic approach to treat the invasiveness of OSCC
(13). A previous study showed
that the upregulation of miR-92b accelerates tumor growth and that
this effect may be associated with activation of the NF-κB
signaling pathway in OSCC (53).
In the present study, the results showed that the upregulation of
miR-199a-5p reduced the levels of key NF-κB pathway proteins by
suppressing IKKβ. The data suggested that miR-199a-5p suppressed
IKKβ to inhibit NF-κB activity and therefore inhibit the malignancy
of OSCC cells.
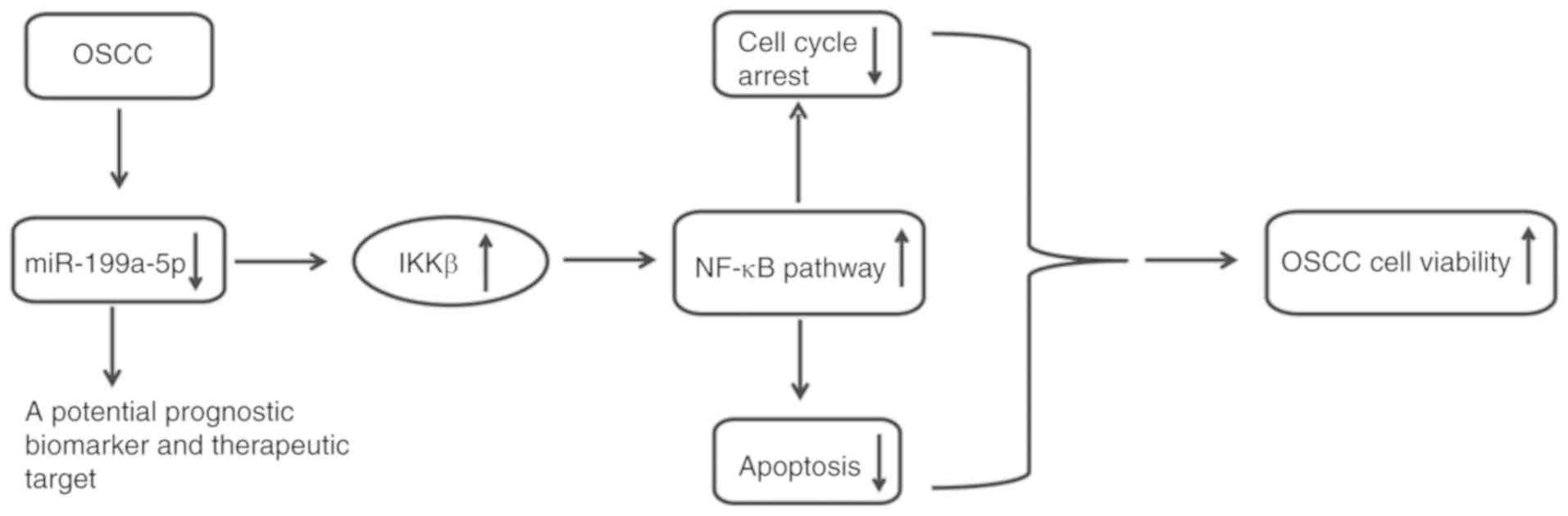
In conclusion, the results of the present study
indicated that miR-199a-5p was markedly downregulated in OSCC
tissues and cell lines, and the decreased level of miR-199a-5p was
relative to tumor differentiation, metastatic lymph nodes and
advanced TNM stage in patients with OSCC. It was also demonstrated
that miR-199a-5p functions as a tumor suppressor via the
suppression of IKKβ, which inhibits activation of the NF-κB pathway
(Fig. 6). These findings indicate
that miR-199a-5p may be a potential target for prognostic
prediction and therapeutic strategies.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
DW, BS, WW, YZ, XY, GL and JY performed the
experiments, contributed to data analysis and wrote the manuscript.
DW, BS, WW, YZ, XY, GL and JY analyzed the data. YS conceptualized
the study design, and contributed to data analysis and experimental
materials. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All individuals provided informed consent for the
use of human specimens for clinical research. The present study was
approved by the First Affiliated Hospital of Xinxiang Medical
University Ethics Committees.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J,
Murray T and Thun MJ: Cancer statistics, 2008. CA. Cancer J Clin.
58:71–96. 2008. View Article : Google Scholar
|
|
2
|
Silverman S Jr: Demographics and
occurrence of oral and pharyngeal cancers. The outcomes, the
trends, the challenge. J Am Dent Assoc. 132(Suppl 132): S7–S11.
2001. View Article : Google Scholar
|
|
3
|
Su L, Wang Y, Xiao M, Lin Y and Yu L:
Up-regulation of survivin in oral squamous cell carcinoma
correlates with poor prognosis and chemoresistance. Oral Surg Oral
Med Oral Pathol Oral Radiol Endod. 110:484–491. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Genden EM, Ferlito A, Bradley PJ, Rinaldo
A and Scully C: Neck disease and distant metastases. Oral Oncol.
39:207–212. 2003. View Article : Google Scholar
|
|
5
|
Menssen A, Häupl T, Sittinger M, Delorme
B, Charbord P and Ringe J: Differential gene expression profiling
of human bone marrow-derived mesenchymal stem cells during
adipogenic development. BMC Genomics. 12:4612011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Perkins ND: Integrating cell-signalling
pathways with NF-kappaB and IKK function. Nat Rev Mol Cell Biol.
8:49–62. 2007. View
Article : Google Scholar
|
|
7
|
Pan MH, Lin-Shiau SY and Lin JK:
Comparative studies on the suppression of nitric oxide synthase by
curcumin and its hydrogenated metabolites through downregulation of
IkappaB kinase and NFkappaB activation in macrophages. Biochem
Pharmacol. 60:1665–1676. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tanaka T, Nakayama H, Yoshitake Y, Irie A,
Nagata M, Kawahara K, Takamune Y, Yoshida R, Nakagawa Y, Ogi H, et
al: Selective inhibition of nuclear factor-kappaB by nuclear
factor-kappaB essential modulator-binding domain peptide suppresses
the metastasis of highly metastatic oral squamous cell carcinoma.
Cancer Sci. 103:455–463. 2012. View Article : Google Scholar
|
|
9
|
Aggarwal BB and Sung B: NF-κB in cancer: A
matter of life and death. Cancer Discov. 1:469–471. 2011.
View Article : Google Scholar
|
|
10
|
Nakayama H, Ikebe T, Beppu M and Shirasuna
K: High expression levels of nuclear factor kappaB, IkappaB kinase
alpha and Akt kinase in squamous cell carcinoma of the oral cavity.
Cancer. 92:3037–3044. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Julien S, Puig I, Caretti E, Bonaventure
J, Nelles L, van Roy F, Dargemont C, de Herreros AG, Bellacosa A
and Larue L: Activation of NF-kappaB by Akt upregulates Snail
expression and induces epithelium mesenchyme transition. Oncogene.
26:7445–7456. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Furuta H, Osawa K, Shin M, Ishikawa A,
Matsuo K, Khan M, Aoki K, Ohya K, Okamoto M, Tominaga K, et al:
Selective inhibition of NF-kappaB suppresses bone invasion by oral
squamous cell carcinoma in vivo. Int J Cancer. 131:E625–E635. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Johnson J, Shi Z, Liu Y and Stack MS:
Inhibitors of NF-kappaB reverse cellular invasion and target gene
upregulation in an experimental model of aggressive oral squamous
cell carcinoma. Oral Oncol. 50:468–477. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Barwari T, Joshi A and Mayr M: MicroRNAs
in cardiovascular disease. J Am Coll Cardiol. 68:2577–2584. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Alipoor SD, Adcock IM, Garssen J, Mortaz
E, Varahram M, Mirsaeidi M and Velayati A: The roles of miRNAs as
potential biomarkers in lung diseases. Eur J Pharmacol.
791:395–404. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
McManus MT: MicroRNAs and cancer. Semin
Cancer Biol. 13:253–258. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang B, Li Y, Hou D, Shi Q, Yang S and Li
Q: MicroRNA-375 inhibits growth and enhances radiosensitivity in
oral squamous cell carcinoma by targeting insulin like growth
factor 1 receptor. Cell Physiol Biochem. 42:2105–2117. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Feng X, Luo Q, Wang H, Zhang H and Chen F:
MicroRNA-22 suppresses cell proliferation, migration and invasion
in oral squamous cell carcinoma by targeting NLRP3. J Cell Physiol.
233:6705–6713. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔC T method. Methods. 25:402–408.
2001. View Article : Google Scholar
|
|
22
|
Rasola A and Geuna M: A flow cytometry
assay simultaneously detects independent apoptotic parameters.
Cytometry. 45:151–157. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Price LC, Caramori G, Perros F, Meng C,
Gambaryan N, Dorfmuller P, Montani D, Casolari P, Zhu J, Dimopoulos
K, et al: Nuclear factor κ-B is activated in the pulmonary vessels
of patients with end-stage idiopathic pulmonary arterial
hypertension. PLoS One. 8:e754152013. View Article : Google Scholar
|
|
24
|
Yu T, Wang XY, Gong RG, Li A, Yang S, Cao
YT, Wen YM, Wang CM and Yi XZ: The expression profile of microRNAs
in a model of 7,12-dimethylbenz[a]anthrance-induced oral
carcinogenesis in Syrian hamster. J Exp Clin Cancer Res. 28:642009.
View Article : Google Scholar
|
|
25
|
He J, Jing Y, Li W, Qian X, Xu Q, Li FS,
Liu LZ, Jiang BH and Jiang Y: Roles and mechanism of miR-199a and
miR-125b in tumor angiogenesis. PLoS One. 8:e566472013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cheng W, Liu T, Wan X, Gao Y and Wang H:
MicroRNA-199a targets CD44 to suppress the tumorigenicity and
multidrug resistance of ovarian cancer-initiating cells. FEBS J.
279:2047–2059. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tsukigi M, Bilim V, Yuuki K, Ugolkov A,
Naito S, Nagaoka A, Kato T, Motoyama T and Tomita Y: Re-expression
of miR-199a suppresses renal cancer cell proliferation and survival
by targeting GSK-3beta. Cancer Lett. 315:189–197. 2012. View Article : Google Scholar
|
|
28
|
Jia XQ, Cheng HQ, Qian X, Bian CX, Shi ZM,
Zhang JP, Jiang BH and Feng ZQ: Lentivirus-mediated overexpression
of microRNA-199a inhibits cell proliferation of human
hepatocel-lular carcinoma. Cell Biochem Biophys. 62:237–244. 2012.
View Article : Google Scholar
|
|
29
|
Huang L, Lin JX, Yu YH, Zhang MY, Wang HY
and Zheng M: Downregulation of six microRNAs is associated with
advanced stage, lymph node metastasis and poor prognosis in small
cell carcinoma of the cervix. PLoS One. 7:e337622012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen R, Alvero AB, Silasi DA, Kelly MG,
Fest S, Visintin I, Leiser A, Schwartz PE, Rutherford T and Mor G:
Regulation of IKKbeta by miR-199a affects NF-kappaB activity in
ovarian cancer cells. Oncogene. 27:4712–4723. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tsuchiya Y, Osaki K, Kanamoto M, Nakao Y,
Takahashi E, Higuchi T and Kamata H: Distinct B subunits of PP2A
regulate the NF-kappaB signalling pathway through dephosphorylation
of IKKβ, IkappaBα and RelA. FEBS Lett. 591:4083–4094. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fang R, Wang C, Jiang Q, Lv M, Gao P, Yu
X, Mu P, Zhang R, Bi S, Feng JM and Jiang Z: NEMO-IKKβ are
essential for IRF3 and NF-κB activation in the cGAS-STING pathway.
J Immunol. 199:3222–3233. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li ZW, Chu W, Hu Y, Delhase M, Deerinck T,
Ellisman M, Johnson R and Karin M: The IKKbeta subunit of IkappaB
kinase (IKK) is essential for nuclear factor kappaB activation and
prevention of apoptosis. J Exp Med. 189:1839–1845. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ma XF, Zhang J, Shuai HL, Guan BZ, Luo X
and Yan RL: IKKbeta/NF-kappaB mediated the low doses of bisphenol A
induced migration of cervical cancer cells. Arch Biochem Biophys.
573:52–58. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
He G, Yu GY, Temkin V, Ogata H, Kuntzen C,
Sakurai T, Sieghart W, Peck-Radosavljevic M, Leffert HL and Karin
M: Hepatocyte IKKbeta/NF-kappaB inhibits tumor promotion and
progression by preventing oxidative stress-driven STAT3 activation.
Cancer Cell. 17:286–297. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lu M, Wang C, Chen W, Mao C and Wang J:
miR-654-5p targets GRAP to promote proliferation, metastasis, and
chemoresistance of oral squamous cell carcinoma through Ras/MAPK
signaling. DNA Cell Biol. 37:381–388. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shiah SG, Hsiao JR, Chang WM, Chen YW, Jin
YT, Wong TY, Huang JS, Tsai ST, Hsu YM, Chou ST, et al:
Downregulated miR329 and miR410 promote the proliferation and
invasion of oral squamous cell carcinoma by targeting Wnt-7b.
Cancer Res. 74:7560–7572. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang K, Jin J, Ma T and Zhai H: MiR-139-5p
inhibits the tumorigenesis and progression of oral squamous
carcinoma cells by targeting HOXA9. J Cell Mol Med. 21:3730–3740.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Murakami Y, Yasuda T, Saigo K, Urashima T,
Toyoda H, Okanoue T and Shimotohno K: Comprehensive analysis of
microRNA expression patterns in hepatocellular carcinoma and
non-tumorous tissues. Oncogene. 25:2537–2545. 2006. View Article : Google Scholar
|
|
40
|
Jiang J, Gusev Y, Aderca I, Mettler TA,
Nagorney DM, Brackett DJ, Roberts LR and Schmittgen TD: Association
of MicroRNA expression in hepatocellular carcinomas with hepatitis
infection, cirrhosis, and patient survival. Clin Cancer Res.
14:419–427. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shen Q, Cicinnati VR, Zhang X, Iacob S,
Weber F, Sotiropoulos GC, Radtke A, Lu M, Paul A, Gerken G and
Beckebaum S: Role of microRNA-199a-5p and discoidin domain receptor
1 in human hepatocellular carcinoma invasion. Mol Cancer.
9:2272010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Su SF, Chang YW, Andreu-Vieyra C, Fang JY,
Yang Z, Han B, Lee AS and Liang G: miR-30d, miR-181a and
miR-199a-5p cooperatively suppress the endoplasmic reticulum
chaperone and signaling regulator GRP78 in cancer. Oncogene.
32:4694–4701. 2013. View Article : Google Scholar :
|
|
43
|
Xu N, Zhang J, Shen C, Luo Y, Xia L, Xue F
and Xia Q: Cisplatin-induced downregulation of miR-199a-5p
increases drug resistance by activating autophagy in HCC cell.
Biochem Biophys Res Commun. 423:826–831. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sun D, Han S, Liu C, Zhou R, Sun W, Zhang
Z and Qu J: Microrna-199a-5p functions as a tumor suppressor via
suppressing connective tissue growth factor (CTGF) in follicular
thyroid carcinoma. Med Sci Monit. 22:1210–1217. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Dai L, Gu L and Di W: MiR-199a attenuates
endometrial stromal cell invasiveness through suppression of the
IKKβ/NF-κB pathway and reduced interleukin-8 expression. Mol Hum
Reprod. 18:136–145. 2012. View Article : Google Scholar
|
|
46
|
Jing H and Lee S: NF-κB in cellular
senescence and cancer treatment. Mol Cells. 37:189–195. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yang J, Kantrow S, Sai J, Hawkins OE,
Boothby M, Ayers GD, Young ED, Demicco EG, Lazar AJ, Lev D and
Richmond A: INK4a/ARF [corrected] inactivation with activation of
the NF-kappaB/IL-6 pathway is sufficient to drive the development
and growth of angiosarcoma. Cancer Res. 72:4682–4695. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Greten FR, Eckmann L, Greten TF, Park JM,
Li ZW, Egan LJ, Kagnoff MF and Karin M: IKKbeta links inflammation
and tumorigenesis in a mouse model of colitis-associated cancer.
Cell. 118:285–296. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang Y, Lapidus RG, Liu P, Choi EY,
Adediran S, Hussain A, Wang X, Liu X and Dan HC: Targeting IκB
kinase β/NF-κB signaling in human prostate cancer by a novel IκB
kinase β inhibitor CmpdA. Mol Cancer Ther. 15:1504–1514. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fan JY, Fan YJ, Wang XL, Xie H, Gao HJ,
Zhang Y, Liu M and Tang H: miR-429 is involved in regulation of
NF-κB activity by targeting IKKβ and suppresses oncogenic activity
in cervical cancer cells. FEBS Lett. 591:118–128. 2017. View Article : Google Scholar
|
|
51
|
Kong XJ, Duan LJ, Qian XQ, Xu D, Liu HL,
Zhu YJ and Qi J: Tumor-suppressive microRNA-497 targets IKKβ to
regulate NF-κB signaling pathway in human prostate cancer cells. Am
J Cancer Res. 5:1795–1804. 2015.
|
|
52
|
Karin M, Cao Y, Greten FR and Li ZW:
NF-kappaB in cancer: From innocent bystander to major culprit. Nat
Rev Cancer. 2:301–310. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
53
|
Liu Z, Diep C, Mao T, Huang L, Merrill R,
Zhang Z and Peng Y: MicroRNA-92b promotes tumor growth and
activation of NF-κB signaling via regulation of NLK in oral
squamous cell carcinoma. Oncol Rep. 34:2961–2968. 2015. View Article : Google Scholar : PubMed/NCBI
|