1. Introduction
The irreversible process of aging is marked by a
decline in the body's physiological functions and adaptation
ability. One of the systems impaired by this process is the
reproductive system. According to recent publications, infertility
affects an estimated 15% of couples globally (1). In 2016, almost 66,000 births in the
United States were achieved from the assisted reproduction
techniques application (2).
According to the statistics of the Society for Assisted
Reproductive Technology (SART), 241,570 cycles for egg retrieval,
frozen embryo transfer and frozen egg thawing were performed
(3). Despite common belief that
infertility factors are usually associated with only the female
sex, male factors have been found to be solely responsible for
20-30% of infertility cases and to contribute to 50% of the cases
overall (4). The deleterious
effects of reproductive aging affect both males and females;
however, an earlier onset is observed among females, as a sharp
decrease in fertility is reported after the age of 35 (5). The process of aging is strongly
influenced by genetics, as well as environmental and lifestyle
factors. Currently, the aging process is divided into two major
components, biological and chronological age, which can differ for
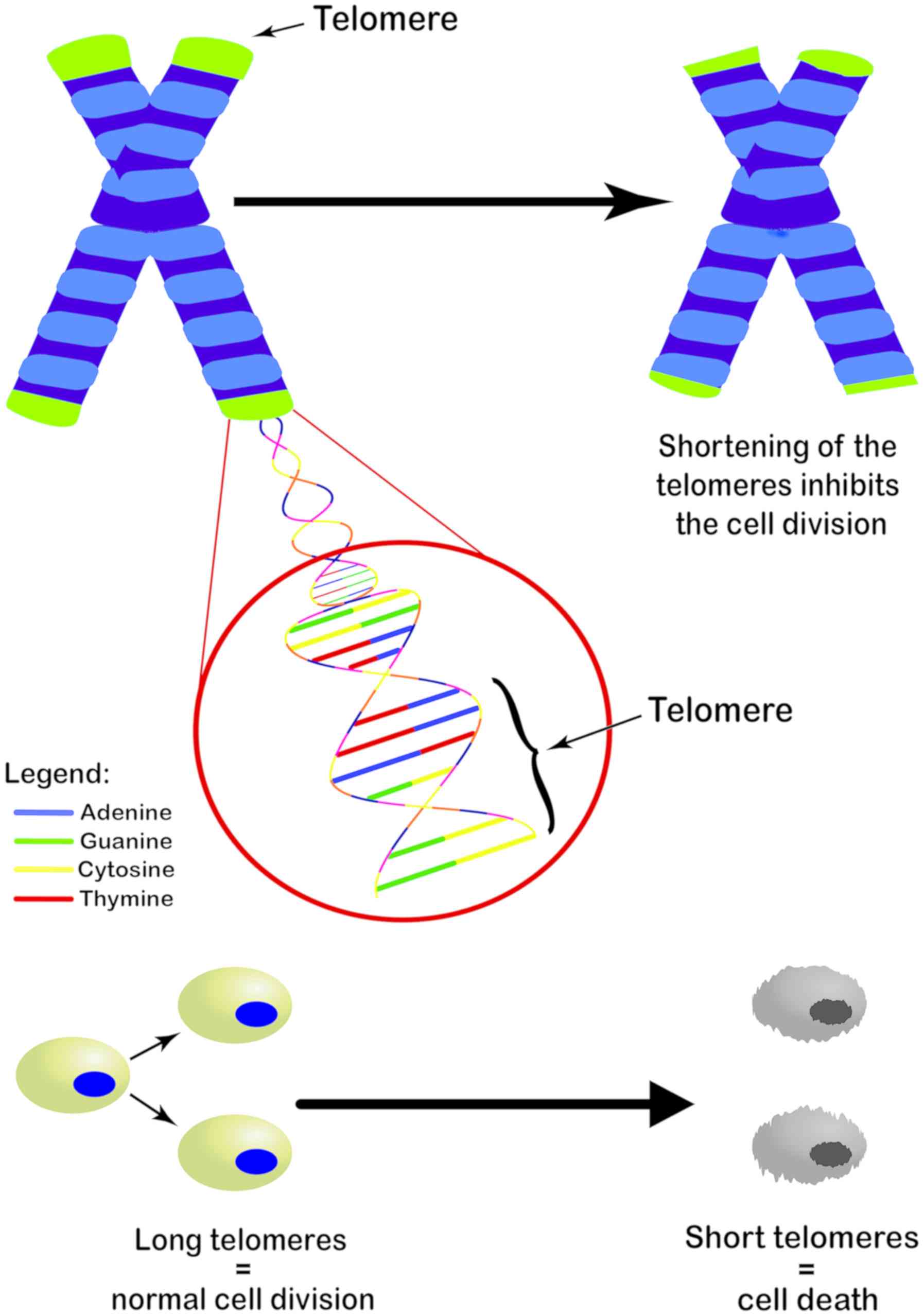
the same individual. Biological aging can be calculated by telomere
length (TL) and DNA methylation levels (6). Telomeres are the end parts of linear
chromosomes and consist of many tandem repeats of 5′-TTAGGG-3′.
When cells divide, they are unable to replicate approximately 50
base pairs to the end of each chromosome. This leads to the
progressive shortening with each round of cell division (7) (Fig.
1), resulting in cell proliferation arrest and cell senescence.
This mechanism is the leading cause of aging and age-related
chronic diseases. In 2009, Blackburn, Greider and Szostak received
the Nobel Prize for discovering the protective role of telomeres
and the enzyme telomerase on chromosomes. These highly significant
discoveries opened the way for researchers to further explore the
role of telomere shortening in aging. By the construction of a
biological age equation, the measuring of TL has come to be a
calculating tool for the biological age of the body, including, but
not limited to gonadal age (8,9)
thus more accurately predicting the fertility status. Recently, a
database was published named 'BIOTEL version 2.4', Telomere Length
Database Project (TLDP), a semi-automated worksheet that calculates
a wide range of TL statistics and it is a useful tool with
applications in research on telomere biology, and in biological age
estimation (10).
Both genetic signature and external factors can
decrease the gonadal pool and can thus subsequently increase male
and female infertility, despite a favorable chronological age
(11). Specifically, such
external factors include human exposure to chemicals and pesticides
used in agriculture and industry that act as endocrine disruptors
(EDs) (11-15), dysregulating normal reproductive
system functions (16-18), as well as drug abuse (19).
It has been claimed that epigenetic factors, such as
nutrition, exercise and tobacco can also affect the rate at which
telomeres shorten and the risk of developing chronic diseases.
Notably, nutrition supplements have been shown to benefit the
length of short telomeres (20).
A method of delaying the aging process has been
proposed to be telomerase activation (21). Telomerase activation by natural
molecules has been suggested to be potent in anti-aging and the
treatment of related diseases. Telomerase activation can be
achieved through natural molecules, synthetic molecules, and
genetic manipulation and intervention. Recently, supplements and
natural extracts were tested for their capacity to enhance
telomerase activity (TA) in human peripheral blood mononuclear
cells. It was demonstrated that formulations containing
Astragalus extract activate telomerase to higher levels than
the reported levels, indicating that the synergistic effects of
nutrients and natural compounds can activate telomerase and can
produce more potent formulations (22). One way in which telomerase can
repair short or dysfunctional telomeres is by the addition of
nucleotides at their ends, further stabilizing them. Indeed, TL and
TA have been proposed as biomarkers of aging and studies have
investigated their implications in other chronic diseases (23,24).
In an effort to identify the causes of age-related
infertility, researchers have managed to link the aging process to
genomic and epigenomic alterations, DNA damage and oxidative stress
(25). However, several human
studies, which will be further reviewed below, have attempted to
investigate the role of telomeres in infertility on a wider
spectrum, by indicating the relative TL in individuals with varying
types of infertility.
The aim of this review was to elucidate the
potential positive or negative association between relative TL and
different factors of infertility, namely male factor, tubal factor,
polycystic ovary syndrome (PCOS), ovarian reserve, endometriosis,
anovulation, cancer related infertility and idiopathic
infertility.
The Medline [Ovid MEDLINE(R) In-Process & Other
Non- Indexed Citations and Ovid MEDLINE(R) (1946 to February,
2018)] electronic database was searched to detect all publications
focusing on the association of TL with human female and male
infertility. The comprehensive literature was searched in
accordance with the Preferred Reporting Items for Systematic
Reviews and Meta-analyses (PRISMA) statement (26), using the following literature
search strategies: i) Telomeres OR TL AND fertility OR infertility;
ii) Male Factor: telomere AND (idiopathic male infertility OR
oligospermia OR azoospermia OR hypospermia OR teratospermia OR
aspermia OR asthenozoospermia OR necrozoospermia OR leukospermia);
iii) Tubal Factor: Telomere AND (hydrosalpinx OR fallopian tube OR
tubal factor); iv) PCOS: Telomere AND PCOS; v) Ovarian Reserve:
Telomere AND (ovary OR ovarian reserve); vi) Endometriosis:
Telomere AND endometriosis; vii) Anovulation: Telomere AND
(anovulation OR ovulation); viii) Unknown: Telomere AND
(unexplained infertility OR idiopathic infertility).
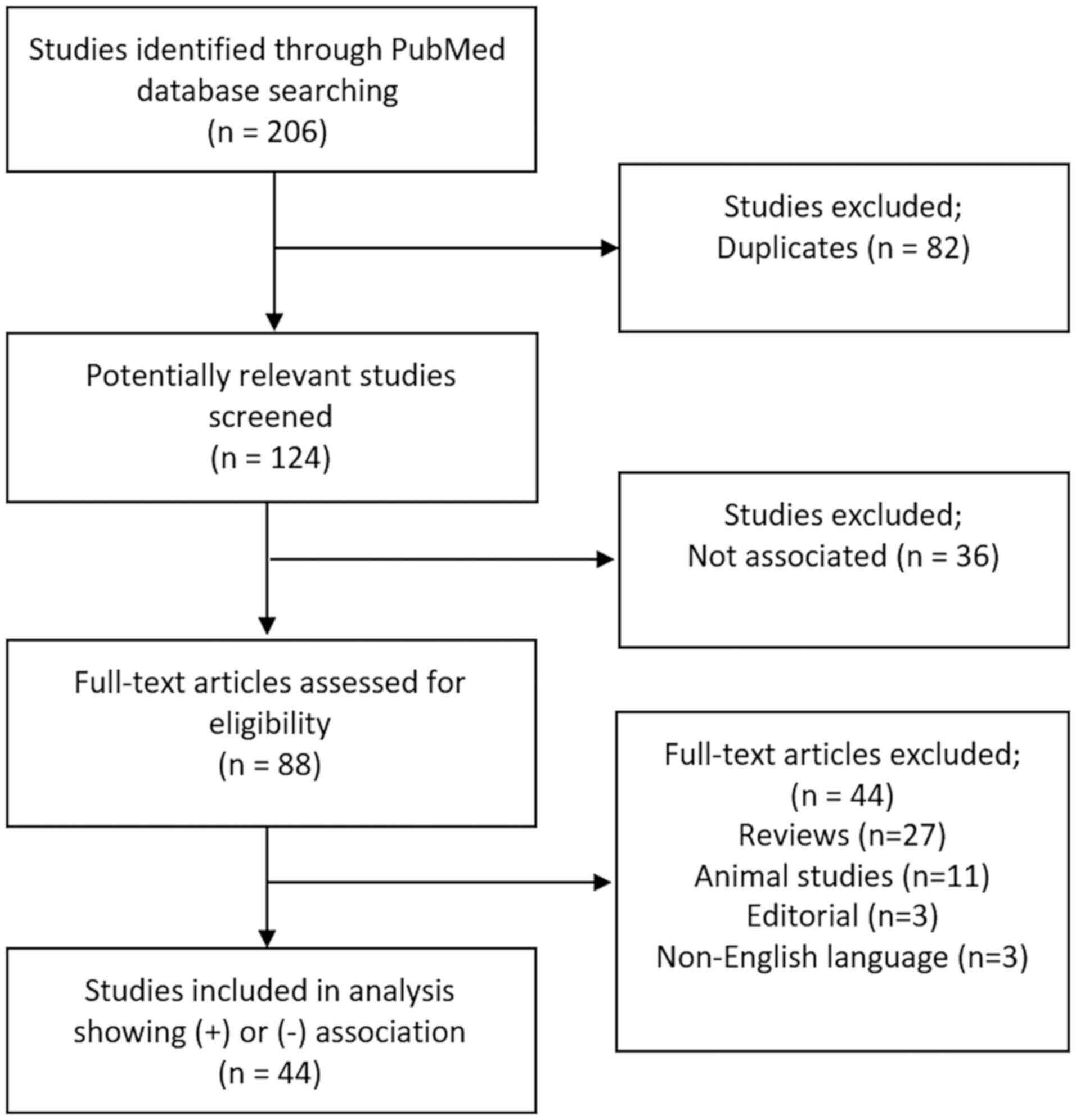
After excluding duplicates, titles and abstracts
were screened for relevance, marked by a link between infertility
cause and relative TL (longer or shorter). Two review authors (E.
Vasilopoulos and C. Kalliora) independently scanned the title or
the abstract content, or both, of every record retrieved to
determine which studies should be assessed further and extracted
all data. Citations in conference abstract form, review articles,
animal studies, editorials, and non-English language articles were
excluded.
For each of these studies, it was determined whether
a positive or negative association between the infertility factor
they are focused on and TL is shown. This specific infertility
factor was noted for each study, along with other important
information (sex, relative TL and the method of measurement,
population size, population age, population region, sample type and
conclusion). The studies were further organized by infertility
factor, allowing collective conclusions to be drawn. A total of 45
studies showing either a positive or negative association between
male or female infertility and TL were included in the present
review (Fig. 2).
2. Telomere length and female
infertility
It is known that shorter telomeres are associated
with a range of chronic diseases, such as cardiovascular disease
(CAD), stroke, cancer, arthritis, osteoporosis, diabetes type 2,
hypertension, mental diseases, chronic obstructive pulmonary
disease and dementia (27-31).
Apart from natural, chronological aging, telomere shortening can be
influenced by physical activity, body mass index (BMI), hormone
replacement therapy, smoking, chronic inflammation, oxidative
stress, dietary antioxidants and vitamins. Previous studies have
found that women following a healthy lifestyle have longer
telomeres (32).
A fundamental factor for the achievement of early
pregnancy is the successful implantation of the embryo. A window of
implantation is in the mid-secretory phase of the menstrual cycle,
which involves the dominant action of progesterone with maximum
cell differentiation. This period is also associated with the
lowest endometrial telomerase activity (TA) and the shortest mean
TL, suggesting a requirement of a low endometrial TA for the
establishment of an early pregnancy (33,34). This suppression of TA in the
endometrium of fertile women has been proposed as a necessary
process in order to allow endometrial cells to undergo
differentiation with cellular apoptosis/senescence required to make
space for the invading embryo (33-35). The underlying mechanisms for this
regulation and differential (dys)regulation are still under
research.
Several studies have thoroughly investigated the
association of female infertility with TL. The majority of studies
exploring the association between female infertility factors and TL
are outlined in Table I and have
revealed a positive association, while only 3 have revealed
negative association (36-38).
Of the 21 positively associated studies, 11 studies demonstrated a
shorter TL, while 10 demonstrated a longer TL. Specifically, a
positive correlation was observed between a short TL and oocyte
maturation, parity, a disrupted gap-junctional inter-cellular
communication (GJIC) in stromal cells and oocytes of poor quality
(39-42). Barha et al measured TL in a
group of 75 Kaqchikel Mayan women over a 13-year study period. They
demonstrated that women with fewer children exhibited shorter TLs
than those who had more children. In the same study, Barha et
al found that estradiol, the levels of which increase during
pregnancy and is a potent antioxidant, protects TL (43). Czamanski-Cohen et al
measured TL in lymphocytes of peripheral blood in women undergoing
in vitro fertilization (IVF) techniques due to infertility;
women undergoing IVF were found to have statistically significant
shorter telomeres compared to the healthy controls in various
phases of the menstrual cycle (44). In addition, lifestyle factors,
such as employment and the work schedule are related to TL in women
(45). Moreover, estradiol
increases TA, maintains TL, and has been related to pregnancy
complications (46).
 | Table IStudies associating telomere length
with female infertility. |
Table I
Studies associating telomere length
with female infertility.
|
Authors/(Refs.) | Sex | Association with
infertility | Relative TL | Method | Population no. | Population age
(years) | Population
country | Sample type | Infertility
factor | Conclusions |
|---|
| Hapangama et
al, 2008 (51) | F | + | Ln | RT-PCR | -(n=29),
endo
-(n=27), non-endo | 18-46
25-44 | UK | Endom Bx | Endo | • TL is longer in
endometria of F with endo during the implantation window. Tissue TL
and LTL are not correlated
• LTL is (+) correlated with circulating estradiol |
| Butts et al,
2009 (52) | F | + | Sh | RT-PCR | -(n=12)
OOI
-(n=42), CNTL (IVF) | 30-37
23-37 | US | G | DOR | • GTL shortening
and diminished TA are associated with occult ovarian
insufficiency |
| Hapangama et
al, 2010 (35) | F | + | Ln | RT-PCR | -(n=38),
endo
-(n=35), non-endo | 18-45
25-38 | UK | Endom Bx | Endo | • (+) correlation
between endometrial TL and both glandular and stromal expression of
nucleolin |
| Hanna et al,
2009 (63) | F | + | Ln | RT-PCR | -(n=34),
POF
-(n=95), SAB history
-(n=108), HC
-(n=46), fert | 21-50
24-45
17-55
37-54 | CA | L | POF | • F with POF have
longer LTL |
| Kuhn et al,
2010 (55) | F | + | Sh | Q-FISH | -(n=22),
STIC
-(n=134) HGSC | N/A | US | Tubal epithelial
cells | TFI/Ca | • Support of the
proposal that STICs are precursors of HGSC potentially an early
event in ovarian HGSC |
| Treff et al,
2011 (53) | F | + | Sh | RT-PCR | -(n=21) | 32-42 | US | Oocytes (polar
bodies) | DOR | • Lower telomere
DNA in aneuploid polar bodies compared to euploid polar bodies
(-3.04-fold, P = 0.016)
• Oocytes with telomere DNA deficiency are prone to aneuploidy
development during meiosis |
| Kuhn et al,
2011 (64) | F | + | Ln | Q-FISH | -(n=219), ovarian
Ca
-(n=106), HGSC
-(n=26), LGSC
-(n=56), OCCC
-(n=31), LGEC | 23-88 | US, TW | SC | Ca/DOR | • TL significantly
differs by histologic type in ovarian Ca
• OCCC has longer mean relative TL compared with the other
histologic types |
| Valentijn et
al, 2013 (62) | F | + | Ln | RT-PCR | -(n=60),
pre-menopausal, normal
-(n=15), post-menopausal
-(n=20), endo | 23-49
46-78
18-49 | UK | Endom cells | Endo | • Showed
significantly higher expression of TA in F with endo
• Longer mean TLs in F with endo |
| Turner and
Hartshorne, 2013 (39) | M/F | + | Sh | Q-FISH | -(n=45),
M
-(n=32), F | 21-49 25-42 | UK | Sperm (semen),
oocytes | C | • STL not crucial
for M fert
• TL reset in embryo after fertilization
• TL shortening during human oocyte maturation
• Oocyte-induced sperm telomere DNA modification, via a
recombination-based TL increase towards the blastocyst stage |
| Cheng et al,
2013 (42) | F | + | Sh | RT-PCR | -(n=45),
<38yo
-(n=35), ≥38yo | 29.2+3.4
41.4+1.9 | TW | Oocyte- cumulus
complex | C | • Mature oocytes
have longer TL than immature oocytes
• Good-quality embryos have longer TL than poor-quality embryos
(cut-off value of the T/S ratio on embryonic Day 3 was 4.235) |
| Dracxler et
al, 2014 (91) | F | + | Ln | q-PCR | -(n=86),
endo
-(n=21), HC | 33
36 | BZ | Lymphoc | Endo | • Lymphoc. TL is
significantly longer in F with endo
• High variety in lymphocyte telomere content in F with endo |
| Li et al,
2014 (58) | F | + | Sh | RT-PCR | -(n=698),
PCOS
-(n=611), HC |
26.42±0.19
28.77±0.27 | CN | L | PCOS | • LTL is shorter in
F with PCOS
• LTL is not associated with biochemical traits in F with
PCOS
• (−) correlation between healthy CNTL LTL and serum DHEAS |
| Yu et al,
2014 (41) | F | + | Sh | RT-PCR | (n=6), no endo/PID)
for laparoscopic tubal ligation | N/A | US | ET (Isolation of
eMSC lines) | C | • Disrupted GJIC in
endometrial SC reduced the mean TL by 45%. Disrupted GJIC causes
ED, which is associated with abnormal uterine bleeding, failed
embryonic implantation, infert, or endom Ca |
| Valentijn et
al, 2015 (62) | F | + | Ln | RT-PCR, Q-FISH | -(n=70), fert
(blood samples)
-(n=74), HC (endom Bx)
-(n=10), endo (ectopic and eutopic endom samples)
-(n=8), HC in mid-secretory phase | 18-46
23-45
24-47
26-45 | UK | L, ET | Endo | • Endom TL is
longer than donor-matched LTL andis (−) correlated with serum
progesterone levels
• Short TL is observed in proliferating EECs in vivo and
in vitro
• Ectopic endom lesions have a higher epithelial telomeric signal
than that in the uterine (eutopic) endom of the same
individuals |
| Czamanski-Cohen
et al, 2015 (44) | F | + | Sh | Q-PCR | -(n=20), IVF
Tx
-(n=10), HC | 29.3±4.3
28.6±3.4 | IL | L | Infert | • Patients seeking
IVF treatment exhibited shorter TL than healthy controls. |
| Pedroso et
al, 2015 (38) | F | − | N/A | RT-PCR | -(n=150),
PCOS
-(n=124), HC | 29.36±5.18 | BZ | L | PCOS | • No difference in
LTL between PCOS and healthy |
| Miranda- Furtado
et al, 2016 (37) | F | − | N/A | RT-PCR | -(n=45),
PCOS
-(n=52), HC | 18-37 | BZ | L | PCOS | • No difference in
telomere content between CNTL and PCOS
• TL shortening after progressive resistance training
intervention |
| Li et al,
2017 (59) | F | + | Sh | RT-PCR | -(n=65), PCOS
(IVF)
-(n=98), non-PCOS (IVF) | 30.3±4.3
31.7±3.8 | CN | L, G | PCOS | • F with PCOS have
shorter GTL
• F with lower TA levels and shorter TL have an earlier onset of
infert symptoms |
| Wei et al,
2017 (56) | F | + | Ln | RT-PCR | -75 F,
PCOS
-81 F, non-PCOS (IVF) |
28.36±2.55
28.09±2.26 | CN | L, G | PCOS | • No difference in
LTL between PCOS and non-PCOS
• GTL is longer in women with PCOS |
| Wang et al,
2017 (57) | F | + | Ln | RT-PCR | -(n=40),
PCOS
-(n=35), HC |
24.78±3.54
26.33±2.15 | CN | L | PCOS | • LTL is
significantly longer in F with PCOS
• (+) correlation between LTL and testosterone levels |
| Kalyan et
al, 2017 (36) | F | - | N/A | RT-PCR | -(n=25),
PCOS
-(n=41), HC | 40.3±3.4
42.5±4.2 | CA | L | PCOS | • No difference in
LTL between PCOS and CNTL |
| Sofiyeva et
al, 2017 (61) | F | + | Ln | (Telo TAGGG) PCR
ELISA | -(n=14),
IE
-(n=17), FE
-(n=16), HC |
30.42±1.36
40.82±1.36
47.9±0.96 | TR | Eutopic endom
cystic wall/ovarian cortex, venous blood | Endome-
triosis | • High TA was found
in the secretory phase of infertile endo women compared to that of
healthy patients |
| Xu et al,
2017 (54) | F | + | Sh | RT-PCR | -(n=120), POI
(IVF)
-(n=279), HC (IVF) |
32.95±4.27
29.98±4.28 | CN | L, G | Premature ovarian
insufficiency | • Shorter LTL and
GTL in women with POI
• LTL decreases 0.067 T/S every year from age 21 up till 39
years
• GTL decreases 0.089 T/S every year from age 23 to 39
years
• Biochemical POI is associated with 0.69 T/S LTL reductions and
0.98T/S GTL reductions |
| Pollack et
al, 2018 (40) | F | + | Sh | RT-PCR | -(n=1505),
parous
-(n=444), nulliparous | 20-44 | US | L | Combination | • Leukocyte T/S
ratio 4.2% (95% CI: 0.9, 7.3)
• TLparous < TLnulliparous
• Parity: 116 fewer base pairs (95% CI: 26, 204) |
However, a late maternal age was reported in
centenarians, supporting that a maternal age over 33 years is a
marker of slow aging, predicting a longer life expectancy (47,48). Fagan et al and others have
shown that the genetic basis between reproductive age, longevity
and biological aging is strongly associated with TL (49,50).
In a nationally representative, cross-sectional
study which included 1,954 women of reproductive age from the
National Health and Nutrition Examination Survey, leukocyte TL
(LTL) was measured by polymerase chain reaction (PCR). It was found
that the women who had at least one live birth versus those that
had biochemical pregnancies, spontaneous abortions, still births,
or other pregnancy complications, had a shorter TL associated with
accelerated cellular aging. The early decline observed in the
reproductive potential of women was suggested to be associated with
TL, which declines earlier in the reproductive system than it does
in somatic cells (40).
According to Hapangama et al, endometrial
telomerase exhibits specific expression patterns in various types
of reproductive failure. Their study included endometrial biopsies
during the window of implantation from women with idiopathic
recurrent loss of empty gestational sacs, fetal loss after cardiac
activity identification and recurrent implantation failure. No
significant differences were observed in the mean TL between the
groups. However, telomerase immunostaining was found to be elevated
in endometrial samples from women with reproductive failure of all
types, particularly those with recurrent implantation failure
(51).
Additionally, Butts et al and others have
revealed a short granulosa cell TL (GTL) and lower telomere DNA in
patients with occult ovarian insufficiency (OOI) (52,53). Premature ovarian insufficiency
(POI) has also been linked to both a shorter LTL and GTL (54). A short TL has been found in tubal
epithelial cells in a study that supported the proposal that serous
tubal intraepithelial carcinomas (STICs) are precursors of
high-grade serous carcinoma (HGSC), potentially an early event in
ovarian high-grade serous carcinogenesis (55). Another factor commonly leading to
infertility is polycystic ovary syndrome (PCOS). Several studies
have focused on the association between TL and PCOS. However,
studies differ in the type of association and correlation. Of the 7
studies reviewed, 3 studies showed a negative association between
LTL and PCOS patients (35-37) while Wei et al demonstrated
significantly longer GTL but a negative association between LTL and
PCOS patients (56). Further, 1
study revealed significantly longer LTL in patients with PCOS
(57). Contrastingly, Li et
al demonstrated that LTL was shorter in women with PCOS and
that a short LTL increased the risk of disease (58). A different group demonstrated
shorter GTL and an earlier onset of infertility symptoms in women
with lower TA levels and PCOS (59).
In addition, a longer TL is widely observed in women
with endometriosis. The tissue-specific regulation of TL in
patients with endometriosis has been suggested, as endometrial TL
is significantly longer, but not is not associated with LTL, which
correlates with circulating estradiol levels (33,60). Additionally, endometrial TL has
been shown to exhibit a positive correlation with the glandular and
stromal expression of nucleolin, involved in the exponential growth
of eukaryotic cells (35). A high
telomerase activity was seen in the secretory phase of infertile
women with endometriosis when compared to healthy women or fertile
women with endometriosis (61).
Similarly, significantly higher expression of telomerase activity,
longer mean TLs, lower expression of genes for steroid receptors
was found in ectopic endometriosis lesions (62). Furthermore, Hanna et al
demonstrated a long LTL in women with premature ovarian failure
(POF) (63). Lastly, telomere
lengths are varied with specific histologic types in ovarian
carcinomas. Specifically, short TLs are observed in tubal
epithelial cells of serus tubal intraepithelial carcinoma (STIC),
an early event in ovarian high-grade serous carcinogenesis
(55). However, stromal cells of
clear cell carcinoma have longer mean relative TL compared to other
histologic types (64). An
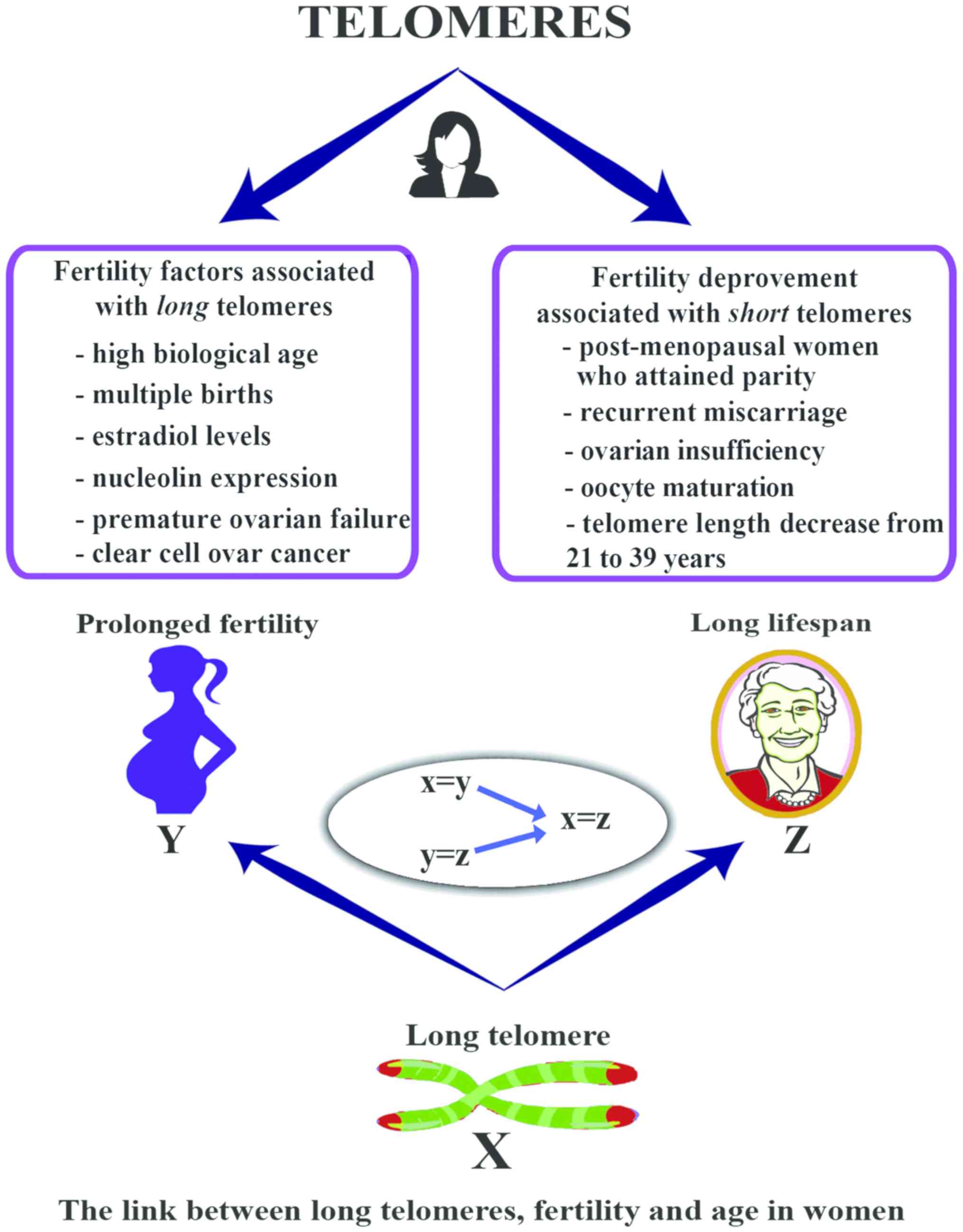
overview of the link between TL, fertility and lifespan in women is
presented in Fig. 3.
The female reproductive system poses certain
paradoxes. Although somatic tissues of the uterus remain
well-functioning and receptive throughout the reproductive years,
oocytes exhibit precocious profound aging. Meiotic dysfunction
increasingly afflicts women as they age, resulting in infertility,
miscarriage, and offspring with congenital abnormalities (65). For normal reproduction, the
biological state of the ovum is of high importance. Apart from TL
in lymphocytes, another factor that appears to be associated with
fertility is the TL of oocytes.
According to the study by Kalmbach et al,
reproductive aging involves declines in both oocyte number and
developmental capacity. In women, the effects of reproductive aging
on oocyte quality are largely explained by telomere shortening
(66). Also paradoxical, is that
despite the reduction of telomeres with aging, their length resets
across generations via the novel mechanism involving recombination
and sister chromatid exchange in the early cell cycles, and with
telomerase activity after the blastocyst stage (67).
3. Telomere length and male infertility
A positive association between male infertility
factors and TL was observed in 19 studies (68-86) and a negative association in 2
studies (39,87) (Table II). In total, 15 of the
positively associated studies detected a sperm TL (STL) association
(68-70,72,74-84), while 3 studies detected an LTL
association (73,85,86), and one study detected both STL and
LTL (71). In several studies,
shorter STL was reported in association to infertility (72,76,78,80) while Vecoli et al reported
an increase in STL in areas of high environmental exposure
(77). Although Turner and
Hartshorne found an association between female fertility and STL,
they did not find a significant association for male fertility
(39). Further, a study focusing
on paternal age at the time of conception reported no significant
change in TL between younger and older males (87). On the other hand, paternal age at
birth has been shown to be positively correlated with LTL (73) and with offspring LTL in the
nurses' health study (8). An
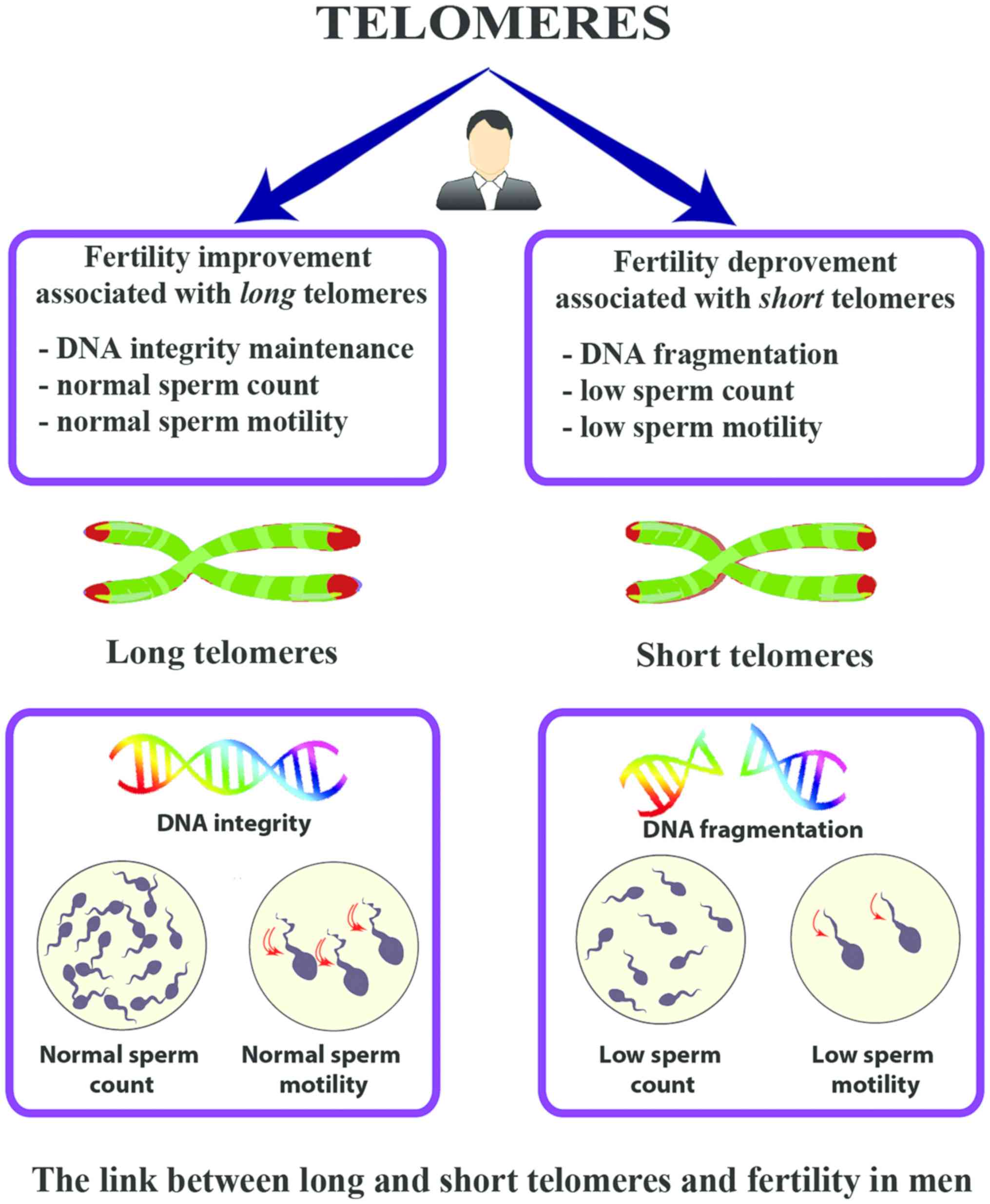
overview of the link between TL, and fertility in men is presented
in Fig. 4.
| Table IIStudies associating telomere length
with male infertility. |
Table II
Studies associating telomere length
with male infertility.
|
Authors/(Refs.) | Sex | Association with
infert | Relative TL | Method | Population no. | Population age
(years) | Population
country | Sample type | Infertility
factor | Conclusions |
|---|
| Baird et al,
2006 (75) | M | + | Sh | RT-PCR, (TRF)
(STELA) | (n=54), 10 samples
analyzed | ~35 | UK | Sperm (semen) | C | • Human telomere
truncation events occur with frequency of 3.6%, while 96.4% of
telomeres at any one chromosomal end are normal
• Telomere truncation in the male germline may contribute to the
observed levels of aneuploidy (up to 1.55%) in human sperm
range
• Telomere truncation can limit replicative potential, and the
subsequent loss of end capping can result in genomic
instability |
| Moskovtsev et
al, 2010 (74) | M | + | Disrupted T-T
ixns | Q-F1SH | -(n=20), DNA
damaged sperm | N/A | CA | Sperm (semen) | C | • Sperm DNA damage
is correlated with disruption of the normal T-T interactions
leading to possible loss of the looped chromosome
configuration |
| Prescott et
al, 2012 (73) | M | + | Ln | RT-PCR | N/A | N/A | US | Genomic DNA | Age | • + correlation for
participant's relative LTL with paternal age at birth
• Inverse relationship between maternal age at birth and relative
LTL |
| Turner and
Hartshorne, 2013 (39) | M/F | | NIA, M | Q-FISH Sh, F | -(n=45),
M
-(n=32), F | 21-49
25-42 | UK | Sperm (semen),
oocytes | C | • STL not crucial
for male fert
• TL reset in embryo after fertilization
• TL shortening during human oocyte maturation
• Oocyte-induced sperm telomere DNA modification, via a
recombination-based TL increase towards the blastocyst stage |
| Ferlin et
al, 2013 (71) | M | + | Sh | RT-PCR | -(n=20),
oligozoosp | 18-19 | IT | Sperm (DGC),
leukocytes | Oligo zoosp | • + correlation
between STL and LTL
• Shorter STL in oligozoosp M
• + correlation between STL and total SC
• + correlation between STL and paternal age at conception |
| Thilagavathi et
al, 2013 (72) | M | + | Sh | RT-PCR | -(n=32), idiopathic
infert
-(n=25), fert | N/A | IN | Sperm (semen) | I MI | • Infert men had
shorter STL
• STL is not correlated with semen parameters |
| J0rgensen et
al, 2013 (87) | M | − | N/A | Q-FISH | -(n=6),
younger
-(n=6), older | 31-40
43-60 | DK | Testicular Bx | Age | • There is no
difference in TL during spermatogenesis between older and younger
men |
| Yan et al,
2014 (70) | M | + | Sh | RT-PCR | -(n=137), infert,
non-OA
-(n=125), infert, oligozoosp
-(n=318), infert, normal SC | 24-42 | CN | Sperm (semen) | Azoosp | • Genetic variants
such as polymorphisms in telomere- associated pathway genes affect
TL and chromosomal stability
• The polymorphism TERT rs2736100 was associated with male infert
risk (adjusted odds ratio (OR)=0.66, 95% Cl: 0.47-0.92;
Ptrend=0.011)
• The polymorphism TEP1 rsl713449 was positively associated with
risk of male infert (adjusted OR=1.39, 95% Cl: 1.20-1.62;
Ptrend<0.001) |
| Reigh-Viader et
al, 2014 (69) | M | + | Sh | TERRA and
telomerase distribution | -(n=4), NO
deficiency of spermatozoa due to unknown causes, seeking fert
treatment
-(n=l), fert, undergoing vasectomy at the time of tissue
retrieval
-CNTL, HeLa cells with known TL | | ES | Testicular Bx | Azoosp | • Telomere
homeostasis is impaired in infert patients, shown by a decrease in
TERRA levels and an alteration of the TERRA-protein component of
telomerase telomeric association in primary
spermatocytes.
• Telomere structure and homeostasis in germ cells is compromised
in infert individuals |
| Yang et al,
2015 (68) | M | + | Sh | RT-PCR | -(n=418), couples,
idiopathic infert (IVF) | 30.3±4.0 | CN | Sperm (DGC) | C | • No significant
correlation between TL and paternal age at the time of conception
(rp=0.18)
• + correlation between STL and total SC
• + correlation between STL and paternal/maternal ages at
conception
• + correlation between STL and embryo quality |
| Yang et al,
2015 (84) | M | + | Sh | RT-PCR | -105 M undergoing
IVF | 31.2±6.1 | CN | Sperm (DGC) | Oligo zoosp | • + correlated
between STL and SC
• Longer TL sperm selected for ART Tx |
| Liu et al,
2015 (82) | M | + | Sh | RT-PCR | -(n=126),
idiopathic infert
-(n=138), fert | 23-57 | CN | Sperm (semen) | SM/SC | • The relative
sperm mean TL of infertile men was significantly shorter than the
relative TL of fertile men (2.894±0.115 vs. 4.016±0.603, P=5.097×10
5)
• TL is associated with SC and SM |
| Antunes et
al, 2015 (83) | M | + | Lg | SCT-qPCR | -(n=10), IVF | 32-47 | US | Sperm (SU) | Age | • Longer and more
variable STL associated with older age |
| Rocca et al,
2016 (81) | M | + | Sh | RT-PCR | -(n=100),
normozoosp | 34.0±8.6 | IT | Sperm (DGC) | SM | • Positive
association between STL and progressive motility, vitality, and
protamination in normozoosp men
• Shorter STL is associated with lower SM |
| Cariati et
al, 2016 (79) | M | + | Sh | RT-PCR | -(n=19),
oligozoosp
-(n=54), normozoosp | 39.4±5.5 | IT | Sperm (DGC) | Oligo zoosp | • STL is negatively
correlated with sperm diploidy. but positively correlated with
SC
• Oligozoosp men have shorter STL |
| Mishra et
al, 2016 (80) | M | + | Sh | RT-PCR | -(n=112),
infert
-(n=102), fert |
31.71±4.45
32.22±4.0 | IN | Sperm (semen) | C | • STL is shorter in
infert males
• Oxidative stress reduces STL |
| Biron-Shental et
al, 2017 (76) | M | + | Sh | Q-FISH | -(n=16), sub-fert,
ICSI
-(n=10), fert | 37.4±5.0
36.5±7.0 | IT | Sperm (semen) | C | • Shorter STL in
sub-fertile men. Sperm from sub-fertile men has a higher percentage
of telomere aggregates and lower TERT expressions |
| Vecoli et
al, 2017 (77) | M | + | Ln | RT-PCR | -(n=112),
normosp
-(n=112), residents of unexposed areas | 18-42 | IT | Sperm (semen) | C | • Young male
residents in areas with high environmental exposure had a
significant increase in TL of their sperm |
| Lafuente et
al, 2017 (78) | M | + | Sh | Q-FISH | (n=30) | N/A | ES | Sperm (pre/post DGC
and SU) | C | • Exposure of sperm
to increasing concentrations of H2O2 is
associated with telomere shortening
• STL of male partners of couples with primary infertility is lower
than that of male partners of fertile couples
• STL is the same pre/post sperm selection (either DGC and
SU)
• STL is the same in DGC and SU-selected sperm with respect to
unselected sperm
• STL is negatively correlated with SM and sperm concentration |
| Heidary et
al, 2018 (86) | M | + | Sh | RT-PCR | -(n=30) idiopathic
NOA -(n=30), HC, fert | 35.4±4.52 | IR | L | Azoosp | • Association
between LTL shortening in a population of Iranian infert men
affected by idiopathic azoosp |
| Yang et al,
2018 (85) | Male | + | Sh | RT-PCR | -(n=270),
normosp
-(n=247), OA
-(n=349), non-OA | 25-38 | CN | L | Azoosp | • Significantly
shorter relative LTL in men with NOA compared to those with OA or
to the normozoosp controls (odds ratio [OR] 0.81,95% [Cl] 0.64-0.98
vs. OR0.92,95% Cl 0.70-1.24 vs. OR 0.99, 95% Cl 0.83-1.22),
respectively)
• Shorter telomeres are significantly associated with higher risk
for NOA
• Shorter LTL is strongly associated with NOA and defective
spermatogenesis |
Antunes et al reported a longer and more
variable STL with an older age (83), and Mishra et al reported
the disruption of normal telomere interactions, leading to the loss
of the looped chromosomal configuration (80). In the latter study on 112
infertile men, seminal ROS and 8-isoprostane levels were reported
to be increased compared to the controls, demonstrating that the
infertile men experienced some form of oxidative stress. When TL
was measured using reverse transcription-polymerase chain reaction
(RT-PCR) and correlated with reactive oxygen species (ROS) levels,
it was suggested that mild oxidative stress caused the lengthening
of telomeres, whereas in severe oxidative stress, TL was shorter in
comparison to TL when normal ROS levels. Therefore, mild oxidative
stress and the progression of meiosis in infertile men is
beneficial to TL as it correlates with their increased lengths,
whereas severe oxidative stress has harmful effects (80). In a previous study, in sperm
samples with high DNA damage based on the DNA fragmentation index
(DFI) that underwent fluorescence in situ hybridization
analysis (FISH) there was an increased number of telomere signals
compared to those with a low DFI. This could be due to abnormal
telomere-telomere interactions in the case of DNA damage, evident
by the fact that in low DFI, almost 71% of the samples had normal
telomere distribution, whereas in high DFI, this number dropped to
42% (74).
A high expression of telomerase has been reported in
undifferentiated spermatogonia and the complete depletion of
telomerase produces telomere shortening and the eventual loss of
germ line cells, since undifferentiated spermatogonia reduce
significantly (88). It has also
been shown that telomerase association at telomeres is reduced in
testes cells of patients with idiopathic infertility. Telomere
repeat-containing RNA (TERRA) was also found to be reduced in the
same study, although TL was not affected. However, the study
pointed out that telomere integrity is thought to be affected with
aberrant telomerase association, leading to reduced fertility
(69).
Additionally, men with oligospermia have been found
to have a shorter STL and a positive correlation has been found
between a shorter STL and total sperm count (68,71,79), LTL and paternal age (71), and a positive correlation between
a shorter STL and sperm diploidy has also been found (79). Moreover, samples that were used
for IVF and had abnormal STLs did not produce a successful
pregnancy, whereas samples that had STLs within the normal range
had a 35.7% success rate (79).
Likewise, males with azoospermia exhibited a shortened STL
(69,70), as well as a shorter LTL (85,86).
Baird et al performed terminal restricted
fragment (TRF) length and single TL (STELA) analysis in 54 human
semen samples. The results from STELA on XpYp coincided with
results from the TRF analysis and revealed that only 19% of germ
cells would at any time have a full set of chromosomes with
genome-wide TL and would not be truncated. Truncated telomeres may
result in abnormal synapsis during meiotic cell division or in
aneuploids observed in human sperm which may explain the
miscarriages and abortions observed in couples trying to have a
baby and the reduced fecundity of humans (75). Previous studies have shown that
men with genotypes containing 2 specific genetic variants of
telomerase reverse transcriptase (TERT) and telomerase-associated
protein 1 (TEP1) have the chance of being infertile increased by
100%. In particular, TERT rs2736100 is negatively associated with
male infertility risk, whereas TEP1 rs1713449 is positively
associated with male infertility. Individuals with the TEP1
rs1713449 variant also exhibit an increased DFI. Therefore, these
genetic variations play a role in the risk of male infertility
(70).
In patients with decreased sperm motility, a shorter
STL has been found and has also been associated with a lower sperm
count, vitality and protamination (81,82), but negatively associated with DNA
fragmentation in normozoospermic individuals (81). Abnormal semen quality, according
to the WHO criteria, has been associated with a shorter TL than
that of semen with normal parameters. Lastly, men with idiopathic
male infertility have been shown to have a short STL (72).
STL is significantly affected by environmental
factors, such as pollution. In a study carried out in Southern
Italy, it was found that STL was higher in individuals that resided
in a highly polluted area (77).
The effect of polycyclic aromatic hydrocarbons (PAHs) on STL was
assessed in a large study with 666 participants. The results
revealed that a high concentration of urinary PAH metabolites was
associated with shorter STLs. However, semen quality or sperm
apoptosis did not appear to be influenced by PAHs. Moreover,
benzo(α)pyrene administration also caused the shortening of STL and
reduced telomerase expression in the germline in a dose-dependent
manner (89). Anticancer agents
have also been investigated for their effects on TL. Of the 4
agents tested, 2 alkylating agents, cisplatin and
4-hydroperoxycyclophosphamide resulted in a shorter TL, a reduced
TA, reduced telomere specific fluorescence in FISH experiments, and
the mRNA expression of two components of the telomerase. All of the
above can lead to reduced fertility and developmental issues in
offspring (90).
Another study demonstrated that aberrant
fertilization and embryo cleavage were the result of fertilization
with either one or both of the gametes being telomerase deficient.
In these telomerase-null gametes, a small subset of telomeres was
not present in some metaphase I chromosomes, suggesting that the
absence of telomerase causes telomere shortening and eventually
loss (84). In cases of
idiopathic recurrent pregnancy loss, LTL was measured in suffering
couples and compared to the controls. A statistically significantly
shorter LTL was found in both males and females of the suffering
couples in addition to the present TL decrease observed in males
due to age. A positive correlation between the LTL and sperm DNA
fragmentation index was also found, but it was not statistically
significant (72).
Collectively, a prominent trend can be observed
towards a shorter TL in both sperm and leukocytes, associated with
the male factor in infertility. An overview of the link between TL
and fertility in males is presented in Fig. 4.
4. Conclusions and future perspectives
In light of the increasing life expectancies of
individuals worldwide, age-related diseases, such as infertility
have progressively become a greater medical and social concern. The
standard laboratory markers for age-related diseases or infertility
have been insufficient. TL has long been a marker of cellular
aging. The question posed is whether TL can be used to predict not
only the biological, but also the reproductive age. Multiple
studies have focused on elucidating this question and have
demonstrated that infertility is in fact most often associated with
shorter telomeres. Specifically, a trend associating female
infertility factors, such as PCOS, DOR, ovarian insufficiency and
tubal factor with shorter TLs was observed in oocyte granulosa
cells, endometrial tissue and leukocytes. On the contrary, longer
TLs were observed in endometrial biopsy, the eutopic endometrium,
cyst and lymphocyte tissue in cases of endometriosis. As regards
studies focusing on male infertility factors, including
azoospermia, oligospermia, abnormal sperm motility and idiopathic
male infertility, these demonstrated a shorter TL in either sperm
or leukocyte samples obtained.
Although this study was able to identify major
trends in the association of several factors of infertility and TL,
limitations apply due to the relatively low number of relevant
studies for each infertility factor published to date.
Additionally, TL in a number of established factors of infertility
has not yet been studied. Specifically, no published literature on
oligospermia, hypospermia, teratosperimia, aspermia,
asthenozospermia, necrozospermia, leukospermia and hydrosalpinx was
found. Although this serves as a limitation, it indicates a
knowledge gap where further research could be undertaken.
Regardless, TL is associated with a sufficient amount of
infertility factors, thus not compromising the external validity of
the review.
As regards female infertility factors, ovarian HGSC,
mature oocytes, low-quality embryos, multiform endometrial
dysfunction, premature ovarian insufficiency, DOR, and nulliparity
were all linked to a shorter TL. Conversely, all studies on
endometriosis and TL agreed that TLs are longer. Additionally, one
study noted that clear cell carcinoma stromal cells exhibit longer
TL that other histological types. A noticeable disagreement is
evident in studies exploring the TL of females with PCOS, as TL has
been negatively and positively associated, and further correlated
to both a shorter and longer TL in different studies. Thus,
although for all other above-mentioned infertility factors a clear
conclusion is drawn, for PCOS, the present evidence is
inconclusive. Of the 21 studies reviewed focusing on male
infertility factors, 1 study showed no association between TL in
the sperm of older versus younger males, while 2 studies showed
that older males had a longer relative TL and one study found STL
not crucial for male fertility. Furthermore, the remaining studies
were in agreement, indicating a shorter TL in infertile
patients.
With recent advances in reproductive technology, we
need powerful predictive biomarkers that will improve the clinical
strategy in individuals with infertility. TL can be a marker used
to identify the reproductive capacity. Additionally, premature
senescence can be avoided by the use of drugs preventing telomere
shortening, thereby increasing the reproductive capacity of
patients. However, many questions must first be answered before the
efficient use of such methods in a clinical setting is possible. It
is critical that further studies are conducted to understand the
bases of TL associations with biological aging and reproductive
capacity.
The strength of this review is that, at least to the
best of our knowledge, this is the first collection of the results
from human epidemiological studies exploring the association of
different factors of infertility and TL. Despite the present
limitations, this review may be a useful aid, and the studies
mentioned, may function as building blocks for further research
needed to establish further associations and to determine how this
knowledge can be used in medical applications.
Abbreviations:
|
ART
|
assisted reproductive techniques
|
|
GJIC
|
gap-junctional intercellular
communication
|
|
GTL
|
granulosa cell telomere length
|
|
HGSC
|
high-grade serous carcinoma
|
|
IVF
|
in vitro fertilization
|
|
LTL
|
leukocyte telomere length
|
|
OOI
|
occult ovarian insufficiency
|
|
PCOS
|
polycystic ovary syndrome
|
|
POI
|
primary ovarian insufficiency
|
|
Q-FISH
|
quantitative fluorescent in situ
hybridization
|
|
RT-PCR
|
reverse transcription-polymerase chain
reaction
|
|
STIC
|
serous tubal intraepithelial
carcinoma
|
|
STL
|
sperm telomere length
|
|
TA
|
telomerase activity
|
|
TEP1
|
telomerase-associated protein 1
|
|
TERT
|
telomerase reverse transcriptase
|
|
TL
|
telomere length
|
|
DOR
|
diminished ovarian reserve
|
Acknowledgments
Not applicable.
Funding
This study was funded by Metabolomic Medicine S.A.
and Spin-Off Toxplus S.A. and supported by the Special Research
Account of University of Crete (ELKE nos. 4602, 4920 and 3963).
Availability of data and materials
Not applicable.
Authors' contributions
All the authors (EVasilopoulos, PF, CK, DF, AOD,
EVakonaki, DT, DC, AMP, GG, CM, AM, DAS and AT) contributed to
conceiving and designing the study. EVasilopoulos and CK searched
the literature for inclusion in the study that was then checked and
reviewed by AOD, EV, DT, DC, AMB, GG and CM. EVasilopoulos, PF, CK,
DF, AOD, EV, DT, DC, AMB and GG drafted and wrote the manuscript.
AM, DAS, AT, CM provided advice on the experimental design,
interpreted the results and critically revised the manuscript. AOD,
DC, AMB, DT, EV, GG designed the figures. CK and EV designed the
tables. All authors have read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
DAS is the Editor-in-Chief for the journal, but had
no personal involvement in the reviewing process, or any influence
in terms of adjudicating on the final decision, for this article.
The other authors declare that they have no competing
interests.
References
|
1
|
Aghajanova L, Hoffman J, Mok-Lin E and
Herndon CN: Obstetrics and gynecology residency and fertility
needs. Reprod Sci. 24:428–434. 2017. View Article : Google Scholar
|
|
2
|
Center for Disease Control and Prevention:
Assisted reproductive technology success rates: National summary
and fertility clinic reports. 2016, http://www.cdc.gov/art/pdf/2016-report/ART-2016-National-Summary-Report.pdf.
|
|
3
|
Society for Assisted Reproductive
Technology: National Summary Report. Society for Assisted
Reproductive Technology; 2016, http://www.sartcorsonline.com/rptCSR_PublicMultYear.aspx?reportingYear=2016.
|
|
4
|
Agarwal A, Mulgund A, Hamada A and Chyatte
MR: A unique view on male infertility around the globe. Reprod Biol
Endocrinol. 13:372015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Artini PG, Obino ME, Vergine F,
Sergiampietri C, Papini F and Cela V: Assisted reproductive
technique in women of advanced fertility age. Minerva Ginecol.
70:738–749. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang WG, Zhu SY, Bai XJ, Zhao DL, Jian
SM, Li J, Li ZX, Fu B, Cai GY, Sun XF, et al: Select aging
biomarkers based on telomere length and chronological age to build
a biological age equation. Age (Dordr). 36:96392014. View Article : Google Scholar
|
|
7
|
Pfeiffer V and Lingner J: Replication of
telomeres and the regulation of telomerase. Cold Spring Harb
Perspect Biol. 5:a0104052013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jones MJ, Goodman SJ and Kobor MS: DNA
methylation and healthy human aging. Aging Cell. 14:924–932. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rizvi S, Raza ST and Mahdi F: Telomere
length variations in aging and age-related diseases. Curr Aging
Sci. 7:161–167. 2014. View Article : Google Scholar
|
|
10
|
Tsatsakis A, Tsoukalas D, Fragkiadaki P,
Vakonaki E, Tzatzarakis M, Sarandi E, Nikitovic D, Tsilimidos G and
Alegakis AK: Developing BIOTEL: A semi-automated spreadsheet for
estimating telomere length and biological age. Front Genet.
10:842019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Petrakis D, Vassilopoulou L, Mamoulakis C,
Psycharakis C, Anifantaki A, Sifakis S, Docea AO, Tsiaoussis J,
Makrigiannakis A and Tsatsakis AM: Endocrine disruptors leading to
obesity and related diseases. Int J Environ Res Public Health.
14:E12822017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mehrpour O, Karrari P, Zamani N, Tsatsakis
AM and Abdollahi M: Occupational exposure to pesticides and
consequences on male semen and fertility: A review. Toxicol Lett.
230:146–156. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kalliora C, Mamoulakis C, Vasilopoulos E,
Stamatiades GA, Kalafati L, Barouni R, Karakousi T, Abdollahi M and
Tsatsakis A: Association of pesticide exposure with human
congenital abnormalities. Toxicol Appl Pharmacol. 346:58–75. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sifakis S, Androutsopoulos VP, Tsatsakis
AM and Spandidos DA: Human exposure to endocrine disrupting
chemicals: Effects on the male and female reproductive systems.
Environ Toxicol Pharmacol. 51:56–70. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Katsikantami I, Sifakis S, Tzatzarakis MN,
Vakonaki E, Kalantzi OI, Tsatsakis AM and Rizos AK: A global
assessment of phthalates burden and related links to health
effects. Environ Int. 97:212–236. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yawson Emmanuel O, Obasi KK and Lawal I:
Spermatogenic and spermatotoxic effects of Telfairia occidentalis
(Ugu) aqueous leaves extract in adult male Wistar rats (Rattus
novergicus). Toxicol Rep. 5:954–958. 2018. View Article : Google Scholar :
|
|
17
|
Acosta IB, Junior ASV, E Silva EF, Cardoso
TF, Caldas JS, Jardim RD and Corcini CD: Effects of exposure to
cadmium in sperm cells of zebrafish, Danio rerio. Toxicol Rep.
3:696–700. 2016. View Article : Google Scholar
|
|
18
|
Mello MSC, Delgado IF, Favareto APA, Lopes
CMT, Batista MM, Kempinas WD and Paumgartten FJR: Sexual maturation
and fertility of mice exposed to triphenyltin during prepubertal
and pubertal periods. Toxicol Rep. 2:405–414. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vakonaki E, Tzatzarakis M, Tsiminikaki K,
Nathena D, Fragkiadaki P, Kalliantasi K, Kanaki K, Vaki G, Plaitis
S, Tsoukalas D, et al: Effect of chronic and heavy drug abuse on
biological aging. World Acad J Sci. 1:67–73. 2019.
|
|
20
|
Tsoukalas D, Fragkiadaki P, Docea AO,
Alegakis AK, Sarandi E, Vakonaki E, Salataj E, Kouvidi E, Nikitovic
D, Kovatsi L, et al: Association of nutraceutical supplements with
longer telomere length. Int J Mol Med. 44:218–226. 2019.PubMed/NCBI
|
|
21
|
Shammas MA: Telomeres, lifestyle, cancer,
and aging. Curr Opin Clin Nutr Metab Care. 14:28–34. 2011.
View Article : Google Scholar
|
|
22
|
Valassi E, Crespo I, Santos A and Webb SM:
Clinical consequences of Cushing's syndrome. Pituitary. 15:319–329.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tedone E, Huang E, O'Hara R, Batten K,
Ludlow AT, Lai TP, Arosio B, Mari D, Wright WE and Shay JW:
Telomere length and telomerase activity in T cells are biomarkers
of high-performing centenarians. Aging Cell. 18:e128592019.
View Article : Google Scholar
|
|
24
|
Kordinas V, Ioannidis A and
Chatzipanagiotou S: The telomere/telomerase system in chronic
inflammatory Diseases Cause or effect? Genes (Basel). 7:E602016.
View Article : Google Scholar
|
|
25
|
Chakravarthi BV, Nepal S and Varambally S:
Genomic and epigenomic alterations in cancer. Am J Pathol.
186:1724–1735. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Moher D, Liberati A, Tetzlaff J and Altman
DG; Group P; PRISMA Group: Preferred reporting items for systematic
reviews and meta-analyses: The PRISMA statement. PLoS Med.
6:e10000972009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ma H, Zhou Z, Wei S, Liu Z, Pooley KA,
Dunning AM, Svenson U, Roos G, Hosgood HD III, Shen M, et al:
Shortened telomere length is associated with increased risk of
cancer: A meta-analysis. PLoS One. 6:e204662011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Haycock PC, Heydon EE, Kaptoge S,
Butterworth AS, Thompson A and Willeit P: Leucocyte telomere length
and risk of cardiovascular disease: Systematic review and
meta-analysis. BMJ. 349. pp. g42272014, View Article : Google Scholar
|
|
29
|
Willeit P, Raschenberger J, Heydon EE,
Tsimikas S, Haun M, Mayr A, Weger S, Witztum JL, Butterworth AS,
Willeit J, et al: Leucocyte telomere length and risk of type 2
diabetes mellitus: New prospective cohort study and
literature-based meta-analysis. PLoS One. 9:e1124832014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Aviv A, Kark JD and Susser E: Telomeres,
atherosclerosis, and human longevity: A causal hypothesis.
Epidemiology. 26:295–299. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang J, Rane G, Dai X, Shanmugam MK,
Arfuso F, Samy RP, Lai MK, Kappei D, Kumar AP and Sethi G: Ageing
and the telomere connection: An intimate relationship with
inflammation. Ageing Res Rev. 25:55–69. 2016. View Article : Google Scholar
|
|
32
|
Parks CG, DeRoo LA, Miller DB, McCanlies
EC, Cawthon RM and Sandler DP: Employment and work schedule are
related to telomere length in women. Occup Environ Med. 68:582–589.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Valentijn AJ, Saretzki G, Tempest N,
Critchley HO and Hapangama DK: Human endometrial epithelial
telomerase is important for epithelial proliferation and glandular
formation with potential implications in endometriosis. Hum Reprod.
30:2816–2828. 2015.PubMed/NCBI
|
|
34
|
Williams CD, Boggess JF, LaMarque LR,
Meyer WR, Murray MJ, Fritz MA and Lessey BA: A prospective,
randomized study of endometrial telomerase during the menstrual
cycle. J Clin Endocrinol Metab. 86:3912–3917. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hapangama DK, Turner MA, Drury J,
Heathcote L, Afshar Y, Mavrogianis PA and Fazleabas AT: Aberrant
expression of regulators of cell-fate found in eutopic endometrium
is found in matched ectopic endometrium among women and in a baboon
model of endometriosis. Hum Reprod. 25:2840–2850. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kalyan S, Patel MS, Kingwell E, Côté HCF,
Liu D and Prior JC: Competing factors link to bone health in
polycystic ovary syndrome: Chronic low-grade inflammation takes a
toll. Sci Rep. 7:34322017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Miranda-Furtado CL, Ramos FK, Kogure GS,
Santana-Lemos BA, Ferriani RA, Calado RT and Dos Reis RM: A
nonrandomized trial of progressive resistance training intervention
in women with polycystic ovary syndrome and its implications in
telomere content. Reprod Sci. 23:644–654. 2016. View Article : Google Scholar
|
|
38
|
Pedroso DC, Miranda-Furtado CL, Kogure GS,
Meola J, Okuka M, Silva C, Calado RT, Ferriani RA, Keefe DL and dos
Reis RM: Inflammatory biomarkers and telomere length in women with
polycystic ovary syndrome. Fertil Steril. 103:542–547.e2. 2015.
View Article : Google Scholar
|
|
39
|
Turner S and Hartshorne GM: Telomere
lengths in human pronuclei, oocytes and spermatozoa. Mol Hum
Reprod. 19:510–518. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pollack AZ, Rivers K and Ahrens KA: Parity
associated with telomere length among US reproductive age women.
Hum Reprod. 33:736–744. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yu J, Berga SL, Zou W, Sun HY,
Johnston-MacAnanny E, Yalcinkaya T, Sidell N, Bagchi IC, Bagchi MK
and Taylor RN: Gap junction blockade induces apoptosis in human
endometrial stromal cells. Mol Reprod Dev. 81:666–675. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cheng EH, Chen SU, Lee TH, Pai YP, Huang
LS, Huang CC and Lee MS: Evaluation of telomere length in cumulus
cells as a potential biomarker of oocyte and embryo quality. Hum
Reprod. 28:929–936. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Barha CK, Hanna CW, Salvante KG, Wilson
SL, Robinson WP, Altman RM and Nepomnaschy PA: Number of children
and telomere length in women: A prospective, longitudinal
evaluation. PLoS One. 11:e01464242016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Czamanski-Cohen J, Sarid O, Cwikel J,
Douvdevani A, Levitas E, Lunenfeld E and Har-Vardi I: Cell-free DNA
and telomere length among women undergoing in vitro fertilization
treatment. J Assist Reprod Genet. 32:1697–1703. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Shalev I, Entringer S, Wadhwa PD,
Wolkowitz OM, Puterman E, Lin J and Epel ES: Stress and telomere
biology: A lifespan perspective. Psychoneuroendocrinology.
38:1835–1842. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fragkiadaki P, Tsoukalas D, Fragkiadoulaki
I, Psycharakis C, Nikitovic D, Spandidos DA and Tsatsakis AM:
Telomerase activity in pregnancy complications (Review). Mol Med
Rep. 14:16–21. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Perls TT, Alpert L and Fretts RC:
Middle-aged mothers live longer. Nature. 389:1331997. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sun F, Sebastiani P, Schupf N, Bae H,
Andersen SL, McIntosh A, Abel H, Elo IT and Perls TT: Extended
maternal age at birth of last child and women's longevity in the
Long Life Family Study. Menopause. 22:26–31. 2015. View Article : Google Scholar
|
|
49
|
Fagan E, Sun F, Bae H, Elo I, Andersen SL,
Lee J, Christensen K, Thyagarajan B, Sebastiani P, Perls T, et al
Long Life Family Study: Telomere length is longer in women with
late maternal age. Menopause. 24:497–501. 2017. View Article : Google Scholar :
|
|
50
|
Gray KE, Schiff MA, Fitzpatrick AL, Kimura
M, Aviv A and Starr JR: Leukocyte telomere length and age at
menopause. Epidemiology. 25:139–146. 2014. View Article : Google Scholar :
|
|
51
|
Hapangama DK, Turner MA, Drury JA,
Martin-Ruiz C, Von Zglinicki T, Farquharson RG and Quenby S:
Endometrial telomerase shows specific expression patterns in
different types of reproductive failure. Reprod Biomed Online.
17:416–424. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Butts S, Riethman H, Ratcliffe S, Shaunik
A, Coutifaris C and Barnhart K: Correlation of telomere length and
telomerase activity with occult ovarian insufficiency. J Clin
Endocrinol Metab. 94:4835–4843. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Treff NR, Su J, Taylor D and Scott RT Jr:
Telomere DNA deficiency is associated with development of human
embryonic aneuploidy. PLoS Genet. 7:e10021612011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Xu X, Chen X, Zhang X, Liu Y, Wang Z, Wang
P, Du Y, Qin Y and Chen ZJ: Impaired telomere length and telomerase
activity in peripheral blood leukocytes and granulosa cells in
patients with biochemical primary ovarian insufficiency. Hum
Reprod. 32:201–207. 2017.
|
|
55
|
Kuhn E, Meeker A, Wang TL, Sehdev AS,
Kurman RJ and Shih IeM: Shortened telomeres in serous tubal
intraepithelial carcinoma: An early event in ovarian high-grade
serous carcinogenesis. Am J Surg Pathol. 34:829–836. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wei D, Xie J, Yin B, Hao H, Song X, Liu Q,
Zhang C and Sun Y: Significantly lengthened telomere in granulosa
cells from women with polycystic ovarian syndrome (PCOS). J Assist
Reprod Genet. 34:861–866. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wang C, Shen F, Zhu Y, Fang Y and Lu S:
Telomeric repeat-containing RNA (TERRA) related to polycystic ovary
syndrome (PCOS). Clin Endocrinol (Oxf). 86:552–559. 2017.
View Article : Google Scholar
|
|
58
|
Li Q, Du J, Feng R, Xu Y, Wang H, Sang Q,
Xing Q, Zhao X, Jin L, He L, et al: A possible new mechanism in the
pathophysiology of polycystic ovary syndrome (PCOS): The discovery
that leukocyte telomere length is strongly associated with PCOS. J
Clin Endocrinol Metab. 99:E234–E240. 2014. View Article : Google Scholar
|
|
59
|
Li Y, Deng B, Ouyang N, Yuan P, Zheng L
and Wang W: Telomere length is short in PCOS and oral contraceptive
does not affect the telomerase activity in granulosa cells of
patients with PCOS. J Assist Reprod Genet. 34:849–859. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Hapangama DK, Turner MA, Drury JA, Quenby
S, Saretzki G, Martin-Ruiz C and Von Zglinicki T: Endometriosis is
associated with aberrant endometrial expression of telomerase and
increased telomere length. Hum Reprod. 23:1511–1519. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Sofiyeva N, Ekizoglu S, Gezer A, Yilmaz H,
Kolomuc Gayretli T, Buyru N and Oral E: Does telomerase activity
have an effect on infertility in patients with endometriosis? Eur J
Obstet Gynecol Reprod Biol. 213:116–122. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Valentijn AJ, Palial K, Al-Lamee H,
Tempest N, Drury J, Von Zglinicki T, Saretzki G, Murray P, Gargett
CE and Hapangama DK: SSEA-1 isolates human endometrial basal
glandular epithelial cells: Phenotypic and functional
characterization and implications in the pathogenesis of
endometriosis. Hum Reprod. 28:2695–2708. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hanna CW, Bretherick KL, Gair JL, Fluker
MR, Stephenson MD and Robinson WP: Telomere length and reproductive
aging. Hum Reprod. 24:1206–1211. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kuhn E, Meeker AK, Visvanathan K, Gross
AL, Wang TL, Kurman RJ and Shih IeM: Telomere length in different
histologic types of ovarian carcinoma with emphasis on clear cell
carcinoma. Mod Pathol. 24:1139–1145. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Keefe DL, Liu L and Marquard K: Telomeres
and aging-related meiotic dysfunction in women. Cell Mol Life Sci.
64:139–143. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kalmbach KH, Antunes DM, Kohlrausch F and
Keefe DL: Telomeres and female reproductive aging. Semin Reprod
Med. 33:389–395. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Keefe DL and Liu L: Telomeres and
reproductive aging. Reprod Fertil Dev. 21:10–14. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Yang Q, Zhao F, Dai S, Zhang N, Zhao W,
Bai R and Sun Y: Sperm telomere length is positively associated
with the quality of early embryonic development. Hum Reprod.
30:1876–1881. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Reig-Viader R, Capilla L, Vila-Cejudo M,
Garcia F, Anguita B, Garcia-Caldés M and Ruiz-Herrera A: Telomere
homeostasis is compromised in spermatocytes from patients with
idiopathic infertility. Fertil Steril. 102:728–738.e1. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Yan L, Wu S, Zhang S, Ji G and Gu A:
Genetic variants in telomerase reverse transcriptase (TERT) and
telomerase-associated protein 1 (TEP1) and the risk of male
infertility. Gene. 534:139–143. 2014. View Article : Google Scholar
|
|
71
|
Ferlin A, Rampazzo E, Rocca MS, Keppel S,
Frigo AC, De Rossi A and Foresta C: In young men sperm telomere
length is related to sperm number and parental age. Hum Reprod.
28:3370–3376. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Thilagavathi J, Kumar M, Mishra SS,
Venkatesh S, Kumar R and Dada R: Analysis of sperm telomere length
in men with idiopathic infertility. Arch Gynecol Obstet.
287:803–807. 2013. View Article : Google Scholar
|
|
73
|
Prescott J, Du M, Wong JY, Han J and De
Vivo I: Paternal age at birth is associated with offspring
leukocyte telomere length in the nurses' health study. Hum Reprod.
27:3622–3631. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Moskovtsev SI, Willis J, White J and
Mullen JB: Disruption of telomere-telomere interactions associated
with DNA damage in human spermatozoa. Syst Biol Reprod Med.
56:407–412. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Baird DM, Britt-Compton B, Rowson J, Amso
NN, Gregory L and Kipling D: Telomere instability in the male
germline. Hum Mol Genet. 15:45–51. 2006. View Article : Google Scholar
|
|
76
|
Biron-Shental T, Wiser A, Hershko-Klement
A, Markovitch O, Amiel A and Berkovitch A: Sub-fertile sperm cells
exemplify telomere dysfunction. J Assist Reprod Genet. 35:143–148.
2018. View Article : Google Scholar :
|
|
77
|
Vecoli C, Montano L, Borghini A, Notari T,
Guglielmino A, Mercuri A, Turchi S and Andreassi MG: Effects of
highly polluted environment on sperm telomere length: A Pilot
Study. Int J Mol Sci. 18:E17032017. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Lafuente R, Bosch-Rue E, Ribas-Maynou J,
Alvarez J, Brassesco C, Amengual MJ, Benet J, Garcia-Peiró A and
Brassesco M: Sperm telomere length in motile sperm selection
techniques: A qFISH approach. Andrologia. 50:e128402018. View Article : Google Scholar
|
|
79
|
Cariati F, Jaroudi S, Alfarawati S, Raberi
A, Alviggi C, Pivonello R and Wells D: Investigation of sperm
telomere length as a potential marker of paternal genome integrity
and semen quality. Reprod Biomed Online. 33:404–411. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Mishra S, Kumar R, Malhotra N, Singh N and
Dada R: Mild oxidative stress is beneficial for sperm telomere
length maintenance. World J Methodol. 6:163–170. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Rocca MS, Speltra E, Menegazzo M, Garolla
A, Foresta C and Ferlin A: Sperm telomere length as a parameter of
sperm quality in normozoospermic men. Hum Reprod. 31:1158–1163.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Liu SY, Zhang CJ, Peng HY, Huang XQ, Sun
H, Lin KQ, Huang K, Chu JY and Yang ZQ: Association study of
telomere length with idiopathic male infertility. Yi Chuan.
37:1137–1142. 2015.In Chinese. PubMed/NCBI
|
|
83
|
Antunes DM, Kalmbach KH, Wang F, Dracxler
RC, Seth-Smith ML, Kramer Y, Buldo-Licciardi J, Kohlrausch FB and
Keefe DL: A single-cell assay for telomere DNA content shows
increasing telomere length heterogeneity, as well as increasing
mean telomere length in human spermatozoa with advancing age. J
Assist Reprod Genet. 32:1685–1690. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Yang Q, Zhang N, Zhao F, Zhao W, Dai S,
Liu J, Bukhari I, Xin H, Niu W and Sun Y: Processing of semen by
density gradient centrifugation selects spermatozoa with longer
telomeres for assisted reproduction techniques. Reprod Biomed
Online. 31:44–50. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Yang Q, Luo X, Bai R, Zhao F, Dai S, Li F,
Zhu J, Liu J, Niu W and Sun Y: Shorter leukocyte telomere length is
associated with risk of nonobstructive azoospermia. Fertil Steril.
110:648–654.e1. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Heidary H, Pouresmaeili F, Mirfakhraie R,
Omrani MD, Ghaedi H, Fazeli Z, Sayban S, Ghafouri-Fard S, Azargashb
E and Shokri F: An association study between longitudinal changes
of leukocyte telomere and the risk of azoospermia in a population
of Iranian infertile men. Iran Biomed J. 22:231–236. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Jørgensen PB, Fedder J, Koelvraa S and
Graakjaer J: Age-dependence of relative telomere length profiles
during spermatogenesis in man. Maturitas. 75:380–385. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Pech MF, Garbuzov A, Hasegawa K, Sukhwani
M, Zhang RJ, Benayoun BA, Brockman SA, Lin S, Brunet A, Orwig KE,
et al: High telomerase is a hallmark of undifferentiated
spermatogonia and is required for maintenance of male germline stem
cells. Genes Dev. 29:2420–2434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Ling X, Zhang G, Chen Q, Yang H, Sun L,
Zhou N, Wang Z, Zou P, Wang X, Cui Z, et al: Shorter sperm telomere
length in association with exposure to polycyclic aromatic
hydrocarbons: Results from the MARHCS cohort study in Chongqing,
China and in vivo animal experiments. Environ Int. 95:79–85. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Liu M, Hales BF and Robaire B: Effects of
four chemotherapeutic agents, bleomycin, etoposide, cisplatin, and
cyclophosphamide, on DNA damage and telomeres in a mouse
spermatogonial cell line. Biol Reprod. 90:722014. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Dracxler RC, Oh C, Kalmbach K, Wang F, Liu
L, Kallas EG, Giret MT, Seth-Smith ML, Antunes D, Keefe DL, et al:
Peripheral blood telomere content is greater in patients with
endometriosis than in controls. Reprod Sci. 21:1465–1471. 2014.
View Article : Google Scholar : PubMed/NCBI
|