Introduction
Cardiovascular diseases have attracted increasing
attention in the research community due to the fact that are the
leading causes of mortalities worldwide, presenting the largest
morbidity and mortality rates (1). Atherosclerosis (AS) is the main
cause of cardiovascular disease (2,3),
which is an attractive topic due to its complex pathogenesis and
the involvement of multiple cell types (4). Inflammation is an important factor
in AS, and it is involved in each stage of the disease (5). Inflammation can oxidize the
low-density lipoproteins, inducing the chemo-taxis of monocyte,
leading to the differentiation of monocytes into macrophage foam
cells and the secretion of active molecules, eventually causing the
formation of atherosclerotic plaques (3). The modified oxidized low-density
lipoproteins can act as antigens, aggravating the inflammatory
response in the arterial vessels, ultimately affecting the
stability of the AS plaque (2,6,7).
Therefore, an increasing number of studies have reported that
anti-inflammatory treatments are effective methods to alleviate AS
(8-10).
Among all the cells that take part in the AS
process, macrophages have an important role, not only in the uptake
of cholesterol lipoproteins, becoming foam cells, but in cytokine
production in the atherosclerotic area (11). As the largest number of
inflammatory cells in the AS lesion, macrophages can produce
pro-inflammatory, such as tumor necrosis factor (TNF)α, and
anti-inflammatory cytokines, such as interleukin (IL)-10. In
addition, previous studies have demonstrated that pro-inflammatory
cytokines can promote AS development, whereas anti-inflammatory
cytokines have an anti-atherosclerotic effect (12,13).
Currently, drug therapy for atherosclerosis
includes, in particular, the use of statins, which were shown to
have a significant effect in reducing the risk of cardiovascular
disease (14). Statins have
anti-inflammatory effects that lower the concentration of lipids,
thus benefiting patients with cardiovascular disease (3). However, many patients cannot
tolerate statin drugs or cannot follow long-term treatments, and do
not show the expected reduction in lipid levels (3). In addition, large clinical trials
have shown that the reduction of cardiovascular risk by statins
treatment is still unsatisfactory (15-17). Some patients continue to suffer
from the expected cardiovascular events under statin treatment
(3). Therefore, it is necessary
to find new methods for the treatment of atherosclerosis aimed to
inhibit the development of atherosclerosis by reducing inflammation
or suppressing the formation of foam cells.
An increasing number of studies have shown that stem
cells have an important role in tissue repair and anti-inflammation
(18). In particular, numerous
experimental studies have proven that mesenchymal stem cells (MSCs)
have anti-inflammatory and immunological properties (19-24). At present, MSCs are used to
control the development of AS (5,25).
Previous studies suggested that MSCs can regulate the role of
various inflammatory cells such as macrophages and inhibit the
formation of plaques by inhibiting inflammatory responses (5,25).
Recently, multiple studies focused on the role of
macrophages in atherosclerosis progression (5,25-27). Macrophages can be classified into
M1 macrophages and M2 macrophages according their phenotype, M1
macrophages show dynamically shifting in a range spanning from a
pro-inflammatory phenotype, and M2 macrophages show a
pro-regenerative phenotype (28).
The functions of macrophages are mediated primarily by a complex
milieu of soluble mediators that influence local cells (29,30). MSCs can induce the polarization to
M2 macrophages, which modulate inflammation and immune response by
secreting a variety of cytokines that inhibit several aspects
involved in AS development (18).
MSCs were first identified in the bone marrow and
subsequently in various tissues such as the synovial membrane
(31-33). Human amnion MSCs (hAMSCs), which
are isolated from amniotic membrane of human placenta, not only
have MSCs characteristics and differentiation potential, but also
have many advantages such as low risk of tumor formation, low
immunogenicity, strong paracrine function and are easily obtained
(34,35). These properties make hAMSCs a
suitable potential resource for clinical applications (36). The immunomodulatory properties of
hAMSCs have been previously identified, including the influence of
T-cell proliferation, the inhibition of dendritic cell
differentiation and maturation (37), and the inhibition of
macrophage-mediated production of inflammatory cytokines (38,39). Therefore, the present study
investigated the effect of hAMSCs on the inhibition of the
formation and progression of atherosclerotic plaque in
apolipoprotein E-knockout (apoE-KO) mice by regulating the function
of inflammatory macrophages, suggesting a therapeutic potential of
hAMSCs for the treatment of AS.
Materials and methods
hAMSCs isolation and cell culture
The amnion samples of normal pregnancies were
obtained after caesarean section. The present study was approved by
the Ethical Committee for Medical Scientific Research of the First
Affiliated Hospital at China Medical University [approval no.
(2016)105] and the informed consent was obtained from all patients.
In the present study, 32 pregnant women (age range, 25-35 years)
were enrolled between July 2016 and December 2016 in the First
Affiliated Hospital at China Medical University. The isolation and
culture of hAMSCs were performed as previously described (40). Briefly, the amnion samples were
treated with collagenase IV (Sigma-Aldrich; Merck KGaA) and DNase I
(Takara Bio, Inc.) after manual separation from the chorion. hAMSCs
were cultured at 37°C in DMEM/F12 (Hyclone; GE Healthcare Life
Sciences) containing 10% FBS (Hyclone; GE Healthcare Life
Sciences), 1% penicillin streptomycin (Gibco; Thermo Fisher
Scientific, Inc.) and 10 mmol/ml fibroblast growth factor-2
(PeproTech, Inc.). hAMSCs were passaged every 72 h for 3-6
times.
Flow cytometric characterization of
hAMSCs
Flow cytometry was used to detect stem cell-related
cell surface markers on hAMSCs. hAMSCs were cultured at 37°C for 48
h prior to analysis. Then, cells were harvested, washed,
resuspended with 1X PBS containing 1% BSA. Cells were counted and
diluted to 5×106 cells/100 ml and incubated for 1 h on
ice. Subsequently, cells were incubated with monoclonal
phycoerythrin-conjugated antibodies for CD44 (cat. no. 338807),
CD90 (cat. no. 32810), CD31 (cat. no. 303105), CD45 (cat. no.
368509), stage-specific embryonic antigen 4 (SSEA-4; cat. no.
330405) and major histocompatibility complex, class II, DR (HLA-DR;
cat. no. 307605). All of the antibodies were purchased from
BioLegend, Inc. Appropriate isotype-matched antibodies were used as
negative controls (BD Biosciences). The data from 10,000 viable
cells were acquired with a flow cytometer and analyzed using the
FACSDiva software (version 6.2; BD Biosciences).
Animals
In total, 36 C57BL/6 apoE-KO male mice (age, 8
weeks; weight, 25±2 g) were obtained from Beijing HFK Bioscience
Co. Ltd. (5). All the animals
were housed in an environment with a temperature of 22±1°C,
relative humidity of 50±1% and a light/dark cycle of 12 h and
received drinking water ad libitum. In addition, mice were
fed with high-fat diet containing 21% fat and 0.15% cholesterol
from 8 weeks of age. Furthermore, all animal studies, including the
mice euthanasia procedure, were done in compliance with the
regulations and guidelines of China Medical University
institutional animal care and conducted according to AAALAC
guideline and IACUC guidelines (approval no. CMU2016028). The
experiments performed on human samples and animals were approved by
the Ethical Committee for Medical Scientific Research of the First
Affiliated Hospital at China Medical University [approval no.
(2016)105]. To examine the role of hAMSCs on the prevention and
treatment of atherosclerosis, hAMSCs were administered by tail vein
injection as previously described (5,27).
For the early-hAMSCs treatment study, 18 apoE-KO mice were treated
with hAMSCs with high-fat diet for 10 weeks. In the delayed-hAMSCs
treatment study, 18 apoE-KO mice were first fed with high-fat diet
for 8 weeks, and were then treated with hAMSCs for 10 weeks. In
each treatment regimen, mice were randomly divided into three
groups (n=6 in each group): i) hAMSCs group, containing mice
treated with hAMSCs (5×105 hAMSCs in 150 µl PBS)
once every other week; ii) PBS group, containing mice treated with
150 µl PBS once every other week; and iii) control group,
without treatment. 'Control 1' group was defined as the control
group in the early-hAMSCs treatment and 'control 2' group was
defined as the control group in the late-hAMSCs treatment.
After the treatment, all mice were anesthetized by
intraperitoneal injection of pentobarbital (50 mg/kg). Lipid
analysis was performed on the serum samples obtained from blood
collected from the left ventricles of mice at the end of the
studies. The serum total cholesterol, triglyceride (TG) and
high-density lipoprotein (HDL)-cholesterol were measured by
enzymatic methods using a commercially available kit (Sekisui
Chemical Co., Ltd.) with a Hitachi Automatic Analyzer 7600
(Hitachi, Ltd.). Low-density lipo-protein (LDL)-cholesterol was
calculated using the Friedewald formula: LDL-cholesterol = total
cholesterol-HDL-cholesterol-(TG/5) (41). Non-HDL-cholesterol was calculated
by subtracting the quantity of HDL-cholesterol from total
cholesterol (42).
Histological and immunohistochemical
analysis
After left ventricle perfusion, the aorta with the
aorta root, the aortic arch and the iliac artery was collected. The
aortic root was removed, fixed with 4% paraformaldehyde at room
temperature and embedded in paraffin. In total, five sections
(thickness, 5 µm) of each aortic root were stained with
hematoxylin and eosin for 5 min at room temperature. Pathological
changes in each group were observed using a light microscope
(Olympus Corporation; magnification, ×200).
Paraffin-embedded sections were exposed to
increasing ethanol concentrations. After incubation with 3%
hydrogen peroxide solution for 15 min at room temperature to
inhibit endogenous peroxidase activity, 0.1 M sodium citrate
solution was used for antigen retrieval at 120°C for 90 sec and 5%
BSA was used to block non-specific signal at room temperature for
20 min, sections were incubated with anti-CD68 rabbit poly-clonal
antibody (1:500; cat. no. GB11067; Wuhan Servicebio Co., Ltd.)
overnight at 4°C. The sections were washed with PBS and incubated
with Goat anti-rabbit IgG (horseradish peroxidase-conjugated;
1:1,000, cat. no. ab6721; Abcam) at room temperature for 30 min.
The sections were then visualized with 3,3′-diaminobenzidine that
was incubated with the samples at room temperature for 1 min. The
nuclei were counterstained with hematoxylin at room temperature for
2 min. The macrophage coverage was determined by the percentage of
the positive stained area in relation to the total plaque area.
Samples were observed using a light microscope (Olympus
Corporation; magnification, ×200).
Western blotting
Total protein lysates were extracted from the aortic
tissue using RIPA lysis buffer (Beyotime Institute of
Biotechnology) supplemented with protease inhibitors, according to
the manufacturer's instructions. The protein concentration was
measured using a bicinchoninic protein assay kit (Beyotime
Institute of Biotechnology). The samples with equal amount of total
protein (20 µg in each lane) were then heated at 100°C for 5
min in loading buffer, separated by 10% SDS-PAGE (Beijing Transgen
Biotech Co., Ltd.) and transferred to PVDF membranes (EMD
Millipore). PVDF membranes were blocked with 5% skim milk for 1 h
at room temperature and incubated overnight at 4°C with the
following primary antibodies: TNFα monoclonal antibody (1:1,000;
cat. no. 60291-1-lg; ProteinTech Group, Inc.), IL-10 antibody
(1:500; cat. no. DF6894; Affinity Biosciences), phosphorylated
(p-)NF-κB p65 rabbit mAb (1:500; cat. no. 3033; Cell Signaling
Technology, Inc.), p-inhibitor of κB-α (IκBα) rabbit (1:500; cat.
no. 2859; Cell Signaling Technology, Inc.), NF-κB p65 rabbit mAb
(1:1,000; cat. no. 8242; Cell Signaling Technology, Inc.), IκBα
rabbit mAb (1:500; cat. no. 4812; Cell Signaling Technology, Inc.)
and mouse anti-β actin monoclonal antibody (1:500; cat. no. TA-09;
OriGene Technologies, Inc.). The membrane was incubated for 1 h at
room temperature with the secondary peroxidase-conjugated antibody
goat anti-rabbit IgG (1:1,000; cat. no. zb2301; OriGene
Technologies, Inc.) or the peroxidase-conjugated goat anti-mouse
IgG (1:1,000; cat. no. zb2305; OriGene Technologies, Inc.), the
membranes were visualized using an ECL kit (Thermo Fisher
Scientific, Inc.) and analyzed using ImageJ software (version
1.5.1; National Institutes of Health).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the aortic tissues
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
RNA concentration was detected using the NanoDrop 2000 (Thermo
Fisher Scientific, Inc.). RT was performed using the PrimeScript RT
Reagent kit (Takara Bio, Inc.). The reaction was performed as
follows: 37°C for 15 min followed by 85°C for 5 sec. qPCR was
performed using the SYBR PrimeScript RT-PCR kit (Takara Bio, Inc.)
with an ABI 7500 Sequence Detection system (Applied Biosystems;
Thermo Fisher Scientific, Inc.). PCR was conducted using the
following thermocycling conditions: Initial denaturation at 95°C
for 30 sec, followed by 40 cycles of 95°C for 5 sec and 60°C for 20
sec. All the gene expression results were normalized to the
expression of GAPDH. The primer sequences were as follows: TFNα
forward, 5′-AGAGTGGTCAGGTTGCCTCTG-3′ and reverse,
5-GGCTCTGTGAGGAAGGCTGT-3′; IL-10 forward,
5′-GGCTCTGTGAGGAAGGCTGT-3′ and reverse, 5′-TTCCGATAAGGCTTGGCAAC-3′;
and GAPDH forward, 5′-GGTTGTCTCCTGCGACTTCA-3′ and reverse,
5′-TGGTCCAGGGTTTCTTACTCC-3′. Normalization and fold changes were
calculated using the 2−∆∆Cq method (43).
Statistical analysis
The results are presented as the mean ± SD from at
least three independent experiments. Statistical analysis was
performed using one-way ANOVA followed by least significant
difference post hoc test using SPSS 22.0 (IBM Corp.). P<0.05 was
considered to indicate a statistically significant difference.
Results
Characterization of hAMSCs
After purification, hAMSCs were identified to be
positive for CD44 and CD90, and negative for CD31 and CD45
(Fig. 1). The present data
indicated that hAMSCs had stem cell characteristics, as they
expressed SSEA-4 (44), and low
immunogenicity, as hAMSCs were negative for HLA-DR (45).
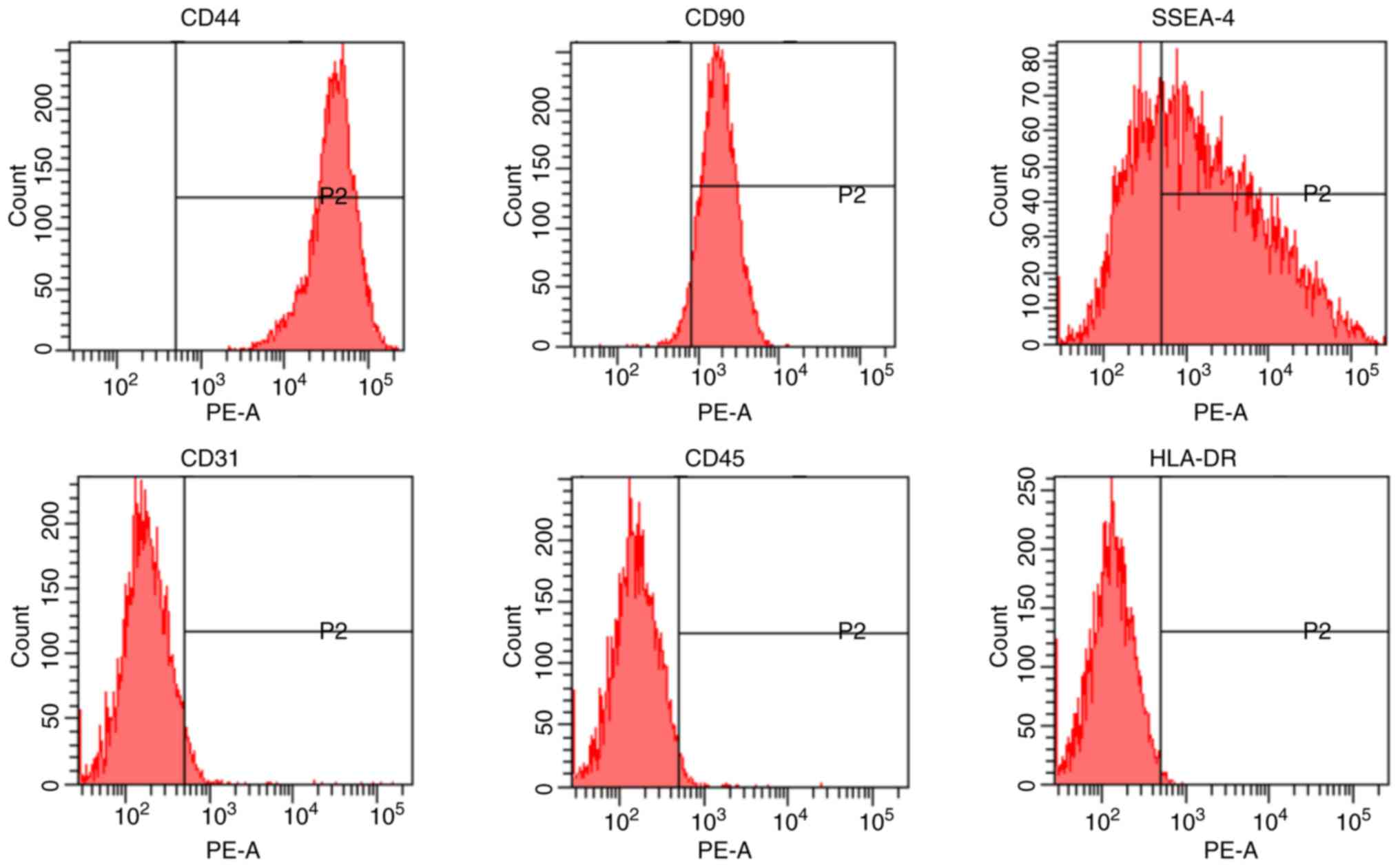 | Figure 1Characterization of hAMSCs. hAMSCs
were characterized by analyzing the expression of the cell surface
markers CD44, CD90, SSEA-4, CD31, CD45 and HLA-DR using flow
cytometry. hAMSCs, human amniotic mesenchymal stem cells; PE,
phycoerythrin; HLA-DR, major histocompatibility complex, class II,
DR; SSEA-4, stage-specific embryonic antigen 4. |
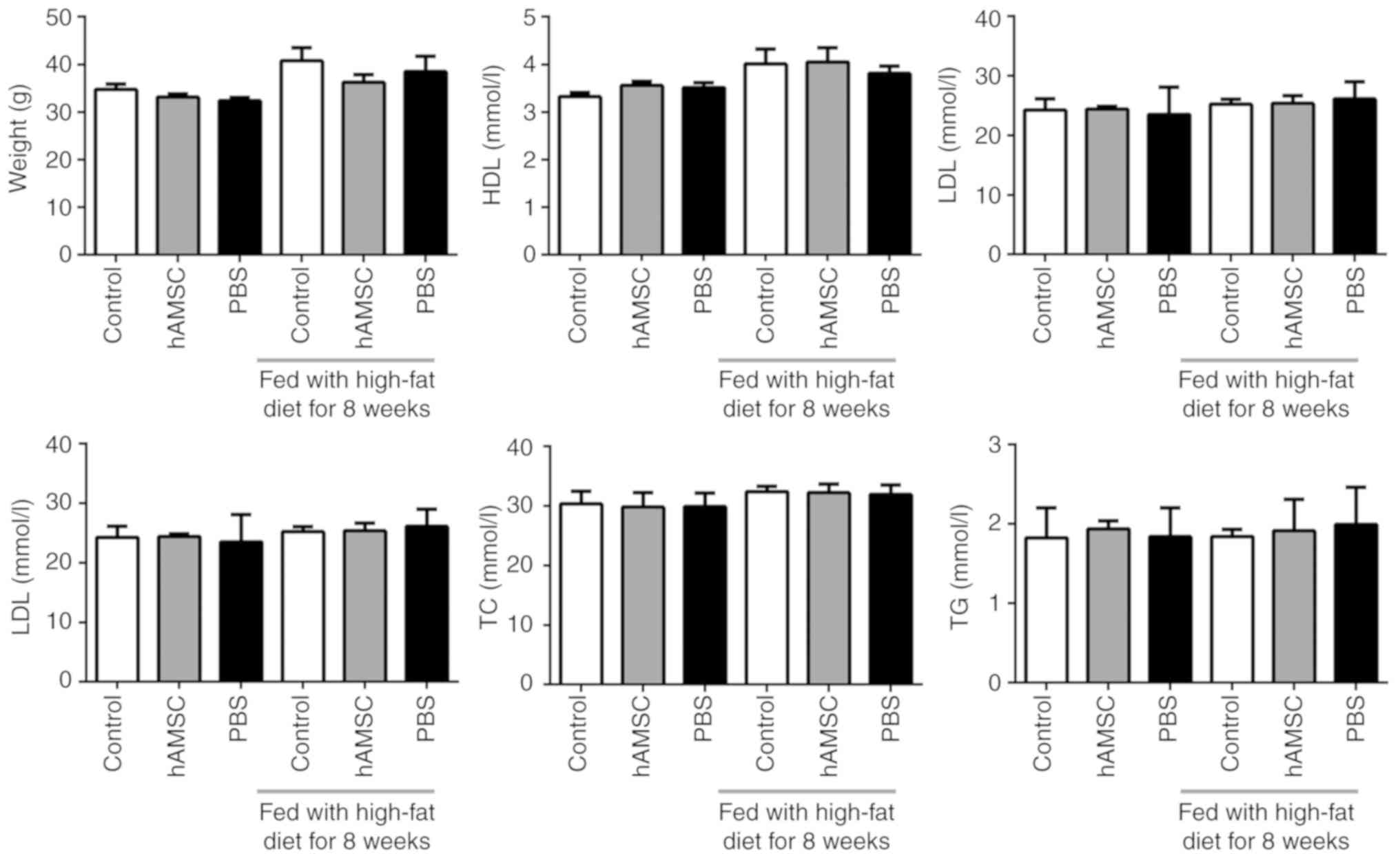
Effects of hAMSCs treatment on weight and
plasma lipid levels
The weight of the mice was calculated. No
significant difference was observed between the hAMSCs treatment
group and the control group either in early hAMSCs treatment study
or in late hAMSCs treatment study (n=6 in each group). In addition,
treatment with hAMSCs did not affect the concentration of plasma
lipids (Fig. 2).
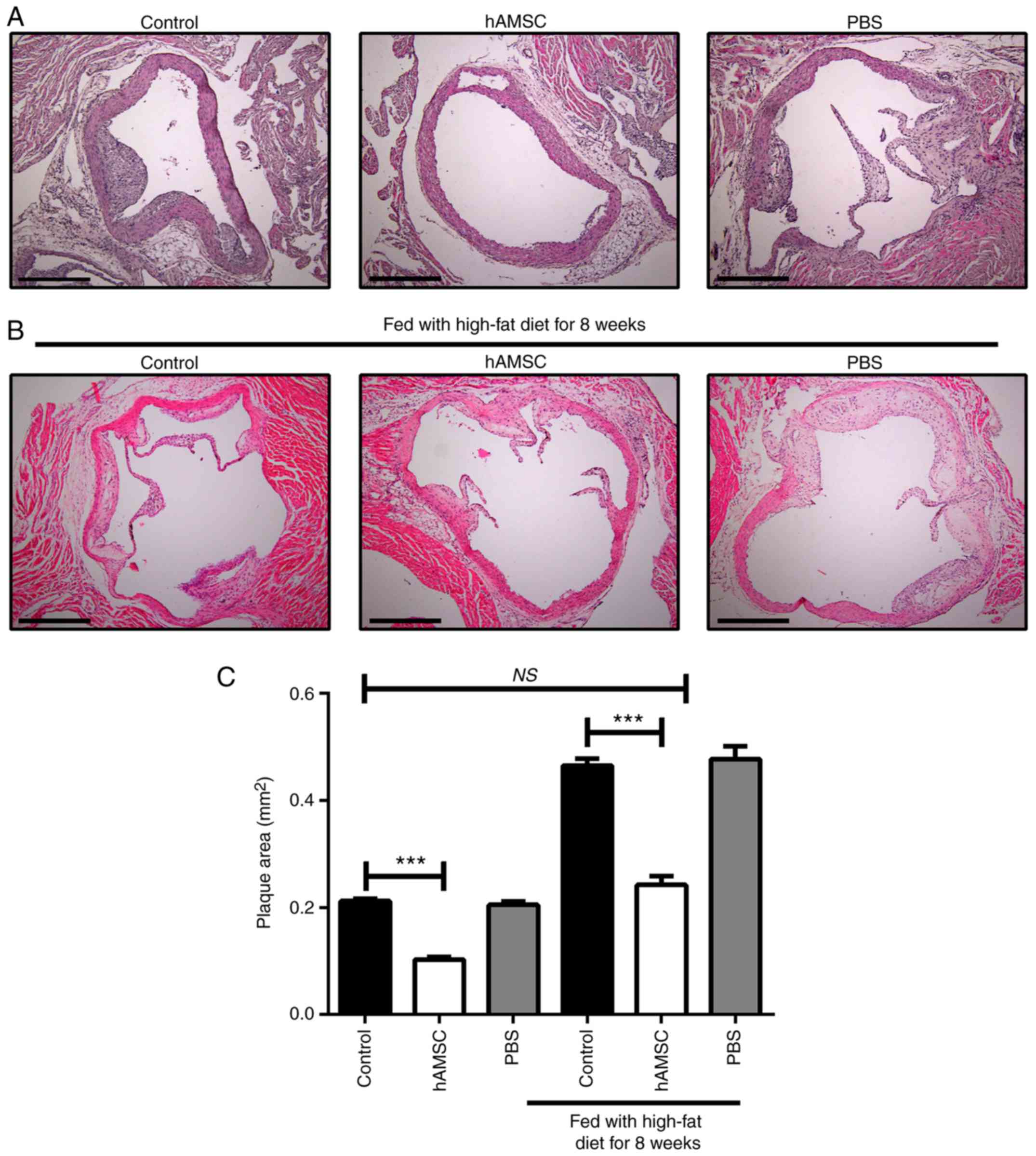
Effects of hAMSCs treatment on the
development of atherosclerotic lesions
After 10 weeks of hAMSCs treatment, histologic
analysis of sections taken from the aortic root was performed. A
significant decrease was found in plaque size in mice treated with
hAMSCs injection compared with control mice in early and late
hAMSCs treatment groups, which suggested that hAMSCs treatment
inhibited the plaque formation and progression of established
lesions. The signifi-cant difference between control 1 and control
2 suggested that prolonging the high-fat diet worsened the
atherosclerotic lesions. Moreover, no significant difference was
observed in lesion size between late hAMSCs treatment group and
early control group, which presented initial signs of
atherosclerotic lesions, indicating that treatment with hAMSCs did
not regress the established lesions (Fig. 3).
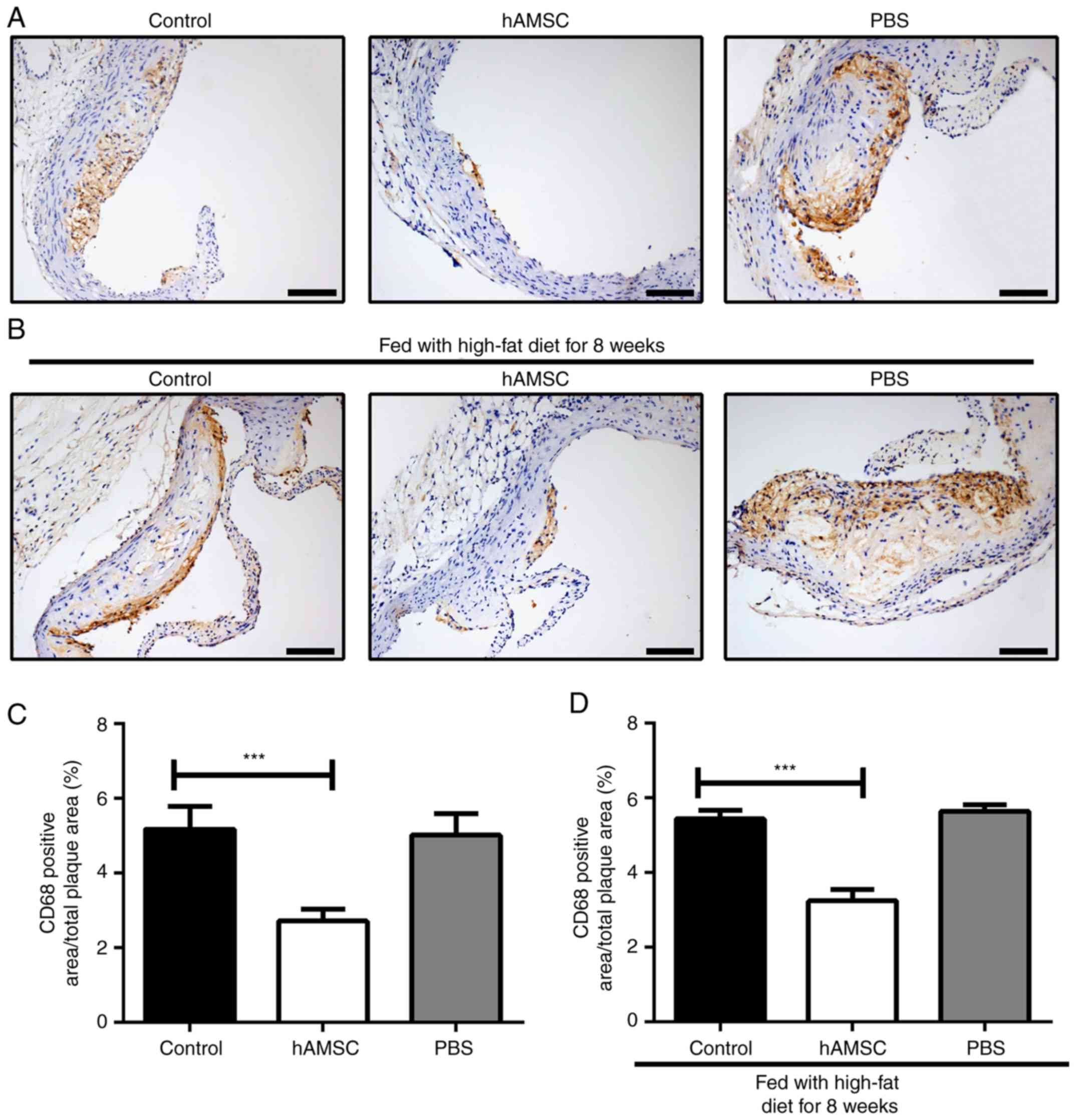
Effects of hAMSCs treatment on macrophage
accumulation in atherosclerotic plaque
To detect the effect of hAMSCs treatment on the
accumulation of macrophages, immunohistochemical analysis was
performed on the aortic root. The present results showed that
hAMSCs treatment suppressed the accumulation of macrophages in both
early and late hAMSCs treatment groups; the number of macrophages
in atherosclerotic plaque was significantly decreased in mice
treated with hAMSCs compared with the PBS group control mice.
However, the accumulation of macrophage may not be associated with
a prolonged high-fat diet, as no significant difference between
control 1 and control 2 was identified (Fig. 4).
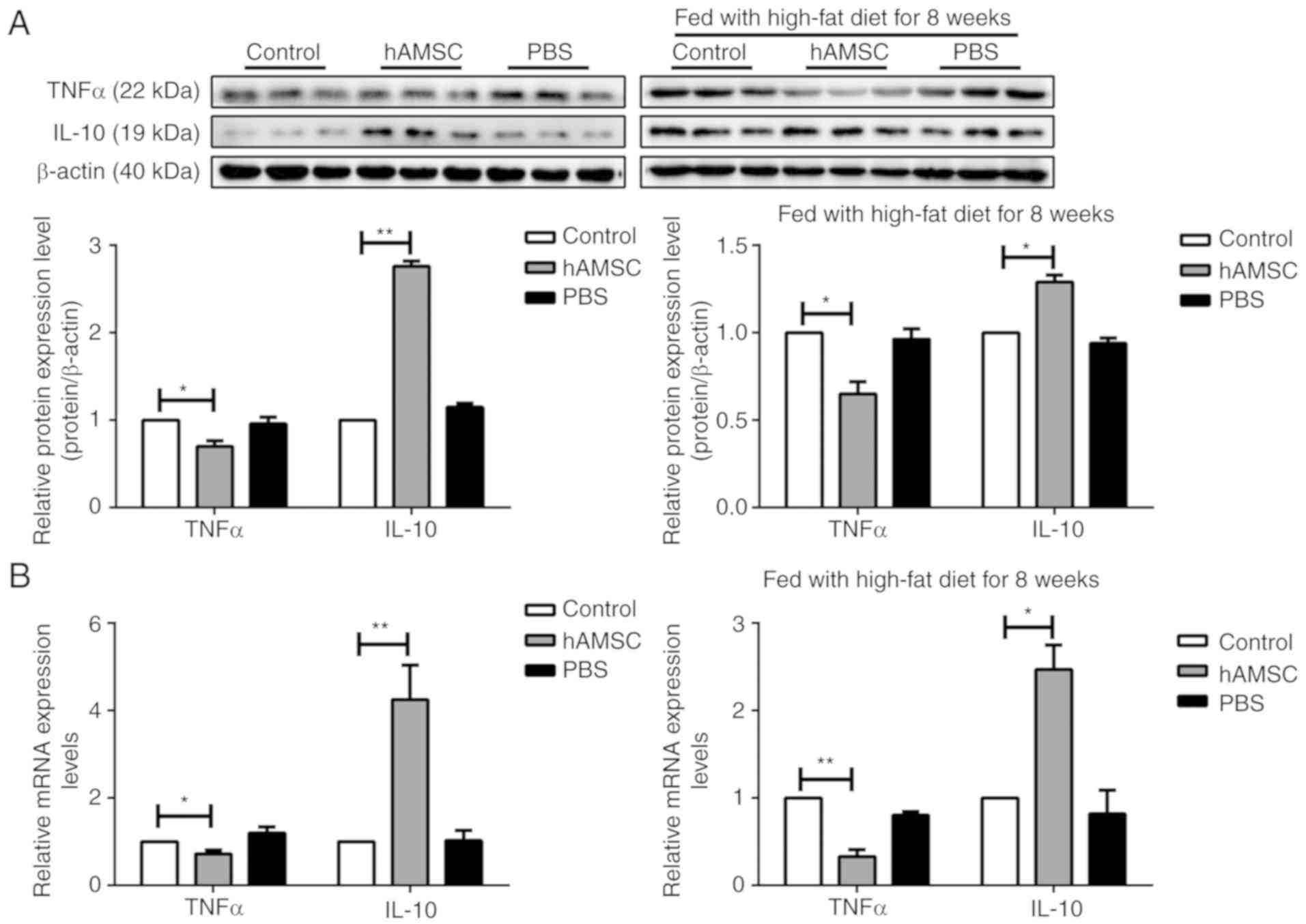
hAMSCs modulates cytokine expression in a
mouse model of AS
Macrophages are the most important source of
cytokine production (46). To
assess the inhibitory effect of hAMSCs on cytokine secretion, the
expression of the pro-inflammatory cytokine TNFα and the
anti-inflammatory cytokine IL-10 was determined from
atherosclerotic arteries by western blotting and RT-qPCR analysis.
In contrast with the control group, TNFα expression was
significantly decreased, whereas IL-10 expression was increased in
the hAMSCs treatment group in both early hAMSCs treatment group and
late hAMSCs treatment group (Fig.
5).
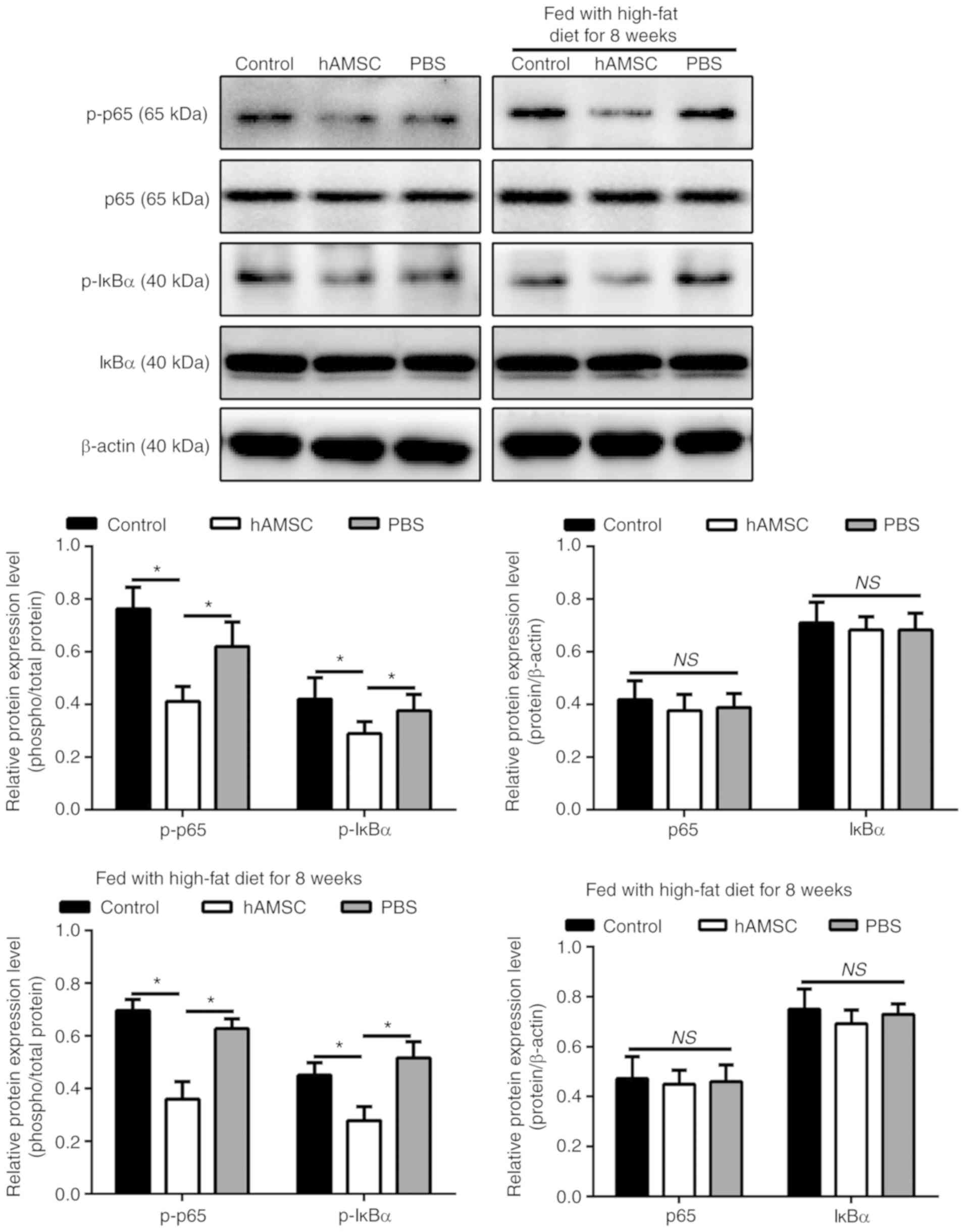
hAMSCs suppresses the phosphorylation of
p65 and IкBα in a mouse model of AS
To determine whether NF-κB signaling pathway was
involved in the regulatory effect of hAMSCs on atherosclerotic
plaque in a mouse model of AS, the phosphorylation level of two
NF-κB signaling molecules, p65 and IкBα, were investigated.
Notably, the phosphorylation level of p65 and IκBα are associated
with the activation of the NF-κB pathway (47). The results of western blot
analysis showed that the levels of p-p65 and IκBα were
downregulated in mice treated with hAMSCs injection compared with
the control mice in both early hAMSCs treatment group and in late
hAMSCs treatment group (Fig.
6).
Discussion
In the present study, hAMSCs injection was
identified to reduce AS development through the decrease of
macrophage accumulation and the inhibition of inflammatory response
of aortic arteries. The present results suggested that the levels
of secreted cytokines, including TNFα and IL-10, was modulated by
hAMSCs through the NF-κB pathway.
AS is a disease in which atherosclerotic plaques
develop inside the arteries (48). Macrophages are the most abundant
immune cell type in atherosclerotic lesions and have an essential
role during all stages of the disease, from lesion initiation to
plaque rupture (49).
Atherosclerotic plaques and their macrophage content can regress,
as previously shown in particular murine models of
hypercholesterolemia reversal (48,50,51). Decrease of macrophage accumulation
and functional regulation in atherosclerotic lesions could
represent a potential target in the treatment of AS. The present
study suggested that systemic administration of hAMSCs decreased AS
development and progression when simultaneously applied with
high-fat diet. hAMSCs inhibited AS progression after 8 weeks when
combined with high-fat diet in apoE-KO mice. In addition, the
therapeutic benefit of hAMSCs could be explained by the modulation
of the inflammatory response, as after hAMSCs treatment the
accumulation of macrophages in the plaque of aortic root was
decreased. Importantly, the function of macrophage in the plaque
was not investigated, and further studies are required to examine
this phenomenon. In addition, the expression of the
pro-inflammatory cytokine TNFα was significantly downregulated,
whereas the anti-inflammatory cytokine IL-10 was upregulated in
aortic arches. However, the present study did not identify direct
and molecular evidence associating hAMSCs and macrophages
recruitment and function in the plaque region, and further studies
are required to examine this process.
Numerous previous studies have shown that MSCs can
reduce AS development by inhibiting the inflammatory response
(5,25,27,52). MSCs might be directly involved in
AS plaque formation, as Li et al (5) observed that MSCs are capable of
migrating to AS plaque and selectively locating near macrophages,
and Fang et al (52)
observed 5-bromo-2-deoxyuridine-positive MSCs in AS plaque.
Nevertheless, Frodermann et al (25) observed that MSCs primarily
accumulate in the lungs after intravenous injection, and only a few
MSCs could migrate to the lymph nodes of the heart and the aorta.
These previous results suggests that MSCs do not necessarily have
an anti-atherosclerotic role due to long-term engraftment, but may
act via the paracrine signaling pathway, since MSCs have a
significant paracrine function (53). Our previous study suggested that
hAMSCs have a strong paracrine function, which could significantly
promote the proliferation and tube formation of endothelial cells
(40). Similar experiments on the
therapeutic improvement of MSCs without significant engraftment
have been reported in various animal models (39,54). Due to the limited time and
research funds, the present study did not investigate the fate of
the injected hAMSCs, which requires to be addressed by further
studies in the future.
The development of atherosclerotic plaque is a
multistep inflammatory process. Macrophages have an important role
in all stages of AS and they serve as effectors in the process of
AS (55). The important
pathological changes in AS include the accumulation of macrophages
in endothelial lesions (56).
These macrophages can become foam cells after uptake of modified
lipoproteins, which trigger the release of cytokines in macrophages
via the activation of a heterodimer formed by Toll-like receptors 4
and 6, aggravating the inflammatory reaction (57). Therefore, reducing the aggregation
of macrophages and the formation of foam cells in atherosclerotic
plaque is essential to control the inflammatory response during AS.
According to previous studies, MSCs could reduce the aggregation of
macrophages in the arterial intima (39), inhibit the formation of
macrophage-foam cells (27),
decrease the expression of inflammatory factor TNFα and increase
the expression of anti-inflammatory factors such as IL-10 (5). In the present study, hAMSCs
treatment was found to decrease the accumulation of macrophages in
atherosclerotic plaque. Notably, the present results may be caused
by the systemic influence of the immune response in response to the
injection of exogenous cells into the bloodstream. In addition,
hAMSCs treatment may influence local tissue micro-environment,
inducing the polarization to M2 macrophages and influencing the
function of macrophages (58).
However, the molecule mechanism underlying the association between
hAMSCs treatment and macrophage recruitment and function require
further investigation. A previous study observed that following
co-culture with hAMSCs, the levels of TNFα and IL-1β in
lipopolysaccha-ride-stimulated macrophages is significantly
reduced, which may be caused by inhibition of the NF-κB pathway
(38).
NF-κB pathway is an important transcription factor
that regulates both innate and adaptive immune responses and is
essential for the expression of a number of genes involved in the
inflammatory response, such as TNFα and IL-6 (38). These genes can directly activate
the NF-κB pathway, promoting the inflammatory response, which
involves the phosphorylation of p65 and IκBα (47,59). Since MSCs have an
anti-inflammatory role in an NF-κB-dependent manner (5), hAMSCs may promote the downregulation
of TNFα and upregulation of IL-10 via the NF-κB pathway. In the
present study, following hAMSCs treatment, the phosphorylation
levels of p65 and IκBα were downregulated, suggesting an inhibition
of the NF-κB pathway. Therefore, the anti-inflammatory function of
hAMSCs may be mediated by the inhibition of the NF-κB pathway.
Previous studies reported that the anti-inflammatory
role of MSCs caused not only the decrease in TNFα level, but also
the increase in IL-10 levels (60,61). Notably, IL-10 was observed to
decrease the synthesis of TNFα (60). Nemeth et al (62) reported the macrophages isolated
from MSCs-treated septic mice produce significantly higher levels
of IL-10 than those isolated from non-treated mice, suggesting a
temporary reprogramming of monocytes and macrophages. IL-10 was
observed to promote atherosclerotic lesion stability (13), and the present results suggested
that MSCs can increase the levels of IL-10 in addition to reducing
AS development by modulating the levels of anti-inflammatory
molecules. However, further studies are required to investigate
whether the blockage of IL-10 could reduce the effects of hAMSCs
treatment. In the present study, an increase in IL-10 in the aortic
vasculature was observed after hAMSCs treatment; therefore, the
effect of hAMSCs on AS stabilization should be further
investigated.
In the present study, no significant effects of
hAMSCs on mice blood lipid levels were observed. To the best of our
knowledge, the report by Frodermann et al (25) is the only study that reported a
decrease in blood lipid levels following MSC treatment. Notably, in
this previous study, MSCs were isolated from the bone marrow and
not from the amniotic fluid, and these previous results may be
associated with the different source of MSCs. Additionally, in the
study by Frodermann et al (25), the decrease in blood lipid levels
in mice was identified 4-5 weeks after MSCs treatment, suggesting
that MSCs may indirectly regulate the level of blood lipids by
regulating other cellular functions. Previous studies observed that
hAMSCs have paracrine function and can secret various types of
cytokines, chemokines and growth factors, which are important for
intercellular interactions (40,53). Therefore, further studies are
necessary to examine the role of these cytokines in AS progression
and development.
In summary, the present study examined the effect of
hAMSCs on AS. In addition, hAMSCs treatment was found to be
effective in reducing the immune response, one of the major
pathways involved in AS, leading to a significant reduction in the
size of atherosclerotic lesions. Cytotherapy has attracted
increasing interest, and finding the most effective methodology to
employ stem cells is an important aspect for identifying the
optimal treatment methods. The present study suggested that hAMSCs
may represent a promising cell source for clinical
applications.
Acknowledgments
Not applicable.
Funding
This study was supported by the National Basic
Research Program of China (grant no. 2012CB518103), the National
Natural Science Foundation of China (grant no. 81450017), the
Science and Technology Planning Project of Liaoning Province (grant
no. 2014305012), the Science and Technology Planning Project of
Shenyang (grant no. F14-201-4-00), the Shenyang Key Laboratory
Project (grant no. F15-157-1-00), and the National Natural Science
Foundation of China (grant no. 81300038).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW and GQ conceived and designed the study and
drafted the manuscript. GS, XZ and QW performed experiments and
interpreted the results. LC, YX and XP analyzed the data. GQ
contributed to the acquisition of funding. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Approval to conduct the study was obtained from
Ethics Committee for Medical Scientific Research in the First
Affiliated Hospital of China Medical University [approval no.
(2016)105]. All animal studies, including the mice euthanasia
procedure, were performed in compliance with the regulations and
guidelines of China Medical University institutional animal care
and conducted according to AAALAC guideline and IACUC guidelines
(63) and were approved by the
Ethics Committee (approval no. CMU2016028).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mozaffarian D, Benjamin EJ, Go AS, Arnett
DK, Blaha MJ, Cushman M, de Ferranti S, Després JP, Fullerton HJ,
Howard VJ, et al: Heart disease and stroke statistics-2015 update:
A report from the american heart association. Circulation. 131. pp.
e29–e322. 2015
|
|
2
|
Frostegård J: Immunity, atherosclerosis
and cardiovascular disease. BMC Med. 11:1172013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang X, Huang F, Chen Y, Qian X and Zheng
SG: Progress and prospect of mesenchymal stem cell-based therapy in
atherosclerosis. Am J Transl Res. 8:4017–4024. 2016.PubMed/NCBI
|
|
4
|
Wu DJ, Xu JZ, Wu YJ, Jean-Charles L, Xiao
B, Gao PJ and Zhu DL: Effects of fasudil on early atherosclerotic
plaque formation and established lesion progression in
apolipoprotein E-knockout mice. Atherosclerosis. 207:68–73. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li Q, Sun W, Wang X, Zhang K, Xi W and Gao
P: Skin-derived mesenchymal stem cells alleviate atherosclerosis
via modulating macrophage function. Stem Cells Transl Med.
4:1294–1301. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
De Jager SC and Pasterkamp G: Crosstalk of
lipids and inflammation in atherosclerosis: The PRO of PGRN?
Cardiovasc Res. 100:4–6. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Johansson ME, Zhang XY, Edfeldt K,
Lundberg AM, Levin MC, Borén J, Li W, Yua XM, Folkersen L, Eriksson
P, et al: Innate immune receptor NOD2 promotes vascular
inflammation and formation of lipid-rich necrotic cores in
hypercholesterolemic mice. Eur J Immunol. 44:3081–3092. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Khan R, Spagnoli V, Tardif JC and L'Allier
PL: Novel anti-inflammatory therapies for the treatment of
atherosclerosis. Atherosclerosis. 240:497–509. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mendel I, Yacov N, Harats D and Breitbart
E: Therapies targeting innate immunity for fighting inflammation in
atherosclerosis. Curr Pharm Des. 21:1185–1195. 2015. View Article : Google Scholar
|
|
10
|
Asciutto G, Dias NV, Edsfeldt A, Alm R,
Fredrikson GN, Gonçalves I and Nilsson J: Low levels of IgG
autoantibodies against the apolipoprotein B antigen p210 increases
the risk of cardiovascular death after carotid endarterectomy.
Atherosclerosis. 239:289–294. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tedgui A and Mallat Z: Cytokines in
atherosclerosis: Pathogenic and regulatory pathways. Physiol Rev.
86:515–581. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Little PJ, Chait A and Bobik A: Cellular
and cytokine-based inflammatory processes as novel therapeutic
targets for the prevention and treatment of atherosclerosis.
Pharmacol Ther. 131:255–268. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Han X and Boisvert WA: Interleukin-10
protects against atherosclerosis by modulating multiple atherogenic
macrophage function. Thromb Haemost. 113:505–512. 2015. View Article : Google Scholar
|
|
14
|
Shapiro MD and Fazio S: From lipids to
inflammation: New approaches to reducing atherosclerotic risk. Circ
Res. 118:732–749. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hague W, Forder P, Simes J, Hunt D, Tonkin
A and Investigators L: Effect of pravastatin on cardiovascular
events and mortality in 1516 women with coronary heart disease:
Results from the long-term intervention with pravastatin in
ischemic disease (LIPID) study. Am Heart J. 145:643–651. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Libby P: The forgotten majority:
Unfinished business in cardiovascular risk reduction. J Am Coll
Cardiol. 46:1225–1228. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Serban MC, Banach M and Mikhailidis DP:
Clinical implications of the IMPROVE-IT trial in the light of
current and future lipid-lowering treatment options. Expert Opin
Pharmacother. 17:369–380. 2016. View Article : Google Scholar
|
|
18
|
Zhang QZ, Su WR, Shi SH, Wilder-Smith P,
Xiang AP, Wong A, Nguyen AL, Kwon CW and Le AD: Human
gingiva-derived mesenchymal stem cells elicit polarization of m2
macrophages and enhance cutaneous wound healing. Stem Cells.
28:1856–1868. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chai NL, Zhang XB, Chen SW, Fan KX and
Linghu EQ: Umbilical cord-derived mesenchymal stem cells alleviate
liver fibrosis in rats. World J Gastroenterol. 22:6036–6048. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tan L, Dai T, Liu D, Chen Z, Wu L, Gao L,
Wang Y and Shi C: Contribution of dermal-derived mesenchymal cells
during liver repair in two different experimental models. Sci Rep.
6:253142016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xie Z, Hao H, Tong C, Cheng Y, Liu J, Pang
Y, Si Y, Guo Y, Zang L, Mu Y and Han W: Human umbilical
cord-derived mesenchymal stem cells elicit macrophages into an
anti-inflammatory phenotype to alleviate insulin resistance in type
2 diabetic rats. Stem Cells. 34:627–639. 2016. View Article : Google Scholar
|
|
22
|
Seebach E, Freischmidt H, Holschbach J,
Fellenberg J and Richter W: Mesenchymal stroma cells trigger early
attraction of M1 macrophages and endothelial cells into fibrin
hydrogels, stimulating long bone healing without long-term
engraftment. Acta Biomater. 10:4730–4741. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Braza F, Dirou S, Forest V, Sauzeau V,
Hassoun D, Chesné J, Cheminant-Muller MA, Sagan C, Magnan A and
Lemarchand P: Mesenchymal stem cells induce suppressive
macrophages-through phagocytosis in a mouse model of asthma. Stem
Cells. 34:1836–1845. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Maria ATJ, Toupet K, Maumus M, Fonteneau
G, Le Quellec A, Jorgensen C, Guilpain P and Noël D: Human adipose
mesenchymal stem cells as potent anti-fibrosis therapy for systemic
sclerosis. J Autoimmun. 70:31–39. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Frodermann V, van Duijn J, van Pel M, van
Santbrink PJ, Bot I, Kuiper J and de Jager SC: Mesenchymal stem
cells reduce murine atherosclerosis development. Sci Rep.
5:155592015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bobryshev YV, Ivanova EA, Chistiakov DA,
Nikiforov NG and Orekhov AN: Macrophages and their role in
atherosclerosis: Pathophysiology and transcriptome analysis. Biomed
Res Int. 2016:95824302016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang ZX, Wang CQ, Li XY, Feng GK, Zhu HL,
Ding Y and Jiang XJ: Mesenchymal stem cells alleviate
atherosclerosis by elevating number and function of CD4(+)CD25
(+)FOXP3 (+) regulatory T-cells and inhibiting macrophage foam cell
formation. Mol Cell Biochem. 400:163–172. 2015. View Article : Google Scholar
|
|
28
|
Murray PJ, Allen JE, Biswas SK, Fisher EA,
Gilroy DW, Goerdt S, Gordon S, Hamilton JA, Ivashkiv LB, Lawrence
T, et al: Macrophage activation and polarization: Nomenclature and
experimental guidelines. Immunity. 41:14–20. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Michaeli S, Dakwar V, Weidenfeld K,
Granski O, Gilon O, Schif-Zuck S, Mamchur A, Shams I and Barkan D:
Soluble mediators produced by pro-resolving macrophages inhibit
angiogenesis. Front Immunol. 9:7682018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Triantafyllou E, Woollard KJ, McPhail MJW,
Antoniades CG and Possamai LA: The role of monocytes and
macrophages in acute and acute-on-chronic liver failure. Front
Immunol. 9:29482018. View Article : Google Scholar
|
|
31
|
Hu Y, Liao L, Wang Q, Ma L, Ma G, Jiang X
and Zhao RC: Isolation and identification of mesenchymal stem cells
from human fetal pancreas. J Lab Clin Med. 141:342–349. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tsai MS, Lee JL, Chang YJ and Hwang SM:
Isolation of human multipotent mesenchymal stem cells from
second-trimester amniotic fluid using a novel two-stage culture
protocol. Hum Reprod. 19:1450–1456. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee OK, Kuo TK, Chen WM, Lee KD, Hsieh SL
and Chen TH: Isolation of multipotent mesenchymal stem cells from
umbilical cord blood. Blood. 103:1669–1675. 2004. View Article : Google Scholar
|
|
34
|
Kmiecik G, Niklinska W, Kuc P,
Pancewicz-Wojtkiewicz J, Fil D, Karwowska A, Karczewski J and
Mackiewicz Z: Fetal membranes as a source of stem cells. Adv Med
Sci. 58:185–195. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim EY, Lee KB and Kim MK: The potential
of mesenchymal stem cells derived from amniotic membrane and
amniotic fluid for neuronal regenerative therapy. BMB Rep.
47:135–140. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xiao J: Human amniotic stem cells: Ideal
seed cells source for regenerative medicine. J Zunyi Med Univ.
38:439–449. 2015.
|
|
37
|
Kronsteiner B, Peterbauer-Scherb A,
Grillari-Voglauer R, Redl H, Gabriel C, van Griensven M and Wolbank
S: Human mesenchymal stem cells and renal tubular epithelial cells
differentially influence monocyte-derived dendritic cell
differentiation and maturation. Cell Immunol. 267:30–38. 2011.
View Article : Google Scholar
|
|
38
|
Shu J, He X, Zhang L, Li H, Wang P and
Huang X: Human amnion mesenchymal cells inhibit
lipopolysaccharide-induced TNF-α and IL-1β production in THP-1
cells. Biol Res. 48:692015. View Article : Google Scholar
|
|
39
|
Shoji M, Oskowitz A, Malone CD, Prockop DJ
and Pochampally R: Human mesenchymal stromal cells (MSCs) reduce
neointimal hyperplasia in a mouse model of flow-restriction by
transient suppression of anti-inflammatory cytokines. J Atheroscler
Thromb. 18:464–474. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu Q, Fang T, Lang H, Chen M, Shi P, Pang
X and Qi G: Comparison of the proliferation, migration and
angiogenic properties of human amniotic epithelial and mesenchymal
stem cells and their effects on endothelial cells. Int J Mol Med.
39:918–926. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Friedewald WT, Levy RI and Fredrickson DS:
Estimation of the concentration of low-density lipoprotein
cholesterol in plasma, without use of the preparative
ultracentrifuge. Clin Chem. 18:499–502. 1972.PubMed/NCBI
|
|
42
|
Frost PH and Havel RJ: Rationale for use
of non-high-density lipoprotein cholesterol rather than low-density
lipoprotein cholesterol as a tool for lipoprotein cholesterol
screening and assessment of risk and therapy. Am J Cardiol.
81:26B–31B. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
44
|
Trusler O, Huang Z, Goodwin J and Laslett
AL: Cell surface markers for the identification and study of human
naive pluripotent stem cells. Stem Cell Res. 26:36–43. 2018.
View Article : Google Scholar
|
|
45
|
Fernandez Vallone VB, Romaniuk MA, Choi H,
Labovsky V, Otaegui J and Chasseing NA: Mesenchymal stem cells and
their use in therapy: What has been achieved? Differentiation.
85:1–10. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Murray RZ and Stow JL: Cytokine secretion
in macrophages: SNAREs, Rabs, and membrane trafficking. Front
Immunol. 5:5382014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liu Z, Han Y, Li L, Lu H, Meng G, Li X,
Shirhan M, Peh MT, Xie L, Zhou S, et al: The hydrogen sulfide
donor, GYY4137, exhibits anti-atherosclerotic activity in high fat
fed apolipo-protein E(−/−) mice. Br J Pharmacol. 169:1795–1809.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Fisher EA: Regression of atherosclerosis:
The journey from the liver to the plaque and back. Arterioscler
Thromb Vasc Biol. 36:226–235. 2016. View Article : Google Scholar
|
|
49
|
Lu X: Impact of macrophages in
atherosclerosis. Curr Med Chem. 23:1926–1937. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Feig JE, Parathath S, Rong JX, Mick SL,
Vengrenyuk Y, Grauer L, Young SG and Fisher EA: Reversal of
hyperlipidemia with a genetic switch favorably affects the content
and inflammatory state of macrophages in atherosclerotic plaques.
Circulation. 123:989–998. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Potteaux S, Gautier EL, Hutchison SB, van
Rooijen N, Rader DJ, Thomas MJ, Sorci-Thomas MG and Randolph GJ:
Suppressed monocyte recruitment drives macrophage removal from
athero-sclerotic plaques of Apoe−/− mice during disease
regression. J Clin Invest. 121:2025–2036. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Fang SM, Du DY, Li YT, van Rooijen N,
Rader DJ, Thomas MJ, Sorci-Thomas MG and Randolph GJ: Allogeneic
bone marrow mesenchymal stem cells transplantation for stabilizing
and repairing of atherosclerotic ruptured plaque. Thromb Res.
131:e253–e257. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yao Y, Huang J, Geng Y, Qian H, Wang F,
Liu X, Shang M, Nie S, Liu N, Du X, et al: Paracrine action of
mesenchymal stem cells revealed by single cell gene profiling in
infarcted murine hearts. PLoS One. 10:e01291642015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Togel F, Weiss K, Yang Y, Hu Z, Zhang P
and Westenfelder C: Vasculotropic, paracrine actions of infused
mesenchymal stem cells are important to the recovery from acute
kidney injury. Am J Physiol Renal Physiol. 292:F1626–F1635. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Perek B, Kowalska K, Kempisty B, Nowicki
A, Jankowski M, Nawrocki MJ and Malińska A: Role of macrophages in
the pathogenesis of atherosclerosis and aortocoronary graft
disease. J Biol Regul Homeost Agents. 32:1055–1059. 2018.PubMed/NCBI
|
|
56
|
Ding Y, Huang L, Xian X, Yuhanna IS,
Wasser CR, Frotscher M, Mineo C, Shaul PW and Herz J: Loss of
Reelin protects against atherosclerosis by reducing
leukocyte-endothelial cell adhesion and lesion macrophage
accumulation. Sci Signal. 9:ra292016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Stewart CR, Stuart LM, Wilkinson K, van
Gils JM, Deng J, Halle A, Rayner KJ, Boyer L, Zhong R, Frazier WA,
et al: CD36 ligands promote sterile inflammation through assembly
of a toll-like receptor 4 and 6 heterodimer. Nat Immunol.
11:155–161. 2010. View Article : Google Scholar :
|
|
58
|
Okabe Y and Medzhitov R: Tissue-specific
signals control reversible program of localization and functional
polarization of macrophages. Cell. 157:832–844. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yamamoto Y and Gaynor RB: IkappaB kinases:
Key regulators of the NF-kappaB pathway. Trends Biochem Sci.
29:72–79. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Putra A, Ridwan FB, Putridewi AI, Kustiyah
AR, Wirastuti K, Sadyah NAC, Rosdiana I and Munir D: The role of
TNF-α induced MSCs on suppressive inflammation by increasing TGF-β
and IL-10. Open Access Maced J Med Sci. 6:1779–1783. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Maiti P, Peruzzaro S, Kolli N, Andrews M,
Al-Gharaibeh A, Rossignol J and Dunbar GL: Transplantation of
mesenchymal stem cells overexpressing interleukin-10 induces
autophagy response and promotes neuroprotection in a rat model of
TBI. J Cell Mol Med. Jun 4;2019(Epub ahead of print). http://doi.org/10.1111/jcmm.14396urisimpledoi.org/10.1111/jcmm.14396.
|
|
62
|
Nemeth K, Leelahavanichkul A, Yuen PS,
Mayer B, Parmelee A, Doi K, Robey PG, Leelahavanichkul K, Koller
BH, Brown JM, et al: Bone marrow stromal cells attenuate sepsis via
prostaglandin E(2)-dependent reprogramming of host macrophages to
increase their interleukin-10 production. Nat Med. 15:42–49. 2009.
View Article : Google Scholar
|
|
63
|
Anderson LC: Institutional and IACUC
responsibilities for animal care and use education and training
programs. ILAR J. 48:90–95. 2007. View Article : Google Scholar : PubMed/NCBI
|