Introduction
The early stage of atherosclerotic plaque formation
is char-acterized by endothelial dysfunction and inflammation. It
has been shown that plaque development occurs at sites with
disturbed blood flow, resulting in non-uniform shear stress acting
on the endothelium (1).
Hemodynamic forces at arterial bifurcations and curvatures have a
marked impact on inflammatory gene expression in endothelial cells
(ECs) (2,3). In addition, several risk factors for
cardiovascular disease (CVD), including hypertension, diabetes
mellitus, hypercholesterol-emia and smoking, contribute to injury
and inflammation of the endothelium.
Non-uniform shear stress induces endothelial
dysfunction characterized by the expression of inflammatory
cytokines and chemokines, increased endothelial permeability and
leukocyte recruitment, and elevated expression of adhesion
molecules, including vascular cell adhesion molecule-1 (VCAM-1) and
E-selectin (4,5). By contrast, laminar shear stress in
straight vessel segments induces laminar cell alignment and a
quiescent phenotype in ECs, which prevents EC activation, and thus
has an atheroprotective effect (6).
For several years it has been known that ECs express
receptors to sense the hemodynamic status. Accordingly, the
endothelium acts as a dynamic interface between hemodynamic factors
and the vascular wall. A mechanosensory complex on the EC surface
is composed of platelet endothelial cell adhesion molecule 1
(PECAM-1), vascular endothelial cadherin (VE-cadherin) and vascular
endothelial growth factor receptor 2 (VEGFR2) transducing the
physical properties of flow into an equivalent intracellular signal
(7).
VEGFR2 is one of the three VEGF receptors, and its
activation affects vascular permeability in addition to the
proliferation, migration, differentiation and survival of ECs
(8,9). Studies using VEGFR2-knockout mice
have revealed that this receptor is indispensable for the
development and formation of a blood vessel network, leading to
fetal lethality in these mice (9,10).
The activation of VEGFR2 triggers the conformational activation of
integrins followed by the stimulation of nuclear factor-κB (NF-κB)
(11), a transcription factor
responsible not only for adaptation to flow but also for the de
novo synthesis of adhesion molecules, including VCAM-1,
E-selectin and ICAM-1. Therefore, the expression of VEGFR2
regulates endothelial activation by influencing the expression of
adhesion molecules, leading to enhanced leukocyte adhesion that
contributes to atherosclerotic plaque formation. Increased plasma
levels of VEGF, which activates VEGFR2, are associated with the
occurrence of atherosclerosis and coronary artery disease (CAD)
(12,13). In our previous study, it was shown
that the knockdown of VEGFR2 in human umbilical vein endothelial
cells (HUVECs) decreased the expression of adhesion molecules and
consequently reduced monocyte adhesion in regions of non-uniform
shear stress. This effect was mediated by inhibition of the
translocation of NF-κB from the cytoplasm to the nucleus (14).
The heritability of CVD is estimated to be 50-60%
(15). In this context, single
nucleotide polymorphisms (SNPs) have been a focus of interest for
several years (16-19). Regarding VEGFR2, which consists of
1,356 amino acids, several hundreds of SNPs have been identified
and are reported to be associated with the angiographic severity of
atherosclerosis in patients with CAD (15). In particular, a study examining
two independent populations showed that three SNPs in
VEGFR2, rs2305948 C>T, rs1870377 T>A and rs2071559
A>G, are associated with an increased risk of CAD (20), most likely due to their role in
blood vessel formation (10,21). Associations have been reported
between rs2071559 A>G and CVD; the G/G genotype is more frequent
than other genotypes in patients with type 2 diabetes and
myocardial infarction (MI) (22),
although the G allele showed a protective effect against CVD by
increasing high-density lipoprotein cholesterol in an Asian
population (23). Therefore,
evidence suggests that SNPs in the VEGF/VEGFR2 system may be
involved in the development of atherosclerosis. Furthermore, SNPs
in PECAM-1, another member of the above-mentioned mechanosensory
complex, have been shown to be associated with MI (24), atherosclerosis (25), ischemic stroke (26) and monocyte adhesion to endothelial
cells (27).
Although the sensitivity of VEGFR2 to shear stress
is well described, no information exists on the impact of the SNPs
in VEGFR2 on shear stress-induced endothelial dysfunction on a
molecular basis.
In terms of the present study, it was hypothesized
that SNPs in VEGFR2 alter the expression of genes that are
associated with endothelial cell activation (i.e., VCAM-1 and
E-selectin) under different shear stress conditions. To test this
hypothesis, the molecular responses in ECs to laminar and
non-uniform shear stress were analyzed using a well-characterized
in vitro flow chamber model. This model mimics hemodynamic
properties at vessel bifurcations comparable to physiological
conditions (4).
Materials and methods
Determination of SNPs in HUVECs
Umbilical cords were collected at the Department of
Obstetrics and Gynecology, Erlangen University Hospital,
Comprehensive Cancer Center Erlangen-EMN, Friedrich-Alexander
University Erlangen-Nürnberg (FAU; Erlangen, Germany), which was
approved by the Ethics Committee of the Medical Department of the
FAU (case no. 246-13B). The HUVECs were isolated from the freshly
collected umbilical cords using standard techniques and cultured in
endothelial cell growth medium (ECGM; Promo Cell) at 37°C in a
humidified 7.5% CO2 atmosphere. In all experiments,
HUVECs at passages 1-2 were used.
Genomic DNA was extracted from 400,000-500,000 cells
using the Promega® DNA-Isolation kit (Promega
Corporation). All DNA samples were genotyped for the three selected
SNPs in VEGFR2: rs1870377 T>A, rs2305948 C>T and
rs2071559 A>G using TaqMan® SNP genotyping assays
(Thermo Fisher Scientific, Inc.; assay nos. rs1870377:
C-11895315_20; rs2305948: C-22271999-20, and rs2071559:
C-15869271-10) with a reaction volume of 5 µl, on the
QuantStudio™ 12K Flex Real-Time PCR system following the
manufacturer's protocol. The thermocy-cling steps were programmed
as follows: Pre-read stage at 60°C for 30 sec; hold stage at 95°C
for 20 sec; 50 cycles of PCR stages at 95°C for 3 sec and at 60°C
for 30 sec; post-read stage at 25°C for 30 sec. The primers are
listed in Table I.
TaqMan® Genotyper software (Thermo Fisher Scientific,
Inc.) was used for accurate genotype calling and visualization of
the data.
 | Table ITaqMan assay probes for analysis of
vascular endothelial growth factor receptor 2 SNPs. |
Table I
TaqMan assay probes for analysis of
vascular endothelial growth factor receptor 2 SNPs.
| NCBI RefSNP ID | SNP | Mutation | Context
sequence |
|---|
| rs1870377
T>A | Exon 11 | Q472H Glu Q
(CAA)>His H (CAT) |
5′-GGTATGGGTTTGTCACTGAGACAGC[A/T]
TGGCTATAAGAAAGAGATAACAGCG-3′ |
| rs2305948
C>T | Exon 7 | I297VVal V
(GTA)>Ile I (ATA) |
5′-TACAATCCTTGGTCACTCCGGGTTA[C/T]
ACCATCTATAGTTAAGGTGCTCAAA-3′ |
| rs2071559
A>G | Intron | - |
5′-GAAAACGCACTTGCCCAGTTCGCCA[A/G]
CATTCCCGCTATTTCCCAAAATATT-3′ |
The linkage disequilibrium was analyzed to determine
whether the frequencies of the three SNPs were related to each
other by utilizing a web-based analysis tool (https://ldlink.nci.nih.gov/; LDpair, LDpop) (28,29), using genome-wide association study
(GWAS) data from the European population as the reference.
Expression levels of VEGFR2 in
HUVECs
The basal expression level of VEGFR2 was determined
semi-quantitatively. The HUVECs were harvested to extract total
protein for a bicinchoninic acid protein assay (Thermo Fisher
Scientific, Inc.) and 30-60 ng protein was used for western
blotting. The samples were separated on 8% SDS-PAGE gels and
transferred onto a PVDF membrane (Macherey-Nagel) via semi-dry
blotting. The membranes were blocked with 5% skim milk (Bio-Rad
Laboratories, Inc.) in Tris-buffered saline with 0.1% Tween (TBS-T)
or 3% bovine serum albumin (Sigma-Aldrich; Merck KGaA) and stained
with either anti-VEGFR2 antibody (cat. no. sc-505, 1:500, Santa
Cruz Biotechnology, Inc.) in 2.5% skim milk, or anti-vinculin
antibody (cat. no. sc-25336, 1:500, Santa Cruz Biotechnology, Inc.)
in TBS-T at 4°C overnight. The secondary antibodies anti-rabbit
(cat. no. RPN4301, 1:10,000) and anti-mouse (cat. no. NX931,
1:50,000) coupled to horseradish peroxidase (Amersham; GE
Healthcare Life Sciences) were incubated for 2 h at room
temperature. The signals were detected with ECL-Prime (GE
Healthcare Life Sciences) using ChemiDoc XRS (Bio-Rad Laboratories,
Inc.). The expression levels of VEGFR2 were semi-quantitatively
analyzed using ImageLab® 5.1 software (Bio-Rad
Laboratories, Inc.) with vinculin as a loading control.
Flow experiments
The HUVECs were seeded at 7×105/ml into
bifurcation flow-through slides (Integrated BioDiagnostics) and
grown until confluence. The cell monolayer was perfused with medium
for 19 h, at a flow rate of 9.6 ml/min, corresponding to a laminar
shear stress of 10.2-10.8 dyne/cm2 in the straight main
channel and a non-laminar shear stress of ~6.3 dyne/cm2
to ~0.5 dyne/cm2 in the channel distal to the
bifurcation (4). The cells were
stimulated with tumor necrosis factor (TNF)-α (2.5 ng/ml; Miltenyi
Biotec) for the final 2 h of flow.
Immunofluorescent staining
Following the flow experiment, the protein
expression of VCAM-1, E-selectin and VEGFR2 was determined by
immunocytochemical staining. The HUVECs were fixed with 4% formalin
(Roth), permeabilized with 0.2% Triton X-100 (Sigma; Merck KGaA) in
PBS, and blocked with 5% horse serum (Gibco; Thermo Fisher
Scientific, Inc.) in PBS. Staining was performed with anti-VCAM-1
(cat no. BBA5, CloneBBIG-V1, 1:100; R&D Systems, Inc.),
anti-E-selectin (cat. no. BBA16, Clone BBIG-E4, 1:100; R&D
Systems, Inc.) and anti-VEGFR2 (cat. no. sc-505, N-931; 1:100;
Santa Cruz Biotechnology, Inc.) at room temperature for 1 h and
visualized using Alexa Fluor 488-conjugated anti-mouse IgG (cat.
no. A11001) or anti-rabbit IgG (cat. no. A11008) for 45 min at room
temperature (1:500 in PBS; Molecular Probes; Thermo Fisher
Scientific, Inc.).
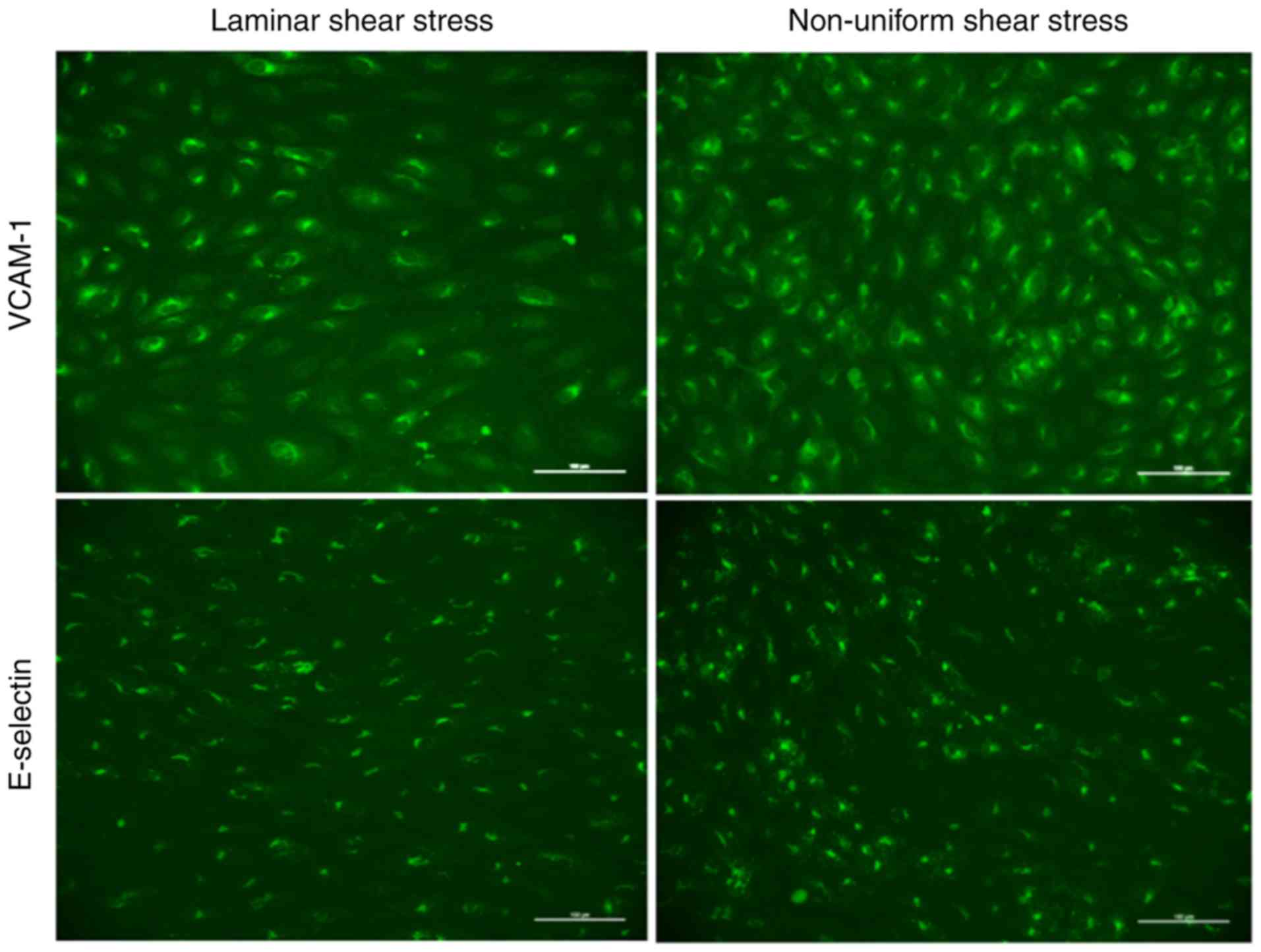
To determine the average protein expression in the
laminar and non-uniform shear stress regions within every slide,
six visual fields at ×200 magnification (0.33 mm2 each)
were selected from the laminar shear stress region and eight visual
fields from the non-uniform shear stress regions (Fig. S1) (4). Digital images were captured using an
inverted fluorescent microscope (Olympus) with image processing
software (NIS Elements® 3.2; Nikon). For every image, a
threshold was set to define the positive fluorescent signal
intensity of the entire image. Analyses were performed using
ImageJ® 1.48v software, and the corrected total cell
fluorescence (CTCF) was calculated as follows: CTCF = integrated
density - (area of selected cell × mean fluorescence of background)
(30).
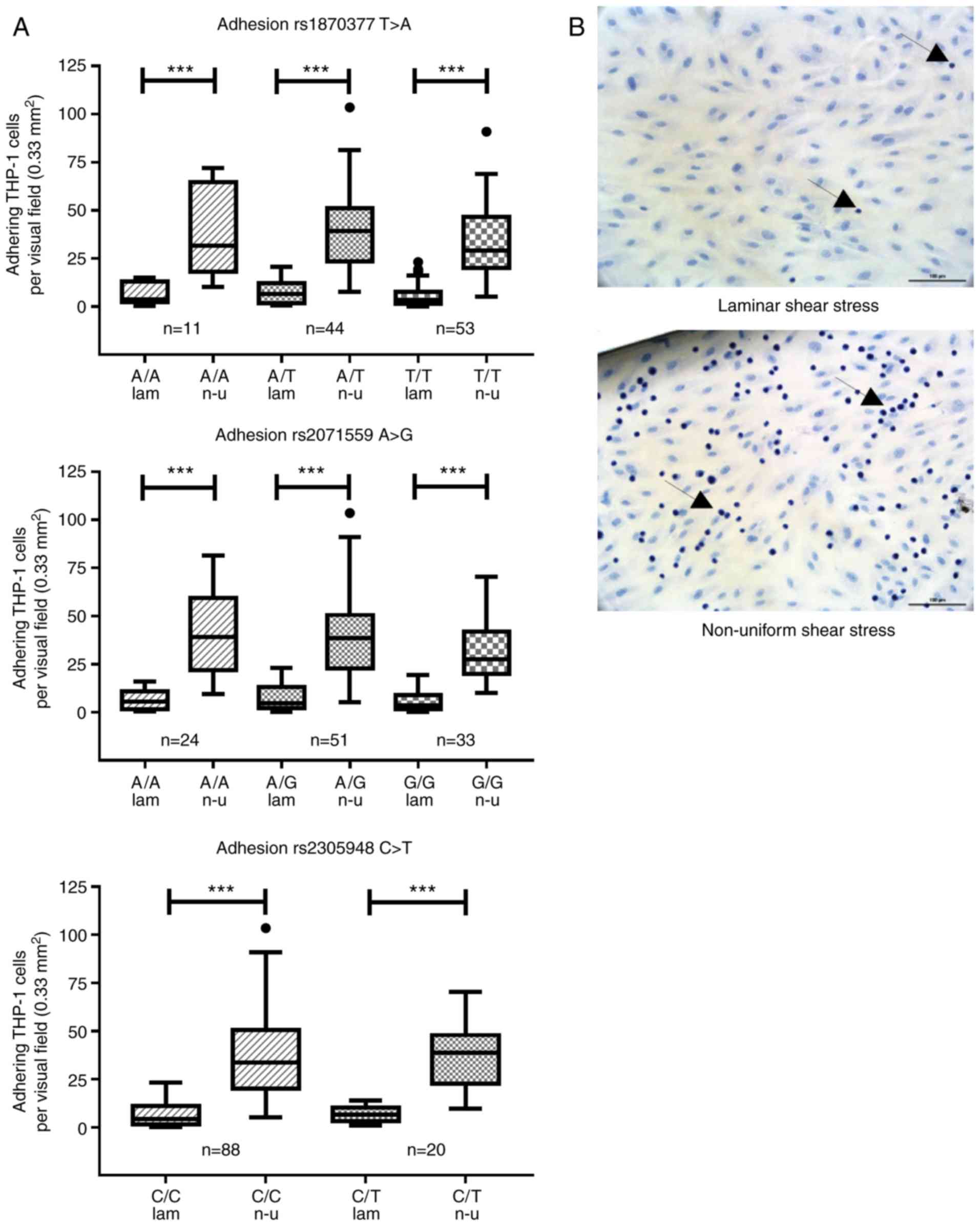
Adhesion assay
Adhesion assays were performed as described in
previous studies (5,31). THP-1 monocytic cells (American
Type Culture Collection; cat. no. TIB-202) were cultured in RPMI
1640 medium supplemented with 10% fetal calf serum, 2 mmol/l
glutamine (Biochrom AG), 100 U/ml penicillin and 100 µg/ml
streptomycin (both antibiotics from Gibco; Thermo Fisher
Scientific, Inc.). The THP-1 cells used for adhesion assays were in
passages 5-25. Following TNF-α stimulation (2.5 ng/ml), the HUVECs
were perfused with 7,500,000 THP-1 monocytic cells for 1 h at 37°C
in ECGM. Subsequently, any non-adhering THP-1 cells were flushed
away with ECGM. The HUVECs with adhering THP-1 cells were fixed
with 4% formalin, treated with 0.2% Triton X-100 in PBS and stained
with hematoxylin and eosin, which stains nuclei dark blue and the
cytoplasm light red. The distinction between HUVECs and THP-1 cells
was clear using this staining (Fig.
1B). For every slide, six visual fields at ×200 magnification
(0.33 mm2) were selected from the laminar shear stress
region and eight visual fields were selected from the non-uniform
shear stress regions. Digital images were captured using an
inverted microscope (Olympus) with image processing software and
the numbers of THP-1 cells per visual field were counted (NIS
Elements® 3.2; Nikon).
Statistical analysis
Statistical analyses were conducted using
SigmaPlot® 12.3 (Systat Software, Inc). The
Hardy-Weinberg equilibrium (HWE) of the occurrence and distribution
in sex for three SNPs were assessed using Fisher's exact test or
the χ2 test, respectively. Differences between the
laminar and non-uniform shear stress regions were analyzed using
the Wilcoxon Signed Rank test. Among the three genotype groups,
differences in adhesion molecule expression, the expression of
VEGFR2 and the number of adhered THP-1 cells were compared using
Kruskal-Wallis one-way ANOVA on ranks with a post hoc test
(Bonferroni). P<0.05 was considered to indicate a statistically
significant difference.
Results
Characteristics of the observed
HUVECs
The HUVECs were collected from 113 individuals,
comprising 61 men (54%) and 52 women (46%). The distribution of
genotypes regarding the three SNPs was in accordance with the
Hardy-Weinberg equilibrium, and the allele frequencies were similar
to those in former studies (Fig.
S2). There was no difference in the ratio of men to women among
genotypes for all SNPs. No significant correlations were observed
between any of the genotypes and being male, which is a
cardiovascular risk factor (Table
II). The homozygous T/T genotype in rs2305948 C>T was
excluded from further analyses due to its sample size being too
small (Fig. S2 and Table II). Even in a T allele dominant
model, comparing the C/C genotype with the C/T + T/T genotypes
revealed no significant differences in the protein expression of
VEGFR2, VCAM-1 or E-selectin (data not shown).
| Table IISNP characteristics. |
Table II
SNP characteristics.
| SNP | Genotype (n) | Sex, n (%)
| P-valuea | Allele
frequency | Global MAFb |
|---|
| Male | Female |
|---|
| rs1870377
T>A | AA (11) | 5 (45.5) | 6 (54.5) | 0.790 | | |
| AT (44) | 24 (54.5) | 20 (45.5) | | A=0.297 | A=0.212 |
| TT (58) | 33 (56.9) | 25 (43.1) | | T=0.703 | |
| rs2071559
A>G | AA (33) | 20 (60.6) | 13 (39.4) | | | |
| AG (53) | 25 (47.2) | 28 (52.8) | 0.235 | A=0.536 | A=0.499 |
| GG (26) | 17 (65.4) | 9 (34.6) | | G=0.464 | |
| rs2305948
C>T | CC (90) | 51 (56.7) | 39 (43.3) | | | |
| CT (21) | 10 (47.6) | 11 (52.4) | 0.612c | C=0.885 | T=0.153 |
| TT (1) | 1 (100) | 0 (0) | | T=0.115 | |
The results of the linkage analyses revealed that
there was no disequilibrium among the three analyzed SNPs compared
with the GWAS data of a European population; therefore, these SNPs
were considered as independent variables (28,29).
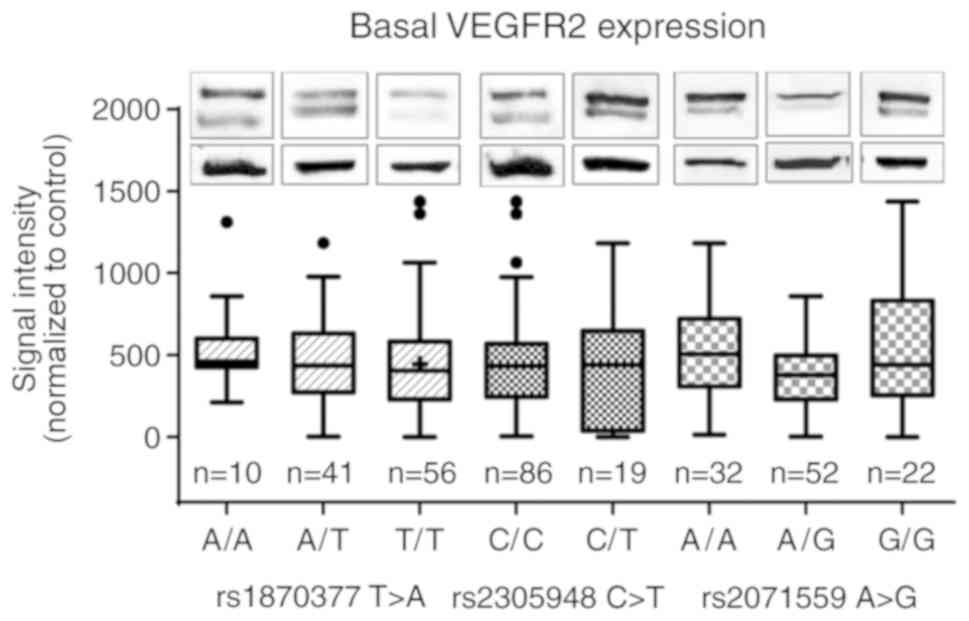
Basal expression of VEGFR2 is not
affected by SNPs in VEGFR2
The basal expression levels of VEGFR2 under static
conditions were semi-quantitatively evaluated by western blotting.
The results revealed no significant differences among the three
genotypes, although the highest median expression of VEGFR2 was
observed in A/A in rs2071559 A>G compared with that in the other
two genotypes (Fig. 2; one-way
ANOVA on ranks 0.096).
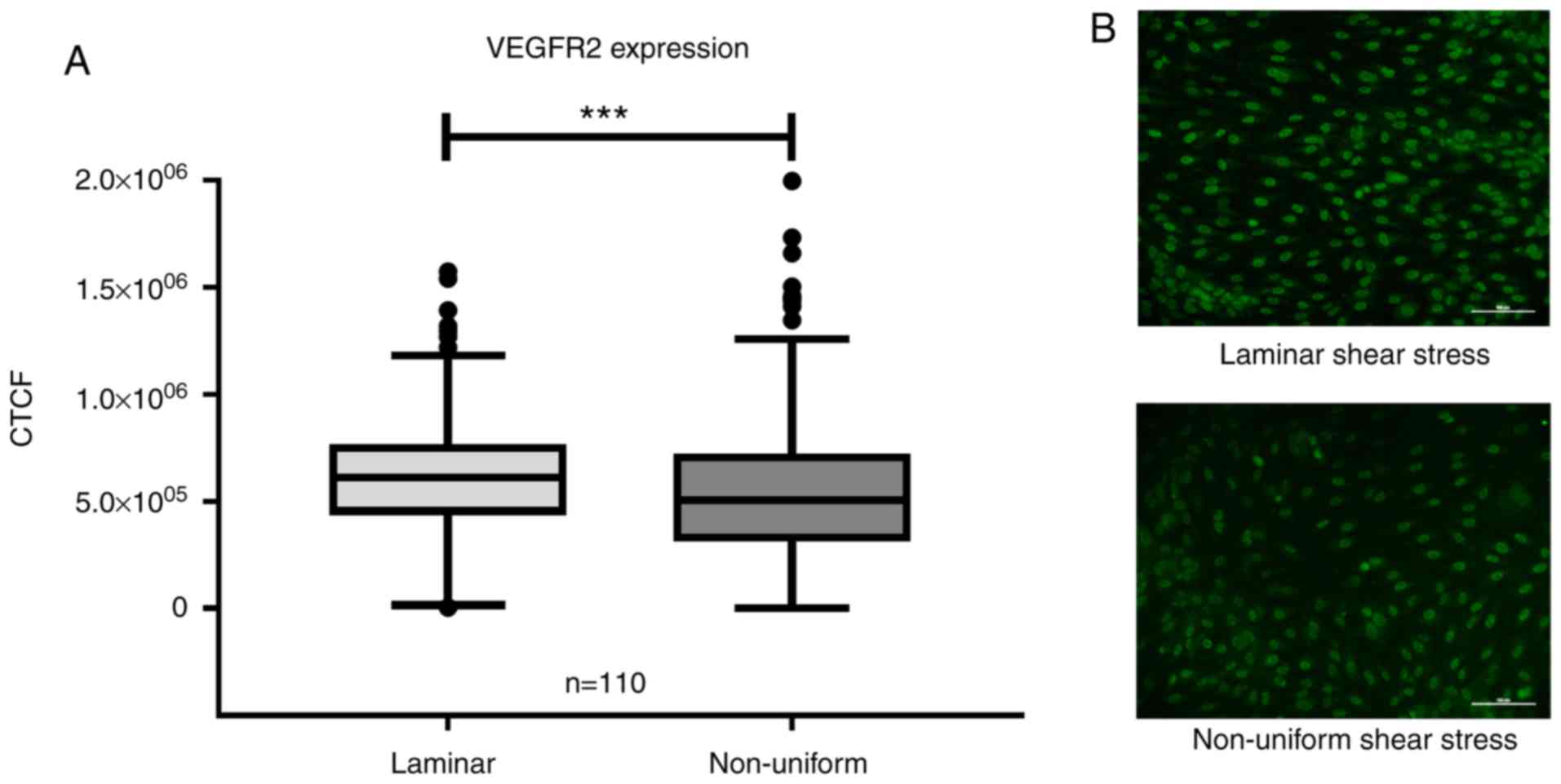
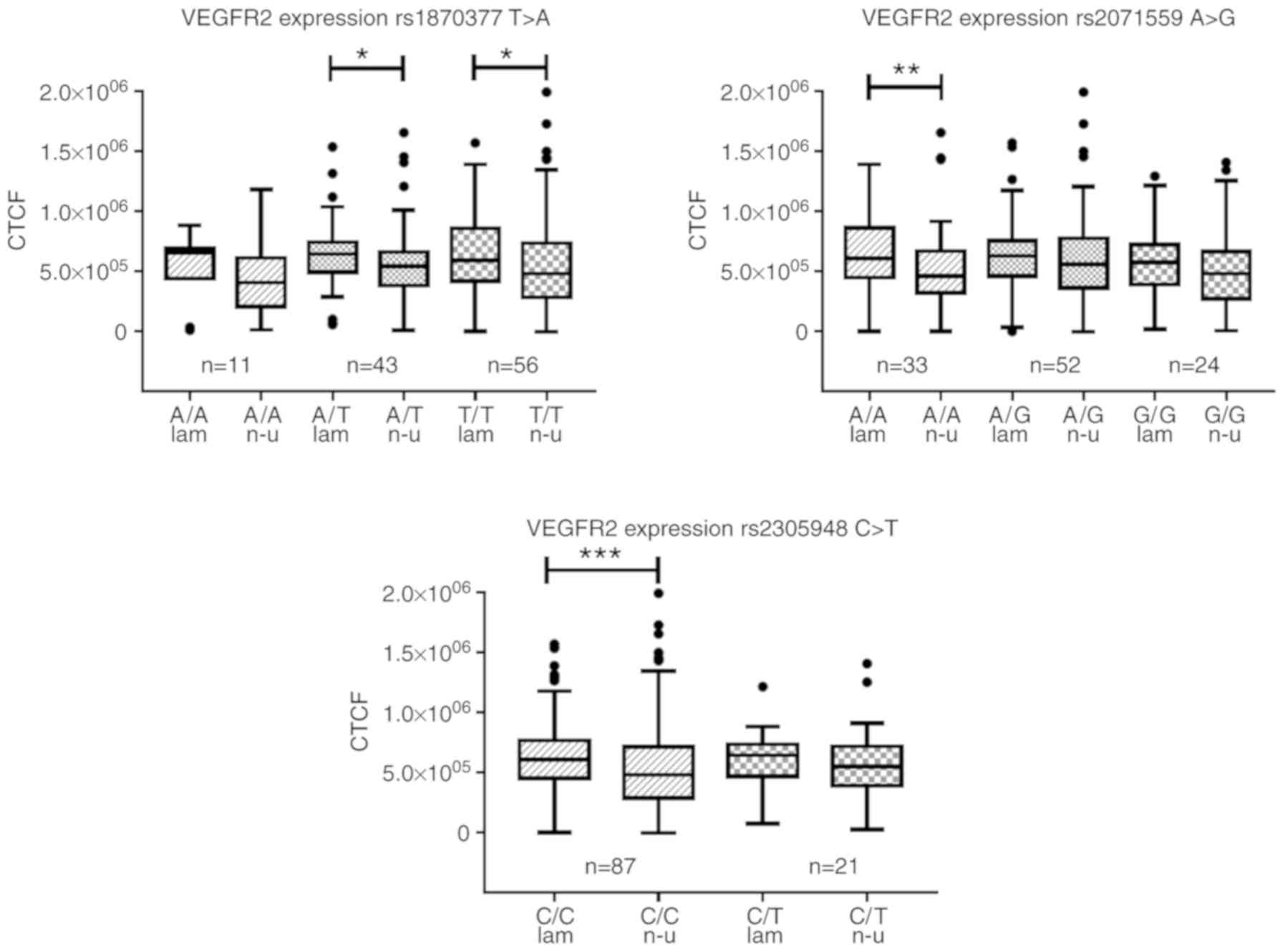
Flow-induced expression of VEGFR2 is not
affected by SNPs
The expression levels of VEGFR2 were evaluated in
HUVECs cultured under non-uniform shear stress and laminar shear
stress for 19 h. The collective VEGFR2 expression data from samples
with any genotype are shown in Fig.
3, grouped according to flow type (Fig. 3A). In regions of non-uniform shear
stress, the HUVECs expressed significantly lower levels of VEGFR2
compared with that in regions of atheroprotective laminar shear
stress (P<0.001; Fig. 3A),
independent of the genotypes of any the SNPs. There was no
significant difference in expression of VEGFR2 under either laminar
or non-uniform shear stress among the three genotypes of any the
SNPs (Fig. 4).
Expression of endothelial cell activation
markers is affected by SNPs in VEGFR2
The HUVECs were cultured under non-uniform or
laminar shear stress conditions and stimulated with TNF-α. The
protein expression levels of adhesion molecules (i.e., VCAM-1 and
E-selectin, markers for endothelial activation) were quantified by
means of fluorescent immuno-cytochemical staining (Fig. 5). As expected, the expression of
VCAM-1 was significantly higher in the non-uniform regions compared
with that in laminar shear stress regions in all data-sets analyzed
using the Wilcoxon Signed Rank test.
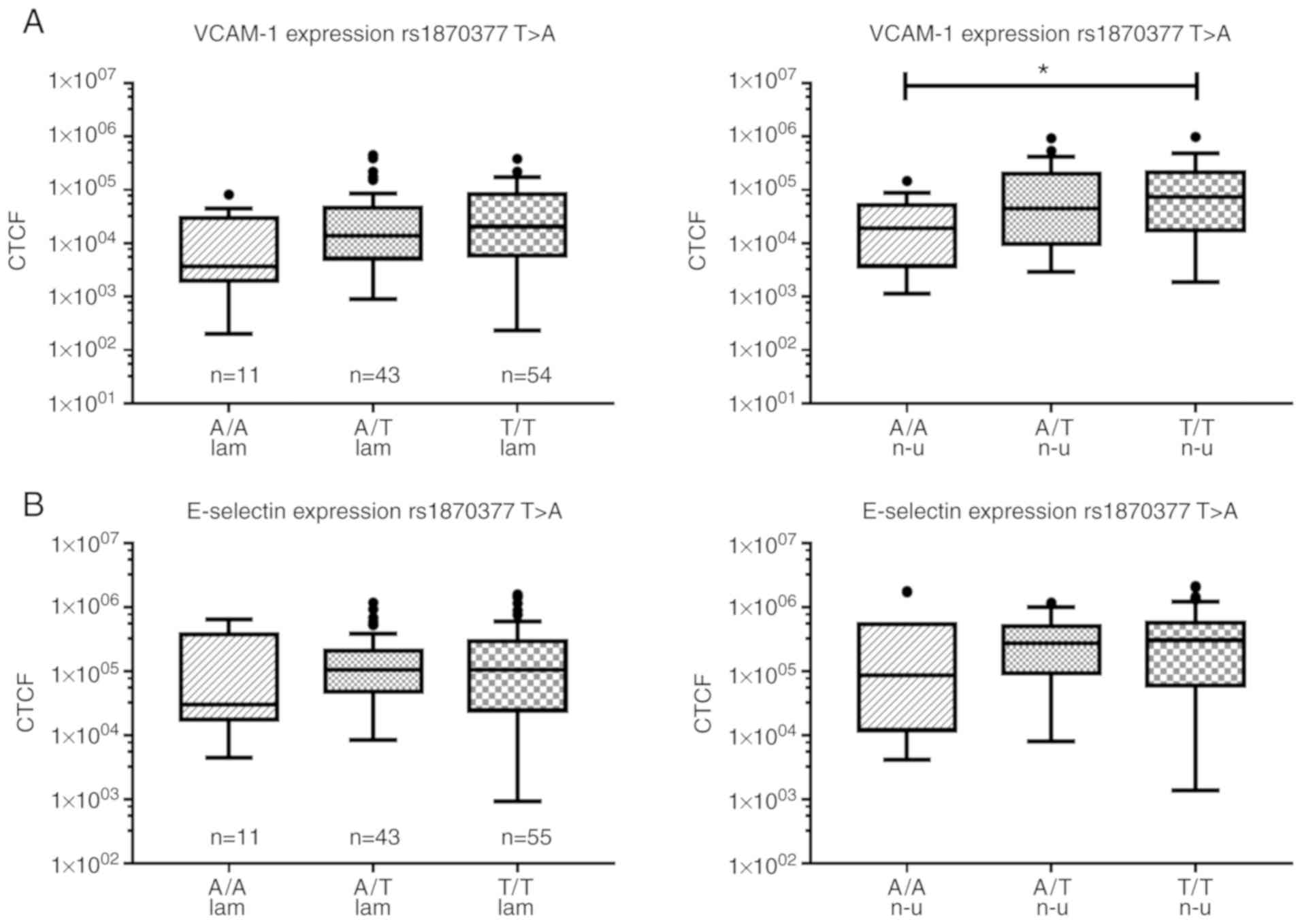
In rs1870377 T>A, significant differences were
observed between the different genotypes: Under laminar shear
stress, the highest median expression of VCAM-1 was observed in
T/T, followed by a lower expression in T/A (67% of T/T), and the
lowest expression in A/A (18% of T/T) genotypes. This showed an
increased expression of VCAM-1 with increasing content of the T
allele (one-way ANOVA on Ranks 0.052). This trend was strengthened
by non-uniform shear stress conditions; the expression of VCAM-1
was significantly lower in the A/A genotype (74% reduction) than
that in the T/T genotype (one-way ANOVA on Ranks 0.023), with
intermediate expression in the heterozygous A/T (40% reduction)
genotype (Fig. 6A). A tendency of
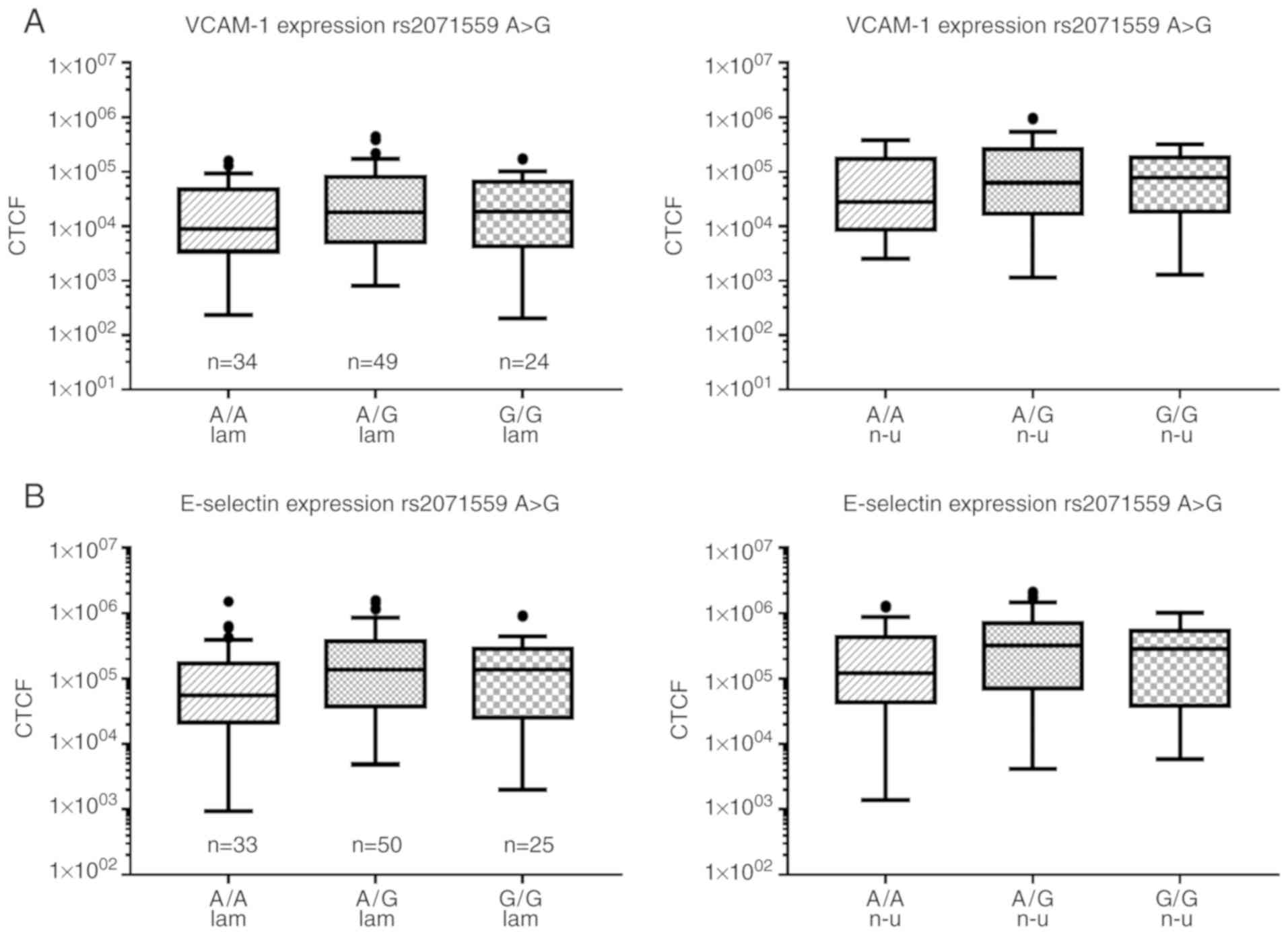
increased median expression of VCAM-1 with increasing G-content was
observed in rs2071559 A>G, but the differences were not
statistically significant (52% reduction in A/A under laminar flow
pattern and 63% under non-uniform shear stress; Fig. 7A). Furthermore, no significant
differences in VCAM-1 were observed among the different genotypes
in rs2305948 C>T (data not shown).
Similarly, significant flow-dependent differences
were observed in the expression of E-selectin. With the exception
of the A/A genotype of rs1870377 T>A, laminar shear stress
regions showed a significant reduction in E-selectin compared with
non-uniform shear stress regions (Fig. 5). Comparable to the expression of
VCAM-1, the lowest expression of E-selectin in rs1870377 T>A was
in the A/A genotype (~70% reduction) under laminar and non-uniform
shear stress conditions, although the difference was not
statistically significant (Fig.
6B). In rs2071559 A>G, the A/A genotype showed similar
expression to VCAM-1, with the lowest expression of E-selectin
under laminar (~60% reduction) and non-uniform (57% reduction)
shear stress conditions (one-way ANOVA on Ranks, 0.093/0.121)
compared with that in the G/G genotype (Fig. 7B). No significant differences were
observed in rs2305948 C>T (data not shown).
SNPs do not influence the adhesion of
THP-1 monocytic cells to an endothelial cell monolayer under flow
conditions
The adhesion of THP-1 monocytic cells was analyzed
in an in vitro flow model after 2 h of TNF-α stimulation. As
expected, the adhesion of THP-1 cells significantly increased in
regions of non-uniform shear stress in all analyzed SNPs compared
with that in atheroprotective laminar shear stress conditions,
although no significant differences in THP-1 adhesion were observed
among the distinct genotypes in any of the SNPs (Fig. 1).
Discussion
The aim of the present study was to investigate the
association between SNPs in VEGFR2 and atherogenesis on a cellular
basis. To date, only a small number of studies have investigated a
possible influence of SNPs in VEGFR2 on the occurrence of
atherosclerosis or associated complications, such as myocardial
infarction or stroke. No previous studies have addressed the
cellular mechanism of the first step of atherogenesis in
endothelial dysfunction. Therefore, the present study performed
flow simulation experiments, in which regions of high laminar shear
stress and non-uniform shear stress were readily differentiated
within an in vitro flow chamber model. The region of laminar
shear stress has a strictly laminar flow pattern, and these regions
were identified throughout the straight main channel, whereas
regions of non-uniform shear stress were located distal to the
bifurcation, characterized by a reduced flow rate and perturbed
shear pattern with a steep shear stress gradient in a direction
transversal to the flow (4).
Quiescent growth factor receptors in the absence of
their ligands are preferentially localized on the plasma membrane
(32,33). Ligand binding and receptor
activation in the majority of cases leads to receptor
internalization (34). Growth
factor receptors, such as VEGFR2, are either recycled back to the
plasma membrane or degraded, via ubiquitinylation by c-Cbl
(35), following signal
transduction (36). Only 60% of
VEGFR2 molecules are located at the plasma membrane, with the
remaining 40% localized in endosomes (37). The process of internalization may
be critical for protecting VEGFR2 against shedding (38). The internalized VEGFR2 interacts
with the transcription factor Sp1, which is known to be implicated
in angiogenesis. It has also been shown that nuclear VEGFR2 is
phosphorylated in proliferating cells. Furthermore, hypoxic
stimulation increases the nuclear localization of total VEGFR2,
whereas phosphorylated VEGFR2 remains predominantly perinuclear
(39-41). Additionally, Domingues et
al showed that nuclear VEGFR2 can act as a transcription factor
regulating its own expression (42).
The phosphorylation and subsequent activation of
VEGFR2 by fluid shear stress are dependent on PECAM-1 and
VE-cadherin via several adapter molecules (43). Shay-Salit et al observed a
rapid nuclear translocation of VEGFR2 following the onset of
laminar flow, in addition to a quiescent mechano-sensory complex
during sustained laminar flow conditions (44). In the present study, the results
obtained by immunofluorescence in the model of different flow
conditions revealed an overall decreased expression of VEGFR2 in
regions of non-uniform shear stress, regardless of genotype.
However, whether the receptor was degraded or internalized was not
elucidated.
Rs1870377 T>A and rs2305948 C>T are located in
the extracellular region at the third and fifth Ig domain. This
affects the primary protein structure and its ligand binding
properties due to polymorphisms of the amino acids (Table II). An increased risk of CVD for
carriers of the A/A genotype of rs1870377 T>A has been
demonstrated in case-control studies, arguing that the SNP impairs
VEGF binding efficiency to its receptor due to conformational
changes of VEGFR2 (20).
Rs2071559 A>G is located at the VEGFR2 promotor
region and leads to structural alterations of the binding site for
the transcription factor E2F, which possibly leads to alterations
in the expression of VEGFR2. Wang et al hypothesized that
the polymorphisms lead to a conformational change, thereby
influencing the binding affinity of VEGFR2 to VEGF, leading to
dysfunctional signal transduction (20). However, the present study did not
analyze VEGF receptor binding. In the population examined, the
basal expression of VEGFR2 in HUVECs showed no significant
differences in any of the SNPs. A comparison of the basal
expression of VEGFR2 in in vitro endothelial cells with an
in vivo study of serum levels in patients should be
considered with caution.
Kariž and Petrovič showed a higher risk for
myocardial infarction (MI) for a genotype with reduced levels of
VEGFR2 (i.e., G/G of rs2071559 A>G) (22) and others have suggested that the
reduced expression of VEGFR2 is associated with a higher risk
profile for CVD (20), which is
in line with the observation in the present study of reduced
expression of VEGFR2 under non-uniform shear stress conditions. The
results indicated that a decreased signal of VEGFR2 may be one
factor underlying the pathologic conditions of non-uniform shear
stress.
In further experimental settings, the present study
analyzed the expression of adhesion molecules, in addition to THP-1
monocyte adhesion, focusing on the different genotypes within the
VEGFR2 SNPs. Our previous study described the direct impact of the
expression of VEGFR2 on the expression of adhesion molecules in
HUVECs under non-uniform shear stress (14). A direct link between polymorphisms
in VEGFR2 and the activation and expression of adhesion molecules
has not been discussed in former studies. However, VEGFR2 is
associated with adhesion molecule expression, as stimulation of
VEGFR2 can induce the phosphoinositide 3-kinase (PI3K)-Akt and
NF-κB pathway (11,45,46), which in turn leads to the de
novo synthesis of molecules, including VCAM-1 and
E-selectin.
From the results of the present study, the presence
of endothelial dysfunction in an in vitro bifurcation model
was confirmed; the A/A genotype for SNP rs1807377 T>A was
identified as having a significantly lower adhesion molecule
expression. Therefore, this genotype may attenuate endothelial
dysfunction in atherosclerosis-prone regions and may have a
protective effect against the development of atherosclerosis and
CVD. This is in line with reports from two Chinese groups: In
rs1870377 T>A, the A allele leads to reduced vulnerability to
CVD, whereas the T allele may have the opposite effect (47,48), particularly when considering the
haplotypes. However, contradictory results have also been reported
in reports: A subpopulation of patients with diabetes mellitus type
2 had a reduced risk of CVD with the T allele, and another
subpopulation of patients who smoke, as the classical risk factor
for CVD, also had a significant reduction in risk of CVD with the T
allele, compared with the in non-smokers (47,48). These contradictory results within
one study indicate the difficulties in interpreting the results of
SNP analyses for a disease like atherosclerosis with a wide variety
of influencing factors, for example, genetic predisposition,
comorbidity factors and lifestyle.
The G/G genotype of rs2071559 A>G showed
increased median expression of VCAM-1 and E-selectin, leading to
increased endothelial activation, thereby supporting the conclusion
that the G allele of the SNP may be a risk allele for CVD. Kariž
and Petrovič found a significant correlation between MI and the G/G
genotype in rs2071559 A>G in patients with type 2 diabetes, with
a 1.6-fold higher risk of MI in G/G carriers (22). Therefore, the results from Kariž
and Petrovič and the present study do not agree with the results
from Li et al, which showed a reduced risk of CVD for
carriers of the G allele in rs2071559 A>G compared with carriers
with the corresponding A allele (47), and those of Zhang et al,
which found a negative correlation between rs2071559 A>G and
intima-media thickness for G allele samples (21).
The third SNP analyzed in the present study was
rs2305948 C>T. Again, the results from previous studies
analyzing potential associations of this SNP with CVD are
conflicting. Zhang et al reported that the T allele was
associated with increased risk for stroke (21), and Li et al confirmed this
from analyzing data of a Chinese population adjusted for
cardiovascular risk factors (47). Additionally, Liu et al
analyzed this SNP regarding the risk for CVD in a Han Chinese
population. Without adjustment for additional risk factors,
individuals with the C allele showed a significantly higher risk
for CVD. However, following regression analyses for the individual
risk factors, such as smoking, diabetes, hypertension or alcohol
abuse, the C allele was shown to be protective against CVD in
smoking and non-smoking patients. The individual analyses of
patients with and without hypertension, and of patients with or
without diabetes, also showed a protective effect of the C allele
against CVD (48). As the T/T
genotype was rare, with only one sample, it was not possible to
compare the two divergent homozygous genotypes in the present
study, where the most pronounced differences were expected. The
comparison between C/C and C/T showed no significant difference in
any of the experimental settings.
Regarding discrepancies between former studies and
the present study, it is important to consider that the in
vitro model used in the present study simulates the early
events of athero-genesis. This may not allow a direct comparison
with the cited association studies in fully developed CVD having
additional risk factors. Atherosclerosis and CVD are complex
diseases with a large number of subtle contributing factors that
may influence the severity or outcome in different patients.
Considering that inherited factors, genetic markers and epigenetics
influence the diverse regulation of VEGFR2 and endothelial cell
homeostasis, the comparability of results in this field of research
is impeded. Taken together, compared with most clinical
case-control studies, the set of 113 samples in the present study
was relatively small. Furthermore, due to the anonymization of the
samples, information about the donors, including ethnic background,
age of the mother, complications at birth, or any accompanying
diseases, are missing. The majority of subjects in the present
study were reportedly central Europeans. Compared with an East
Asian population, a European population appears to have a higher
frequency of T in rs1870377 T>A and C in rs2305948 C>T
(49).
As the results of the present study are conflicting
with those of former studies in a Chinese population (16,48), there may be variations in gene
expression due to a variety of factors, including ethnicity, which
affected the results obtained (50). This may explain the large
variability within the population; and despite a large proportional
reduction of adhesion molecule expression, statistical significance
was not detected.
In conclusion, the present study provides new
evidence that the three investigated SNPs may offer potential as
novel risk markers for CVD and warrant further investigation in the
pathogenesis of atherosclerosis and CVD. The A/A genotype of
rs1870377 A>T appeared to be atheroprotective, whereas the G/G
genotype of rs2071559 A>G may enhance endothelial dysfunction.
The discrepant findings between the observations in the present
study at a cellular level and those from clinical studies suggest
that any genetic polymorphism would be strongly affected by
additional risk factors for CVD, including type 2 diabetes,
hypertension, hypercholesterolemia and smoking. In addition, SNPs
may influence collateral development not only individually but also
when acting together with other SNPs, through gene haplotype
networks.
Supplementary Data
Abbreviations:
|
CAD
|
coronary artery disease
|
|
CVD
|
cardiovascular disease
|
|
EC
|
endothelial cell
|
|
HUVECs
|
human umbilical vein endothelial
cells
|
|
HWE
|
Hardy-Weinberg equilibrium
|
|
MI
|
myocardial infarction
|
|
NF-κB
|
nuclear factor-κB
|
|
PECAM-1
|
platelet endothelial cell adhesion
molecule-1
|
|
PI3K
|
phosphoinositide 3-kinase
|
|
SNP
|
single nucleotide polymorphism
|
|
TNF-α
|
tumor necrosis factor-α
|
|
VCAM-1
|
vascular cell adhesion molecule-1
|
|
VEGFR2
|
vascular endothelial growth factor
2
|
Acknowledgments
The authors would like to thank Mrs. Jaziri, Mrs.
Flick and Mrs. Kloos (Laboratory for Molecular and Experimental
Cardiology, Department of Medicine-2, University Hospital Erlangen)
for their technical support in this study, and Mrs. Zwenger
(Institute of Human Genetics, University Hospital Erlangen) for SNP
sequencing.
Funding
The present study was funded by the ELAN fund,
Friedrich-Alexander-Universität Erlangen-Nürnberg (grant. no.
ELAN-13-1171-Urschel). The study was designed, conducted and
written independently from the funding institution.
Availability of data and materials
The datasets are available from the corresponding
author upon reasonable request.
Authors' contributions
KU and BD conceived and designed the study. NMS and
KU collected and analyzed the data. KU and NMS wrote the
manuscript. MT and DRS substantively revised the researched
articles and were involved in the interpretation of results. FMS
provided the umbilical cords and contributed to the interpretation
of results. SA revised the results from a clinicians view, thereby
being involved in analyses and interpretation of the results in the
specific professional context. SA, BD and MT critically revised the
manuscript for important intellectual content. FP was involved in
data collection and interpretation of the results. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
The collection of human material and the study
protocol were approved by the Ethics Committee of the Medical
Department of the Friedrich-Alexander-Universität Erlangen-Nürnberg
(case no. 246_13B). Written consent was obtained from all women
prior to donation.
Patient consent for participation
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Slager CJ, Wentzel JJ, Gijsen FJ,
Schuurbiers JC, van der Wal AC, van der Steen AF and Serruys PW:
The role of shear stress in the generation of rupture-prone
vulnerable plaques. Nat Clin Pract Cardiovasc Med. 2:401–407. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li YS, Haga JH and Chien S: Molecular
basis of the effects of shear stress on vascular endothelial cells.
J Biomech. 38:1949–1971. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wasserman SM, Mehraban F, Komuves LG, Yang
RB, Tomlinson JE, Zhang Y, Spriggs F and Topper JN: Gene expression
profile of human endothelial cells exposed to sustained fluid shear
stress. Physiol Genomics. 12:13–23. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cicha I, Beronov K, Ramirez EL, Osterode
K, Goppelt-Struebe M, Raaz D, Yilmaz A, Daniel WG and Garlichs CD:
Shear stress preconditioning modulates endothelial susceptibility
to circulating TNF-alpha and monocytic cell recruitment in a
simplified model of arterial bifurcations. Atherosclerosis.
207:93–102. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Urschel K, Worner A, Daniel WG, Garlichs
CD and Cicha I: Role of shear stress patterns in the TNF-α-induced
atherogenic protein expression and monocytic cell adhesion to
endothelium. Clin Hemorheol Microcirc. 46:203–210. 2010.
|
|
6
|
Pan S: Molecular mechanisms responsible
for the atheropro-tective effects of laminar shear stress. Antioxid
Redox Signal. 11:1669–1682. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tzima E, Irani-Tehrani M, Kiosses WB,
Dejana E, Schultz DA, Engelhardt B, Cao G, DeLisser H and Schwartz
MA: A mecha-nosensory complex that mediates the endothelial cell
response to fluid shear stress. Nature. 437:426–431. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Holmes K, Roberts OL, Thomas AM and Cross
MJ: Vascular endothelial growth factor receptor-2: Structure,
function, intracellular signalling and therapeutic inhibition. Cell
Signal. 19:2003–2012. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shalaby F, Rossant J, Yamaguchi TP,
Gertsenstein M, Wu XF, Breitman ML and Schuh AC: Failure of
blood-island formation and vasculogenesis in Flk-1-deficient mice.
Nature. 376:62–66. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fong GH, Rossant J, Gertsenstein M and
Breitman ML: Role of the Flt-1 receptor tyrosine kinase in
regulating the assembly of vascular endothelium. Nature. 376:66–70.
1995. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Y, Flores L, Lu S, Miao H, Li YS and
Chien S: Shear stress regulates the Flk-1/Cbl/PI3K/NF-κB pathway
via actin and tyrosine kinases. Cell Mol Bioeng. 2:341–350. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lim HS, Blann AD, Chong AY, Freestone B
and Lip GY: Plasma vascular endothelial growth factor,
angiopoietin-1, and angiopoietin-2 in diabetes: Implications for
cardiovascular risk and effects of multifactorial intervention.
Diabetes Care. 27:2918–2924. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Petrovic D: The role of vascular
endothelial growth factor gene as the genetic marker of
atherothrombotic disorders and in the gene therapy of coronary
artery disease. Cardiovasc Hematol Agents Med Chem. 8:47–54. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Urschel K, Garlichs CD, Daniel WG and
Cicha I: VEGFR2 signalling contributes to increased endothelial
susceptibility to TNF-α under chronic non-uniform shear stress.
Atherosclerosis. 219:499–509. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Howell W, Ali S, Rose-Zerilli M and Ye S:
VEGF polymorphisms and severity of atherosclerosis. J Med Genet.
42:485–490. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang LJ, Zhang YQ, Han X, Zhang ZT and
Zhang ZQ: Association of VEGFR-2 Gene polymorphisms with
clopidogrel resistance in patients with coronary heart disease. Am
J Ther. 23:e1663–e1670. 2016. View Article : Google Scholar
|
|
17
|
Hermann MM, van Asten F, Muether PS,
Smailhodzic D, Lichtner P, Hoyng CB, Kirchhof B, Grefkes C, den
Hollander AI and Fauser S: Polymorphisms in vascular endothelial
growth factor receptor 2 are associated with better response rates
to ranibizumab treatment in age-related macular degeneration.
Ophthalmology. 121:905–910. 2014. View Article : Google Scholar
|
|
18
|
Jain L, Sissung TM, Danesi R, Kohn EC,
Dahut WL, Kummar S, Venzon D, Liewehr D, English BC, Baum CE, et
al: Hypertension and hand-foot skin reactions related to VEGFR2
genotype and improved clinical outcome following bevacizumab and
sorafenib. J Exp Clin Cancer Res. 29:952010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim DH, Xu W, Kamel-Reid S, Liu X, Jung
CW, Kim S and Lipton JH: Clinical relevance of vascular endothelial
growth factor (VEGFA) and VEGF receptor (VEGFR2) gene polymorphism
on the treatment outcome following imatinib therapy. Ann Oncol.
21:1179–1188. 2010. View Article : Google Scholar
|
|
20
|
Wang Y, Zheng Y, Zhang W, Yu H, Lou K,
Zhang Y, Qin Q, Zhao B, Yang Y and Hui R: Polymorphisms of KDR gene
are associated with coronary heart disease. J Am Coll Cardiol.
50:760–767. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang W, Sun K, Zhen Y, Wang D, Wang Y,
Chen J, Xu J, Hu FB and Hui R: VEGF receptor-2 variants are
associated with susceptibility to stroke and recurrence. Stroke.
40:2720–2726. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kariž S and Petrovič D: Minor association
of kinase insert domain-containing receptor gene polymorphism
(rs2071559) with myocardial infarction in Caucasians with type 2
diabetes mellitus: Case-control cross-sectional study. Clin
Biochem. 47:192–196. 2014. View Article : Google Scholar
|
|
23
|
Yap RW, Shidoji Y, Hon WM and Masaki M:
Association and interaction between dietary pattern and VEGF
receptor-2 (VEGFR2) gene polymorphisms on blood lipids in Chinese
Malaysian and Japanese adults. Asia Pac J Clin Nutr. 21:302–311.
2012.PubMed/NCBI
|
|
24
|
Sahebkar A, Morris DR, Biros E and
Golledge J: Association of single nucleotide polymorphisms in the
gene encoding platelet endothelial cell adhesion molecule-1 with
the risk of myocardial infarction: A systematic review and
meta-analysis. Thromb Res. 132:227–233. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Listi F, Caruso C, Di Carlo D, Falcone C,
Boiocchi C, Cuccia M and Candore G: Association between platelet
endothelial cellular adhesion molecule-1 polymorphisms and
atherosclerosis: Results of a study on patients from northern
Italy. Rejuvenation Res. 13:237–241. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wei YS, Lan Y, Liu YG, Meng LQ, Xu QQ and
Xie HY: Platelet-endothelial cell adhesion molecule-1 gene
polymorphism and its soluble level are associated with ischemic
stroke. DNA Cell Biol. 28:151–158. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Goodman RS, Kirton CM, Oostingh GJ, Schon
MP, Clark MR, Bradley JA and Taylor CJ: PECAM-1 polymorphism
affects monocyte adhesion to endothelial cells. Transplantation.
85:471–477. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Machiela MJ and Chanock SJ: LDlink: A
web-based application for exploring population-specific haplotype
structure and linking correlated alleles of possible functional
variants. Bioinformatics. 31:3555–3557. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Machiela MJ and Chanock SJ: LDassoc: An
online tool for interactively exploring genome-wide association
study results and prioritizing variants for functional
investigation. Bioinformatics. 34:887–889. 2018. View Article : Google Scholar :
|
|
30
|
Fitzpatrick M: Measuring Cell Fluorescence
using ImageJ. Science Tech Blog. 2011, https://theolb.readthedocs.io/en/latest/imaging/measuring-cell-fluorescence-using-imagej.html.
|
|
31
|
Cicha I, Urschel K, Daniel WG and Garlichs
CD: Telmisartan prevents VCAM-1 induction and monocytic cell
adhesion to endothelium exposed to non-uniform shear stress and
TNF-α. Clin Hemorheol Microcirc. 48:65–73. 2011.
|
|
32
|
Hopkins CR, Miller K and Beardmore JM:
Receptor-mediated endocytosis of transferrin and epidermal growth
factor receptors: A comparison of constitutive and ligand-induced
uptake. J Cell Sci Suppl. 3:173–186. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Goh LK and Sorkin A: Endocytosis of
receptor tyrosine kinases. Cold Spring Harb Perspect Biol.
5:a0174592013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sorkin A and von Zastrow M: Endocytosis
and signalling: Intertwining molecular networks. Nat Rev Mol Cell
Biol. 10:609–622. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Murdaca J, Treins C, Monthouel-Kartmann
MN, Pontier-Bres R, Kumar S, Van Obberghen E and Giorgetti-Peraldi
S: Grb10 prevents Nedd4-mediated vascular endothelial growth factor
receptor-2 degradation. J Biol Chem. 279:26754–26761. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sigismund S, Confalonieri S, Ciliberto A,
Polo S, Scita G and Di Fiore PP: Endocytosis and signaling: Cell
logistics shape the eukaryotic cell plan. Physiol Rev. 92:273–366.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gampel A, Moss L, Jones MC, Brunton V,
Norman JC and Mellor H: VEGF regulates the mobilization of
VEGFR2/KDR from an intracellular endothelial storage compartment.
Blood. 108:2624–2631. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Basagiannis D and Christoforidis S:
Constitutive endocytosis of VEGFR2 protects the receptor against
shedding. J Biol Chem. 291:16892–16903. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang Y, Pillai G, Gatter K, Blazquez C,
Turley H, Pezzella F and Watt SM: Expression and cellular
localization of vascular endothelial growth factor A and its
receptors in acute and chronic leukemias: An immunohistochemical
study. Hum Pathol. 36:797–805. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Blazquez C, Cook N, Micklem K, Harris AL,
Gatter KC and Pezzella F: Phosphorylated KDR can be located in the
nucleus of neoplastic cells. Cell Res. 16:93–98. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Stewart M, Turley H, Cook N, Pezzella F,
Pillai G, Ogilvie D, Cartlidge S, Paterson D, Copley C, Kendrew J,
et al: The angiogenic receptor KDR is widely distributed in human
tissues and tumours and relocates intracellularly on
phosphorylation. An immunohistochemical study Histopathology.
43:33–39. 2003.
|
|
42
|
Domingues I, Rino J, Demmers JA, de
Lanerolle P and Santos SC: VEGFR2 translocates to the nucleus to
regulate its own transcription. PLoS One. 6:e256682011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Conway D and Schwartz MA: Lessons from the
endothelial junctional mechanosensory complex. F1000 Biol Rep.
4:12012.PubMed/NCBI
|
|
44
|
Shay-Salit A, Shushy M, Wolfovitz E, Yahav
H, Breviario F, Dejana E and Resnick N: VEGF receptor 2 and the
adherens junction as a mechanical transducer in vascular
endothelial cells. Proc Natl Acad Sci USA. 99:9462–9467. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang Y, Chang J, Li YC, Li YS, Shyy JY and
Chien S: Shear stress and VEGF activate IKK via the Flk-1/Cbl/Akt
signaling pathway. Am J Physiol Heart Circ Physiol. 286:H685–H692.
2004. View Article : Google Scholar
|
|
46
|
Jin ZG, Ueba H, Tanimoto T, Lungu AO,
Frame MD and Berk BC: Ligand-independent activation of vascular
endothelial growth factor receptor 2 by fluid shear stress
regulates activation of endothelial nitric oxide synthase. Circ
Res. 93:354–363. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li L, Pan Y, Dai L, Liu B and Zhang D:
Association of genetic polymorphisms on vascular endothelial growth
factor and its receptor genes with susceptibility to coronary heart
disease. Med Sci Monit. 22:31–40. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu D, Song J, Ji X, Liu Z, Cong M and Hu
B: Association of genetic polymorphisms on VEGFA and VEGFR2 with
risk of coronary heart disease. Medicine (Baltimore). 95:e34132016.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Medicine USNLo: SNP Database. https://www.ncbi.nlm.nih.gov/snp.
|
|
50
|
Zhang H, Zhai Q, Zhang Z, Cai B, Cai H,
Zhou S, Sun L, Xie Y, Kong D, Xu Z, et al: Association of
GWAS-Supported Variants rs556621 on Chromosome 6p21.1 with large
artery atherosclerotic stroke in a Southern Chinese Han Population.
Neuromolecular Med. 19:94–100. 2017. View Article : Google Scholar
|