Introduction
Pulmonary hypertension generally results from
hypoxic lung diseases, such as chronic obstructive pulmonary
disease (COPD), cystic fibrosis and bronchiectasis (1). Chronic hypoxia results in pulmonary
hypertension, which is characterized by progressively elevated
pulmonary arterial pressure, increased right ventricular afterload
leading to right ventricular hypertrophy, ultimately resulting in
heart failure (2,3). Pulmonary hypertension is
independently associated with increased morbidity and reduced
survival in patients suffering from hypoxic lung disease. Pulmonary
vascular remodeling accompanies and is considered to be the primary
cause of pulmonary hypertension by increasing pulmonary vascular
resistance (4).
The pulmonary vascular wall is a heterogeneous
three-layered structure composed of adventitia, media and intima.
Pulmonary artery remodeling is a complicated pathological process
involving disorders of all three layers of the vascular wall,
including adventitial thickening, medial hypertrophy, neointima
formation and plexiform lesions (5). Resident adventitial fibroblasts are
considered to be the principal sensor tissue of the vessel wall in
response to stress or injury, and they can be activated and undergo
phenotypic changes to exhibit various structural and functional
alterations (6,7). Our previous study demonstrated that
hypoxia induces functional alterations in lung fibroblasts and it
may be involved in pulmonary vascular remodeling (8). Evidence from recent studies
indicates that mast cells are also associated with non-allergic
chronic lung diseases. The presence of abundant perivascular mast
cells, as well as increased secretion of inflammatory substances,
was observed in the lungs of patients with pulmonary arterial
hypertension (PAH) and experimental models of pulmonary
hypertension (9,10). Despite evidence showing that mast
cells contribute to vascular remodeling and the development of
pulmonary hypertension under pathological conditions, the
underlying mechanism has not been fully elucidated.
The aim of the present study was to investigate the
effect of mast cells on the viability, function and phenotype of
fibroblasts under hypoxic conditions. It was hypothesized that
hypoxia-activated mast cells affect the characteristics of lung
fibroblasts and are involved in pulmonary vascular remodeling. To
test this hypothesis, the viability and secretion of cytokines by
human mast cells (HMC-1) under hypoxia were first examined.
Subsequently, the effects of hypoxic conditioned medium from HMC-1
cells on human fetal lung fibroblasts (HFL-1) were examined using
RNA sequencing (RNA-seq) analysis and molecular biology
experiments. Finally, using a rat model of hypoxic pulmonary
hypertension, the distribution of mast cells, extracellular matrix
remodeling and myofibroblast transition in the lung were
explored.
Materials and methods
Cell culture
The human mast cell line HMC-1 (Cellbio) was
cultured in Iscove's Modified Dubecco's Medium (IMDM; Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.), 100 mg/ml
streptomycin and 100 U/ml penicillin (Sigma-Aldrich; Merck KGaA).
HMC-1 cells were incubated at 37°C in a 5% CO2
atmosphere at 95% relative humidity. For hypoxic exposure, HMC-1
cells were placed in a modulator incubator in an atmosphere of 94%
N2, 5% CO2 and 1% O2, and normoxic
conditions were defined as 21% O2. Subsequently, cells
and supernatants were collected for subsequent experiments. For the
preparation of conditioned medium, the supernatants were
centrifuged for 10 min at 500 × g at room temperature to remove
detached cells and used directly. HFL-1 cells (Type Culture
Collection Center, Chinese Academy of Sciences) were cultured in
Ham's F-12K (Kaighn's) medium with 10% FBS, 100 mg/ml streptomycin
and 100 U/ml penicillin until confluent.
Cell proliferation assay
Cells at 70% confluence were serum-starved in 0.1%
FBS for 24 h for synchronization, plated at 5×103 cells
per well in 96-well plates and cultured for 6 h at 37℃ in 5%
CO2 in order to allow the cells to attach. Subsequently,
HMC-1 cells were incubated with fresh IMDM (Gibco; Thermo Fisher
Scientific, Inc.) containing 10% FBS under hypoxic conditions, and
HFL-1 cells were cultured in conditioned medium from hypoxic or
normoxic HMC-1 cells for different times. The Cell Counting Kit-8
(CCK-8, Dojindo Molecular Technologies, Inc.) was used to determine
cell proliferation at various time points. For the CCK-8 assay,
cells were incubated with 10 µl CCK-8 solution for 1 h, and
absorbance was measured at 450 nm.
Measurement of cytokines
Secreted cytokines from HMC-1 cells, including tumor
necrosis factor (TNF)-α (cat. no. KE00068, ProteinTech Group,
Inc.), interleukin (IL)-1β (cat. no. KE00021, ProteinTech Group,
Inc.), IL-6 (cat. no. KE00007, ProteinTech Group, Inc.) and IL-15
(cat. no. KE00102, ProteinTech Group, Inc.) were measured with the
use of ELISA kits. ELISA was performed according to the
manufacturer's protocol (ProteinTech Group, Inc.). Briefly, the
supernatants or standards were added to 96-well microplates and
incubated for 60 min at room temperature. After washing 3 times
with wash buffer, the corresponding detection antibodies were added
for 60 min at 37°C. The plate was washed with wash buffer, 100
µl horseradish peroxidase (HRP)-conjugated antibodies were
added and the reaction was incubated for 30 min at 37°C in a
light-resistant container. Subsequently, 100 µl
tetramethylbenzidine substrate (TMB) solution was added and
incubated for 15 min at 37°C. Finally, stop solution was added to
each well to terminate the reaction. Absorbance was calculated by
measurement at 450 nm, and the levels of cytokines were calculated
using the standard curve.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was extracted from HFL-1 and HMC-1 cells
by use of TRIzol reagent (Tiangen), and cDNA was acquired by use of
reverse transcriptase (Thermo Fisher Scientific, Inc.). qPCR with
the use of Maxima SYBR-Green I qPCR Master Mix (Thermo Fisher
Scientific, Inc.) was performed as follows: 95°C for 5 min,
followed by 35 cycles at 95°C for 30 sec, 60°C for 30 sec and 72°C
for 1 min. cDNA was amplified with the primer sequences for TNF-α
(forward 5′-ccttcctctctccagatgtttc-3′, reverse
5′-cccggtctcccaaataaataca-3′), IL-1β (forward
5′-cctg-gactttcctgttgtctac-3′, reverse
5′-aagtgagtaggagaggtgagag-3′), IL-6 (forward
5′-cagctatgaactccttctccac-3′, reverse
5′-cgtcgag-gatgtaccgaattt-3′), IL-15 (forward
5′-ccactgtgtccggaattgat-3′, reverse 5′-gaacccaccagaaggaagaaa-3′),
matrix metallopro-teinase (MMP)-9 (forward
5′-ggtaaggagtactcgacctgta-3′, reverse
5′-cggcactgaggaatgatctaag-3′), MMP-13 (forward
5′-ggcgacttc-tacccatttga-3′, reverse 5′-cttggagtggtcaagacctaag-3′),
tissue inhibitor of metalloproteinase 1 (TIMP-1; forward
5′-gatg-gtgggtggatgagtaatg-3′, reverse 5′-gggttctctggtgtctctct-3′),
collagen type I (forward 5′-cccagccaagaactggtatag-3′, reverse
5′-ggtgatgttctgagaggcatag-3′) and III (forward
5′-ttggaagtcctg-gtccaaag-3′, reverse 5′-caccaccttcacccttatctc-3′),
α-smooth muscle actin (α-SMA; forward 5′-ccgaatgcagaaggagatca-3′,
reverse 5′-gtggacagagaggccaggat-3′) and β-tubulin (forward
5′-gaggctgagagcaacatgaa-3′, reverse 5′-cagttgagtaagacg-gctaagg-3′).
For quantification, target gene expression was normalized to that
of β-actin in each sample. Data analysis involved the -ΔΔCq method
(11).
Western blot analysis
Total proteins were extracted from treated cells
using lysis buffer containing protease inhibitors (Beyotime
Institute of Biotechnology). The protein concentration of the
lysates was quantified with a bicinchoninic acid assay (Beyotime
Institute of Biotechnology). Total protein in lysates (50 µg
per lane) was separated by 10% SDS-PAGE and then transferred to
nitrocellulose membranes (Thermo Fisher Scientific, Inc.). The
membranes were incubated with primary antibodies, including
polyclonal rabbit anti-hypoxia inducible factor (HIF)-1α (dilution
1:1,000; cat. no. 20960-1-AP, ProteinTech Group, Inc.), polyclonal
rabbit anti-collagen type I (dilution 1:1,000; cat. no. 14695-1-AP,
ProteinTech Group, Inc.), polyclonal rabbit anti-collagen type III
(dilution 1:1,000; cat. no. 22734-1-AP, ProteinTech Group, Inc.),
polyclonal rabbit anti-MMP-9 (dilution 1:1,000; cat. no.
10375-2-AP, ProteinTech Group, Inc.), polyclonal rabbit anti-MMP-13
(dilution 1:1,000; cat. no. 18165-1-AP, ProteinTech Group, Inc.),
polyclonal rabbit anti-TIMP-1 (dilution 1:1,000; cat. no.
16644-1-AP, ProteinTech Group, Inc.), polyclonal rabbit anti-α-SMA
(dilution 1:1,000; cat. no. 55135-1-AP, ProteinTech Group, Inc.) or
polyclonal rabbit anti-β-tubulin (dilution 1:1,000; cat. no.
10068-1-AP, ProteinTech Group, Inc.). After washing, the membranes
were incubated with HRP-conjugated secondary antibody (mouse
anti-rabbit IgG, dilution 1:5,000; cat. no. sc-2357, Santa Cruz
Biotechnology, Inc.). The blots were observed using an enhanced
chemiluminescence western blotting detection system (Bio-Rad
Laboratories, Inc.).
RNA-seq and data analysis
RNA integrity was examined by 1% formaldehyde
denaturing gel electrophoresis. RNA-seq was performed by Illumina
HiSeq sequencer (Illumina, Inc.) at CapitalBio Corporation. The raw
data of RNA-seq were aligned to the human genome version hg19.
Differentially expressed genes with absolute fold change >2.0
and P<0.05 were considered statistically significant in the
RNA-seq analysis. To better understand the roles of the
differentially expressed mRNAs, biological process analysis was
performed using the Gene Ontology (GO; www.geneontology.org) and Kyoto Encyclopedia of Genes
and Genomes (KEGG; http://www.genome.jp/kegg/) databases.
Migration assay
HFL-1 cells were seeded at 5×106 cells
per well in 6-well plates and then incubated at 37℃ in 5%
CO2 until confluence. Following preincubation with F12K
medium with 0.1% FBS for 24 h for synchronization, a scratch was
created in each well by using a 200-µl pipette tip. The
cells were gently washed twice with PBS and cultured in normoxic
and hypoxic conditioned medium from HMC-1 cells at 37℃ in 5%
CO2 for another 24 h. Images of the cells along the
scratch were captured at ×100 magnification using an inverted
optical microscope. The migrated cells were then counted to
evaluate the migration efficiency of the HFL-1 cells.
Immunofluorescence analysis
The slides of HFL-1 cells were fixed in 4%
paraformaldehyde for 30 min at room temperature. After washing with
PBS three times, they were permeabilized with 0.1% Triton X-100 in
PBS for 5 min on ice and then blocked with 3% BSA in PBS for 30 min
at room temperature. Subsequently, HFL-1 cells were incubated with
rabbit anti-α-SMA antibody (dilution, 1:1,000; cat. no. 55135-1-AP,
ProteinTech Group, Inc.) or corresponding serum as a negative
control at 4℃ overnight. Unbound antibodies were removed by washing
with PBS three times, and the slides were incubated with goat
anti-rabbit IgG-FITC antibody (1:500 dilution; cat. no. ab6717,
Abcam) in the dark for 60 min. After washing with PBS three times,
HFL-1 cells were stained with 4',6-diamidino-2-phenylindole (DAPI;
10 µg/ml, Sigma-Aldrich; Merck KGaA) in the dark for 5 min,
and finally examined under a Zeiss LSM laser confocal microscope
(Carl Zeiss AG).
Animal experimental protocol
All animal experiments were performed according to
the Guide for the Care and Use of Laboratory Animals and approved
by the Peking University First Hospital Ethical Review Committee
(approval no. J201533). Male Sprague-Dawley rats (n=12, age, 8-10
weeks, weight, 220±10 g) were obtained from Vital River Laboratory
Animal Technology Co. Ltd. and randomly divided into normoxic and
hypoxic groups (n=6/group). A hypoxic rat model was established as
described in our previous study (8). Briefly, rats in the normoxic group
were placed in a chamber with normobaric normoxia (21%
O2), while rats in the hypoxic group were placed in an
automatic hypobaric chamber (10% O2) continuously for 3
weeks. All rats were housed in an air-conditioned room (22-24℃)
with a 12-h light/dark cycle and free access to food and water. At
the end of the treatment period, the rats were anesthetized using
sodium pentobarbital (50 mg/kg, i.p.) and cervical dislocation was
performed quickly by an experienced researcher; no rats exhibited
clinical signs of suffering, and death was verified by the lack of
heartbeat and breathing. After euthanizing the anesthetized rats,
the lungs were harvested for hematoxylin and eosin (H&E)
staining and immunohistochemistry.
Histological analysis and
immunohistochemistry assay
The lung specimens were fixed in 4%
paraformaldehyde, embedded in paraffin and cut into 5-µm
sections. Subsequently, the sections were mounted on glass slides
and deparaffinized. Serial sections were stained with H&E for
morphological analysis using a routine protocol. Mast cells were
stained with toluidine blue (Sigma-Aldrich; Merck KGaA) in 1%
sodium chloride. For immunohistochemistry staining, paraffin
sections were autoclaved in a citrate buffer (pH 6.0) and blocked
in 3% H2O2. The slides were incubated at 4°C
overnight with primary rabbit antibodies, including anti-collagen
typeI(dilution, 1:1,000; cat. no. 14695-1-AP, ProteinTech Group,
Inc.), anti-collagen type III (dilution, 1:200; cat. no.
22734-1-AP, ProteinTech Group, Inc.), anti-MMP-9 (dilution, 1:200;
cat. no. 10375-2-AP, ProteinTech Group, Inc.), anti-MMP-13
(dilution, 1:200; cat. no. 18165-1-AP, ProteinTech Group, Inc.),
anti-TIMP-1 (dilution, 1:100; cat. no. 16644-1-AP, ProteinTech
Group, Inc.). Subsequently, the slides were incubated with
HRP-conjugated goat anti-rabbit IgG secondary antibody (provided at
working dilution, cat. no. PV-6001; Zhongshan Golden Bridge
Biotechnology). The immune complexes were detected by a
3,3′-diaminobenzidine detection system (Zhongshan Golden Bridge
Biotechnology). The sections were counterstained with hematoxylin
and dehydrated before mounting. The stained sections were observed
using an optical microscope.
Statistical analysis
Statistical analysis involved the use of SPSS 16.0
(SPSS, Inc.). Quantitative data are presented as the mean ±
standard deviation of at least 3 independent experiments. Student's
t-test or one-way ANOVA followed by Scheffe's post hoc test was
used to compare multiple groups. P<0.05 was considered to
indicate statistically significant differences.
Results
Increased proliferation of HMC-1 cells
under hypoxic conditions
To examine the effects of hypoxia on the viability
of human mast cells, HMC-1 cells were cultured under hypoxic
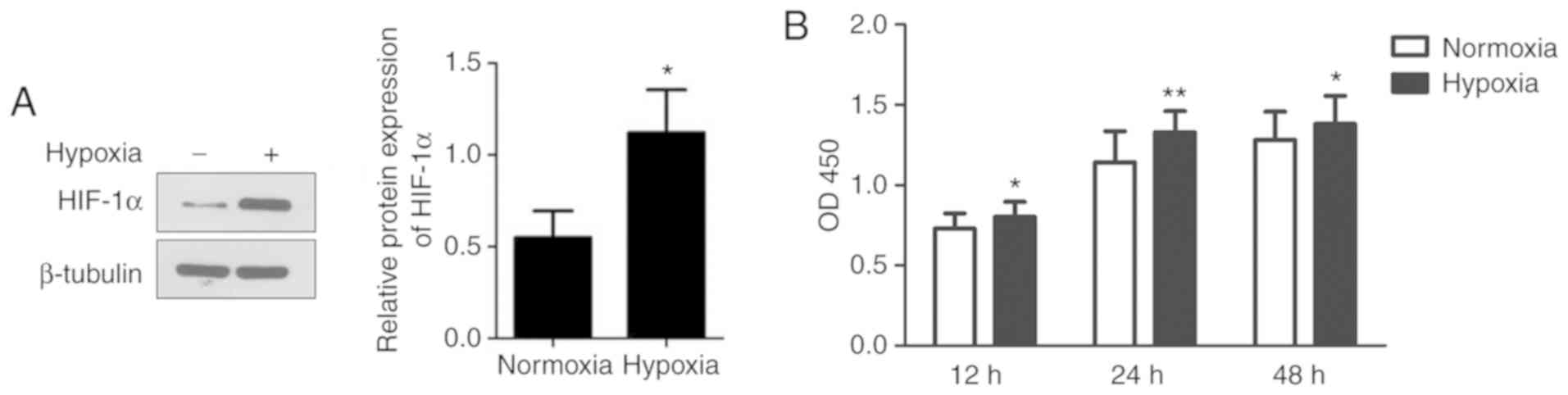
conditions and their proliferation was detected. As shown in
Fig. 1A, exposure to hypoxia
significantly increased the expression of HIF-1α, suggesting that
HMC-1 cells exist within a hypoxic environment. Compared to
normoxic conditions, hypoxia significantly increased the
proliferation of HMC-1 cells by 10.2 and 16.2% at 12 and 24 h,
respectively (Fig. 1B). However,
there was no significant difference between the hypoxic and
normoxic groups at 48 h.
Effects of hypoxia on the secretion of
cytokines by HMC-1 cells
To explore the changes in cytokines in human mast
cells under hypoxia, we next cultured HMC-1 cells under hypoxia for
the indicated time periods. The expression of IL-1β and IL-6
increased substantially after 12 h and peaked after 24 h, by almost
three- and six-fold, respectively, in the medium of HMC-1 cells
exposed to hypoxia compared with the normoxic controls. The
expression of TNF-α mRNA was increased in a time-dependent manner,
while the expression of IL-15 mRNA did not exhibit obvious changes
(Fig. 2A). Subsequently, the
resulting supernatant from HMC-1 cells under different conditions
was collected and the levels of cytokines were measured. As shown
in Fig. 2B-E, the levels of
IL-1β, IL-6 and TNF-α were higher in the hypoxic medium of HMC-1
cells compared with the normoxic control medium, while the IL-15
content remained unchanged.
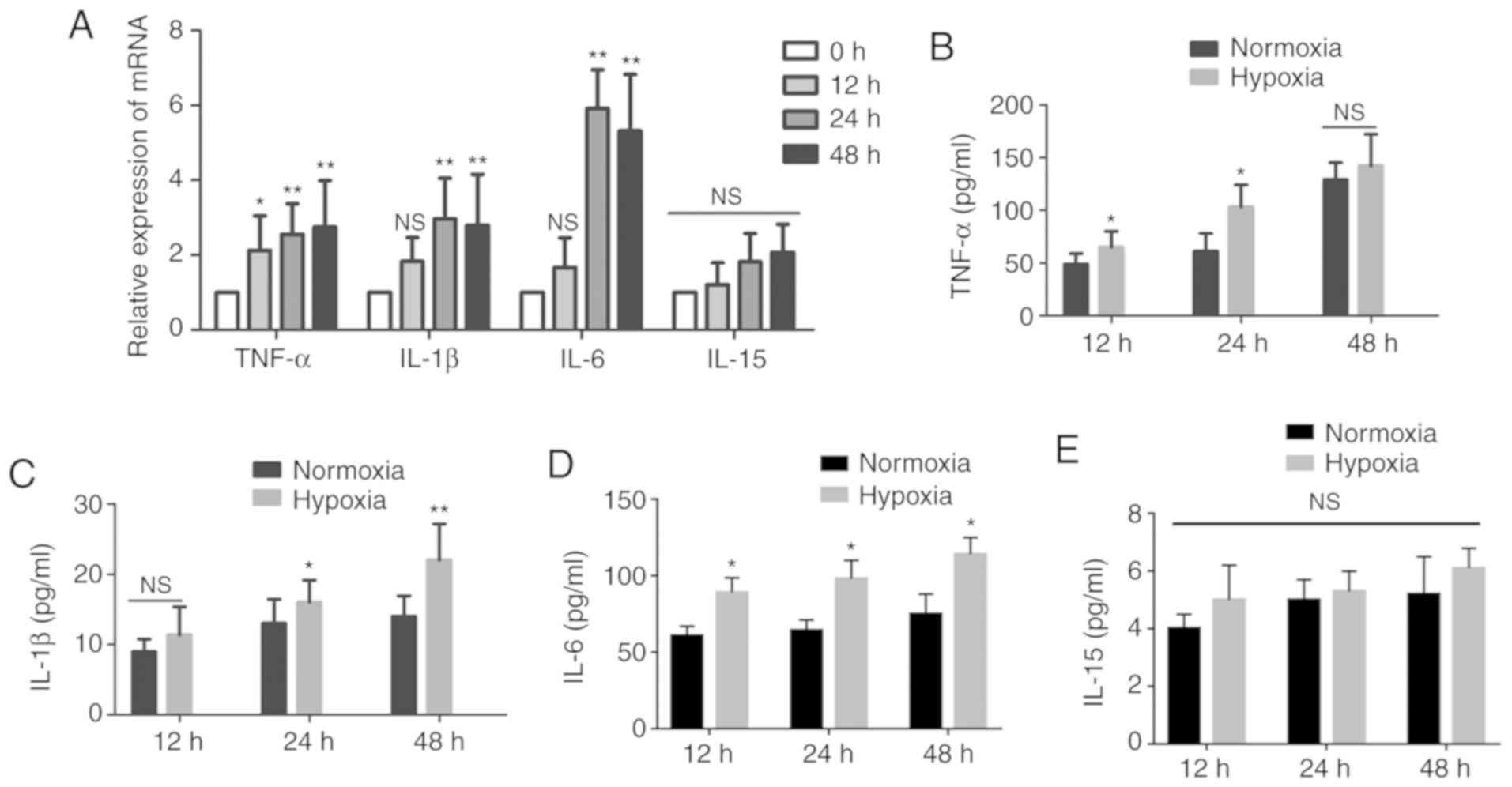 | Figure 2Effects of hypoxia on the secretion
of cytokines by HMC-1 cells. (A) HMC-1 cells were cultured under
hypoxic conditions for the indicated time periods and then
subjected to reverse transcription-quantitative polymerase chain
reaction to detect the expression of cytokines IL-1β, IL-6, IL-15
and TNF-α. β-tubulin was used as an internal control. (B-E)
Following culture under hypoxic conditions, the levels of the
secreted cytokines IL-1β, IL-6, IL-15 and TNF-α in the conditioned
medium from HMC-1 cells were determined by ELISA. There were 4-6
biological replicates/condition. Error bars depict the standard
deviation. *P<0.05, **P<0.01, NS, not
significant. IL, interleukin; TNF, tumor necrosis factor; HMC-1,
human mast cells. |
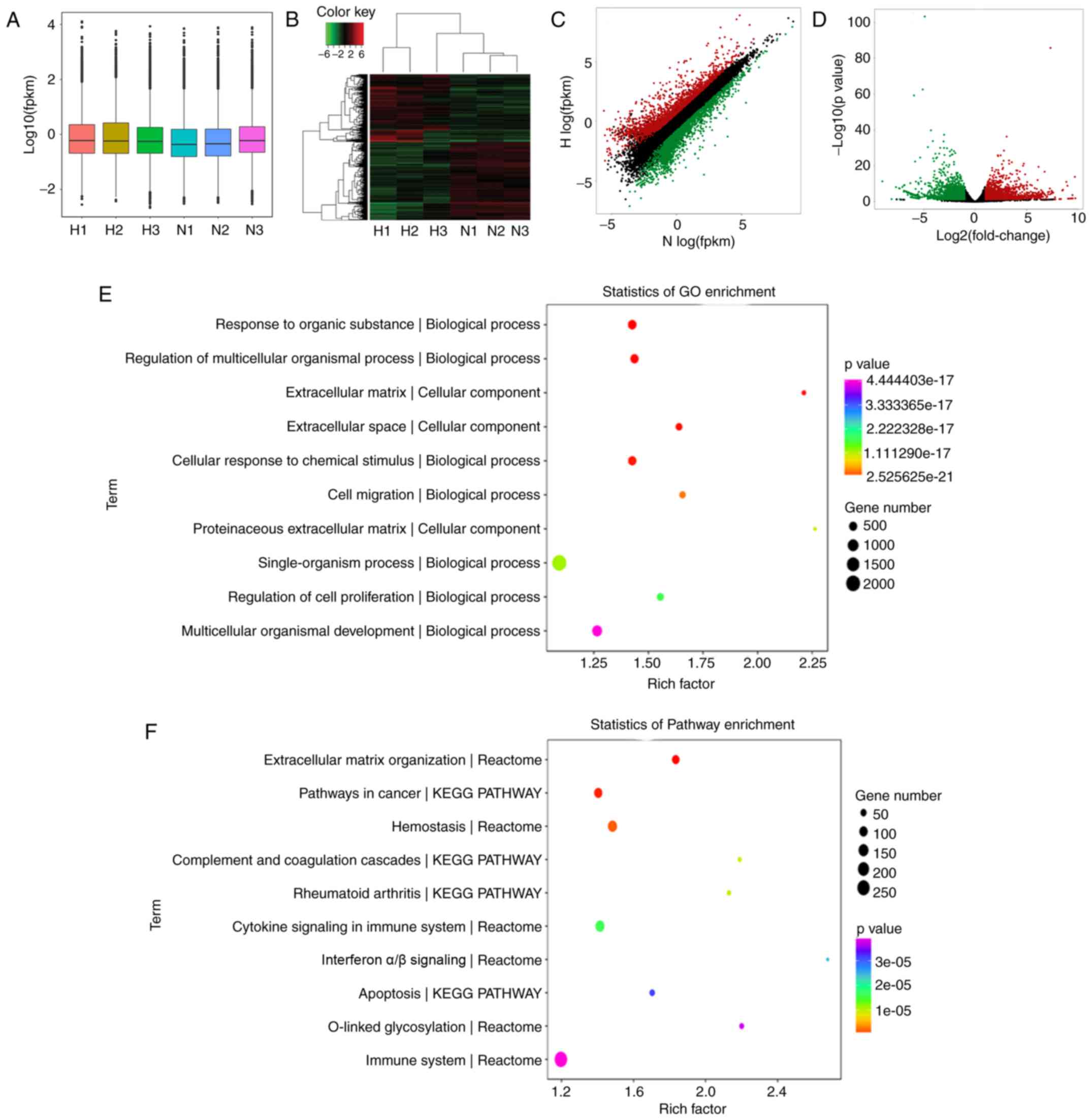
RNA-seq analysis of HFL-1 cells cultured
in conditioned medium from HMC-1 cells
To evaluate the effects of mast cells on lung
fibroblasts under hypoxia, the HFL-1 cells were cultured in hypoxic
and normoxic conditioned medium from HMC-1 cells and RNA-seq
analysis was performed to compare the global mRNA expression
profile. A box plot was drawn to visualize the distribution of the
intensities of all the samples after normalization and the
distribution of log2 ratios was found to be similar in all the
samples (Fig. 3A). Hierarchical
clustering revealed that there is a distinguishable mRNA expression
profiling between the hypoxic group and controls, and microarray
identified 4,495 differentially expressed mRNAs in total (fold
change >2.0, P-value <0.05), including 2,077 upregulated and
2,418 downregulated in HFL-1 cells cultured in hypoxic conditioned
medium (Fig. 3B). The top 40
differentially expressed mRNA are listed in Table I. Additionally, scatter and
volcano plots were constructed to visualize the significant
variation between the two groups (Fig. 3C and D). To understand the
function of these differentially expressed genes, we performed a
bioinformatic pathway analysis of GO and KEGG data. As shown in
Fig. 3E, the highest enriched GO
pathways targeted by differentially expressed genes were
extracellular matrix organization and cell migration, among others.
Moreover, the potential module function was predicted by KEGG
pathway analysis. Various pathways, including extracellular matrix
organization, cytokine signaling in immune system and collagen
formation, which are closely associated with pulmonary
hypertension, were significantly enriched in HFL-1 cells cultured
in hypoxic conditioned medium from HMC-1 cells (Fig. 3F).
 | Table ITop 20 upregulated and downregulated
mRNAs in human fetal lung fibroblasts (HFL-1) cultured in hypoxic
conditioned medium from human mast cells (HMC-1) and normoxic
controls. |
Table I
Top 20 upregulated and downregulated
mRNAs in human fetal lung fibroblasts (HFL-1) cultured in hypoxic
conditioned medium from human mast cells (HMC-1) and normoxic
controls.
A, Upregulated
mRNAs
|
|---|
| mRNAs | P-value | log2 fold
change |
|---|
| TNFSF8 | 0.000 | 9.4533 |
| IL36RN | 0.020 | 9.1130 |
| CYP19A1 | 0.000 | 8.5295 |
| BEST3 | 0.001 | 7.0775 |
| IL24 | 0.000 | 7.0084 |
| WTAPP1 | 0.000 | 6.7566 |
| STC1 | 0.000 | 6.6111 |
| ESM1 | 0.002 | 6.6003 |
| CCL3L3 | 0.002 | 6.5711 |
| MMP10 | 0.002 | 6.5556 |
| IL36B | 0.002 | 6.5535 |
| THBD | 0.000 | 6.4407 |
| CSF2 | 0.026 | 6.3719 |
| DCSTAMP | 0.000 | 6.3002 |
| IL1B | 0.000 | 6.2982 |
| SHISA2 | 0.005 | 6.2159 |
| LCE1C | 0.000 | 6.1842 |
| EREG | 0.000 | 6.1640 |
| PTGS2 | 0.000 | 6.1336 |
| SERPINB2/10 | 0.000 | 6.0740 |
|
| B, Downregulated
mRNAs |
|
| mRNAs | P-value | log2 fold
change |
|
| MUC19 | 0.038 − | 8.1010 |
| USH2A | 0.000 − | 7.3792 |
| TACR3 | 0.000 − | 7.3221 |
| ADH1B | 0.000 | −7.0604 |
| RSPO2 | 0.000 | −6.8458 |
| CDSN | 0.000 | −6.6503 |
| ITIH5 | 0.000 | −6.6445 |
| SMTNL2 | 0.000 | −6.6222 |
| RNU2-52P | 0.000 | −6.5828 |
| PTGDR2 | 0.000 | −6.5349 |
| ASTN1 | 0.000 | −6.5022 |
| SPTB | 0.000 | −6.4692 |
| OGN | 0.000 | −6.4411 |
| LAD | 0.000 | −6.3950 |
| NFE2 | 0.000 | −6.3828 |
| GPR20 | 0.000 | −6.2826 |
| MUC22 | 0.000 | −6.2617 |
| TMEM37 | 0.000 | −6.2537 |
| ABCA9 | 0.000 | −6.2412 |
| DNAH2 | 0.000 | −6.2343 |
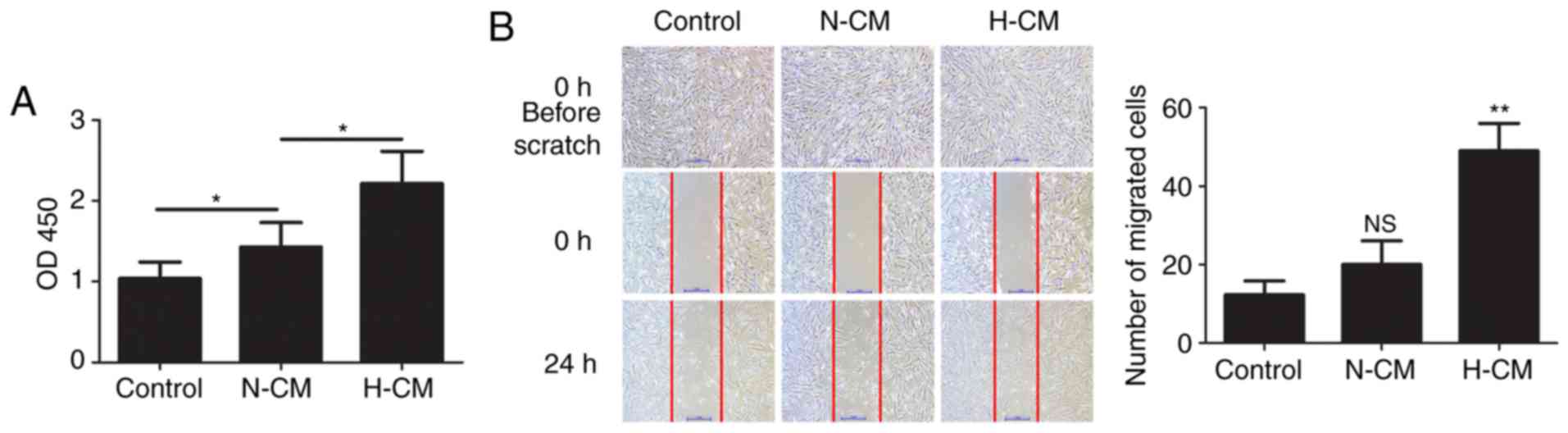
Hypoxic conditioned medium from HMC-1
cells promotes the proliferation and migration of HFL-1 cells
The proliferation and migration of lung fibroblasts
plays an important role in hypoxic pulmonary hypertension. To
assess the effect of the secretion of paracrine factors from mast
cells on human lung fibroblasts, HFL-1 cells were cultured in
hypoxic and normoxic conditioned medium from HMC-1 cells and their
proliferation and the migration were examined. Compared with
normoxic conditioned medium from HMC-1 cells, hypoxic conditioned
medium significantly increased the proliferation of HFL-1 cells
(Fig. 4A). The migration of HFL-1
cells was examined by scratch assay and the results revealed that
treatment with hypoxic conditioned medium significantly facilitated
the migration of HFL-1 cells (Fig.
4B).
Hypoxic conditioned medium from HMC-1
cells regulates the production of collagen and collagenase in HFL-1
cells
To further characterize the role of mast cells in
fibrogenesis, the expression of collagen I and III was first
detected in HFL-1 cells under different culture conditions. The
mRNA expression levels of collagen type I and III were both higher
in the hypoxic conditioned medium compared with those in the
control medium, and the protein expression displayed the same trend
(Fig. 5). The expression of the
proteolytic enzymes MMP-9 and MMP-13 at the mRNA and protein levels
was upregulated in HFL-1 cells treated with hypoxic conditioned
medium from HMC-1 cells, while TIMP-1 expression remained
unchanged.
 | Figure 5Effects of conditioned medium from
HMC-1 cells on the production of collagens and MMPs by HFL-1 cells.
(A) Following culture in normoxic or hypoxic conditioned medium
from HMC-1 cells, the mRNA expression of collagen type I and III,
MMP-9, MMP-13 and TIMP-1 in HFL-1 cells were assessed by reverse
transcription-quantitative polymerase chain reaction analysis,
normalized to the expression of β-tubulin, and expressed as fold
change relative to the expression of the control group. (B) Western
blot analysis of collagen type I and III, MMP-9, MMP-13 and TIMP-1
protein expression in HFL-1 cells cultured in conditioned medium
from HMC-1 cells. There were 4 biological replicates/condition.
Error bars depict the standard deviation. *P<0.05,
**P<0.01, NS, not significant. COL I, collagen type
I; COL III, collagen type III; MMP-9, matrix metalloproteinase-9;
TIMP-1, tissue inhibitor of metalloproteinase-1; N-CM, normoxic
conditioned medium; H-CM, hypoxic conditioned medium; HMC-1, human
mast cells; HFL-1, human fetal lung fibroblasts. |
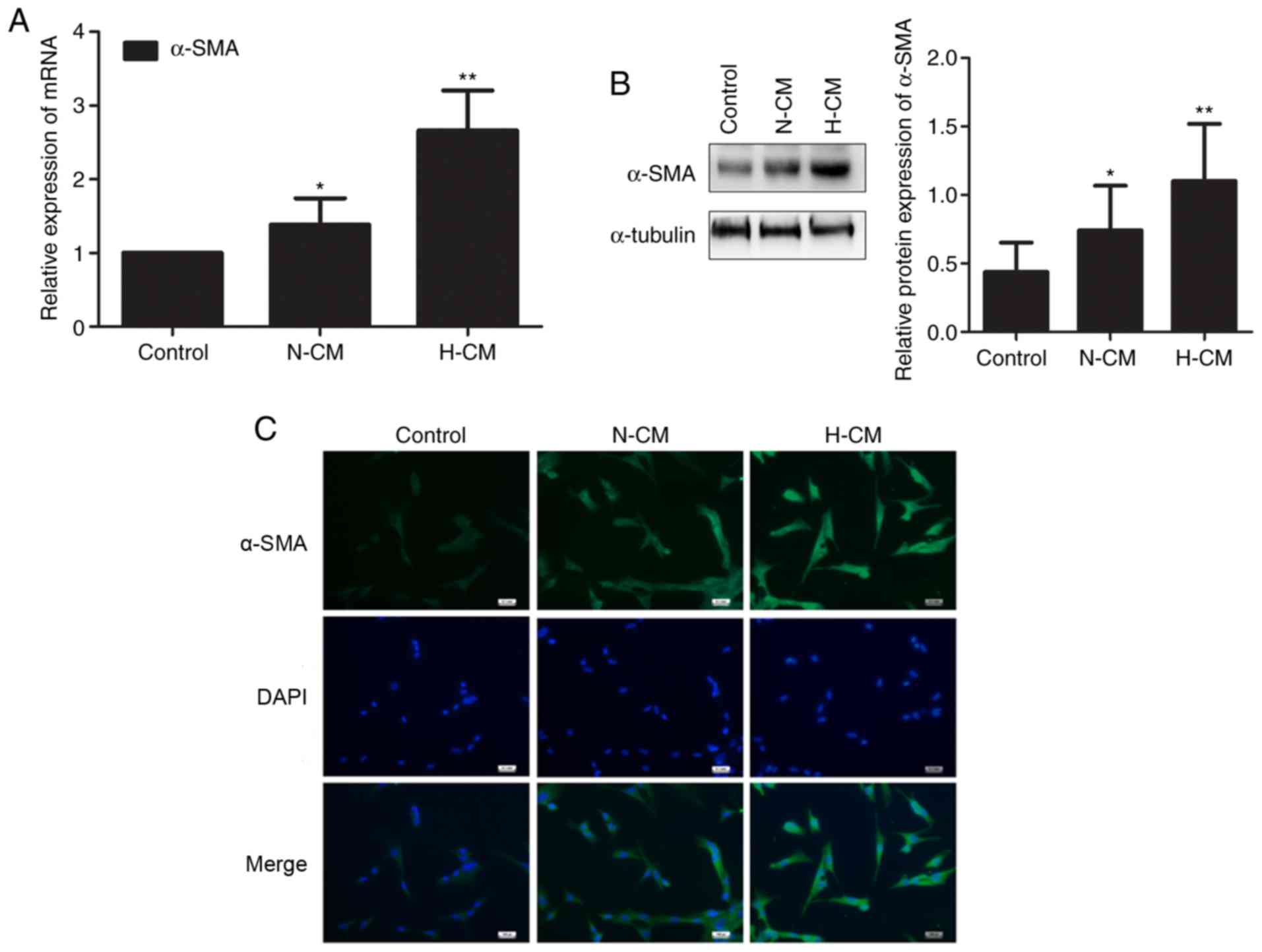
Hypoxic conditioned medium from HMC-1
cells induces the transition from fibroblast to myofibroblast in
HFL-1 cells
We investigated whether human mast cells affect the
constitutive characteristics of lung fibroblasts under hypoxia. The
expression of α-SMA mRNA and protein was upregulated in HFL-1 cells
cultured in hypoxic conditioned medium from HMC-1 cells compared
with HFL-1 cells cultured in normoxic conditioned medium (Fig. 6A and B). Similarly, the transition
from fibroblast to myofibroblast induced by hypoxic conditioned
medium from HMC-1 cells was identified by immunofluorescence,
suggesting that mast cells are involved in the phenotypic switch of
lung fibroblasts under hypoxia (Fig.
6C).
Increased mast cell numbers and activated
extracellular matrix remodeling in the lungs of hypoxic rats
Histological analysis revealed that the thickness of
pulmonary vessels in rats in the hypoxic group was increased
compared with the normoxic controls, while their vascular lumens
were narrowed (Fig. 7A).
According to the toluidine blue in situ staining of lung
specimens, an increased accumulation of mast cells was observed in
the lungs of hypoxic rats (Fig.
7B). Immunohistochemistry results revealed that the hypoxic
group exhibited increased deposition of collagen I and III compared
with the control group (Fig. 7C).
Moreover, the protein expression of the proteolytic enzymes MMP-9,
MMP-13 and TIMP-1 was also increased in the lung tissues of hypoxic
rats, indicating that hypoxia facilitates the accumulation of mast
cells, and activates collagen metabolism and vascular remodeling
in vivo (Fig. 7D).
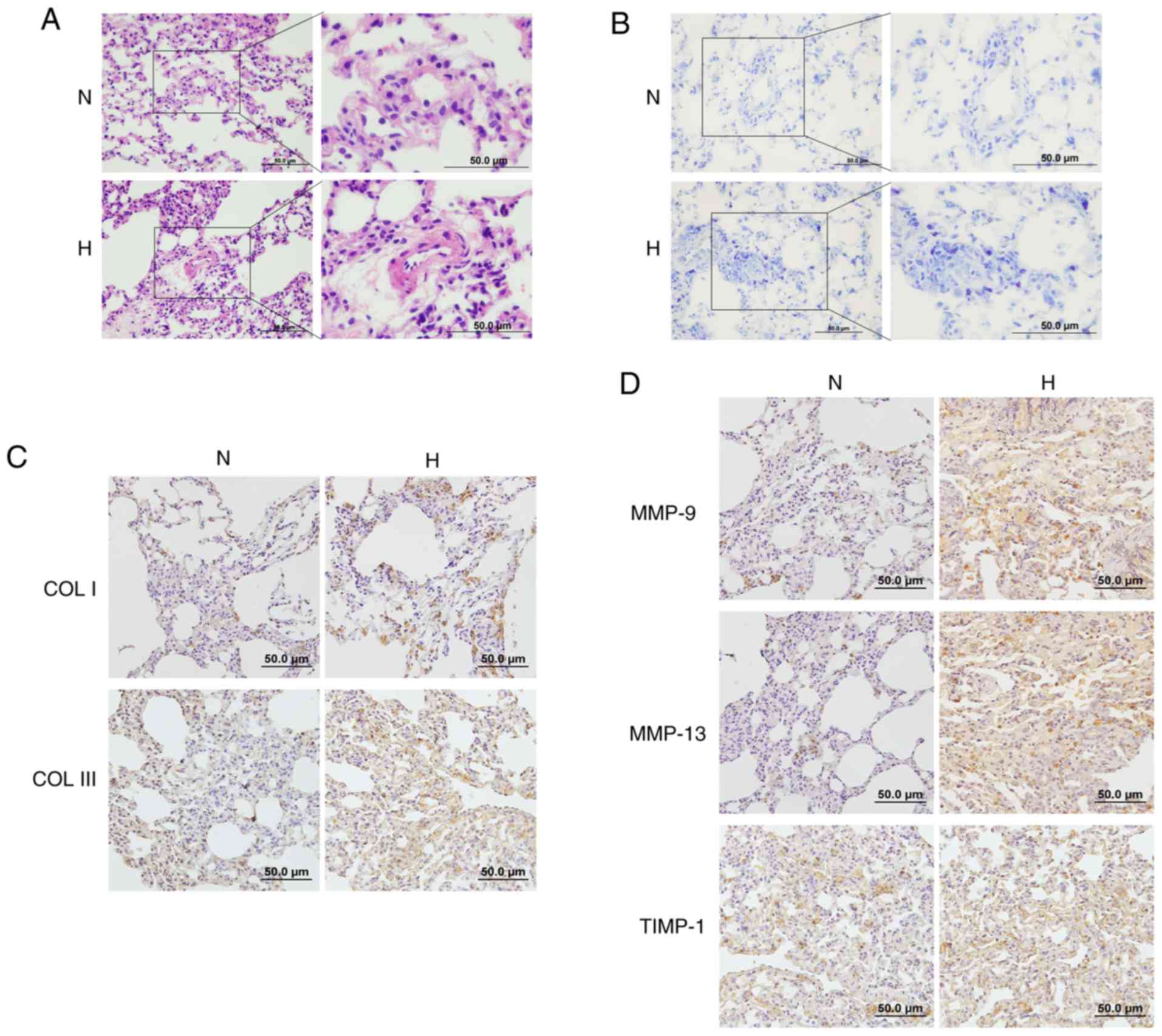 | Figure 7Effects of hypoxia on mast cells and
ECM remodeling in rat lungs. (A) The pathological changes of
pulmonary vessels in the rat lungs after exposure to hypoxia were
examined by hematoxylin and eosin staining (magnification, ×200 and
400). (B) Toluidine blue staining demonstrated increased
accumulation of mast cells in the lungs of hypoxic rats
(magnification, ×200 and 400). (C and D) The expression of collagen
I, collagen III, MMP-9, MMP-13 and TIMP-1 in the lungs of rats
after exposure to hypoxia was determined by immunohistochemistry
staining (magnification, ×200). ECM, extracellular matrix; MMP,
matrix metalloproteinase; TIMP, tissue inhibitor of
metalloproteinase; N, normoxic; H, hypoxic. |
Discussion
The normal pulmonary vascular system is
well-organized and exhibits high compliance, but its physical
structure is destroyed by pulmonary vascular remodeling in patients
with pulmonary hypertension, which is characterized by increased
vascular stiffness and reduced pulmonary arterial compliance
(4). Resident pulmonary arterial
fibroblasts are considered one of the key cells initiating and
potentiating chronic hypoxic pulmonary vascular remodeling
(6,12). The mechanism responsible for the
structural and functional changes in fibroblasts remains unclear.
In the present study, increased proliferation and proinflammatory
cytokine secretion by mast cells was observed under hypoxic
conditions. RNA-seq identified 2,077 upregulated and 2,418
downregulated mRNAs in HFL-1 cells cultured in hypoxic conditioned
medium from HMC-1 cells compared with normoxic controls, which are
involved in various pathways, including extracellular matrix
organization, cell proliferation and migration. To verify the
effects of HMC-1 on HFL-1 cells, conditioned medium from hypoxic
mast cells was found to increase the proliferation, migration
capacity and collagen production in lung fibroblasts and trigger
the phenotypic transition from fibroblasts to myofibroblasts.
Similar pathological phenomena, including vascular remodeling,
accumulation of mast cells and excessive deposition of collagen
were observed in the lungs of hypoxic rats.
Mast cells are derived from CD34-expressing
hematopoietic stem cells and have traditionally been recognized as
sentinel cells in allergic and non-allergic immune responses under
physiological conditions (13-15). Of note, accumulating evidence
demonstrates that mast cells are involved in vascular remodeling
through secretion of proangiogenic proteases and interaction with
endothelial and smooth muscle cells, and they may be responsible
for the development of pulmonary hypertension (16,17). In the lungs of left heart disease
model rats, mast cell activation is the most notable change in the
pathological process of vascular remodeling and pulmonary
hypertension, and these effects are largely attenuated in mast
cell-deficient rats (18,19). In the present study, hypoxia
induced the proliferation of mast cells and increased mast cell
infiltration in rat lungs with hypoxic pulmonary hypertension,
which is in accordance with previous studies. Mast cells are found
to be highly viable for up to 3 days under hypoxic conditions, and
then the viability decreases for 5 to 7 days; it was also observed
that hypoxia increases the secretion of IL-6 by mast cells, but
does not affect their degranulation (20). Vajner et al reported a
significant increase in the number of mast cells in the walls of
prealveolar arteries in the early stage of hypertension in the
lungs of rats exposed to chronic hypoxia, and additional mast cell
accumulation was detected within the walls of conduit arteries and
subpleurally (21). IL-6 plays a
critical role in pulmonary vascular remodeling and hypertension in
hypoxic and monocrotaline-induced PAH models, and mast cells have
been identified as the primary source of IL-6 (22). In the present study, it was
observed that hypoxia facilitated the production and secretion of
various cytokines, including IL-1β, IL-6 and TNF-α, by mast cells.
The early use of mast cell stabilizers may inhibit the release of
all mediators, including proteases and proinflammatory factors, and
reduce vascular remodeling in hypoxia-induced rodent pulmonary
hypertension, suggesting that mast cells may be a potential target
to arrest and reverse pulmonary vascular remodeling (23).
Under physiological conditions, the balance between
proliferation and apoptosis of vascular cells contributes to the
maintenance of the thickness of the vascular wall. If the
equilibrium is disturbed, the overproliferating cells may obstruct
the vessel lumen, resulting in vascular remodeling and increased
pulmonary pressure. The adventitial fibroblast is a pivotal
regulator of vascular wall function and performs biological
functions, including proliferation, differentiation, synthesis and
secretion of extracellular matrix and other mediators (24,25). The activation of pulmonary
adventitial fibroblasts contributes substantially to pulmonary
vascular remodeling (8). It was
reported that mast cells promote the viability of fibroblasts in
several physiological and pathological processes. In idiopathic
pulmonary fibrosis, tryptase from mast cells activates the protein
kinase C-α/Raf-1/p44/42 signaling pathway in human lung fibroblasts
and thus promotes their proliferation. In turn, fibroblasts induce
mast cell activation and stimulate tryptase release (26). In the present study, we found that
hypoxia-induced cytokine secretion from mast cells stimulated the
proliferation and migration of lung fibroblasts. The proliferation
of adventitial fibroblasts increases within hours of hypoxic
exposure and fibroblasts can transform into myofibroblasts and
migrate into the medial layer, or even into the intima,
subsequently resulting in increased thickness of the vascular wall
and reduced pulmonary arterial compliance. Moreover, it is
hypothesized that certain angiogenic factors, including VEGF and
PDGFR, may be involved in the pathological process of vascular
remodeling and pulmonary hypertension. Further studies with in
vivo and in vitro experiments to detect the expression
of angiogenic factors are warranted. The transition from fibroblast
to myofibroblast is a critical pathophysiological process in a
variety of fibrotic diseases, including pulmonary hypertension
(27). Accumulated myofibroblasts
have been observed in the intimal lesions of patients with
pulmonary hypertension (28). We
herein observed that, compared with normoxic control medium,
hypoxic conditioned medium from mast cells induced the
proliferation and phenotypic transition of lung fibroblasts,
indicating that the hypoxic microenvironment is another important
trigger for the activation of fibroblasts and pulmonary vascular
remodeling.
The imbalance of extracellular matrix synthesis and
degradation also contributes to the structural remodeling of
peripheral pulmonary arteries and aggravates the pathological
process (29,30). Active collagen synthesis,
increased collagen accumulation and deposition in perivascular and
intravascular compartments has been observed in patients and
experimental models of pulmonary hypertension. It was reported that
type I collagen was the earliest expressed gene and exhibited the
largest increase in expression in the lungs of mice exposed to
hypoxia, followed by type III collagen, type IV basement membrane
collagen, MMPs and TIMPs, which increased with prolonged hypoxic
exposure (31). Wang et al
reported that the content of collagen not cross-linking in the
pulmonary vasculature is associated with pulmonary artery
remodeling and leads to proximal vascular stiffening and
ventricular pulsatile overload in early pulmonary hypertension
(32). The N-terminal propeptide
of type III procollagen was found to be significantly elevated in
the serum of patients with pulmonary hypertension and may predict
the severity of the disease (33). In the present study, we found that
hypoxic conditioned medium from mast cells stimulated fibroblasts
to produce more type I and type III collagen. Moreover,
disorganized expression of proteolytic enzymes, including MMP-9,
MMP-13 and TIMP-1, in the pulmonary arteries is involved in
pulmonary vascular remodeling. However, the change tendency of
TIMP-1 expression in vivo differs from the in vitro
results, which requires further investigation. Transgenic mice
overexpressing MMP-9 exhibit aggravated pulmonary vascular
remodeling and pulmonary hypertension following treatment with
monocrotaline compared with wild-type mice (34). The level of circulating
proteolytic enzymes, including MMP-9 and TIMP-1, was found to be
dysregulated in patients with pulmonary hypertension (35). The present study demonstrated that
hypoxic mast cells contribute to dysregulated extracellular matrix
synthesis and degradation in vitro and in vivo.
The phenotypic alteration of lung fibroblasts
activated by hypoxic conditioned medium from mast cells was
consistent with our hypothesis that the multiple effects of the
hypoxic microenvironment and mast cells on fibroblasts contribute
to pulmonary vascular remodeling, and this process is likely one of
the important mechanisms underlying the development of hypoxic
pulmonary hypertension.
Abbreviations:
|
TNF-α
|
tumor necrosis factor-α
|
|
IL
|
interleukin
|
|
MMP
|
matrix metalloproteinase
|
|
COPD
|
chronic obstructive pulmonary
disease
|
|
IMDM
|
Iscove's modified Dulbecco's
medium
|
|
FBS
|
fetal bovine serum
|
|
CCK8
|
Cell Counting Kit-8
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
TIMP
|
tissue inhibitor of
metalloproteinase
|
|
COL I
|
collagen type I
|
|
COL III
|
collagen type III
|
|
HIF
|
hypoxia-inducible factor
|
|
PAH
|
pulmonary arterial hypertension
|
|
α-SMA
|
α-smooth muscle actin
|
|
GO
|
Gene Ontology
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
H&E
|
hematoxylin and eosin
|
|
PBS
|
phosphate-buffered saline
|
|
DAPI
|
4′,6-diamidino-2-phenylindole
|
Acknowledgments
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81670043).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW, LL and XL conceived and designed the experiment.
XW, XC, YW and YL performed the experiments. XW and XC performed
data analysis. XW, LL and XL prepared and wrote the manuscript. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were performed according to
the Guide for the Care and Use of Laboratory Animals and were
approved by the Peking University First Hospital Ethical Review
Committee (approval no. J201533). All efforts were made to minimize
animal suffering and the number of animals used.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Grimminger J, Ghofrani HA, Weissmann N,
Klose H and Grimminger F: COPD-associated pulmonary hypertension:
Clinical implications and current methods for treatment. Exp Rev
Respir Med. 10:755–766. 2016. View Article : Google Scholar
|
|
2
|
Wang Z and Chesler NC: Pulmonary vascular
mechanics: Important contributors to the increased right
ventricular afterload of pulmonary hypertension. Exp Physiol.
98:1267–1273. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nakagawa Y, Kishida K, Kihara S, Funahashi
T and Shimomura I: Adiponectin ameliorates hypoxia-induced
pulmonary arterial remodeling. Biochem Biophys Res Commun.
382:183–188. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thenappan T, Ormiston ML, Ryan JJ and
Archer SL: Pulmonary arterial hypertension: Pathogenesis and
clinical management. BMJ. 360:j54922018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sluiter I, van Heijst A, Haasdijk R,
Kempen MB, Boerema-de Munck A, Reiss I, Tibboel D and Rottier RJ:
Reversal of pulmonary vascular remodeling in pulmonary hypertensive
rats. Exp Mol Pathol. 93:66–73. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Di Wang H, Ratsep MT, Chapman A and Boyd
R: Adventitial fibroblasts in vascular structure and function: The
role of oxidative stress and beyond. Canadian J Physiol Pharmacol.
88:177–186. 2010. View
Article : Google Scholar
|
|
7
|
Lerner CA, Rutagarama P, Ahmad T, Sundar
IK, Elder A and Rahman I: Electronic cigarette aerosols and copper
nanoparticles induce mitochondrial stress and promote DNA
fragmentation in lung fibroblasts. Biochem Biophys Res Commun.
477:620–625. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chai X, Sun D, Han Q, Yi L, Wu Y and Liu
X: Hypoxia induces pulmonary arterial fibroblast proliferation,
migration, differentiation and vascular remodeling via the
PI3K/Akt/p70S6K signaling pathway. Int J Mol Med. 41:2461–2472.
2018.PubMed/NCBI
|
|
9
|
Marsh LM, Jandl K, Grunig G, Foris V,
Bashir M, Ghanim B, Klepetko W, Olschewski H, Olschewski A and
Kwapiszewska G: The inflammatory cell landscape in the lungs of
patients with idiopathic pulmonary arterial hypertension. Eur
Respir J. 51:pii: 1701214. 2018. View Article : Google Scholar
|
|
10
|
Kosanovic D, Dahal BK, Peters DM, Seimetz
M, Wygrecka M, Hoffmann K, Antel J, Reiss I, Ghofrani HA, Weissmann
N, et al: Histological characterization of mast cell chymase in
patients with pulmonary hypertension and chronic obstructive
pulmonary disease. Pulm Circ. 4:128–136. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang Y, Zhang Z, Zhang H, Hong K, Tang W,
Zhao L, Lin H, Liu D, Mao J, Wu H and Jiang H: Effects of maternal
acrolein exposure during pregnancy on testicular testosterone
production in fetal rats. Mol Med Rep. 16:491–498. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kasmi El KC, Pugliese SC, Riddle SR, Poth
JM, Anderson AL, Frid MG, Li M, Pullamsetti SS, Savai R, Nagel MA,
et al: Adventitial fibroblasts induce a distinct
proinflammatory/profibrotic macrophage phenotype in pulmonary
hypertension. J Immunol. 193:597–609. 2014. View Article : Google Scholar
|
|
13
|
Mukai K, Tsai M, Saito H and Galli SJ:
Mast cells as sources of cytokines, chemokines, and growth factors.
Immunol Rev. 282:121–150. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shi GP, Bot I and Kovanen PT: Mast cells
in human and experimental cardiometabolic diseases. Nat Rev
Cardiol. 12:643–658. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ko SC, Lee DS, Park WS, Yoo JS, Yim MJ,
Qian ZJ, Lee CM, Oh J, Jung WK and Choi IW: Anti-allergic effects
of a nonameric peptide isolated from the intestine gastrointestinal
digests of abalone (Haliotis discus hannai) in activated HMC-1
human mast cells. Int J Mol Med. 37:243–250. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pugliese SC, Poth JM, Fini MA, Olschewski
A, El Kasmi KC and Stenmark KR: The role of inflammation in hypoxic
pulmonary hypertension: From cellular mechanisms to clinical
phenotypes. Am J Physiol Lung Cell Mol Physiol. 308:L229–L252.
2015. View Article : Google Scholar :
|
|
17
|
Moiseeva EP, Roach KM, Leyland ML and
Bradding P: CADM1 is a key receptor mediating human mast cell
adhesion to human lung fibroblasts and airway smooth muscle cells.
PLoS One. 8:e615792013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhu YJ, Kradin R, Brandstetter RD, Staton
G, Moss J and Hales CA: Hypoxic pulmonary hypertension in the mast
cell-deficient mouse. J Appl Physiol Respir Enviroc Exerci Physiol.
54:680–686. 1983.
|
|
19
|
Hoffmann J, Yin J, Kukucka M, Yin N,
Saarikko I, Sterner-Kock A, Fujii H, Leong-Poi H, Kuppe H,
Schermuly RT and Kuebler WM: Mast cells promote lung vascular
remodelling in pulmonary hypertension. Eur Respir J. 37:1400–1410.
2011. View Article : Google Scholar
|
|
20
|
Gulliksson M, Carvalho RF, Ulleras E and
Nilsson G: Mast cell survival and mediator secretion in response to
hypoxia. PLoS One. 5:e123602010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vajner L, Vytásek R, Lachmanová V, Uhlík
J, Konrádová V, Novotná J, Hampl V and Herget J: Acute and chronic
hypoxia as well as 7-day recovery from chronic hypoxia affects the
distribution of pulmonary mast cells and their MMP-13 expression in
rats. Int J Exp Pathol. 87:383–391. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Breitling S, Hui Z, Zabini D, Hu Y,
Hoffmann J, Goldenberg NM, Tabuchi A, Dos Santos C and Kuebler WM:
The mast cell-B cell axis in lung vascular remodeling and pulmonary
hypertension. Am J Physiol Lung Cell Mol Physiol. 312:L710–L721.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bartelds B, van Loon RLE, Mohaupt S,
Wijnberg H, Dickinson MG, Boersma B, Takens J, van Albada M and
Berger RMF: Mast cell inhibition improves pulmonary vascular
remodeling in pulmonary hypertension. Chest. 141:651–660. 2012.
View Article : Google Scholar
|
|
24
|
Haurani MJ and Pagano PJ: Adventitial
fibroblast reactive oxygen species as autacrine and paracrine
mediators of remodeling: Bellwether for vascular disease? Cardiovas
Res. 75:679–689. 2007. View Article : Google Scholar
|
|
25
|
Stenmark KR, Gerasimovskaya E, Nemenoff RA
and Das M: Hypoxic activation of adventitial fibroblasts: Role in
vascular remodeling. Chest. 122(6 Suppl): S326–S334. 2002.
View Article : Google Scholar
|
|
26
|
Wygrecka M, Dahal BK, Kosanovic D,
Petersen F, Taborski B, von Gerlach S, Didiasova M, Zakrzewicz D,
Preissner KT, Schermuly RT and Markart P: Mast cells and
fibroblasts work in concert to aggravate pulmonary fibrosis: Role
of transmembrane SCF and the PAR-2/PKC-alpha/Raf-1/p44/42 signaling
pathway. Am J Pathol. 182:2094–2108. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Short M, Nemenoff RA, Zawada WM, Stenmark
KR and Das M: Hypoxia induces differentiation of pulmonary artery
adventitial fibroblasts into myofibroblasts. Am J Physiol Cell
Physiol. 286:C416–C425. 2004. View Article : Google Scholar
|
|
28
|
Farkas L, Gauldie J, Voelkel NF and Kolb
M: Pulmonary hypertension and idiopathic pulmonary fibrosis: A tale
of angiogenesis, apoptosis, and growth factors. Am J Respir Cell
Mol Biol. 45:1–15. 2011. View Article : Google Scholar
|
|
29
|
Thenappan T, Chan SY and Weir EK: Role of
extracellular matrix in the pathogenesis of pulmonary arterial
hypertension. Am J Physiol Heart Circ Physiol. 315:H1322–H1331.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu P, Yan S, Chen M, Chen A, Yao D, Xu X,
Cai X, Wang L and Huang X: Effects of baicalin on collagen Iota and
collagen IotaIotaIota expression in pulmonary arteries of rats with
hypoxic pulmonary hypertension. Int J Mol Med. 35:901–908. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Estrada KD and Chesler NC:
Collagen-related gene and protein expression changes in the lung in
response to chronic hypoxia. Biomech Model Mechanobiol. 8:263–272.
2009. View Article : Google Scholar :
|
|
32
|
Wang Z, Schreier DA, Abid H, Hacker TA and
Chesler NC: Pulmonary vascular collagen content, not cross-linking,
contributes to right ventricular pulsatile afterload and overload
in early pulmonary hypertension. J Appl Physiol (1985).
122:253–263. 2017. View Article : Google Scholar :
|
|
33
|
Safdar Z, Tamez E, Frost A, Guffey D,
Minard CG and Entman ML: Collagen metabolism biomarkers and health
related quality of life in pulmonary arterial hypertension. Int J
Cardiovas Res. 4:2015.
|
|
34
|
George J and D'Armiento J: Transgenic
expression of human matrix metalloproteinase-9 augments
monocrotaline-induced pulmonary arterial hypertension in mice. J
Hypertens. 29:299–308. 2011. View Article : Google Scholar
|
|
35
|
Safdar Z, Tamez E, Chan W, Arya B, Ge Y,
Deswal A, Bozkurt B, Frost A and Entman M: Circulating collagen
biomarkers as indicators of disease severity in pulmonary arterial
hypertension. JACC Heart Fail. 2:412–421. 2014. View Article : Google Scholar : PubMed/NCBI
|