Introduction
Heart failure (HF) is caused by any cardiac disorder
that leads to an increased intracardiac pressure or a decreased
cardiac output (1). Myocardial
infarction (MI) is a common and increasingly recognized
predisposition to HF (2,3). Following MI, neurohormonal systems
are activated, via decreased cardiac output, to maintain
hemodynamic stability (2).
However, the sustained activation of neurohormonal systems results
in maladaptive alterations, including cardiac hypertrophy and
cardiomyocyte death, and can lead to loss of cardiomyocytes and
contractile dysfunction (2,4).
Slow and maintained cardiomyocyte death occurs in failing hearts
and is a major pathophysiologic mechanism that underlies
progressive cardiac dysfunction (2). Significant progress has been made in
determining postinfarction therapeutic strategies. However, the
incidence of HF continues to increase (5,6).
Therefore, it is important to understand the underlying mechanisms
associated with cardiomyocyte death to develop novel therapies for
use in HF prevention and treatment.
Apoptosis is a highly choreographed form of cell
death that is mediated by caspases (4). Gradual but sustained cardiomyocyte
apoptosis occurs in failing hearts, leading to the progressive loss
of cardiomyocytes and lethal HF (4,7-12).
Angiotensin-converting enzyme inhibitors and β-receptor blockers
are used in clinical practice, and the benefits of these treatments
presumably result in part from reductions in cardiomyocyte
apoptosis (10). Following MI,
the activation of protein synthesis in response to neuroendocrine
or growth factors serves an important role in cardiomyocyte
hypertrophy and apoptosis (13,14). The endoplasmic reticulum (ER) is
the main site of synthesized protein folding (13). Any cellular stressor leading to
the accumulation of unfolded proteins in the ER will trigger ER
stress (also known as the unfolded protein response) (13). Multiple stimuli, including
hypoxia, oxidative injury and protein aggregates, have been
identified in failing hearts and can trigger ER stress (15). The initial aim of the unfolded
protein response is to reduce the accumulation of unfolded proteins
and restore normal ER function (16). However, prolonged or severe ER
stress can trigger processes leading to apoptosis (16). The initiation of ER stress is
characterized by the upregulation of chaperones, including
glucose-regulated protein 78 kDa (GRP78) (16), while prolonged or excessive ER
stress promotes apoptosis through the transcriptional induction of
DNA damage-inducible transcript 3 (DDIT3), or the activation of
c-JUN NH2-terminal kinase (JNK)- and-or
caspase-12-dependent pathways (17,18).
Autophagy is another type of cell death that is
primarily a prosurvival mechanism by which intracellular components
are digested and recycled by lysosomal degradation to maintain
energy production and protein synthesis (4,19-22). Numerous forms of HF are implicated
by the accumulation of aberrant proteins, which occurs due to the
insufficient protein degradation (5,10,23). Autophagy is a major intracellular
protein degradation mechanism that is often upregulated in
cardio-myocytes within a failing heart (4,23).
The protective effect of autophagy under conditions of stress,
especially in ischemia, starvation and b-adrenergic stimulation, is
exhibited by the remove of damaged or misfolded proteins,
organelles and aggregates (4,19,24-27). Additionally, impaired autophagy
has been indicated to serve a pathological role in the progression
of HF (23,27,28). The mechanistic target of rapamycin
(mTOR) kinase and Beclin-1 are two major regulators of autophagy
(4). Under nutrient-rich
conditions, mTOR inhibits autophagy by inactivating
autophagy-associated genes. During starvation, the activity of mTOR
is reduced by the class I PI3K-Akt pathway and autophagy is
inhibited (4,23). Beclin-1 signaling positively
regulates autophagy by facilitating autophagosome formation
(4).
mTOR kinase is an important regulator that is
associated with protein synthesis, autophagy and survival (29,30). The mTOR kinase is present in two
distinct multiprotein complexes, mTOR complex 1 (mTORC1) and mTOR
complex 2 (mTORC2) (29). mTORC1
detects upstream inputs and promotes protein synthesis, cell growth
and ribosomal biogenesis through downstream effectors, including S6
kinase and eIF4E-binding protein-1, which phosphorylate ribosomal
S6 protein (29,30). Stimuli of neurohormonal systems,
including angiotensin II and b-adrenoceptor agonists, can cause the
rapid and sustained activation of S6 kinase (31,32). Additionally, mTORC1 inhibits
catabolic processes, including autophagy (30). The inhibition of mTORC1 signaling,
by pharmacological or genetic approaches, increases cardiomyocyte
autophagy (33,34). In comparison, the function of
mTORC2 is not fully determined. Previous findings have demonstrated
that mTORC2 triggers an anti-apoptotic response by phosphorylating
the prosurvival kinase Akt-protein kinase B on Ser473 (30,35,36). Furthermore, the activities of
mTORC1 and mTORC2 are closely linked, and they may be differently
influenced by rapamycin (30).
Rapamycin is an allosteric inhibitor of mTORC1 (37). However, mTORC2 is insensitive to
rapamycin (29,35,36). Short-term rapamycin treatment has
been indicated to acutely inhibit the activity of mTORC1 and
stimulate mTORC2 signaling through a negative-feedback mechanism.
However, prolonged rapamycin treatment reduces the level of mTORC2
by inhibiting its assembly (35,37). Due to its ability to suppress cell
growth and proliferation, rapamycin has been used as an
immunosuppressant, anticancer drug, and anti-restenosis agent in
coronary arterial stents (37).
In acute MI, rapamycin, or its analogue, limit cardiomyocyte death
and infarct size and attenuate cardiomyocyte hypertrophy and
cardiac remodeling (33,36,38). Although prolonged rapamycin
pretreatment disrupts the pro-survival effect of mTORC2 on
cardiomyocytes (36), rapamycin
has been revealed to inhibit cardiomyocyte apoptosis (38). In clinical practice, HF develops
from an old MI and follows a chronic course. Therefore, it is
important to determine whether prolonged rapamycin treatment
exhibits a potential therapeutic value in chronic postinfarction HF
through the regulation of cardiomyocyte death.
Recent studies have demonstrated that inhibiting
apoptosis and promoting autophagy in cardiomyocytes represent two
potential therapeutic strategies for use in HF (19,39,40). Additionally, ER stress and the
mTOR signaling network (mTORC1 and mTORC2 pathways) have been
indicated to act in a coordinative manner to regulate a variety of
cellular processes, including cell growth and survival (41-43). In the current study, using in
vivo (MI-induced chronic HF rat model) and in vitro
(angiotensin II-induced cardiomyocyte apoptosis model) experimental
approaches, whether rapamycin impacts cardiomyocyte apoptosis and
autophagy by affecting the crosstalk between mTOR signaling and ER
stress pathways was assessed.
Materials and methods
Reagents
Rapamycin and chloroquine (diphosphate salt) were
purchased from Sigma Aldrich (Merck KGaA). Angiotensin II was
purchased from Phoenix Pharmaceuticals (Burlingame).
Animals
All animal procedures were conducted in accordance
with the institutional guidelines for the care and use of
laboratory animals by Jilin University, Jilin, China. All
experimental procedures were approved by the Ethical Review Board
of China-Japan Union Hospital of Jilin University. Male Wistar rats
(age, 8 weeks; weight, 240-270 g) were obtained from the Center for
Laboratory Animals, Medical College, Jilin University, China.
Postinfarction HF was generated following a method
as previously described (39,44). Rats were subjected to sham surgery
or surgery involving the ligation of the left anterior descending
artery. Rats were then anesthetized using 100% oxygen containing 3%
isoflurane, which was supplied using a rodent respirator. Following
anesthetization, the thorax was opened in the left parasternal
area, and MI was induced by ligating the left anterior descending
coronary artery using a 3-0 suture between the pulmonary cone and
the left atrium. Following surgery, rats were randomly divided into
six groups, including the sham-, vehicle- and rapamycin-operated
groups, at 8 weeks (n=6, n=8 and n=8, respectively) or 12 weeks
post-MI (n=6, n=8 and n=8, respectively). After a period of 4
weeks, the successful induction of HF was confirmed using
echocardiography, and the animals in the rapamycin- and
vehicle-operated groups, at 8 weeks or 12 weeks post-MI, received
an intraperitoneal injection of rapamycin (1.4 mg-kg-day) dissolved
in dimethyl sulfoxide or vehicle control (equivalent volumes of
dimethyl sulfoxide diluted in normal saline) for 4 weeks. The dose
of rapamycin was selected based on the body surface area, as
described previously, and this dose has been indicated to be
effective and well tolerated in previous studies (45,46). At 8 and 12 weeks post-MI
induction, body weight and echocardiography were recorded. Animals
were then anesthetized using 100% oxygen containing 3% isoflurane
and euthanized via a rapid exsanguination from the abdominal aorta
and the removal of the hearts. Exsanguination was performed via an
abdominal aortic catheter, which permitted the free flow of blood,
and blood with a total volume of 7-9 ml per rat was rapidly removed
until no longer bleeding. The hearts were then quickly harvested
and washed with ice-cold normal saline, and subsequently blotted
with medical gauze. The left ventricle was dissected and fixed in
4% paraformaldehyde for histological evaluation, or snap frozen for
biochemical measurements.
Echocardiography
Rats were mildly anesthetized using 3% isoflurane,
and transthoracic echocardiography was performed using a Vivid-i
echocardiography machine (General Electric Company) equipped with
an 11.5-MHz transducer. The investigators who conducted the
echocardiography exam were blinded to the treatment groups.
Pathology
Histologic studies were conducted with 4%
paraformaldehyde-fixed and paraffin-embedded left ventricular
samples from rats of all six groups. Hematoxylin-eosin staining,
Masson's trichrome staining and immunohistochemical staining for
caspase-3 were conducted on cross sections of the left ventricle.
Images were acquired with a microscope (Olympus Corporation)
equipped with a digital camera and cellSens Dimension software
(version 1.16; Olympus Corporation).
Cell culture
The rat cardiomyocyte line H9c2 cells were purchased
from American Type Culture Collection (Manassas). H9c2 cells were
cultured in Dulbecco's modified Eagle's medium with a 1:1 mixture
of Ham's F-12 (DME-F-12 1:1; HyClone; GE Healthcare) supplemented
with 10% fetal bovine serum (Biological Industries,), 100 U-ml
penicillin (Gibco; Thermo Fisher Scientific, Inc.), and 100 mg-ml
streptomycin (Gibco; Thermo Fisher Scientific, Inc.). The cells
were cultured in a moist atmosphere with 5% CO2 and 95%
air at 37°C.
Cell viability measurement
Cell viability was assessed with a Cell Counting Kit
8 assay (Dojindo Laboratories). In brief, H9c2 cells were plated in
96-well plates at a density of 10,000 cells-well. After 24 h, the
cells were treated with graded concentrations of angiotensin II (0,
50, 100, 200, and 500 nM) for an additional 12, 24, or 36 h. Then,
10 µl of Cell Counting Kit 8 reagent was added to the
culture medium. After incubating the cells for 30 min, the plates
were read at OD 450 nm in a microplate spectrophotometer (BioTek
Instruments, Inc.).
Caspase-3 activity assay
The caspase-3 activity assay kit (Beyotime
Biotechnology) was used to confirm caspase-3 activity, which
recognizes the substrate acetyl-Asp-Glu-Val-Asp
p-nitroanilide (Ac-DEVD-pNA). The assay is mainly
based on quantification of the yellow p-nitroaniline at a
wavelength of 405 nm with a microplate spectrophotometer after
cleavage from the substrate Ac-DEVD-pNA. The assay was
carried out according to the kit instructions after preparing cell
lysates from H9c2 cardiomyocytes.
Flow cytometry
Cell survival and apoptosis were assessed by flow
cytometry with Annexin V-fluorescein isothiocyanate and propidium
iodide (KeyGEN BioTECH) staining. Analysis of phosphatidylserine on
the outer leaflet of apoptotic cell membranes was performed using
Annexin V-fluorescein isothiocyanate and propidium iodide to
identify apoptotic and necrotic cells, respectively. Briefly, the
cells were collected with ethylenediaminetetraacetic acid-free
trypsin and washed with PBS three times. Then, the cells were
resuspended in 500 µl of HEPES buffer, mixed with 5
µl of Annexin V-fluorescein isothiocyanate and 5 µl
of propidium iodide, and incubated at room temperature for 5 min in
the dark. The cells were analyzed using a flow cytometer (BD
Biosciences).
Immunofluorescence staining
Cells were seeded, treated, fixed, and stained
directly in 24-well plates. The cells were then washed twice with
PBS and fixed in ice-cold 100% methanol for 15 min at -20°C. After
rinsing with PBS three times for 5 min each, the samples were
incubated with 0.1% Triton X-100 at room temperature for 10 min,
blocked with 1% bovine serum albumin at room temperature for 60
min, and incubated with rabbit anti-light chain 3B (1:200; Cell
Signaling Technology, Inc.; cat. no. 3868) antibody at 4°C
overnight. The samples were then washed three times with PBS,
followed by incubation with Alexa Fluor 488-conjugated secondary
antibodies (1:1,000; Cell Signaling Technology, Inc.; cat. no.
4412) at room temperature for 60 min after
4′,6-diamidino-2-phenyl-indole counterstaining for 5 min. Images
were captured with a fluorescence microscope system (Olympus
Corporation).
Western blot analysis
Heart samples (left ventricular tissue of an area
far from the infarction site) and cardiomyocytes were homogenized
in ice-cold lysis buffer consisting of Tris 50 mM (pH 7.4), 150 mM
NaCl, 1% NP-40, 0.5% sodium deoxycholate and 0.1% sodium dodecyl
sulfate. The protein concentrations were determined by a
bicinchoninic acid assay (KeyGEN BioTECH) using bovine serum
albumin as a standard. Equal amounts of total protein were loaded
for resolution by sodium dodecyl sulfate-polyacrylamide gels under
reducing conditions. The primary antibodies against caspase-3
(1:1,000; cat. no. 9662), cleaved caspase-3 (1:500; cat. no. 4412),
phospho-S6 ribosomal protein (Ser235-236) (1:1,000; cat. no. 4412),
S6 ribosomal protein (1:1,000; cat. no. 4412), phospho-Akt (Ser473)
(1:1,000; cat. no. 4412), light chain 3B (1:1,000; cat. no. 4412),
SQSTM1-p62 (1:1,000; cat. no. 4412), Beclin-1 (1:1,000; cat. no.
3495), phospho-SAPK-JNK (Thr183-Tyr185) (1:1,000; cat. no. 4668),
SAPK-JNK (1:1,000; cat. no. 9252) and GAPDH (1:1,000; cat. no.
4412) were purchased from Cell Signaling Technology, Inc. The
primary antibody against caspase-12 (0.25 µg/ml; cat. no.
AF1456-SP) was purchased from R&D Systems, Inc. The primary
antibodies against DDIT3 (1:1,000; cat. no. ab179823) and GRP78
(1:2,000; cat. no. ab108613) were purchased from Abcam. The
HRP-conjugated secondary antibody (1:5,000; cat. no. BS13278) was
purchased from Bioworld Technology, Inc.
Statistical analysis
Data are expressed as the means ± S.E.M. Statistical
analysis was performed with PASW Statistics 18 software (version
18.0.0; IBM, Inc.). Differences between two groups (the sham- and
MI-operated groups at 4 weeks post-MI) were tested by two
independent samples t-test. Differences among groups were tested by
one-way ANOVA followed by post hoc comparisons with
Student-Newman-Keuls (S) and Tukey's post hoc tests. Differences
among groups for the long-term observations were tested by two-way
ANOVA followed by post hoc comparisons with Student-Newman-Keuls
(S) and Tukey's post hoc tests. Statistical graphing was performed
using GraphPad Prism software (version 7.04; GraphPad Software,
Inc., San Diego, CA, USA). For all analyses, P<0.05 was
considered significantly different.
Results
Rapamycin improves cardiac function in
rats with postinfarction HF
At 4 weeks following MI, a reduction in the
echocardiographic ejection fraction and fractional shortening
measurements was observed in MI animals compared with sham animals,
which indicated the damage to cardiac function and initiation of
chronic HF (Fig. 1A). At 8 weeks
post-MI, impaired cardiac function was observed in vehicle-treated
animals, and this impairment was prevented by rapamycin treatment
(Fig. 1B). Ejection fraction
measurements were significantly lower in the vehicle group compared
with the sham group, whereas in the rapamycin group, ejection
fraction measurements were higher compared with the vehicle group
(Fig. 1B). Similar results were
observed for fractional shortening measurements (Fig. 1B). The left ventricular internal
end diastolic dimension and left ventricular internal end systolic
dimension values were smaller in the rapamycin group compared with
the vehicle group (Fig. 1B). To
investigate the delayed effects of rapamycin on cardiac function,
the same set of animal groups were observed at 12 weeks following
MI (4 weeks after the end of rapamycin treatment). A significant
improvement in cardiac function was still present in the
rapamycin-treated group as assessed using the ejection fraction and
fractional shortening measurements (Fig. 1C).
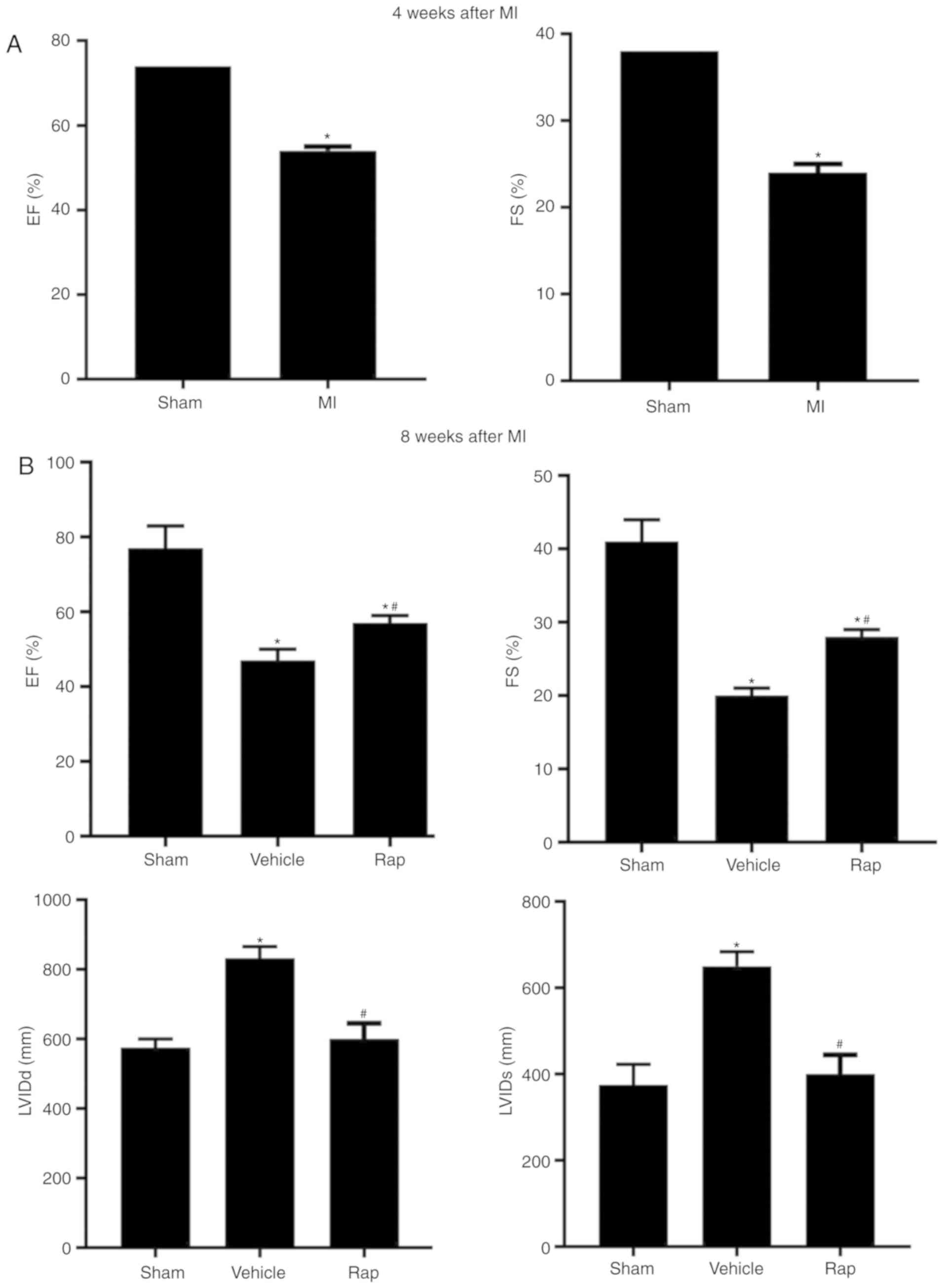 | Figure 1Echocardiographic measurements of
rats. (A) Echocardiographic EF and FS measurements of the sham- and
MI-operated groups at 4 weeks following MI. (B) EF, FS, LVIDd and
LVIDs values of the sham-, vehicle- and rapamycin-operated groups
at 8 weeks following MI. *P<0.05 vs. the sham group;
#P<0.05 vs. the vehicle group. Echocardiographic measurements of
rats. (C) EF and FS measurements of rats at 8 and 12 weeks
following MI. (D) Gross morphology of the hearts in the sham-,
vehicle- and rapamycin-operated groups at 8 weeks following MI. (E)
Hematoxylin and eosin staining of areas far from the infarction
site. (F) Masson's trichrome staining of left ventricles. (G)
Masson's trichrome staining of areas far from the infarction sites
and cardiomyocyte size measurements. *P<0.05 vs. the
sham group; #P<0.05 vs. the vehicle group. MI,
myocardial infarction; Rap, rapamycin; EF, ejection fraction; FS,
fractional shortening; LVIDd, left ventricular internal end
diastolic dimension; LVIDs, left ventricular internal end systolic
dimension. |
At 8 weeks following MI, the size of the heart was
slightly larger in the vehicle group compared with the sham group,
and myocardial dilation was less significant in the rapamycin group
compared with the vehicle group (Fig.
1D). Histopathology indicated that the left ventricle exhibited
increased extracellular matrix synthesis in the vehicle group
compared with the sham group, and this change was reduced in the
rapamycin group (Fig. 1E).
Rapamycin treatment also attenuated myocardial fibrosis of the left
ventricle when compared with vehicle treatment at 8 weeks following
MI (Fig. 1F). The results also
demonstrated that cardiomyocyte size in an area distant from the
site of infarction was largest in the vehicle group among all
groups, and this hypertrophy was prevented by the rapamycin
treatment (Fig. 1G).
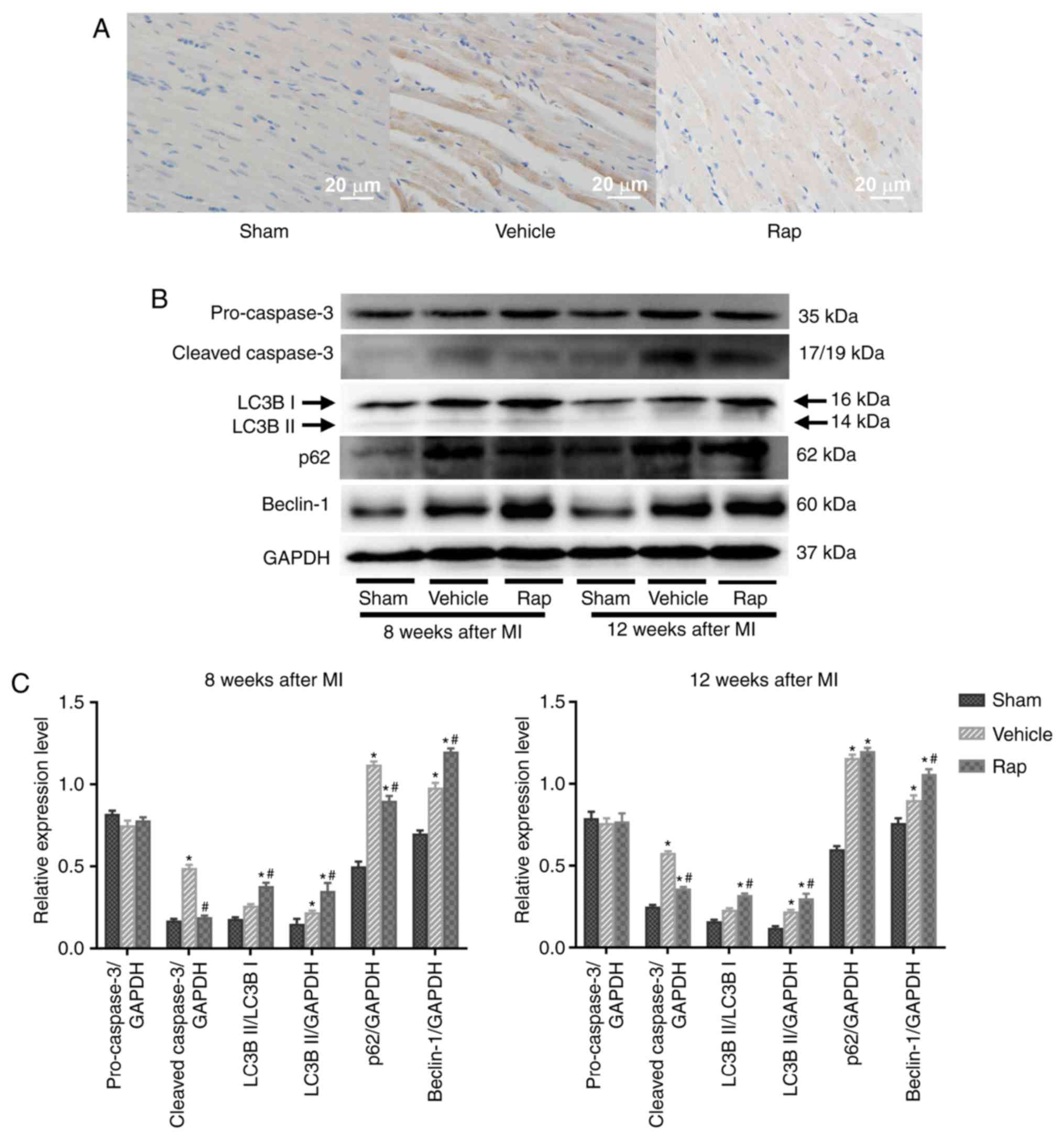
Rapamycin inhibits the expression of
cleaved caspase-3 and promotes cardiomyocyte autophagy in failing
hearts
Cleaved caspase-3 serves a critical role during
apoptosis. Cardiomyocyte apoptosis was evaluated in areas that were
distant from the infarction site of left ventricles, using
immunohistochemical staining for cleaved caspase-3. At 8 weeks
following MI, the level of cleaved caspase-3 was higher in the
vehicle group compared with the sham group, and this upregulation
was partly prevented by rapamycin treatment (Fig. 2A). This result was also identified
using western blot analysis (Fig. 2B
and C). Additionally, the levels of cleaved caspase-3 in the
vehicle group continued to increase up to 12 weeks following MI
(Fig. 2B). During the 4 weeks
after the end of rapamycin treatment, low but sustained inhibition
of cleaved caspase-3 was observed in the rapamycin treatment group,
as evaluated by immunoblot-ting for cleaved caspase-3 (Fig. 2B and C). Whether prolonged
rapamycin treatment promoted sustained and protective autophagy was
subsequently assessed by determining the conversion of light chain
3B I to light chain 3B II, an autophagosomal membrane protein used
as a marker of autophagy, and the protein levels of p62 and
Beclin-1. Beclin-1 is known to signal the onset of autophagy and
p62 is a well-known substrate of autophagy, and is reduced by the
lysosomal fusion of autophagosomes (23,47). The results indicated that the
baseline levels of light chain 3B II and Beclin-1 were increased in
the vehicle group, and enhanced by rapamycin treatment at 8 and 12
weeks post-MI (Fig. 2B and C).
Additionally, the levels of p62 were increased in the vehicle
group, and were decreased by rapamycin treatment at 8 weeks
post-MI. No significant difference in the levels of p62 was
identified between the vehicle- and rapamycin-operated groups at 12
weeks following MI.
Rapamycin inhibits the mTOR and ER stress
pathways in rats with chronic HF
To further explore the mechanisms underlying the
inhibitory effect of rapamycin on cardiomyo-cyte apoptosis, and the
stimulatory effect on cardiomyocyte autophagy, the involvement of
the mTOR signaling network and ER stress pathway was investigated.
The activities of mTORC1 and mTORC2 were assessed as indicated by
the phosphorylation states of their downstream targets, ribosomal
S6 protein and Akt, respectively. At 8 weeks following MI, the
phosphorylation of ribosomal S6 protein was activated in the
vehicle group, indicating the activation of protein synthesis and
cell growth following MI (Fig.
3A). Rapamycin administration inhibited the phosphorylation of
ribosomal S6 protein compared with vehicle treatment at 8 weeks
following MI (Fig. 3A).
Furthermore, treatment with rapamycin for 4 weeks decreased the
phosphorylation of Akt on Ser473 compared with the vehicle
treatment, indicating that the prosurvival activity of mTORC2 was
inhibited by rapamycin treatment (Fig. 3A). As shown in Fig. 2, although the prosurvival activity
of mTORC2 was inhibited, the level of cleaved caspase-3 was still
reduced by rapamycin treatment at 8 weeks following MI. After
discontinuing rapamycin treatment for 4 weeks, the level of
phosphoribosomal S6 protein was still decreased in the
rapamycin-treated group compared with the vehicle group, whereas
the inhibited phosphorylation of Akt on Ser473 was restored to
baseline level at 12 weeks following MI (Fig. 3A). Additionally, the level of
cleaved caspase-3 was still reduced by rapamycin treatment at 12
weeks following MI (Fig. 2).
Therefore, the anti-apoptotic effects of rapamycin may be mediated
through the inhibition of mTOR pathways, especially the mTORC1
pathway. Additionally, the inhibition of phosphoribosomal S6
protein by rapamycin was accompanied by the activation of
cardiomyocyte autophagy by rapamycin at 8 and 12 weeks following MI
(Fig. 2B and C, and Fig. 3A). The inhibitory effect of
rapamycin on cardiomyocyte apoptosis and the promoting effect on
cardiomyocyte autophagy may be mediated through the inhibition of
mTOR pathways.
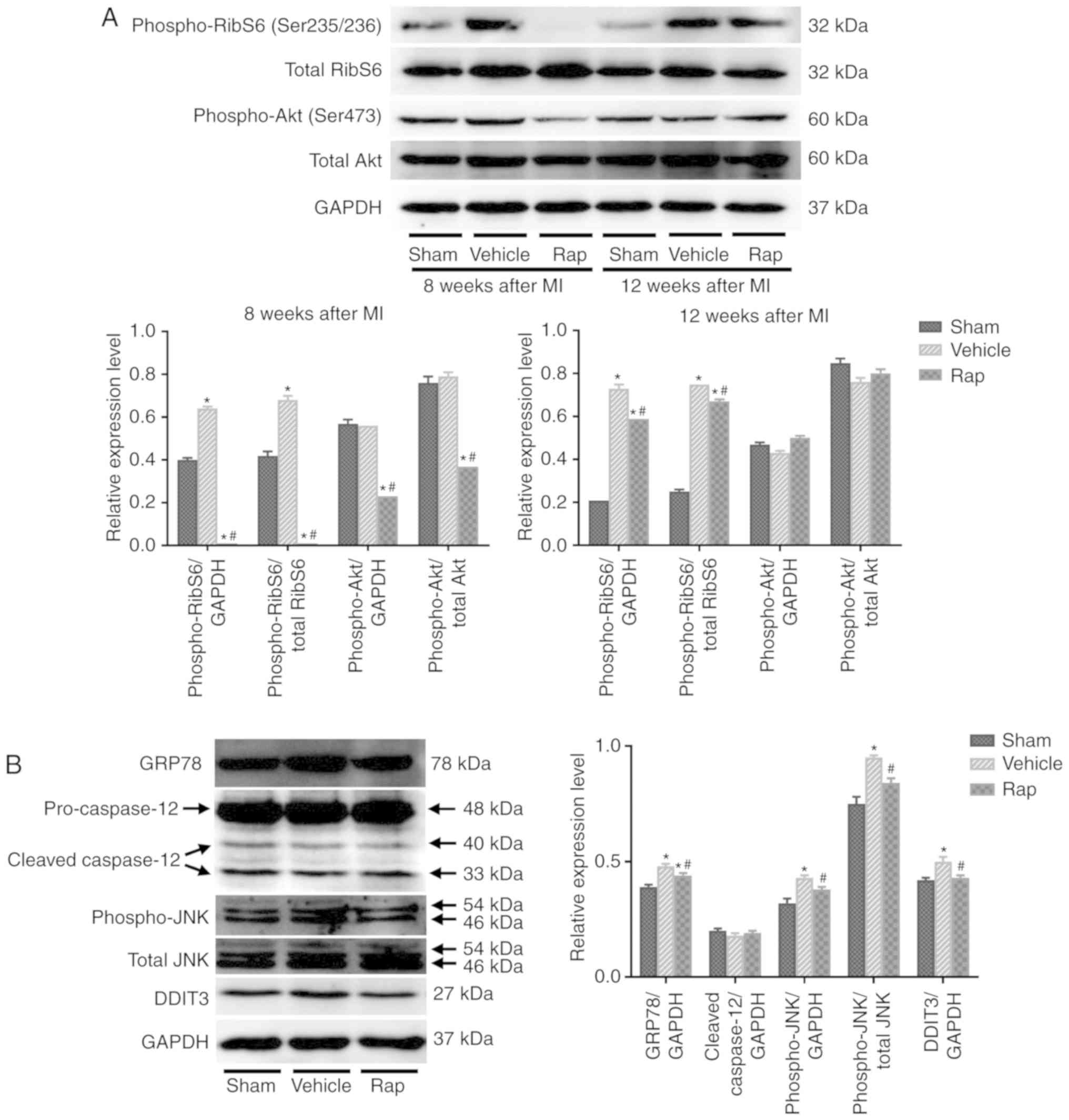 | Figure 3Rapamycin inhibits the mTOR and
endoplasmic reticulum stress pathways in hearts of HF rats at 8 and
12 weeks following MI. (A) Western blot of phospho-ribosomal S6
protein, total ribosomal S6, phospho-Akt and total Akt. (B) Western
blot of GRP78, cleavage of caspase-12, phospho-JNK, total JNK and
DDIT3. *P<0.05 vs. the corresponding sham group;
#P<0.05 vs. the corresponding vehicle group. HF,
heart failure; Rap, rapamycin; MI, myocardial infarction; RibS6,
ribosomal S6 protein; GRP78, glucose-regulated protein 78 kDa;
DDIT3, DNA damage-inducible transcript 3; JNK, c-JUN NH2-terminal
kinase. |
The mTOR and ER stress pathways are linked and they
serve important roles in cell growth and cell survival (42). Prolonged or severe ER stress that
is induced by the accumulation of unfolded proteins in the ER can
lead to cell apoptosis (16).
Whether the inhibition of mTOR by rapamycin prevented cardiomyocyte
apoptosis through the regulation of the ER stress pathway was
investigated in the current study. The effects of rapamycin on the
ER stress pathway were determined using the levels of GRP78,
caspase-12, phospho-JNK and DDIT3. At 8 weeks following MI,
increased levels of GRP78, phospho-JNK and DDIT3 were demonstrated
in the vehicle group, which coincided with the appearance of
increased cleaved caspase-3 expression (Figs. 2 and 3B). Furthermore, rapamycin treatment
reduced the levels of GRP78, phospho-JNK and DDIT3 compared with
vehicle treatment (Fig. 3B). No
significant differences in the levels of cleaved caspase-12 were
revealed among the sham-, vehicle- and rapamycin-operated groups.
The changes in the mTOR and ER stress pathways in hearts at 8 weeks
following MI indicated that rapamycin may prevent cardiomyocyte
apoptosis by inhibiting the ER stress pathway downstream of the
mTORC1 pathway, which concomitantly promoted pro-survival
cardiomyocyte autophagy.
Rapamycin prevents angiotensin II-induced
H9c2 cell apoptosis
As a neuroendocrine factor, angiotensin II serves an
important role in the pathogenesis of HF and induces protein
synthesis and cardiomyocyte hypertrophy, which causes the
activation of the ER stress and mTOR pathways (17,31, 48). Previous findings have demonstrated
that angiotensin II induces modest levels of cardiomyocyte
apoptosis (39,49), which is similar to the slight but
persistent cardiomyocyte apoptosis observed in failing hearts.
Therefore, in the current study, angiotensin II was used to create
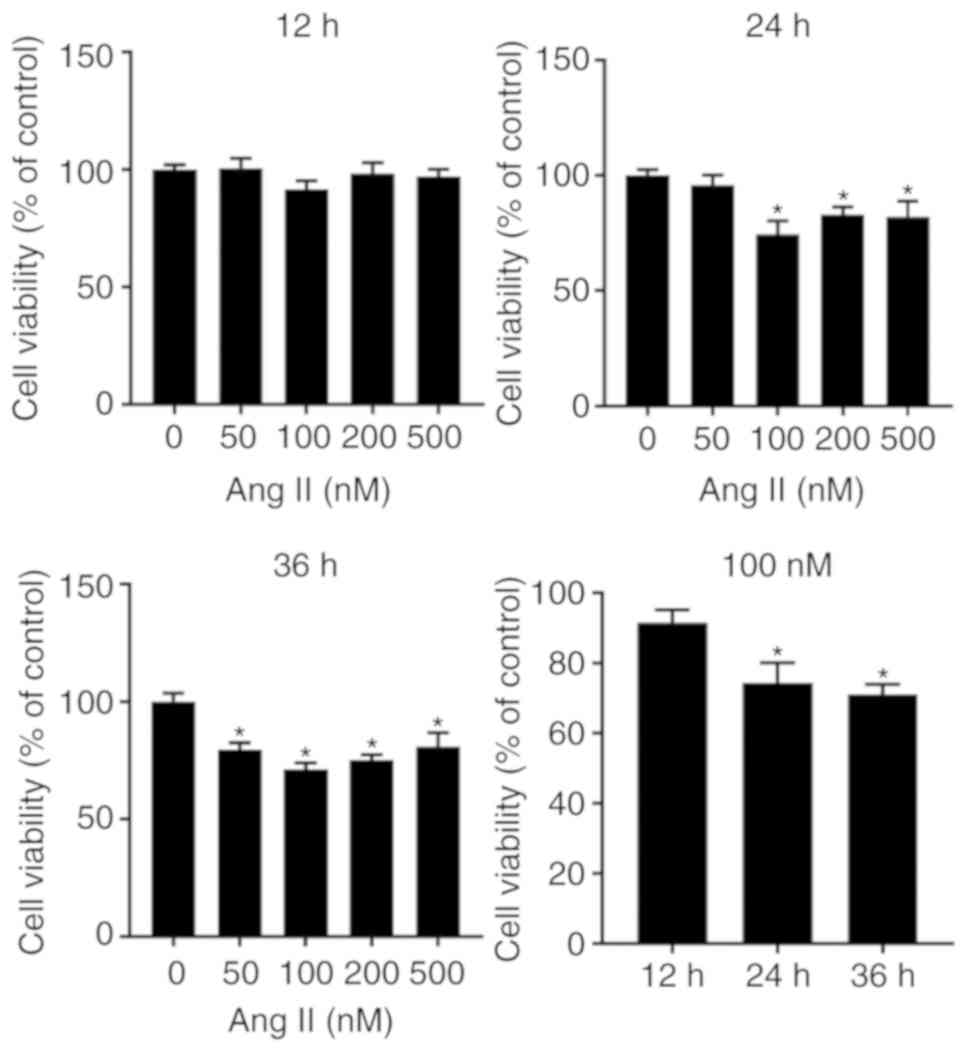
a pathological humoral environment for H9c2 cells. To determine the
optimal concentration of angiotensin II for inducing apoptosis, an
in vitro cell viability assay was performed, in which H9c2
cells were treated with graded concentrations of angiotensin II
(50-500 nM) for 12, 24 and 36 h. Angiotensin II was indicated to
reduce cell viability in a dose-independent and time-dependent
manner, and the peak effect was identified at a dose of 100 nM and
a time of 24-36 h (Fig. 4).
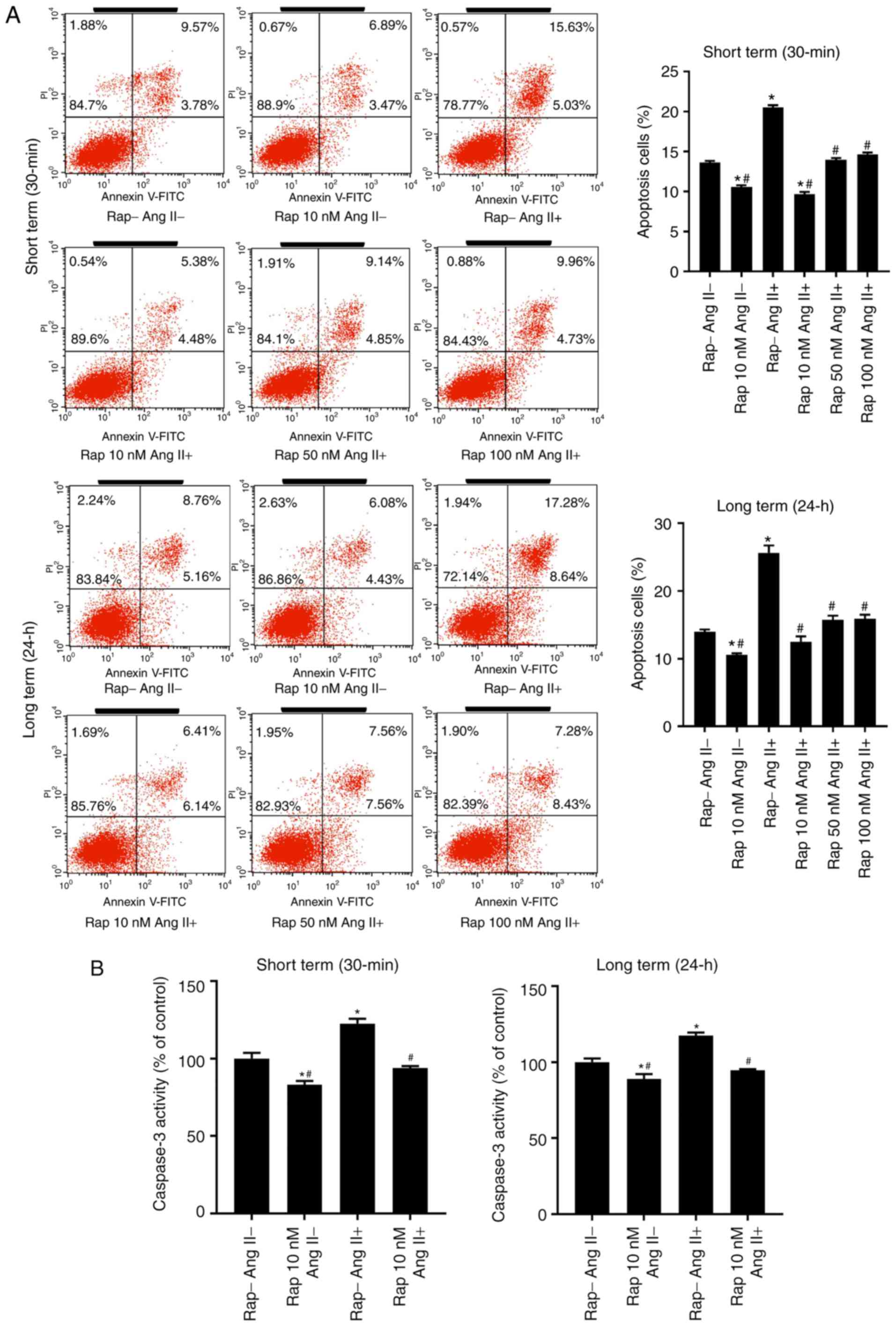
The effect of rapamycin on angiotensin II-induced
H9c2 cell apoptosis was subsequently assessed. Long-term rapamycin
treatment has been reported to inhibit mTORC2 function in a variety
of cell types (35,36). Therefore, to investigate the roles
of mTORC1 and mTORC2 in cell survival, H9c2 cells were pretreated
with various concentrations of rapamycin (10,50 and 100 nM) or
vehicle control (0.05% dimethyl sulfoxide) for 30 min (short term)
or 24 h (long term) prior to treatment with angiotensin II (100 nM)
for 24 h. Flow cytometry analysis demonstrated that rapamycin
administration decreased basal and angiotensin II-induced H9c2 cell
apoptosis in the groups treated for 30 min and 24 h, with a maximal
inhibition observed at a dose of 10 nM (Fig. 5A). This antiapoptotic effect of
rapamycin (10 nM) was reproduced in the caspase-3 activity assay
(Fig. 5B) and western blot
analysis of cleaved caspase-3 (Fig.
5C).
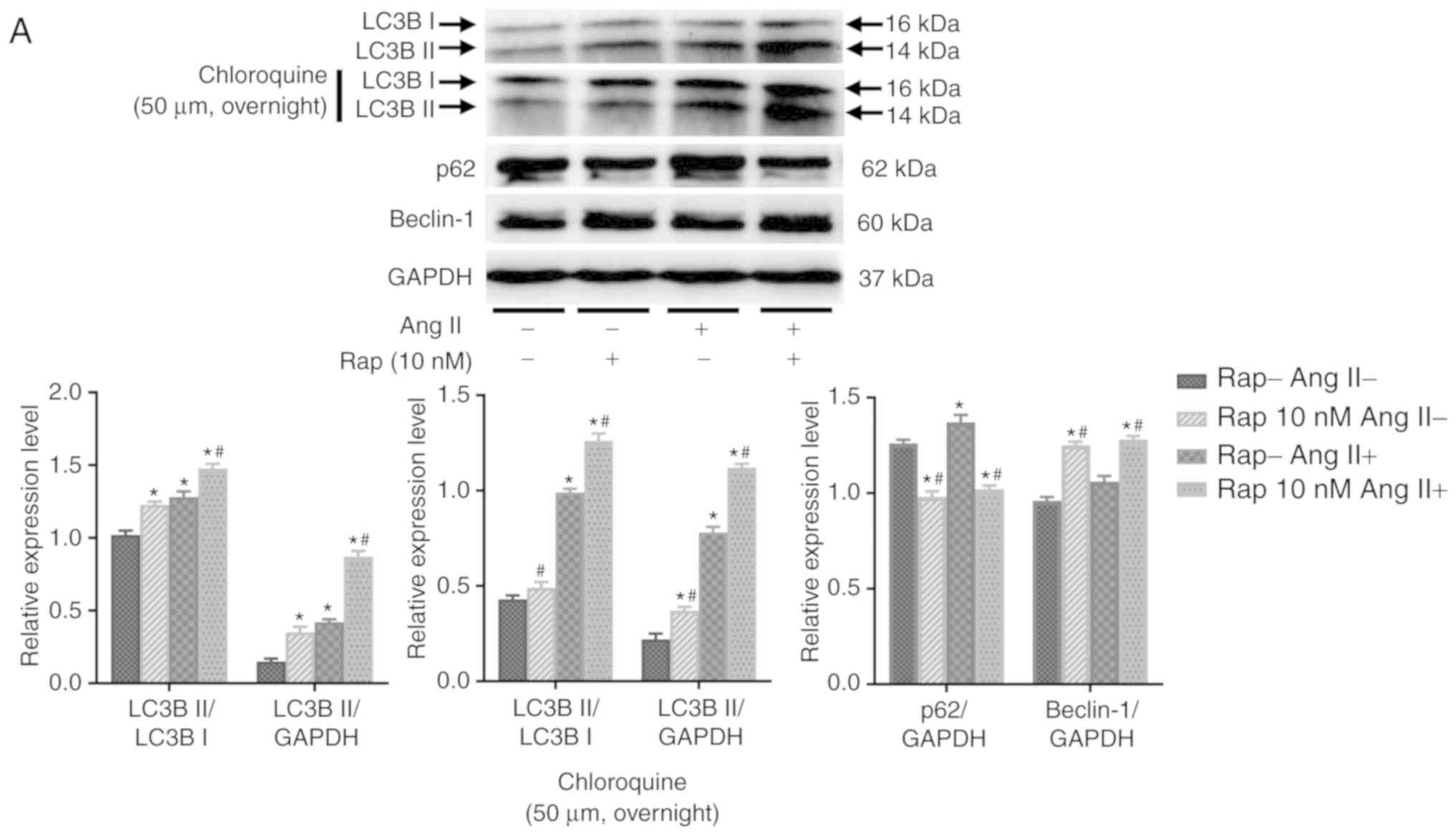
Effects of rapamycin on H9c2 cell
autophagic flux
To determine whether rapamycin-induced enhancement
in cardiomyocyte autophagy is mediated by the upregulation of
autophagosome formation or the inhibition of lysosomal degradation
of autophagosomes, the effects of rapamycin on H9c2 cell autophagic
flux were examined in an angio-tensin II-induced H9c2 cell
apoptosis model in the presence or absence of chloroquine (50
µM, overnight), which is an inhibitor of lysosomal activity.
As determined by the levels of light chain 3B II, rapamycin
treatment increased basal and angiotensin II-induced autophagy, and
these effects of rapamycin were further potentiated when
chloroquine was added in the group treated for 30 min (Fig. 6A). These results indicated that
the effect of rapamycin may not be due to the decreased degradation
of autophagosomes. This is also supported by the results of the
H9c2 cell autophagic flux assay using immuno-fluorescence staining
for light chain 3B, which indicated that autophagy was
significantly increased by rapamycin treatment when compared with
vehicle treatment with or without the administration of angiotensin
II in the 30 min group (Fig. 6B).
Additionally, p62 expression was decreased and the levels of
Beclin-1 were increased in rapamycin treatment group compared with
the vehicle control group with or without angiotensin II
pretreatment for 30 min (Fig.
6A).
Rapamycin inhibits the mTOR and ER stress
pathways in H9c2 cells
As presented in Fig.
7A, the phosphorylation of ribosomal S6 protein was stimulated
by angiotensin II alone (100 nM) for 24 h in 30 min and 24 h
rapamycin pretreatment groups, indicating that angiotensin II
increased protein synthesis in H9c2 cells via activation of the
mTORC1 pathway (Fig. 7A).
Pretreatment with rapamycin (10 nM) for 30 min or 24 h inhibited
the basal and angiotensin II-induced phosphorylation of ribosomal
S6 protein in H9c2 cells (Fig.
7A). Furthermore, treatment with rapamycin for 30 min
stimulated the phosphorylation of Akt on Ser473 in H9c2 cells
(Fig. 7A), and this stimulation
was not present after 24 h of treatment with rapamycin (Fig. 7A). These results indicated that
rapamycin treatment may prevent H9c2 cell apoptosis and promote
autophagy through the inhibition of mTORC1 pathways. In the current
study, it was revealed that administration of angiotensin II (100
nM) for 24 h induced GRP78 activation and an ER stress response in
H9c2 cells, which coincided with H9c2 cell apoptosis (Fig. 7B). Furthermore, rapamycin
treatment reduced basal and angiotensin II-induced GRP78 activation
in the 30 min group (Fig. 7B).
Rapamycin treatment also suppressed angiotensin II-induced DDIT3
activation and JNK phosphorylation in the 30 min group (Fig. 7B). The levels of phospho-JNK in
H9c2 cells did not change following the administration of
angiotensin II, and no caspase-12 cleavage was observed in H9c2
cells after the administration of angiotensin II or rapamycin
(Fig. 7B).
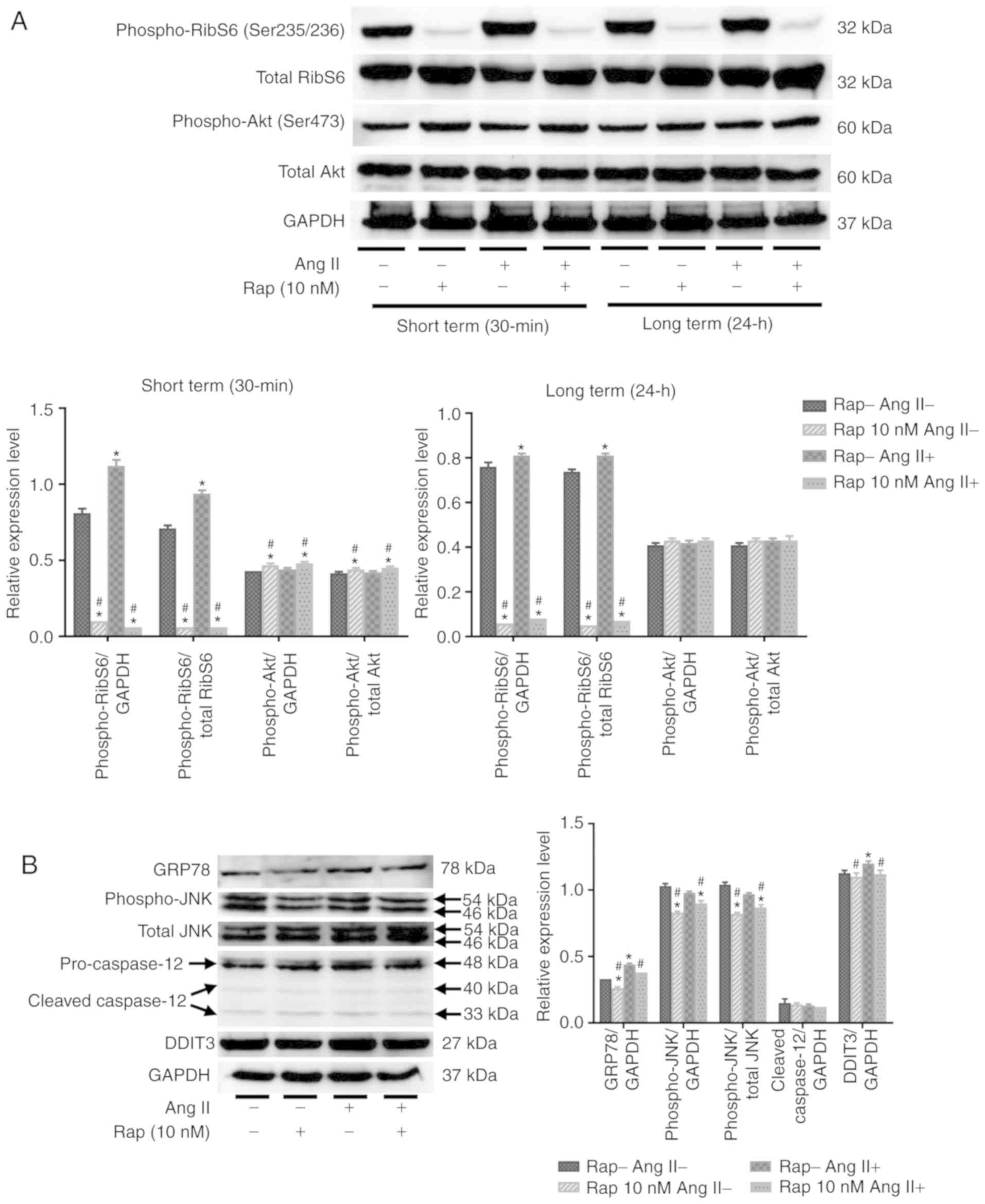 | Figure 7Rapamycin inhibits the mTOR and
endoplasmic reticulum stress pathways in H9c2 cells. (A) Western
blot analysis of phospho-ribosomal S6 protein, total ribosomal S6,
phospho-Akt and total Akt in H9c2 cells. (B) Western blot of GRP78,
cleavage of caspase-12, phospho-JNK, total JNK and DDIT3 in H9c2
cells. *P<0.05 vs. the corresponding control group;
#P<0.05 vs. the corresponding angiotensin II-treated
group. RibS6, ribosomal S6 protein; Ang II, angiotensin II; Rap,
rapamycin; GRP78, glucose-regulated protein 78 kDa; DDIT3, DNA
damage-inducible transcript 3; JNK, c-JUN NH2-terminal kinase. |
Discussion
Following MI, thinning and distension are initially
observed in infarcted and non-infarcted myocardium (44). Thinning of the infarcted wall is
followed by resultant scar formation, while hypertrophy develops in
non-infarcted myocardium in response to the increased stress and
neuroendocrine or growth factors (44). Cardiomyocyte apoptosis occurs
during the progression of myocardial hypertrophy and leads to
cardiomyocyte loss and HF (4,7,10,11,17,50). Recently, the involvement of
protein synthesis in cardiac remodeling and cardiomyocyte apoptosis
has received considerable attention (5,10,23). It may be suggested that the
sustained activation of protein synthesis with or without
insufficient protein degradation can lead to ER stress and ER
stress-induced cardiomyocyte apoptosis (15, 23). mTORC1 is a major regulator
associated with protein synthesis and degradation (30). On the one hand, mTORC1 promotes
protein synthesis by increasing ribosome production (30). On the other hand, mTORC1 is a key
complex that inhibits autophagy, a major mechanism associated with
cellular protein degradation, during stress and physiological and
pathological conditions (23,51). The binding of rapamycin to its
intracellular receptor, FKBP12, inhibits mTORC1 allosterically
(37). Therefore, rapamycin is
often used as an inhibitor of protein synthesis and cell growth to
examine its effects on myocardial hypertrophy or cardiac remodeling
in pressure-overloaded heart disease (52-55). However, the effects of rapamycin
on cardiac function and cardiomyocyte apoptosis in chronic HF
remain to be determined. The current study indicated that treatment
with rapamycin can inhibit the phosphorylation of ribosomal S6
protein and promote cardiomyocyte autophagy, protecting
cardiomyocytes from apoptosis by inhibiting the ER stress pathway
and improving cardiac function in chronic HF. It can be suggested
that the mTORC1 pathway may closely link to the ER stress pathway
via regulating the balance between cellular protein synthesis and
degradation. The inhibition of mTORC1 with rapamycin may inhibit ER
stress by reducing the newly synthesized protein load and
accelerating autophagy-induced unfolded protein degradation in the
ER, and in turn protect cardiomyocytes from apoptosis, improving
cardiac function in chronic HF.
ER is a highly dynamic organelle that serves
important roles in protein synthesis and apoptosis (17). The increased protein synthesis in
hypertrophic and failing hearts can lead to the accumulation of
unfolded proteins in the ER, which in turn trigger ER stress and
apoptosis (17). To prevent
unfolded protein accumulation, cells upregulate the transcription
of chaperones, including GRP78 and GRP94, reduce translation to
attenuate the burden from newly synthesized proteins and degrade
unfolded proteins that are accumulated in the ER (56). In the present study, ER stress was
determined by assessing the upregulation of GRP78. In rats with HF
and angiotensin II-treated H9c2 cells, GRP78 was upregulated, which
coincided with cardiomyocyte apoptosis, and the inhibition of mTOR
with rapamycin reduced the levels of GRP78 and cardiomyocyte
apoptosis in the in vitro and in vivo setting. These
results indicated that rapamycin may inhibit the ER stress pathway
downstream of the mTOR pathway and prevent cardiomyocyte apoptosis
caused by ER stress. Furthermore, prolonged or severe ER stress
induces apoptosis through the DDIT3-, JNK- and-or
caspase-12-dependent pathways (17,18). DDIT3, also known as CHOP, is a
component downstream of ER stress pathways (16). DDIT3 protein overexpression has
been indicated to induce apoptosis (16). JNK is a kinase that participates
in signal transduction in response to stressors (18). ER stress activates JNK, and the
sustained activation of the JNK pathway leads to apoptosis
(18). Caspase-12 is localized in
the ER membrane and is activated by ER stress but not death
receptor- or mitochondrion-mediated apoptotic pathways (18). The results of the current study
indicated that DDIT3 was induced in rats with failing hearts and in
angiotensin II-treated H9c2 cells, and was suppressed by rapamycin
treatment. Furthermore, the JNK pathway is closely associated with
the mTORC1 pathway and has been indicated to be preferentially
activated downstream of mTORC1 over other ER stress pathways
(43). Although the activation of
JNK was observed in failing hearts but not in angiotensin
II-treated H9c2 cells, rapamycin treatment inhibited the
phosphorylation of JNK in both cases. However, caspase-12 cleavage
was not induced in failing hearts and angiotensin II-treated H9c2
cells. Although different ER stress inducers or courses may
differentially impact the modulation of the three different ER
stress pathways, failing hearts and angiotensin II-treated H9c2
cells increased ER stress, which contributed to cardiomyocyte
apoptosis, while the inhibition of mTORC1 with rapamycin
significantly prevented ER stress-induced cardiomyocyte apoptosis
in vivo and in vitro.
The potential cardioprotective effects of rapamycin
against cardiomyocyte hypertrophy during pressure overload have
been previously demonstrated (52-55). In the context of chronic ischemic
injury, Buss et al (33).
demonstrated that everolimus prevented myocardial hypertrophy and
cardiac remodeling through inhibition of the mTORC1 pathway.
However, Völkers et al (36). revealed that prolonged rapamycin
treatment (18 h) inhibited the prosurvival mTORC2 signaling and
impaired the cell death-inhibiting effect of rapamycin in neonatal
ventricular cardiomyocytes following an acute oxidative injury (50
µM H2O2 for 4 h). Although prolonged
rapamycin treatment (1 week prior and 2 days following MI)
inhibited mTORC2 signaling, a study indicated that the
cardioprotective effects of rapamycin against myocardial
hypertrophy and cardiomyocyte apoptosis were still observed in the
mice suffering from chronic ischemic injury (38). In the present study, it was
revealed that short-term rapamycin treatment inhibited the activity
of mTORC1 and stimulated the activity of mTORC2 in H9c2 cells,
while prolonged rapamycin treatment inhibited the mTORC1 and mTORC2
pathways in rats with chronic HF. Additionally, the activity of
mTORC2 was restored to baseline level, and the activity of mTORC1
was still partly inhibited by rapamycin after discontinuing
rapamycin treatment for 4 weeks in rats with chronic HF. This
delayed inhibitory effect of rapamycin on mTORC1 may be associated
with its allosteric inhibition of mTORC1 (mTORC1 needs to be newly
synthesized). The cardioprotective effects of rapamycin on cardiac
function and cardiomyocyte apoptosis were continuously observed in
the rapamycin-treated groups at 8 and 12 weeks post-MI. These
results indicated that mTORC1 inhibition and mTORC2 activation may
contribute to the anti-apoptotic effect of short-term rapamycin
treatment, but the inhibition of mTORC1 may be the primary
mechanism for the cardioprotective effects of prolonged rapamycin
treatment. The inhibition of protein synthesis and the activation
of autophagy-induced protein renewal may be the major mechanism
underlying the anti-apoptotic effect of rapamycin in chronic
HF.
In conclusion, although the inhibitors of the
neurohormonal systems, including angiotensin-converting enzyme
inhibitors, β-receptor blockers and aldosterone antagonists have
been widely used over the past 20 years, the incidence of chronic
postinfarction HF remains high. The results of the current study
demonstrated that rapamycin effectively prevented cardiomyocyte
apoptosis, promoted cardiomyocyte autophagy and improved cardiac
function via regulating the crosstalk between the mTORC1 and ER
stress pathways in chronic postinfarction HF. However, the
potential clinical application of rapamycin requires further
investigation.
Abbreviations:
|
HF
|
heart failure
|
|
MI
|
myocardial infarction
|
|
mTOR
|
mechanistic target of rapamycin
|
|
mTORC1
|
mechanistic target of rapamycin
complex 1
|
|
mTORC2
|
mechanistic target of rapamycin
complex 2
|
|
ER
|
endoplasmic reticulum
|
|
GRP78
|
glucose-regulated protein 78 kDa
|
|
DDIT3
|
DNA damage-inducible transcript 3
|
|
JNK
|
c-JUN NH2-terminal kinase
|
Acknowledgments
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant number 81570360); and the Fund
of Key Laboratory of Myocardial Ischemia, Ministry of Education
(grant number KF201815).
Availability of data and materials
The datasets used and-or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PY and GG conceived and designed the study. GG, WC
and MY performed the experiments. JL, HL and CW collected the data
and prepared the Figures. GG analyzed the data and drafted the
manuscript. PY edited the language of the manuscript. All authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
All animal procedures were conducted in accordance
with the institutional guidelines for the care and use of
laboratory animals by Jilin University, Jilin, China. All
experimental procedures were approved by the Ethical Review Board
of China-Japan Union Hospital of Jilin University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Metra M and Teerlink JR: Heart failure.
Lancet. 390:1981–1995. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cahill TJ, Choudhury RP and Riley PR:
Heart regeneration and repair after myocardial infarction:
Translational opportunities for novel therapeutics. Nat Rev Drug
Discov. 16:699–717. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Teringova E and Tousek P: Apoptosis in
ischemic heart disease. J Transl Med. 15:872017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Whelan RS, Kaplinskiy V and Kitsis RN:
Cell death in the pathogenesis of heart disease: Mechanisms and
significance. Annu Rev Physiol. 72:19–44. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tham YK, Bernardo BC, Ooi JY, Weeks KL and
McMullen JR: Pathophysiology of cardiac hypertrophy and heart
failure: Signaling pathways and novel therapeutic targets. Arch
Toxicol. 89:1401–1438. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ziaeian B and Fonarow GC: Epidemiology and
aetiology of heart failure. Nat Rev Cardiol. 13:368–378. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Narula J, Haider N, Virmani R, DiSalvo TG,
Kolodgie FD, Hajjar RJ, Schmidt U, Semigran MJ, Dec GW and Khaw BA:
Apoptosis in myocytes in end-stage heart failure. N Engl J Med.
335:1182–1189. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hein S, Arnon E, Kostin S, Schönburg M,
Elsässer A, Polyakova V, Bauer EP, Klövekorn WP and Schaper J:
Progression from compensated hypertrophy to failure in the
pressure-overloaded human heart: Structural deterioration and
compensatory mechanisms. Circulation. 107:984–991. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wencker D, Chandra M, Nguyen K, Miao W,
Garantziotis S, Factor SM, Shirani J, Armstrong RC and Kitsis RN: A
mechanistic role for cardiac myocyte apoptosis in heart failure. J
Clin Invest. 111:1497–1504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Abbate A, Biondi-Zoccai GG and Baldi A:
Pathophysiologic role of myocardial apoptosis in post-infarction
left ventricular remodeling. J Cell Physiol. 193:145–153. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Olivetti G, Abbi R, Quaini F, Kajstura J,
Cheng W, Nitahara JA, Quaini E, Di Loreto C, Beltrami CA, Krajewski
S, et al: Apoptosis in the failing human heart. N Engl J Med.
336:1131–1141. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kostin S, Pool L, Elsässer A, Hein S,
Drexler HC, Arnon E, Hayakawa Y, Zimmermann R, Bauer E, Klövekorn
WP and Schaper J: Myocytes die by multiple mechanisms in failing
human hearts. Circ Res. 92:715–724. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dickhout JG, Carlisle RE and Austin RC:
Interrelationship between cardiac hypertrophy, heart failure, and
chronic kidney disease: Endoplasmic reticulum stress as a mediator
of pathogenesis. Circ Res. 108:629–642. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kolwicz SC Jr, Purohit S and Tian R:
Cardiac metabolism and its interactions with contraction, growth,
and survival of cardiomyocytes. Circ Res. 113:603–616. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Minamino T, Komuro I and Kitakaze M:
Endoplasmic reticulum stress as a therapeutic target in
cardiovascular disease. Circ Res. 107:1071–1082. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim I, Xu W and Reed JC: Cell death and
endoplasmic reticulum stress: Disease relevance and therapeutic
opportunities. Nat Rev Drug Discov. 7:1013–1030. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Okada K, Minamino T, Tsukamoto Y, Liao Y,
Tsukamoto O, Takashima S, Hirata A, Fujita M, Nagamachi Y, Nakatani
T, et al: Prolonged endoplasmic reticulum stress in hypertrophic
and failing heart after aortic constriction: Possible contribution
of endoplasmic reticulum stress to cardiac myocyte apoptosis.
Circulation. 110:705–712. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Oyadomari S, Araki E and Mori M:
Endoplasmic reticulum stress-mediated apoptosis in pancreatic
beta-cells. Apoptosis. 7:335–345. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Takemura G, Kanamori H, Okada H, Miyazaki
N, Watanabe T, Tsujimoto A, Goto K, Maruyama R, Fujiwara T and
Fujiwara H: Anti-apoptosis in nonmyocytes and pro-autophagy in
cardio-myocytes: Two strategies against postinfarction heart
failure through regulation of cell death-degeneration. Heart Fail
Rev. 23:759–772. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kanamori H, Takemura G, Goto K, Maruyama
R, Ono K, Nagao K, Tsujimoto A, Ogino A, Takeyama T, Kawaguchi T,
et al: Autophagy limits acute myocardial infarction induced by
permanent coronary artery occlusion. Am J Physiol Heart Circ
Physiol. 300:H2261–H2271. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kanamori H, Takemura G, Goto K, Maruyama
R, Tsujimoto A, Ogino A, Takeyama T, Kawaguchi T, Watanabe T,
Fujiwara T, et al: The role of autophagy emerging in postinfarction
cardiac remodelling. Cardiovasc Res. 91:330–339. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sciarretta S, Zhai P, Shao D, Maejima Y,
Robbins J, Volpe M, Condorelli G and Sadoshima J: Rheb is a
critical regulator of autophagy during myocardial ischemia:
Pathophysiological implications in obesity and metabolic syndrome.
Circulation. 125:1134–1146. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ghosh R and Pattison JS: Macroautophagy
and chaperone-mediated autophagy in heart failure: The known and
the unknown. Oxid Med Cell Longev. 2018:86020412018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang C, Yitzhaki S, Perry CN, Liu W,
Giricz Z, Mentzer RM Jr and Gottlieb RA: Autophagy induced by
ischemic preconditioning is essential for cardioprotection. J
Cardiovasc Transl Res. 3:365–373. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
De Meyer GR and Martinet W: Autophagy in
the cardiovascular system. Biochim Biophys Acta. 1793:1485–1495.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gurusamy N, Lekli I, Gorbunov NV,
Gherghiceanu M, Popescu LM and Das DK: Cardioprotection by
adaptation to ischaemia augments autophagy in association with
BAG-1 protein. J Cell Mol Med. 13:373–387. 2009. View Article : Google Scholar
|
|
27
|
Bhuiyan MS, Pattison JS, Osinska H, James
J, Gulick J, McLendon PM, Hill JA, Sadoshima J and Robbins J:
Enhanced autophagy ameliorates cardiac proteinopathy. J Clin
Invest. 123:5284–5297. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nakai A, Yamaguchi O, Takeda T, Higuchi Y,
Hikoso S, Taniike M, Omiya S, Mizote I, Matsumura Y, Asahi M, et
al: The role of autophagy in cardiomyocytes in the basal state and
in response to hemodynamic stress. Nat Med. 13:619–624. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Laplante M and Sabatini DM: mTOR signaling
in growth control and disease. Cell. 149:274–293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sciarretta S, Forte M, Frati G and
Sadoshima J: New insights into the role of mTOR signaling in the
cardiovascular system. Circ Res. 122:489–505. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sadoshima J and Izumo S: Rapamycin
selectively inhibits angiotensin II-induced increase in protein
synthesis in cardiac myocytes in vitro. Potential role of 70-kD S6
kinase in angiotensin II-induced cardiac hypertrophy. Circ Res.
77:1040–1052. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Simm A, Schlüter K, Diez C, Piper HM and
Hoppe J: Activation of p70(S6) kinase by beta-adrenoceptor agonists
on adult cardio-myocytes. J Mol Cell Cardiol. 30:2059–2067. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Buss SJ, Muenz S, Riffel JH, Malekar P,
Hagenmueller M, Weiss CS, Bea F, Bekeredjian R, Schinke-Braun M,
Izumo S, et al: Beneficial effects of Mammalian target of rapamycin
inhibition on left ventricular remodeling after myocardial
infarction. J Am Coll Cardiol. 54:2435–2446. 2009. View Article : Google Scholar
|
|
34
|
Li Q, Xie J, Li R, Shi J, Sun J, Gu R,
Ding L, Wang L and Xu B: Overexpression of microRNA-99a attenuates
heart remodelling and improves cardiac performance after myocardial
infarction. J Cell Mol Med. 18:919–928. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sarbassov DD, Ali SM, Sengupta S, Sheen
JH, Hsu PP, Bagley AF, Markhard AL and Sabatini DM: Prolonged
rapamycin treatment inhibits mTORC2 assembly and Akt-PKB. Mol Cell.
22:159–168. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Völkers M, Konstandin MH, Doroudgar S,
Toko H, Quijada P, Din S, Joyo A, Ornelas L, Samse K, Thuerauf DJ,
et al: Mechanistic target of rapamycin complex 2 protects the heart
from ischemic damage. Circulation. 128:2132–2144. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Benjamin D, Colombi M, Moroni C and Hall
MN: Rapamycin passes the torch: A new generation of mTOR
inhibitors. Nat Rev Drug Discov. 10:868–880. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Di R, Wu X, Chang Z, Zhao X, Feng Q, Lu S,
Luan Q, Hemmings BA, Li X and Yang Z: S6K inhibition renders
cardiac protection against myocardial infarction through PDK1
phos-phorylation of Akt. Biochem J. 441:199–207. 2012. View Article : Google Scholar
|
|
39
|
Liu M, Mao C, Li J, Han F and Yang P:
Effects of the Activin A-follistatin system on myocardial cell
apoptosis through the endoplasmic reticulum stress pathway in heart
failure. Int J Mol Sci. 18:2017.
|
|
40
|
Kanamori H, Takemura G, Goto K, Tsujimoto
A, Ogino A, Takeyama T, Kawaguchi T, Watanabe T, Morishita K,
Kawasaki M, et al: Resveratrol reverses remodeling in hearts with
large, old myocardial infarctions through enhanced
autophagy-activating AMP kinase pathway. Am J Pathol. 182:701–713.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ji Y, Luo X, Yang Y, Dai Z, Wu G and Wu Z:
Endoplasmic reticulum stress-induced apoptosis in intestinal
epithelial cells: A feed-back regulation by mechanistic target of
rapamycin complex 1 (mTORC1). J Anim Sci Biotechnol. 9:382018.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Appenzeller-Herzog C and Hall MN:
Bidirectional crosstalk between endoplasmic reticulum stress and
mTOR signaling. Trends Cell Biol. 22:274–282. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kato H, Nakajima S, Saito Y, Takahashi S,
Katoh R and Kitamura M: mTORC1 serves ER stress-triggered apoptosis
via selective activation of the IRE1-JNK pathway. Cell Death
Differ. 19:310–320. 2012. View Article : Google Scholar
|
|
44
|
Goldman S and Raya TE: Rat infarct model
of myocardial infarction and heart failure. J Card Fail. 1:169–177.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Shioi T, McMullen JR, Tarnavski O,
Converso K, Sherwood MC, Manning WJ and Izumo S: Rapamycin
attenuates load-induced cardiac hypertrophy in mice. Circulation.
107:1664–1670. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bishu K, Ogut O, Kushwaha S, Mohammed SF,
Ohtani T, Xu X, Brozovich FV and Redfield MM: Anti-remodeling
effects of rapamycin in experimental heart failure: Dose response
and interaction with angiotensin receptor blockade. PLoS One.
8:e813252013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Klionsky DJ, Abdalla FC, Abeliovich H,
Abraham RT, Acevedo-Arozena A, Adeli K, Agholme L, Agnello M,
Agostinis P, Aguirre-Ghiso JA, et al: Guidelines for the use and
interpretation of assays for monitoring autophagy. Autophagy.
8:445–544. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Schunkert H, Sadoshima J, Cornelius T,
Kagaya Y, Weinberg EO, Izumo S, Riegger G and Lorell BH:
Angiotensin II-induced growth responses in isolated adult rat
hearts. Evidence for load-independent induction of cardiac protein
synthesis by angiotensin II. Circ Res. 76:489–497. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yan M, Yang S, Meng F, Zhao Z, Tian Z and
Yang P: MicroRNA 199a-5p induces apoptosis by targeting JunB. Sci
Rep. 8:66992018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang D, Contu R, Latronico MV, Zhang J,
Rizzi R, Catalucci D, Miyamoto S, Huang K, Ceci M, Gu Y, et al:
MTORC1 regulates cardiac function and myocyte survival through
4E-BP1 inhibition in mice. J Clin Invest. 120:2805–2816. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jung CH, Ro SH, Cao J, Otto NM and Kim DH:
mTOR regulation of autophagy. FEBS Lett. 584:1287–1295. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
McMullen JR, Sherwood MC, Tarnavski O,
Zhang L, Dorfman AL, Shioi T and Izumo S: Inhibition of mTOR
signaling with rapamycin regresses established cardiac hypertrophy
induced by pressure overload. Circulation. 109:3050–3055. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Song X, Kusakari Y, Xiao CY, Kinsella SD,
Rosenberg MA, Scherrer-Crosbie M, Hara K, Rosenzweig A and Matsui
T: mTOR attenuates the inflammatory response in cardiomyocytes and
prevents cardiac dysfunction in pathological hypertrophy. Am J
Physiol Cell Physiol. 299:C1256–C1266. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ikeda M, Ide T, Fujino T, Matsuo Y, Arai
S, Saku K, Kakino T, Oga Y, Nishizaki A and Sunagawa K: The
Akt-mTOR axis is a pivotal regulator of eccentric hypertrophy
during volume overload. Sci Rep. 5:158812015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Harston RK, McKillop JC, Moschella PC, Van
Laer A, Quinones LS, Baicu CF, Balasubramanian S, Zile MR and
Kuppuswamy D: Rapamycin treatment augments both protein
ubiquitination and Akt activation in pressure-overloaded rat
myocardium. Am J Physiol Heart Circ Physiol. 300:H1696–H1706. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mori K: Tripartite management of unfolded
proteins in the endoplasmic reticulum. Cell. 101:451–454. 2000.
View Article : Google Scholar : PubMed/NCBI
|