Introduction
Platelets are non-nucleated cells that are produced
in the cytoplasm of intramedullary megakaryocytes (1). These platelets circulate throughout
the bloodstream, performing hemostasis and preventing bleeding in
conjunction with coagulation factors. However, these platelet
functions produce blood clots that potentially block arterial,
microvascular and capillary blood flows, causing cardiovascular
diseases (CVDs), including angina pectoris, myocardial infarction
and atherosclerosis (2).
Collagen is a protein abundantly present in the
extracellular matrix of the blood vessel wall, providing
flexibility to withstand the outward forces exerted by arterial
pressure (3). Under normal blood
flow situations, collagen is not exposed to the lumen. However,
when the collagen leaks into the lumen, due to damage or sclerosis
of the blood vessel walls by CVD risk factors, glycoprotein (GP)-VI
and GP-IX-V complex receptors on the surface of platelets bind to
the collagen (4). Fc receptor-γ
chain and Src family kinase associated with GP-VI lead to the
phosphorylation and activation of phospholipase Cγ2,
catalyzing the hydrolysis of phosphatidylinositol 4,5-bisphosphate
to diacylglycerol and inositol trisphosphate (IP3). In
such instances, agonist-binding receptors induce granule release,
integrin activation, thromboxane A2 (TXA2)
production, and as a consequence, platelet aggregation (5,6).
Deep sea water (DSW) is an abundant sea resource
located at depths as deep as 200 m below sea level. It contains
high concentrations of minerals, such as magnesium (Mg), calcium
(Ca), potassium (K) and sodium (Na), as well as several trace
minerals. Currently, DSW is being developed in Korea, Japan and
Taiwan for use in various applications in the aquaculture,
agriculture and food industries (7,8).
Although various industries are trying to adopt and use DSW, such
as the food industry (e.g., mineral water, deep sea water salt,
confectioneries, etc.), there is a lack of evidence that DSW has
biologically beneficial functions. The biological effects of DSW on
obesity, hypertension, inflammation, atopic eczema/dermatitis
syndrome and cancer have been reported in several studies (7,9-12).
Moreover, DSW has been shown to exert suppressive effects on
hyperlipidemia and atherosclerosis in animals fed a high
cholesterol diet (13-15). Previously, it was demonstrated
that DSW inhibited doxorubicin-induced epithelial-mesenchymal
transition by inhibiting the extracellular signal-activated protein
kinase (ERK1/2), p38 and PI3K/Akt signaling pathways (15). Mitogen-activated protein kinases
(MAPKs), such as the ERK, c-Jun N-terminal kinase (JNK) and p38,
and Akt signaling pathways, play primary roles in the amplification
of platelet aggregation, granule secretion and integrin
αIIbβ3 activation induced by agonists
(16-18). However, to the best of our
knowledge, there is no study available to date describing a role of
DSW in the inhibition of platelet activation, which is another
cause of CVDs, including angina pectoris, myocardial infarction and
atherosclerosis.
In this study, since the original DSW is high in Na
and requires hardness control, experiments were performed using a
mineral-balanced DSW (MBDSW). The anti-platelet activity of MBDSW
was investigated using collagen- and thrombin-induced platelet
aggregation. Furthermore, whether MBDSW inhibits granule secretion,
platelet-activating molecules, integrin and the MAPK and Akt
signaling pathways was also evaluated.
Materials and methods
Materials and reagents
Collagen and thrombin were purchased from the
Chrono-Log Co. The thromboxane B2 (TXB2)
ELISA kit, Cyclic AMP ELISA kit and dipyridamole were purchased
from Cayman Chemical Co. The CytoTox 96® Non-Radioactive
Cytotoxicity assay (cat. no. G1780) was purchased from Promega. The
serotonin ELISA kit was purchased from Labor Diagnostika Nord GmbH
& Corp. Fura-2/acetoxymethyl ester (AM) and Alexa Fluor
488-conjugated fibrinogen were purchased from Molecular Probes. APC
anti-human CD62P (P-selectin, cat. no. 304910) antibody was
purchased from BioLegend Inc. The adenosine triphosphate (ATP)
assay kit was purchased from Biomedical Research Service Center.
Protease inhibitor cocktail and phosphatase inhibitor cocktail were
purchased from GenDEPOT, and antibodies against ERK1/2 (cat. no.
4695), phospho-ERK1/2 (Thr202/Tyr204; cat.
no. 4370), p38 (cat. no. 8690), phospho-p38 (cat. no. 4511), JNK
(cat. no. 9258), phospho-JNK (cat. no. 4668), Akt (cat. no. 4691),
phospho-Akt (Ser473; cat. no. 9271), phospho-Akt
(Thr308; cat. no. 2965), vasodilator-stimulated
phosphoprotein (VASP; cat. no. 3112), phospho-VASP
(Ser157; cat. no. 3111), phospho-inositol
1,4,5-triphosphate receptor (IP3R; cat. no. 8548) and
glyceraldehyde 3-phosphate dehydrogenase (GAPDH; cat. no. 5174)
were purchased from Cell Signaling Technology. β-actin antibody
(cat. no. sc-69879) was purchased from Santa Cruz Biotechnology.
LY294002 and U0126 were purchased from LC laboratories.
HRP-conjugated goat anti-rabbit IgG (cat. no. 31460) and
bi-cinchoninic acid (BCA) protein assay kits were purchased from
Pierce Biotechnology. Polyvinylidene fluoride (PVDF) membrane was
purchased from PallLife Sciences.
Preparation of MBDSW
MBDSW was obtained from the Seawater Utilization
Plant Research Center in the Research Institute of Ships and Ocean
Engineering. DSW was pumped from a depth of 500 m in the East Sea
of Korea. To remove salts and minerals, it underwent reverse
osmosis filtration and electrodialysis, as previously described
(14). Refined minerals were then
added to the treated MBDSW to generate a hardness of 3,428 (H3,428)
containing 711.72 mg/l Mg, 203.84 mg/l Ca, 39.65 mg/l Na and 17.31
mg/l K (Table I). After
manufacturing the MBDSW (Mg:Ca ratio, 3:1), there is a slight
difference in the analysis (Mg:Ca ratio, 3.49:1). MBDSW (H3,428)
was then diluted with filtered MBDSW (H0) to prepare MBDSW of
various hardness levels. The hardness (H) was calculated using the
following formula: Hardness=[Mg (mg/l) ×4.1] + [Ca (mg/l) ×2.5], as
previously described (14).
 | Table IMineral content of original DSW and
MBDSW used in this study. |
Table I
Mineral content of original DSW and
MBDSW used in this study.
| Major elements | MBDSW (mg/l) | Original DSW
(mg/l) |
|---|
| Mg | 711.72 | 1,269 |
| Ca | 203.84 | 421 |
| K | 17.31 | 396 |
| Na | 39.65 | 10,567 |
| Pb | ND | <0.005 |
| Hg | ND | ND |
| Cd | ND | <0.001 |
| Sr | 3.55 | 1.68 |
Preparation of washed human platelets and
measurement of platelet aggregation
Once the platelet plug has been formed by the
activated platelets, the coagulation factors presented in plasma
are activated in a sequence of events known as the 'coagulation
cascade', which leads to the formation of fibrin from fibrinogen
plasma protein (19). Therefore,
this study evaded all response to any secondary hemostasis effects
using the washed platelet aggregation method. This study used blood
samples obtained from healthy volunteers (17 to 59 years of age).
This study was approved by the Dongguk University Gyeongju
Institutional Review Board (DUG IRB 20180014-04). Volunteer written
consent was waivered in this study. A total of 13 human
platelet-rich plasma (PRP) with an acid-citrate-dextrose solution
(0.8% citric acid, 2.2% sodium citrate and 2.45% glucose) was
collected and obtained from the Korean Red Cross Blood Center
(Daegu, Korea) from July, 2018 to December, 2018. Human PRP was
centrifuged for 10 min at 125 × g at 25°C to remove other cells and
then centrifuged for 10 min at 1,300 × g at 25°C to obtain platelet
pellets. The platelet pellets were washed twice with washing buffer
(138 mM NaCl, 2.7 mM KCl, 12 mM NaHCO3, 0.36 mM
NaH2PO4, 5.5 mM glucose and 1 mM
Na2EDTA, pH 6.5) and then resuspended in a suspension
buffer (138 mM NaCl, 2.7 mM KCl, 12 mM NaHCO3, 0.36 mM
NaH2PO4, 0.49 mM MgCl2, 5.5 mM
glucose and 0.25% gelatin, pH 6.9) to a final concentration of
1×108 cells/ml, as previously described (20). All of the above-mentioned
procedures were performed at 25°C to avoid any effects (i.e.,
platelet activation) related to temperature.
Platelet aggregation was measured using an
aggregometer (Chrono-Log, Corp.) at a constant stirring speed of 50
× g. In the experiment, platelets were pre-incubated with MBDSW at
37°C in the presence of 2 mM CaCl2 prior to the addition
of collagen (3 µg/ml) or thrombin (0.05 U/ml). Each
aggregation rate (%) was recorded for 5 min.
Measurement of lactate dehydrogenase
Platelet cytotoxicity was determined by the leakage
of lactate dehydrogenase (LDH) from the cytosol. Washed human
platelets (108 cells/ml) were incubated with various
concentrations of MBDSW (H250 to H2,000) for 30 min at 37°C, and
then centrifuged with 2,000 × g for 5 min at room temperature. The
supernatant (50 µl) incubated with cytotoxicity assay
reagent mix (50 µl) for 30 min at room temperature was
measured using a SpectraMax M2e (Molecular Devices). A maximal
value of cytotoxicity was recorded in 0.05% triton X-100.
Percentage cytotoxicity was calculated as follows: Cytotoxicity
(%)=100× (sample LDH release/maximum LDH release).
Measurement of cytosolic calcium ion
concentration
Human PRP was incubated with Fura-2/AM (5 µM)
at 37°C for 1 h in the dark. Washed, Fura-2/AM-loaded human
platelets were prepared using the procedure described above, and
1×108 platelets/ml were pre-treated with MBDSW for 2 min
at 37°C and then stimulated with collagen (3 µg/ml) for 5
min. Fluorescence was recorded with a spectrofluorometer (SFM 25;
Bio-Teck Instruments) with an excitation wavelength that changed
every 0.5 sec from 340 to 380 nm; the emission wavelength was set
at 510 nm. The calcium ion concentration
([Ca2+]i) was calculated using the method
developed by Schaeffer and Blaustein (21). The following formula was used:
[Ca2+]i=224 nM ×
(F−Fmin)/(Fmax−F), where 224 nM
is the dissociation constant of Fura-2-Ca2+ complex, and
Fmin and Fmax are fluorescence
intensities at low and high Ca2+ concentrations,
respectively.
Measurement of serotonin release
Platelet aggregation was terminated after 5 min of
stimulation by the addition of 5 mM ice-cold EDTA followed by
centrifuging the samples (5,000 × g, 3 min, at 4°C). The
supernatant was used to determine the level of serotonin release.
Samples were reacted with acylation buffer for 15 min at room
temperature. Acylated samples were incubated with serotonin
antiserum in the serotonin microtiter strips for 15 min at room
temperature. The level of serotonin release was measured using a
SpectraMax M2e (Molecular Devices) microplate reader.
Measurement of ATP release
Platelet aggregation was stopped after 5 min of
stimulation, by adding 5 mM ice-cold EDTA followed by centrifuging
the samples (5,000 × g, 3 min, at 4°C). The supernatant was used to
determine the level of ATP release. Samples were reacted with ATP
assay solution. The level of ATP release was measured using a
SpectraMax M2e (Molecular Devices) microplate reader.
Measurement of P-selectin expression and
fibrinogen binding to integrin αIIbβ3
Platelets, pre-incubated with MBDSW (H250 to
H2,000), were stimulated with collagen (3 µg/ml) at 37°C in
the presence of 2 mM CaCl2. The platelets were
centrifuged (1,500 × g, 3 min, at 4°C) consecutively followed by
fixation using 0.5% paraformaldehyde. The fixed platelets were then
washed 3 times via centrifugation at 2,000 × g for 5 min, at 4°C
and resuspended in cool phosphate-buffered saline (PBS) containing
5% bovine serum albumin (BSA) and 1% sodium azide. Platelets were
incubated with FITC-conjugated CD62P antibody (4 µg/ml) and
Alexa Fluor 488-conjugated human fibrinogen (200 µg/ml) in
3% bovine serum albumin/PBS for 30 min at 4°C in the dark.
Subsequently, flow cytometric analysis was performed on a
FACSCalibur II flow cytometer using CellQuest software.
Measurement of cyclic AMP level
Platelets, pre-incubated with MBDSW (H250 to H2,000)
or dipyridamole (40 µM) for 2 min at 37°C, were stimulated
with collagen (3 µg/ml) for 5 min at 37°C in the presence of
2 mM CaCl2. A sample was then added 80% ice-cold ethanol
and underwent vortexing for pre-treatment. The sample was left at
room temperature for 5 min and centrifuged at 2,000 × g for 10 min
at 4°C. Samples were reacted with cAMP antiserum and cAMP tracer in
a mouse anti-rabbit IgG-coated plate overnight at 4°C. The level of
cAMP in the supernatant was determined using a SpectraMax M2e
(Molecular Devices) microplate reader.
Measurement of TXA2
production
Platelets, pre-incubated with MBDSW, were stimulated
with collagen (3 µg/ml) at 37°C in the presence of 2 mM
CaCl2. Platelet aggregation was terminated after 5 min
of stimulation by the addition of 5 mM ice-cold EDTA including 0.2
mM indomethacin followed by centrifuging the samples (5,000 × g, 3
min, at 4°C). Supernatants were diluted 1:500 and used in assays.
Samples were reacted with TXB2 antiserum and
TXB2 tracer in a mouse anti-rabbit IgG-coated plate
overnight at 4°C. The level of TXB2 in the supernatant
was determined using a SpectraMax M2e (Molecular Devices)
microplate reader.
Western blot analysis
Platelets, pre-incubated with MBDSW (H250 to
H2,000), dipyridamole (40 µM), LY294002 (2 and 20 µM)
or U0126 (2 and 20 µM) for 2 min at 37°C, were stimulated
with collagen (3 µg/ml) for 5 min at 37°C in the presence of
2 mM CaCl2. Platelet aggregation was terminated after 5
min of stimulation by the addition of protease and phosphatase
inhibitor-containing RIPA lysis buffer (50 mM pH 7.5 Tris-HCl, 150
mM NaCl, 1% Triton X-100, 1% sodium deoxycholate, 0.1% SDS and 2 mM
EDTA). The protein concentration was determined by BCA assay; an
equal amount of protein in platelet lysates was separated via 8 to
12% SDS-polyacrylamide gel electrophoresis and then transferred
onto a PVDF membrane. The PVDF membrane was blocked with 1% BSA in
TBS buffer. The primary antibody (1:3,000) was reacted overnight at
4°C, after which, a secondary antibody (1:5,000) reaction was
performed for 2 h at room temperature. The protein bands were
visualized with a chemiluminescence substrate and photographed
using a LAS-4000 (Fujifilm) luminescent image analyzer.
Densitometric analysis was performed using image J version 1.6.0_20
(National Institutes of Health) with data from at least 3
independent experiments.
Statistical analysis
The experimental results are presented as the means
± standard deviation (SD) values accompanied by the number of
observations. Statistical significance was determined by analyzing
variance. Significant differences among groups in experiments were
assessed by undertaking analysis of variance (ANOVA). When the
ANOVA results indicated significant differences among group means,
the groups were compared by the Newman-Keuls method. A value of
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of MBDSW on collagen- and
thrombin-induced human platelet aggregation, and cytotoxicity
Platelet aggregation was determined using a light
transmission aggregometer. The concentration of collagen used was 3
µg/ml, which was the threshold concentration required to
induce maximal aggregation (Fig.
S1). As the threshold concentration of thrombin (0.2 U/ml) is a
very high concentration, the concentration of thrombin used was
0.05 U/ml. The effects of various hardness levels of MBDSW on
platelet aggregation induced by collagen and thrombin were examined
and it was observed that MBDSW inhibited platelet aggregation
induced by collagen (3 µg/ml) and thrombin (0.05 U/ml) in a
hardness-dependent manner (Fig. 1A
and B). Subsequently, it was confirmed that the presence of
calcium in MBDSW did not automatically affect platelet aggregation
(Fig. 1C). Furthermore, Ca plus
Mg, equivalent to MBDSW (H250 and H1,000), inhibited platelet
aggregation induced by collagen (3 µg/ml) (Fig. S2). In addition, LDH assay revealed
that MBDSW (H250 to H2,000) incubated with the platelets for 30 min
did not increase LDH leakage in the platelets (Fig. 1D). These results suggested that
MBDSW inhibited platelet aggregation without affecting
cytotoxicity.
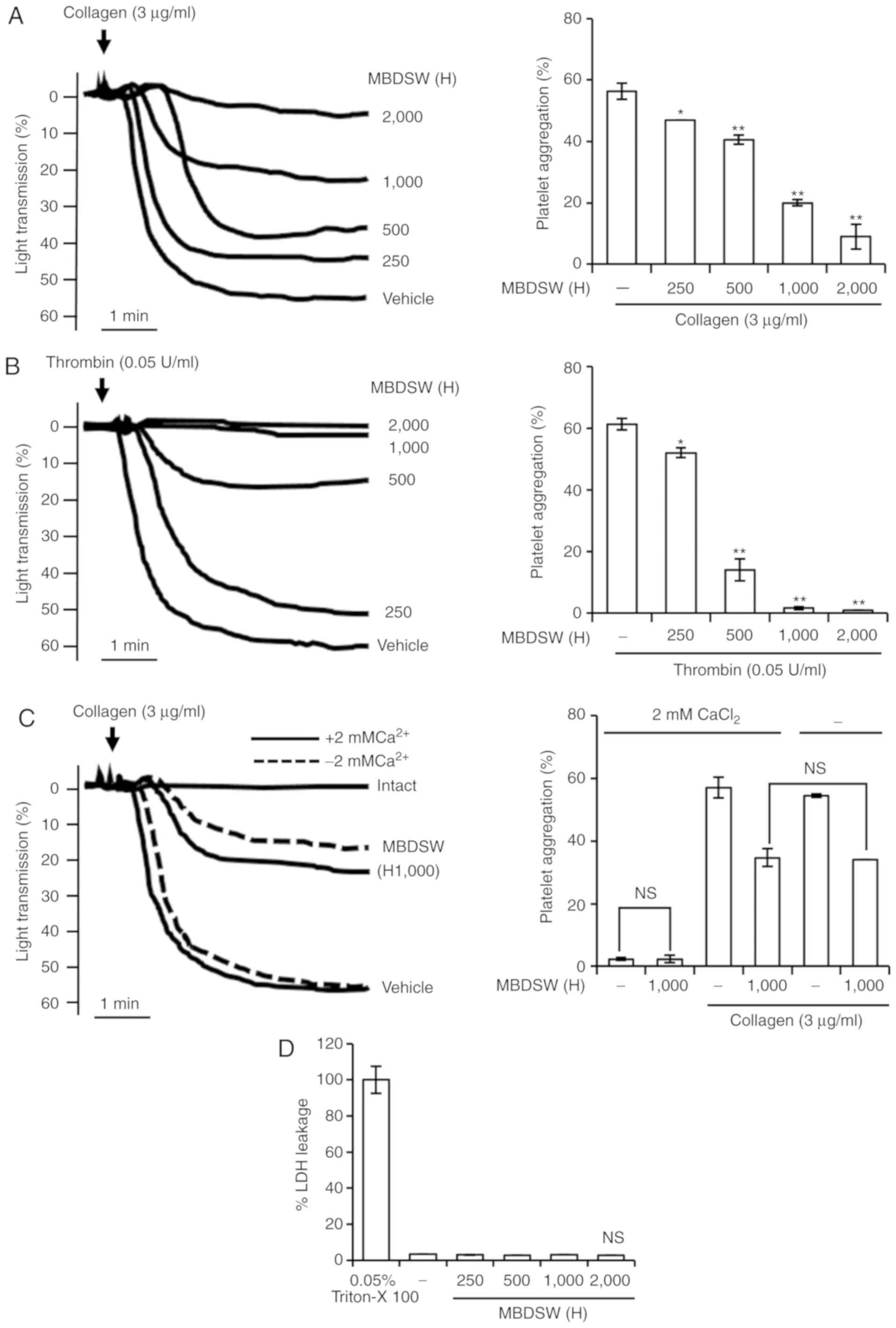 | Figure 1Inhibitory activity of MBDSW on
agonist-induced human platelet aggregation, and cytotoxicity. (A)
Washed human platelets (1×108 cells/ml) were
pre-incubated with MBDSW at various levels of hardness (H250, 500,
1,000 and 2,000) and subsequently treated with collagen (3
µg/ml) to induce platelet aggregation. (B) Washed human
platelets (1×108 cells/ml) were pre-incubated with MBDSW
at various levels of hardness (H250, 500, 1,000 and 2,000) and
subsequently treated with thrombin (0.05 U/ml) to induce platelet
aggregation. (C) Washed human platelets (1×108 cells/ml)
were pre-incubated with MBDSW (H1,000) or the vehicle in the
presence or absence of 2 mM CaCl2 and subsequently
treated with collagen (3 µg/ml). (D) Effects of MBDSW on
cytotoxicity. MBDSW (H) was at the ratio of Mg:Ca, 3:1, and
hardness. All data are presented as the means ± SD (n=3). Vehicle
is collagen only. NS, not significant. *P<0.05 and
**P<0.001 vs. the vehicle. H, hardness; MBDSW,
mineral-balanced deep sea water. |
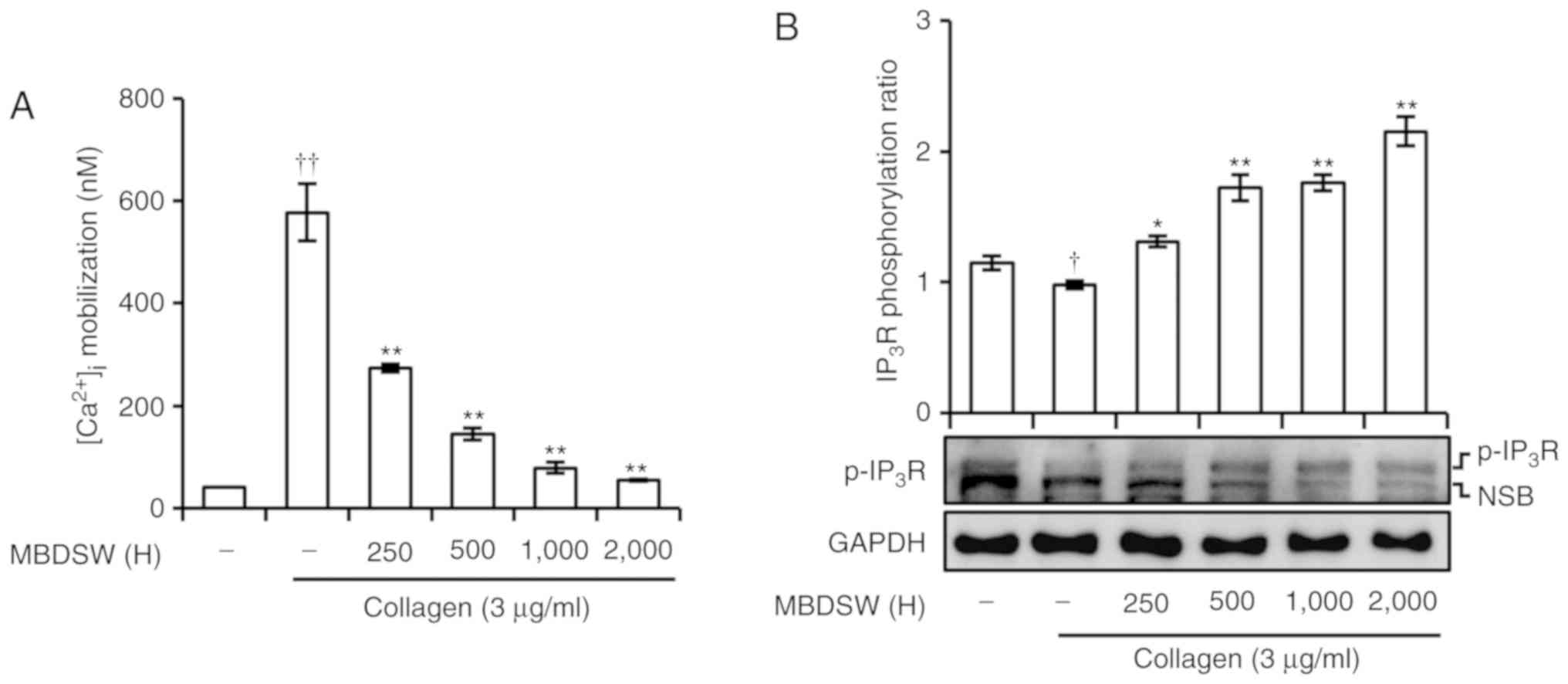
MBDSW reduces collagen-induced
[Ca2+]i mobilization and increases
IP3R phosphorylation
Intracellular calcium
([Ca2+]i) is considered an important molecule
for platelet activation, such as the inside-out signaling
activation of integrin αIIbβ3 and thrombus
formation (22). Thus, the
inhibitory effects of MBDSW on the collagen-induced increase in
[Ca2+]i levels were investigated.
Pre-treatment of the platelets with various hardness levels of
MBDSW significantly prevented the collagen-induced increase in
[Ca2+]i levels (Fig. 2A). To reveal the inhibitory
mechanisms of MBDSW associated with [Ca2+]i
mobilization, the association between IP3R and
[Ca2+]i mobilization was evaluated. As shown
in Fig. 2B, MBDSW increased
IP3R phosphorylation in a hardness-dependent manner.
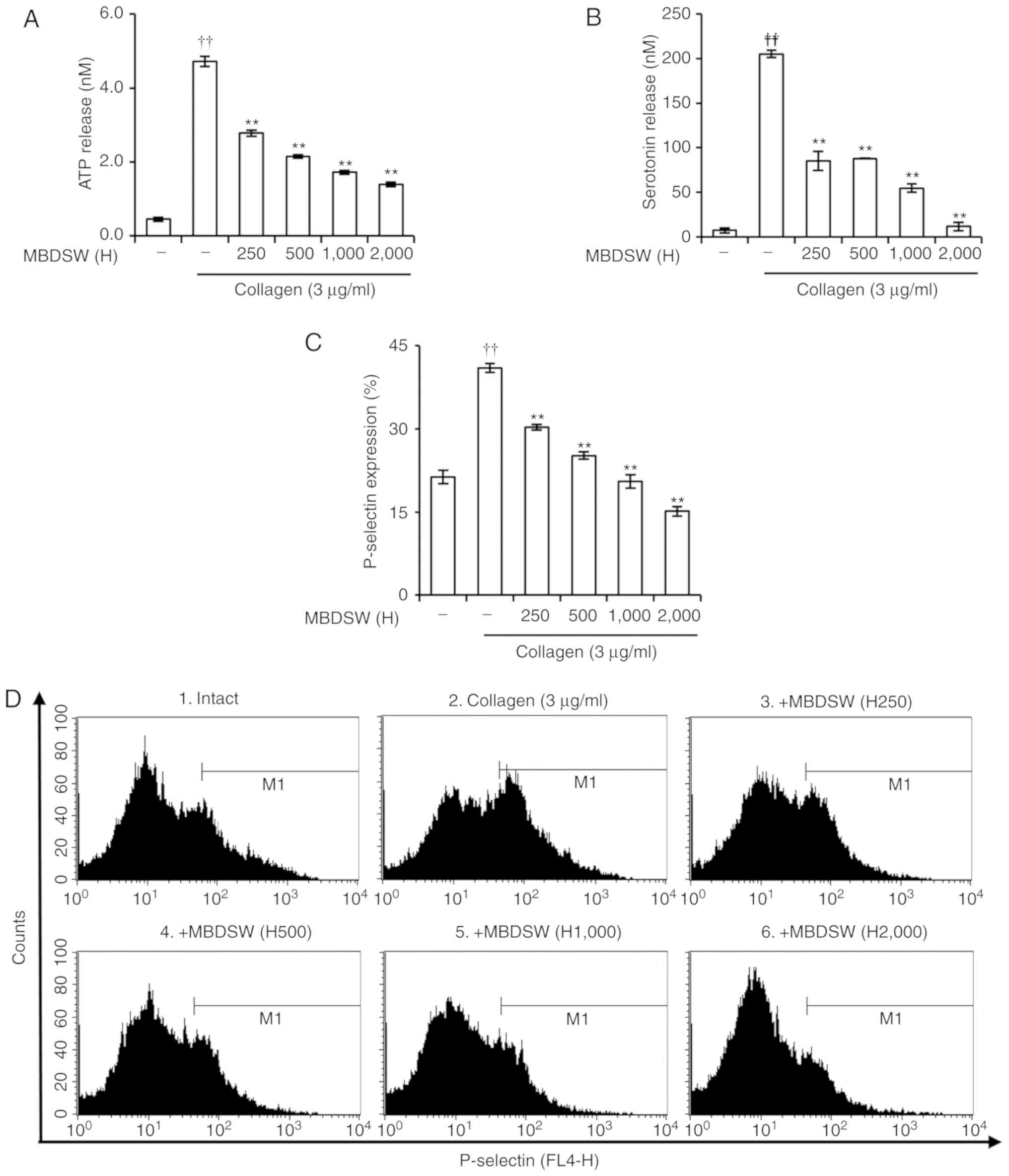
MBDSW reduces granule secretion
When platelets are activated via agonist
stimulation, platelets release granules. Therefore, it was
investigated whether MBDSW affects α-granule and dense granule
secretion in collagen-induced platelet aggregation. Collagen (3
µg/ml) alone markedly increased ATP and serotonin secretion
from dense granules. However, the ATP and serotonin secretions were
significantly decreased by MBDSW in a hardness-dependent manner
(Fig. 3A and B). To determine
whether MBDSW inhibits α-granule secretion, P-selectin expression
was examined by flow cytometry. The results revealed that MBDSW
significantly inhibited P-selectin expression in a
hardness-dependent manner (Fig. 3C
and D).
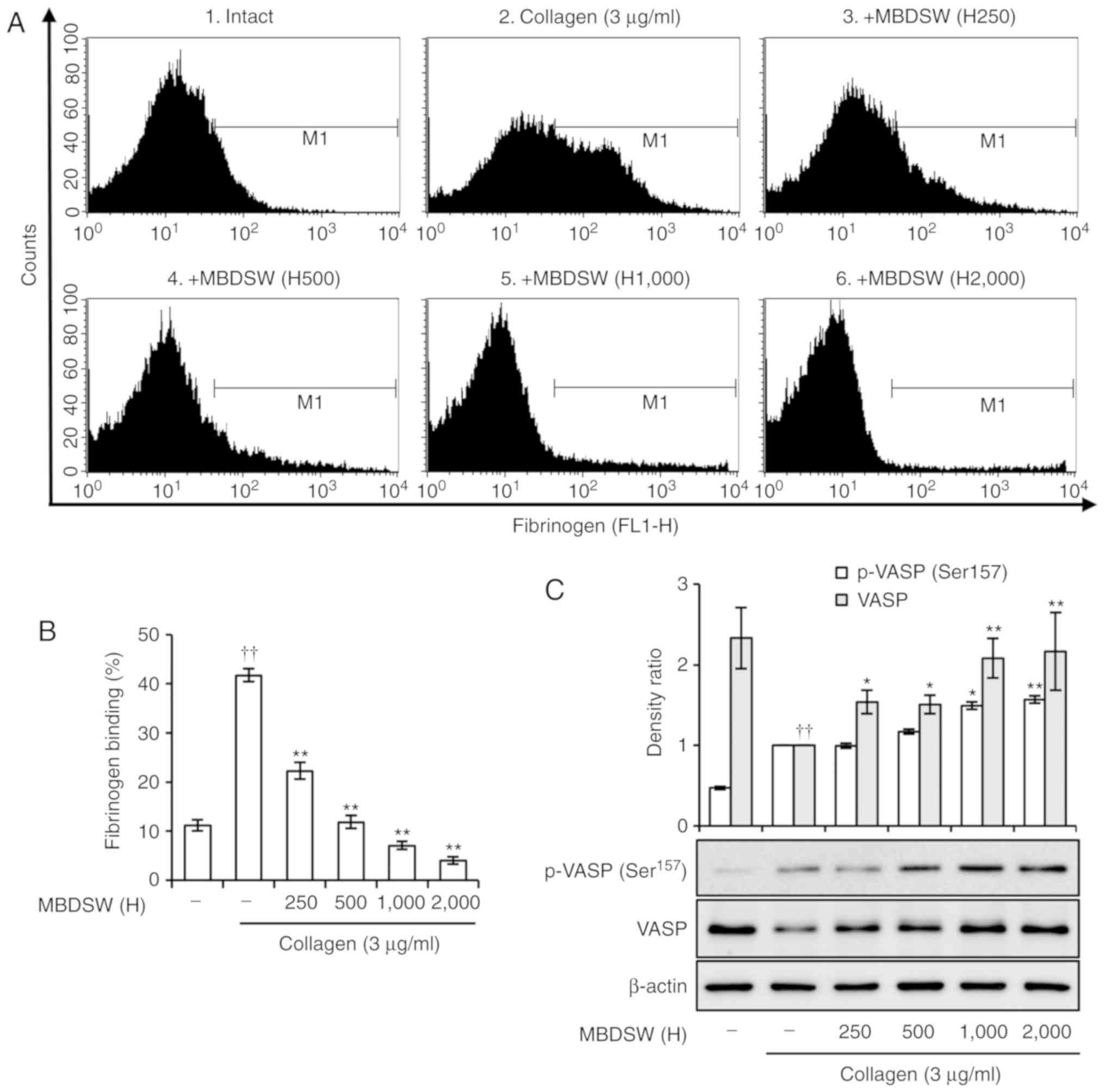
MBDSW inhibits activation of integrin
αIIbβ3
Integrin αIIbβ3 undergoes a
conformational transition upon platelet activation and binds to
fibrinogen (23). To determine
whether MBDSW inhibits the activation of integrin
αIIbβ3, the binding of fibrinogen to the
activated integrin αIIbβ3 was examined by
flow cytometry. The binding of fibrinogen to the activated integrin
αIIbβ3 was significantly inhibited in
MBDSW-treated platelets (Fig. 4A and
B). Furthermore, we examined the effect of MBDSW on VASP
phosphorylation as a regulator of integrin
αIIbβ3 activation (24). In collagen-activated platelets,
VASP (ser157) was shown to be directly phosphorylated
(Fig. 4C), an action that
involves a collagen-related feedback inhibitory signaling pathway
(25). However, MBDSW markedly
increased the phosphorylation of VASP (Ser157) to a
level greater than that observed with collagen only (Fig. 4C).
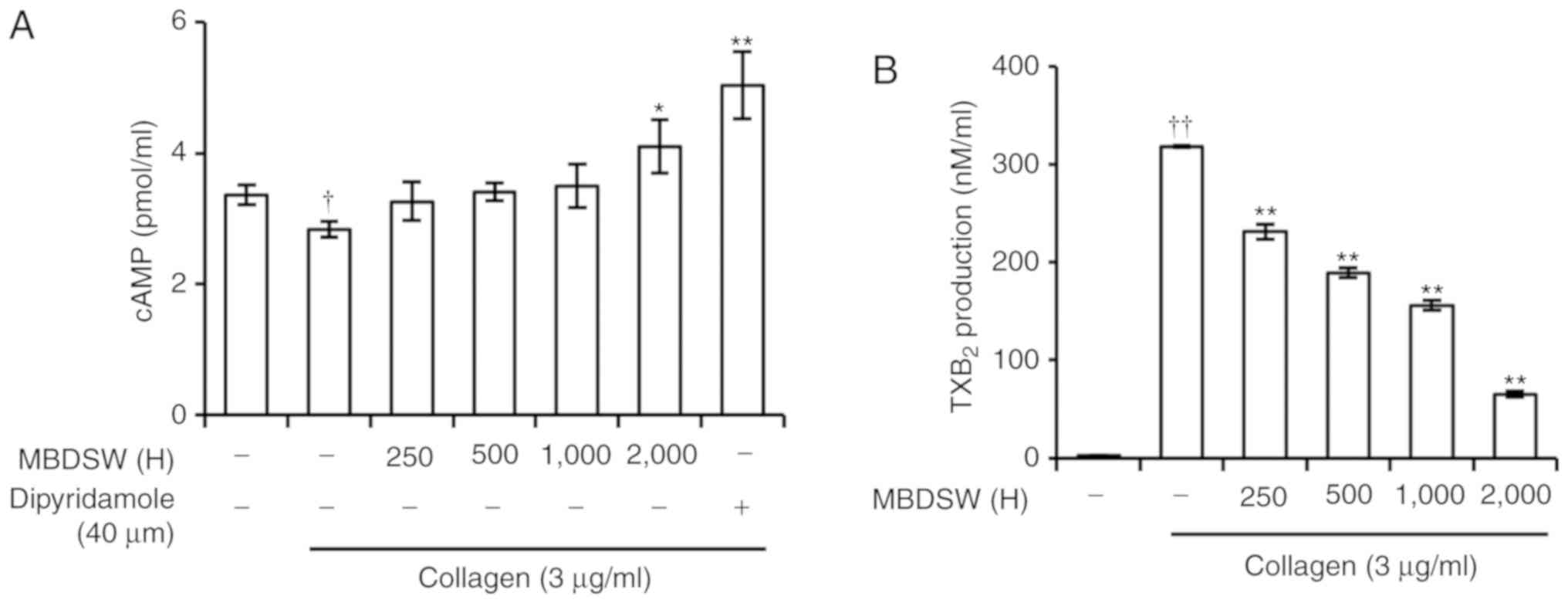
MBDSW alters the cAMP level
Since it has been well established that cAMP
inhibits the activation of platelets and stimulates VASP
phosphorylation in activated platelets, the effects of MBDSW on the
cAMP level in collagen-activated platelets were investigated. As
shown in Fig. 5A, MBDSW (H2,000)
increased the cAMP level. In addition, dipyridamole (40 µM),
a PDE inhibitor, significantly increased the cAMP level and VASP
(ser157) phosphorylation. (Figs. 5A and S3).
MBDSW decreases TXA2
production
Upon exposure to collagen, the platelet membrane is
catabolized from a phospholipid to TXA2. The produced
TXA2 acts as a positive feedback mediator in the
activation of nearby platelets (26). Therefore, in this study, whether
MBDSW decreases the production of TXA2 under collagen
exposure was then examined. In this experiment, TXA2 was
detected using TXB2, a stable metabolite of
TXA2. The results revealed that treatment with collagen
(3 µg/ml) markedly increased TXB2 production;
however, the platelets pre-treated with MBDSW exhibited
significantly decreased TXB2levels, with the reduction
occurring in a hardness-dependent manner (Fig. 5B).
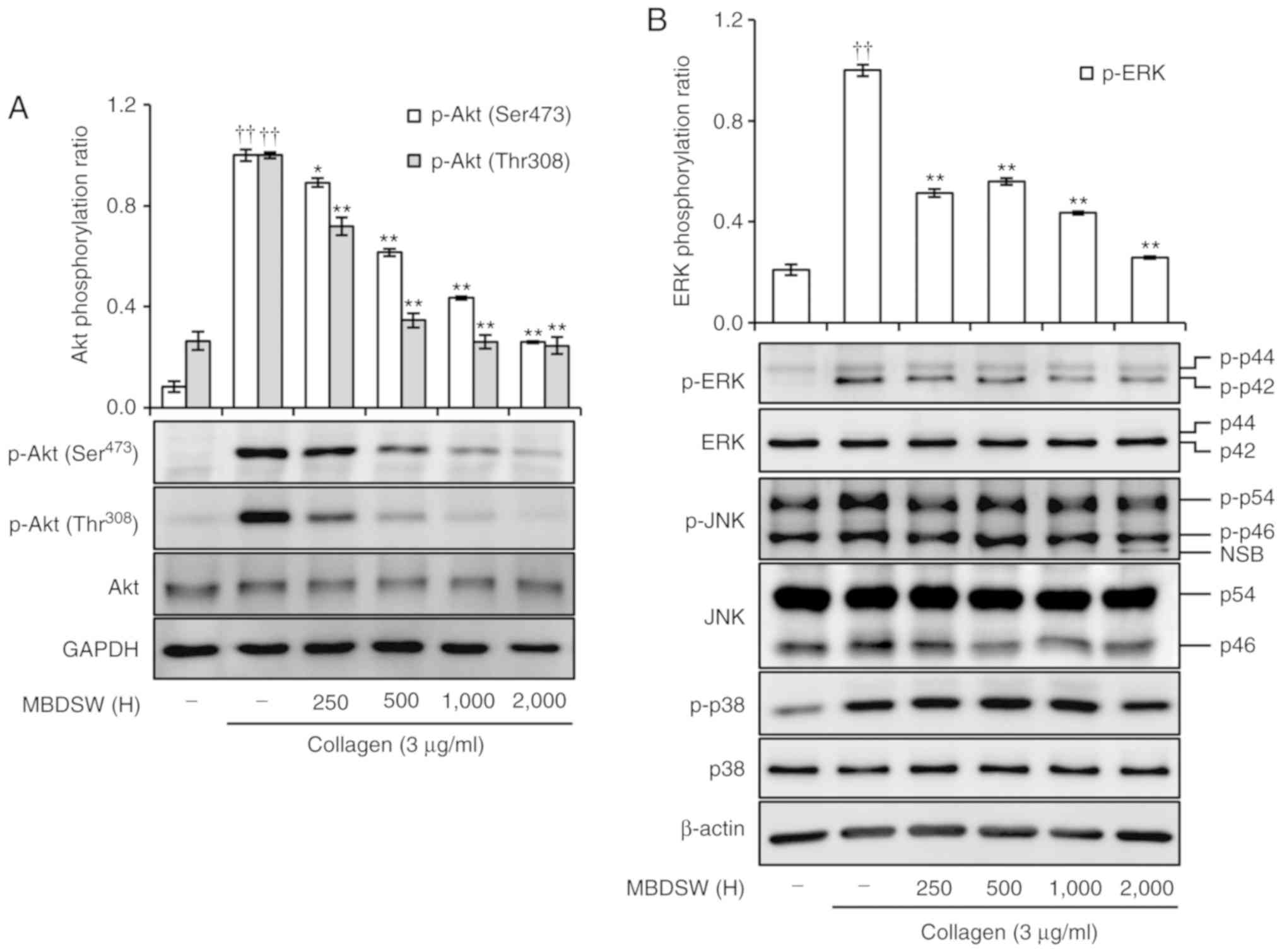
MBDSW inhibits Akt and ERK
phosphorylation
Previous studies have demonstrated that Akt plays a
role in the amplification of platelet aggregation, granule
secretion and integrin αIIbβ3 activation
induced by agonists (16,17); therefore, this study investigated
whether MBDSW inhibits the collagen-stimulated phosphorylation of
Akt; the results revealed that the inhibition of Akt
phosphorylation occurred in a hardness-dependent manner (Fig. 6A).
MAPKs, including ERK, JNK and p38, are activated and
phosphorylated in platelets following treatment with various
agonists (18). Thus, this study
whether MBDSW downregulates MAPKs. The results revealed that MBDSW
inhibits the phosphorylation of ERK, in a hardness-dependent
manner; however, it did not inhibit the phosphorylation of JNK or
p38 (Fig. 6B).
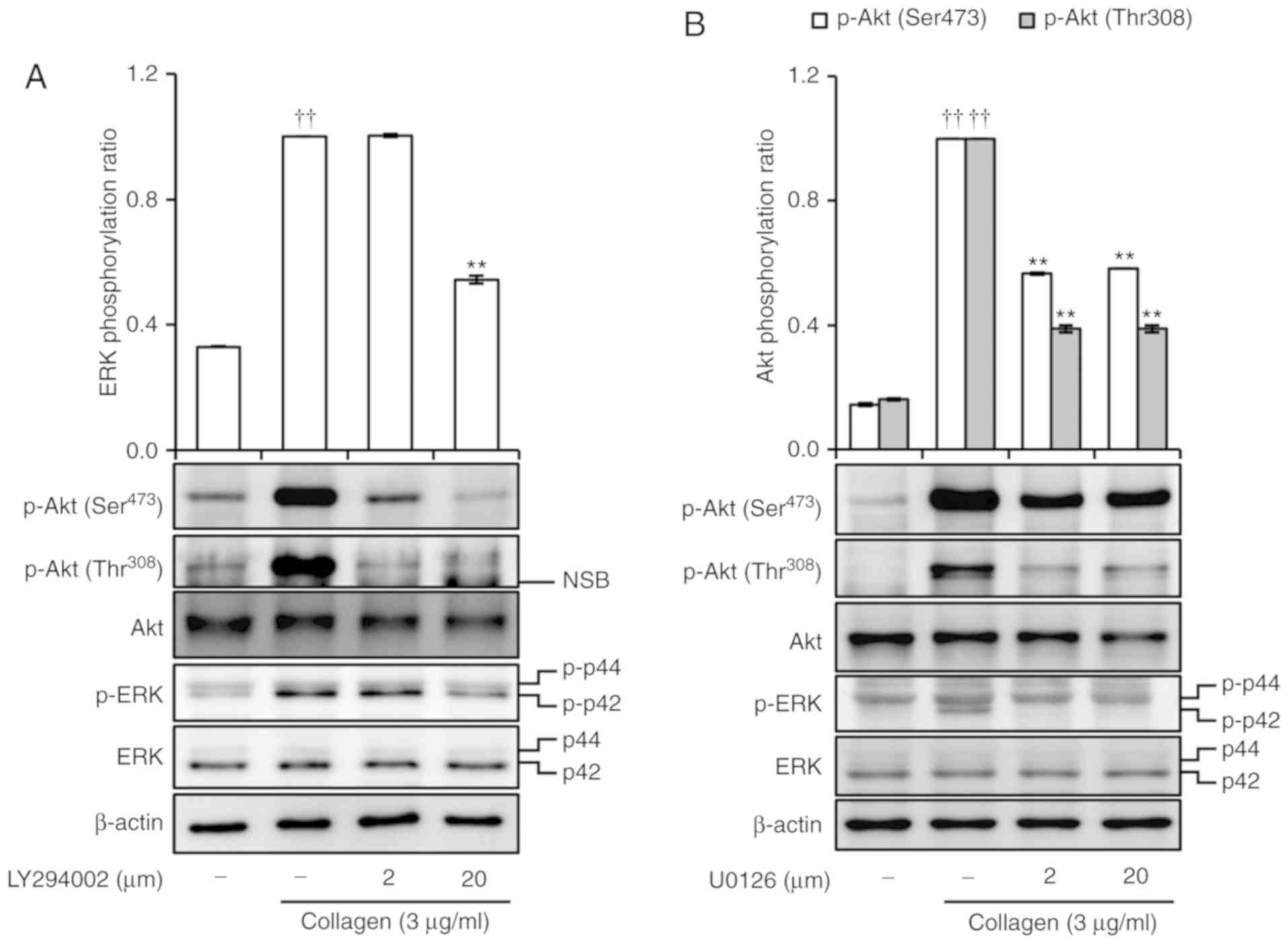
LY294002 and U0126 inhibit Akt and ERK
phosphorylation
In addition, it was investigated whether LY294002,
an Akt inhibitor, or U0126, an ERK inhibitor, can cross-inhibit the
phosphorylation of Akt and ERK. Collagen (3 µg/ml) was shown
to extensively increase phosphorylation of Akt and ERK. By
contrast, both LY294002 and U0126 markedly inhibited the
phosphorylation of Akt and ERK (Fig.
7A and B). In addition, dipyridamole inhibited the
phosphorylation of Akt and ERK (Fig.
S3).
Discussion
CVDs are the most common underlying cause of
mortality in developed nations and are associated with various risk
factors, such as abnormal cholesterol levels, high blood pressure,
diabetes, smoking and obesity (27,28). In addition, the activation of
platelets, which is caused by a series of processes, can increase
the risk of CVDs, when combined with risk factors (29). Previous studies have demonstrated
that MBDSW can regulate various risk factors for CVDs (7,9,11,30-32); however, there is incomplete
information on its effects on platelet activation. Therefore, the
present study evaluated the inhibitory effects of MBDSW on platelet
activity by analyzing granule release, integrin
αIIbβ3 activation, calcium mobilization,
TXA2 production, cAMP level, and the functioning of the
Akt and MAPK pathways.
When the extracellular matrix of the blood vessel
wall is damaged, the released collagen induces platelet
aggregation; thrombin, a coagulation protein produced during the
coagulation process, also causes platelet aggregation (4,33).
Although these mechanisms differ, they ultimately produce the same
result; platelet aggregation and activation. Therefore, this study
initially examined whether MBDSW can inhibit both collagen- and
thrombin-induced platelet aggregation. As shown in Fig. 1A and B, MBDSW markedly inhibited
collagen- and thrombin-induced platelet aggregation in a
hardness-dependent manner. In addition, the presence of calcium in
MBDSW did not automatically affect platelet aggregation (Fig. 1C). This indicates that calcium,
which is essential for platelet activation, is abundant in MBDSW
but does not affect for platelet aggregation. In addition, MBDSW
contains Mg, K, Na and Sr. Mg and K are known to inhibit platelet
aggregation and Na intake tends to increase platelet aggregation
(34,35). However, the suspension buffer
contains much higher Na and K concentrations than MBDSW. Therefore,
Na and K in MBDSW are not affected [suspension buffer, 138 mM NaCl,
2.7 mM KCl and MBDSW (H2,000): 1 mM Na, 0.25 mM K]. Sr is an
antagonist of calcium and is known to act similar to calcium in the
platelet aggregation (36).
However, MBDSW contains a very small amount of strontium and will
not be affected. In this study Fig.
S2 shows Ca and Mg information for platelet aggregation.
However, more detailed research is required and thus further
studies need to be carried out on this matter.
To elucidate the underlying inhibitory mechanisms of
MBDSW, platelet activation markers and downstream molecules were
investigated. To determine whether MBDSW regulates the
intracellular calcium levels, a calcium assay was carried out using
a fluorescence spectrophotometer. The data revealed that MBDSW
significantly decreased the collagen-induced intracellular calcium
levels in a hardness-dependent manner (Fig. 2A). The IP3 generated by
phospholipase C is able to release [Ca2+]i
from the endoplasmic reticulum by directly activating
IP3R channels. In platelets, the regulation of this
receptor is considered to occur through receptor phosphorylation.
Previous studies have reported the inhibition of IP3R
function following phosphorylation (37,38). This study demonstrated that MBDSW
markedly increased the phosphorylation of IP3R (Fig. 2B). In platelets, functions
including degranulation, adhesion and thrombus formation are
activated by an elevation in the intracellular calcium level
(22). Hence, these results
suggest that MBDSW containing calcium inhibits collagen-induced
calcium mobilization via the phosphorylation of IP3R.
Moreover, it is also suggested that MBDSW can prevent
calcium-dependent signaling in platelets. A previous study
suggested that the calcium/calmodulin-induced phosphorylation of
the myosin light chain contributes to platelet granule secretion
(39).
Platelet α-granules contain protein molecules, such
as coagulation factors (e.g., fibrinogen, factor V), growth factors
(e.g., platelet-derived growth factor, epidermal growth factor) and
adhesion molecules (P-selectin). Dense granules contain small
non-protein molecules, such as adenosine diphosphate, ATP and
serotonin that amplify the activation of platelets (40). We found that MBDSW significantly
inhibited P-selectin expression and dense granule secretion
(Fig. 3). These results indicate
that MBDSW, by suppressing P-selectin expression, inhibits
platelet-mediated inflammation and leukocyte migration during
thrombus formation and prevents platelet aggregation.
In platelets, abundant integrin
αIIbβ3 plays an essential role in both
inside-out and outside-in signaling. Integrin
αIIbβ3-mediated signaling transduction is
achieved when it binds to fibrinogen, which leads to platelet
aggregation (41). In this study,
MBDSW markedly inhibited its binding to fibrinogen, blocking
inside-out signaling (Fig. 4A and
B). VASP links the extracellular matrix through integrin to the
actin cytoskeleton and is involved in the inactivation of
inte-grin, and Akt is required for full activation of integrin and
granule secretion (16,17). In the present study, it was
demonstrated that MBDSW activated VASP (Ser157) and
inactivated the phosphorylation of Akt (Figs. 4C and 6A). This result indicates that MBDSW
inhibits the activation of integrin αIIbβ3 by
regulating both VASP and Akt. In addition, the activation of
integrin αIIbβ3 and granule secretion are
known to be inhibited by the cAMP pathway (42,43). As the cAMP pathway inhibits the
activation of integrin αIIbβ3 and granule
secretion through VASP and Akt proteins, this study examined
whether MBDSW increases the cAMP level. MBDSW (H250-H1,000)
restored the cAMP level to that of unstimulated platelets, although
this change was not significant. However, MBDSW did increase the
cAMP level at H2,000 (Fig.
5A).
Further investigation of the molecular mechanisms
revealed that MBDSW may target ERK (Fig. 6B). Indeed, human platelets contain
the MAPK family and Akt proteins, and these proteins are involved
in the platelet activation, adhesion, and granule secretion
(16-18). However, this study demonstrated
that p38 and JNK are not the targets of MBDSW, based on the
observation that MBDSW failed to inhibit the phosphorylation of p38
and JNK (Fig. 6B). Since MBDSW
selectively suppressed the phosphorylation of Akt and ERK, whether
the cross-inhibition of Akt and ERK was possible was investigated.
The phosphorylation of Akt and ERK was assessed by using LY294002,
an Akt inhibitor, and U0126, an ERK inhibitor, to confirm the
association between Akt and ERK pathway signaling. As shown in
Fig. 7, LY294002 inhibited the
phosphorylation of ERK and U0126 inhibited the phosphorylation of
Akt. Furthermore, it was determined whether dipyridamole can
regulate the phosphorylation of Akt, ERK and VASP
(Ser157). The results revealed that dipyridamole
increased the cAMP levels and regulated the phosphorylation of Akt,
ERK and VASP (Ser157) (Figs. 5A and S3). These results suggest that MBDSW can
cross-inhibit ERK and Akt via an increase in the cAMP level.
Moreover, the activation of ERK increases TXA2
production through cytosolic phospholipase A2 (44). MBDSW significantly inhibited the
production of TXA2 (Fig.
5B), suggesting that MBDSW more effectively inhibits the
production of TXA2 through the inhibition of ERK
phosphorylation. The released TXA2, serotonin, and ATP
act as positive feedback mediators in the activation of more
platelets to participated in platelet aggregation (31,45,46). Therefore, the inhibition of
secretion of these molecules by MBDSW is a source of effective
inhibition of platelet aggregation.
Platelet aggregation influences the development of
CVDs. Of note, the majority of risk factors of CVDs, including
hypercholesterolemia, hypertension, smoking and diabetes are able
to increase platelet activation (47-50). In other study, DSW has been shown
to be effective in reducing CVD risk factors (7,9,11,13,14). The present study demonstrated that
platelet aggregation was markedly inhibited by MBDSW that contains
abundant Mg. Therefore, the results suggest that Mg in MBDSW is a
key component in determining the anti-platelet aggregation activity
of MBDSW.
In conclusion, this study demonstrated that the
inhibitory effect of MBDSW on collagen- and thrombin-induced
platelet aggregation occurred in a hardness-dependent manner. In
addition, MBDSW markedly suppressed intracellular downstream signal
transduction; that is, MBDSW significantly inhibited
collagen-induced [Ca2+]i mobilization,
granule secretion and TXA2 production. Furthermore, it
was found that MBDSW regulated the cAMP level and the
phosphorylation of ERK, Akt and VASP (Ser157). Finally,
the activity of integrin αIIbβ3 was inhibited
by MBDSW. Anti-platelet drugs used for the prevention and treatment
of thrombosis target TXA2 production, integrin
inhibition and increases in cAMP levels (51); therefore, MBDSW can be regarded as
a potential prophylactic agent for application in the prevention of
CVDs.
Supplementary Data
Funding
This study was financially supported by the
National R&D project 'Development of new application technology
for deep seawater industry' supported by the Ministry of Oceans and
Fisheries of the Republic of Korea.
Availability of data and materials
All data generated or analyzed during this study
are included in this published article or are available from the
corresponding author on reasonable request.
Authors' contributions
KSN and GSN designed the experiments. KSN, GSN and
KSL analyzed and interpreted the data. GSN performed the
experiments and wrote the manuscript. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by Dongguk University
Gyeongju Institutional Review Board (DUG IRB 20180014-04).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Kuter DJ: The physiology of platelet
production. Stem Cells. 14(Suppl 1): S88–S101. 1996. View Article : Google Scholar
|
|
2
|
Huo Y and Ley KF: Role of platelets in the
development of atherosclerosis. Trends Cardiovasc Med. 14:18–22.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Farndale RW, Sixma JJ, Barnes MJ and de
Groot PG: The role of collagen in thrombosis and hemostasis. J
Thromb Haemost. 2:561–573. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Barnes MJ and Farndale RW: Collagens and
atherosclerosis. Exp Gerontol. 34:513–525. 1999. View Article : Google Scholar
|
|
5
|
Jin J, Quinton TM, Zhang J, Rittenhouse SE
and Kunapuli SP: Adenosine diphosphate (ADP)-induced thromboxane
A(2) generation in human platelets requires coordinated signaling
through integrin alpha(IIb)beta(3) and ADP receptors. Blood.
99:193–198. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Flaumenhaft R: Molecular basis of platelet
granule secretion. Arterioscler Thromb Vasc Biol. 23:1152–1160.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hwang HS, Kim HA, Lee SH and Yun JW:
Anti-obesity and anti-diabetic effects of deep sea water on ob/ob
mice. Mar Biotechnol (NY). 11:531–539. 2016. View Article : Google Scholar
|
|
8
|
Hataguchi Y, Tai H, Nakajima H and Kimata
H: Drinking deep-sea water restores mineral imbalance in atopic
eczema/dermatitis syndrome. Eur J Clin Nutr. 59:1093–1096. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee KS, Kwon YS, Kim S, Moon DS, Kim HJ
and Nam KS: Regulatory mechanism of mineral-balanced deep sea water
on hypocholesterolemic effects in HepG2 hepatic cells. Biomed
Pharmacother. 86:405–413. 2017. View Article : Google Scholar
|
|
10
|
Lee KS, Chun SY, Lee MG, Kim S, Jang TJ
and Nam KS: The prevention of TNF-α/IFN-γ mixture-induced
inflammation in human keratinocyte and atopic dermatitis-like skin
lesions in Nc/Nga mice by mineral-balanced deep sea water. Biomed
Pharmacother. 97:1331–1340. 2018. View Article : Google Scholar
|
|
11
|
Sheu MJ, Chou PY, Lin WH, Pan CH, Chien
YC, Chung YL, Liu FC and Wu CH: Deep sea water modulates blood
pressure and exhibits hypolipidemic effects via the AMPK-ACC
pathway: An in vivo study. Mar Drugs. 11:2183–2202. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee KS, Shin JS, Kwon YS, Moon DS and Nam
KS: Suppression of cancer progression and metastasis in HT-29 human
colorectal adenocarcinomas by deep sea water. Biotechnol Bioprocess
Eng. 18:194–200. 2013. View Article : Google Scholar
|
|
13
|
Miyamura M, Yoshioka S, Hamada A, Takuma
D, Yokota J, Kusunose M, Kyotani S, Kawakita H, Odani K, Tsutsui Y
and Nishioka Y: Difference between deep seawater and surface
seawater in the preventive effect of atherosclerosis. Biol Pharm
Bull. 27:1784–1787. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee KS, Chun SY, Kwon YS, Kim S and Nam
KS: Deep sea water improves hypercholesterolemia and hepatic lipid
accumulation through the regulation of hepatic lipid metabolic gene
expression. Mol Med Rep. 15:2814–2822. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chun SY, Kim S and Nam KS: The inhibitory
effects of deep-sea water on doxorubicin-induced
epithelial-mesenchymal transition. Oncol Rep. 38:1163–1171. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen J, De S, Damron DS, Chen WS, Hay N
and Byzova TV: Impaired platelet responses to thrombin and collagen
in AKT-1-deficient mice. Blood. 104:1703–1710. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stojanovic A, Marjanovic JA, Brovkovych
VM, Peng X, Hay N, Skidgel RA and Du X: A phosphoinositide
3-kinase-AKT-nitric oxide-cGMP signaling pathway in stimulating
platelet secretion and aggregation. J Biol Chem. 281:16333–16339.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Flevaris P, Li Z, Zhang G, Zheng Y, Liu J
and Du X: Two distinct roles of mitogen-activated protein kinases
in platelets and a novel Rac1-MAPK-dependent integrin outside-in
retractile signaling pathway. Blood. 113:893–901. 2009. View Article : Google Scholar :
|
|
19
|
Heemskerk JW, Bevers EM and Lindhout T:
Platelet activation and blood coagulation. Thromb Haemost.
88:186–193. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rittenhouse-Simmons S and Deykin D:
Isolation of membranes from normal and thrombin treated
gel-filtered platelets using a lectin marker. Biochim Biophys Acta.
426:688–696. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Schaeffer J and Blaustein MP: Platelet
free calcium concentrations measured with fura-2 are influenced by
the transmembrane sodium gradient. Cell Calcium. 10:101–113. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nesbitt WS, Giuliano S, Kulkarni S,
Dopheide SM, Harper IS and Jackson SP: Intercellular calcium
communication regulates platelet aggregation and thrombus growth. J
Cell Biol. 160:1151–1161. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shattil SJ and Newman PJ: Integrins:
Dynamic scaffolds for adhesion and signaling in platelets. Blood.
104:1606–1615. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Reinhard M, Jarchau T and Walter U:
Actin-based motility: Stop and go with Ena/VASP proteins. Trends
Biochem Sci. 26:243–249. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gambaryan S, Kobsar A, Rukoyatkina N,
Herterich S, Geiger J, Smolenski A, Lohmann SM and Walter U:
Thrombin and collagen induce a feedback inhibitory signaling
pathway in platelets involving dissociation of the catalytic
subunit of protein kinase A from an NFkappaB-IkappaB complex. J
Biol Chem. 285:18352–18363. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Paul BZ, Jin J and Kunapuli SP: Molecular
mechanism of throm-boxane A(2)-induced platelet aggregation
Essential role for p2t(ac) and alpha(2a) receptors. J Biol Chem.
274:29108–29114. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mathur RK: Role of diabetes, hypertension,
and cigarette smoking on atherosclerosis. J Cardiovasc Dis Res.
1:64–68. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
O'Donnell CJ and Elosua R: Cardiovascular
risk factors. Insights from Framingham Heart Study. Rev Esp
Cardiol. 61:299–310. 2008.In Spanish. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Willoughby S, Holmes A and Loscalzo J:
Platelets and cardiovascular disease. Eur J Cardiovasc Nurs.
1:273–288. 2002. View Article : Google Scholar
|
|
30
|
Ha BG, Shin EJ, Park JE and Shon YH:
Anti-diabetic effect of balanced deep-sea water and its mode of
action in high-fat diet induced diabetic mice. Mar Drugs.
11:4193–4212. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Katsuda S, Yasukawa T, Nakagawa K, Miyake
M, Yamasaki M, Katahira K, Mohri M, Shimizu T and Hazama A:
Deep-sea water improves cardiovascular hemodynamics in kurosawa and
kusanagi-hypercholesterolemic (KHC) rabbits. Biol Pharm Bull.
31:38–44. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yoshioka S, Hamada A, Cui T, Yokota J,
Yamamoto S, Kusunose M, Miyamura M, Kyotani S, Kaneda R, Tsutsui Y,
et al: Pharmacological activity of deep-sea water: Examination of
hyperlipemia prevention and medical treatment effect. Biol Pharm
Bull. 26:1552–1559. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Borissoff JI, Spronk HM, Heeneman S and
ten Cate H: Is thrombin a key player in the
'coagulation-atherogenesis' maze? Cardiovasc Res. 82:392–403. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gawaz M, Ott I, Reininger AJ and Neumann
FJ: Effects of magnesium on platelet aggregation and adhesion
Magnesium modulates surface expression of glycoproteins on
platelets in vitro and ex vivo. Thromb Haemost. 72:912–918. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gow IF, Padfield PL, Reid M, Stewart SE,
Edwards CR and Williams BC: High sodium intake increases platelet
aggregation in normal females. J Hypertens. (Suppl 5)–S246.
1987.
|
|
36
|
Togna G, Gallozzi S and Caprino L:
Influence of strontium chloride on blood platelet function. Arch
Toxicol. 63:366–369. 1989. View Article : Google Scholar
|
|
37
|
Komalavilas P and Lincoln TM:
Phosphorylation of the inositol 1,4,5-trisphosphate receptor by
cyclic GMP-dependent protein kinase. J Biol Chem. 269:8701–8707.
1994.PubMed/NCBI
|
|
38
|
Supattapone S, Danoff SK, Theibert A,
Joseph SK, Steiner J and Snyder SH: Cyclic AMP-dependent
phosphorylation of a brain inositol trisphosphate receptor
decreases its release of calcium. Proc Natl Acad Sci USA.
85:8747–8750. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Nishikawa M, Tanaka T and Hidaka H:
Ca2+-calmodulin- dependent phosphorylation and platelet secretion.
Nature. 287:863–865. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Reed GL, Fitzgerald ML and Polgár J:
Molecular mechanisms of platelet exocytosis: Insights into the
'secrete' life of thrombocytes. Blood. 96:3334–3342.
2000.PubMed/NCBI
|
|
41
|
Payrastre B, Missy K, Trumel C, Bodin S,
Plantavid M and Chap H: The integrin alpha IIb/ beta 3 in human
platelet signal transduction. Biochem Pharmacol. 60:1069–1074.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Smolenski A: Novel roles of
cAMP/cGMP-dependent signaling in platelets. J Thromb Haemost.
10:167–176. 2012. View Article : Google Scholar
|
|
43
|
Kim S, Jee K, Kim D, Koh H and Chung J:
Cyclic AMP inhibits Akt activity by blocking the membrane
localization of PDK1. J Biol Chem. 276:12864–12870. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lin LL, Wartmann M, Lin AY, Knopf JL, Seth
A and Davis RJ: cPLA2 is phosphorylated and activated by MAP
kinase. Cell. 72:269–278. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Birk AV, Broekman MJ, Gladek EM, Robertson
HD, Drosopoulos JH, Marcus AJ and Szeto HH: Role of extracellular
ATP metabolism in regulation of platelet reactivity. J Lab Clin
Med. 140:166–175. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cerrito F, Lazzaro MP, Gaudio E, Arminio P
and Aloisi G: 5HT2-receptors and serotonin release: Their role in
human platelet aggregation. Life Sci. 53:209–215. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bröijersén A, Hamsten A, Eriksson M,
Angelin B and Hjemdahl P: Platelet activity in vivo in
hyperlipoproteinemia-importance of combined hyperlipidemia. Thromb
Haemost. 79:268–275. 1998. View Article : Google Scholar
|
|
48
|
Nityanand S, Pande I, Bajpai VK, Singh L,
Chandra M and Singh BN: Platelets in essential hypertension. Thromb
Res. 72:447–454. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Nowak J, Murray JJ, Oates JA and
FitzGerald GA: Biochemical evidence of a chronic abnormality in
platelet and vascular function in healthy individuals who smoke
cigarettes. Circulation. 76:6–14. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Manduteanu I, Calb M, Lupu C, Simionescu N
and Simionescu M: Increased adhesion of human diabetic platelets to
cultured valvular endothelial cells. J Submicrosc Cytol Pathol.
24:539–547. 1992.PubMed/NCBI
|
|
51
|
Eikelboom JW, Hirsh J, Spencer FA, Baglin
TP and Weitz JI: Antiplatelet drugs: Antithrombotic therapy and
prevention of thrombosis, 9th ed: American college of chest
physicians evidence-based clinical practice Guidelines. Chest.
141(2 Suppl): e89S–e119S. 2012. View Article : Google Scholar : PubMed/NCBI
|