Introduction
Coronary heart disease is the most common type of
cardiovascular disease. Myocardial infarction is the most dangerous
pathological process, and reperfusion of the coronary artery is the
most fundamental treatment (1).
With the development of coronary thrombolysis, percutaneous
coronary intervention, coronary artery bypass grafting and other
treatments, myocardial necrosis may be prevented, saving the lives
of numerous patients with myocardial infarction (2). However, the reperfusion of the
coronary vessels subsequent to myocardial ischemia and infarction
itself may also cause reperfusion injury that aggravates myocardial
cell damage or even causes necrosis (3,4).
This phenomenon is myocardial ischemic-reperfusion injury (MIRI)
(5). The mechanism of MIRI is
very complex. Previous studies have reported that
ischemia-reperfusion is able to induce oxidative stress,
intracellular calcium overload, the rapid recovery of physiological
pH, an inflammatory response, further active cell apoptosis,
necrosis and autophagy, eventually resulting in further damage to
the myocardium (6-8). However, it is recognized that MIRI
may be alleviated through regulation of the apoptotic pathway.
Multiple apoptotic pathways, including the
phosphoinositide-3-kinase (PI3K)/protein kinase B (AKT) pathway,
are activated and involved in MIRI (9,10).
Research on the molecular mechanisms of MIRI is still being
conducted; the effective prevention and treatment of MIRI is a
difficult problem and a focus of research (11). Current research trends in the
field of life sciences have shifted from the protein level to the
gene regulation level, and from protein-coding genes to
non-protein-coding protein genes (12).
Long-non-coding RNA (lncRNA) is most commonly
defined as a transcript with a length of >200 nucleotides and no
protein encoding function, and is distributed in the nucleus and
the cytoplasm (13). lncRNA has
been revealed to be involved in the pathophysiological processes of
various tumor types, including in cell growth, proliferation,
invasion and apoptosis (14,15). A number of studies have considered
the effects of lncRNAs that serve functions in myocardial ischemia
(16,17). A number of lncRNAs have been
reported to be involved in the regulation of myocardial ischemia by
exerting their biological mechanisms (18,19). However, there are few reports on
the function of lncRNAs in an MIRI model. A previous study
performed biological analyses of a MIRI model in mice, and revealed
that various lncRNAs are differentially expressed, and it was
predicted that MIRI may be regulated by their participation in
biological processes (20). The
investigation of individual lncRNA functions is still ongoing. It
is important to study the function of lncRNAs in MIRI.
Schneider et al (21) discovered upregulated lncRNA growth
arrest specific 5 (GAS5) in growth-arrested cells screened by
subtractive hybridization in 1988. The coding gene for GAS5 is a
member of the 5′ terminal oligopyrimidine family, containing 12
exons, and its potential open reading frame is very limited
(22,23). Various studies have suggested that
lncRNA GAS5 is expressed in a variety of tumor tissues and serves
the function of a tumor suppressor gene (24-26).
Overexpression of lncRNA GAS5 may inhibit the
growth, invasion and metastasis of tumor cells and induce apoptosis
(27). However, the mechanism of
action of lncRNA GAS5 in MIRI is still unclear. The present study
focused on the potential regulatory function of lncRNA GAS5 in
MIRI, particularly in regulating apoptosis. It was proposed that
lncRNA GAS5 may regulate apoptosis induced by MIRI via the PI3K/AKT
pathway by binding to microRNA (miR)-532-5p. The aim of the present
study was to provide a preventive strategy for MIRI.
Materials and methods
Cell culture and hypoxia-reoxygenation
(HR) model
Rat cardiomyocte H9c2 cells were purchased from the
Shanghai Institutes for Biological Sciences, Chinese Academy of
Sciences (Shanghai, China). H9c2 cells were cultured in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc.) containing 10% fetal bovine serum (FBS; Tianjin Hao Yang
Biological Company) at 37°C in an incubator with 95% air and 5%
CO2. Then, a H9c2 HR injury model was prepared to mimic
the MIRI model in vitro. In brief, the cells were cultured
with glucose-free DMEM and incubated in a low-oxygen atmosphere
(90% N2, 5% CO2 and 5% O2; 37°C)
for 8 h. Subsequently, the medium was changed to DMEM containing
10% FBS in a normal atmosphere (95% air and 5% CO2;
37°C) for 2 h.
Animals and MIRI model
A total of 20 healthy male (age, 6-8 weeks) specific
pathogen-free (SPF) Wistar rats weighing 300±20 g, were purchased
from Liaoning Changsheng Biotechnology Co., Ltd. The rats were
treated according to the Guide for the Care and Use of Laboratory
Animals (Federal Register Doc. 2011.11490; National Institutes of
Health, Bethesda, MD, USA) (28).
The rats were maintained in a clean and ventilated environment at a
temperature of 22-24°C indoors. Fresh food and drinking water were
available ad libitum, with a circadian rhythm of 12 h. Food
and bedding were changed for the rats every other day, and the
health of the animals was monitored. The 20 rats were randomly
divided into a control group (without any treatment) and an
ischemia-reperfusion group. Preparation of the isolated rat heart
MIRI model was performed using the Langendorff isolated cardiac
perfusion system (29). A
peritoneal injection of pentobarbital sodium (30 mg/kg) was
administered for anesthesia; the anesthetized rats were fixed to a
table, the chest was quickly opened, the intact heart was removed
and placed in a 4°C KH solution (NaCl 0.15 mol/l, KCl 0.006 mol/l,
CaCl2 0.002 mol/l and NaHCO3 0.002 mol/l)
with pre-filled 95% O2 + 5% CO2 mixed gas,
and then the surrounding tissue was carefully removed and the aorta
exposed. The aorta was carefully lifted with forceps in order to
insert the perfusion tube. Once the heart began to fluctuate, it
was fixed to the cardiac perfusion device with a 7-0 surgical line
near the upper edge of the aorta. The isolated rat heart was
perfused with KH solution using a perfusion device and stabilized
for 10 min prior to the induction of ischemia. The perfusion
conditions were maintained at a constant temperature of 37°C and a
constant pressure of 75 mmHg. Then, the perfusion was stopped, and
the MIRI model was prepared by stopping the perfusion for 30 min
and reperfusing for 120 min. Rats were sacrificed by cervical
dislocation at the end of reperfusion for 120 min, and no
additional anesthetic dose was required. At the end of the
experiment, the rat had the heart removed and no heartbeat. In
addition, the rats had no breathing, no pupils and nerve reflexes,
and had finally succumbed to mortality. Animal experiments were
ethically approved by the experimental animal welfare and ethics
committee of China Medical University (Liaoning, China; approval
no. kt2018009). Animal studies were conducted according to the
checklist of the ARRIVE guidelines (30).
Cell transfection
Small interfering RNAs (siRNAs) targeting GAS5
(siGAS5) and the negative control siRNAs (si-NC) were designed and
synthesized by Shanghai GenePharma Co., Ltd. The sequences were as
follows: siGAS5 (rat) sense, 5′-TCT CAC AGG CAG TTC TGT GG-3′ and
antisense, 5′-ATC CAT CCA GTC ACC TCT GG-3; si-NC (rat) sense,
5′-UUC UCC GAA CGU GUC ACG UTT-3′ and antisense, 5′-CGU GAC ACG UUC
GGA GAA TT-3′. miR-532-5p mimics/inhibitors and their negative
control were synthesized by Shanghai GenePharma Co., Ltd. The
sequences were as follows: miR-532-5p mimics (rat), 5′-CAU GCC UUG
AGU GUA GGA CUG UAG UCC UAC ACU CAA GGC AUG UU-3′; and miR-532-5p
inhibitor (rat), 5′-ACA GUC CUA CUC AAG GCA UG-3′; miRNA inhibitor
NC, 5′-CAG UAC UUU UGU GUA GUA CAA-3 and microRNA mimic NC, 5′-UUC
UCC GAA CGU GUC ACG UTT-3. For transfection, H9c2 cells were
cultured to achieve a density of 70%, and transfected using
Lipofectamine® 3000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. The
quantity of siRNA transfected was as follows: 75 pmol/well in
6-well plates, 30 pmol/well in 12-well plates and 3 pmol/well in
96-well plates. After transfection for 24 or 48 h at 37°C in an
incubator containing 5% CO2, the cells were subjected to
HR experiments, followed by the subsequent experiments.
Transfections were performed in 6-, 12- or 96-well plates according
to the requirements of each experiment.
Cell counting kit-8 (CCK-8) assay
To investigate the effect of lncRNA GAS5 on cell
viability in MIRI, a CCK-8 assay was used. The H9c2 cells
(3×103 cells/well) were cultured in 96-well plates
according to the experimental groups, and then cell viability was
determined using a CCK-8 assay (Dojindo Molecular Technologies,
Inc.), according to the manufacturer's protocol. Subsequent to
treating the cells according to the predetermined experimental
groups, 10 µl CCK-8 solution was added to each well and
incubated for 1 h at 37°C in a 5% CO2 incubator. The
absorbance value was detected at a wavelength of 450 nm by an
ultra-micro microporous plate spectrophotometer (BioTek
Instruments, Inc.).
Measurement of myocardial injury
markers
The H9c2 cells (3×103 cells/well) were
cultured in 96-well culture plates to detect myocardial enzyme
markers. Lactate dehydrogenase (LDH) and creatine kinase isoenzyme
(CK-MB) may be released into the culture medium following
myocardial cell injury and death, and their levels may be tested to
indicate the degree of damage to cardiomyocytes (31). LDH was determined using an LDH
assay kit (Shenyang Wan Biotechnology Co., Ltd.) according to the
manufacturer's protocol. The levels of CK-MB in the culture medium
were measured using a CK-MB enzyme-linked immunosorbent assay kit
(cat. no. MB-6930A; Jiangsu MB Biotechnology Co., Ltd.) according
to the manufacturer's protocol. The absorbance value was detected
at a wavelength of 450 nm by an ultra-micro microporous plate
spectrophotometer (BioTek Instruments, Inc.).
Measurement of cell apoptosis
The degree of cell apoptosis was visually measured
using a Hoechst staining kit (Beyotime Institute of Biotechnology)
according to the manufacturer's protocol. H9c2 cells
(1×105 cells/well) were plated in 12-well plates. The
cells were transfected and treated by HR according to the
experimental group. The treated cells were fixed with 4%
paraformaldehyde at room temperature for 20 min, washed with PBS
and stained with a Hoechst stain for 20 min at room temperature.
Then, the nuclear morphology of the cells was detected at a ×400
magnification under a fluorescence microscope (Image-Pro Plus
version 6.0; Media Cybernetics, Inc., Rockville, MD, USA). The
number of apoptotic cells was counted with the naked eye, and the
quantitative index of apoptosis was expressed as the percentage of
apoptotic cells in the total cells. A more accurate rate of cell
apoptosis was determined using flow cytometry analysis
(LSRFortessa; BD Biosciences, Franklin Lakes, NJ, USA) using the
Annexin V fluorescein isothiocyanate/propidium iodide kit (Nanjing
KeyGen Biotech Co., Ltd., Nanjing, China) to label the cells,
according to the manufacturer's protocol. SPSS version 23.0 (SPSS,
Inc., Chicago, IL, USA) was used for data analysis.
Target identification and luciferase
reporter assay
RNAhybrid and Targetscan online software were used
to identify the downstream target genes of lncRNA GAS5, RNAhybrid
software (RNAhybrid-2.1.2; https://bibiserv.cebitec.uni-bielefeld.de/rnahybrid/)
can predict the binding of lncRNA and miRNA, and Targetscan
(release 3.1; http://www.targetscan.org) can predict the binding of
miRNA and mRNA. 293T cells purchased from The Cell Bank of Type
Culture Collection of the Chinese Academy of Sciences and cultured
in 96-well plate (1.0×104 cells/well) with DMEM (Gibco;
Thermo Fisher Scientific, Inc.) at 37°C and 5% CO2 for
24 h. In order to detect the binding of GAS5 and miR-532-5p, 293T
cells were co-transfected with pmiR-RB-Report™r-Gas5(NR_002704.1
complete sequence)-WT(r-Gas5-WT)-or pmiR-RB-Report™
r-Gas5(NR_002704.1 complete sequence)-MUT(249-255:AA
GGCAT>TTCCGTA) (r-Gas5-MUT) luciferase reporter plasmids
(Guangzhou RiboBio Co., Ltd., Guangzhou, China) along with
miR-532-5p mimic (50 nM) or miR-532-5p mimic negative control (50
nM) using Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at room temperature. To detect the binding of
phosphatase and tensin homolog (PTEN) and miR-532-5p, 293T cells
were co-transfected with pmiR-RB-Rep ort™r-Pten(3′UTR:1310-3809)-WT
(r-Pten-WT) or pmiR-RB-R
eport™r-Pten(3′UTR:1310-3809)-MUT(1352-1358 AAGGCAT
>TTCCGTA2584-2590AAGGCAT>TTCCGTA3730-3736AA GGCAT>TTCCGTA)
(r-Pten-MUT) luciferase reporter plasmid along with miR-532-5p
mimic (50 nM) or miR-532-5p mimic negative control (50 nM). A total
of 48 h after transfection, the luciferase activities were examined
using the Dual-Luciferase Reporter Assay system (Promega
Corporation, Madison, WI, USA) according to the manufacturer's
protocol.
Western blotting
At 48 h post-transfection, H9c2 cells were treated
with HR, and then protein was extracted with
radioim-munoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology). The total protein was extracted from lysates by
centrifugation at 13,000 × g and 4°C for 15 min. The concentration
of protein was measured using a BCA protein concentration kit
(Beyotime Institute of Biotechnology). For the western blotting, 40
µg protein denatured by heating was subjected to 10%
SDS-polyacrylamide gel electrophoresis for separation and then
transferred to polyvinylidene difluoride (PVDF) membranes. The
membrane was blocked using 5% skim milk for 1 h at room temperature
and then incubated overnight at 4°C with the specific primary
antibodies, including anti-B-cell lymphoma 2 (Bcl-2; cat. no.
WL01556; 1:500; Shenyang Wan Biotechnology Co., Ltd.),
anti-Bcl-2-associated X protein (Bax; cat. no. WL01637; 1:500;
Shenyang Wan Biotechnology Co., Ltd.), anti-cleaved-caspase-3 (cat.
no. WL01992; 1:500; Shenyang Wan Biotechnology Co., Ltd.),
anti-PTEN (cat. no. WL01901; 1:500; Shenyang Wan Biotechnology Co.,
Ltd.), anti-PI3K (cat. no. WL01169; 1:500; Shenyang Wan
Biotechnology Co., Ltd.), anti-phosphorylated (p-) PI3K (cat. no.
ab182651; 1:1,000; Abcam), anti-AKT (cat. no. WLP001a; 1:500;
Shenyang Wan Biotechnology Co., Ltd.), anti-p-AKT (cat. no.
WL0003b; 1:1,000; Shenyang Wan Biotechnology Co., Ltd.) and
anti-β-actin (cat. no. WL01845; 1:1,000; Shenyang Wan Biotechnology
Co., Ltd.). The following day, the PVDF membranes were incubated
with horseradish peroxidase-labeled goat anti-rabbit immunoglobulin
G secondary antibodies (cat. no. WLA023; 1:5,000; Shenyang Wan
Biotechnology Co., Ltd.) at 37°C for 45 min. Detection of the
protein bands was performed using an enhanced chemiluminescence for
western blotting kit (Shenyang Wan Biotechnology Co., Ltd.),
according to the manufacturer's protocol. The levels of
phosphorylated proteins were normalized to their corresponding
total protein levels. Relative densitometry was calculated using
Image J2x analysis software (National Institutes of Health).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the myocardium and
cardiomyocytes using TRIzol® reagent (Life Technologies;
Thermo Fisher Scientific, Inc.) at 48 h after transfection.
Subsequent to detecting the quality of the total RNA, RNA was
reverse transcribed into cDNA using the PrimeScript RT reagent kit
with gDNA Eraser (Takara Bio, Inc.), according to the
manufacturer's protocol. The RNA was heated at 37°C for 15 min and
at 85°C for 5 sec, and then cooled to 4°C to obtain the cDNA.
RT-qPCR was then performed using SYBR Premix Ex Taq II (Takara Bio,
Inc.), according to the manufacturer's protocol. The cDNA was
amplified under the following thermocycling conditions: 95°C for 5
min, followed by 8 cycles of 95°C for 30 sec, 60°C for 45 sec and
72°C for 20 sec; and 35 cycles of 95°C for 30 sec, 56°C for 45 sec,
72°C for 20 sec, 95°C for 1 min, 55°C for 30 sec and 95°C for 30
sec. For miRNA RT-qPCR, RNA was reverse transcribed into cDNA using
the Super RT kit (Bioteke Biotechnology Co., Ltd.) according to the
manufacturer's protocol. The RNA was heated at 37°C for 30 min and
at 42°or 30 min, and then 70°C for 10 min to obtain the cDNA.
RT-qPCR was then performed using SYBR Taq PCR MasterMix (Bioteke
Biotechnology Co., Ltd.) according to the manufacturer's protocol.
The cDNA was amplified under the following thermocycling
conditions: 94°C for 2 min, followed by 94°C for 15 sec, 60°C for
15 sec and 72°C for 15 sec; 40 cycles of 72°C for 2 min 30 sec and
40°C for 1 min 30 sec; melting from 60°C to 94°C, with 1.0°C every
1 sec; and 25°C for 1 min. All oligonucleotide primer pairs and the
reference primers (β-actin and U6) were designed by Sangon Biotech
Co., Ltd. Relative gene expression was analyzed using the
2−ΔΔCq method (32).
The primer sequences in the present study are presented in Table I.
 | Table IPrimer sequences in the present
study. |
Table I
Primer sequences in the present
study.
| Gene | Primer sequence
(5′-3′) |
|---|
| lncRNA-GAS5
forward |
TCTCACAGGCAGTTCTGTGG |
| lncRNA-GAS5
reverse |
ATCCATCCAGTCACCTCTGG |
| miR-532-5p
forward |
CGCCCCATGCCTTGAGTGTA |
| miR-532-5p
reverse |
GTGCAGGGTCCGAGGTATTC |
| PTEN forward |
TAGAGCGTGCGGATAATGAC |
| PTEN reverse |
GATGGCTCCTCTACTGTTTT |
Statistical analysis
The experimental data are expressed as the mean ±
standard deviation and were statistically analyzed using SPSS 23.0
software (SPSS, Inc.). Differences between the control and HR
groups were evaluated using an independent sample Student's t-test.
Differences between multiple groups were initially evaluated using
a one-way analysis of variance followed by Bonferroni's test was
performed to assess the comparisons between each two groups.
P<0.05 was considered to indicate a statistically significant
difference. GraphPad Prism 5.0 software (GraphPad Software, Inc.)
was used to draw the diagrams.
Results
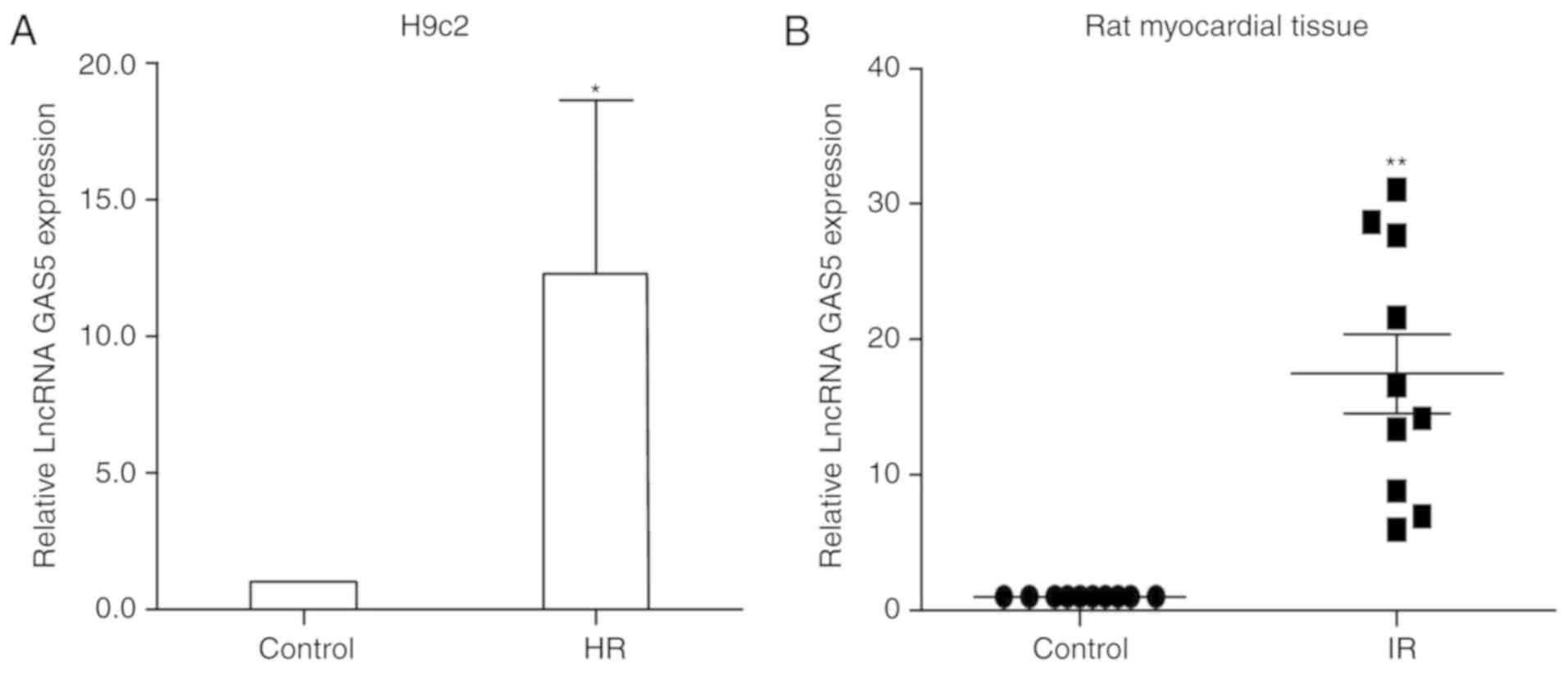
LncRNA GAS5 expression is significantly
upregulated following ischemia/reperfusion myocardial injury
In the present study, H9c2 cells treated with HR
were used to simulate ischemia-reperfusion injury. The change in
the expression levels of lncRNA GAS5 were detected between
normally-cultured cells and HR-induced cells by RT-qPCR. It was
revealed that lncRNA GAS5 was significantly upregulated following
HR compared with the normal control cells (P<0.05). The data
indicated that the expression levels of lncRNA GAS5 were increased
by twelve times in the HR group compared with the normal cell group
(Fig. 1A). Then, 10 pairs of
normal myocardial tissues and myocardial tissues subjected to
ischemia-reperfusion injury from male SPF Wistar rats were
collected. RNA was extracted from 10 groups of myocardial tissues
and the expression of lncRNA GAS5 was measured using RT-qPCR.
Similarly, the aforementioned conclusions were also confirmed in
the myocardial tissues of rats with ischemia-reperfusion injury
(P<0.01; Fig. 1B). The present
study reviewed the results of studies using lncRNAs that have been
reported to be associated with myocardial ischemia-reperfusion
injury. It has been reported that metastasis associated lung
adenocarcinoma transcript 1 is upregulated ~4-fold in the context
of ischemia-reperfusion mediation. Subsequent to intervention,
significant positive results were revealed in subsequent functional
tests (33). In addition, the
data revealed that the expression levels of lncRNA-regulator of
reprogramming was significantly increased by nearly 2-fold compared
with the control group in the ischemia-reperfusion injury group
(34). Therefore, the
upregulation of lncRNA is notable. The present results suggest that
lncRNA GAS5 may serve an important function in the process of
MIRI.
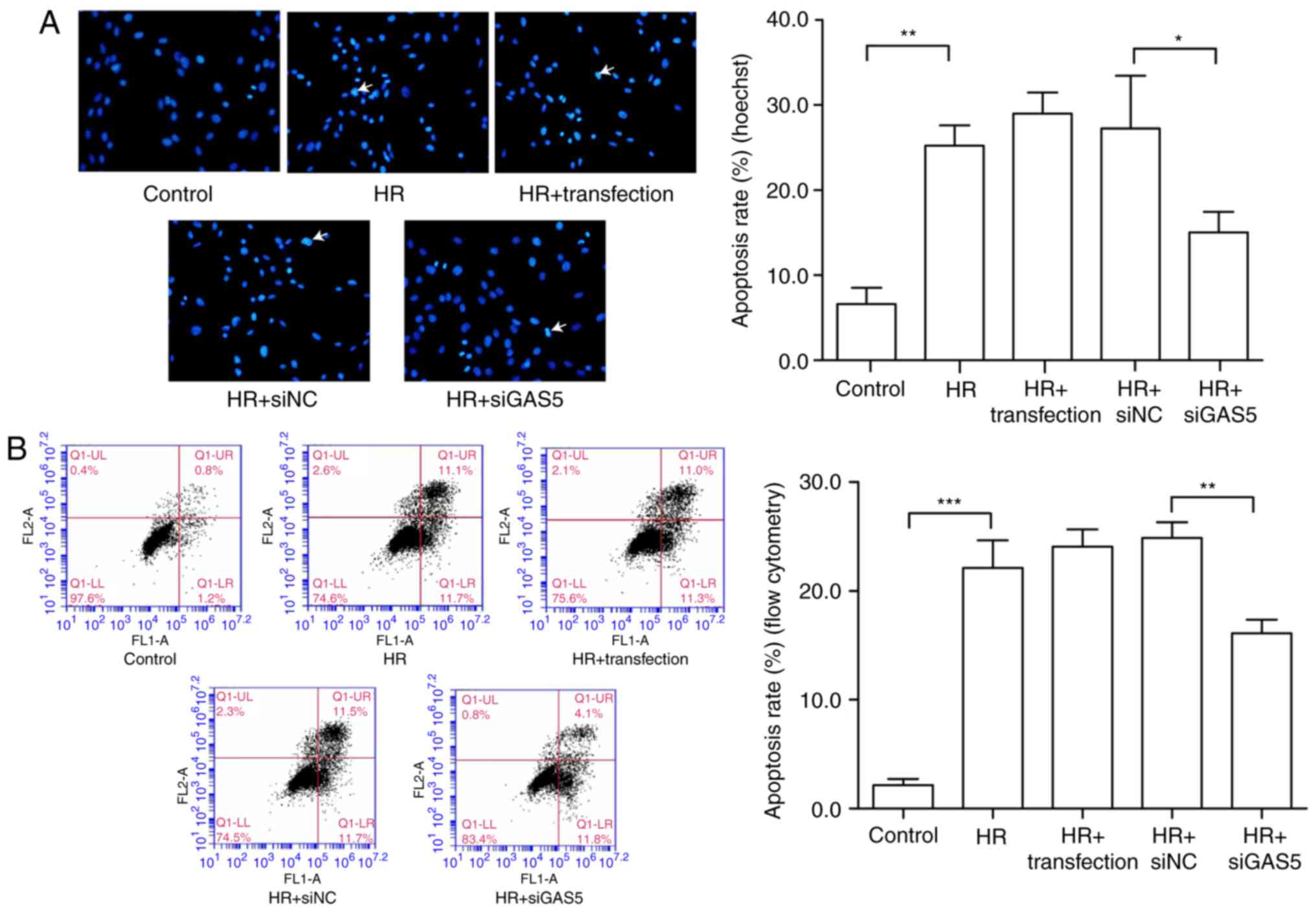
Silencing of lncRNA GAS5 may protect H9c2
cells against HR injury in H9c2 cells
The present study further investigated the
functional effects of lncRNA GAS5 in the HR model by altering the
expression of lncRNA GAS5. siRNA was transfected to successfully
silence the expression of lncRNA GAS5, as presented in Fig. 2A. The silencing efficiency of the
siRNA sequences was ~80%. The experimental group with transfection
reagent alone was used to confirm that the transfection reagent had
no significant additional damaging effect on the cells. Cell
viability was tested using a CCK-8 assay. The data revealed that
subsequent to silencing the highly expressed lncRNA GAS5, the cell
viability was restored significantly, and the cell damage caused by
HR was alleviated (P<0.05; Fig.
2B). Furthermore, the detection of changes in the levels of
myocardial necrosis markers, including LDH and CK-MB, was
performed. It was revealed that the silencing of lncRNA GAS5
significantly reduced myocardial enzyme release (P<0.01). This
further proved that the degree of cell damage was reduced (Fig. 2C and D). Therefore, the present
study preliminarily demonstrated that the silencing of lncRNA GAS5
may alleviate myocardial HR damage.
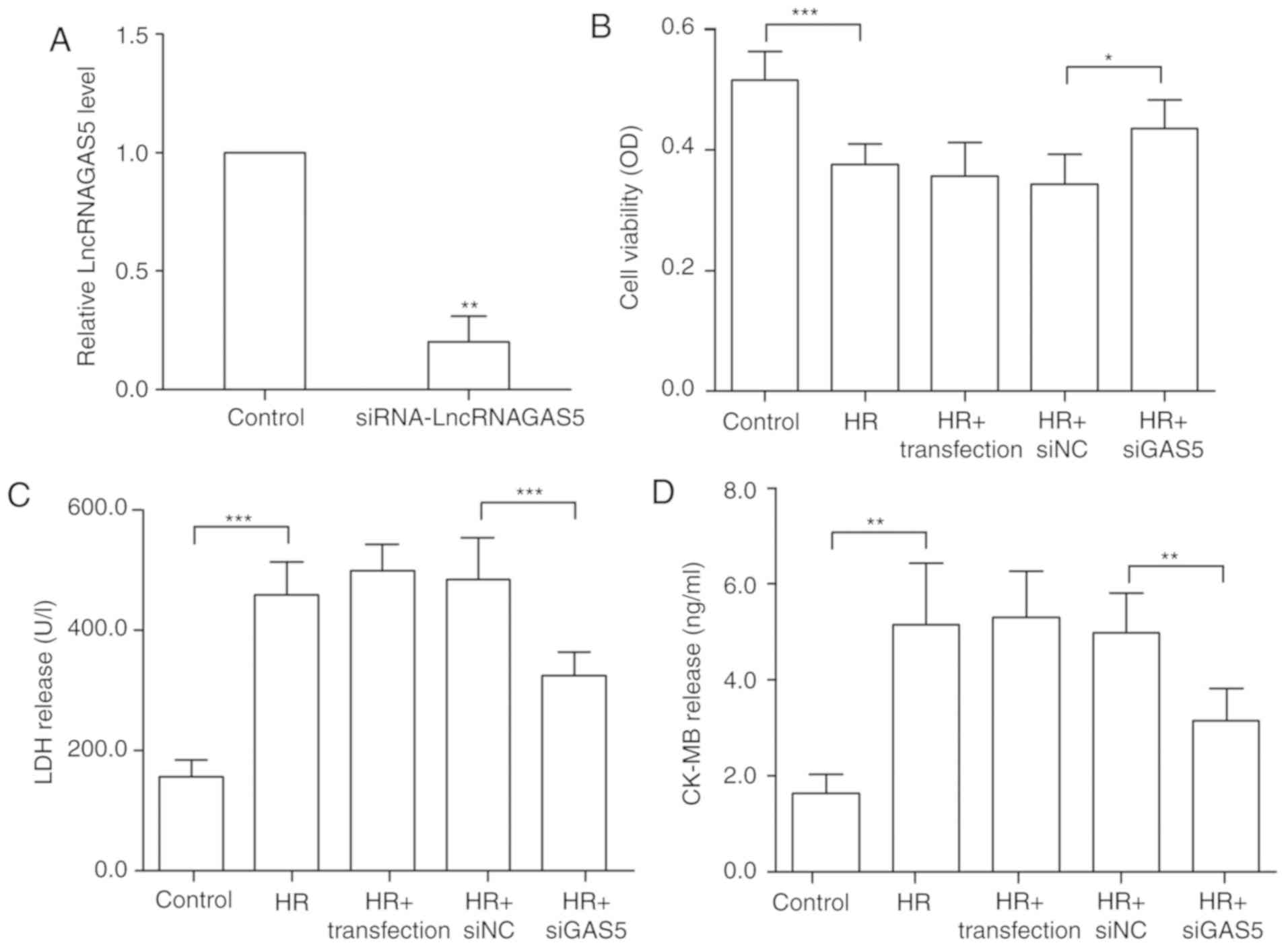 | Figure 2Silencing of lncRNA GAS5 is able to
protect H9c2 cells against HR injury. (A) H9c2 cells were
transfected using siGAS5, and it was verified that the silencing
efficiency reached 80%. **P<0.01 vs. the control. (B)
Silencing of lncRNA GAS5 significantly improves cell viability as
demonstrated by Cell Counting Kit 8 assays. (C) Release levels of
LDH. (D) Release levels of CK-MB. *P<0.05,
**P<0.01 and ***P<0.001 with
comparisons shown by lines. n=3. Data are presented as the mean ±
standard deviation. Control, cells with a normal culture; HR, cells
with hypoxia/reoxygenation injury treatment; HR+transfection, cells
with hypoxia/reoxygenation injury and transfection reagent only;
HR+siNC, cells with hypoxia/reoxygenation injury and negative
control transfection; HR+siGAS5, cells with hypoxia/reoxygenation
injury and siRNA-GAS5 transfection; lncRNA, long non-coding RNA;
GAS5, growth arrest specific 5; si/siRNA, small interfering RNA;
NC, negative control; HR, hypoxia-reoxygenation; LDH, lactate
dehydrogenase; CK-MB, creatine kinase isoenzyme; OD, optical
density. |
Targeted regulation of lncRNA GAS5 is
able to attenuate apoptosis induced by myocardial HR injury in H9c2
cells
To investigate the regulatory function of lncRNA
GAS5 in myocardial HR injury, the present study further examined
the regulation of apoptosis. The cells were first stained via
Hoechst staining, and apoptotic changes were visually observed with
the naked eye. Under a fluorescence microscope, the living cells
were diffusely and uniformly fluorescent, and a dense agglomerated
granular block fluorescence was observed in the apoptotic cell
nucleus or cytoplasm. As presented in Fig. 3A, the blue mass was the nucleus;
once apoptosis occurred, blue clumps were observed, the chromatin
shrank and highlighting occurred. It was revealed that the number
of apoptotic cells induced by HR was reduced following the
silencing of lncRNA GAS5. The apoptosis rate was expressed by
calculating the apoptotic cells and the total number of cells in
the same visual field with the naked eye. The results of the
experiment preliminarily demonstrated that lncRNA GAS5 is involved
in the regulation of apoptosis in myocardial HR injury. Then, flow
cytometry was used to further analyze cell apoptosis in the HR
model. The early and late apoptotic rates of cells were compared
prior to and following the silencing of lncRNA GAS5. The results
indicated that the silencing of lncRNA GAS5 significantly reduced
the apoptotic rates of the cells (P<0.01; Fig. 3B). As a result of the previous
experiments, it was demonstrated that lncRNA GAS5 serves a function
in regulating apoptosis in cardiomyocytes. Next, the expression of
major apoptosis-associated proteins was assessed during myocardial
ischemia-reperfusion, including the anti-apoptotic factors Bcl2 and
Bax, in addition to the pro-apoptotic factor caspase-3. The results
revealed that the protein expression levels of Bcl2 were
significantly downregulated following HR treatment (P<0.001),
while the protein expression level of Bax was significantly
upregulated (P<0.001), and the ratio of Bcl2/Bax decreased.
Protein expression exhibited a trend towards recovery subsequent to
the silencing of lncRNA GAS5 (Fig.
3C-G). Caspase-3 is a pro-apoptotic protein downstream of the
apoptotic pathway. It was revealed that there was the significantly
upregulated expression of caspase-3 following HR treatment compared
with the control (P<0.001), but the expression levels
significantly decreased following the silencing of the expression
of lncRNA GAS5 (P<0.01; Fig. 3H
and I). The results suggested that lncRNA GAS5 may serve a
function in MIRI by participating in the regulation of
apoptosis.
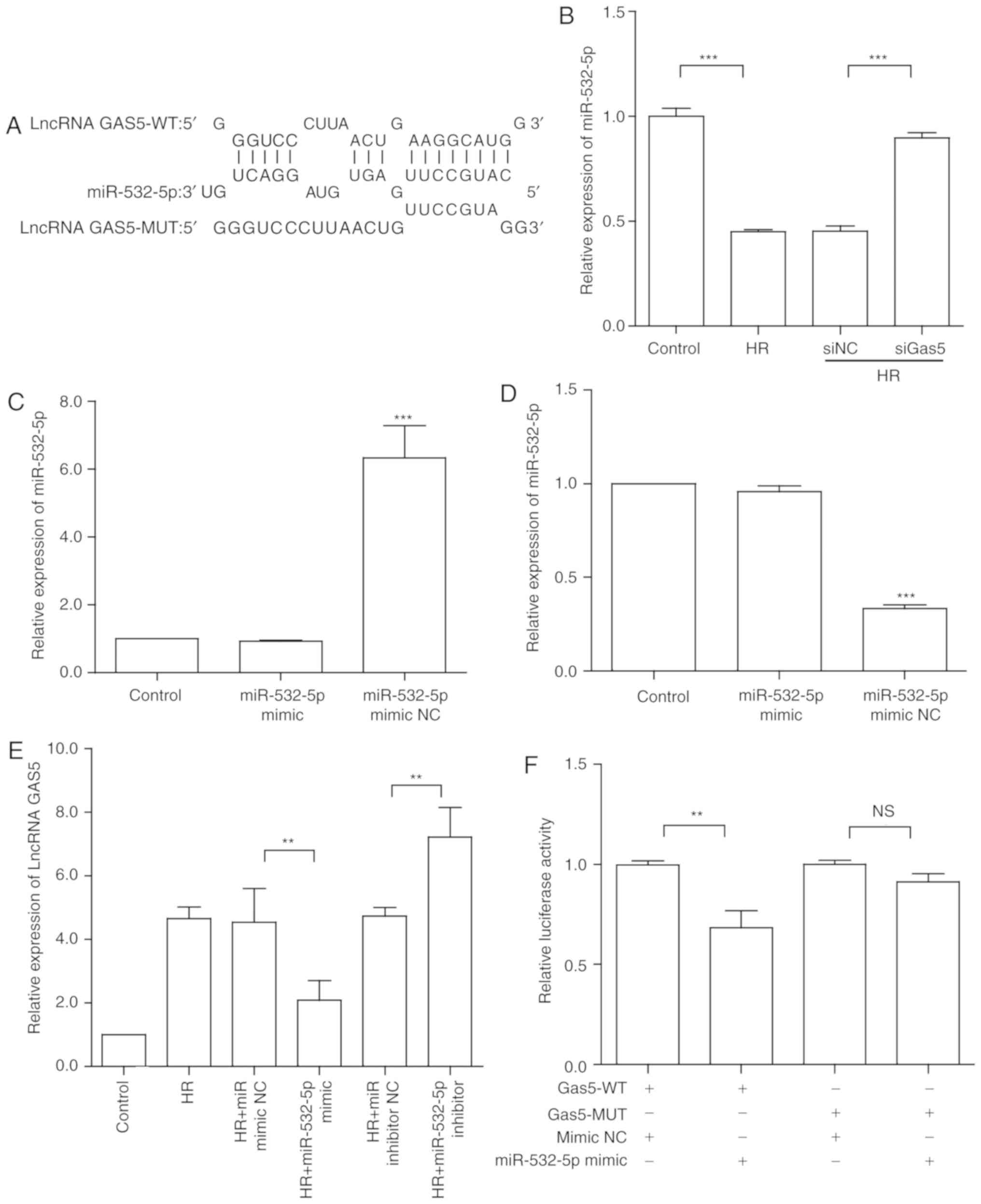
lncRNA GAS5 functions as a molecular
sponge for miR-532-5p in H9c2 cells
In order to clarify the downstream regulation of
lncRNA GAS5, previous studies were assessed, and it was revealed
that it is able to function as a competing endogenous (ceRNA) to
regulate apoptosis (35,36). To further investigate the
regulatory mechanisms of lncRNA GAS5 in MIRI, bioinformatics
analysis was used to predict target genes. miRNAs that may have
binding sites for lncRNA GAS5 were identified via predictive
software (RNAhybrid online) (37). Through screening, the present
study focused on miR-532-5p, which had the highest score. The
binding site of miR-532-5p in the lncRNA GAS5 sequence was obtained
by using bioinformatics analysis (Fig. 4A). Following that, the expression
of miR-532-5p in a model of myocardial HR was detected by RT-qPCR.
It was revealed that the expression of miR-532-5p was significantly
downregulated under HR treatment compared with the control
(P<0.001), contrary to lncRNA GAS5 expression. Furthermore,
miR-532-5p was significantly upregulated in response to lncRNA GAS5
silencing (P<0.001; Fig. 4B).
From another perspective, the expression of lncRNA GAS5 following
the silencing of miR-532-5p was assessed by RT-qPCR. miRNA mimic
and inhibitor were used to perform miRNA overexpression and
silencing, respectively. The transfection efficiency is presented
in Fig. 4C and D. The results
were consistent with the predictions; when the expression of
miR-532-5p was altered, lncRNA GAS5 also exhibited significant
corresponding expression changes (P<0.01; Fig. 4E). The results further illustrated
that miR-532-5p and lncRNA GAS5 may combine and serve a function in
myocardial HR injury. To validate the accuracy of the targets
predicted by bioinfor-matics analysis, luciferase reporter assays
were performed to further investigate the combination of lncRNA
GAS5 and miR-532-5p. The luciferase activity of GAS5-WT was
significantly reduced by miR-532-5p mimics (P<0.01). However,
the luciferase activity of GAS5-MUT was not significantly altered
(Fig. 4F). These results
confirmed that miR-532-5p is a target of lncRNA GAS5 and that
lncRNA GAS5 may function as a ceRNA in HR-induced apoptosis.
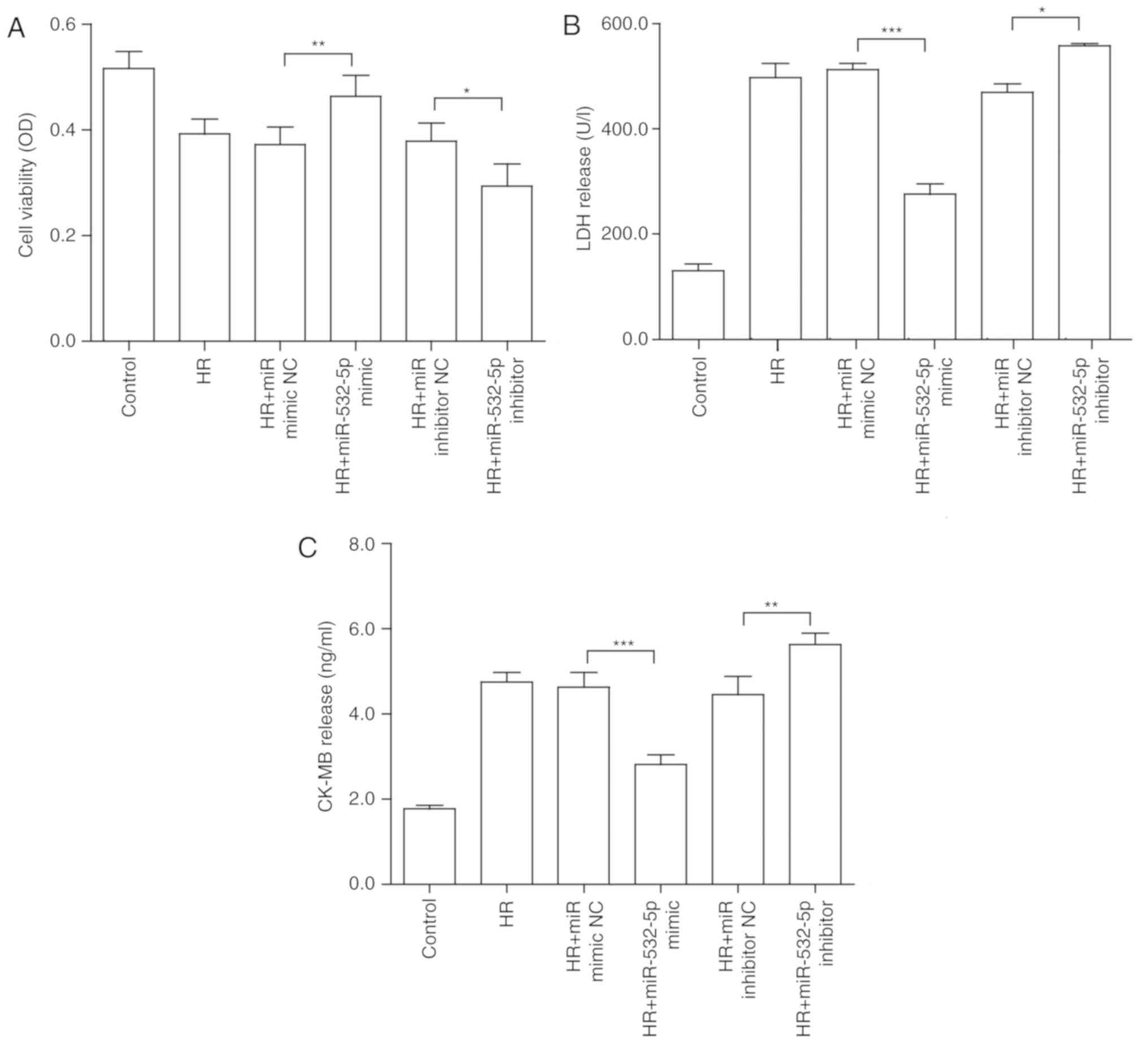
miR-532-5p serves a function in HR injury
in H9c2 cells
Ma et al (38) reported that miR-532-5p may
alleviate hypoxia-induced cardiomyocyte apoptosis. To further
confirm whether miR-532-5p serves a function in myocardial HR, the
present study investigated the biological functions of miR-532-5p
in H9c2 cells by gain- and loss-of-function analyses. miRNA mimic
and miRNA inhibitor were transfected into H9c2 cells to overexpress
and inhibit the expression of miR-532-5p, respectively. The
aforementioned cellular HR model was established. Similarly, cell
viability and myocardial enzyme indexes were analyzed. The results
of the CCK-8 assay revealed that when the miRNA mimic was
transfected, the original decreased cell viability was
significantly reversed (P<0.01). Conversely, inhibition
significantly reduced cell viability (P<0.05; Fig. 5A). In terms of myocardial
enzymology, the results for CK-MB (Fig. 5B) and LDH (Fig. 5C) were similar, further
demonstrating that overexpression may alleviate the damage to some
extent, and that inhibition may aggravate the damage.
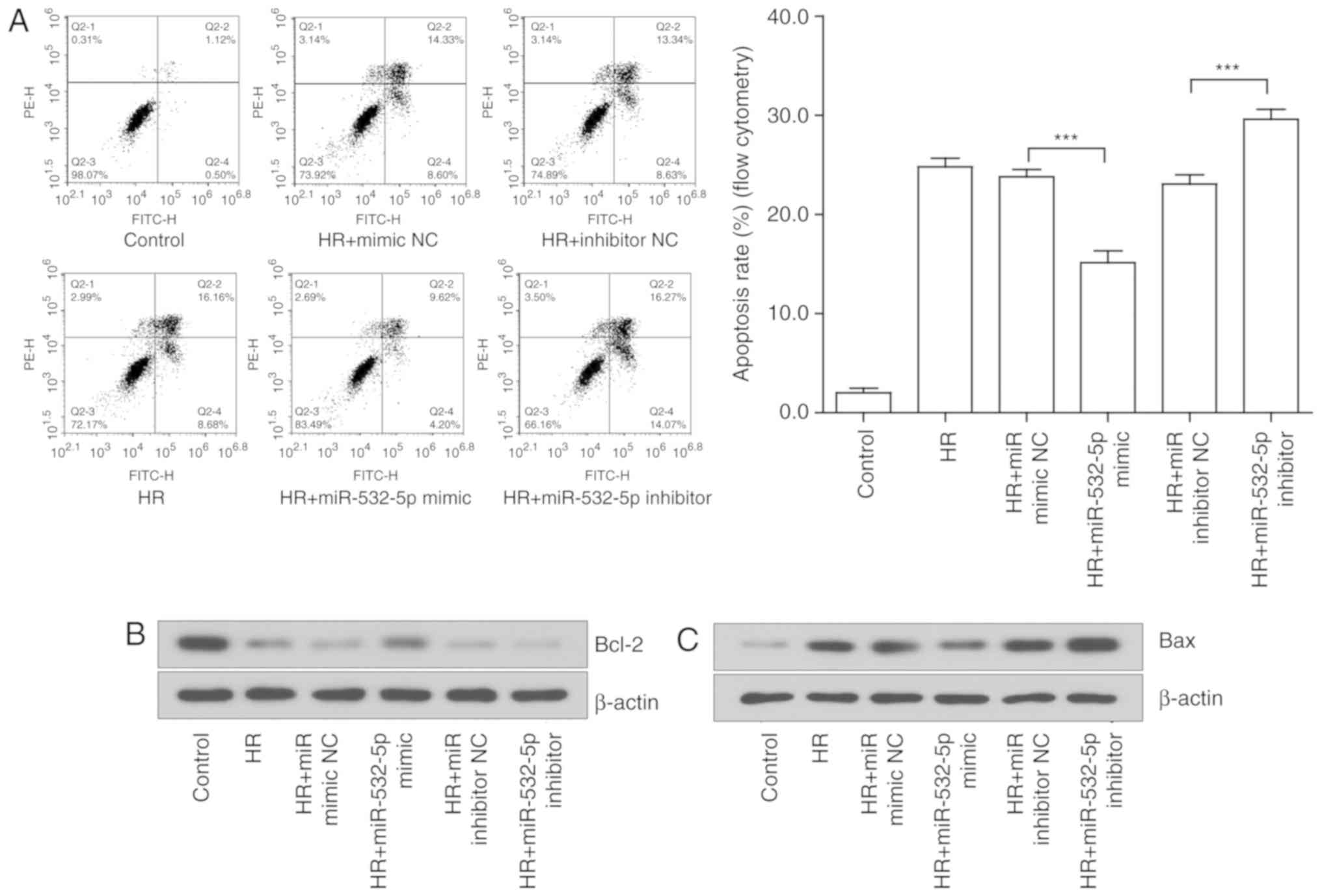
Alterations in miR-532-5p expression may
regulate the degree of apoptosis induced by HR in H9c2 cells
A flow cytometry was used to initially detect the
effects on cell apoptosis. The early and late apoptotic rates of
cells were compared prior and subsequent to altering the expression
of miR-532-5p. The results suggested that the overexpression of
miR-532-5p significantly suppressed cell apoptosis, while the
inhibition of miR-532-5p significantly aggravated cell apoptosis
(P<0.001; Fig. 6A). The
changes at the protein level further revealed the effect of
miR-532-5p on cell apoptosis. The expression of classical apoptotic
proteins was examined by western blotting. The expression trends of
Bcl-2 (Fig. 6B and D), Bax
(Fig. 6C and E) and caspase-3
(Fig. 6G and H) were consistent
with the predictions. The expression of the pro-apoptotic proteins,
which were notably induced by injury, were significantly decreased
under the action of the miRNA mimic and overexpressed under the
action of the miRNA inhibitor (P<0.05). However, the
anti-apoptotic proteins exhibited the opposite trend. Further
analysis of the ratio of Bcl-2 to Bax indicated that the effect of
the miRNA mimic was more substantial compared with that of the
miRNA inhibitor; the results were not statistically significant
(P>0.05), but a certain trend was observed (Fig. 6F). These results confirmed the
hypothesis that lncRNA GAS5 may function in HR-induced apoptosis by
binding to miR-532-5p to regulate target gene expression.
lncRNA GAS5 regulates PTEN through
miR-532-5p in HR injury
The PI3K/AKT signaling pathway is an important
pathway in the regulation of apoptosis, and it has also been
reported to serve an important anti-apoptotic function in MIRI
(9). Firstly, it was revealed
that in the H9c2 cells subjected to HR, PTEN was also significantly
upregulated compared with the control (P<0.01), consistent with
the trend of lncRNA GAS5 expression. Subsequent to further
intervention with siGAS5, PTEN also exhibited a significant
decrease (P<0.05; Fig. 7A),
and protein expression was also reduced (P<0.05; Fig. 7C and D). Bioinformatics analysis
was used to predict target binding sites between miR-532-5p and
PTEN. It was revealed using the TargetScan online database that
they have potential binding sites (Fig. 7G) (39). In addition, the same trend was
observed following the alteration of miR-532-5p alone. With the
change in miR-532-5p expression, the gene expression levels of PTEN
also changed accordingly (P<0.05; Fig. 7B). Protein expression was
consistent with the trend in gene expression (Fig. 7E and F). The results suggested
that PTEN may be a target gene for miR-532-5p, and that lncRNA GAS5
regulated PTEN expression by binding to miR-532-5p. Then, a
luciferase reporter assay was performed to verify the predicted
results. The data in Fig. 7H
revealed that the relative luciferase activity was significantly
inhibited by co-transfection using PTEN-WT and miR-532-5p mimics,
when compared with co-transfection with PTEN-WT and miR-NC
(P<0.05). The luciferase activity of PTEN-MUT was almost
unchanged. The aforementioned results suggested that PTEN is a
target gene of miR-532-5p and that lncRNA GAS5 may regulate PTEN
through miR-532-5p in HR injury.
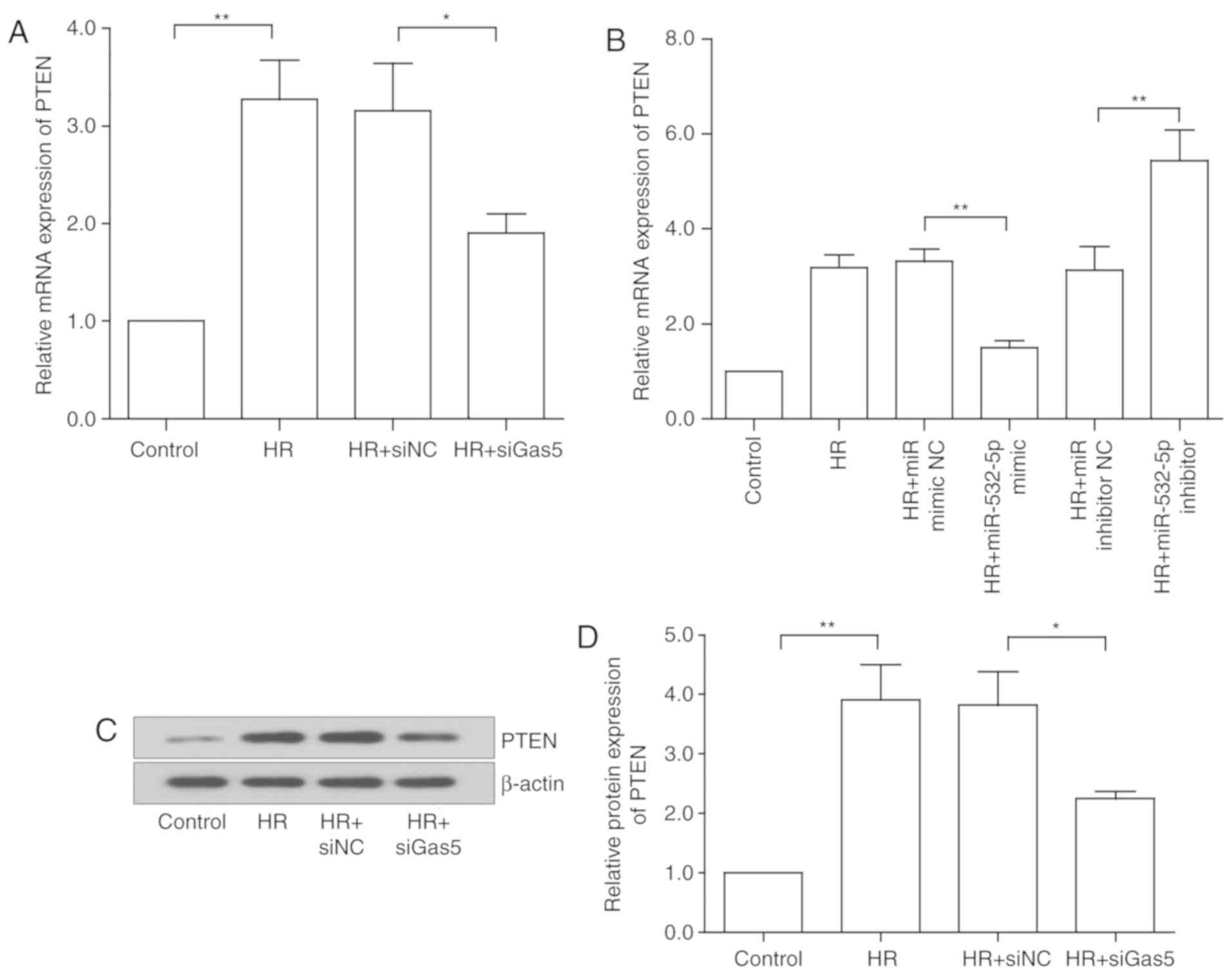 | Figure 7LncRNA GAS5 regulates PTEN through
miR-532-5p in HR injury. (A) Gene expression of PTEN following
lncRNA GAS5 intervention. (B) Gene expression of PTEN following
miR-532-5p intervention. (C) Protein expression of PTEN following
lncRNA GAS5 intervention. (D) Quantitative value of relative
protein expression levels of PTEN. (E) Protein expression of PTEN
following miR-532-5p intervention. (F) Quantitative value of
relative protein expression levels of PTEN. (G) Prediction of
miR-532-5p binding sites on the PTEN transcript. Luciferase assay
mutated sequence. (H) Association between PTEN and miR-532-5p was
demonstrated using a luciferase reporter assay.
*P<0.05, **P<0.01 and
***P<0.001 with comparisons shown by lines. Data are
presented as the mean ± standard deviation. n=3. LncRNA, long
non-coding RNA; GAS5, growth arrest specific 5; PTEN, phosphatase
and tensin homolog; miR, microRNA; HR, hypoxia-reoxygenation; si,
small interfering RNA; NC, negative control; WT, wild type; MUT,
mutant; 3′UTR, 3′ untranslated region. |
lncRNA GAS5 modulates the PI3K/AKT
signaling pathway in HR injury
Based on previous experimental results and the
conclusion of the bioinformatics analysis, a hypothesis was
proposed: lncRNA GAS5 targets PTEN by binding to miR-532-5p,
thereby inhibiting the activation of the PI3K/AKT pathway. In order
to further confirm the regulatory mechanism of the lncRNA
GAS5/miR-532-5p/PTEN axis in HR-induced apoptosis, a rescue
experiment was designed. The expression of PTEN was upregulated
during the HR of cardiomyocytes, and decreased following lncRNA
GAS5 inhibition. In the next experiment, H9c2 cells were
transfected with siGAS5 and miR-532-5p inhibitor, individually and
together, and then treated with HR. Transfection of siGAS5 alone
reduced the gene expression of PTEN. When the cells were
simultaneously transfected with miR-532-5p inhibitor, the
downregulation of PTEN was significantly reversed (P<0.05;
Fig. 8A). Regarding the protein
expression of PTEN, the results were consistent with the RT-qPCR
results (P<0.05; Fig. 8B).
Following the same test groups, the upstream and downstream
molecules involved in the action of PTEN were examined. The
expression of proteins including PI3K, AKT and their phosphorylated
forms was assessed using western blotting. The protein expression
results of PI3K and p-PI3K are presented in Fig. 8C and D. The protein expression
results of AKT and p-AKT are presented in Fig. 8E and F. The results indicated that
silencing lncRNA GAS5 activated the PI3K/AKT pathway, which may be
inhibited by miR-532-5p inhibitor. Thus, it was preliminarily
concluded that the silencing of lncRNA GAS5 promotes the activation
of the PI3K/AKT signaling pathway, potentially by sponging
miR-532-5p.
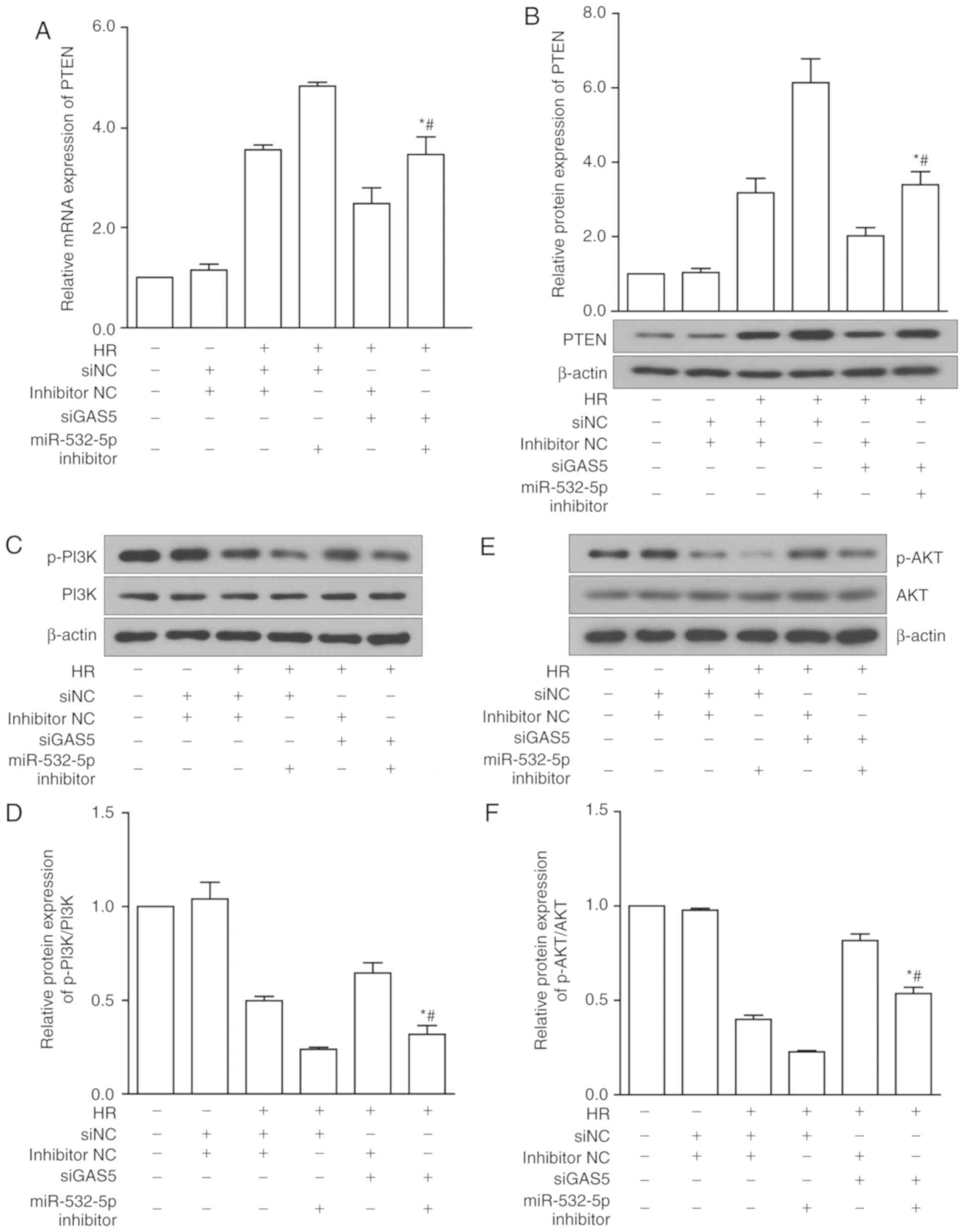 | Figure 8LncRNA GAS5 modulates the PI3K/AKT
signaling pathway in HR injury. (A) Gene expression of PTEN under
siGAS5 and miR-532-5p inhibitor alone and with co-intervention. (B)
Protein expression levels of PTEN under siGAS5 and miR-532-5p
inhibitor alone and with co-intervention. (C) Protein expression of
PI3K and p-PI3K under siGAS5 and miR-532-5p inhibitor alone and
with co-intervention. (D) Quantitative value of relative protein
expression levels of p-PI3K/PI3K. (E) Protein expression of AKT and
p-AKT under siGAS5 and miR-532-5p inhibitor alone and with
co-intervention. (F) Quantitative value of relative protein
expression levels of p-AKT/AKT. *P<0.05 vs.
HR+siNC+miR-532-5p inhibitor. #P<0.05 vs.
HR+siGAS5+inhibitor NC. Data are presented as the mean ± standard
deviation. n=3. LncRNA, long non-coding RNA; GAS5, growth arrest
specific 5; PI3K, phosphoinositide-3-kinase; AKT, protein kinase B;
p-, phosphorylated; HR, hypoxia-reoxygenation; si, small
interfering RNA; NC, negative control; PTEN, phosphatase and tensin
homolog; miR, microRNA; NC, negative control. |
Discussion
Although the study of MIRI has a long history,
researchers are still continually trying to investigate this
phenomenon. Apoptosis is the main cause of cell death caused by
ischemia-reperfusion injury, which induces the activation of the
apoptotic pathway (40,41). The PI3K/AKT signaling pathway is a
classical cell survival pathway that has been suggested to be
involved in the regulation of apoptosis induced by MIRI, mainly via
the promotion of cell survival (9,42).
The PI3K/AKT signaling pathway is regulated by a variety of
factors. PTEN is a key enzyme, functioning as an inhibitor of the
activity of AKT in this pathway, and thereby inhibiting downstream
effector molecules. It also serves an important function in MIRI
(43,44). In previous years, an increasing
number of studies focusing on the upstream regulatory factors of
PTEN have revealed that certain lncRNAs and miRNAs may regulate
their expression in a variety of ways. Ke et al (45) indicated that miR-93 may protect
against HR-induced cardiomyocyte apoptosis by inhibiting the
PI3K/AKT/PTEN signaling pathway. It has also been reported that
lncRNAs may directly or indirectly regulate PTEN during apoptosis
in tumor cells (46).
lncRNAs were previously thought to be
transcriptional noise with no biological function. With continuous
development in the life sciences, epigenetics and genomics, an
increasing number of lncRNAs have been discovered and named, and
their potential functions in the regulation of a variety of
biological processes have been discovered simultaneously (47). Although lncRNAs themselves do not
have protein-coding functions, they may indirectly exert biological
effects by binding to miRNAs, protein-coding RNA or proteins, and
the mechanism of action is very complicated (48). Tumors were an early focus of
lncRNA research (49). Gradually,
other lncRNAs have been revealed to serve a function in systemic
diseases with biological processes similar to those of a tumor
(50,51). Due to their pathophysiological
characteristics, certain lncRNAs have also been revealed in
cardiovascular diseases. The lncRNA metastasis-associated lung
adenocarcinoma transcript 1 has been reported to be involved in the
regulation of apoptosis (33) and
inflammatory responses (52)
induced by MIRI.
Previous research established that lncRNA GAS5 is
abundant in cells whose growth has been arrested due to a lack of
nutrients or growth factors (23). The function of lncRNA GAS5 in the
regulation of apoptosis was initially discovered in a functional
cloning experiment, in which the researchers isolated a portion of
the lncRNA GAS5 sequence that inhibits apoptosis from γ-irradiated
mouse thymoma cells (53). Kino
et al (23) revealed that
lncRNA GAS5 may sensitize cells to apoptosis by suppressing the
glucocorticoid-mediated induction of a number of responsive genes,
including the one encoding the cellular inhibitor of apoptosis
protein. In the past two decades, a number of researchers have
reported that lncRNA GAS5 participates in the processes of
proliferation, migration and invasion in numerous tumor types,
including colorectal cancer, breast cancer, prostate cancer,
non-small-cell lung cancer and thyroid cancer (24,25,27,54). However, there is very little
published research in the field of ischemic heart disease
associated with lncRNA GAS5. One previous study on cardiovascular
disease was on myocardial fibrosis. LncRNA GAS5 was revealed to be
expressed at low levels in cardiac fibrosis tissues, in addition to
activated cardiac fibroblasts. Overexpression of lncRNA GAS5
inhibited the proliferation of cardiac fibroblasts. It was further
discovered that lncRNA GAS5 may regulate the expression of PTEN by
binding to miRNA-21 to affect myocardial fibrosis (36).
As mentioned earlier, the process of
ischemia-reperfusion may involve multiple mechanisms. However,
apoptosis is the main cause of cell death induced by
ischemia-reperfusion. Furthermore, previous studies have reported
that lncRNA GAS5 has been revealed to be expressed at low levels in
tumor tissues, which promotes apoptosis and inhibits cell growth
(27). Therefore, a hypothesis
was proposed, that lncRNA GAS5 may be involved in the regulation of
apoptosis induced by MIRI. By comparing and detecting the
difference in lncRNA GAS5 expression in cardiomyocytes and tissue
samples mediated by ischemia-reperfusion injury, it was revealed to
be increased following ischemia-reperfusion, and these results
supported the hypothesis. The present study then further
investigated the potential function of lncRNA GAS5 in MIRI. By
intervening in the expression of lncRNA GAS5, changes in myocardial
necrosis markers, cell viability and cell apoptosis were observed.
Through Hoechst staining and flow cytometry, it was revealed that
silencing lncRNA GAS5 reduced the apoptosis rate. Therefore, the
present study focused on the assessment of the apoptotic regulatory
function of lncRNA GAS5. Apoptosis signal transduction molecules
Bcl-2, Bax and caspase-3 are important signaling molecules in
apoptosis (55). The expression
levels of the above apoptotic proteins were further examined, and
the conclusions were consistent with the aforementioned
assumptions.
The present results are consistent with the
experimental results from Liu et al (56); however, they did not conduct
in-depth research on the regulatory mechanism, and the specific
mechanism remains unclear. According to the reported mechanism of
action of lncRNA GAS5 in other diseases, the present study focused
on the regulation of PTEN by lncRNA GAS5. As previously stated,
PTEN itself is a relatively classical protein that promotes
apoptosis (45). Although the
mechanisms of lncRNAs are complex, the biological function of
lncRNA GAS5 is mostly mediated through the ceRNA mechanism,
according to previous studies (57,58). lncRNA GAS5 has also been reported
to be involved in biological processes including apoptosis, through
a variety of mechanisms associated with PTEN (36). It was hypothesized that lncRNA
GAS5 may function through PTEN in apoptosis induced by MIRI.
Following the intervention of lncRNA GAS5, the present study
examined the expression of PTEN, and it was revealed that the
inhibition of lncRNA GAS5 did reduce the high expression of PTEN in
MIRI. Furthermore, through bioinformatics analysis, combined with
the ceRNA mechanism, it was revealed that lncRNA GAS5 may function
as an endogenous RNA, binding to miR-532-5p and regulating the
expression of PTEN. To demonstrate this prediction, a luciferase
reporter assay was performed on the predicted binding targets. The
results confirmed the predictions. The expression levels of
miR-532-5p were examined, and it was revealed that it was
downregulated under HR treatment and had the opposite trend to the
expression of lncRNA GAS5 and PTEN. miR-532-5p has been
demonstrated to be able to alleviate hypoxia-induced apoptosis by
targeting programmed cell death 4 in H9c2 cells (37). Furthermore, following miR-532-5p
mimic and inhibitor treatment, it was revealed that HR-induced
apoptosis was notably altered in the present study. In order to
confirm the ceRNA mechanism, a rescue experiment was conducted. The
present experiments further confirmed the regulatory function and
mechanism of lncRNA GAS5 in MIRI.
In conclusion, as a pro-apoptotic factor, lncRNA
GAS5 serves a function in MIRI, and it may reduce the apoptosis
induced by injury to a certain extent. In-depth study suggested
that lncRNA GAS5 may bind to miR-532-5p, so that miR-532-5p loses
its regulatory effect on its target PTEN, resulting in the
overexpression of PTEN and consequent inhibition of the PI3K/AKT
pathway, thereby promoting apoptosis. Therefore, the inhibition of
lncRNA GAS5 may improve cell survival, reduce the occurrence of
apoptosis and ultimately alleviate MIRI. MIRI is a common clinical
reperfusion complication. The present study may provide novel ideas
for protecting against myocardial injury, but further research is
still required.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (grant nos. 81670320 and
31741047) and China Medical University Youth Backbone Support
Program (grant no. QGZ2018021).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DJ, YH and NW conceived and designed the
experiments. YH, FX and SL performed the experiments. YH and FX
analyzed the data. YH wrote the paper. YH and FX revised the
manuscript. All the authors read and approved the final paper.
Ethics approval and consent to
participate
The experimental protocol was approved by the
Institutional Ethics Committee of China Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Gerczuk PZ and Kloner RA: An update on
cardioprotection: A review of the latest adjunctive therapies to
limit myocardial infarction size in clinical trials. J Am Coll
Cardiol. 59:969–978. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pollack A, Mohanty BD, Handa R, Looser PM,
Fuster V, King S III and Sharma SK: Preventive stenting in acute
myocardial infarction. JACC Cardiovasc Interv. 8:131–138. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Majidi M, Kosinski AS, Al-Khatib SM,
Lemmert ME, Smolders L, van Weert A, Reiber JH, Tzivoni D, Bär FW,
Wellens HJ, et al: Reperfusion ventricular arrhythmia 'bursts'
predict larger infarct size despite TIMI 3 flow restoration with
primary angioplasty for anterior ST-elevation myocardial
infarction. Eur Heart J. 30:757–764. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Taggart DP and Neubauer S: Incidence,
predictors, and significance of abnormal cardiac enzyme rise in
patients treated with bypass surgery in the arterial
revascularization therapies study (ARTS). Circulation. 106:e55–e56.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Eltzschig HK and Eckle T: Ischemia and
reperfusion-from mechanism to translation. Nat Med. 17:1391–1401.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Garcia-Dorado D, Ruiz-Meana M, Inserte J,
Rodriguez-Sinovas A and Piper HM: Calcium-mediated cell death
during myocardial reperfusion. Cardiovasc Res. 94:168–180. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Koshinuma S, Miyamae M, Kaneda K, Kotani J
and Figueredo VM: Combination of necroptosis and apoptosis
inhibition enhances cardioprotection against myocardial
isch-emia-reperfusion injury. J Anesth. 28:235–241. 2014.
View Article : Google Scholar
|
|
8
|
Vilahur G and Badimon L:
Ischemia/reperfusion activates myocardial innate immune response:
The key role of the toll-like receptor. Front Physiol. 5:4962014.
View Article : Google Scholar
|
|
9
|
Tang L, Mo Y, Li Y, Zhong Y, He S, Zhang
Y, Tang Y, Fu S, Wang X and Chen A: Urolithin A alleviates
myocardial ischemia/reperfusion injury via PI3K/Akt pathway.
Biochem Biophys Res Commun. 486:774–780. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ren GD, Cui Y, Li WL, Li FF and Han XY:
Research on cardioprotective effect of irbesartan in rats with
myocardial ischemia-reperfusion injury through MAPK-ERK signaling
pathway. Eur Rev Med Pharmacol Sci. 23:5487–5494. 2019.PubMed/NCBI
|
|
11
|
Bainey KR and Armstrong PW: Clinical
perspectives on reperfusion injury in acute myocardial infarction.
Am Heart J. 167:637–645. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ong SB, Katwadi K, Kwek XY, Ismail NI,
Chinda K, Ong SG and Hausenloy DJ: Non-coding RNAs as therapeutic
targets for preventing myocardial ischemia-reperfusion injury.
Expert Opin Ther Targets. 22:247–261. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hombach S and Kretz M: Non-coding RNAs:
Classification, biology and functioning. Adv Exp Med Biol.
937:3–17. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qiu MT, Hu JW, Yin R and Xu L: Long
noncoding RNA: An emerging paradigm of cancer research. Tumour
Biol. 34:613–620. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Vitiello M, Tuccoli A and Poliseno L: Long
non-coding RNAs in cancer: Implications for personalized therapy.
Cell Oncol (Dordr). 38:17–28. 2015. View Article : Google Scholar
|
|
16
|
Zangrando J, Zhang L, Vausort M, Maskali
F, Marie PY, Wagner DR and Devaux Y: Identification of candidate
long non-coding RNAs in response to myocardial infarction. BMC
Genomics. 15:4602014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Saddic LA, Sigurdsson MI, Chang TW,
Mazaika E, Heydarpour M, Shernan SK, Seidman CE, Seidman JG, Aranki
SF, Body SC and Muehlschlegel JD: The long noncoding RNA landscape
of the ischemic human left ventricle. Circ Cardiovasc Genet.
10:pii: e001534. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gong LC, Xu HM, Guo GL, Zhang T, Shi JW
and Chang C: Long non-coding RNA H19 protects H9c2 cells against
hypoxia-induced injury by targeting MicroRNA-139. Cell Physiol
Biochem. 44:857–869. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu CY, Zhang YH, Li RB, Zhou LY, An T,
Zhang RC, Zhai M, Huang Y, Yan KW, Dong YH, et al: LncRNA CAIF
inhibits autophagy and attenuates myocardial infarction by blocking
p53-mediated myocardin transcription. Nat Commun. 9:292018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Y, Li G, Lu H, Li W, Li X, Liu H, Li
X, Li T and Yu B: Expression profiling and ontology analysis of
long noncoding RNAs in post-ischemic heart and their implied roles
in isch-emia/reperfusion injury. Gene. 543:15–21. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Schneider C, King RM and Philipson L:
Genes specifically expressed at growth arrest of mammalian cells.
Cell. 54:787–793. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Smith CM and Steitz JA: Classification of
gas5 as a multi-small-nucleolar-RNA (snoRNA) host gene and a member
of the 5′-terminal oligopyrimidine gene family reveals common
features of snoRNA host genes. Mol Cell Biol. 18:6897–6909. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kino T, Hurt DE, Ichijo T, Nader N and
Chrousos GP: Noncoding RNA gas5 is a growth arrest- and
starvation-associated repressor of the glucocorticoid receptor. Sci
Signal. 3:ra82010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pickard MR, Mourtada-Maarabouni M and
Williams GT: Long non-coding RNA GAS5 regulates apoptosis in
prostate cancer cell lines. Biochim Biophys Acta. 1832:1613–1623.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li J, Wang Y, Zhang CG, Xiao HJ, Hu JM and
He JD: Effect of long non-coding RNA Gas5 on proliferation,
migration, invasion and apoptosis of colorectal cancer HT-29 cell
line (vol 18, 4, 2018). Cancer Cell Int. 18:42018. View Article : Google Scholar
|
|
26
|
Cao Q, Wang N, Qi J, Gu Z and Shen H: Long
noncoding RNAGAS5 acts as a tumor suppressor in bladder
transitional cell carcinoma via regulation of chemokine (CC motif)
ligand 1 expression. Mol Med Rep. 13:27–34. 2016. View Article : Google Scholar
|
|
27
|
Pickard MR and Williams GT: Molecular and
cellular mechanisms of action of tumour suppressor GAS5 LncRNA.
Genes (Basel). 6:484–499. 2015. View Article : Google Scholar
|
|
28
|
Kastenmayer RJ, Moore RM, Bright AL,
Torres-Cruz R and Elkins WR: Select agent and toxin regulations:
Beyond the Eighth edition of the guide for the care and use of
laboratory animals. J Am Assoc Lab Anim Sci. 51:333–338.
2012.PubMed/NCBI
|
|
29
|
Wu N, Li WN, Shu WQ and Jia DL: Protective
effect of picroside II on myocardial ischemia reperfusion injury in
rats. Drug Des Dev Ther. 8:545–554. 2014.
|
|
30
|
Kilkenny C, Browne WJ, Cuthill IC, Emerson
M and Altman DG: Improving bioscience research reporting: The
ARRIVE guidelines for reporting animal research. Vet Clin Path.
41:27–31. 2012. View Article : Google Scholar
|
|
31
|
Murphy E and Steenbergen C: Mechanisms
underlying acute protection from cardiac ischemia-reperfusion
injury. Physiol Rev. 88:581–609. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(T)(-Delta Delta C) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
33
|
Sun R and Zhang L: Long non-coding RNA
MALAT1 regulates cardiomyocytes apoptosis after hypoxia/reperfusion
injury via modulating miR-200a-3p/PDCD4 axis. Biomed Pharmacother.
111:1036–1045. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang W, Li Y and Wang P: Long non-coding
RNA-ROR aggravates myocardial ischemia/reperfusion injury. Braz J
Med Biol Res. 51:e65552018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zeng B, Li Y, Jiang F, Wei C, Chen G,
Zhang W, Zhao W and Yu D: LncRNA GAS5 suppresses proliferation,
migration, invasion, and epithelial-mesenchymal transition in oral
squamous cell carcinoma by regulating the miR-21/PTEN axis. Exp
Cell Res. 374:365–373. 2019. View Article : Google Scholar
|
|
36
|
Tao H, Zhang JG, Qin RH, Dai C, Shi P,
Yang JJ, Deng ZY and Shi KH: LncRNA GAS5 controls cardiac
fibroblast activation and fibrosis by targeting miR-21 via
PTEN/MMP-2 signaling pathway. Toxicology. 386:11–18. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rehmsmeier M, Steffen P, Hochsmann M and
Giegerich R: Fast and effective prediction of microRNA/target
duplexes. RNA. 10:1507–1517. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ma J, Zhang J, Wang Y, Long K, Wang X, Jin
L, Tang Q, Zhu L, Tang G, Li X and Li M: MiR-532-5p alleviates
hypoxia-induced cardiomyocyte apoptosis by targeting PDCD4. Gene.
675:36–43. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:2015. View Article : Google Scholar
|
|
40
|
Han X, Shi H, Liu K, Zhong L, Wang F and
You Q: Protective effect of gastrodin on myocardial
ischemia-reperfusion injury and the expression of Bax and Bcl-2.
Exp Ther Med. 17:4389–4394. 2019.PubMed/NCBI
|
|
41
|
Liu S, Wu N, Miao J, Huang Z, Li X, Jia P,
Guo Y and Jia D: Protective effect of morin on myocardial
ischemia-reperfusion injury in rats. Int J Mol Med. 42:1379–1390.
2018.PubMed/NCBI
|
|
42
|
Zhang BF, Jiang H, Chen J, Guo X, Li Y, Hu
Q and Yang S: Nobiletin ameliorates myocardial ischemia and
reperfusion injury by attenuating endoplasmic reticulum
stress-associated apoptosis through regulation of the PI3K/AKT
signal pathway. Int Immunopharmacol. 73:98–107. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Siddall HK, Warrell CE, Yellon DM and
Mocanu MM: Ischemia-reperfusion injury and cardioprotection:
Investigating PTEN, the phosphatase that negatively regulates PI3K,
using a congenital model of PTEN haploinsufficiency. Basic Res
Cardiol. 103:560–568. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ruan H, Li J, Ren S, Gao J, Li G, Kim R,
Wu H and Wang Y: Inducible and cardiac specific PTEN inactivation
protects ischemia/reperfusion injury. J Mol Cell Cardiol.
46:193–200. 2009. View Article : Google Scholar
|
|
45
|
Ke ZP, Xu P, Shi Y and Gao AM: MicroRNA-93
inhibits ischemia-reperfusion induced cardiomyocyte apoptosis by
targeting PTEN. Oncotarget. 7:28796–28805. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wei H, Yang Z and Lin B: Overexpression of
long non coding RNA CA3-AS1 suppresses proliferation, invasion and
promotes apoptosis via miRNA-93/PTEN axis in colorectal cancer.
Gene. 687:9–15. 2019. View Article : Google Scholar
|
|
47
|
Cipolla GA, de Oliveira JC, Salviano-Silva
A, Lobo-Alves SC, Lemos DS, Oliveira LC, Jucoski TS, Mathias C,
Pedroso GA, Zambalde EP and Gradia DF: Long non-coding RNAs in
multifactorial diseases: Another layer of complexity. Noncoding
RNA. 4:pii: E13. 2018.PubMed/NCBI
|
|
48
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sanchez Calle A, Kawamura Y, Yamamoto Y,
Takeshita F and Ochiya T: Emerging roles of long non-coding RNA in
cancer. Cancer Sci. 109:2093–2100. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Smolle MA and Pichler M: The role of long
non-coding RNAs in osteosarcoma. Noncoding RNA. 4:pii: E7.
2018.PubMed/NCBI
|
|
51
|
Weirick T, Militello G and Uchida S: Long
non-coding RNAs in endothelial biology. Front Physiol. 9:5222018.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang S, Yu W, Chen J, Yao T and Deng F:
LncRNA MALAT1 sponges miR-203 to promote inflammation in myocardial
ischemia-reperfusion injury. Int J Cardiol. 268:2452018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Williams GT, Hughes JP, Stoneman V,
Anderson CL, McCarthy NJ, Mourtada-Maarabouni M, Pickard M, Hedge
VL, Trayner I and Farzaneh F: Isolation of genes controlling
apoptosis through their effects on cell survival. Gene Ther Mol
Biol. 10:255–261. 2006.
|
|
54
|
Esmatabadi MJD, Motamedrad M and
Sadeghizadeh M: Down-regulation of lncRNA, GAS5 decreases
chemotherapeutic effect of dendrosomal curcumin (DNC) in breast
cancer cells. Phytomedicine. 42:56–65. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Scorrano L and Korsmeyer SJ: Mechanisms of
cytochrome c release by proapoptotic BCL-2 family members. Biochem
Biophys Res Commun. 304:437–444. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Liu SD, Meng WX, Xu L, Chi C, Sun X and
Liu HY: GAS5 promotes myocardial apoptosis in myocardial
ischemia-reperfusion injury via upregulating LAS1 expression. Eur
Rev Med Pharmacol Sci. 22:8447–8453. 2018.PubMed/NCBI
|
|
57
|
Xue D, Zhou C, Lu H, Xu R, Xu X and He X:
LncRNA GAS5 inhibits proliferation and progression of prostate
cancer by targeting miR-103 through AKT/mTOR signaling pathway.
Tumor Biol. 37:16187–16197. 2016. View Article : Google Scholar
|
|
58
|
Yang W, Hong L, Xu X, Wang Q, Huang J and
Jiang L: LncRNA GAS5 suppresses the tumorigenesis of cervical
cancer by downregulating miR-196a and miR-205. Tumor Biol.
39:10104283177113152017. View Article : Google Scholar
|