Introduction
Cerebral ischemia/reperfusion (I/R) injury is a
leading cause of mortality and permanent adult disability
worldwide, causing a significant clinical and socioeconomic impact
(1). Cerebral I/R injury is a
complex pathophysiologic process, during which apoptosis, oxidative
stress, excitotoxicity, inflammation and mitochondrial dysfunction
are all involved (2,3). Among these, oxidative stress caused
by the excess production of reactive oxygen species (ROS) can
induce protein dysfunction, DNA damage and lipid peroxidation,
resulting in neuronal apoptosis and death, playing a pivotal role
in the process of cerebral I/R injury (4,5).
To date, significant efforts have been made to mitigate neuronal
injury following cerebral injury; however, no effective treatment
strategies are currently available (6,7).
Therefore, the search for suitable therapeutic agents to alleviate
oxidative stress may be an effective means of treating cerebral
I/R-induced nervous system injury, which continues to be a major
research endeavor.
In recent years, the benefits of traditional Chinese
medicine conferring neuroprotective effects have been increasingly
investigated in cerebral I/R injury (8-10).
Curcumin, a natural antioxidant mainly extracted from the root of
turmeric (Curcuma longa), exhibits a range of
pharmacological effects, including anticancer, anti-apoptosis,
ant-inflammatory and antioxidant effects (11-13). Curcumin can cross the blood-brain
barrier and exerts neuroprotective effects in central neurological
disease (14). Accumulating
evidence has indicated that the most vital biological function of
curcumin associated with neuroprotection is its antioxidant effect,
which can improve the antioxidant ability of neurons in cerebral
ischemia (15,16). Although emerging studies have
indicated the neuroprotective effects of curcumin against cerebral
I/R injury, the exact mechanisms underlying these effects are not
yet fully understood.
Apurinic/apyrimidinic endonuclease 1 (APE1) is a
multifunctional protein involved in the base-excision repair of
oxidative DNA damage and in the redox activation of transcription
factors (17). Recently, several
studies have demonstrated that APE1 is implicated in the
development and progression of cerebral I/R injury, which is
conversely associated with the reduction of oxidative DNA damage,
while APE1 expression has been reported to decrease following
ischemic injury (18,19). In addition, the overexpression of
APE1 induced by an APE-mimicking peptide or adenovirus-mediated
APE1 following reperfusion increased DNA repair, attenuated
apoptosis and reduced the infarct volume (20,21). It has also been demonstrated that
APE1 is an attractive target for neuroprotection and is involved in
neuroprotection against cerebral I/R injury (22,23). Stetler et al (23) revealed that APE1 was required for
pituitary adenylate cyclase-activating polypeptide (PACAP)-induced
neuroprotection against global cerebral ischemia. However, the
direct contribution of APE1 to the neuroprotective effects of
curcumin has yet to be established.
The present study was undertaken to determine the
roles of APE1 in the protective effects of curcumin against
cerebral I/R injury, as well as to identify the molecular
mechanisms through which curcumin affects SH-SY5Y neuronal cells
subjected to oxygen-glucose deprivation/reperfusion (OGD/R), a
commonly used in vitro model of cerebral I/R injury
(24,25). It was observed that curcumin
protected the SY-SH5Y cells against OGD/R injury by upregulating
APE1 expression, which is associated with the regulation of the
phosphatidylinositol 3-kinase/protein kinase B (PI3K/AKT) signaling
pathway. The results indicated that APE1 is essential for
curcumin-induced neuroprotection.
Materials and methods
Reagents and antibodies
Curcumin was dissolved in 0.01% dimethyl sulfoxide
(both purchased from Sigma-Aldrich; Merck KGaA) to prepare a 10
mmol/l stock solution, which was then stored at -20̊C. Drug stocks
were prepared for use to avoid repeated freeze-thaw cycles and
diluted to the desired concentration, as needed. High-glucose
Dulbecco's modified Eagle's medium (DMEM; Gibco), fetal bovine
serum (FBS; Gibco), TRIzol reagent (Invitrogen) and Lipofectamine
RNAiMAX (Invitrogen) were obtained from Thermo Fisher Scientific,
Inc. The Cell Counting Kit-8 (CCK-8) assay kit was supplied by
Dojindo Molecular Technologies, Inc., while the Cytotoxicity
Detection kit was purchased from Roche Applied Science. The BCA
protein assay kit, RIPA lysis buffer and BeyoECL Star were obtained
from Beyotime Institute of Biotechnology. The reverse transcription
(RT) kit and ABI Prism 7500 Real-Time PCR system with the
SYBR® RT-PCR kit were obtained from Takara Bio, Inc. and
Thermo Fisher Scientific, Inc. Antibodies against GAPDH (cat. no.
5174), B-cell lymphoma 2 (Bcl-2; cat. no. 3498), Bcl-2-associated X
protein (Bax; cat. no. 14796), p-AKT (cat. no. 13038), AKT (cat.
no. 4685), p-PI3K (cat. no. 17366) and PI3K (cat. no. 4249) were
provided by Cell Signaling Technology. The antibody against NADPH
oxidase 2 (NOX2) was obtained from Abcam (cat. no. ab131083).
Cells and cell culture
SH-SY5Y neuroblastoma cells were obtained from the
American Type Culture Collection (cat. no. CRL-2266) and cultured
in high-glucose DMEM supplemented with 10% (v/v) FBS, 1% (v/v)
penicillin (100 µg/ml) and 100 U/ml streptomycin at 37̊C in
a humidified incubator containing an atmosphere of 5%
CO2 and 95% air. The cell culture medium was changed
every 2-3 days.
OGD and OGD/R models and treatment
In order to establish an in vitro cerebral
ischemia model, OGD exposure was performed by replacing the culture
medium of the cells with glucose-free DMEM without serum, and
placing the cells in a controlled humidified hypoxic glove box (Coy
Laboratory Products, Inc.) supplemented with a 0% O2, 5%
CO2 and 95% N2 gas mixture for 1 h at 37̊C.
At the end of the OGD process, the anoxic glucose-free medium was
replaced with normal DMEM, and the cells were incubated at 37°C in
a humidified incubator containing an atmosphere of 5%
CO2 and 95% air for 24 h of reperfusion, establishing an
OGD/R model. To illustrate the protection of curcumin against OGD/R
injury, SH-SY5Y cells were treated with curcumin (1, 5, 10 and 15
µl) for 24 h immediately after 1 h of OGD exposure during
reperfusion. To investigate the role of APE1 in this process,
SH-SY5Y cells were transfected with chemically synthesized small
interfering RNA (siRNA) for 6 h, followed by treatment with
curcumin (10 µl) for 24 h immediately after 1 h of OGD
exposure during reperfusion. To demonstrate the function of the
PI3K/Akt signaling pathway, SH-SY5Y cells were pretreated with 10
µM LY294002 (a PI3K/Akt inhibitor) for 2 h, and then treated
with curcumin (10 µl) for 24 h immediately after 1 h of OGD
exposure during reperfusion.
APE1 siRNA transfection
siRNA targeting APE1 (APE1t) and scrambled siRNA
(APE1s) were purchased from Bioneer Corporation. Before SH-SY5Y
neuronal cells were subjected to OGD injury, they were transfected
with 20 nM chemically synthesized APE1t (forward, 5′-GUC UGG UAC
GAC UGG AGU ACC-3′, and reverse, 5′-UAC UCC AGU CGU ACC AGA CCU-3′)
or APE1s (forward, 5′-CCA UGA GGU CAG CAU GGU CUG-3′, and reverse,
5′GAC CAU GCU GAC CUC AUG GAA-3′) using Lipofectamine RNAiMAX
according to the manufacturer protocol. Following transfection for
6 h, the culture medium was changed with fresh DMEM, and the cells
were used in subsequent experiments. The transfection efficiency
was determined by western blot analysis.
Cell viability assay
Cell viability was assessed using a CCK-8 assay kit,
according to the manufacturer's protocol. Briefly, the SH-SY5Y
cells were plated into 96-well plates at a density of
3×104 cells per well and treated as described earlier.
At the end of the experiment, CCK-8 regent (10 µl) was added
to each well and co-incubated with the cells for 3 h at 37°C. The
absorbance was measured at a wavelength of 450 nm using a
microplate reader (Bio-Rad Laboratories, Inc.). The assays were
independently performed in triplicate, and cell viability was
expressed as a percentage of the control.
Lactate dehydrogenase (LDH) activity
assay
Cytotoxicity was evaluated by determining the LDH
activity in the culture supernatant using the Cytotoxicity
Detection kit, according to the manufacturer's protocol. Following
the indicated treatments, the extracellular medium was collected
and centrifuged at 400 × g at room temperature for 5 min. The
supernatant (20 µl) was then mixed with
2,4-dinitrophenylhydrazine (20 µl) at 37°C for 15 min, and
NaOH (0.4 M, 250 µl) was then added to the mixture and
co-incubated for a further 15 min at 37°C. The LDH concentration
was quantified by measuring the absorbance at 490 nm. The protein
concentration from the cell lysates was determined using the BCA
protein assay kit, and the cytotoxicity was normalized to the
protein concentration.
RNA isolation and RT-quantitative
polymerase chain reaction (RT-qPCR) assay
Total RNA was isolated from the cells using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific) in accordance with
the manufacturer's protocol. The RNA quality and purity were
determined using the NanoDrop-2000 spectrophotometer at an
A260/A280 ratio. Next, total RNA (2 µg) was reverse
transcribed into cDNA using an RT kit. The qPCR reactions were
performed with the SYBR® RT-PCR kit on the ABI 7500
Real-Time PCR System, according to the manufacturer's protocol. The
following qPCR conditions were set: Reverse transcription (1 cycle)
at 61°C for 20 sec, denaturation (1 cycle) at 95°C for 1 min, and
annealing (45 cycles) at 95̊C for 5 sec, followed by extension at
72°C for 20 sec. The amount of amplified GAPDH was used as an
external reference gene. The mRNA levels were calculated by the
2-ΔΔCq method (26)
and are presented as fold changes relative to the expression levels
of internal control (GAPDH). All reactions were performed in
triplicate. The primers used in qPCR were synthesized by Sigma
Genosys, and their sequences were as follows: APE1/Ref-1 forward,
5′-CTG CCT GGA CTC TCT CAT CAA TAC-3′, and reverse, 5′-GAA TGC CGT
ATC CGC TAC TCC-3′; GAPDH forward, 5-ACG GCA AGT TCA ACG GCA C-3′,
and reverse, 5′-CGC CAG TAG ACT CCA CGA C A TA-3′.
Flow cytometric detection of
apoptosis
The apoptosis rate was determined using an Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) apoptosis
detection kit (BD Biosciences) according to the manufacturer's
protocol, in combination with a flow cytometer (Beckman Coulter,
Inc.). Briefly, SH-SY5Y cells were seeded in 6-well plates at a
density of 1×106 cells/well. Subsequent to the different
treatments discussed earlier, cells were washed twice with cold PBS
and re-suspended with 1X binding buffer. Next, the cells were
stained with Annexin V-FITC (5 µl) and PI (10 µl),
and co-incubated for 20 min at room temperature in the dark.
Finally, the flow cytometer was used to detect the apoptotic
cells.
8-Hydroxy-2′-deoxyguanosine (8-OHdG)
level determination
The 8-OHdG level was determined using an ELISA kit
from Oxis Health Products, according to the manufacturer's
protocol. Briefly, genomic DNA was isolated from the SH-SY5Y cells
using the Genomic DNA Isolation kit (RayBiotech). Standards
included in the kit and DNA samples were incubated with
biotinylated 8-OHdG in microtitration wells coated with other
8-OHdG molecules with defined and unique epitope specificity.
Subsequent to washing, streptavidin labeled with enzyme horseradish
peroxidase (HRP) was added to each well, followed by the addition
of a TMB substrate solution, and color development in proportion to
the concentration of 8-OHdG. Finally, stop solution was added, and
the color changed from blue to yellow. The intensity was measured
at 450 nm using a microplate reader. Sample DNA assays were
performed in duplicate.
DNA damage assays
The abasic sites, also known as
apurinic/apyrimidinic (AP) sites, in the DNA extracts from the
SH-SY5Y cells were determined with a biotin-labeled aldehyde
reactive probe in a colorimetric assay (Dojindo Molecular
Technologies, Inc.) according to the manufacturer's protocol.
Caspase-3 activity assessment
The activity of caspase-3 was measured with a
caspase-3 activity assay kit (Beyotime Institute of Biotechnology),
following the protocol provided by the manufacturer.
Acetyl-Asp-Glu-Val-Asp p-nitroanilide (Ac-DEVD-рNA) is catalyzed by
caspase-3 to produce a yellow p-nitroaniline (pNA) compound. For
caspase-3 activity assessment, the SH-SY5Y cells were collected,
washed twice with PBS and then lysed in lysis buffer on ice for 15
min. Following centrifugation at 16,000 × g for 15 min at 4°C, the
supernatant was incubated with Ac-DEVD-рNA at 37°C for 2 h. The
absorbance at a wavelength of 405 nm was then measured. The
activity of caspase-3 is expressed as the relative percentage of
the optical density in the control group.
Measurement of intracellular reactive
oxygen species
Dichlorodihydrofluorescein diacetate (DCFH-DA;
Invitrogen; Thermo Fisher Scientific, Inc.), a membrane-permeable
non-fluorescent dye, was used in the present study to detect the
endogenous ROS generation in SH-SY5Y neuronal cells according to
the protocol provided by the manufacturer. DCFH-DA can be converted
by intracellular esterases into 2′,7′-dichlorodihydrofluorescein,
which is then oxidized by ROS into fluorescent
2′,7′-dichlorofluorescein. In brief, following treatment, the
SH-SY5Y cells were washed twice with ice-cold PBS and then
co-incubated in DMEM solution containing 10 µM DCFH-DA at
37°C for 20 min. Subsequent to washing twice with PBS, the
fluorescence was observed using a fluorescence microplate reader
(H1m; BioTek Instruments, Inc.) at an excitation wavelength of 488
nm and an emission wavelength of 528 nm. In addition, the cells
were collected, centrifuged at 800 × g for 5 min at room
temperature and washed twice with PBS, and then the fluorescence
intensity was be quantitatively analyzed using a flow cytometer (BD
Biosciences).
Superoxide dismutase (SOD) activity
determination
At the end of the treatment period, the SH-SY5Y
cells were harvested, washed with ice-cold PBS, and then incubated
for 30 min with lysis buffer containing 50 mM Tris-HCl (pH 7.4), 1%
Triton X-100, 150 mM NaCl, 1 mM EDTA and 0.2% sodium dodecyl
sulfate (SDS). The cell lysates were centrifuged at 10,000 × g at
4°C for 10 min, and the protein concentration was detected using a
BCA kit. SOD activity in the cell lysates was finally evaluated
using a SOD determination kit (Assay Designs) as per the
manufacturer's protocol, and is expressed as U/mg of protein.
Glutathione (GSH) content
determination
The total GSH content in the cellular lysates was
determined using a glutathione measurement kit (Assay Designs; Enzo
Life Sciences, Inc.). Following treatment, cell lysates were
centrifuged at 10,000 × g for 15 min at 4°C, and the supernatant
obtained was used for measuring the GSH content. The protein
concentration was detected using a BCA kit. The absorbance of the
samples was determined using a microplate reader (Infinite 200;
Tecan) at 405 nm. The amount of GSH was detected by means of a
calibration curve and normalized to the protein concentration. The
results are presented as a percentage of the control cells.
Western blot analysis
Following the indicated treatments, the SH-SY5Y
cells were collected and lysed in RIPA buffer containing 1%
phenylmethylsulfonyl fluoride. The lysates were centrifuged at
10,000 × g at 4̊C for 30 min, and the concentration of the protein
samples was detected using a BCA protein quantification kit.
Subsequently, equal amounts of protein (30 µg) were
separated by SDS-PAGE (12% gel) and transferred onto polyvinylidene
difluoride membranes (Millipore). Subsequently, the membranes were
blocked with 5% non-fat milk at room temperature for 2 h, and then
incubated at 4°C overnight with primary antibodies, including
anti-GAPDH (1:1,000), anti-APE1 (1:1,000), anti-Bax (1:1,000),
anti-Bcl-2 (1:1,000), anti-NOX2 (1:1,000), anti-p-AKT (1:1,000),
anti-AKT (1:1,000), anti-p-PI3K (1:1,000) and anti-PI3K (1:1,000).
Following washing three times for 10 min with Tris-buffered saline
with Tween-20, the membranes were incubated with a HRP-conjugated
secondary antibody (1:5,000; Cell Signaling Technology; cat. no.
7076) at room temperature for 2 h. The blots were then developed
with an BeyoECL Star according to the manufacturer's protocol, and
visualized using Image Lab™ Software (Bio-Rad Laboratories,
Inc.).
Statistical analysis
The results are presented as the mean ± standard
deviation values. The statistical significance of differences
between the mean values was assessed by analysis of variance and
post-hoc Bonferroni/Dunn's tests using SPSS software (version 16.0;
SPSS, Inc.). A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
Curcumin mitigates OGD/R-induced SH-SY5Y
cell injury
First, the protective effects of curcumin on OGD/R
injury in SH-SY5Y cells were investigated in the present study. The
results of CCK-8 assay revealed that treatment with curcumin alone
at various concentrations (1, 5, 10 and 15 µl) exerted no
cytotoxicity (Fig. 1A), while
treatment with curcumin (5, 10 and 15 µM) markedly reversed
the OGD/R-induced the downregulation of cell viability (Fig. 1B). In addition, the results from
LDH activity assay revealed that OGD/R significantly increased the
LDH activity, and this effect was also blocked by curcumin (5, 10
and 15 µM) treatment (Fig.
1C). According to the aforementioned results, curcumin
treatment at 10 µl significantly increased cell viability
and decreased LDH activity; thus, this concentration was selected
for use in subsequent experiments.
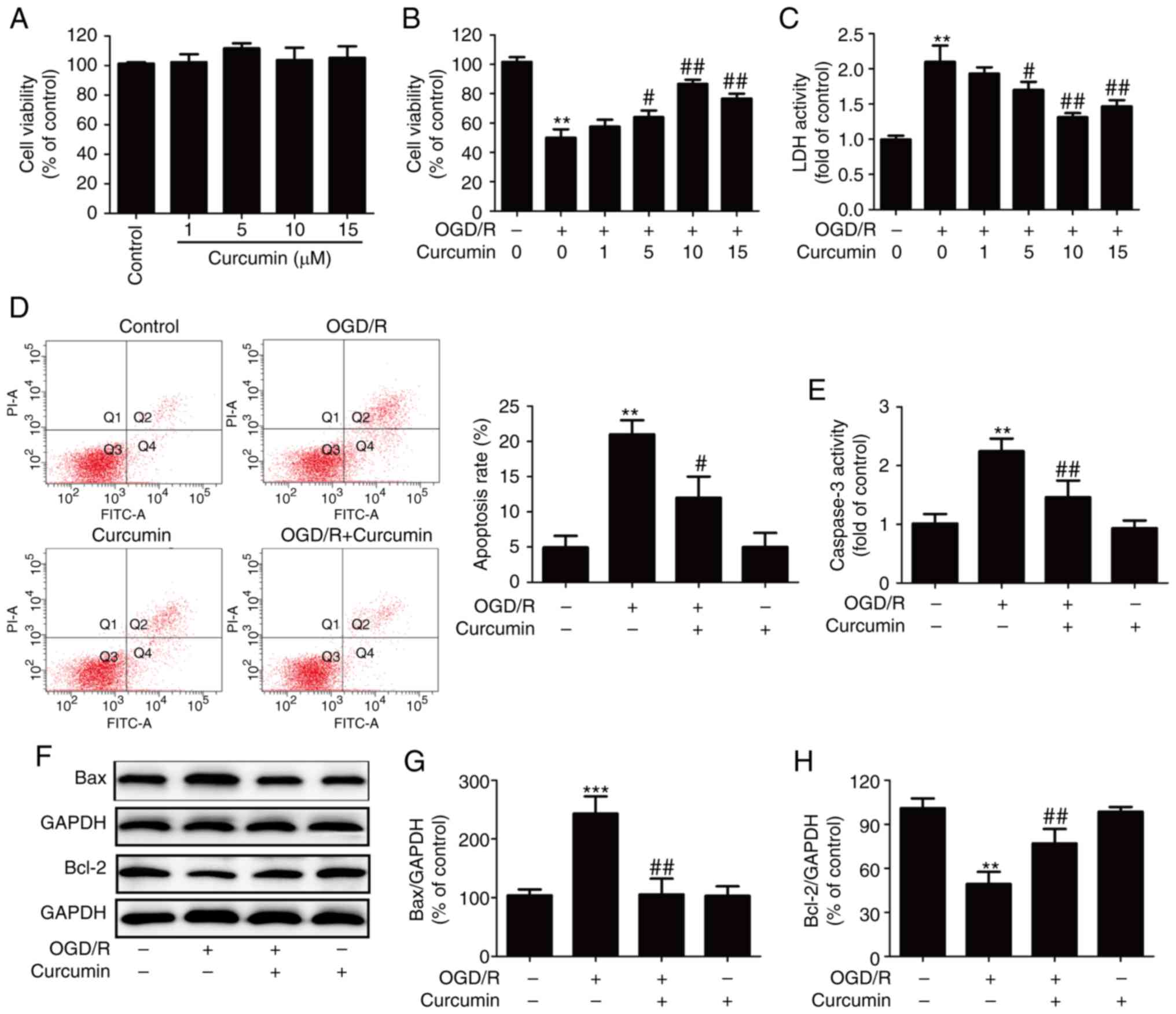 | Figure 1Effects of curcumin on cytotoxicity
and the apoptosis of OGD/R-injured SH-SY5Y cells. (A) SH-SY5Y cells
were treated with various concentrations of curcumin (1, 5, 10 and
15 µl) for 24 h, and the cell viability was measured by
CCK-8 assay. Data were normalized to the control as 100%. (B) Cell
viability examined by CCK-8 assay and (C) LDH activity in the
culture supernatant were tested in SH-SY5Y cells with curcumin (1,
5, 10 and 15 µl) for 24 h immediately after 1 h of OGD
exposure. LDH activity data were normalized to the control, which
was set as 1. Subsequently, SH-SY5Y cells were treated with
curcumin (10 µl) for 24 h immediately after 1 h of exposure
to OGD and then assessed for apoptosis. (D) Apoptosis was measured
by Annexin V-FITC/PI double staining, followed by flow cytometry.
(E) Caspase-3 activity was measured with an assay kit. (F) Protein
expression was detected by western blot analysis. (G) Bax and (H)
Bcl-2 protein levels were quantitatively analyzed. Data are
expressed as the mean ± standard deviation of at least three
independent experiments. **P<0.01 and
***P<0.001, vs. control; #P<0.05 and
##P<0.01, vs. OGD/R exposure alone. OGD/R,
oxygen-glucose deprivation/reperfusion; CCK-8, Cell Counting Kit-8;
LDH, lactate dehydrogenase; Bax, Bcl-2-associated X protein; Bcl-2,
B-cell lymphoma 2. |
The study further investigated the effects of
curcumin on the apoptosis of OGD/R-injured SY-SH5Y cells. Annexin
V-FITC/PI double staining followed by flow cytometry revealed that
curcumin significantly reversed the OGD/R-induced increase in the
apoptotic rate of the SH-SY5Y cells (Fig. 1D). In addition, OGD/R exposure
resulted in marked elevation of caspase-3 activity, which was also
blocked by curcumin (Fig. 1E).
Consistently, the results of western blot analysis (Fig. 1F) demonstrated that OGD/R promoted
the pro-apoptotic protein Bax expression (Fig. 1G) and inhibited the anti-apoptotic
protein Bcl-2 expression (Fig.
1H) in the SY-SH5Y cells; however, treatment with curcumin
reversed these effects. These results thus indicated that curcumin
protected the SH-SY5Y cells against OGD/R-induced injury by
inhibiting apoptosis.
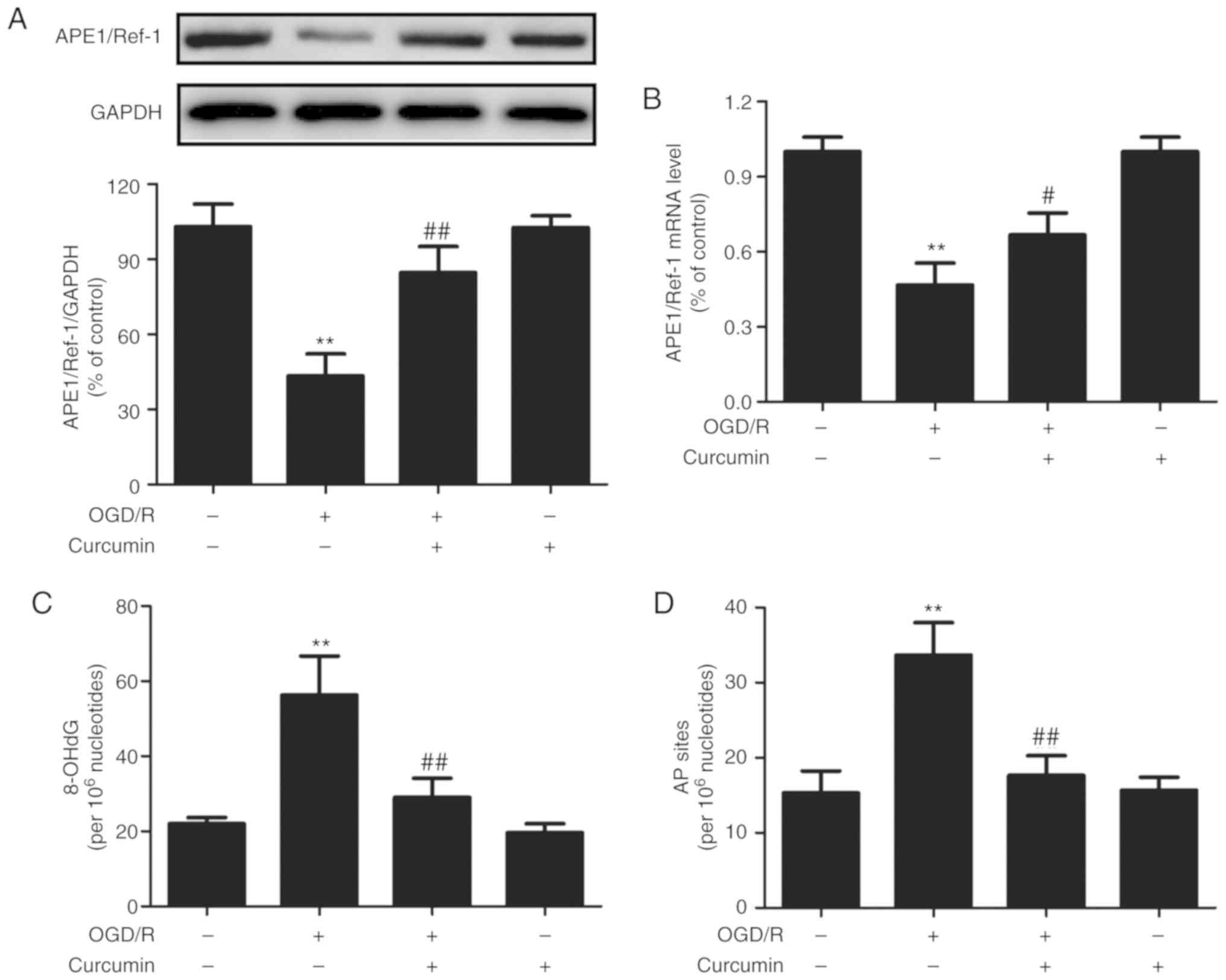
Curcumin increases the APE1 level and
activity in OGD/R-injured SH-SY5Y cells
To determine whether APE1 is involved in the
curcumin-induced neuroprotective effects, the effects of curcumin
on the APE1 level and activity under OGD/R conditions were
examined. As presented in Fig. 2,
the levels of APE1 protein (Fig.
2A) and mRNA (Fig. 2B) in
cells subjected to OGD/R were significantly lower as compared with
those in the control cells. However, post-treatment of the cells
with curcumin (10 µM) immediately after OGD exposure
markedly blocked these effects. Treatment with curcumin alone had
no effect on the APE1 protein and mRNA levels.
APE1 is a multifunctional protein involved in DNA
repair and redox regulation, serving an essential role in repairing
oxidative DNA damage involving single-strand breaks, and formation
of 8-OHdG and AP sites (27,28). The present study further observed
that OGD/R significantly increased the 8-OHdG (Fig. 2C) and AP site (Fig. 2D) levels compared with those in
the control group. However, these effects were abolished by
curcumin post-treatment. Curcumin alone did not affect the 8-OHdG
and AP site levels. Taken together, these results suggested that
curcumin enhanced the level and DNA repair capacity of APE1 under
OGD/R conditions in the SH-SY5Y cells.
APE1 knockdown abrogates the
neuroprotective effects of curcumin against OGD/R-induced SH-SY5Y
cell injury
To further address the role of APE1 in
curcumin-induced neuroprotection, the SH-SY5Y cells were
transfected with chemically synthesized APE1t or APE1s siRNAs. The
results of western blot analysis (Fig. 3A) revealed that APE1 expression in
the APE1t transfection group was significantly reduced to ~35% of
that in the APE1s transfection group (Fig. 3B), indicating that APE1 knockdown
was successfully induced by APE1t transfection. Subsequently, it
was observed that APE1 knockdown further reduced the viability of
the OGD/R-injured SH-SY5Y cells. However, APE1t transfection
reversed the curcumin-induced upregulation of cell viability
(Fig. 3C) and downregulation of
LDH activity (Fig. 3D), as
compared with the effect of APE1s transfection in the curcumin +
OGD/R-treated SH-SY5Y cells. In addition, the effects of APE1 on
cell apoptosis in the presence or absence of curcumin under OGD/R
conditions were further investigated. The results revealed that
APE1t transfection exacerbated the effect of OGD/R on apoptosis,
resulting in the upregulation of caspase-3 activity (Fig. 3E) and Bax protein expression
(Fig. 3F and G), as well as in
the downregulation of Bcl-2 protein expression (Fig. 3F and H), as compared with those in
APE1s-transfected cells. Notably, APE1t transfection attenuated the
curcumin-induced inhibition on these apoptosis-associated indexes.
In the control group, APE1t transfection did not affect the cell
viability, LDH activity, and Bax and Bcl-2 expression levels as
compared with the APE1s transfection group. These results indicated
that APE1 contributed to the neuroprotective effects of curcumin
against cerebral I/R injury in vitro.
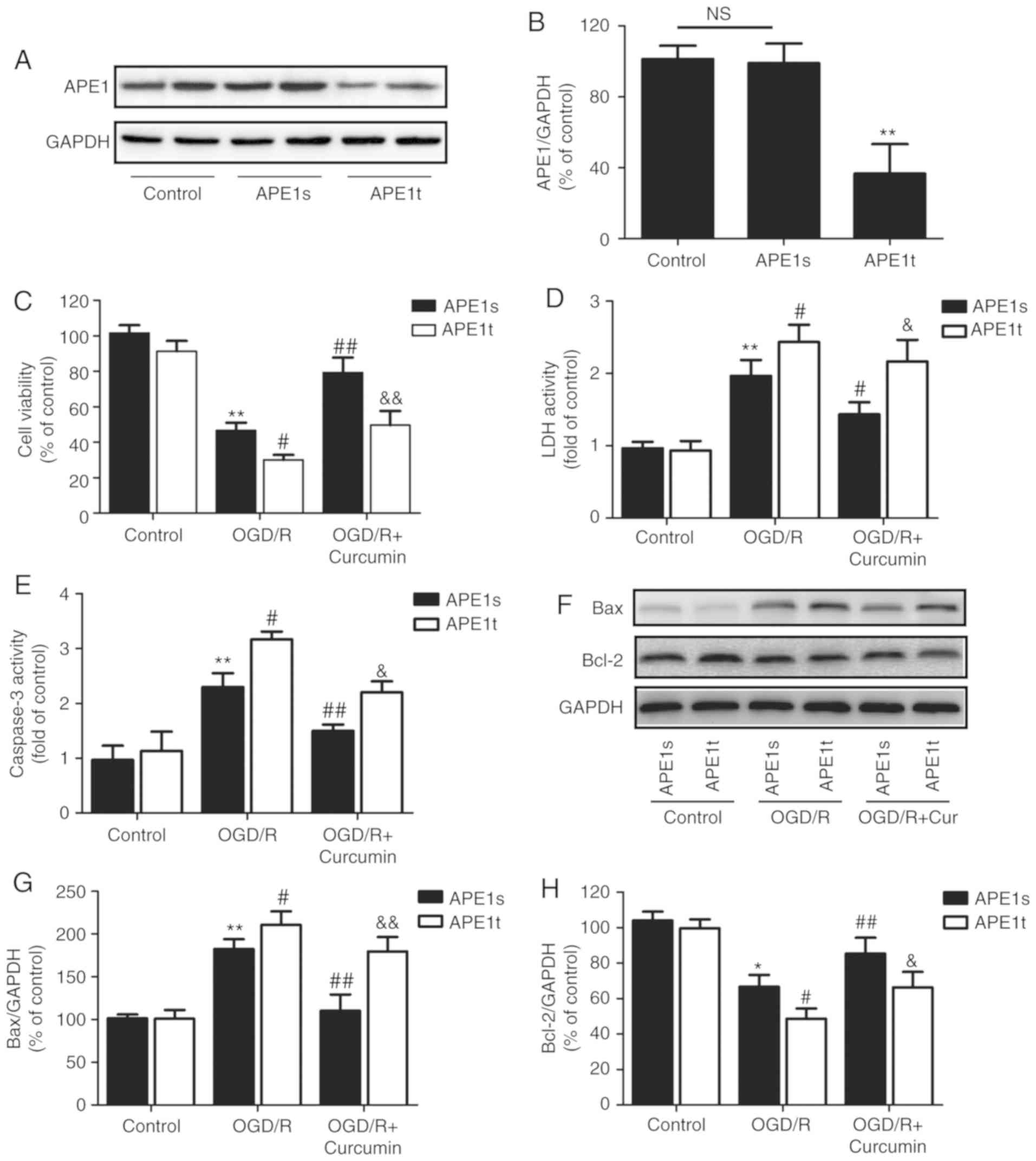 | Figure 3Effects of APE1 knockdown on
cytotoxicity and apoptosis in the presence or absence of curcumin
in SH-SY5Y cells exposed to OGD/R. SH-SY5Y cells were transfected
with APE1t or APE1s, followed by treatment with curcumin (10
µl) for 24 h immediately after 1 h of OGD exposure. (A)
Western blot and (B) quantified protein expression of APE1.
**P<0.01 vs. control. (C) Cell viability was measured
by Cell Counting Kit-8 assay. (D) LDH activity was detected with a
cytotoxicity detection kit. (E) Caspase-3 activity was measured
using an assay kit. (F) The protein expression levels of Bax and
Bcl-2 were determined by western blot analysis. Quantitative
analysis of the (G) Bax and (H) Bcl-2 protein bands. Data are
expressed as the mean ± standard deviation of at least three
independent experiments. *P<0.05 and
**P<0.01, vs. APE1s + control; #P<0.05
and ##P<0.01, vs. APE1s + OGD/R exposure alone;
&P<0.05 and &&P<0.01, vs. APE1s +
OGD/R + curcumin treatment. OGD/R, oxygen-glucose
deprivation/reperfusion; APE1, apurinic/apyrimidinic endonuclease
1; APE1t, siRNA targeting APE1; APE1s, scrambled siRNA; LDH,
lactate dehydrogenase; Bax, Bcl-2-associated X protein; Bcl-2,
B-cell lymphoma 2; NS, no significant difference. |
APE1 knockdown eliminates the inhibitory
effects of curcumin on OGD/R-induced oxidative stress in SH-SY5Y
cells
Next, the effects of curcumin on oxidative stress in
OGD/R-injured SH-SY5Y cells and the role of APE1 in this process
were further examined. As illustrated in Fig. 4, in the APE1s-transfected SH-SY5Y
cells, OGD/R significantly increased ROS production (Fig. 4A and B) and NADPH oxidase 2 (NOX2)
expression (Fig. 4C), which is
one of the most important sources of ROS production in the brain,
compared with the control group; these effects were then reversed
by curcumin post-treatment. Notably, APE1t transfection further
increased ROS generation and NOX2 expression compared with those
observed in the APE1s-transfeced OGD/R-injured SH-SY5Y cells. Upon
curcumin treatment, APE1t transfection blocked the curcumin-induced
decrease in ROS production and NOX2 expression in the OGD/R-injured
SH-SY5Y cells.
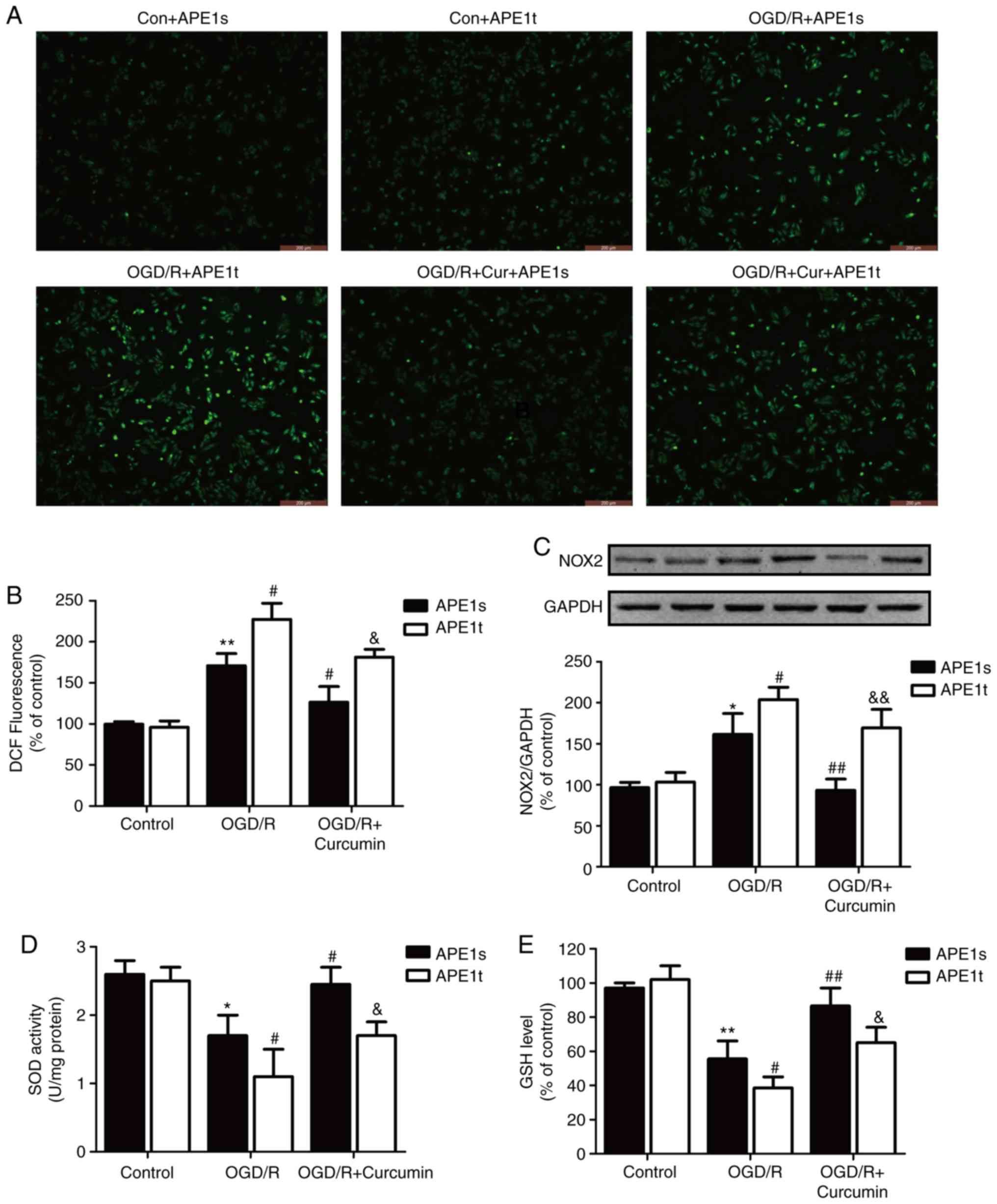 | Figure 4Effects of APE1 knockdown on
oxidative stress in the presence or absence of curcumin in SH-SY5Y
cells exposed to OGD/R. SH-SY5Y cells were transfected with APE1t
or APE1s followed by treatment with curcumin (10 µl) for 24
h immediately after 1 h of OGD exposure. (A) Intracellular ROS
generation determined by the DCFH-DA method (magnification, ×200).
(B) The fluorescence intensity was analyzed by flow cytometry. (C)
NOX2 expression was determined by western blot analysis. (D) SOD
activity and (E) GSH levels were measured using commercial assay
kits. Data are expressed as the mean ± standard deviation of at
least three independent experiments. *P<0.05 and
**P<0.01, vs. APE1s + control; #P<0.05
and ##P<0.01 vs. APE1s + OGD/R exposure alone;
&P<0.05 and &&P<0.01, vs.
APE1s + OGD/R + curcumin treatment. OGD/R, oxygen-glucose
deprivation/reperfusion; APE1, apurinic/apyrimi-dinic endonuclease
1; APE1t, siRNA targeting APE1; APE1s, scrambled siRNA; NOX2, NADPH
oxidase 2; SOD, superoxide dismutase; GSH, glutathione. |
The imbalance of antioxidant endogenous enzymatic
activity is known to result in oxidative stress. Thus, the levels
of endogenous enzymes, including SOD and GSH, were measured, and it
was observed that OGD/R led to a decrease in SOD activity (Fig. 4D) and in GSH levels (Fig. 4E). The OGD/R-induced decrease in
SOD and GSH levels was exacerbated by APE1t transfection, while it
was attenuated by curcumin treatment; however, the effect of
curcumin was eliminated by APE1t transfection. APE1t transfection
alone did not affect these oxidative stress and antioxidant stress
indexes compared with the APE1s transfection group. Taken together,
these results indicated that APE1 mediated the protective effects
of curcumin against oxidative stress in OGD/R-injured SH-SY5Y
cells.
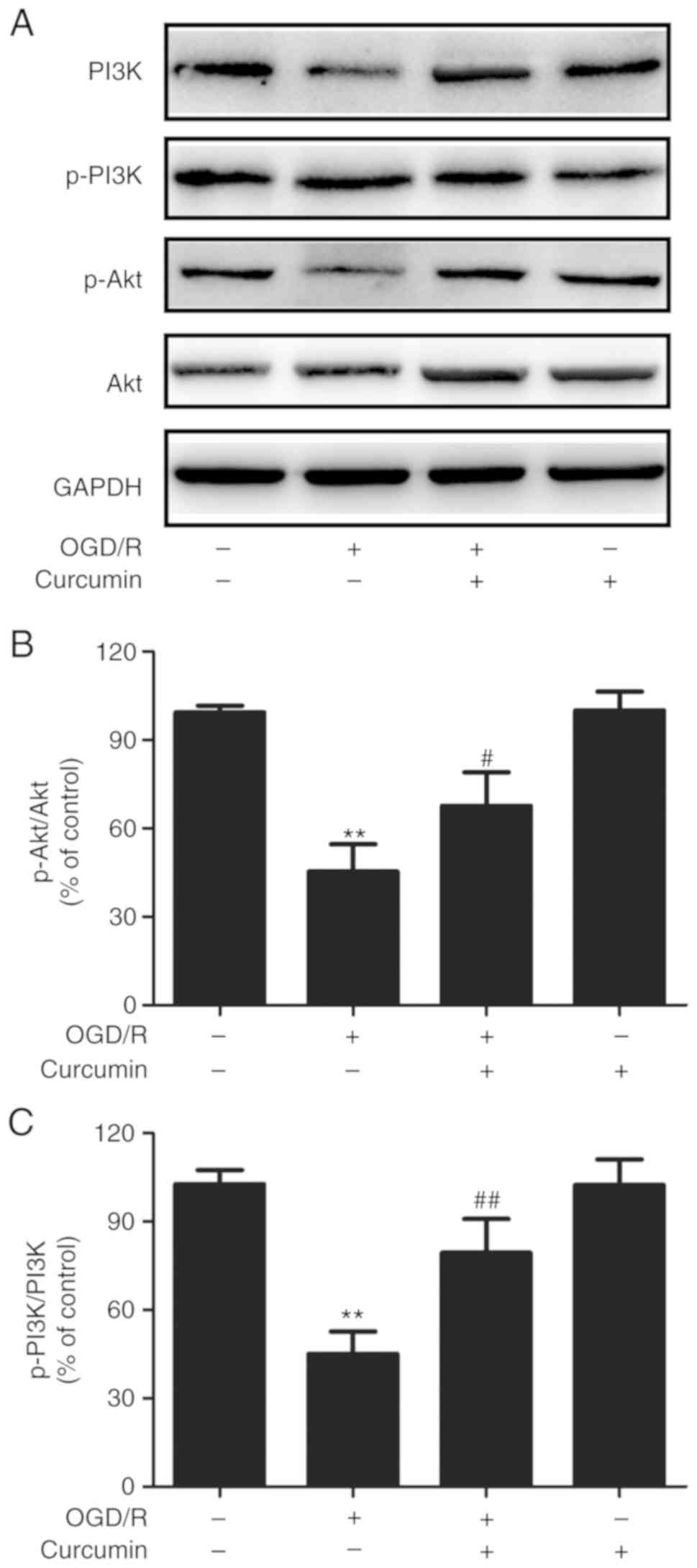
Curcumin activates the PI3K/AKT signaling
pathway in OGD/R-injured SH-SY5Y cells
The PI3K/AKT pathway is known to be involved in
neuronal death associated with cerebral ischemia (29). To further evaluate the mechanisms
underlying the neuroprotective effects of curcumin under OGD/R
conditions, the effects of curcumin on the PI3K/AKT signaling
pathway were then examined. It was observed that the ratios of
p-PI3K/PI3K (Fig. 5A) and
p-AKT/AKT (Fig. 5B) were
significantly decreased in SH-SY5Y cells exposed to OGD/R, as
compared with those in cells exposed to normal conditions. However,
these effects were reversed by curcumin post-treatment. Curcumin
treatment alone had no effect on the expression levels of p-PI3K
and p-AKT proteins. These results suggested that curcumin promotes
the activation of the PI3K/AKT pathway in OGD/R-injured SH-SY5Y
cells.
Inhibition of the PI3K/AKT signaling
pathway attenuates the curcumin-induced protective effects against
OGD/R injury in SH-SY5Y cells
In order to further determine the roles of the
PI3K/AKT pathway in the neuroprotective effects of curcumin, the
cells were pre-treated with LY294002, an inhibitor of the PI3K/AKT
pathway. The results of CCK-8 assay revealed that LY294002
abolished the curcumin-induced increase in cell viability (Fig. 6A) and the decrease in LDH release
(Fig. 6B) in the OGD/R-injured
SH-SY5Y cells. In addition, it was observed that LY294002 reversed
the curcumin-induced inhibition of apoptosis, as evidenced by the
increase in caspase-3 activity (Fig.
6C) and Bax expression (Fig.
6D), as well as by the decrease in Bcl-2 expression (Fig. 6E), compared with the curcumin +
OGD/R co-treatment group. These results indicated that the PI3K/AKT
pathway contributes to the protective effects of curcumin against
OGD/R-induced injury in SH-SY5Y cells. Notably, the data revealed
that LY294002 also blocked the curcumin-induced upregulation of
APE1 in the OGD/R-injured SH-SY5Y cells (Fig. 6F). Taken together, the
aforementioned results suggested that curcumin activates the
PI3K/AKT pathway and further increases APE1 expression, resulting
in neuroprotective effects against OGD/R-induced SH-SY5Y cell
injury.
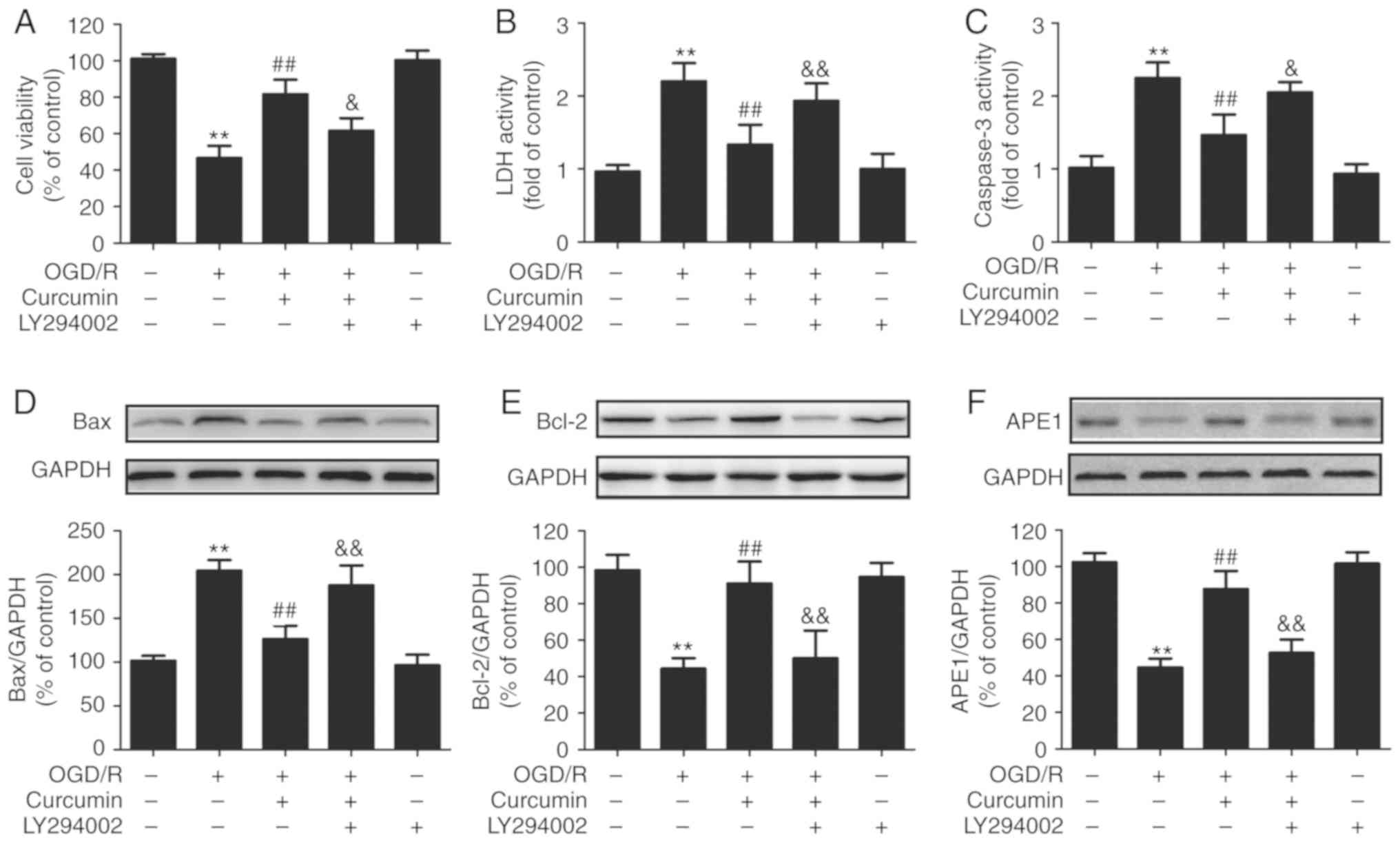 | Figure 6Effects of LY294002 on cytotoxicity,
apoptosis and APE1 expression in OGD/R-injured SH-SY5Y cells. Cells
were pre-treated with a PI3K/Akt inhibitor (LY294002; 10 µM)
for 2 h following incubation with curcumin (10 µl) for 24 h
immediately after OGD exposure for 1 h. (A) Cell viability was
measured by Cell Counting Kit-8 assay. (B) LDH activity in the
culture supernatant was detected using a Cytotoxicity Detection
kit. (C) Caspase-3 activity was determined by a caspase-3 activity
assay kit. (D) Bax, (E) Bcl-2 and (F) APE1 protein expression
levels were detected by western blot analysis. Data are expressed
as the mean ± standard deviation of at least three independent
experiments. **P<0.01 vs. control;
##P<0.01 vs. OGD/R exposure alone;
&P<0.05 and &&P<0.01, vs.
curcumin + OGD/R co-treatment. OGD/R, oxygen-glucose
deprivation/reperfusion; APE1, apurinic/apyrimidinic endonuclease
1; PI3k, phosphatidylinositol 3-kinase; AKT, protein kinase B; LDH,
lactate dehydrogenase; Bax, Bcl-2-associated X protein; Bcl-2,
B-cell lymphoma 2. |
Discussion
The present study revealed that curcumin protected
the SH-SY5Y neuronal cells against apoptosis and oxidative stress
induced by OGD/R injury by increasing the APE1 levels. In addition,
the results demonstrated that curcumin activated the PI3K/AKT
signaling pathway, exerting protective effects against
OGD/R-induced SH-SY5Y cell injury. Notably, the PI3K/AKT pathway
mediated the curcumin-induced upregulation of APE1. To the best of
our knowledge, these results confirm for the first time that
curcumin exerts neuroprotective effects by enhancing the expression
and activity of APE1, which is involved in the activation of the
PI3K/AKT pathway following OGD/R injury in SH-SY5Y neuronal
cells.
Owing to the high mortality and severe neurological
disorder of cerebral I/R injury, the development of effective
therapeutic drugs for the prevention of ischemic injury is of
utmost importance (30,31). Curcumin, a naturally occurring
polyphenolic compound isolated from the root of the Curcuma
longa Linn., can pass through the blood-brain barrier, and has
been recommended for the prevention and treatment of
cerebrovascular disease due to its anti-apoptotic, antioxidant and
anti-inflammatory effects, its limited toxicity and minimal
side-effects (32-34). The results of the present study
revealed that curcumin markedly increased the viability and
decreased LDH activity in OGD/R-injured SH-SY5Y cells, which is in
accordance with the findings of studies by Xie et al
(35) and Zhang et al
(36). These results suggested
that curcumin exerts protective effects against OGD/R-induced
injury in SH-SY5Y cells. While a number of studies have attempted
to elucidate the possible mechanisms underlying the neuroprotective
effects of curcumin, the complete mechanisms through which curcumin
protects cells against cerebral I/R injury have not yet been
elucidated.
A growing number of studies have suggested that
oxidative stress refers to the elevated production of intracellular
ROS, which may lead to damage in tissue, lipids, proteins and DNA,
involved in the pathophysiological processes of cerebral ischemia
(37,38). It is well known that the
antioxidant activity of curcumin is critical for its
neuroprotective effects (39).
APE1 is a master regulator of the cellular response to oxidative
stress, and is involved in gene transcriptional regulation during
the adaptive cellular response to oxidative stress, as well as in
the base excision repair pathway of oxidative DNA lesions, which
consist of DNA-protein crosslinks, AP sites, 8-OHdG formation and
single-strand breaks (40,41).
Several studies have examined APE1 in the context of cerebral I/R
injury, and APE1 expression has long been known to decrease
following ischemic injury (19).
In line with these findings, the present study observed that the
APE1 protein and mRNA levels were decreased by OGD/R. In addition,
OGD/R increased 8-OHdG and AP site formation, which was in
accordance with the findings of a study by Kim et al
(21). However, these effects
were all blocked by curcumin, and the results indicated that
curcumin increased APE1 expression and activity in cerebral I/R
injury. However, an earlier study revealed that curcumin is an
inhibitor of the APE1 redox function that affects numerous genes
and pathways, which is contrary to the results of the current study
(42). These differences may be a
result of the different research systems used: The previous study
examined a virus system, while the present study investigated
SH-SY5Y cells. Consistent with our present research, a number of
other studies have also confirmed that curcumin exerts biological
activities via activation of APE1 (43,44). Subsequently, it was revealed that
the knockdown of APE1 by transfection with APE1 siRNA reversed the
curcumin-induced protective effects against OGD/R injury in SH-SY5Y
cells, suggesting the contribution of APE1 to the neuroprotective
effects of curcumin. Taken together, these results suggest that
APE1 upregulation contributes to the neuroprotective effects of
curcumin.
Apoptosis and oxidative stress are prominent
features of cerebral ischemia (38,45). Zhao et al (46) have proven that curcumin attenuated
focal cerebral ischemic injury via anti-oxidative stress and
anti-apoptotic mechanisms in rats. The results of a study by Wang
et al (47) also revealed
that the neuroprotective effects of curcumin against I/R-induced
neuronal damage were attributed to its antioxidant capacity in
decreasing oxidative stress and the associated signaling cascade,
leading to apoptotic cell death. Notably, APE1 has been studied for
its extended repertoire in controlling the cellular response to
apoptosis and oxidative stress (48). APE1 upregulation, either induced
endogenously or through transgene overexpression, reduces oxidative
DNA damage and prevents hippocampal neuronal apoptosis due to
ischemic injury (19).
Furthermore, APE1 is required for PACAP-mediated neuroprotection
(23). Consistent with these
findings, in the present study, it was observed that APE1 knockdown
induced by APE1 siRNA transfection further aggravated the
OGD/R-induced apoptosis and oxidative stress, while curcumin
mitigated the effects of OGD/R. Notably, these protective effects
of curcumin were all abolished by APE1 knockdown induced by APE1
siRNA. Overall, these results revealed that APE1 mediates the
protective effects of curcumin against apoptosis and oxidative
stress in cerebral I/R injury. APE1 has two major functions in
mammalian cells: DNA repair (49)
and the redox regulation of gene transcription (50). However, the present study did not
determine whether the redox activity of APE1 was less instrumental
in the neuroprotective effects of curcumin than its DNA repair
activity. These aspects remain to be investigated in the future
using APE1 functional mutant (51,52).
The PI3K/AKT pathway has been documented as an
essential pathway for cell survival in response to apoptosis and
oxidative stress induced by a series of physiological and
pathological or exogenous stimuli (53,54), while this pathway is also
significantly involved in cerebral ischemia (55). To date, a number of studies have
confirmed that the PI3K/AKT pathway serves an important role in the
neuroprotective effects of curcumin by inhibiting apoptosis,
oxidative stress and inflammation (56-58). However, there are few reports
available to date on the roles of the PI3K/AKT pathway in the
neuroprotective effects of curcumin against cerebral I/R injury. In
the present study, it was further revealed that curcumin increased
the ratio of p-PI3K/PI3K and p-AKT/AKT proteins, whereas
pharmacological treatment with LY294002, a PI3K inhibitor, reversed
the curcumin-induced cytoprotective and anti-apoptotic effects on
OGD/R-injured SY-SH5Y cells. It should be noted that LY294002 was
also found to block the curcumin-induced upregulation of APE1
expression under OGD/R conditions. These results suggested that the
PI3K/AKT pathway contributes to the neuroprotective effects of
curcumin by regulating APE1 in cerebral I/R injury.
However, the present study also has certain
limitations. Firstly, only one cell line was used to demonstrate
the potential mechanism of neuroprotection of curcumin, and the use
of other cell lines or cultured neurons would provide further
convincing results. Secondly, no data from an in vivo animal
model were provided, which is important to verify the validity of
curcumin concentration in vitro in the present study. These
two issues will be addressed in our future experiments. Finally,
the present study lacked the results from caspase-3 expression and
cleaved caspase-3 protein expression, which should be noted in
future research.
In conclusion, the results of the present study
demonstrated that curcumin confers neuroprotective effects against
apoptosis and oxidative stress in cerebral I/R injury by increasing
the APE1 level and activity, which may be mediated via the PI3K/AKT
signaling pathway. To the best of our knowledge, the present study
is the first to suggest that curcumin may be a potential
therapeutic agent against cerebral I/R injury. The present study
also identified APE1 as a major therapeutic target of the
neuroprotective effects of curcumin.
Funding
The present study was supported by the Natural
Science Foundation of Zhejiang Province (grant. no.
LY16H090003).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
LW participated in performing the experiments and
writing of the paper. CJ and YK contributed to data analysis. YD
and WF performed the experiments. PH designed the study and revised
the paper. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Hausenloy DJ and Yellon DM: Ischaemic
conditioning and reperfusion injury. Nat Rev Cardiol. 13:193–209.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee RHC, Lee MHH, Wu CYC, Couto E, Silva
A, Possoit HE, Hsieh TH, Minagar A and Lin HW: Cerebral ischemia
and neuroregeneration. Neural Regen Res. 13:373–385. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Millar LJ, Shi L, Hoerder-Suabedissen A
and Molnar Z: Neonatal hypoxia ischaemia: Mechanisms, models, and
therapeutic challenges. Front Cell Neurosci. 11:782017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sun MS, Jin H, Sun X, Huang S, Zhang FL,
Guo ZN and Yang Y: Free radical damage in ischemia-reperfusion
injury: An obstacle in acute ischemic stroke after
revascularization therapy. Oxid Med Cell Longev. 2018:38049792018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thornton C, Leaw B, Mallard C, Nair S,
Jinnai M and Hagberg H: Cell death in the developing brain after
hypoxia-ischemia. Front Cell Neurosci. 11:2482017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Leiva-Salinas C, Patrie JT, Xin W, Michel
P, Jovin T and Wintermark M: Prediction of early arterial
recanalization and tissue fate in the selection of patients with
the greatest potential to benefit from intravenous tissue-type
plasminogen activator. Stroke. 47:397–403. 2016. View Article : Google Scholar
|
|
7
|
Rother J, Ford GA and Thijs VN:
Thrombolytics in acute ischaemic stroke: Historical perspective and
future opportunities. Cerebrovasc Dis. 35:313–319. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cheng CY and Lee YC: Anti-inflammatory
effects of traditional chinese medicines against ischemic injury in
in vivo models of cerebral ischemia. Evid Based Complement Alternat
Med. 2016:57394342016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Feigin VL: Herbal medicine in stroke: Does
it have a future? Stroke. 38:1734–1736. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ip FC, Zhao YM, Chan KW, Cheng EY, Tong
EP, Chandrashekar O, Fu GM, Zhao ZZ and Ip NY: Neuroprotective
effect of a novel Chinese herbal decoction on cultured neurons and
cerebral ischemic rats. BMC Complement Altern Med. 16:4372016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lestari ML and Indrayanto G: Curcumin.
Profiles Drug Subst Excip Relat Methodol. 39:113–204. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Maiti P and Dunbar GL: Use of curcumin, a
natural polyphenol for targeting molecular pathways in treating
age-related neurodegenerative diseases. Int J Mol Sci.
19:E16372018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rahmani AH, Alsahli MA, Aly SM, Khan MA
and Aldebasi YH: Role of curcumin in disease prevention and
treatment. Adv Biomed Res. 7:382018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee WH, Loo CY, Bebawy M, Luk F, Mason RS
and Rohanizadeh R: Curcumin and its derivatives: Their application
in neuropharmacology and neuroscience in the 21st century. Curr
Neuropharmacol. 11:338–378. 2013. View Article : Google Scholar :
|
|
15
|
Wicha P, Tocharus J, Janyou A, Jittiwat J,
Changtam C, Suksamrarn A and Tocharus C: Hexahydrocurcumin protects
against cerebral ischemia/reperfusion injury, attenuates
inflammation, and improves antioxidant defenses in a rat stroke
model. PLoS One. 12:e01892112017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu JX, Zhang LY, Chen YL, Yu SS, Zhao Y
and Zhao J: Curcumin pretreatment and post-treatment both improve
the antioxidative ability of neurons with oxygen-glucose
deprivation. Neural Regen Res. 10:481–489. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xanthoudakis S and Curran T:
Identification and characterization of Ref-1, a nuclear protein
that facilitates AP-1 DNA-binding activity. EMBO J. 11:653–665.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lan J, Li W, Zhang F, Sun FY, Nagayama T,
O'Horo C and Chen J: Inducible repair of oxidative DNA lesions in
the rat brain after transient focal ischemia and reperfusion. J
Cereb Blood Flow Metab. 23:1324–1339. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Leak RK, Li P, Zhang F, Sulaiman HH, Weng
Z, Wang G, Stetler RA, Shi Y, Cao G, Gao Y and Chen J:
Apurinic/apyrimidinic endonuclease 1 upregulation reduces oxidative
DNA damage and protects hippocampal neurons from ischemic injury.
Antioxid Redox Signal. 22:135–148. 2015. View Article : Google Scholar :
|
|
20
|
Kim HW, Cho KJ, Lee BI, Kim HJ and Kim GW:
Post-ischemic administration of peptide with apurinic/apyrimidinic
endonu-clease activity inhibits induction of cell death after focal
cerebral ischemia/reperfusion in mice. Neurosci Lett. 460:166–169.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim HW, Cho KJ, Park SC, Kim HJ and Kim
GW: The adenoviral vector-mediated increase in
apurinic/apyrimidinic endonuclease inhibits the induction of
neuronal cell death after transient ischemic stroke in mice. Brain
Res. 1274:1–10. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jia JY, Tan ZG, Liu M and Jiang YG:
Apurinic/apyrimidinic endonuclease 1 (APE1) contributes to
resveratrol-induced neuroprotection against oxygen-glucose
deprivation and re-oxygenation injury in HT22 cells: Involvement in
reducing oxidative DNA damage. Mol Med Rep. 16:9786–9794. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Stetler RA, Gao Y, Zukin RS, Vosler PS,
Zhang L, Zhang F, Cao G, Bennett MV and Chen J:
Apurinic/apyrimidinic endo-nuclease APE1 is required for
PACAP-induced neuroprotection against global cerebral ischemia.
Proc Natl Acad Sci USA. 107:3204–3209. 2010. View Article : Google Scholar
|
|
24
|
Liu Y, Eaton ED, Wills TE, McCann SK,
Antonic A and Howells DW: Human ischaemic cascade studies using
SH-SY5Y cells: A systematic review and meta-analysis. Transl Stroke
Res. 9:564–574. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu Q, Jin Z, Xu Z, Yang H, Li L, Li G, Li
F, Gu S, Zong S, Zhou J, et al: Antioxidant effects of ginkgolides
and bilobalide against cerebral ischemia injury by activating the
Akt/Nrf2 pathway in vitro and in vivo. Cell Stress Chaperones.
24:441–452. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
Relative Gene Expression Data Using Real-Time Quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
27
|
Li M and Wilson DM III: Human
apurinic/apyrimidinic endonuclease 1. Antioxid Redox Signal.
20:678–707. 2014. View Article : Google Scholar
|
|
28
|
Thakur S, Sarkar B, Cholia RP, Gautam N,
Dhiman M and Mantha AK: APE1/Ref-1 as an emerging therapeutic
target for various human diseases: Phytochemical modulation of its
functions. Exp Mol Med. 46:e1062014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shioda N, Han F and Fukunaga K: Role of
Akt and ERK signaling in the neurogenesis following brain ischemia.
Int Rev Neurobiol. 85:375–387. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nagy Z and Nardai S: Cerebral
ischemia/repefusion injury: From bench space to bedside. Brain Res
Bull. 134:30–37. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou G, Li MH, Tudor G, Lu HT, Kadirvel R
and Kallmes D: Remote ischemic conditioning in cerebral diseases
and neuro-interventional procedures: Recent researchprogress. Front
Neurol. 9:3392018. View Article : Google Scholar
|
|
32
|
Kalani A, Kamat PK, Kalani K and Tyagi N:
Epigenetic impact of curcumin on stroke prevention. Metab Brain
Dis. 30:427–435. 2015. View Article : Google Scholar
|
|
33
|
Kaur C and Ling EA: Blood brain barrier in
hypoxic-ischemic conditions. Curr Neurovasc Res. 5:71–81. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Marchiani A, Rozzo C, Fadda A, Delogu G
and Ruzza P: Curcumin and curcumin-like molecules: From spice to
drugs. Curr Med Chem. 21:204–222. 2014. View Article : Google Scholar
|
|
35
|
Xie CJ, Gu AP, Cai J, Wu Y and Chen RC:
Curcumin protects neural cells against ischemic injury in N2a cells
and mouse brain with ischemic stroke. Brain Behav. 8:e009212018.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang Y, Fang M, Sun Y, Zhang T, Shi N, Li
J, Jin L, Liu K and Fu J: Curcumin attenuates cerebral ischemia
injury in Sprague-Dawley rats and PC12 cells by suppressing
overactivated autophagy. J Photochem Photobiol B. 184:1–6. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li P, Stetler RA, Leak RK, Shi Y, Li Y, Yu
W, Bennett MVL and Chen J: Oxidative stress and DNA damage after
cerebral ischemia: Potential therapeutic targets to repair the
genome and improve stroke recovery. Neuropharmacology. 134:208–217.
2018. View Article : Google Scholar
|
|
38
|
Zhao M, Zhu P, Fujino M, Zhuang J, Guo H,
Sheikh I, Zhao L and Li XK: Oxidative stress in hypoxic-ischemic
encephalopathy: Molecular mechanisms and therapeutic strategies.
Int J Mol Sci. 17:E20782016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bavarsad K, Barreto GE, Hadjzadeh MA and
Sahebkar A: Protective effects of curcumin against
ischemia-reperfusion injury in the nervous system. Mol Neurobiol.
56:1391–1404. 2019. View Article : Google Scholar
|
|
40
|
Chen H, Yoshioka H, Kim GS, Jung JE, Okami
N, Sakata H, Maier CM, Narasimhan P, Goeders CE and Chan PH:
Oxidative stress in ischemic brain damage: Mechanisms of cell death
and potential molecular targets for neuroprotection. Antioxid Redox
Signal. 14:1505–1517. 2011. View Article : Google Scholar
|
|
41
|
Li P, Hu X, Gan Y, Gao Y, Liang W and Chen
J: Mechanistic insight into DNA damage and repair in ischemic
stroke: Exploiting the base excision repair pathway as a model of
neuro-protection. Antioxid Redox Signal. 14:1905–1918. 2011.
View Article : Google Scholar
|
|
42
|
Li H, Zhong C, Wang Q, Chen W and Yuan Y:
Curcumin is an APE1 redox inhibitor and exhibits an antiviral
activity against KSHV replication and pathogenesis. Antiviral Res.
167:98–103. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sarkar B, Dhiman M, Mittal S and Mantha
AK: Curcumin revitalizes Amyloid beta (25-35)-induced and
organophosphate pesticides pestered neurotoxicity in SH-SY5Y and
IMR-32 cells via activation of APE1 and Nrf2. Metab Brain Dis.
32:2045–2061. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zaky A, Mahmoud M, Awad D, El Sabaa BM,
Kandeel KM and Bassiouny AR: Valproic acid potentiates
curcumin-mediated neuroprotection in lipopolysaccharide induced
rats. Front Cell Neurosci. 8:3372014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen SD, Yang DI, Lin TK, Shaw FZ, Liou CW
and Chuang YC: Roles of oxidative stress, apoptosis, PGC-1α and
mitochondrial biogenesis in cerebral ischemia. Int J Mol Sci.
12:7199–7215. 2011. View Article : Google Scholar
|
|
46
|
Zhao J, Yu S, Zheng W, Feng G, Luo G, Wang
L and Zhao Y: Curcumin improves outcomes and attenuates focal
cerebral ischemic injury via antiapoptotic mechanisms in rats.
Neurochem Res. 35:374–379. 2010. View Article : Google Scholar
|
|
47
|
Wang Q, Sun AY, Simonyi A, Jensen MD,
Shelat PB, Rottinghaus GE, MacDonald RS, Miller DK, Lubahn DE,
Weisman GA and Sun GY: Neuroprotective mechanisms of curcumin
against cerebral ischemia-induced neuronal apoptosis and behavioral
deficits. J Neurosci Res. 82:138–148. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lee YR, Kim KM, Jeon BH and Choi S:
Extracellularly secreted APE1/Ref-1 triggers apoptosis in
triple-negative breast cancer cells via RAGE binding, which is
mediated through acetylation. Oncotarget. 6:23383–23398.
2015.PubMed/NCBI
|
|
49
|
Muller UR, Dudler T, Schneider T, Crameri
R, Fischer H, Skrbic D, Maibach R, Blaser K and Suter M: Type I
skin reactivity to native and recombinant phospholipase A2 from
honeybee venom is similar. J Allergy Clin Immunol. 96:395–402.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Tell G, Quadrifoglio F, Tiribelli C and
Kelley MR: The many functions of APE1/Ref-1: Not only a DNA repair
enzyme. Antioxid Redox Signal. 11:601–620. 2009. View Article : Google Scholar
|
|
51
|
Shan JL, He HT, Li MX, Zhu JW, Cheng Y, Hu
N, Wang G, Wang D, Yang XQ, He Y, et al: APE1 promotes antioxidant
capacity by regulating Nrf-2 function through a redox-dependent
mechanism. Free Radic Biol Med. 78:11–22. 2015. View Article : Google Scholar
|
|
52
|
Xie J, Zhang L, Li M, Du J, Zhou L, Yang
S, Zeng L, Li Z, Wang G and Wang D: Functional analysis of the
involvement of apurinic/apyrimidinic endonuclease 1 in the
resistance to melphalan in multiple myeloma. BMC Cancer. 14:112014.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Fan YY, Hu WW, Nan F and Chen Z:
Postconditioning-induced neuroprotection, mechanisms and
applications in cerebral ischemia. Neurochem Int. 107:43–56. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Matsuda S, Nakagawa Y, Tsuji A, Kitagishi
Y, Nakanishi A and Murai T: Implications of PI3K/AKT/PTEN signaling
on superoxide dismutases expression and in the pathogenesis of
Alzheimer's disease. Diseases. 6:E282018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Davis SM and Pennypacker KR: Targeting
antioxidant enzyme expression as a therapeutic strategy for
ischemic stroke. Neurochem Int. 107:23–32. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Sang Q, Sun D, Chen Z and Zhao W: NGF and
PI3K/Akt signaling participate in the ventral motor neuronal
protection of curcumin in sciatic nerve injury rat models. Biomed
Pharmacother. 103:1146–1153. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Srivastava P, Dhuriya YK, Kumar V,
Srivastava A, Gupta R, Shukla RK, Yadav RS, Dwivedi HN, Pant AB and
Khanna VK: PI3K/Akt/GSK3beta induced CREB activation ameliorates
arsenic mediated alterations in NMDA receptors and associated
signaling in rat hippocampus: Neuroprotective role of curcumin.
Neurotoxicology. 67:190–205. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhong W, Qian K, Xiong J, Ma K, Wang A and
Zou Y: Curcumin alleviates lipopolysaccharide induced sepsis and
liver failure by suppression of oxidative stress-related
inflammation via PI3K/AKT and NF-KB related signaling. Biomed
Pharmacother. 83:302–313. 2016. View Article : Google Scholar : PubMed/NCBI
|