Introduction
Hypoxia occurs during numerous common phenomena,
including embryogenesis, exposure to high-altitude areas, physical
exercise and diseases (1,2). Pathological hypoxia can impair
skeletal muscle function, a characteristic that it shares with
chronic hypoxia-related diseases, such as chronic heart failure
(3), chronic obstructive
pulmonary disease (4), type 2
diabetes (5) and obstructive
sleep apnea syndrome (6,7). Hypoxia leads to continuous muscle
contraction, which in turn leads to muscle fatigue and
abnormalities in the morphological structure of muscle cells, which
inhibits myogenic differentiation (8–10).
Moreover, several studies have reported that hypoxia induces the
overproduction of reactive oxygen species (ROS) in skeletal muscle.
ROS are a powerful initiator of cellular oxidative stress and
apoptosis, which in turn contribute to skeletal muscle injury
(11,12). Therefore, it is crucial to
mitigate hypoxic injury in the repair of damaged muscle fibers, in
order to improve muscle function and resistance to fatigue.
CoCl2, the most commonly used hypoxia
mimic, substitutes Fe2+ in prolyl hydroxylases to
inhibit its hydroxylation, resulting in hypoxia-inducible factor-1α
(HIF-1α) stabilization (13).
Studies have shown that CoCl2-mimicked hypoxia inhibits
cell proliferation and differentiation and induces autophagy,
apoptosis and myotube atrophy of C2C12 (14–18).
In general, the cornerstone of treatment for muscle
dysfunction in hypoxia-related diseases is rehabilitation-based
exercises (such as small muscle mass and inspiratory muscle
training, optimized nutrition and electrical stimulation) (19–21). In addition, in vitro and
in vivo results have demonstrated that certain
small-molecule compounds are effective in alleviating
hypoxia-induced skeletal muscle damage (16,22). In previous years, mesenchymal stem
cell (MSC) therapy has become an alternative treatment in the field
of skeletal muscle repair (23–25). MSCs have biological
characteristics such as self-renewal, multidirectional directional
differentiation potential and low immunogenicity, making them
highly attractive in clinical applications for a variety of
diseases. Dental pulp stem cells (DPSCs) are excellent candidates
for MSC therapy. Compared with other MSC tissue sources, such as
bone marrow, adipose tissue and peripheral blood, DPSCs present
some favorable advantages, including their convenient, non-invasive
harvesting, induction of less trauma, and the absence of ethical
concerns (26). However, the
therapeutic potential of MSCs is highly dependent on their
secretome (27,28). The involved mechanism of the MSCs
secretome includes immunomodulation, angiogenesis, anti-apoptosis,
anti-oxidative stress and anti-inflammatory functions (29). In vitro, cells secrete
factors into the supernatant, also referred to as conditioned
medium (CM). Several studies have demonstrated that MSC-CM exerts a
marked protective effect against skeletal muscle dysfunction
(30–32). However, the potential protective
effects of DPSC-CM against hypoxia-induced skeletal muscle injury
and the underlying mechanisms remain unclear. In the present study,
human DPSCs were used and their CM was co-cultured with murine
C2C12 myoblasts rather than human skeletal muscle cells. Mouse
cells are considered as the starting point for investigating the
effects of DPSCs and their CM, which will subsequently be injected
into mice to verify their efficacy in vivo in a future
study, and finally into human subjects. The mouse cells mirror the
biology of human cells well in various aspects (33). Previous studies (34–36) have also demonstrated that human
cells and/or their CM have protective effects in mouse and rat
cells. Therefore, C2C12 cells were used in the present study.
Wnt/β-catenin signaling plays an important role in
satellite cell self-renewal, myoblast proliferation, fusion and
myofiber homeostasis in skeletal muscle (37). Skeletal muscle injury initiates
Wnt signaling, thereby activating satellite cells, promoting cell
proliferation and differentiation and repairing damaged muscle
fibers.
The present study sought to investigate whether the
secretome of human (h)DPSCs can alleviate hypoxic injury in C2C12
myoblasts and determine whether the underlying mechanism is
associated with regulation of the Wnt/β-catenin signaling
pathway.
Materials and methods
hDPSC isolation and CM preparation
Normal human third molar teeth (free of caries
and/or periodontitis) indicated for extraction were collected from
adults (patient characteristics are summarized in Table I) at the Shanghai Stomatological
Hospital. The Shanghai Stomatological Hospital Ethics Association
approved the study (approval no. 2019-003) and all methods were
implemented in accordance with relevant regulations. All patients
provided written informed consent to participate in the study.
Immediately following tooth extraction, the teeth were placed in
cold PBS containing 5% penicillin-streptomycin (Gibco; Thermo
Fisher Scientific, Inc.) and sent to the lab within 1 h. The tooth
surfaces were washed ten times with PBS and cut on the
cementoenamel junction. The pulp was gently separated from the
teeth, cut into 1 mm3 pieces and then digested in
collagenase type I (Gibco; Thermo Fisher Scientific, Inc.) and
dispase (Gibco; Thermo Fisher Scientific, Inc.) for 45 min to 1 h
at 37°C with occasional vortexing. Tissues and cells were cultured
in α-MEM (Gibco; Thermo Fisher Scientific, Inc.) supplemented with
10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.)
and 1% penicillin-streptomycin at 37°C in 5% CO2, and
the medium was changed every 3 days. All cells used in the present
study had undergone three to five passages. hDPSCs from up to three
donors were cultured separately and used to conduct the various
assays.
 | Table IDonor information. |
Table I
Donor information.
| Age, years | Sex | Recruitment
date |
|---|
| 18 | Male | 30/04/2018 |
| 25 | Female | 28/04/2018 |
| 25 | Female | 28/04/2018 |
A total of 2×105 cells were seeded in
100-mm dishes and, when the hDPSCs had reached 70–80% confluence,
the medium was removed, the cells were washed three times with PBS,
and the medium was replaced with 10 ml serum-free DMEM (Gibco;
Thermo Fisher Scientific, Inc.). After 48 h, the culture medium was
centrifuged at 1,000 × g for 3 min at room temperature; the
supernatant was collected and filtered through a 0.22-μm filter;
subsequently, the CM was concentrated 30-fold using an
ultrafiltration unit with a 10-kDa molecular weight cutoff
(Ultracel-10 membrane; EMD Millipore). All ultrafiltration units
were centrifuged at 4,000 × g for 25 min at 4°C. Each concentrated
CM was diluted 2-, 5- and 10-fold, and the unconcentrated medium
was 1-fold. The CM was stored at −80°C until further use (Fig. 1A). Serum-free DMEM without hDPSCs
was used as control CM, and it was incubated, collected and stored
in a similar manner.
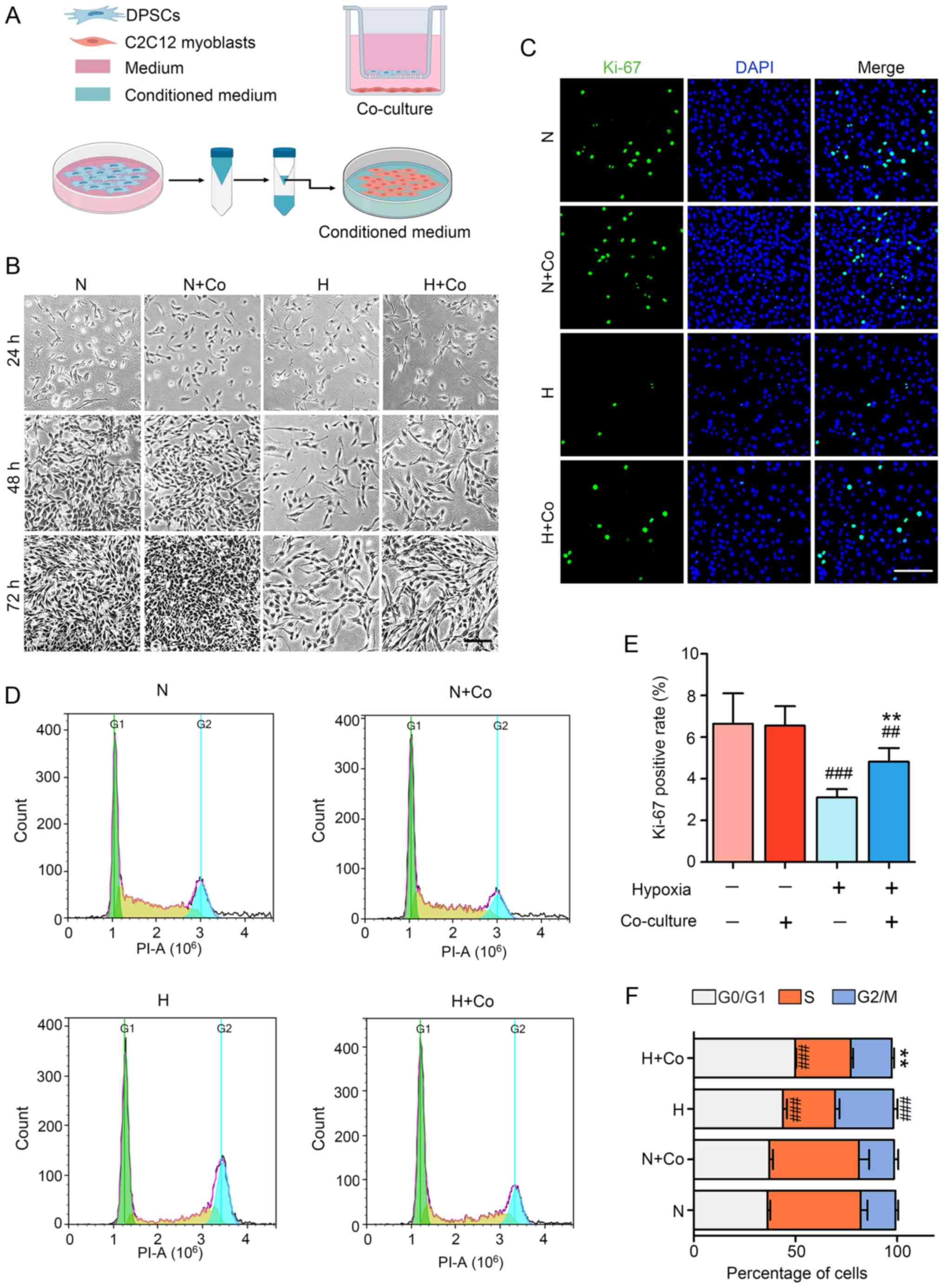 | Figure 1hDPSCs alleviate hypoxia-induced
injury of C2C12 myoblasts in the coculture model. (A) Establishing
the hDPSCs-C2C12 coculture system and preparing the hDPSC-CM.
Upper: The indirect coculture system with hDPSCs and C2C12
myoblasts was established. C2C12 and cultured hDPSCs were seeded in
6-well plates and Transwell-Clear inserts, respectively, to study
the paracrine effects. Lower: The hDPSC growth medium was replaced
with serum-free medium, after which time it was collected and
concentrated using an ultrafiltration unit (48 h) and then added to
regular medium after diluting to different concentrations. (B)
Optical micrographs of C2C12 cell morphology and quantity change at
24–72 h. Scale bar, 200 μm. (C) Cell proliferation was assessed
through Ki-67 immunofluorescence staining and (E) statistical
analysis of the Ki-67 (green)-positive rate. Scale bar, 100 μm. (D)
Cell cycle distribution was monitored using flow cytometry. (F)
Statistical analysis of the cell cycle in C2C12 myoblasts.
##P<0.01 and ###P<0.001 vs. N,
**P<0.01 vs. H. hDPSCs, human dental pulp stem cells;
DAPI, 4′,6-diamidino-2-phenylindole; PI, propidium iodide; N,
normoxia; H, hypoxia; Co, coculture. |
Cell culture and treatment
The murine myoblast cell line C2C12 was cultured in
DMEM with 10% FBS and 1% penicillin-streptomycin (growth medium,
GM) at 37°C in 5% CO2, and the medium was changed every
2 days. To introduce myogenic differentiation, the myoblasts were
transferred at 90% confluence to DMEM supplemented with 2% horse
serum (Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin (differentiation medium, DM), which was
replenished every other day. Hypoxia was induced in C2C12 cells by
employing the widely used hypoxia mimic 200 μM CoCl2
(Sigma-Aldrich; Merck KGaA), as detailed in the manufacturer’s
protocol. C2C12 myoblasts were treated as follows: Normoxia (N,
control medium), hypoxia (H, CoCl2 treatment with
control medium), normoxia + CM (N + CM), and hypoxia + CM (H + CM,
CoCl2 treatment with CM).
hDPSCs-C2C12 coculture system
In a typical experimental coculture system, as
illustrated in Fig. 1A,
2.5×103 C2C12 cells/well were seeded in 6-well plates
and Transwell-Clear inserts (3-μm pore size; Corning, Inc.) covered
with 5×103 hDPSCs/well were placed in another well.
After 24 h of cell attachment, the Transwell was assembled with
hDPSCs and wells containing C2C12. For C2C12 differentiation, C2C12
cells were first seeded in 6-well plates at a density of
4×105 cells/well in GM, and when the C2C12 cells reached
90% confluence, the GM was replaced with DM for 2 days to initiate
differentiation. The hDPSCs-C2C12 cells were further incubated for
2, 4 and 6 days in the same coculture experiments. The hDPSCs-C2C12
cells were treated as follows: Normoxia group (N), hypoxia group
(H), normoxia + coculture group (N + Co) and hypoxia + coculture
group (H + Co).
Flow cytometry
After treatment for 24 h, the cells were harvested
and centrifuged at 1,500 × g for 3 min at room temperature, and the
supernatant was discarded. The cells were fixed in 70% precooled
ethanol overnight at 4°C. The next day, the cells were centrifuged
at 1,500 × g for 3 min at room temperature and washed thrice with
PBS. RNase-containing PI solution (BioTime) was then added, and the
cells were incubated for 30 min at 37°C. Finally, cell cycle
analysis was performed using flow cytometry (ACEA NovoCyte) with
NovoExpress software version 1.4.1 (ACEA NovoCyte) within 24 h.
RNA extraction and gene expression
analysis using quantitative PCR
Total RNA was extracted using TRIzol reagent (Thermo
Fisher Scientific, Inc.) after different treatments in each group.
A total of 1 μg extracted RNA was reverse-transcribed into cDNA
using a FastQuant RT kit (Tiangen Biotech Co., Ltd.) at 42°C for 15
min and at 95°C for 3 min. SuperReal PreMix (Tiangen Biotech Co.,
Ltd.) was used for amplification of cDNA to the relative mRNA of
genes in 10 μl of the final volume using a Real-Time PCR System
(Biometra Biomedizinische Analytik GmbH). The thermocycling
conditions were as follows: 95°C for 15 min, followed by 40 cycles
at 95°C for 10 sec and at 60°C for 32 sec. The results were
normalized against the housekeeping gene 18S. The forwards and
reverse primers used are listed in Table II. The relative mRNA expression
was calculated by using 2−ΔΔCq method (38).
| Table IISequence of primers used for PCR
amplification. |
Table II
Sequence of primers used for PCR
amplification.
| Gene | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| 18s |
GTAACCCGTTGAACCCCATT |
CCATCCAATCGGTAGTAGCG |
| Myogenin |
GAGACATCCCCCTATTTCTACCA |
GCTCAGTCCGCTCATAGCC |
| MHC |
GCGAATCGAGGCTCAGAACAA |
GTAGTTCCGCCTTCGGTCTTG |
| Ccna2 |
AGAAGCTCAAGACTCGACGG |
AATGGTGAAGGCAGGCTGTT |
| Ccnd1 |
GCGTACCCTGACACCAATCTC |
ACTTGAAGTAAGATACGGAGGGC |
| Tcf7 |
CCCTCAATGCGTTCATGCTTT |
CTTGCGGGCCAGTTCATAGT |
| Lef1 |
GCCACCGATGAGATGATCCC |
TTGATGTCGGCTAAGTCGCC |
Western blotting
Cultured cells were lysed in RIPA buffer (Santa Cruz
Biotechnology, Inc.) with protease and phosphatase inhibitors.
Protein concentration was determined by the BCA Protein Assay kit
(Thermo Fisher Scientific, Inc). Subsequently, total protein
samples were prepared using 2X SDS loading buffer and equal amounts
of protein (30 μg) were denatured by boiling for 10 min. These
samples were loaded into a 10% SDS-PAGE gel and transferred onto
PVDF membranes (EMD Millipore). Ponceau (P0012; Beijing Solarbio
Science & Technology Co., Ltd.)-stained membranes were used for
detection of total protein. The membranes were blocked in 5%
non-fat milk diluted in TBS at room temperature for 1 h, and then
incubated overnight at 4°C with the following primary antibodies:
Rabbit anti-β-catenin (1:1,000, cat. no. 8480; Cell Signaling
Technology, Inc.), mouse anti-β-actin (1:5,000, cat. no. abs830031;
Absin Bioscience Inc.), mouse anti-myosin heavy chain (MHC; 1:60,
cat. no. MF20; Developmental Studies Hybridoma Bank), mouse
anti-myogenin (cat. no. sc-52903), mouse anti-MyoD (cat. no.
sc-32758), mouse anti-Wnt1 (cat. no. sc-5630), mouse anti-Wnt4
(cat. no. sc-376279), mouse anti-Wnt7a (cat. no. sc-365665), mouse
anti-glycogen synthase kinase (GSK)-3β (cat. no. sc-24563), mouse
anti-phosphorylated (p)-GSK-3β (cat. no. sc-11757) (all from Santa
Cruz Biotechnology, Inc., except where otherwise indicated, and
used at 1:1,000) and mouse anti-HIF-1α (1:500, cat. no. NB100-105;
Novus Biologicals). The membranes were washed with TBS-T (0.1%
Tween-20 in TBS) four times for 6 min each and then incubated with
horseradish peroxidase-conjugated secondary antibodies (1:10,000;
cat. no. 7076 for anti-mouse IgG, cat. no. 7074 for anti-rabbit
IgG, both from Cell Signaling Technology, Inc.) for 1.5 h at room
temperature. After four washes for 6 min with TBS-T, the blots were
visualized using an enhanced chemiluminescent substrate kit (ECL
Advance; Thermo Fisher Scientific, Inc.). The bands were detected
with Amersham Imager 600 (GE Healthcare) and then quantified using
the ImageJ program version 1.50i (National Institute of
Health).
Immunofluorescence staining
C2C12 cells were seeded in 24-well plates at
2×104/well. After treatment, the medium was carefully
removed and the cells were washed twice with PBS. The cells were
then fixed in precooled 4% paraformaldehyde for 15 min at room
temperature. Then, the cells were permeabilized in the presence of
PBS with 0.25% Triton X-100 for 15 min and blocked with 5% BSA at
room temperature for 1 h. The cells were incubated overnight at 4°C
with mouse anti-MHC (1:500; cat. no. MAB4470, R&D Systems,
Inc.), rabbit anti-Ki-67 (1:1,000; cat. no. MA5-14520, Thermo
Fisher Scientific, Inc.), or HIF-1α in 2.5% BSA. The cells were
then washed three times in PBS with 0.1% Tween-20 (PBST) and
stained for 1 h at room temperature with anti-mouse FITC-conjugated
secondary antibody (1:1,000, cat. no. F-2761, Thermo Fisher
Scientific, Inc.) in 2.5% BSA. After washing five times with PBST,
the cells were counterstained with 4′,6-diamidino-2-phenylindole
(1:10,000) at room temperature for 10 min. The cells were viewed
under a fluorescence microscope (Leica Microsystems, Inc.) and
assessed for myogenesis by measuring the number of MHC-positive
nuclei in the myotubes in 3–5 randomly selected fields to quantify
the differentiation index.
Schematic production
The diagrams, including co-culture model and
molecular mechanism, were created using the software available at
BioRender (https://app.biorender.com/).
Statistical analysis
All data were processed using GraphPad Prism 5
(GraphPad Software, Inc.) and ImageJ software version 1.50i
(National Institute of Health), and the measurement data were
analyzed as the mean ± standard deviation using one-way analysis of
variance and Tukey’s multiple comparison post hoc test with SPSS
Statistical 22.0 software (IBM, Corp.). P<0.05 was considered to
indicate a statistically significant difference. All assays were
repeated three times independently.
Results
hDPSCs exert cytoprotective effects by
enhancing C2C12 viability under hypoxic conditions in a coculture
model
To investigate whether the secretome of hDPSCs was
effective, an indirect coculture system of hDPSCs and C2C12 was
performed (Fig. 1A). After
treatment for 24–72 h, hypoxia in the other groups progressively
induced a decline in cell viability in a time-dependent manner,
compared with the control group. Hypoxia induction caused an
increase in the characteristic cellular extensions and surface area
in myoblasts, revealing the stressed state of the cells. After 24
h, there was no apparent difference between the groups under the
microscope, whereas the number of cells in the hDPSC coculture
group was visibly increased compared with that in the hypoxia group
after 48–72 h (Fig. 1B).
Furthermore, Ki-67 staining revealed that the rate of cell
proliferation significantly increased from 3.1±0.5 to 4.8±0.7%
after hDPSC coculture in hypoxic cells (Fig. 1C and E). In addition, the cell
cycle distribution indicated that hDPSCs blocked the G2/M phase
arrest caused by hypoxia (Fig. 1D and
F). Thus, hDPSCs in coculture with C2C12 were shown to
ameliorate hypoxia-related injury of the skeletal muscle myoblasts
through paracrine effects.
hDPSC-CM attenuates hypoxia-induced
injury of C2C12 myoblasts in a HIF-1α-independent manner
As MSCs secrete protein or growth factors in their
medium, whether hDPSC-CM was beneficial to hypoxia-exposed C2C12
myoblasts was next investigated. In this study, different
fold-increases of hDPSC-CM were used for treatment. Compared with
the hypoxia group, it was observed that 5-, 8- and 10-fold CM
achieved obvious effects after 48 h (Fig. 2A). After cell counting, it was
found that, compared with the hypoxia group, there was no
significant difference for the use of 2-fold CM, while there were
significant differences for the use of 5-, 8- and 10-fold CM. There
was no significant difference among the 5-, 8- and 10-fold CM
groups (Fig. 2C). Therefore, 5X
hDPSC-CM was selected for the subsequent experiments. The
proliferation of cells was also improved in the H + CM group
compared with that of cells in the hypoxia group (Fig. 2B and E). Furthermore, whether the
effect of hDPSC-CM was associated with changes in HIF-1α was
tested. The results demonstrated that hypoxia increased the
expression of HIF-1α in the nucleus, but there was no significant
difference in the effect of hDPSC-CM on HIF-1α compared with that
in the H group (Fig. 2D and F).
The results suggested that hDPSC-CM markedly improved the
proliferation and viability of C2C12, but this protective effect
appeared to be unrelated to the level of HIF-1α.
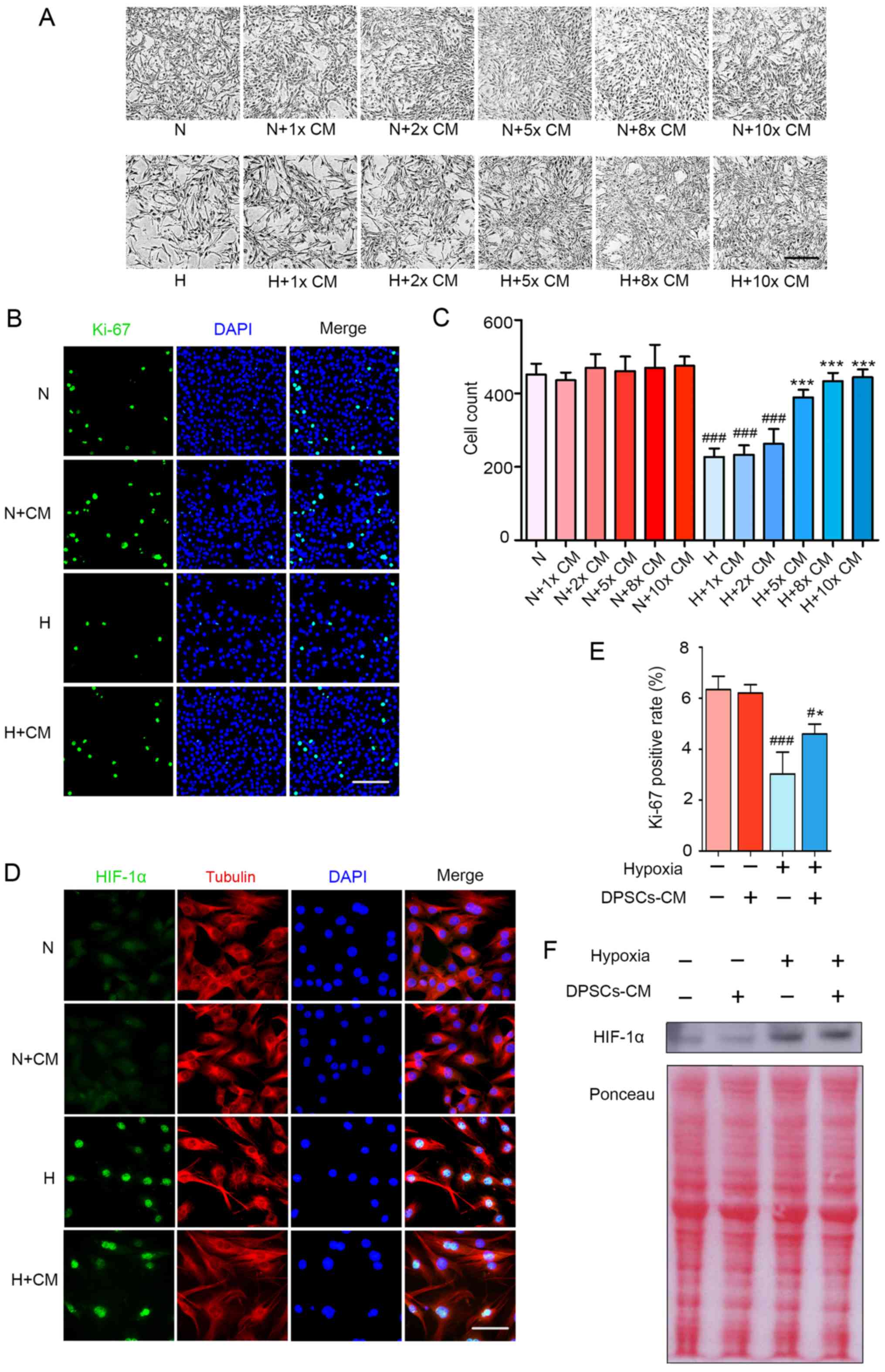 | Figure 2hDPSC-CM improves the hypoxia-induced
decrease in the proliferation and viability of C2C12 cells in a
HIF-1α-independent manner. (A) Optical micrographs of C2C12 cell
morphology and quantity change for 48 h. Scale bar, 200 μm. (B)
Cell proliferation was assessed through Ki-67 immunofluorescence
staining. Scale bar, 100 μm. (C) The number of cells per field were
counted at a magnification of ×100. (D) After 24 h of hypoxia
treatment and immunofluorescence staining of HIF-1α (green)
translocation to the nucleus, the cytoplasm was stained with
tubulin (red) and the cell nuclei with DAPI (blue). Scale bar, 50
μm. (E) Cell proliferation was assessed via statistical analysis of
the Ki-67-positive rate. (F) Nucleoprotein expression of HIF-1α was
detected using western blotting. #P<0.05 and
###P<0.001 vs. N, *P<0.05 and
***P<0.001 vs. H. HIF, hypoxia inducible factor;
hDPSCs, human dental pulp stem cells; CM, conditioned media; N,
normoxia; H, hypoxia; DAPI, 4′,6-diamidino-2-phenylindole. |
hDPSCs improve hypoxia-induced injury of
myogenic differentiation in a cell coculture model
To assess whether hDPSCs affected the physiological
function of myoblasts, C2C12 cells were induced to differentiate
for 2 days under normoxic conditions followed by hypoxic treatment
for 2, 4 and 6 days. Myogenin and MHC are specific middle and late
differentiation markers during the process of myoblast fusion to
form myotubes (39). The myotubes
stained positive for MHC by immunofluorescence. These results
demonstrated that the H group significantly inhibited C2C12
differentiation, myotube rupture and atrophy (Fig. 3A). The data demonstrated that the
H + Co group stimulated the myogenesis of C2C12 after 2 days, with
a 2.8-fold increase in the index of differentiation compared with
the H group, whereas there were no significant differences observed
between the N and the N + Co groups (Fig. 3D). Furthermore, hDPSCs induced an
increase in myogenin mRNA expression and relative protein levels
that were 1.66- and 3.3-fold higher after 2 days of
differentiation, respectively, compared with the H group (Fig. 3B, E and F). Analysis of the
terminal differentiation marker MHC revealed that its mRNA
expression and relative protein level were 2.02- and 2.7-fold
higher, respectively, in the H + Co group compared with the H group
(Fig. 3C, E and G). These results
suggest that hDPSCs protected myotubes against hypoxia-induced
injury by restoring myogenic differentiation and myotube
morphology. However, it was also observed that the protective
effect of coculture gradually weakened after 4–6 days and was not
significantly different with or without hDPSCs. This may be an
experimental limitation of coculture in the study, as both C2C12
and the hDPSCs were subjected to hypoxic conditions over a long
period of time. Damaged hDPSCs may lead to an attenuation of
protection due to their decreased paracrine function, even though
they continue to achieve significant results in the first 2 days of
treatment.
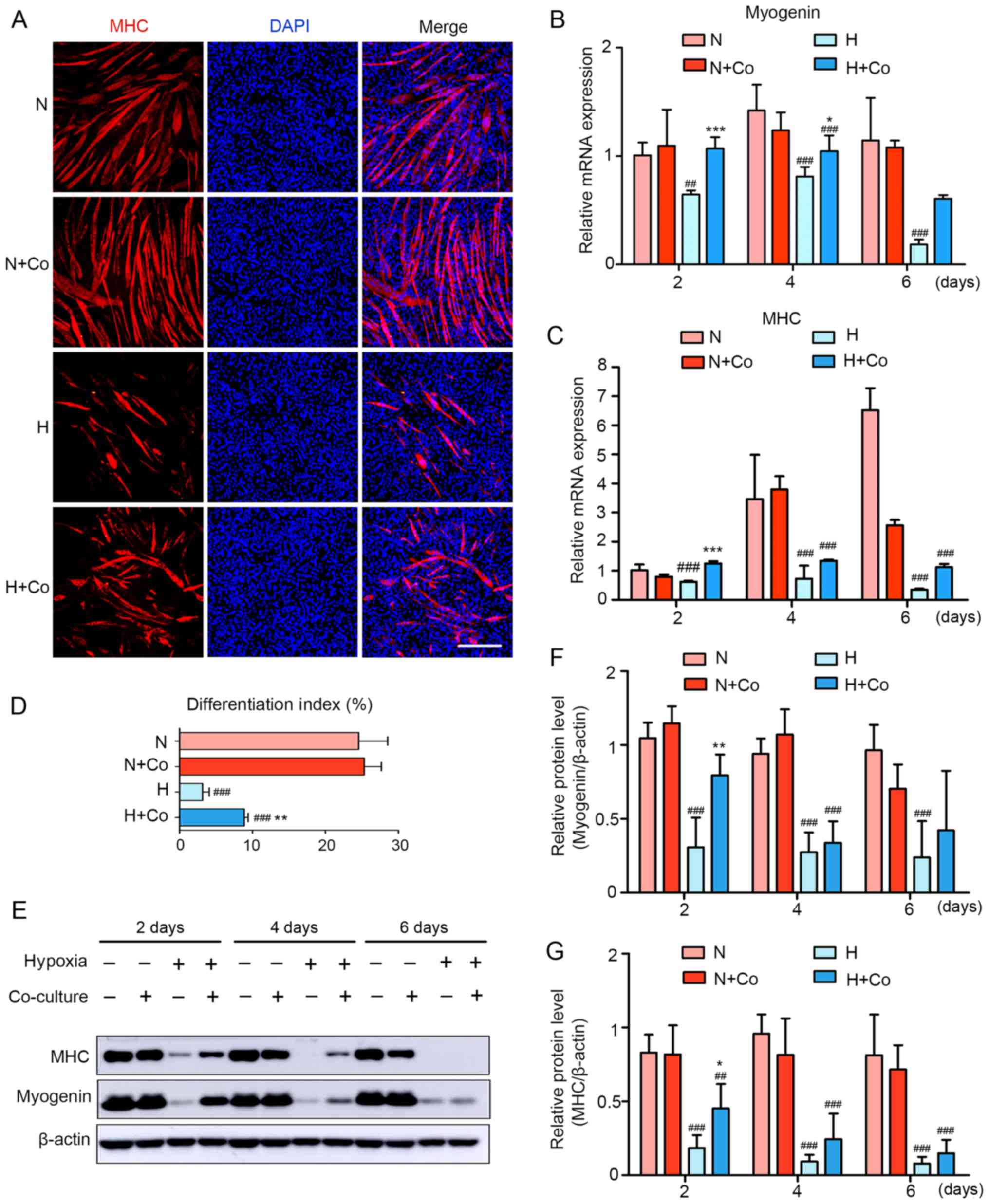 | Figure 3hDPSCs improve hypoxia-induced
inhibition of myogenic differentiation in the coculture model. (A)
Two days after hypoxia treatment, the formation of myotubes and
cell nuclei was assessed using immunofluorescence staining with MHC
(red) and DAPI (blue), respectively. Scale bar, 200 μm. (B and C)
The mRNA expression of (B) myogenin and (C) MHC was assessed using
quantitative PCR. Data are presented as the fold-change of the
normoxia group at 2 days. (D) The differentiation index is
presented as the ratio of MHC-positive nuclei to total nuclei. (E)
Protein expression of myogenin and MHC was assessed using western
blotting. Semiquantitative analysis of the expression ratio of (F)
myogenin/β-actin and (G) MHC/β-actin. ##P<0.01 and
###P<0.001 vs. N, *P<0.05,
**P<0.01 and ***P<0.001 vs. H. hDPSCs,
human dental pulp stem cells; CM, conditioned media; DAPI,
4′,6-diamidino-2-phenylindole; N, normoxia; H, hypoxia; Co,
coculture; MHC, myosin heavy chain. |
hDPSC-CM ameliorates hypoxia-induced
injury of myogenic differentiation
Whether hDPSC-CM was beneficial to myogenic
differentiation was next investigated. In the present study,
different folds of hDPSC-CM were selected for treatment (1-, 2-, 5-
and 10-). As shown in Fig. 4A–C,
hypoxia decreased the relative expression of myogenic
differentiation genes (MyoD, myogenin and MHC) for 48 h, whereas a
moderate concentration of hDPSC-CM reversed the inhibition of
myogenic differentiation genes under hypoxia, particularly 2X
hDPSC-CM. The protein level of MyoD was inconsistent with mRNA
expression. Compared with other groups, the relative protein level
of MyoD in the normoxia group was significantly downregulated
(Fig. 4D and F). The authors
considered this result to be partly due to the temporal expression
pattern during myogenesis. MyoD is an early expressed gene, while
myogenin and MHC are expressed in the middle and late stages.
Moreover, it was also possible that MyoD is regulated by
post-transcriptional modifications (40). However, analysis of myogenin and
MHC after 48 h revealed that their relative protein levels were
upregulated in H + 2X hDPSC-CM compared with the hypoxia group, but
1X, 5X and 10X hDPSC-CM was not associated with a significant
change (Fig. 4E, G and H). These
results demonstrated that treatment with hDPSC-CM at a moderate
concentration was capable of attenuating the inhibition of myogenic
differentiation, thereby protecting cells and myotubes.
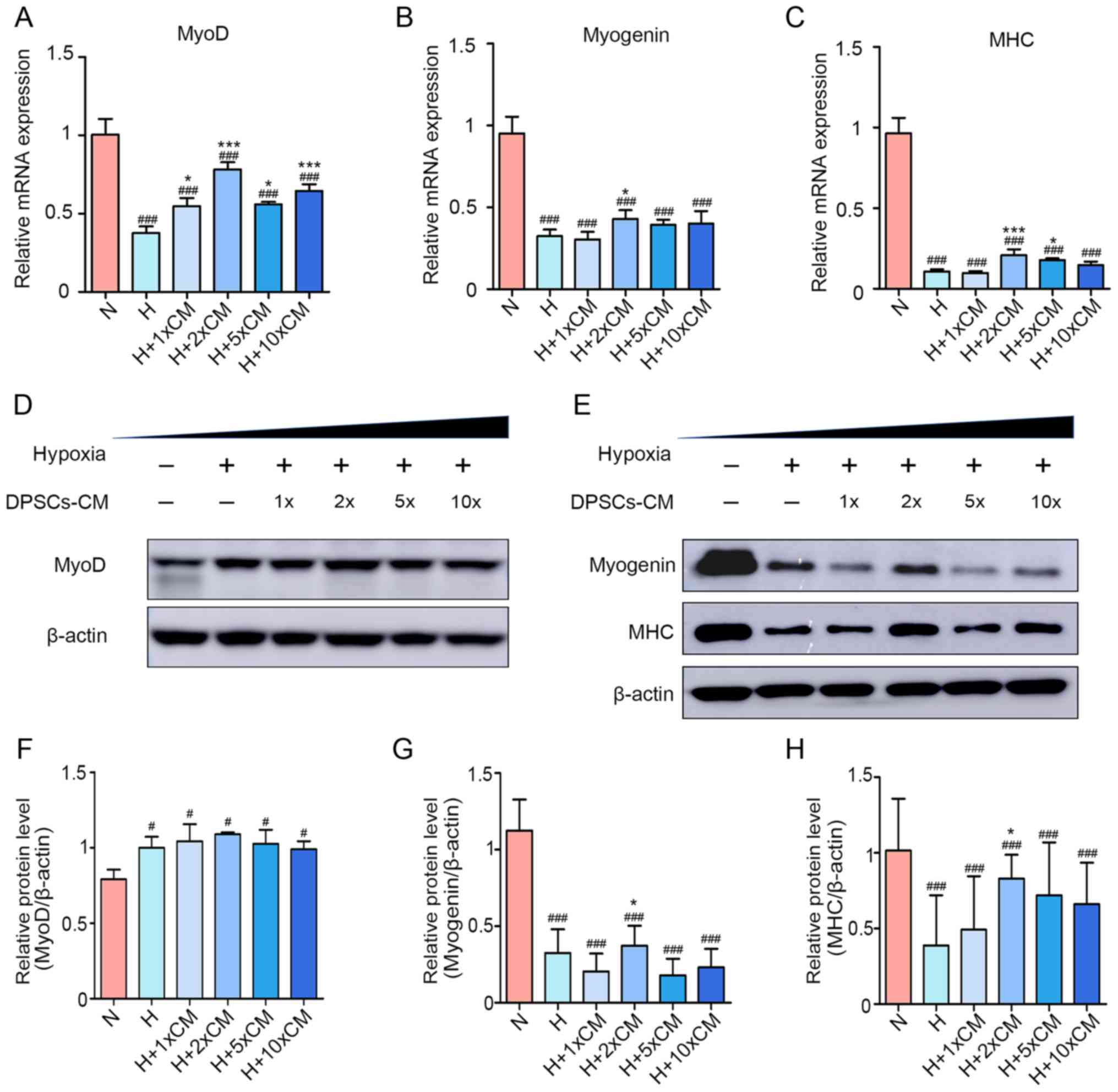 | Figure 4Appropriate concentration of hDPSC-CM
improves the inhibition of myogenic differentiation. mRNA
expression of (A) MyoD, (B) myogenin and (C) MHC. Protein levels of
(D) MyoD, (E) myogenin and MHC during the differentiation process.
Semiquantitative analysis of the expression ratios of (F)
MyoD/β-actin, (G) myogenin/β-actin and (H) MHC/β-actin.
#P<0.05 and ###P<0.001 vs. N,
*P<0.05 and ***P<0.001 vs. H. hDPSCs,
human dental pulp stem cells; CM, conditioned media; DAPI,
4′,6-diamidino-2-phenylindole; N, normoxia; H, hypoxia; MHC, myosin
heavy chain. |
hDPSC-CM mediates Wnt/β-catenin signaling
in hypoxia-treated C2C12 myoblasts
Wnt/β-catenin signaling has been shown to be
involved in hypoxia-induced cell injury (41,42). To further explore the mechanisms
of myoblast damage and the potential protective role of hDPSC-CM
against hypoxia, the protein levels of total GSK-3β, p-GSK-3β (S9)
and β-catenin were detected between 6 and 24 h. As shown in
Fig. 5A–C, the relative protein
levels of p-GSK-3β (S9) and β-catenin in the hypoxia group were
lower compared with those in the normoxia group for 12 and 24 h,
but there were no significant differences at 6 h. In the presence
of hDPSC-CM, the H + CM group exhibited upregulated levels of
p-GSK-3β (S9) and β-catenin. In addition, hypoxia inhibited the
downstream target genes of Wnt/β-catenin, including lymphoid
enhancer binding factor 1 (Lef1), transcription factor 7 (Tcf7),
cyclin D1 (Ccnd1) and cyclin A2 (Ccna2). After hDPSC-CM treatment,
the expression of target genes was restored under hypoxic
conditions (Fig. 5D–G).
Furthermore, due to the possible involvement of the Wnt/β-catenin
pathway in the therapeutic effect of hDPSC-CM, the protein levels
of Wnt ligands that mediate Wnt signaling activation in C2C12
myoblasts were analyzed. These ligands included Wnt1, Wnt4 and
Wnt7a. It was observed that Wnt1 was decreased by hypoxia and then
recovered after hDPSC-CM treatment, whereas there was no
statistically significant difference in Wnt4 and Wnt7a (Fig. 5H–K). The present results suggested
that hDPSC-CM alleviated C2C12 myoblast injury at least partly
through activation of Wnt1/β-catenin signaling.
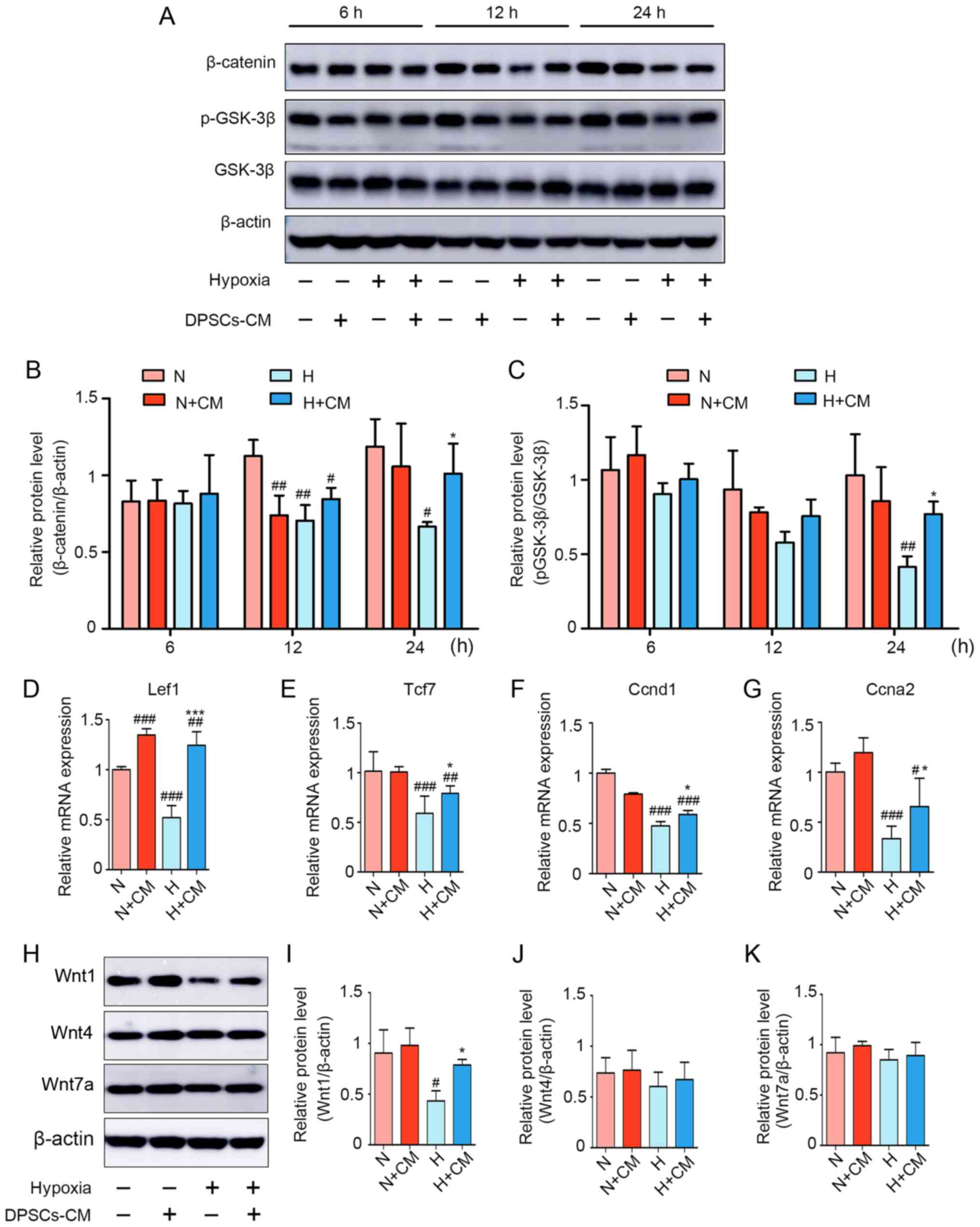 | Figure 5Wnt/β-catenin signaling pathway is
involved in hypoxia-induced C2C12 myoblast injury and is activated
by hDPSC-CM. (A) Expression of β-catenin, p-GSK-3β (S9) and total
GSK-3β was detected using western blotting for 6, 12 and 24 h.
Semiquantitative analysis of the expression ratios of (B)
β-catenin/β-actin and (C) p-GSK-3β/GSK-3β. The levels of the
downstream effectors of β-catenin-related mRNAs, including (D)
Lef1, (E) Tcf7, (F) Ccnd1 and (G) Ccna2, were assessed using
quantitative PCR. (H) Expression of Wnt signaling ligands,
including Wnt1, Wnt4 and Wnt7a. Semiquantitative analysis of the
expression ratio of (I) Wnt1/β-actin, (J) Wnt4/β-actin and (K)
Wnt7a/β-actin. #P<0.05, ##P<0.01 and
###P<0.001 vs. N, *P<0.05 and
***P<0.001 vs. H. hDPSCs, human dental pulp stem
cells; CM, conditioned media; p-GSK, phosphorylated-glycogen
synthase kinase; Lef1, lymphoid enhancer binding factor 1; Tcf7,
transcription factor 7; Ccnd1, cyclin D1; Ccna2, cyclin A2; N,
normoxia; H, hypoxia. |
The protective effect of hDPSC-CM is
blocked by inhibition of Wnt/β-catenin signaling
To further support the present results on the
protective role of the Wnt/β-catenin pathway in hDPSC-CM, XAV939
(20 μM) was used a specific inhibitor of Wnt/β-catenin that
promotes the phosphorylation and degradation of β-catenin. The
results revealed that XAV939 induced a decrease in both the
differentiation indices and the Ki-67-positive rate compared with
the H + CM group (Fig. 6A–D).
Moreover, the inhibitor XAV939 induced a reduction in the
expression of myogenin and MHC compared with the H + CM group
(Fig. 6E and F).
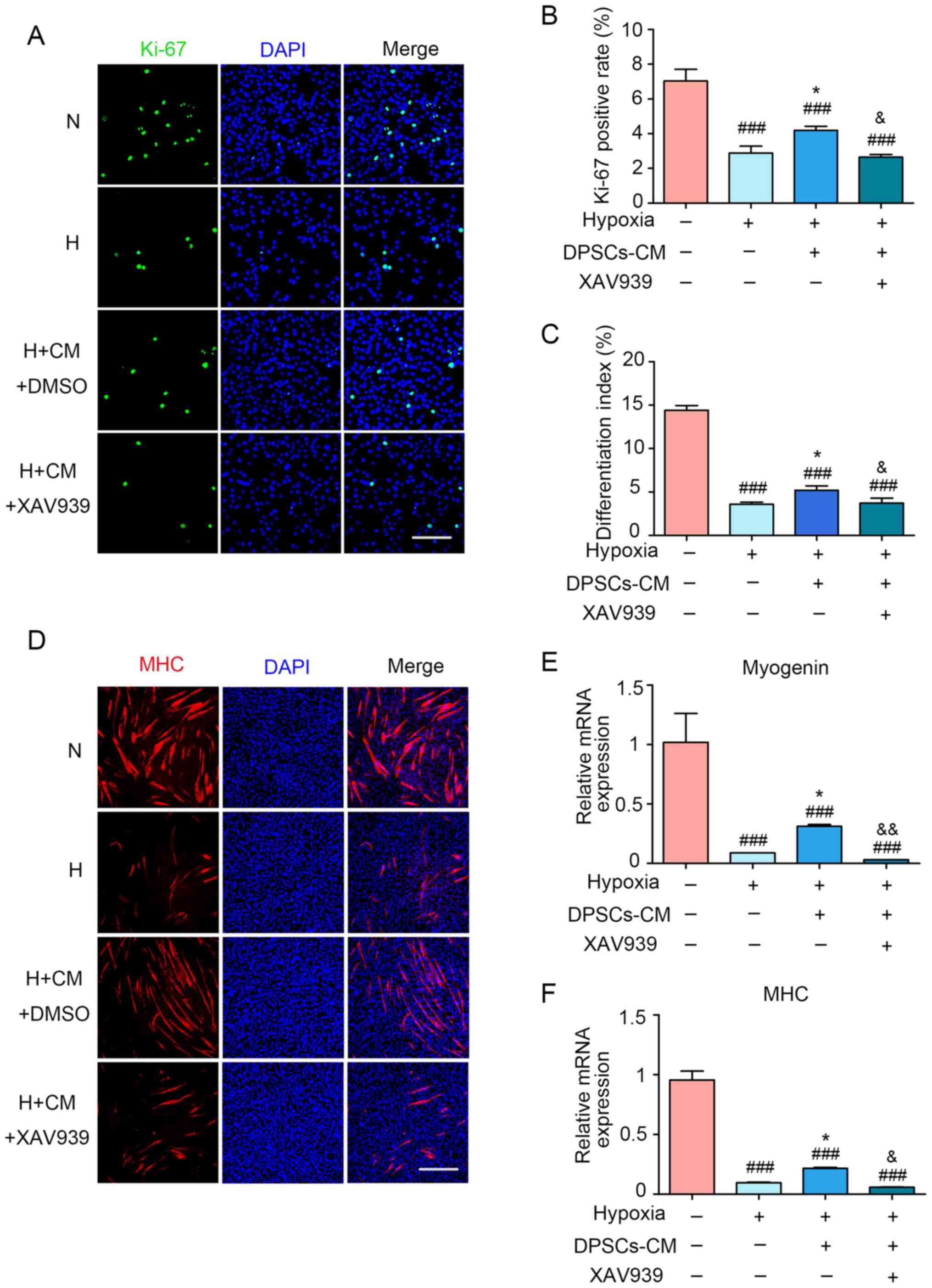 | Figure 6The protective effect of hDPSC-CM was
blocked by using XAV939. (A) Cell proliferation was assessed
through Ki-67 immunofluorescence staining. Scale bar, 100 μm. (B)
Statistical analysis of the Ki-67-positive rate (%). (C) The
differentiation index was presented as the ratio between
MHC-positive nuclei and the total number of nuclei. (D) Two days
after hypoxia treatment, the formation of myotubes and cell nuclei
was assessed via immunofluorescence staining with MHC (red) and
DAPI (blue), respectively. Scale bar, 200 μm. The mRNA expression
of (E) myogenin and (F) MHC was detected using quantitative PCR.
###P<0.001 vs. N, *P<0.05 vs. H.
&P<0.05 and &&P<0.01 vs. H
+ CM. hDPSCs, human dental pulp stem cells; CM, conditioned media;
DAPI, 4′,6-diamidino-2-phenylindole; N, normoxia; H, hypoxia; MHC,
myosin heavy chain. |
Discussion
The present study revealed that hDPSCs attenuated
hypoxia-induced myoblast injury by improving viability and myogenic
differentiation. It was demonstrated that hDPSCs activated the
Wnt1/GSK-3β/β-catenin signaling pathway by paracrine factors, which
was at least partly responsible for these protective effects.
Previous studies have demonstrated pathological
hypoxia-induced skeletal muscle myoblast injury, including the
inhibition of viability and differentiation (43–45). Regarding viability,
CoCl2 produced changes typical of cell death, such as
characteristic cell extension, increased volume, chromatin
condensation and G2/M phase cell cycle arrest, ultimately leading
to reduced proliferation (13).
Regarding myogenic differentiation, hypoxia, by degrading early and
intermediate markers, such as MyoD and myogenin, prevented terminal
differentiation (46). In the
present study, following exposure to hypoxia, myoblasts exhibited a
decrease in viability, arrest at the G2/M phase and inhibition of
myogenin and MHC expression, which was in agreement with the
aforementioned reports on skeletal muscle hypoxic injury.
MSCs and their derivatives are known to positively
affect skeletal myogenesis and repair injury. In particular,
fibroblast growth factor, hepatocyte growth factor, insulin-like
growth factor, vascular endothelial growth factor and members of
the Wnt family are involved in differentiation (29,47,48). Therefore, the indirect coculture
system was first adopted. Upon observing a beneficial effect, it
was hypothesized that these secretions were present in the CM, and
the results were then verified with hDPSC-CM. In previous
applications of MSC-CM, however, most studies (49–52) did not compare the different
concentrations of CM. Nagata et al (53) demonstrated that MSC-CM enhanced
tissue regeneration and repair, depending on the concentration
ratio of CM. However, different cells and tissues may have
different sensitivities to different CM concentrations. In the
present study, it was demonstrated that different hDPSC-CM
concentrations produced different effects. In the process of
myoblast proliferation, an ~5-fold concentration of CM is
considered to have a significant effect; however, such a high
concentration does not appear to be required during myoblast
differentiation. In the present experiment, a protective effect was
achieved using only an ~2-fold concentration during
differentiation. This difference was considered to be caused by the
differing demand for paracrine substances during the repair
processes of proliferation and differentiation. Convincing evidence
was presented herein that hDPSCs exert beneficial effects on
myoblast hypoxic injury. However, there were several limitations to
the present study. Primary DPSCs and their CM were derived from
humans, while the C2C12 myoblasts were from murine lines.
Therefore, interactions between the two cells may be affected by
structural differences of cytokines and proteins due to the
different species. Although numerous soluble factors from humans
and mice may interact, it is not true for all of them and the level
of responsiveness may not be equal to that to cytokines from the
same species. In the present study, C2C12 myoblasts cocultured with
human DPSCs achieved good cellular growth under normoxia, and even
better under hypoxia, when compared with the C2C12 alone groups.
The interactions between the two types of cells appeared to be
positive. However, the protective effect may be compromised.
Additionally, the present study was unable to mimic the dynamic
changes of real tissue hypoxia. Thus, further studies are warranted
to investigate the effect of hDPSCs on human primary skeletal
myoblasts in hypoxia-related diseases.
The findings of the present study provide
theoretical support for the exploration of the repair of hypoxic
damage to myoblasts by hDPSCs; however, a deeper understanding of
the underlying mechanism of these findings requires further
investigation. Wnt signaling also plays a crucial role in the
regulation of myogenic differentiation, as Wnt is induced and
promotes myoblast differentiation and myotube fusion (54). Wnt ligands bind to low-density
lipoprotein-related protein/frizzled complexes on the cell membrane
and after phosphorylation, inactivation of GSK-3β leads to
stabilization of β-catenin. This gradually accumulates and is
transferred into the nucleus to promote transcription of downstream
specific target genes (such as Lef1 and Ccnd1), or directly
activates MyoD and upregulates myogenic regulatory factor
coactivators (42). The Wnt
signaling pathway is weakly activated in mature skeletal muscle.
However, after injury, satellite cells are activated in the
skeletal muscle, where numerous Wnt ligands (such as Wnt1, Wnt3a,
Wnt4 and Wnt7a/b) are expressed and secreted (37). Moreover, Leroux et al
(48) reported that MSCs, via the
Wnt4 pathway, improved skeletal muscle fiber regeneration following
ischemic injury. Other studies have also demonstrated activation of
the Wnt pathway by MSCs (55–57). The present study found that, in
the presence of hDPSC-CM, the levels of Wnt/β-catenin
pathway-related proteins, including p-GSK-3β and β-catenin, were
upregulated. Furthermore, the downstream target genes of
Wnt/β-catenin, including Lef1, Tcf7, Ccnd1 and Ccna2, were
inhibited by hypoxia, whereas their expression levels were restored
after hDPSC-CM treatment. To confirm these data, in the presence of
hDPSC-CM, myoblasts were cultured with 20 μM of the Wnt/β-catenin
inhibitor XAV939 and the protective effects were blocked. These
results suggest that the protective role of hDPSCs on C2C12
hypoxia-induced injury requires the participation of the Wnt
signaling pathway.
Under hypoxic conditions, the Wnt/β-catenin pathway
is directly or indirectly modulated. A previous study reported that
HIF-1α blocked the Wnt/β-catenin signaling pathway by inhibiting
hARD1-mediated β-catenin (58).
Indeed, increased expression of HIF-1α inhibits canonical Wnt
signaling during skeletal muscle repair, as reflected by the
increased target genes of β-catenin expression after silencing
HIF-1α (42). Thus, the present
study hypothesized that the paracrine protective effect of hDPSCs
through activation of the Wnt/β-catenin pathway was related to the
prevention of HIF-1α accumulation. However, in the current study,
hDPSC-CM did not decrease hypoxia-induced HIF-1α stabilization.
During the various stages of skeletal muscle repair
and regeneration, correct activation of the Wnt signaling pathways
is crucial. Wnt1 and Wnt4 mainly activate the canonical Wnt pathway
(37). However, Wnt7a did not
activate β-catenin in myoblasts or muscle myofibers. Indeed, by
regulating the PCP pathway, Wnt7a signaling can be described as a
non-canonical Wnt pathway (42).
C2C12, in which the Wnt1/β-catenin pathway was able to enhance
myogenic differentiation, indicates one of the potential roles of
canonical Wnt signaling in skeletal muscle (59). Consistent with reports mentioned
above, the present study demonstrated that hDPSC-CM activated Wnt1
and β-catenin expression and regulated GSK-3β at S9, leading to its
inactivation. Therefore, hDPSC-CM may restore the Wnt1/β-catenin
pathway in myoblasts to alleviate hypoxia-induced injury.
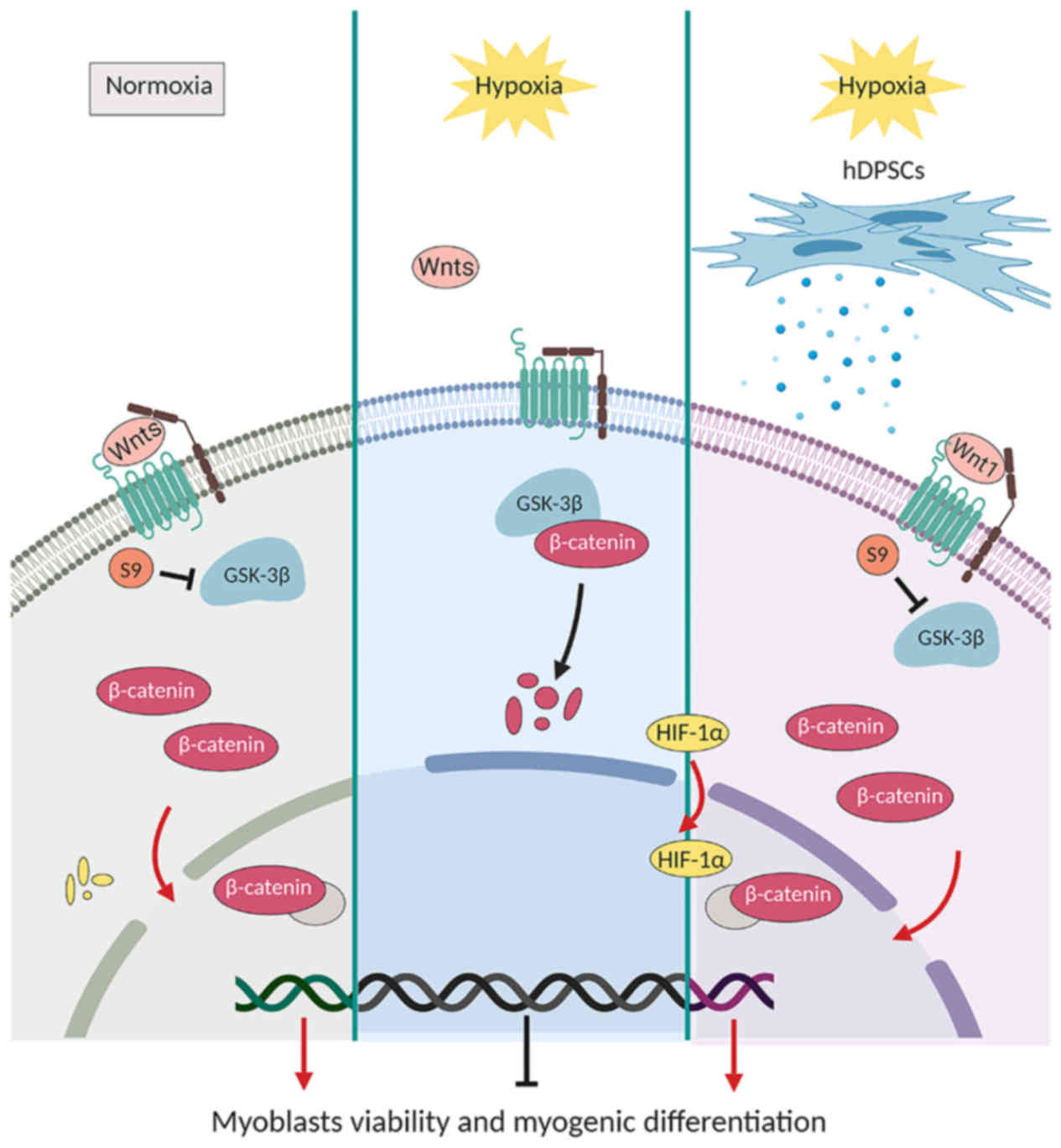
In summary, the findings of the present study
suggest that hDPSCs may alleviate hypoxia-induced injury in C2C12
myoblasts and the underlying mechanism may be associated with
regulation of the Wnt1/β-catenin signaling pathway (Fig. 7). The next step would be to assess
which paracrine factors of hDPSCs are effective in repairing
injured myoblasts, and determine whether another mechanism could be
involved in vitro or in an animal model.
Acknowledgements
Not applicable.
Funding
The present study was funded by the National Natural
Science Foundation of China (grant nos. 81771109 and 81600897); the
General Program Shanghai Municipal Health and Family Planning
Commission (grant nos. 201640023 and 201740091; and the Natural
Science Foundation of Shanghai (grant no. 19ZR1445400).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors’ contributions
WZ: Conception and design, data analysis and
interpretation, manuscript writing and figure editing. LY:
Experiment operation, data collection, analysis and interpretation.
XH and YLu: Financial support, data analysis and interpretation.
JP, JD, LZ, WH and SL: Provision of study materials, collection and
assembly of data. YLiu and QL: Conception and design, financial
support, data analysis and interpretation. WZ and LY contributed
equally. All the authors have read and approved the final version
of the manuscript.
Ethics approval and consent to
participate
The protocols were approved by the Shanghai
Stomatological Hospital Ethics Association (Shanghai, China).
Written informed consent was obtained from all participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Koh MY and Powis G: Passing the baton: The
HIF switch. Trends Biochem Sci. 37:364–372. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chaillou T: Skeletal muscle fiber type in
hypoxia: Adaptation to high-altitude exposure and under conditions
of pathological hypoxia. Front Physiol. 9:14502018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Adams V, Linke A and Winzer E: Skeletal
muscle alterations in HFrEF vs. HFpEF Current Heart Failure
Reports. 14:489–497. 2017. View Article : Google Scholar
|
|
4
|
Gea J, Agusti A and Roca J:
Pathophysiology of muscle dysfunction in COPD. J Appl Physiol.
1985. 114:1222–1234. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Putti R, Migliaccio V, Sica R and Lionetti
L: Skeletal muscle mitochondrial bioenergetics and morphology in
high fat diet induced obesity and insulin resistance: Focus on
dietary fat source. Front Physiol. 6:4262016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lu Y, Liu Y and Li Y: Comparison of
natural estrogens and synthetic derivative on genioglossus function
and estrogen receptors expression in rats with chronic intermittent
hypoxia. J Steroid Biochem Mol Biol. 140:71–79. 2014. View Article : Google Scholar
|
|
7
|
Williams R, Lemaire P, Lewis P, McDonald
FB, Lucking E, Hogan S, Sheehan D, Healy V and O’Halloran KD:
Chronic intermittent hypoxia increases rat sternohyoid muscle NADPH
oxidase expression with attendant modest oxidative stress. Front
Physiol. 6:152015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Beaudry M, Hidalgo M, Launay T, Bello V
and Darribère T: Regulation of myogenesis by environmental hypoxia.
J Cell Sci. 129:2887–2896. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chaillou T, Koulmann N, Meunier A, Chapot
R, Serrurier B, Beaudry M and Bigard X: Effect of hypoxia exposure
on the recovery of skeletal muscle phenotype during regeneration.
Mol Cell Biochem. 390:31–40. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Favier FB, Britto FA, Freyssenet DG,
Bigard XA and Benoit H: HIF-1-driven skeletal muscle adaptations to
chronic hypoxia: Molecular insights into muscle physiology. Cell
Mol Life Sci. 72:4681–4696. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Quadrilatero J, Alway SE and
Dupont-Versteegden EE: Skeletal muscle apoptotic response to
physical activity: Potential mechanisms for protection. Appl
Physiol Nutr Metab. 36:608–617. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
L’honoré A, Commère PH, Ouimette JF,
Montarras D, Drouin J and Buckingham M: Redox regulation by Pitx2
and Pitx3 is critical for fetal myogenesis. Dev Cell. 39:7562016.
View Article : Google Scholar
|
|
13
|
Muñoz-Sánchez J and Chánez-Cárdenas ME:
The use of cobalt chloride as a chemical hypoxia model. J Appl
Toxicol. 39:556–570. 2019. View
Article : Google Scholar
|
|
14
|
Hayot M, Rodriguez J, Vernus B, Carnac G,
Jean E, Allen D, Goret L, Obert P, Candau R and Bonnieu A:
Myostatin up-regulation is associated with the skeletal muscle
response to hypoxic stimuli. Mol Cell Endocrinol. 332:38–47. 2011.
View Article : Google Scholar
|
|
15
|
Chen R, Xu J, She Y, Jiang T, Zhou S, Shi
H and Li C: Necrostatin-1 protects C2C12 myotubes from
CoCl2-induced hypoxia. Int J Mol Med. 41:2565–2572. 2018.PubMed/NCBI
|
|
16
|
Baskaran R, Kalaiselvi P, Huang CY and
Padma VV: Neferine, a bisbenzylisoquinoline alkaloid, offers
protection against cobalt chloride-mediated hypoxia-induced
oxidative stress in muscle cells. Integr Med Res. 4:231–241. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen R, Jiang T, She Y, Xu J, Li C, Zhou
S, Shen H, Shi H and Liu S: Effects of cobalt chloride, a
hypoxia-mimetic agent, on autophagy and atrophy in skeletal C2C12
myotubes. Biomed Res Int. 2017:7097580. 2017.
|
|
18
|
Rovetta F, Stacchiotti A, Faggi F,
Catalani S, Apostoli P, Fanzani A and Aleo MF: Cobalt triggers
necrotic cell death and atrophy in skeletal C2C12 myotubes. Toxicol
Appl Pharmacol. 271:196–205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jaitovich A and Barreiro E: Skeletal
muscle dysfunction in chronic obstructive pulmonary disease. What
we know and can do for our patients. Am J Respir Crit Care Med.
198:175–186. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hirai DM, Musch TI and Poole DC: Exercise
training in chronic heart failure: Improving skeletal muscle O2
transport and utilization. Am J Physiol Heart Circ Physiol.
309:H1419–H1439. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guimarães KC, Drager LF, Genta PR,
Marcondes BF and Lorenzi-Filho G: Effects of oropharyngeal
exercises on patients with moderate obstructive sleep apnea
syndrome. Am J Respir Crit Care Med. 179:962–966. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chaudhary P, Sharma YK, Sharma S, Singh SN
and Suryakumar G: High altitude mediated skeletal muscle atrophy:
Protective role of curcumin. Biochimie. 156:138–147. 2019.
View Article : Google Scholar
|
|
23
|
Kerkis I, Ambrosio CE, Kerkis A, Martins
DS, Zucconi E, Fonseca SA, Cabral RM, Maranduba CM, Gaiad TP,
Morini AC, et al: Early transplantation of human immature dental
pulp stem cells from baby teeth to golden retriever muscular
dystrophy (GRMD) dogs: Local or systemic. J Transl Med. 6:352008.
View Article : Google Scholar
|
|
24
|
Nakatsuka R, Nozaki T, Uemura Y, Matsuoka
Y, Sasaki Y, Shinohara M, Ohura K and Sonoda Y:
5-Aza-2′-deoxycytidine treatment induces skeletal myogenic
differentiation of mouse dental pulp stem cells. Arch Oral Biol.
55:350–357. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Spath L, Rotilio V, Alessandrini M,
Gambara G, De Angelis L, Mancini M, Mitsiadis TA, Vivarelli E, Naro
F, Filippini A and Papaccio G: Explant-derived human dental pulp
stem cells enhance differentiation and proliferation potentials. J
Cell Mol Med. 14:1635–1644. 2010. View Article : Google Scholar
|
|
26
|
Kichenbrand C, Velot E, Menu P and Moby V:
Dental pulp stem cell-derived conditioned medium: An attractive
alternative for regenerative therapy. Tissue Eng Part B Rev.
25:78–88. 2019. View Article : Google Scholar
|
|
27
|
Madrigal M, Rao KS and Riordan NH: A
review of therapeutic effects of mesenchymal stem cell secretions
and induction of secretory modification by different culture
methods. J Transl Med. 12:2602014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Assoni A, Coatti G, Valadares MC, Beccari
M, Gomes J, Pelatti M, Mitne-Neto M, Carvalho VM and Zatz M:
Different donors mesenchymal stromal cells secretomes reveal
heterogeneous profile of relevance for therapeutic use. Stem Cells
Dev. 26:206–214. 2017. View Article : Google Scholar
|
|
29
|
Liang X, Ding Y, Zhang Y, Tse HF and Lian
Q: Paracrine mechanisms of mesenchymal stem cell-based therapy:
Current status and perspectives. Cell Transplant. 23:1045–1059.
2014. View Article : Google Scholar
|
|
30
|
Park CM, Kim MJ, Kim SM, Park JH, Kim ZH
and Choi YS: Umbilical cord mesenchymal stem cell-conditioned media
prevent muscle atrophy by suppressing muscle atrophy-related
proteins and ROS generation. In Vitro Cell Dev Biol Anim. 52:68–76.
2016. View Article : Google Scholar
|
|
31
|
Kim MJ, Kim ZH, Kim SM and Choi YS:
Conditioned medium derived from umbilical cord mesenchymal stem
cells regenerates atrophied muscles. Tissue Cell. 48:533–543. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cho KA, Park M, Kim YH, Woo SY and Ryu KH:
Conditioned media from human palatine tonsil mesenchymal stem cells
regulates the interaction between myotubes and fibroblasts by
IL-1Ra activity. J Cell Mol Med. 21:130–141. 2017. View Article : Google Scholar
|
|
33
|
Mouse Genome Sequencing Consortium.
Waterston RH, Lindblad-Toh K, Birney E, Rogers J, Abril JF, Agarwal
P, Agarwala R, Ainscough R, Alexandersson M, et al: Initial
sequencing and comparative analysis of the mouse genome. Nature.
420:520–562. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Naskar S, Kumaran V, Markandeya YS, Mehta
B and Basu B: Neurogenesis-on-Chip: Electric field modulated
transdifferentiation of human mesenchymal stem cell and mouse
muscle precursor cell coculture. Biomaterials. 226:1195222020.
View Article : Google Scholar
|
|
35
|
Zhao Y, Li N, Li Z, Zhang D, Chen L, Yao Z
and Niu W: Conditioned medium from contracting skeletal muscle
cells reverses insulin resistance and dysfunction of endothelial
cells. Metabolism. 82:36–46. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kwon S, Ki SM, Park SE, Kim MJ, Hyung B,
Lee NK, Shim S, Choi BO, Na DL, Lee JE and Chang JW: Anti-apoptotic
effects of human Wharton’s Jelly-derived mesenchymal stem cells on
skeletal muscle cells mediated via secretion of XCL1. Mol Ther.
24:1550–1560. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Girardi F and Le Grand F: Wnt signaling in
skeletal muscle development and regeneration. Prog Mol Biol Transl
Sci. 153:157–179. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
39
|
Comai G and Tajbakhsh S: Molecular and
cellular regulation of skeletal myogenesis. Curr Top Dev Biol.
110:1–73. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cho OH, Mallappa C, Hernández-Hernández
JM, Rivera-Pérez JA and Imbalzano AN: Contrasting roles for MyoD in
organizing myogenic promoter structures during embryonic skeletal
muscle development. Dev Dyn. 244:43–55. 2015. View Article : Google Scholar
|
|
41
|
Zhu X and Lu X: MiR-423-5p inhibition
alleviates cardiomyocyte apoptosis and mitochondrial dysfunction
caused by hypoxia/reoxygenation through activation of the
wnt/β-catenin signaling pathway via targeting MYBL2. J Cell
Physiol. 234:22034–22043. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Majmundar AJ, Lee DS, Skuli N, Mesquita
RC, Kim MN, Yodh AG, Nguyen-McCarty M, Li B and Simon MC: HIF
modulation of Wnt signaling regulates skeletal myogenesis in vivo.
Development. 142:2405–2412. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Drouin G, Couture V, Lauzon MA, Balg F,
Faucheux N and Grenier G: Muscle injury-induced hypoxia alters the
proliferation and differentiation potentials of muscle resident
stromal cells. Skelet Muscle. 9:182019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rahar B, Chawla S, Pandey S, Bhatt AN and
Saxena S: Sphingosine-1-phosphate pretreatment amends
hypoxia-induced metabolic dysfunction and impairment of myogenic
potential in differentiating C2C12 myoblasts by stimulating
viability, calcium homeostasis and energy generation. J Physiol
Sci. 68:137–151. 2018. View Article : Google Scholar
|
|
45
|
Pagé M, Maheux C, Langlois A, Brassard J,
Bernatchez É, Martineau S, Henry C, Beaulieu MJ, Bossé Y,
Morissette MC, et al: CD34 regulates the skeletal muscle response
to hypoxia. J Muscle Res Cell Motil. 40:309–318. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Di Carlo A, De Mori R, Martelli F,
Pompilio G, Capogrossi MC and Germani A: Hypoxia inhibits myogenic
differentiation through accelerated MyoD degradation. J Biol Chem.
279:16332–16338. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Aziz A, Sebastian S and Dilworth FJ: The
origin and fate of muscle satellite cells. Stem Cell Rev.
8:609–622. 2012. View Article : Google Scholar
|
|
48
|
Leroux L, Descamps B, Tojais NF, Séguy B,
Oses P, Moreau C, Daret D, Ivanovic Z, Boiron JM, Lamazière JM, et
al: Hypoxia preconditioned mesenchymal stem cells improve vascular
and skeletal muscle fiber regeneration after ischemia through a
Wnt4-dependent pathway. Mol Ther. 18:1545–1552. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gao T, Yu Y, Cong Q, Wang Y, Sun M, Yao L,
Xu C and Jiang W: Human mesenchymal stem cells in the tumour
microenvironment promote ovarian cancer progression: The role of
platelet-activating factor. BMC Cancer. 18:9992018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chen B, Ni Y, Liu J, Zhang Y and Yan F:
Bone marrow-derived mesenchymal stem cells exert diverse effects on
different macrophage subsets. Stem Cells Int. 2018:8348121. 2018.
View Article : Google Scholar
|
|
51
|
Azhdari Tafti Z, Mahmoodi M, Hajizadeh MR,
Ezzatizadeh V, Baharvand H, Vosough M and Piryaei A: Conditioned
media derived from human adipose tissue mesenchymal stromal cells
improves primary hepatocyte maintenance. Cell J. 20:377–387.
2018.PubMed/NCBI
|
|
52
|
Periasamy R, Surbek DV and Schoeberlein A:
In vitro-microenvironment directs preconditioning of human chorion
derived MSC promoting differentiation of OPC-like cells. Tissue
Cell. 52:65–70. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Nagata M, Iwasaki K, Akazawa K, Komaki M,
Yokoyama N, Izumi Y and Morita I: Conditioned medium from
periodontal ligament stem cells enhances periodontal regeneration.
Tissue Eng Part A. 23:367–377. 2017. View Article : Google Scholar :
|
|
54
|
von Maltzahn J, Chang NC, Bentzinger CF
and Rudnicki MA: Wnt signaling in myogenesis. Trends Cell Biol.
22:602–609. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wang L, Qing L, Liu H, Liu N, Qiao J, Cui
C, He T, Zhao R, Liu F, Yan F, et al: Mesenchymal stromal cells
ameliorate oxidative stress-induced islet endothelium apoptosis and
functional impairment via Wnt4-β-catenin signaling. Stem Cell Res
Ther. 8:1882017. View Article : Google Scholar
|
|
56
|
Guo X, Gu X, Hareshwaree S, Rong X, Li L
and Chu M: Induced pluripotent stem cell-conditional medium
inhibits H9C2 cardiomyocytes apoptosis via autophagy flux and
Wnt/β-catenin pathway. J Cell Mol Med. 23:4358–4374. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Guo X, Chen Y, Hong T, Chen X, Duan Y, Li
C and Ge R: Induced pluripotent stem cell-derived conditional
medium promotes Leydig cell anti-apoptosis and proliferation via
autophagy and Wnt/β-catenin pathway. J Cell Mol Med. 22:3614–3626.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Lim JH, Chun YS and Park JW:
Hypoxia-inducible factor-1alpha obstructs a Wnt signaling pathway
by inhibiting the hARD1-mediated activation of beta-catenin. Cancer
Res. 68:5177–5184. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Rochat A, Fernandez A, Vandromme M, Molès
JP, Bouschet T, Carnac G and Lamb NJ: Insulin and wnt1 pathways
cooperate to induce reserve cell activation in differentiation and
myotube hypertrophy. Mol Biol Cell. 15:4544–4555. 2004. View Article : Google Scholar : PubMed/NCBI
|