Introduction
Chronic obstructive pulmonary disease (COPD) is
becoming a significant cause of mortality and disability worldwide
due to the aging population, the increasing number of smokers and
worsening air pollution (1).
Among the numerous symptoms of COPD, the decline of the respiratory
function is a crucial factor affecting the quality of life of
patients (2), and the respiratory
failure caused by respiratory muscle dysfunction is the most severe
symptom of COPD, given that respiratory failure is closely related
to the mortality rate of patients with COPD. The maximum
inspiratory pressure, which is closely associated with the function
of inspiratory muscle, also represents one of the independent
factors affecting the survival rate of these patients (3). A previous study demonstrated that
the respiratory muscle dysfunction observed in patients with COPD
is caused by chronic increased mechanical load (4). The decline in respiratory muscle
function has also been demonstrated to be induced by multiple
mechanisms, such as chronic inflammatory infiltration (5) and oxidative stress (6), which severely decrease the
ventilation capacity of patients with COPD. The diaphragm, as the
most important source of mechanical power in respiratory movement,
is significantly affected in this process and has also the primary
subject of previous studies (7,8).
In patients with early COPD, the diaphragm may compensate for poor
ventilation (9), although it
becomes gradually damaged by the increasing ventilation load
combined with systemic inflammation (10), oxidative stress (11) and other factors. The decline in
diaphragm muscle strength and endurance in patients with COPD makes
it impossible for these patients to generate enough pressure to
obtain normal ventilation levels, which further worsens respiratory
distress (12). Improving the
diaphragm function of patients with COPD is of great significance
in improving their respiratory status and quality of life.
As the understanding of the significance of
diaphragm function in COPD improves, various interventions have
been proposed to improve diaphragm function; however, only one
intervention was demonstrated to be effective in reversing such
abnormalities, exercise training (13). As an important means of lung
rehabilitation, exercise training is considered the cornerstone of
COPD management. A large number of clinical studies have
demonstrated that the effects of exercise training on the muscle
function of patients with COPD are not only observed in the
skeletal muscles of limbs, but also in the respiratory muscles such
as the diaphragm (14).
Simultaneously, how exercise training induces the improvement of
COPD muscle function has been demonstrated, such as the effect of
long-term aerobic exercise on the improvement of skeletal muscle
stamina, in which fatigue-prone IIB and IIX fibers are transformed
into fatigue-resistant type I fibers, enhancing the mitochondrial
content and activity, as well as the muscle sugar transport
capacity (15). Different types
of exercise training may activate independent signaling pathways
(16), leading to different
skeletal muscle adaptation. Concomitantly, as a complex stimulus on
the body (17), exercise may have
effects on diaphragm dysfunction in multiple ways. Unfortunately,
the existing data are not sufficient to explain the specific
mechanism of exercise-dependent enhancement of diaphragm function.
Therefore, further exploration of the underlying molecular
mechanisms of this improvement in diaphragm function in COPD may
lead to the improvement of lung rehabilitation.
Rapid advances in technology have allowed the
identification of candidate biomarkers associated with different
clinical phenotypes. The emergence of genomics, transcriptomics,
proteomics and metabolomics has provided new methods of exploring
the pathogenesis and therapeutic targets of diseases (18). However, the low concordance
between gene-level and protein-level analysis limits this top-down
mechanism approach (19); in
general, transcriptome data do not generally reflect the true
metabolic level of tissues and organs, which is associated with
multiple complex and undetermined factors that are involved in the
generation of metabolites from genes. Therefore, analysis of
gene-level data cannot fully reveal the specific changes associated
with the physiological and biochemical statuses of a biosystem
relative to phenotype. Fortunately, the emergence of metabolomics
addressed this issue.
Normal physiological processes and external
stimulation cause changes in the types and quantities of
metabolites, which may be quantitatively and qualitatively analyzed
by metabolomics (20). The
physiological and biochemical phenotypes can be determined by
identifying changes in metabolites, and these data can infer the
mechanism of disease progression or potential therapeutic targets.
Therefore, metabolomics has been widely used in disease diagnosis,
medical research and development, nutritional food science and
other fields (21). Considering
that exercise intervention is a complex form of stimulation,
non-target processing in metabolomics could be used to screen a
wide range of different metabolites prior and subsequent to
exercise intervention. Based on existing data (22) and our previous study (23), underwater exercise was considered
to be a more suitable exercise mode for patients with COPD compared
with land activities. Previously, long-term cigarette smoke
exposure (CSE) was considered to be a method to establish a COPD
model with respiratory muscle dysfunction (24-26), and it caused systemic changes in
rats, including oxidative stress (27) and systemic inflammation (28), and also induced a dysregulation of
skeletal muscle protein synthesis and decomposition, resulting in
COPD muscle dysfunction (29). In
the present study, CSE was used to establish a COPD rat model,
while aerobic exercise in water was used as an exercise
intervention method in animal models. The potential mechanism of
action of an exercise intervention on diaphragm dysfunction in COPD
was explored by comparing the changes in pulmonary function,
diaphragm muscle strength and diaphragm differential metabolites
between three groups: A COPD + exercise group (CEG); COPD model
group (CMG); and control group (CG). Gas chromatography tandem
time-of-flight mass spectrometry (GC-TOF-MS) was used to detect the
different metabolites in the diaphragm tissues of rats of the
different groups.
Materials and methods
Animals and management
Healthy male 2-month-old Sprague-Dawley rats with an
average body weight of 200±20 g were purchased from Beijing Vital
River Laboratory Animal Technology Co., Ltd. All the experimental
animals had ad libitum access to food and water in a 12:12 h
light:dark cycle. The temperature in the feeding room was
maintained at 21±2°C and the humidity was stable at 60±5%. The
experimental protocol was approved by the Institutional Animal Care
and Use Committee of Shanghai University of Sport (approval no.
2018026), and was conducted following the Animal Research
Guidelines of the Shanghai University of Sport. During the
experiment, all rats were anesthetized and executed according to
the experimental scheme, and all measures were taken to decrease
the pain of the experimental animals.
Establishment and verification of the
COPD model
A total of 38 rats (Fig. 1) were randomly divided into two
groups: COPD rats (CR; n=24) and CG (n=14). Animals from the CR
group were exposed to cigarette smoke for 16 weeks to establish a
COPD model, while the CG rats were exposed to fresh air and did not
undergo any intervention. The passive smoking poisoning system
(PAB-S200; Beijing Bestlab High-Tech Co., Ltd.) was used to
generate and expose the animals to cigarette smoke during the
experimental process. The method for establishing the model was
based on those previously described (24-26,30,31). The similar total number of
cigarettes, as described by Caron et al (31), was adopted in the present study,
and the modeling duration was shortened to 16 weeks, using
progressive CSE. A total of 20 rats were placed in the passive
smoking poisoning system at a time. The establishment of the model
was completed in 4 stages. The first week was the adaptive modeling
period: For 7 days, the rats were passively exposed to a dose of
smoke from 10 cigarettes once a day for 1 h. During weeks 2-8, the
rats received twice the daily dose of smoking, in which 10
cigarettes were burned during each exposure for 1 h. From weeks
9-12, rats were exposed to smoke twice a day, in which 15
cigarettes were burned each time for 1 h. In weeks 13-16, rats were
exposed to smoke twice a day, in which 20 cigarettes were burned
each time for 1 h. A total of 2 rats died of acute respiratory
tract inflammation during the establishment of the model.
A total of 6 rats in CR were randomly selected and
compared with 6 CG rats to validate the COPD model after 4 months
of smoke exposure. The main parameters of model validation were
pulmonary function tests and histological changes in the lung
tissues. The pulmonary function procedure was as follows: Rats were
weighed and anesthetized using intraperitoneal injection of 10%
chloral hydrate (400 mg/kg body weight; Sinopharm Chemical Reagent
Co., Ltd.) solution. A total of 10 min later, the righting reflex
disappeared and the response to pain stimulus disappeared. Rats
entered a deep anesthesia state without peritonitis symptoms, such
as abdominal muscle tension. Following skin preparation, the neck
skin was cut longitudinally. The subcutaneous tissue and
sternohyoid muscle were separated, and the trachea was exposed. The
trachea was then cut, and the rats were intubated and connected to
the Buxco Pulmonary Function Testing System (Data Sciences
International). Forced vital capacity (FVC), 100 milliseconds
forced expiratory volume (FEV100), functional residual capacity
(FRC), vital capacity (VC) and peak expiratory flow (PEF) were used
to identify the lung function.
Following the pulmonary function test, rats were
euthanized by rapid decapitation under deep anesthesia. The
heartbeat and nerve reflex were then examined to ensure rat death:
Cardiac arrest and loss of the nerve reflex were used as
confirmation of animal death. The lung lobes of the rats were
removed for histological observation to confirm pathological
changes. The lung tissues were fixed in 4% paraformaldehyde for 24
h at room temperature and dehydrated using an increasing ethanol
gradient. Then, the lungs were submerged in xylene, and embedded in
paraffin using the Leica EG1160 paraffin embedding machine (Leica
Microsystems GmbH). Serial sagittal sections (5 µm) were obtained
for histological analyses using the Leica RM2235 paraffin microtome
(Leica Microsystems GmbH). Finally, the lung tissues were stained
with hematoxylin & eosin and observed using a light microscope
(magnification, ×200).
Training protocols
Following model identification, the remaining COPD
rats (n=16) were randomly divided into CEG (n=8) and CMG (n=8). The
CEG rats underwent aerobic exercise intervention for the subsequent
9 weeks, while the CMG and CG rats did not.
The original training program was described by Totou
et al (32), and was
subsequently modified in the present study. Swimming was used as
aerobic exercise, taking place in a tank. The water level was
initially set at 45 cm, and then adjusted according to the growth
of the animals and the water temperature was set at 33-35°C.
Swimming training included a 60 min swimming course. The whole
intervention process was divided into 3 stages. The 17th week was
the adaptive intervention stage. On the first day, swimming
training lasted 10 min, and was then increased for 10 min every day
until it reached 60 min on the 5th day. The training lasted for 6
days. Starting from the 18th week, training was conducted every
other day, 3 days a week. At weeks 18-21, rats under-went swimming
sessions without load, while at weeks 22-25, rats gradually adapted
to a body mass overload of 5%. The present study aimed to set the
intensity of exercise to near the sub-maximum, according to maximal
lactate steady states (6% of body weight for swimming load) noted
in the literature (33).
Stage-by-stage exercise intervention was beneficial for the rats,
to ensure gradual adaption to the intensity of the exercise and to
decrease the adverse effects caused by sudden increases in exercise
intensity. During the experiment, the rats were progressively
challenged with exercises at a higher intensity compared with that
of the exercises they had just become adapted to, as once the rats
adapt to the intensity of the exercise, the effects of the
intervention are not reliable.
Evaluation of pulmonary function and
diaphragm function
In the 26th week, the pulmonary function, as
aforementioned, and diaphragm function were evaluated in all rats.
The isolated diaphragm muscle strength, measured in a tissue
perfusion system (Harvard Apparatus), was used to reflect the
diaphragm function in rats, as described previously (34). Following thoracotomy, 1.5×9.0 mm
diaphragm strips were cut from the edge of a blood vessel
perpendicular to the left side and the middle of the diaphragm and
placed into Krebs-Hanseleit buffer (117 mmol/l NaCl, 4.7 mmol/l
KCl, 2.5 mmol/l CaCl2, 1.2 mmol/l
KH2PO4, 1.2 mmol/l MgSO4, 24.6
mmol/l NaHCO and, 10.1 mmol/l D-glucose) at 37°C. The mixture of
95% O2 and 5% CO2 was used to balance the
perfusion fluid for 15 min prior to perfusion, and the gas flow was
maintained during the perfusion. After a 2 g weight preload
calibration, the diaphragm tissue strips were put into the glass
bath; one end of the strip was fixed at the bottom of the bath and
the other end of the strip was attached to the tension sensor. A
tungsten wire electrode was used to stimulate the muscle strip at
10 V with a square wave of 1 Hz and 2 ms wavelength along the long
axis of the muscle. The diaphragm contraction strength was recorded
using PowerLab 4/35 data acquisition hardware (ADInstruments Ltd.).
The mean values of the 5 maximum peaks of muscle contraction were
measured and corrected by the cross-sectional area (CSA) in
mg/cm2. Following the measurement of diaphragm strength
in vitro, the non-muscle tissue was removed, and the length
of the muscle strip was measured. The water was removed with
absorbent filter paper, and the muscle strip was weighed. CSA was
calculated using the following formula: CSA (mm2)=strip
weight (mg)/[strip length (mm) × muscle density 105.6 (mg/mm)].
The histological observations of the lung and
diaphragm were also performed as aforementioned. In addition, the
lung tissues of the three groups of rats were stained using a
reticulin kit (Genmed Scientifics, Inc.) to further observe the
structure of alveoli. The specific steps are described in a
previous study (35). Briefly,
sections were cut at a 5-µm thickness from routinely processed
paraffin blocks. They were deparaffinized, rehydrated, rinsed in
distilled water, and immersed in 1% potassium permanganate (2 min).
The sections were rinsed in the following solutions: 2.5% oxalic
acid (1 min), 2% iron alum (1 min), Gomori's solution (3 min), 10%
formalin (2 min), gold chloride (1:500; 5 min), 3% potassium
metabisulfite (1 min), and 3% sodium thiosulfate (1 min); sections
were rinsed with distilled water before immersion in each solution.
Then, the sections were dehydrated with a graded series of ethanol,
cleared in xylene, and fixed with neutral gum. The above operations
were performed in a dark room at room temperature.
Sample preparation
The remaining diaphragm muscle tissues of rats were
collected and 50±1 mg diaphragm tissue was extracted for
metabolomic analysis. The samples were placed into 2 ml Eppendorf
(EP) tubes and 450 µl extraction liquid
(VMethanol:VChloroform=3:1) was then added. A total of
10 µl L-2-chlorophenylalanine (1 mg/ml stock in double-distilled
H2O; Shanghai Hengbai Biotech Co., Ltd.) was used as the
internal standard. The mix was vortexed for 30 sec, homogenized in
a ball mill for 4 min at 45 Hz, and ultrasound-treated at 40 kHz
for 5 min. The sample was incubated in ice water during the
ultrasound treatment. Then, the sample was centrifuged at 13,800 ×
g for 15 min at 4°C, and 300 µl supernatant was transferred into a
fresh 1.5 ml EP tube. A total of 50 µl from each sample was taken
and pooled as the QC sample, which was dried completely in a vacuum
concentrator without heating. Then, 40 µl methoxyamination
hydrochloride [20 mg/ml in pyridine; TCI (Shanghai) Development
Co., Ltd.] was added, and the QC sample was incubated for 30 min at
80°C. Afterward, 60 µl BSTFA regent (1% trimethyl chlorosilane,
v/v; Regis Technologies, Inc.) was added to the QC sample, which
was further incubated for 1.5 h at 70°C. A total of 5 µl FAMEs (in
chloroform; Dr. Ehrenstorfer GmbH) was added to the QC sample while
cooling to room temperature. All samples were analyzed by a gas
chromatography system coupled with a Pegasus HT time-of-flight mass
spectrometer (GC-TOF-MS).
GC-TOF-MS analysis
GC-TOF-MS analysis was performed using an Agilent
7890 gas chromatograph system (Agilent Technologies, Inc.) coupled
with a Pegasus® HT time-of-flight mass spectrometer
(LECO Corporation). The system utilized a DB-5MS capillary column
(Agilent Technologies, Inc.) coated with 5% diphenyl cross-linked
with 95% dimethylpolysiloxane (30 m × 250 µm inner diameter,
0.25 µm film thickness; J&W Scientific). A 1 µl
aliquot of the analyte was injected in the splitless mode. Helium
was used as the carrier gas, the front inlet purge flow was 3 ml
min−1, and the gas flow rate through the column was 1 ml
min−1. The initial temperature was maintained at 50°C
for 1 min, and it was then raised to 310°C at a rate of 10°C
min−1 and subsequently maintained at 310°C for 8 min.
The injection, transfer line, and ion source temperatures were 280,
280 and 250°C, respectively. The energy was −70 eV in the electron
impact mode. The mass spectrometry data were acquired in full-scan
mode with the m/z range of 50-500 at a rate of 12.5 spectra/second
following a solvent delay of 6.17 min.
Data preprocessing
Pulmonary function and muscle strength
in vitro
The data distribution was examined by the
Kolmogorov-Smirnov test and homogeneity test of variance (Levene's
test). The normally distributed data are expressed as mean ±
standard deviation, while skewed data are expressed as median
(interquartile range). A one-way analysis of variance was used for
comparisons among multiple groups, and the Least Significant
Difference method was used for further comparison. Dunnett's T3 was
used for multiple comparisons among groups if the variance was not
uniform. P<0.05 was considered to indicate a statistically
significant difference. The software package SPSS v.21.0 (IBM Corp)
was used for statistical analyses.
Metabolomics results processing
Chroma TOF 4.3X software (LECO Corporation) and
LECO-Fiehn Rtx5 database were used for raw peaks exacting, the data
baselines filtering and calibration of the baseline, peak
alignment, deconvolution analysis, peak identification and
integration of the peak area (36). Both the mass spectrum match and
retention index match values were measured during metabolite
identification. Peaks detected in <50% of QC samples or RSD
>30% in QC samples were removed, as described previously
(37).
The original data were preprocessed. The deviation
values were filtered, and the missing values in the original data
were simulated. Finally, the data were standardized using the
internal standard. Following processing, the three-dimensional data
associated with peak number, sample name and standardized peak area
were input into SIMCA v.14.1 software package (MKS Instruments,
Inc.) for the orthogonal projection of principal component analysis
(PCA) and orthogonal projections to latent structures-discriminant
analysis (OPLS-DA). PCA revealed the internal structure of data.
Concomitantly, OPLS-DA was used to improve visualization and for
follow-up analysis of the differential variables identified in the
large metabolomics dataset. In addition, univariate statistical
analysis was used for data analysis to avoid false-positive errors
or model over-fitting caused by using only one type of statistical
analysis. The variable importance for the projection (VIP) >1.0
and P<0.05 were used as the cut-off values for significantly
different metabolites.
Differential metabolites obtained from the analysis
were identified to be biologically similar or complementary in
functions, or regulated positively or negatively by the same
metabolic pathway, showing either similar or opposing expression
profiles among the different experimental groups. Therefore, a
hierarchical cluster analysis of the differentiated metabolites was
performed using Euclidean distance matrix (MATLAB 2018b; https://www.mathworks.com/), as described previously
(38). Moreover, the annotations
identified from the Kyoto Encyclopedia of Genes and Genomes
(39,40) (KEGG; https://www.genome.jp/kegg/) were analyzed, and the
metabolic pathways of differentiated metabolites were further
analyzed using MetaboAnalyst 3.0 (https://www.metaboanalyst.ca).
Results
Effect of 16-week CSE on pulmonary
function and tissue structure in rats
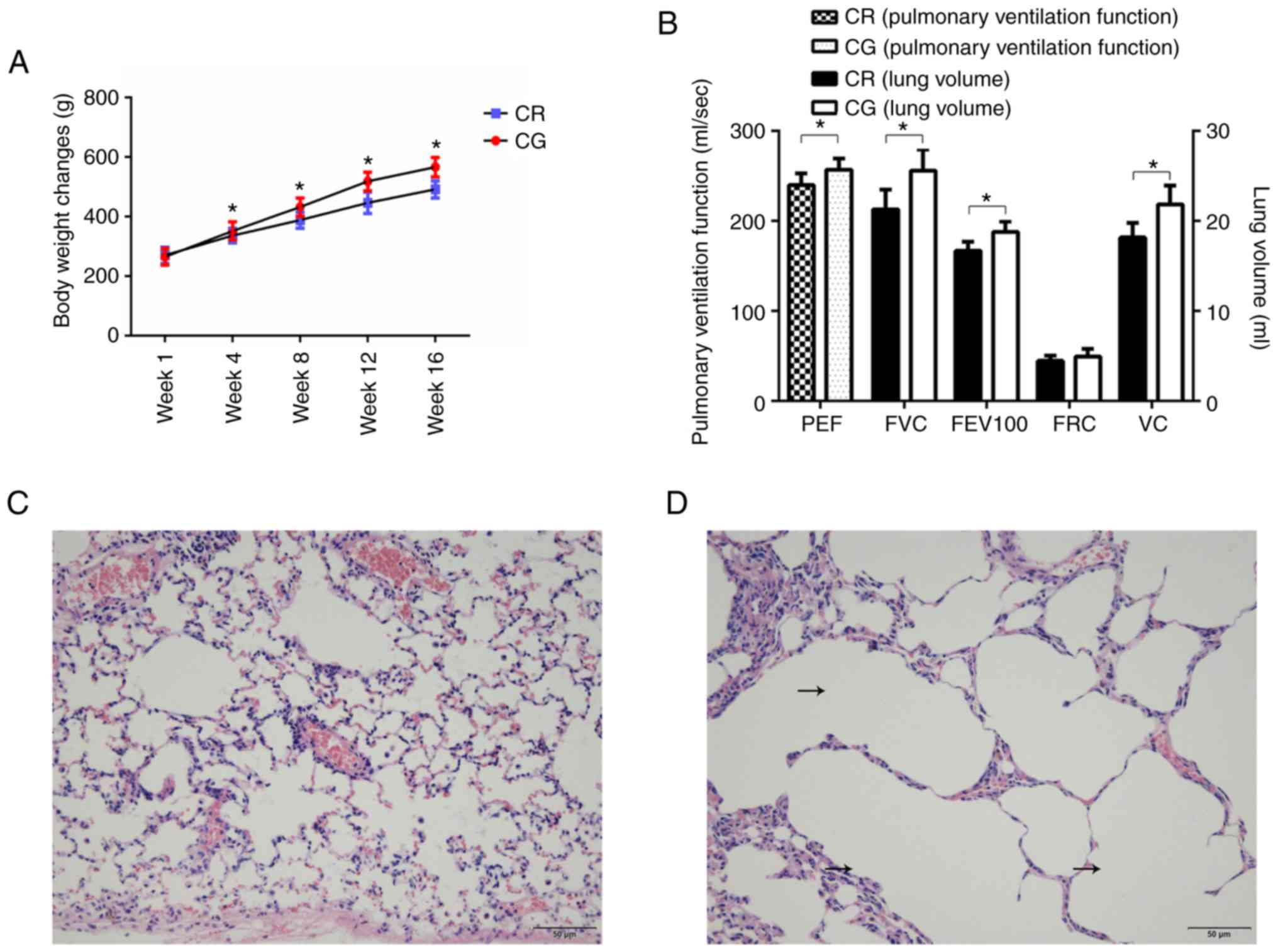
The results of model verification demonstrated that
there were significant differences in lung function and structure
between the COPD rats and control rats, which suggested that the
model was successfully established (Fig. 2). The 16-week model-building
process inhibited weight gain of the model rats (Fig. 2A). The pulmonary ventilation
function of model rats decreased significantly, including PEF,
FEV100 and FVC (Fig. 2B).
Concomitantly, the histological examination demonstrated that,
compared with the control rats (Fig.
2C), the pulmonary structure of the model rats was
reconstructed: the alveoli were enlarged, and fusions between the
terminal bronchus and the alveoli were observed (Fig. 2D).
Changes in pulmonary function and
histology following exercise
The effects of the CSE treatment on the respiratory
tract and the general condition of rats were different between the
treatment groups. During the entire study, the animals in the CG
were allowed to move freely for an extended period of time, with
good breathing conditions, a smooth coat and gradual weight gain.
The COPD model animals (CEG and CMG) were affected by cigarette
smoke during the modeling period (0-16 weeks). Their fur was dry
and yellowish, and they exhibited symptoms of respiratory
discomfort, including shortness of breath, hypersonic breathing and
increased secretions of the respiratory tract. The weight of the
model animals increased slowly. During the intervention period
(17-25 weeks), the respiratory symptoms of CEG and CMG gradually
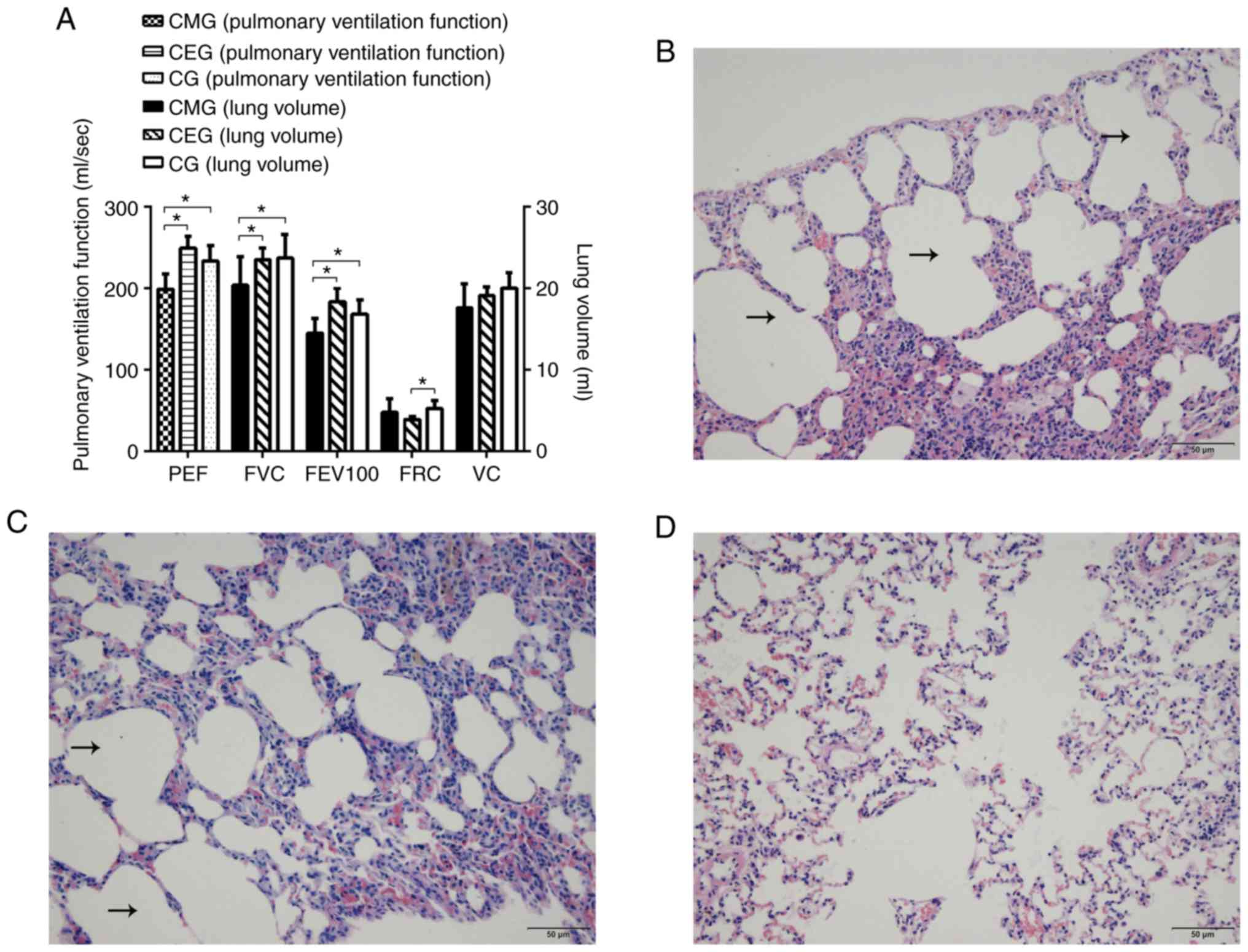
disappeared, but the weight gain of CEG rats was slow. In addition,
the 9-week swimming intervention significantly improved the lung
function in the COPD rats: Compared with the CMG, the pulmonary
function of CEG was significantly improved (P<0.05), and there
were increases in FVC, FEV100 and PEF values. Furthermore, the
difference between CEG and CG was primarily reflected in the
decrease of FRC (P<0.05; Fig.
3A).
Histological staining of lung tissue demonstrated a
significant difference among the three groups following
intervention. In the CMG, alveolar dilatation, narrowing and
rupture of alveolar septum were observed under microscopy, and the
adjacent alveoli had fused into larger cysts (Fig. 3B). The morphological structure of
lung tissue in CEG was similar to that in the CMG (Fig. 3C). In the CG, the rats exhibited
healthy lung tissue structure with uniform alveolar size and within
a reasonable range (Fig. 3D). As
it was not possible to determine under the microscope whether the
dark stained sections were aggregated inflammatory cells, the
effect of an exercise intervention on the inflammatory response
could not be determined under the microscope. The reticulin stained
results are presented as supplementary materials (Fig. S1).
Effects of aerobic exercise on diaphragm
muscle strength and histology in rats
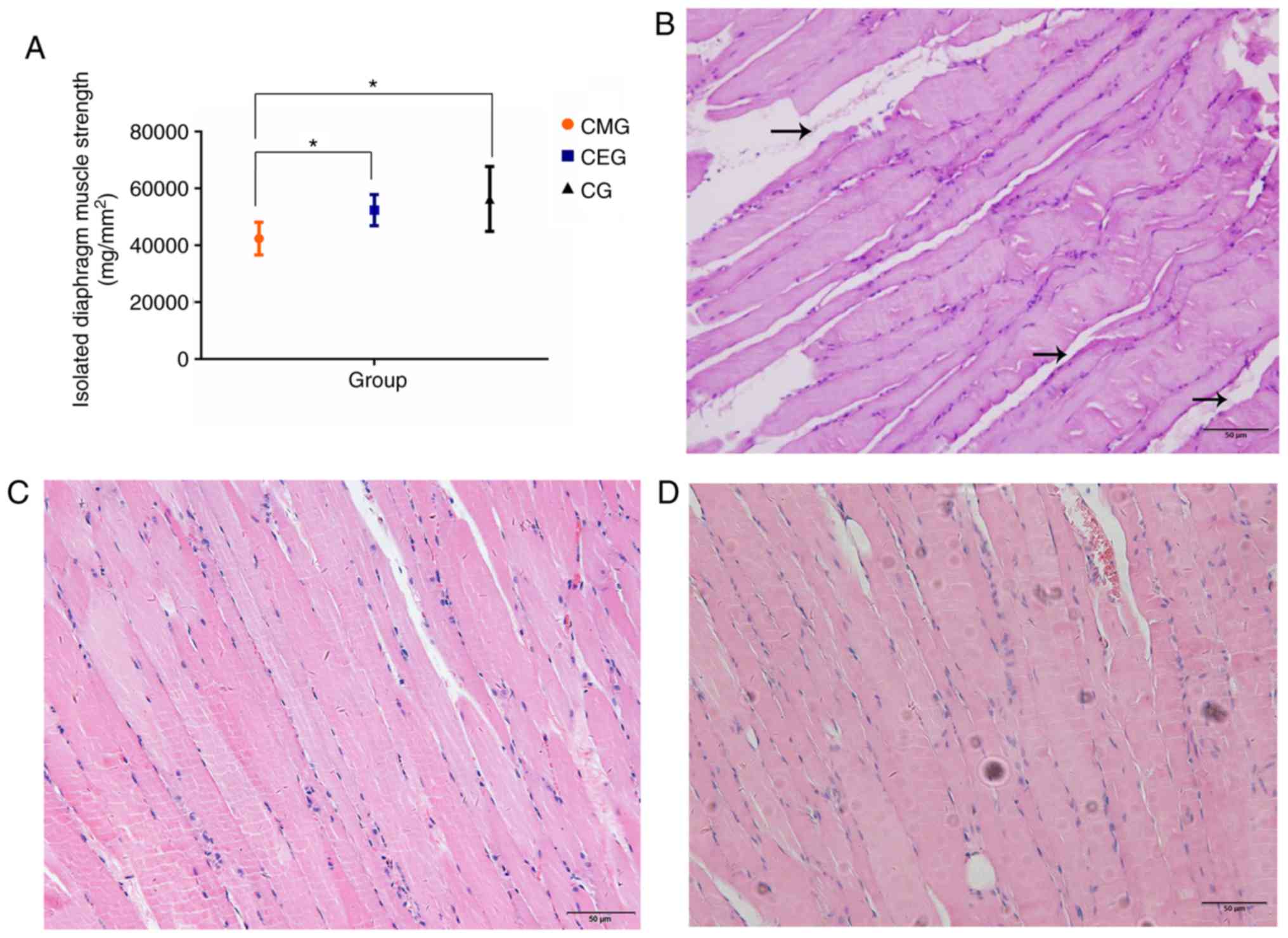
The measurement of the diaphragm muscle strength
in vitro closely reflected the contractile function of the
diaphragm, and the results of the present study indicated that
aerobic exercise improved the diaphragm function of COPD rats. It
was demonstrated that the diaphragm muscle strength of the CMG was
significantly weaker compared with that of the CG (P<0.05), and
the diaphragm muscle strength of CEG rats was stronger compared
with that of the CMG (P<0.05). There was no significant
difference in diaphragm muscle strength between the CEG and CG
(Fig. 4A). The diaphragms of the
three groups of rats exhibited different histological morphologies.
In the CMG, the structure of muscle fibers in the longitudinal
section of the diaphragm was not uniform, and the muscle fibers
were not close together (Fig.
4B). The proliferation of interstitial fibers could be observed
in certain visual fields. In turn, the muscle fibers in the CEG
(Fig. 4C) and the CG (Fig. 4D) were arranged neatly and
tightly, and no interstitial fibers and capillary proliferation
were observed.
Multivariate analysis of the metabolomics
data
Of the 24 samples, 742 peaks were extracted from the
original data. Following data preprocessing, 599 peaks remained.
The results of the multivariate pattern recognition analysis
indicated that the data were reliable. Firstly, from the results of
the PCA score chart, all the samples were in the 95% Hotelling's
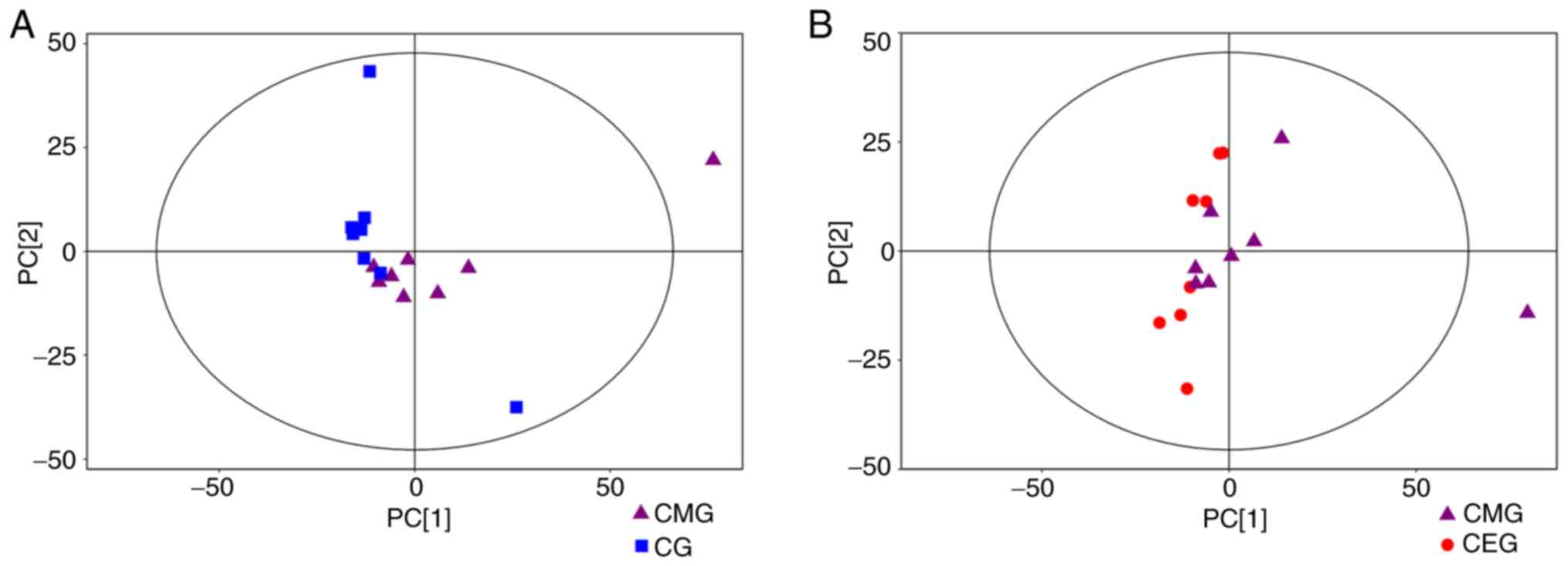
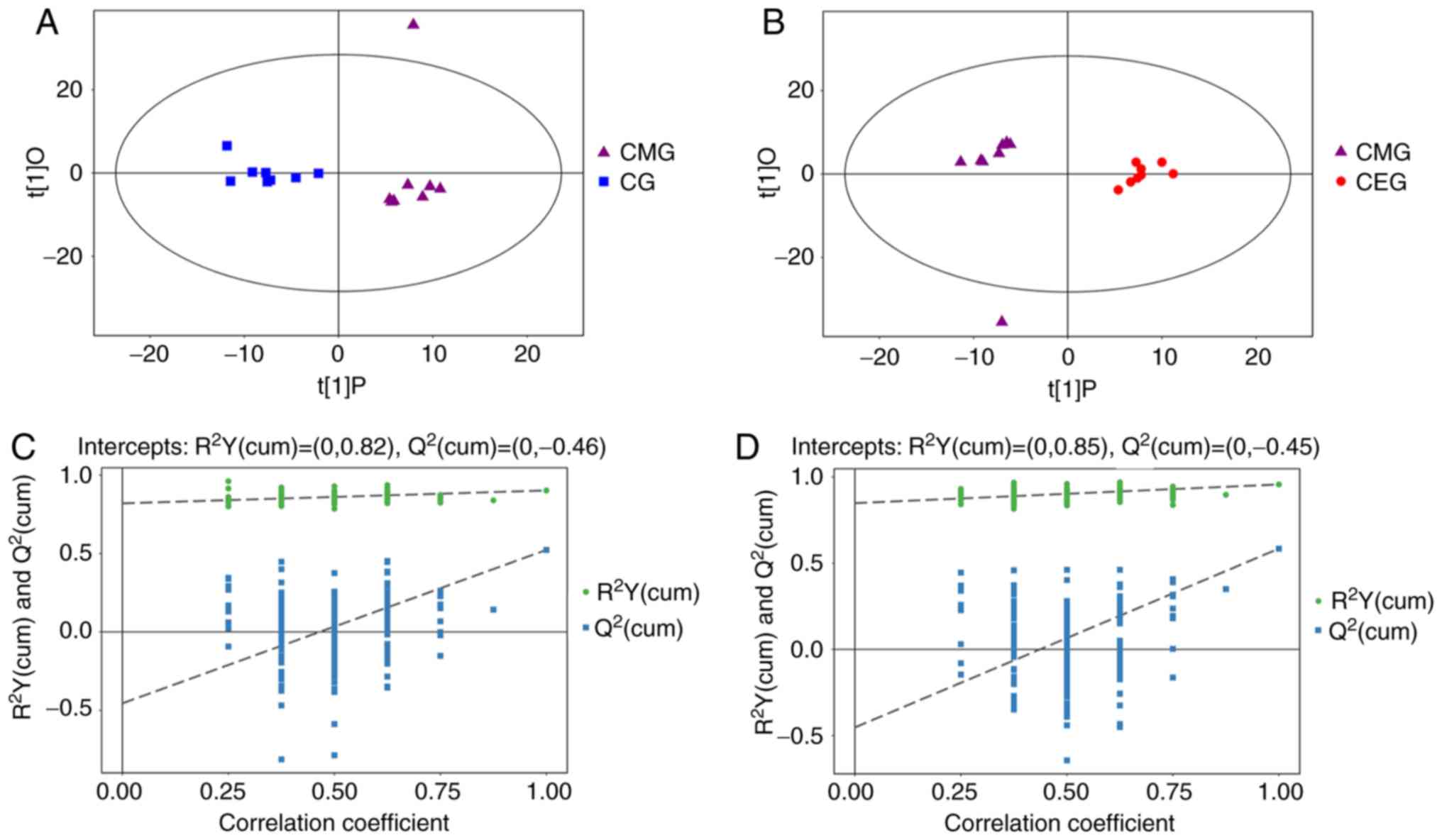
T-squared ellipse, with the exception of one outlier (Fig. 5). Secondly, OPLS-DA was used to
analyze the model (Fig. 6) to
obtain more reliable information about the correlation between the
significantly different metabolites and the experimental groups.
The OPLS-DA score plot demonstrated significantly separated
clusters between the three different groups, although a sample from
the CMG was not in the 95% Hotelling's T-squared ellipse in 2
comparisons (Fig. 6A and B).
Compared with the CG, the two evaluating parameters of the OPLS-DA
model were R2Y= 0.902 and Q2= 0.523. Compared
with CMG, the parameters were R2Y=0.956 and
Q2=0.584. Fig. 6C
demonstrates the results from 200 permutation tests, with intercept
R2=(0, 0.82) and Q2=(0, −0.46) in the
comparison of the CMG and the CG, while Fig. 6D shows the permutation test
results of the comparison between the CEG and CMG, with intercept
R2=(0, 0.85) and Q2=(0, −0.45). These results
confirmed that the original model had good robustness and that
there was no over-fitting phenomenon, indicating that the model was
appropriate for follow-up analysis. Therefore, the outlier was
retained in subsequent analysis, given that the results of
multivariate analysis were stable and reliable.
Biomarkers associated with exercise
intervention
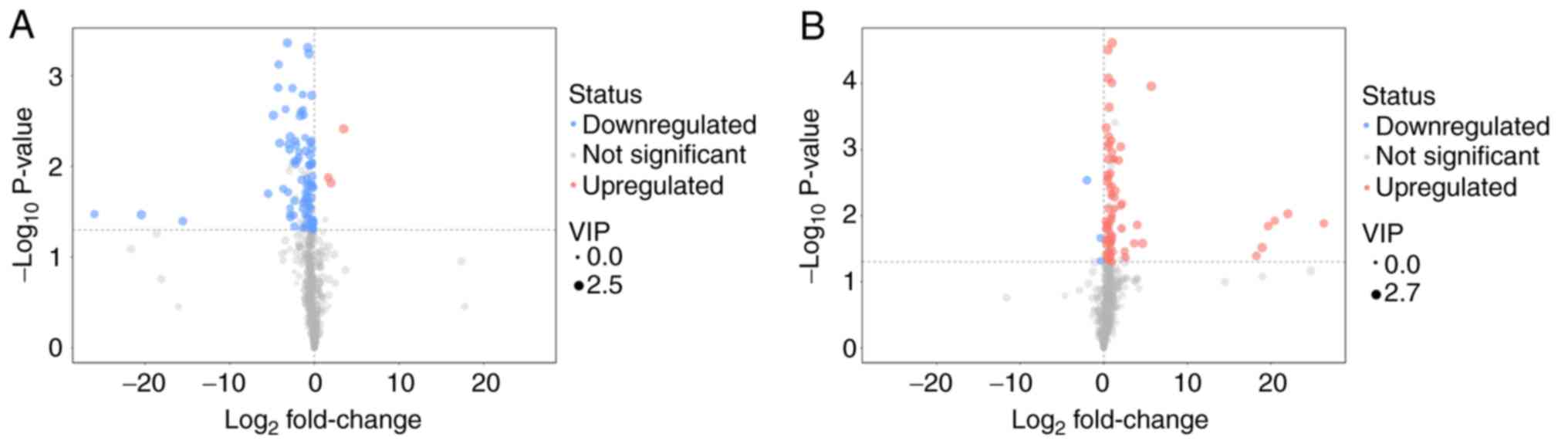
The distribution of all differential metabolites was
grouped into upregulated and downregulated metabolites. Compared
with the diaphragm metabolites of the CMG and the CG, 85 different
metabolites were screened out, of which 82 metabolites were
identified as downregulated. In the comparison between CEG and CMG,
80 different metabolites were identified, 77 of which were
identified as upregulated. The screened differential metabolites
are presented in the form of volcano plots (Fig. 7). Compared with the CG, excluding
unknown metabolites, 36 metabolites in the diaphragm of the CMG
were identified to be significantly different. In addition,
compared with the CMG, the expression of 29 metabolites in the
diaphragm of rats in CEG was significantly different following
exercise intervention. To identify any candidate biomarkers
associated with exercise training, the differentially expressed
metabolites that exhibited contrasting expression profiles between
the CMG and the CG, and between the CEG and the CMG, were selected.
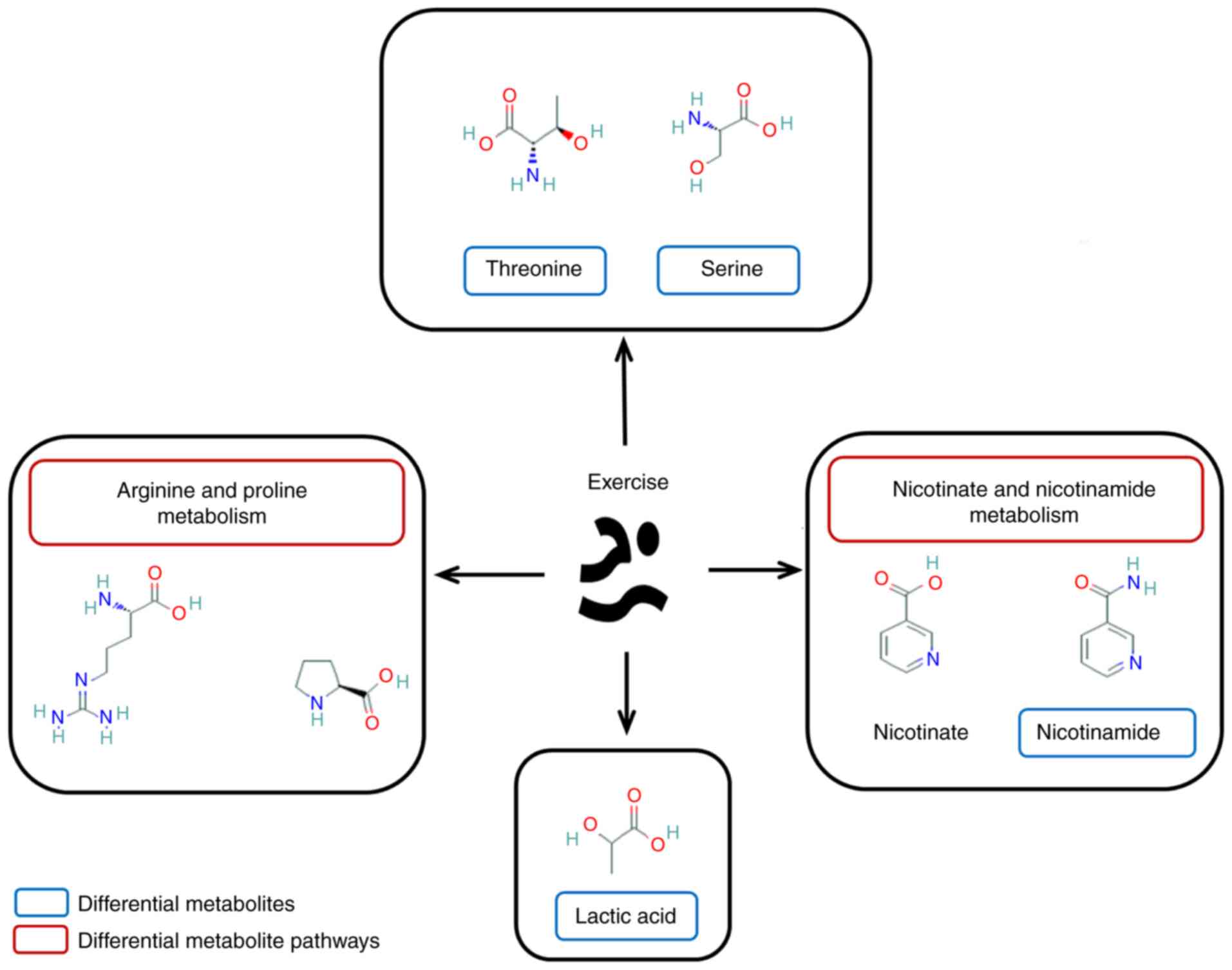
A total of 8 metabolites were identified (Table I), including threonine, serine,
nicotinamide, lactic acid, citraconic acid, 3-cyanoalanine, dioctyl
phthalate and 2-hydroxypyridine.
 | Table IMetabolites with contrasting
expression profiles between CMG/CG and CEG/CMG groups. |
Table I
Metabolites with contrasting
expression profiles between CMG/CG and CEG/CMG groups.
| Metabolites | Trends in
CMG/CG | Changing trends in
CEG/CMG | Fold-change in
CMG/CG | P-value in
CMG/CG | Fold-change in
CEG/CMG | P-value in
CEG/CMG |
|---|
| Threonine | Down | Up | 0.778 | 0.005 | 1.569 | 0.001 |
| Serine | Down | Up | 0.275 | 0.007 | 1.433 | 0.001 |
| Nicotinamide | Down | Up | 0.672 | 0.028 | 1.504 | 0.022 |
| Lactic acid | Down | Up | 1.507 | 0.033 | 83147489.53 | 0.013 |
| Citraconic
acid | Down | Up | 0.808 | 0.039 | 1.426 | 0.001 |
|
2-Hydroxypyridine | Down | Up | 0.814 | 0.006 | 1.700 | 0.002 |
| 3-Cyanoalanine | Down | Up | 0.656 | 0.009 | 1.451 | 0.002 |
| Dioctyl
phthalate | Down | Up | 0.022 | 0.019 | 24.811 | 0.026 |
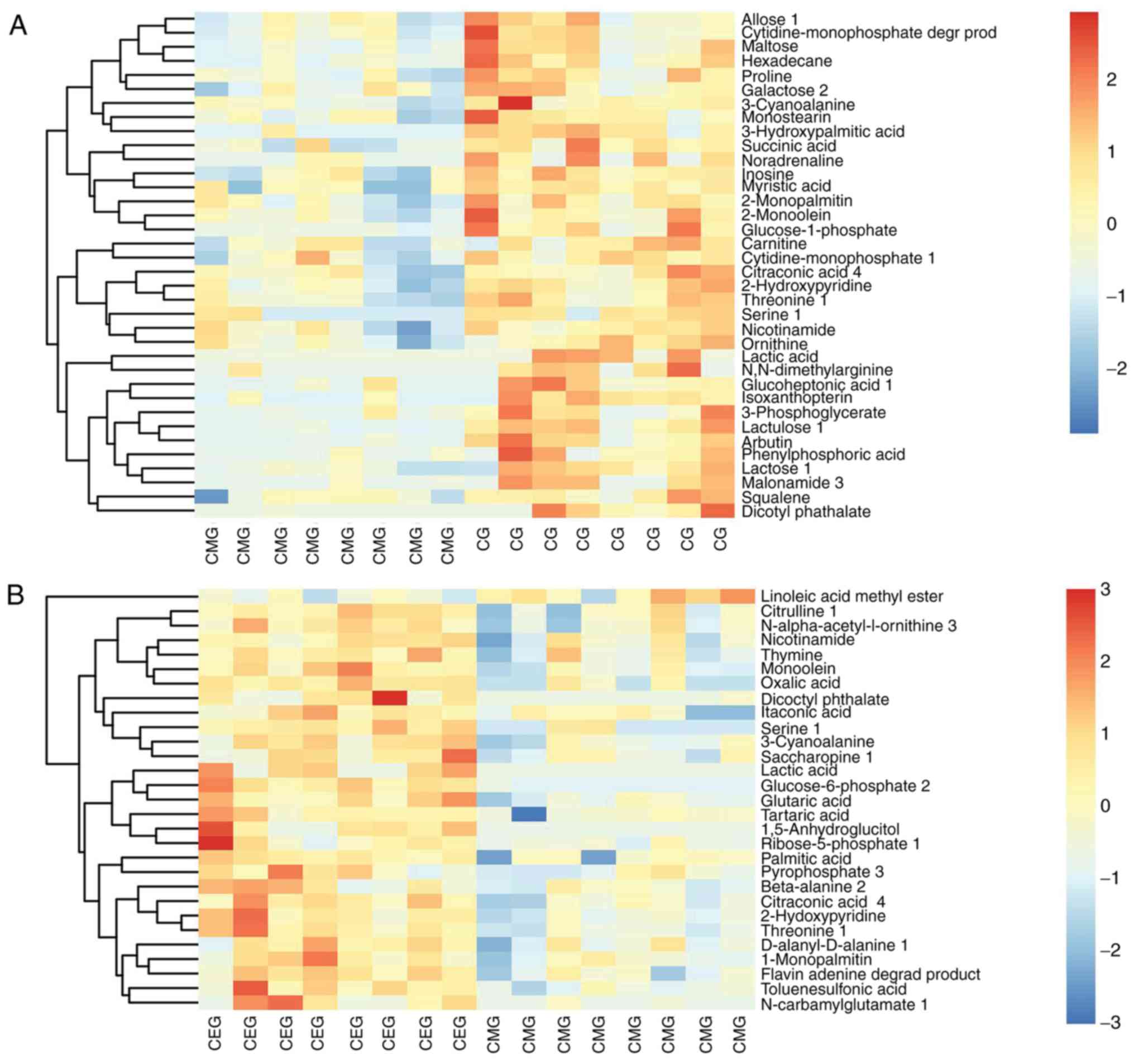
Pathway analysis of biomarkers
The Euclidean distance matrix was used for
differential metabolite analysis, to explore the correlation
between the different metabolites identified and to explore the
characteristics of metabolite changes among experimental groups
(Fig. 8). In addition, KEGG
annotation of differential metabolites and pathway analysis of
differential metabolites were employed for further analysis,
considering the complex metabolic reactions and associated
regulatory mechanisms. When the metabolites different in CMG and CG
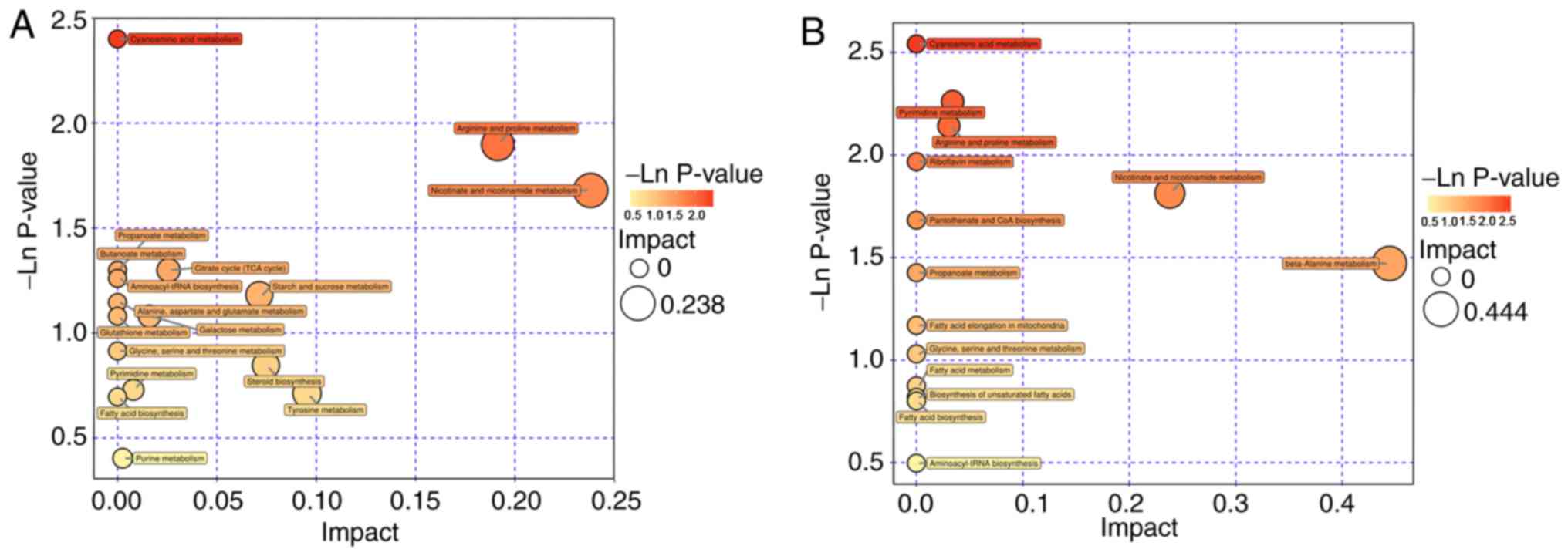
were analyzed using MetaboAnalyst, 17 metabolite pathways were
identified to be significant. Based on their-ln (P-value) and
pathway impact scores [-ln (P-value) >1 and impact scores >0]
and the biological roles of metabolic pathways in associated
fields, five pathways were selected. These were: Arginine and
proline metabolism; nicotinate and nicotin-amide metabolism;
tricarboxylic acid cycle; starch and sucrose metabolism; and
galactose metabolism (Fig. 9A). A
total of 4 metabolite pathways were identified to be significantly
different between CEG and CMG: Pyrimidine metabolism; arginine and
proline metabolism; nicotinate and nicotinamide metabolism; and
beta-alanine metabolism (Fig.
9B). Notably, the metabolic pathways that were identified in
both comparisons deserve further discussion. The substances in
these pathways may contribute to the occurrence of diseases, but
they could also be involved in the potential mechanisms underlying
the exercise-based improvement of the condition. Considering the
source and function of these metabolites, the exercise-dependent
enhancement of diaphragm function in COPD rats is potentially
mediated by these shared pathways, namely, the nicotinate and
nicotinamide metabolism, and arginine and proline metabolism
pathways. In addition, the glycine-serine-threonine metabolism,
fatty acid biosynthesis and metabolism-associated pathways also
appeared in 2 comparisons, even though the impact scores of these
pathways were not high (Fig.
10).
Discussion
Changes in pulmonary function and
histology in COPD rats following exercise
Exercise intervention inhibited the adverse effects
of respiratory tract exposure to smoke (41,42) and improved the general condition
of rats (43). Considering the
association between the modeling process and self-repair and the
smoking habits of some patients with COPD [long-term smoking and
difficult to quit smoking (44,45)], a progressive long-term exposure
to cigarette smoke was adopted. Rodents have a demonstrated ability
for self-repair (46).
Maintaining the intensity of a constant stimulus results in a
gradual increase in tolerance and tissue repair (47,48). Incremental exposure to cigarette
smoke decreased the effect of self-repair in rats (49,50). Concomitantly, long-term
incremental exposure to cigarette smoke also imitates the smoking
habits of patients with COPD, compounding the effects of cigarettes
on the disease (30,51). Following smoking cessation, the
changes in the respiratory symptoms in the CMG reflected the effect
of self-repair, while the recovery process in the CEG rats may have
benefited from the exercise intervention; the change of pulmonary
function results in this group reflected this hypothesis. In
addition, in the entire cohort of rats, exercise elicited an effect
on the rate of weight gain, revealing that the effect of exercise
on the body is multifaceted.
To evaluate the effect of exercise on pulmonary
function in COPD rats, the present study primarily focused on the
improvement of ventilation function. As the gold standard of COPD
diagnosis, the improvement in the pulmonary function has a
significant effect on the prognosis of COPD. Clinical pulmonary
function examination is primarily divided into two aspects; namely,
the examination of lung volume and the examination of pulmonary
ventilation function (52). The
abnormality of pulmonary function in patients with COPD is
reflected in both pulmonary volume and pulmonary ventilation
function (53). In patients with
COPD, excessive inflation of the lungs results in an increase in
the residual volume, which leads to a marked decrease in the
ability of inhalation. In pulmonary function tests, abnormal lung
capacity was observed, such as decreased complementary expiratory
volume, increased residual volume or decreased vital capacity.
Airway obstruction, such as decreased ventilation function, is the
primary symptom in the lungs of patients with COPD. Abnormal
ventilation function can be manifested as a decrease of FVC, forced
expiratory volume in the first second or PEF, or an increase of FRC
(54).
The results of the pulmonary function tests
indicated that FVC, FEV100 and PEF, as indexes of pulmonary
ventilation function, were significantly different in the three
groups. The difference between the CMG and the CG indicated that
the pulmonary ventilation function of CMG did decrease following
exposure to cigarette smoke, which was one of the successful
manifestations of the establishment of the model. Meanwhile, the
difference between the CEG and CMG groups indicated that the
ventilation function of the rats was significantly improved
following exercise intervention, particularly the FRC and PEF
indexes. These 2 indicators reflected the important index of
respiratory muscle strength, which also implied that aerobic
exercise in water effectively exercised the respiratory muscles,
enhanced muscle strength and improved pulmonary ventilation
function. However, no significant difference in the lung volume VC
index observed between the three groups. The effects of lung volume
are associated with the changes in the physiological structure of
lung tissue caused by different diseases, and also factors such as
body weight and body shape (55).
After 16 weeks of smoke exposure and 9 weeks of exercise
intervention, the body shape of the rats in different groups
changed significantly. The body weight of rats in the CEG was
decreased compared with that of the CMG rats, which is likely to
affect certain lung volume indexes, such as VC.
In the present study, the mechanism of
exercise-dependent improvement of the lung function in rats with
COPD was not fully investigated. Results of the histological
analyses indicated that there were no specific differences in the
structures of the lung tissues between the CEG and CMG. Both groups
exhibited the pathological characteristics of COPD, which suggested
that the 9-week exercise intervention did not significantly reverse
the pathological changes of lung tissue in COPD rats. It should be
noted that the levels of inflammation in these sections were not
analyzed in the present study. Although a previous study has
confirmed that exercise does regulate inflammation in COPD airways
and lung tissues (56), the
markers of inflammation were not investigated and therefore it is
not able to be concluded. In addition, although some studies have
demonstrated that aerobic exercise reversed the airway remodeling
in asthmatic mice (57,58), the results of the present study
suggested that the benefits of exercise on COPD were more
significant in terms of respiratory muscle strength, exercise
capacity and exercise-induced dyspnea, which is consistent with the
conclusions of other previous studies (59-61).
Effects of exercise on the diaphragm of
rats with COPD
A marked difference in the diaphragm function of the
different groups was observed. The contractility of the diaphragm
tissues in vitro was measured as a proxy for the muscle
function (62), and the
corresponding results indicated that the COPD rats exhibited a
decline in diaphragm muscle strength. For patients with COPD, the
respiratory muscles are exposed to an inhalation load for an
extended period of time, and the diaphragm has to be in an active
state to ensure survival, so it is less affected compared with the
lower limb muscles. However, with the advances in COPD diaphragm
research, the impairment of diaphragm function in these patients
has been confirmed, and the associated pathological process has
been gradually clarified (63).
Due to a change in thoracic geometry, the diaphragm can be placed
at an adverse contraction length. Concomitantly, the increasing
ventilation load, exacerbations, nutritional abnormalities and
aging would make the diaphragm undergo a compensatory period by
functioning faster (12), which
was also the final result observed in many diaphragms in the
compen-satory period (64,65).
Histological analysis of the model rats demonstrated a morphology
similar to muscle atrophy, which was consistent with previous
clinical studies (66,67). Changes in muscle structure
affected the normal function of the muscle. The comparison of
diaphragm function and structure between the CEG and the CMG
indicated that exercise significantly improved the diaphragm
function, which was also consistent with previous studies (68,69). Although other studies (70,71) have indicated that physical
exercise or respiratory re-education increased the strength of
respiratory muscles, the effect of specific muscle training has
been widely debated (72). In
addition, the effectiveness of exercise training observed in the
present study does not indicate that all types of exercise have
beneficial effects on the diaphragms of patients with COPD. The
ventilation load caused by conventional exercise intensity is
insufficient to induce training adaptation (73). To decrease dyspnea and improve
exercise ability, a specific ventilation load should be imposed on
the respiratory muscles to improve the respiratory muscle function
in COPD. Compared with land-based exercise, aquatic aerobic
exercise has better suitability and acceptability for moderate to
severe COPD (22,74). Simultaneously, when exercising in
water, the effects of water pressure and other factors ensure that
the respiratory muscles and thorax of patients with COPD undergo
stronger load stimulation, and these factors promote training
adaption. In combination with the results concerning the effects of
exercise on pulmonary function, these data suggest that exercise
may serve an essential role in improving diaphragm function.
Biomarkers for improving diaphragmatic
function by exercise
Multivariate statistical analysis ensured the
validity of biomarker screening. Firstly, the PCA results
demonstrated that, following dimensionality reduction, the
principal components of the three groups of samples were quite
different compared with when they were analyzed in pairs. Secondly,
the OPLS-DA scatter plot results indicated that the samples in the
different groups were significantly different, while the
permutation test results excluded the possibility of false-positive
data. The biomarkers screened from the established model had
relatively high reliability. Although there was an outlier in the
PCA and OPLS-DA data, the OPLS-DA model had good validity, and the
physiological indexes (pulmonary function and diaphragm
contractility) and histological data of this rat were not abnormal.
Therefore, this sample was not eliminated in the subsequent
analysis.
The improvement of diaphragm function by exercise
was associated with the exercise-dependent promotion of the
expression of various metabolites. The volcano maps indicated that
the majority of the significantly different metabolites in the CMG
were downregulated compared with the CG, while the majority of the
metabolites with significant differences were upregulated following
the exercise intervention. Concomitantly, the decrease in diaphragm
function in the COPD rats was likely to be associated with the
downregulation of certain metabolites in the diaphragm. Following
exercise training, the improvement of diaphragm function in the CEG
rats was identified to be associated with the upregulation of a
large number of metabolites in the diaphragm. Differential
metabolites involved in both comparisons may be biomarkers
associated with the exercise-dependent enhancement of diaphragm
function. A total of 8 metabolites were identified: Threonine;
serine; nicotinamide; lactic acid; citraconic acid; 3-cyanoalanine;
dioctyl phthalate; and 2-hydroxypyridine.
Threonine is an essential amino acid for the human
body. It is an important nutrient fortifier, which decreases
fatigue and promotes growth and development. Muscle growth is known
to be regulated by signal transduction pathways, which sense and
compute local and systemic signals and regulate various cellular
functions. Threonine, which is phosphorylated by phosphokinase,
activates the mTOR signaling pathway, regulates the function of
insulin-like growth factor-1/mechano growth factor/insulin and/or
mechanical signals, amino acids and muscles, and correspondingly
regulates protein synthesis to promote muscle growth (75). It has been suggested that exercise
improved the level of threonine in vivo (76). In the present study, the increase
in threonine levels in the diaphragms of the CEG rats and the
decrease in threonine levels in the diaphragms of the CMG rats
suggested that the improvement of the function of the diaphragm by
exercise may be associated with the regulation of threonine
levels.
Serine also serves a vital role in the human body.
Serine is involved in the formation of cell membranes and the
synthesis of myoprotein. It has been demonstrated that the carrier
proteins serine incorporator 1-5 incorporates a polar amino acid
serine into the membrane and promotes the synthesis of 2
serine-derived lipids, namely phosphatidylserine and sphingolipid
(77). Pantaleo et al
(78) suggested that the
phosphorylation of serine residues is involved in the construction
of cell membranes, based on the study of the erythrocyte membrane.
A previous study has demonstrated that serine participates in the
regulation of the mTOR pathway on skeletal muscle protein
synthesis: Phosphorylation of serine residues on eukaryotic
translation initiation factor 4G promotes muscle protein synthesis,
and this process is regulated by the mTOR pathway (79). Exercise increases the levels of
serine phosphorylation in the body, while supplementation of serine
enhances exercise ability (80).
Considering the results of the present study, the decline in
respiratory muscle function in the COPD rats may be associated with
the decrease in serine expression, and the increase in serine
levels following exercise is also a potential mechanism for
improving respiratory muscle function.
Nicotinamide is the amide form, and the primary
form, of nicotinic acid. It is also the precursor of human
synthetic coenzyme I dihydrouracil dehydrogenase (NAD+) and
coenzyme II nicotinamide-adenine dinucleotide phosphate (NADP),
which are key molecules widely present in cellular signaling
pathways regulating cell metabolism and homeostasis. The presence
of nicotinic acid is significant in regulating the level of
oxidative stress in vivo. It inhibits phosphatidylserine
externalization and late DNA damage in a unique way during
oxidative stress (81). It is
thought that nicotinamide-based protein secretion regulation
removes reactive oxygen species and protects against oxidation. In
addition, nicotinamide also regulates cell inflammation and
inhibits inflammatory mediators (82,83) such as interleukin (IL)-1β, IL-6,
IL-8, transforming growth factor β-2 and macrophage chemotactic
protein-1 in liver cells. Nicotinamide also inhibits the production
of tumor necrosis factor (83).
Therefore, nicotinamide was demonstrated to participate in cellular
metabolism processes and metabolic diseases, and it serves an
important role in maintaining normal cell activities (84). Both intra- and extrapulmonary
injuries in COPD are considered to be associated with oxidative
stress and systemic inflammation. Whether this exercise-mediated
elevation affects oxidative stress and inflammation through
nicotinic acid requires further investigation.
Lactic acid is the intermediate product of
carbohydrate metabolism in the anaerobic state and has been
considered to be closely associated with exercise fatigue (85). Following ≥1 intensive training
sessions, a large amount of lactic acid is produced, and
H+, produced by lactic acid dissociation, will decrease
the environmental pH value, thereby inhibiting the excitability of
muscle fibers and decreasing muscle contraction ability. However,
follow-up studies have disputed the causal association between
lactic acid-acidosis and muscle fatigue, and additional associated
data demonstrated that lactic acid was a key component of local and
global metabolism (86). It was
identified that lactic acid elicits an anti-fatigue effect, and can
be used as a 'mobile fuel' for other tissues and muscle fibers to
maintain the blood sugar homeostasis (87). Lactic acid also maintains ATP
production for cellular metabolic demands by stabilizing the redox
potential and decreasing the difference in the H+
concentration gradient inside and outside active muscle cells,
decreasing K+ release (88). In addition, lactic acid was
suggested to have a certain antioxidant effect (89) and was associated with tissue
damage repair (90). In the
present study, the results indicated that the level of lactic acid
in the diaphragm of the model rats decreased compared with the
control rats, while the expression of lactic acid in the diaphragm
of the exercise rats increased, which may involve a variety of
roles for lactic acid. Future studies should be performed to
elucidate these roles.
3-cyanoalanine belongs to the class of organic
compounds known as the l-alpha-amino acids (91). Citraconic acid, also called
2-methylmaleate or methylmaleic acid, belongs to the class of
organic compounds known as methyl-branched fatty acids (92). 2-hydroxypyridine is used in
peptide synthesis (93). To the
best of our knowledge, there have only been a few studies on these
3 substances in COPD and exercise-induced human metabolite changes,
and further research is required. Dioctyl phthalate does not serve
a role in the metabolism of mammals. Although this compound was
qualitatively identified in the present study, the specific level
of dioctyl phthalate and the reason for its appearance require
further examination.
Biomarker pathway analysis of improving
diaphragm function through exercise
The mechanism by which exercise improves diaphragm
function may involve multiple metabolites and associated metabolic
pathways. Pathway analysis of differential metabolites is useful in
analyzing the specific regulation mechanism of exercise. By
studying the pathways of different metabolites between different
groups, the present study identified that the pathways involved in
the exercise-dependent improvement of diaphragm function are most
likely the nicotinate and nicotinamide metabolism and arginine and
proline metabolism pathways. Notably, combined with previously
published data, the glycine-serine-threonine metabolism and fatty
acid biosynthesis and metabolism pathways may also be associated
with the exercise-dependent enhancement of the diaphragm function.
The nicotinate and nicotinamide pathways are potentially part of
the mechanism by which exercise enhances diaphragm function.
Nicotinate is oxidized by tryptophan and converted to nicotinamide
in the small intestinal mucosa, which eventually binds to proteins
to form NAD+ and NADP. A previous study demonstrated that moderate
exercise accelerates nicotinic acid metabolism in vivo
(94). The effects of exercise on
the expression of NAD+ and NADP in vivo has been an ongoing
topic of investigation (95). The
results of the present study are also consistent with these
studies: The training effect of exercise on the diaphragm may be
associated with the local changes in the nicotinamide metabolic
pathway.
The arginine and proline metabolic pathway serves a
crucial role in the formation of cytoplasm and nucleic acid
proteins, and in controlling protein renewal (96). Supplementary arginine increases
the production of nitric oxide, and then induces the expression of
utrophin and the activation of muscle satellite cells, thereby
improving the pathological process of muscular dystrophy (97). Proline is not only the result of
arginine metabolism but also the raw material for the synthesis of
arginine (98). This process
depends on the ornithine cycle, which renders the synthesis and
metabolism of arginine recyclable. The pulmonary matrix is degraded
in patients with COPD, and matrix metalloproteinases further
destroy collagen fibers and degrade them into small fragments of
proline and other amino acids. Hypoxic damage in patients with COPD
is likely to be accompanied by the presence of methylated arginine
products, which means that arginine may represent a biomarker of an
acute attack of COPD (99). These
methylated arginine products lead to airway obstruction through the
decomposition of arginine (100). The pathways may also involve
multiple extra-pulmonary injuries (101,102). For example, associated amino
acids may contribute to the imbalance of protein synthesis and
degradation in COPD skeletal muscle dysfunction and muscle atrophy
(103). Early studies indicated
that exercise affects the metabolism of arginine and proline,
although in a manner that is dependent on exercise patterns and
intensity (94,95). Therefore, the effect of exercise
on the metabolic pathway of arginine and proline may be one of the
mechanisms through which exercise affects the function of the
diaphragm muscle.
In the present study, it was identified that
threonine and serine may contribute to exercise-dependent
improvement of diaphragm function. The interaction between glycine,
serine and threonine metabolism is close: Glycine and serine can be
transformed into each other under the action of
hydroxylmethyltransferase (104), while threonine can be decomposed
into serine or glycine under the action of specific enzymes, such
as hydroxymethyl transferase and threonine deaminase. The
metabolism of these amino acids requires further study based on
their vital biological functions.
Finally, lipid-associated processes, such as fatty
acid synthesis and metabolism, were also identified in the
comparison of the differential metabolite-associated pathways,
although these factors were not significantly involved. With the
in-depth studies on adipose tissue, in particular the
identification of leptin (105),
adiponectin (106), resistin
(107) and chemokines (108), the multiple roles of adipose
tissue in chronic diseases are gradually being revealed. Adipose
tissue may serve a regulatory role in the body through multiple
pathways (109). At present,
exercise training has demonstrated a unique stimulating effect on
the adipose tissue: Moderate exercise accelerates lipid metabolism
(110,111). Simultaneously, as the basic
component of lipids, fatty acids are considered to be associated
with COPD inflammation (112,113), which suggests that changes in
fatty acids are likely to be a potential factor affecting COPD
disease progression. Adding a fatty acid supplement to the
treatment regimen of patients with COPD on the basis of exercise
training can improve the regulation of the immune response of the
body, and the change in immune function can further affect the
inflammatory response of the disease, thereby achieving effective
exercise rehabilitation (114).
According to the results of the present study, exercise may also
affect the role of adipose tissue in the human body, producing
therapeutic effects.
In the present study, CSE was used in the process of
establishing animal models of COPD, which has been applied in
numerous studies. However, the factors leading to COPD formation
are complex: Multiple etiologies are considered to be associated
with the occurrence of COPD, such as cigarette smoke, air
pollution, metabolic derangements, genetics and aging (1,115). Unfortunately, the present study
only simulated a COPD animal model using cigarette smoke, which is
one of the study limitations. Pulmonary function tests and lung
histology were used to identify COPD models. Following verification
of the model, the remaining rats underwent an exercise intervention
or were allowed to move freely. A 9-week intervention indicated the
effectiveness of regular exercise and caused changes in multiple
metabolites. In the present study, certain physiological values
were measured, such as diaphragm function and pulmonary function
tests, to measure diaphragm function directly or indirectly,
respectively. Nevertheless, these indicators are not sufficient,
and more assessments of the function of the target muscles and
physiological studies should be performed, which is one of the
limitations of the present study, as the closer the relationship
between biological alternations and pathophysiological phenomena,
the more meaningful the results of biological analysis will be.
Future studies should focus on the quantitative changes of
differential metabolites and their metabolic pathways. Furthermore,
the present study group will conduct knockout/overexpression
studies of the metabolic pathways of the metabolites identified in
the present study, to observe whether the effect of exercise on the
improvement of diaphragm function is affected. Whether the
biomarkers identified in the present study are also relevant in
patients with COPD remains to be established, as the present study
focused on a COPD animal model.
In conclusion, physiological studies and biological
analyses were used in the present study. Following confirmation of
the improvement in diaphragm function, metabolomics research
methods were applied to explore the potential molecular biology
mechanism of exercise training to improve diaphragm function in
COPD. A total of 4 metabolites (threonine, serine, nicotinamide and
lactic acid) were considered as potential biomarkers to enhance the
diaphragm function in COPD rats. Concomitantly, through the
analysis of the pathways of different metabolites, 2 pathways, the
nicotinic acid and nicotinamide metabolism and arginine and proline
metabolism pathways, were identified to be the most likely
metabolic pathways involved in the exercise-based improvement of
diaphragm function. The present study provides a reference for
exploring the mechanism of pulmonary rehabilitation through
screening biomarkers and associated pathways by metabolomics, but
further studies are required.
Supplementary Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81902307).
Availability of data and materials
The data used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JL, WW and XL made substantial contributions to the
conception and design of the study. YL, NL, PL, ZW, TW, ZY and YY
acquired, analyzed and interpreted the data. JL, JS, HC, LX and HD
drafted the article and revised it critically for important
intellectual content. All authors read and approved the final
manuscript. All authors agree to be held accountable for all
aspects of the work in ensuring that questions related to the
accuracy or integrity of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
The experimental protocol was approved by the
Institutional Animal Care and Use Committee of Shanghai University
of Sport (approval no. 2018026), and was conducted following the
Animal Research Guidelines of the Shanghai University of Sport.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Horner A, Soriano JB, Puhan MA, Studnicka
M, Kaiser B, Vanfleteren LEGW, Gnatiuc L, Burney P, Miravitlles M,
García-Rio F, et al: Altitude and COPD prevalence: Analysis of the
PREPOCOL-PLATINO-BOLD-EPI-SCAN study. Respir Res. 18:1622017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mesquita R, Donária L, Genz IC, Pitta F
and Probst VS: Respiratory muscle strength during and after
hospitalization for COPD exacerbation. Respir Care. 58:2142–2149.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hodgev VA and Kostianev SS: Maximal
inspiratory pressure predicts mortality in patients with chronic
obstructive pulmonary disease in a five-year follow-up. Folia Med.
48:36–41. 2006.
|
|
4
|
Newell SZ, McKenzie DK and Gandevia SC:
Inspiratory and skeletal muscle strength and endurance and
diaphragmatic activation in patients with chronic airflow
limitation. Thorax. 44:903–912. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fabbri LM and Rabe KF: From COPD to
chronic systemic inflammatory syndrome? Lancet. 370:797–799. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kirkham PA and Barnes PJ: Oxidative stress
in COPD. Chest. 144:266–273. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rochester DF: The diaphragm in COPD.
Better than expected, but not good enough. New Engl J Med.
325:961–962. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Scott A, Wang X, Road JD and Reid WD:
Increased injury and intramuscular collagen of the diaphragm in
COPD: Autopsy observations. Eur Respir J. 27:51–59. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Doucet M, Debigare R, Joanisse DR, Côté C,
Leblanc P, Grégoire J, Deslauriers J, Vaillancourt R and Maltais F:
Adaptation of the diaphragm and the vastus lateralis in
mild-to-moderate COPD. Eur Respir J. 24:971–979. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Haegens A, Schols AM, Gorissen SH, van
Essen AL, Snepvangers F, Gray DA, Shoelson SE and Langen RC: NF-κB
activation and polyubiquitin conjugation are required for pulmonary
inflammation-induced diaphragm atrophy. Am J Physiol Lung Cell Mol
Physiol. 302:L103–L110. 2012. View Article : Google Scholar
|
|
11
|
Wijnhoven HJ, Heunks LM, Geraedts MC,
Hafmans T, Viña JR and Dekhuijzen PN: Oxidative and nitrosative
stress in the diaphragm of patients with COPD. Int J Chron Obstruct
Pulmon Dis. 1:173–179. 2006.
|
|
12
|
Barreiro E and Gea J: Respiratory and limb
muscle dysfunction in COPD. COPD. 12:413–426. 2015. View Article : Google Scholar
|
|
13
|
Nici L, Donner C, Wouters E, Zuwallack R,
Ambrosino N, Bourbeau J, Carone M, Celli B, Engelen M, Fahy B, et
al: American thoracic society/European respiratory society
statement on pulmonary rehabilitation. Am J Respir Crit Care Med.
173:1390–1413. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ricci C, Terzoni S, Gaeta M, Sorgente A,
Destrebecq A and Gigliotti F: Physical training and noninvasive
ventilation in COPD patients: A meta-analysis. Respir Care.
59:709–717. 2014. View Article : Google Scholar
|
|
15
|
Polkey MI, Kyroussis D, Hamnegard CH,
Mills GH, Green M and Moxham J: Diaphragm strength in chronic
obstructive pulmonary disease. Am J Respir Crit Care Med.
154:1310–1317. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Koulmann N and Bigard AX: Interaction
between signalling pathways involved in skeletal muscle responses
to endurance exercise. Pflugers Arch. 452:125–139. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rowell LB, Sheriff DD, Wyss CR and Scher
AM: The nature of the exercise stimulus. Acta Physiol Scand Suppl.
556:7–14. 1986.PubMed/NCBI
|
|
18
|
Casado-Vela J, Cebrián A, Gómez del Pulgar
MT and Lacal JC: Approaches for the study of cancer: Towards the
integration of genomics, proteomics and metabolomics. Clin Transl
Oncol. 13:617–628. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cox J and Mann M: Is proteomics the new
genomics? Cell. 130:395–398. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lindon JC, Holmes E and Nicholson JK:
Metabonomics techniques and applications to pharmaceutical research
& development. Pharm Res. 23:1075–1088. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lindon JC, Holmes E, Bollard ME, Stanley
EG and Nicholson JK: Metabonomics technologies and their
applications in physiological monitoring, drug safety assessment
and disease diagnosis. Biomarkers. 9:1–31. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wadell K: Water-based exercise is more
effective than land-based exercise for people with COPD and
physical comorbidities. J Physiother. 60:572014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu W, Liu X, Liu J, Li P and Wang Z:
Effectiveness of water-based Liuzijue exercise on respiratory
muscle strength and peripheral skeletal muscle function in patients
with COPD. Int J Chron Obstruct Pulmon Dis. 13:1713–1726. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Carlos SP, Dias AS, Forgiarini Júnior LA,
Patricio PD, Graciano T, Nesi RT, Valença S, Chiappa AM, Cipriano G
Jr, Souza CT and Chiappa GR: Oxidative damage induced by cigarette
smoke exposure in mice: Impact on lung tissue and diaphragm muscle.
J Bras Pneumol. 40:411–420. 2014.In English, Portuguese. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vieira Ramos G, Choqueta de Toledo-Arruda
A, Maria Pinheiro-Dardis C, Liyoko Suehiro C, Luiz de Russo T,
Vieira RP, Arruda Martins M, Salvini TF and Durigan JLQ: Exercise
prevents diaphragm wasting induced by cigarette smoke through
modulation of antioxidant genes and metalloproteinases. Biomed Res
Int. 2018:59090532018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bowen TS, Aakerøy L, Eisenkolb S, Kunth P,
Bakkerud F, Wohlwend M, Ormbostad AM, Fischer T, Wisloff U, Schuler
G, et al: Exercise training reverses extrapulmonary impairments in
smoke-exposed mice. Med Sci Sports Exerc. 49:879–887. 2017.
View Article : Google Scholar
|
|
27
|
Barreiro E, del Puerto-Nevado L,
Puig-Vilanova E, Pérez-Rial S, Sánchez F, Martínez-Galán L, Rivera
S, Gea J, González-Mangado N and Peces-Barba G: Cigarette
smoke-induced oxidative stress in skeletal muscles of mice. Respir
Physiol Neurobiol. 182:9–17. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Krüger K, Dischereit G, Seimetz M, Wilhelm
J, Weissmann N and Mooren FC: Time course of cigarette
smoke-induced changes of systemic inflammation and muscle
structure. Am J Physiol Lung Cell Mol Physiol. 309:L119–L128. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wüst RCI and Degens H: Factors
contributing to muscle wasting and dysfunction in COPD patients.
Int J Chron Obstruct Pulmon Dis. 2:289–300. 2007.
|
|
30
|
Nie YC, Wu H, Li PB, Luo YL, Zhang CC,
Shen JG and Su WW: Characteristic comparison of three rat models
induced by cigarette smoke or combined with LPS: To establish a
suitable model for study of airway mucus hypersecretion in chronic
obstructive pulmonary disease. Pulm Pharmacol Ther. 25:349–356.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Caron MA, Morissette MC, Thériault ME,
Nikota JK, Stämpfli MR and Debigaré R: Alterations in skeletal
muscle cell homeostasis in a mouse model of cigarette smoke
exposure. PLoS One. 8:e664332013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Totou NL, Moura SS, Coelho DB, Oliveira
EC, Becker LK and Lima WG: Swimming exercise demonstrates
advantages over running exercise in reducing proteinuria and
glomerulosclerosis in spontaneously hypertensive rats. Physiol Int.
105:76–85. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Barbosa Neto O, Abate DT, Marocolo Júnior
M, Mota GR, Orsatti FL, Rossi e Silva RC, Reis MA and da Silva VJ:
Exercise training improves cardiovascular autonomic activity and
attenuates renal damage in spontaneously hypertensive rats. J
Sports Sci Med. 12:52–59. 2013.PubMed/NCBI
|
|
34
|
Egawa T, Tsuda S, Goto A, Ohno Y, Yokoyama
S, Goto K and Hayashi T: Potential involvement of dietary advanced
glycation end products in impairment of skeletal muscle growth and
muscle contractile function in mice. Br J Nutr. 117:21–29. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yao S, Zhang J, Chen H, Sheng Y, Zhang X,
Liu Z and Zhang C: Diagnostic value of immunohistochemical staining
of GP73, GPC3, DCP, CD34, CD31, and reticulin staining in
hepatocellular carcinoma. J Histochem Cytochem. 61:639–648. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kind T, Wohlgemuth G, Lee DY, Lu Y,
Palazoglu M, Shahbaz S and Fiehn O: FiehnLib: Mass spectral and
retention index libraries for metabolomics based on quadrupole and
time-of-flight gas chromatography/mass spectrometry. Anal Chem.
81:10038–10048. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dunn WB, Broadhurst D, Begley P, Zelena E,
Francis-McIntyre S, Anderson N, Brown M, Knowles JD, Halsall A,
Haselden JN, et al: Procedures for large-scale metabolic profiling
of serum and plasma using gas chromatography and liquid
chromatography coupled to mass spectrometry. Nat Protoc.
6:1060–1083. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Alfakih A, Khandani A and Wolkowicz H:
Solving Euclidean distance matrix completion problems via
semidefinite programming. Comput Optim Appl. 12:13–30. 1999.
View Article : Google Scholar
|
|
39
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar
|
|
40
|
Kanehisa M, Sato Y, Kawashima M, Furumichi
M and Tanabe M: KEGG as a reference resource for gene and protein
annotation. Nucleic Acids Res. 44(D1): D457–D462. 2016. View Article : Google Scholar :
|
|
41
|
Toledo AC, Magalhaes RM, Hizume DC, Vieira
RP, Biselli PJ, Moriya HT, Mauad T, Lopes FD and Martins MA:
Aerobic exercise attenuates pulmonary injury induced by exposure to
cigarette smoke. Eur Respir J. 39:254–264. 2012. View Article : Google Scholar
|
|
42
|
Ma WL, Cai PC, Xiong XZ and Ye H: Exercise
training attenuated chronic cigarette smoking-induced up-regulation
of FIZZ1/RELMα in lung of rats. J Huazhong Univ Sci Technolog Med
Sci. 33:22–26. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Nishii Y, Kawata S, Fujita N, Tomoda K and
Imagita H: Moderate exercise attenuated airway resistance and
inflammation induced by cigarette smoke solution and endotoxin in
rats. Sport Sci Health. 12:91–97. 2016. View Article : Google Scholar
|
|
44
|
Eklund BM, Nilsson S, Hedman L and
Lindberg I: Why do smokers diagnosed with COPD not quit smoking?-a
qualitative study. Tob Induc Dis. 10:172012. View Article : Google Scholar
|
|
45
|
Taghizadeh N, Vonk JM and Boezen HM:
Lifetime smoking history and cause-specific mortality in a cohort
study with 43 years of follow-up. PLoS One. 11:e01533102016.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Klaude M, Gedik CM and Collins AR: DNA
damage and repair after low doses of UV-C radiation; comparable
rates of repair in rodent and human cells. Int J Radiat Biol.
67:501–508. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Rees DC, Wood RW and Laties VG: Stimulus
control and the development of behavioral tolerance to daily
injections of d-amphetamine in the rat. J Pharmacol Exp Ther.
240:65–73. 1987.PubMed/NCBI
|
|
48
|
Siegel S and Sdao-Jarvie K: Attenuation of
ethanol tolerance by a novel stimulus. Psychopharmacology.
88:258–261. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Swanny A, Morton RF and Lee LY: Acute
effect of cigarette smoke on breathing is attenuated by chronic
smoking in rats. J Appl Physiol (1985). 74:333–338. 1993.
View Article : Google Scholar
|
|
50
|
Thames HD Jr, Withers HR and Peters LJ:
Tissue repair capacity and repair kinetics deduced from
multifractionated or continuous irradiation regimens with
incomplete repair. Br J Cancer Suppl. 6:263–269. 1984.PubMed/NCBI
|
|
51
|
Rinaldi M, Maes K, De Vleeschauwer S,
Thomas D, Verbeken EK, Decramer M, Janssens W and Gayan-Ramirez GN:
Long-term nose-only cigarette smoke exposure induces emphysema and
mild skeletal muscle dysfunction in mice. Dis Model Mech.
5:333–341. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Berry CE and Wise RA: Interpretation of
pulmonary function test: Issues and controversies. Clin Rev Allergy
Immunol. 37:173–180. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Cukic V, Lovre V and Ustamujic A: The
changes of pulmonary function in COPD during four-year period.
Mater Sociomed. 25:88–92. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wang J, Zhou XM, Yang X, Zhao ST, Ma QL
and Wang CZ: A three years longitudinal follow-up study of
pulmonary function changes in patients with chronic obstructive
pulmonary disease. Chin J Intern Med. 55:302–306. 2016.In
Chinese.
|
|
55
|
Talaminos Barroso A, Márquez Martin E, Roa
Romero LM and Ortega Ruiz F: Factors affecting lung function: A
review of the literature. Arch Bronconeumol. 54:327–332. 2018.In
English, Spanish. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Davidson WJ, Verity WS, Traves SL, Leigh
R, Ford GT and Eves ND: Effect of incremental exercise on airway
and systemic inflammation in patients with COPD. J Appl Physiol
(1985). 112:2049–2056. 2012. View Article : Google Scholar
|
|
57
|
Silva RA, Almeida FM, Olivo CR,
Saraiva-Romanholo BM, Martins MA and Carvalho CR: Airway remodeling
is reversed by aerobic training in a murine model of chronic
asthma. Scand J Med Sci Sports. 25:e258–e266. 2015. View Article : Google Scholar
|
|
58
|
Vieira RP, Claudino RC, Duarte AC, Santos
AB, Perini A, Faria Neto HC, Mauad T, Martins MA, Dolhnikoff M and
Carvalho CR: Aerobic exercise decreases chronic allergic lung
inflammation and airway remodeling in mice. Am J Respir Crit Care
Med. 176:871–877. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lan CC, Chu WH, Yang MC, Lee CH, Wu YK and
Wu CP: Benefits of pulmonary rehabilitation in patients with COPD
and normal exercise capacity. Respir Care. 58:1482–1488. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Ambrosino N, Foglio K, Balzano G, Paggiaro
PL, Lessi P and Kesten S; Tiotropium Multicentric Italian Study
Group: Tiotropium and exercise training in COPD patients: Effects
on dyspnea and exercise tolerance. Int J Chron Obstruct Pulmon Dis.
3:771–780. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Leelarungrayub J, Pinkaew D, Puntumetakul
R and Klaphajone J: Effects of a simple prototype respiratory
muscle trainer on respiratory muscle strength, quality of life and
dyspnea, and oxidative stress in COPD patients: A preliminary
study. Int J Chron Obstruct Pulmon Dis. 12:1415–1425. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Armbruster C, Dassow C, Gamerdinger K,
Guttmann J, Schneider M and Schumann S: In vitro muscle contraction
force measurements on isolated and entire rat diaphragms. Critical
Care. 14:P2042010. View
Article : Google Scholar :
|
|
63
|
Gea J, Agusti A and Roca J:
Pathophysiology of muscle dysfunction in COPD. J Appl Physiol
(1985). 114:1222–1234. 2013. View Article : Google Scholar
|
|
64
|
Levine S, Bashir MH, Clanton TL, Powers SK
and Singhal S: COPD elicits remodeling of the diaphragm and vastus
lateralis muscles in humans. J Appl Physiol (1985). 114:1235–1245.
2013. View Article : Google Scholar
|
|
65
|
Gayan-Ramirez G and Decramer M: Mechanisms
of striated muscle dysfunction during acute exacerbations of COPD.
J Appl Physiol (1985). 114:1291–1299. 2013. View Article : Google Scholar
|
|
66
|
Levine S, Kaiser L, Leferovich J and
Tikunov B: Cellular adaptations in the diaphragm in chronic
obstructive pulmonary disease. N Engl J Med. 337:1799–1806. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Marin-Corral J, Minguella J,
Ramirez-Sarmiento AL, Hussain SN, Gea J and Barreiro E: Oxidised
proteins and superoxide anion production in the diaphragm of severe
COPD patients. Eur Respir J. 33:1309–1319. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Spruit MA, Gosselink R, Troosters T, De
Paepe K and Decramer M: Resistance versus endurance training in
patients with COPD and peripheral muscle weakness. Eur Respir J.
19:1072–1078. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Franssen FM, Broekhuizen R, Janssen PP,
Wouters EF and Schols AM: Effects of whole-body exercise training
on body composition and functional capacity in normal-weight
patients with COPD. Chest. 125:2021–2028. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Cortopassi F, Castro AA, Porto EF, Colucci
M, Fonseca G, Torre-Bouscoulet L, Iamonti V and Jardim JR:
Comprehensive exercise training improves ventilatory muscle
function and reduces dyspnea perception in patients with COPD.
Monaldi Arch Chest Dis. 71:106–112. 2009.PubMed/NCBI
|
|
71
|
Wada JT, Borges-Santos E, Porras DC,
Paisani DM, Cukier A, Lunardi AC and Carvalho CR: Effects of
aerobic training combined with respiratory muscle stretching on the
functional exercise capacity and thoracoabdominal kinematics in
patients with COPD: A randomized and controlled trial. Int J Chron
Obstruct Pulmon Dis. 11:2691–2700. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Ries AL, Bauldoff GS, Carlin BW, Casaburi
R, Emery CF, Mahler DA, Make B, Rochester CL, Zuwallack R and
Herrerias C: Pulmonary rehabilitation: Joint ACCP/AACVPR
evidence-based clinical practice guidelines. Chest. 131(5 Suppl):
4S–42S. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Wanke T, Formanek D, Lahrmann H, Brath H,
Wild M, Wagner C and Zwick H: Effects of combined inspiratory
muscle and cycle ergometer training on exercise performance in
patients with COPD. Eur Respir J. 7:2205–2211. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
de Souto Araujo ZT, de Miranda Silva
Nogueira PA, Cabral EE, de Paula Dos Santos L, da Silva IS and
Ferreira GM: Effectiveness of low-intensity aquatic exercise on
COPD: A randomized clinical trial. Respir Med. 106:1535–1543. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Wackerhage H and Ratkevicius A: Signal
transduction pathways that regulate muscle growth. Essays Biochem.
44:99–108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Chaouloff F, Kennett GA, Serrurrier B,
Merino D and Curzon G: Amino acid analysis demonstrates that
increased plasma free tryptophan causes the increase of brain
tryptophan during exercise in the rat. J Neurochem. 46:1647–1650.
1986. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Inuzuka M, Hayakawa M and Ingi T: Serinc,
an activity-regulated protein family, incorporates serine into
membrane lipid synthesis. J Biol Chem. 280:35776–35783. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Pantaleo A, Ferru E, Carta F, Mannu F,
Giribaldi G, Vono R, Lepedda AJ, Pippia P and Turrini F: Analysis
of changes in tyro-sine and serine phosphorylation of red cell
membrane proteins induced by P. falciparum growth. Proteomics.
10:3469–3479. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Hu SI, Katz M, Chin S, Qi X, Cruz J,
Ibebunjo C, Zhao S, Chen A and Glass DJ: MNK2 inhibits eIF4G
activation through a pathway involving serine-arginine-rich protein
kinase in skeletal muscle. Sci Signal. 5:ra142012. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Talanian JL, Tunstall RJ, Watt MJ, Duong
M, Perry CG, Steinberg GR, Kemp BE, Heigenhauser GJ and Spriet LL:
Adrenergic regulation of HSL serine phosphorylation and activity in
human skeletal muscle during the onset of exercise. Am J Physiol
Regul Integr Comp Physiol. 291:R1094–R1099. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Li F, Chong ZZ and Maiese K: Cell life
versus cell longevity: The mysteries surrounding the NAD+ precursor
nicotinamide. Curr Med Chem. 13:883–895. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Ungerstedt JS, Blömback M and Söderström
T: Nicotinamide is a potent inhibitor of proinflammatory cytokines.
Clin Exp Immunol. 131:48–52. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Traister A, Breitman I, Bar-Lev E, Zvibel
I, Harel A, Halpern Z and Oren R: Nicotinamide induces apoptosis
and reduces collagen I and pro-inflammatory cytokines expression in
rat hepatic stellate cells. Scand J Gastroenterol. 40:1226–1234.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Chlopicki S, Swies J, Mogielnicki A,
Buczko W, Bartus M, Lomnicka M, Adamus J and Gebicki J:
1-Methylnicotinamide (MNA), a primary metabolite of nicotinamide,
exerts anti-thrombotic activity mediated by a
cyclooxygenase-2/prostacyclin pathway. Br J Pharmacol. 152:230–239.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Surenkok O, Kin-Isler A, Aytar A and
Gültekin Z: Effect of trunk-muscle fatigue and lactic acid
accumulation on balance in healthy subjects. J Sport Rehabil.
17:380–386. 2008. View Article : Google Scholar
|
|
86
|
Robergs RA, Ghiasvand F and Parker D:
Biochemistry of exercise-induced metabolic acidosis. Am J Physiol
Regul Integr Comp Physiol. 287:R502–R516. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Gladden LB: Lactate metabolism: A new
paradigm for the third millennium. J Physiol. 558:5–30. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Lewis SF and Haller RG: The
pathophysiology of McArdle's disease: Clues to regulation in
exercise and fatigue. J Appl Physiol (1985). 61:391–401. 1986.
View Article : Google Scholar
|
|
89
|
Hunt TK, Aslam R, Hussain Z and Beckert S:
Lactate, with oxygen, incites angiogenesis. Adv Exp Med Biol.
614:73–80. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Groussard C, Morel I, Chevanne M, Monnier
M, Cillard J and Delamarche A: Free radical scavenging and
antioxidant effects of lactate ion: An in vitro study. J Appl
Physiol (1985). 89:169–175. 2000. View Article : Google Scholar
|
|
91
|
Thiele I, Swainston N, Fleming RM, Hoppe
A, Sahoo S, Aurich MK, Haraldsdottir H, Mo ML, Rolfsson O, Stobbe
MD, et al: A community-driven global reconstruction of human
metabolism. Nat Biotechnol. 31:419–425. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Stoltz JF and Nicolas A: Study of amino
groups of the human platelet membrane. Acta Haematol. 60:304–309.
1978. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Menon R, Tolbert D and Cefali E: The
comparative bioavailability of an extended-release niacin and
lovastatin fixed dose combination tablet versus extended-release
niacin tablet, lovastatin tablet and a combination of
extended-release niacin tablet and lovastatin tablet. Biopharm Drug
Dispos. 28:297–306. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Shibata K, Matsumoto K, Fushiki T and
Sugimoto E: Effects of exercise on the metabolism of NAD in rats.
Biosci Biotech Bioch. 58:1763–1766. 1994. View Article : Google Scholar
|
|
95
|
White AT and Schenk S: NAD(+)/NADH and
skeletal muscle mitochondrial adaptations to exercise. Am J Physiol
Endocrinol Metab. 303:E308–E321. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Wu G and Morris SM Jr: Arginine
metabolism: Nitric oxide and beyond. Biochem J. 336:1–17. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Barton ER, Morris L, Kawana M, Bish LT and
Toursel T: Systemic administration of L-arginine benefits mdx
skeletal muscle function. Muscle nerve. 32:751–760. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Wu G, Bazer FW, Datta S, Johnson GA, Li P,
Satterfield MC and Spencer TE: Proline metabolism in the conceptus:
Implications for fetal growth and development. Amino Acids.
35:691–702. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Ruzsics I, Nagy L, Keki S, Sarosi V, Illes
B, Illes Z, Horvath I, Bogar L and Molnar T: L-arginine pathway in
COPD patients with acute exacerbation: A new potential biomarker.
COPD. 13:139–145. 2016. View Article : Google Scholar
|
|
100
|
Dohm GL, Beecher GR, Warren RQ and
Williams RT: Influence of exercise on free amino acid
concentrations in rat tissues. J Appl Physiol Respir Environ Exerc
Physiol. 50:41–44. 1981.PubMed/NCBI
|
|
101
|
Parmaksız ET, Inal A, Salepci B, Comert S,
Fidan A, Kiral N, Doǧan C and Caglayan B: Relationship of
asymmetric dimethylarginine levels with disease severity and
pulmonary hypertension in chronic obstructive pulmonary disease.
Lung India. 35:199–203. 2018. View Article : Google Scholar
|
|
102
|
van den Berg MP, Meurs H and Gosens R:
Targeting arginase and nitric oxide metabolism in chronic airway
diseases and their comorbidities. Curr Opin Pharmacol. 40:126–133.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Hörster I, Weigt-Usinger K, Carmann C,
Chobanyan-Jürgens K, Köhler C, Schara U, Kayacelebi AA, Beckmann B,
Tsikas D and Lücke T: The L-arginine/NO pathway and homoarginine
are altered in Duchenne muscular dystrophy and improved by
glucocorticoids. Amino Acids. 47:1853–1863. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Wang W, Wu Z, Dai Z, Yang Y, Wang J and Wu
G: Glycine metabolism in animals and humans: Implications for
nutrition and health. Amino Acids. 45:463–477. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Bruno A, Chanez P, Chiappara G, Siena L,
Giammanco S, Gjomarkaj M, Bonsignore G, Bousquet J and Vignola AM:
Does leptin play a cytokine-like role within the airways of COPD
patients? Eur Respir J. 26:398–405. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Bianco A, Mazzarella G, Turchiarelli V,
Nigro E, Corbi G, Scudiero O, Sofia M and Daniele A: Adiponectin:
An attractive marker for metabolic disorders in chronic obstructive
pulmonary disease (COPD). Nutrients. 5:4115–4125. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Al Mutairi SS, Mojiminiyi OA,
Shihab-Eldeen A, Al Rammah T and Abdella N: Putative roles of
circulating resistin in patients with asthma, COPD and cigarette
smokers. Dis Markers. 31:1–7. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Tomaki M, Sugiura H, Koarai A, Komaki Y,
Akita T, Matsumoto T, Nakanishi A, Ogawa H, Hattori T and Ichinose
M: Decreased expression of antioxidant enzymes and increased
expression of chemokines in COPD lung. Pulm Pharmacol Ther.
20:596–605. 2007. View Article : Google Scholar
|
|
109
|
Tilg H and Moschen AR: Adipocytokines:
Mediators linking adipose tissue, inflammation and immunity. Nat
Rev Immunol. 6:772–783. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Horowitz JF and Klein S: Lipid metabolism
during endurance exercise. Am J Clin Nutr. 72(2 Suppl): 558S–563S.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Gill JM, Frayn KN, Wootton SA, Miller GJ
and Hardman AE: Effects of prior moderate exercise on exogenous and
endogenous lipid metabolism and plasma factor VII activity. Clin
Sci (Lond). 100:517–527. 2001. View Article : Google Scholar
|
|
112
|
de Batlle J, Sauleda J, Balcells E, Gómez
FP, Méndez M, Rodriguez E, Barreiro E, Ferrer JJ, Romieu I, Gea J,
et al: Association between Ω3 and Ω6 fatty acid intakes and serum
inflammatory markers in COPD. J Nutr Biochem. 23:817–821. 2012.
View Article : Google Scholar
|
|
113
|
Warnakulasuriya SN, Ziaullah and
Rupasinghe HP: Novel long chain fatty acid derivatives of
quercetin-3-O-glucoside reduce cytotoxicity induced by cigarette
smoke toxicants in human fetal lung fibroblasts. Eur J Pharmacol.
781:128–138. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Garrod R, Ansley P, Canavan J and Jewell
A: Exercise and the inflammatory response in chronic obstructive
pulmonary disease (COPD)-Does training confer anti-inflammatory
prop-erties in COPD? Med Hypotheses. 68:291–298. 2007. View Article : Google Scholar
|
|
115
|
Spurzem JR and Rennard SI: Pathogenesis of
COPD. Semin Respir Crit Care Med. 26:142–153. 2005. View Article : Google Scholar : PubMed/NCBI
|