Introduction
Liver fibrosis results from chronic liver injury,
for example persistent hepatitis virus infection, excessive alcohol
intake, non-alcoholic steatohepatitis, autoimmune disorders,
intra-hepatic cholestasis, drug abuse and metabolic abnormalities.
Accumulation of excessive extracellular matrix (ECM) proteins leads
to liver cirrhosis (LC), which is a leading cause of morbidity and
mortality worldwide (1).
Furthermore, progression of cirrhotic stage liver disease
frequently occurs with hepatocellular carcinoma (HCC) (2,3).
Therefore, anti-fibrotic therapies are urgently required to
decrease the mortality associated with liver injury and HCC.
There are ~140 clinical anti-fibrotic trials
currently registered at ClinicalTrials.gov (http://clinicaltrials.gov). To develop an effective
anti-fibrotic agent, an understanding of the precise mechanism
involved in liver fibrosis progression is required. Transforming
growth factor-β (TGF-β) is generally considered to be a key
cytokine in hepatic fibrosis and an inhibitor of hepatocyte growth
(4,5). TGF-β is also the most potent
stimulus for the synthesis of type-I collagen and other matrix
constituents. Therefore, agents that could inhibit the action of
TGF-β would contribute to suppression of liver fibrosis by lowering
levels of circulating TGF-β, antagonizing its receptors, and/or
blocking its activation via intracellular signaling pathways. We
previously reported that the inhibition of TGF-β signaling using
adenovirus-mediated transfer of a mutated TGF-β receptor gene to
rats with dimethylni-trosamine-induced liver injury prevented the
progression of liver fibrosis and promotes hepatic regeneration
(6-8). Our results suggested that anti-TGF-β
agents, i.e., serine/threonine kinase inhibitors, antibodies,
antisense or fusion proteins of TGF-β receptors, may be a
potentially effective therapy for liver fibrosis; however, there
are no specific inhibitors of TGF-β signaling being tested in
clinical settings to date.
TGF-β is also involved in the progression and
metastasis of cancer. The anti-tumor action of TGF-β signaling
inhibitors, including galunisertib, has attracted significant
attention and is being tested in clinical trials (9). Galunisertib is a small-molecule
inhibitor that targets the TGF-β receptor type-I kinase (10). This agent demonstrates potent and
selective inhibition of TGF-β receptor type-I and also affects
downstream signaling by inhibiting Smad phosphorylation (11). More recently, a Phase Ib study
involving Japanese patients with unresectable HCC (NCT02240433)
assessed the safety and anti-tumor potential of galunisertib in
combination with the receptor tyrosine kinase inhibitor sorafenib
(12). Galunisertib also
demonstrated significant antifibrotic potency in a human ex
vivo model using precision-cut liver sections from both healthy
controls and patients with cirrhosis (13). To examine the potential
therapeutic application of galunisertib for the treatment of liver
fibrosis, the present study investigated the anti-fibrogenic and
regenerative effects of galunisertib on liver cirrhosis induced by
carbon tetrachloride (CCl4) in mice.
Materials and methods
Cell lines and culture conditions
The immortalized human hepatic stellate cell (HSC)
line LX-2 was purchased from EMD Millipore Corporation (EMD
Millipore), the human hepatoblastoma HepG2 cell line was purchased
from Cellular Engineering Technologies (Cellular Engineering
Technologies Inc.), and human umbilical vein endothelial cells
(HUVEC) were purchased from PromoCell GmbH. LX-2 and HepG2 cells
were cultured in DMEM (Wako Pure Chemical Industries, Ltd.)
supplemented with 10% FBS (Biowest SAS). HUVECs were cultured in
EGM2 (Lonza Group, Ltd.) lacking hydrocortisone and supplemented
with 5% FBS, growth factors [human fibroblast growth factor β
(hFGFβ), human vascular endothelial growth factor (hVEGF), human
insulin-like growth factor-1 (hIGF-1) and human epidermal growth
factor (hEGF)], penicillin (100 U/ml) and streptomycin (100
µg/ml; Nacalai Tesque, Inc.) in a humidified atmosphere
containing 5% CO2 at 37°C.
To evaluate the effect of galunisertib on LX-2 cells
using reverse transcription-quantitative (RT-q) PCR and western
blot analysis, the cells were cultured under serum-starved
conditions at 37°C overnight and then treated with the indicated
concentration of galunisertib (20, 200 and 2,000 nM) or vehicle
under stimulation with 5 ng/ml TGF-β1 (R&D Systems, Inc.) for 2
and 48 h for the western blot analysis and RT-qPCR assays,
respectively. Galunisertib was dissolved in dimethyl sulfoxide
(DMSO) and diluted with culture medium.
Cell proliferation assay
HepG2 cells, LX-2 cells and HUVECs were plated onto
uncoated 96-well plates at a density of 1×103 cells/well for HepG2 and HUVEC or
3×103 cells/well for LX-2 and
cultured for 24 h. After overnight at 37°C incubation in serum-free
medium, the medium was changed to medium containing 10% FBS for
HepG2 cells, serum-free medium for LX-2 cells, or EGM2 medium
supplemented with growth factors (hFGFβ, hVEGF, hIGF-1 and hEGF)
and 5% FBS for HUVEC. The indicated concentrations of galunisertib
and human TGF-β1 (5 ng/ml) were then added to the culture media.
The cells were incubated for another 2, 3 or 5 days for LX-2, HepG2
or HUVEC cells, respectively. Cell proliferation rates were
determined using Cell Count Reagent SF (Nacalai Tesque, Inc.)
according to the manufacturer's protocol.
Tube formation assay
Tube formation assays were performed using a
µ-slide angiogenesis kit (ibidi GmbH) according to the
manufacturer's protocol. Chilled liquid Matrigel was applied to the
inner wells of a µ-slide (10 µl per well) and allowed
to solidify for 1 h at 37°C. HUVECs were suspended at a density of
1×103 cells/µl with EGM2
medium containing several growth factors (hFGFβ, hVEGF, hIGF-1 and
hEGF) and 10% FBS with or without galunisertib (2,000 nM) and
TGF-β1 (5 ng/ml). Cell suspensions (50 µl) were applied into
the upper well of the µ-slide and incubated at 37°C for 16 h
prior to staining with Calcein AM (5 µg/ml; PromoCell GmbH)
at 37°C for 15 min and capture using the BZ-X 710 computer-assisted
inverted light microscope (Keyence Corporation) at magnification,
×2. The cavities were counted using computer-assisted image
software (Adobe Photoshop CC 2017 version 18.1.1; Adobe Systems,
Inc.).
Animal model
Male C57BL/6 mice (6-week-old) were purchased from
Japan SLC, Inc. Mice were maintained at room temperature (21±2°C)
with a 12:12 h dark: light cycle and had ad libitum access
to food (standard laboratory chow) and water. The experimental
protocol was approved by the Ethics Review Committee for Animal
Experimentation of the Kurume University School of Medicine.
Experimental conditions and
administration of galunisertib
Galunisertib was provided by Eli Lilly and Company.
Liver fibrosis was induced by CCl4 (Wako Pure Chemical
Industries; 1 µl/g BW in olive oil) that was administered to
mice intraperitoneally under inhalational anesthesia using 3%
isoflurane (Wako Pure Chemical Industries) twice a week for 8
weeks. Following the procedure, breathing and changes in
circulation once a day were checked by appearance, and one of mice
in vehicle-treated group was euthanized by cervical dislocation
under 3% isoflurane anesthesia due to intestinal rupture. In week
5, the mice were divided randomly into four treatment groups:
Vehicle-treated (vehicle group; n=7) and treated with low-(50
mg/kg/day; n=7) middle-(150 mg/kg/day; n=7) and high-dose (300
mg/kg/day; n=4) galunisertib, which was administered by oral gavage
twice daily from week 5 to week 8. The vehicle group received the
control mixture (1 w/v% carboxymethylcellulose, 0.5 w/v% sodium
lauryl sulfate and 0.085 w/v% povidone) without galunisertib.
CCl4 administration was continued twice weekly until
week 8. Mice in the intact group (n=4) were intraperitoneally
injected with olive oil alone and orally received the control
mixture without galunisertib. On day 56, 72 h after the final
CCl4 injection, ~800 µl of whole blood samples
were obtained from inferior vena cava under 3% isoflurane
inhalational anesthesia. Mice were euthanized by cervical
dislocation under 3% isoflurane anesthesia. Following euthanasia,
whole liver samples were collected for histological and molecular
analyses.
RT-qPCR
Total RNA was extracted using an Isogen kit (Nippon
Gene Co., Ltd) following the manufacturer's protocol. Single-strand
cDNA was synthesized using a high-capacity RNA-to-cDNA kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.). qRT-PCR was performed
using TaqMan® Universal Master Mix (Applied Biosystems;
Thermo Fisher Scientific, Inc.) with technical triplicates using a
StepOnePlus™ Real-Time PCR System (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The thermocycling parameters were as
follows: 2 min at 50°C and 10 min at 95°C, followed by 45 cycles of
15 sec at 95°C and 1 min at 60°C. Relative quantification of gene
expression was performed according to the 2−ΔΔCq method
(14) using StepOne Software 2.0
(Applied Biosystems; Thermo Fisher Scientific, Inc.), and
Gapdh (as a housekeeping gene in mice) or GAPDH (as a
housekeeping gene in human) was used as the reference for
normalization of the results. Primer sequences are listed in
Table SI.
The expression of genes involved in liver
regeneration was analyzed by PCR array using a Mouse Growth Factors
RT2 Profiler PCR Array Plate
(cat. no. PAMM-041Z; Qiagen, Inc.) and the StepOnePlus™ Real-Time
PCR System according to the manufacturer's protocol. The data were
analyzed using SDS Software 2.3 and RT2 Profiler PCR Array Data Analysis
version 3.5 (www.sabiosciences.com/pcrarraydataanalysis.php;
SABiosciences Corporation; Qiagen, Inc.). Gene expression was
normalized to the mean of house-keeping genes in the array.
Histological examination
The liver tissues fixed in 10% buffered formalin for
24 h at room temperature were embedded in paraffin and sliced to
form 5 µm-thick sections. The sections were either processed
using Mallory's Azan staining using 0.1% azocarmine G (cat. no.
1A266; Waldeck, Division Chroma) for 50 min at 60°C and Aniline
Blue-Orange G (cat. no. 4005-2; Muto Pure Chemicals co., LTD.) for
15 min at room temperature, or were analyzed histochemically using
antibodies against α-smooth muscle actin (α-SMA; cat. no. 19245;
1:1,000; Cell Signaling Technology, Inc.), proliferating cell
nuclear antigen (PCNA; cat. no. sc-7907; 1:300; Santa Cruz
Biotechnology), cleaved caspase-3 (cat. no. 9664; 1:50; Cell
Signaling Technology, Inc.) and FITC conjugated isolectin B4 (cat.
no. FL-1201; 1:300; Vector Laboratories, Inc.). The tissues were
incubated with primary antibodies overnight at 4°C.
Immunoreactivity was visualized using EnVision+ system
HRP labelled polymer anti-rabbit (cat. no. K4003; Dako; Agilent
Technologies, Inc.) for 30 min at room temperature and a DAB
commercial kit (Liquid DAB+ Substrate Chromogen System;
cat. no. K3468; Dako; Agilent Technologies, Inc.). For
double-immunofluorescence examination, slides were incubated with
Alexa Fluor 568-conjugated anti-rabbit IgG (cat. no. A11036; 1:200;
Invitrogen; Thermo Fisher Scientific, Inc.) for 30 min at room
temperature and mounted with DAPI (VECTASHIELD mounting medium with
DAPI; cat. no. H1200; Vector Laboratories, Inc.) to label the
nuclei. Apoptosis was detected by modifying DNA fragments using
terminal deoxynucleotidyl transferase mediated d-UTP nick end
labeling (TUNEL) staining. Digoxigenin-labeled UTP was detected
with an antibody to digoxigenin. An in situ apoptosis
detection kit (cat. no. S7101; EMD Millipore) was used for TUNEL
staining according to the manufacturer's protocol. In brief, the
tissue sections were pretreated with proteinase K (20 µg/ml;
Wako Pure Chemical Industries, Ltd.) for 15 min at room
temperature. Following washing with distilled water for 3 min
trice, 3% hydrogen peroxide (Wako Pure Chemical Industries, Ltd.)
was used to quench endogenous peroxidase for 5 min at room
temperature. The sections were incubated with Working Strength TdT
Enzyme (EMD Millipore) for 1 h at 37°C, followed by Stop/Wash
buffer (EMD Millipore) for 10 min at room temperature.
Immunoreactivity was visualized using Anti-Digoxigenin Conjugate
(EMD Millipore) for 30 min at room temperature and Peroxidase
Substrate (EMD Millipore). The nuclei were stained with hematoxylin
for 30 sec at room temperature. Blue-stained areas in Mallory's
Azan staining, the immunoreactive areas for α-SMA, and PCNA-,
TUNEL- and cleaved caspase-3-positive hepatocytes were identified
with computer-assisted image software (Adobe Photoshop CC 2017
version 18.1.1) by a technician who was blinded to the treatment
regimens. The proliferation/apoptosis (PA) index was calculated by
% PCNA-positive hepatocytes/% TUNEL-positive hepatocytes. A total
of two fields were assessed at magnification, ×10 for each sample
for the Mallory's Azan and α-SMA staining. For the PCNA, TUNEL and
cleaved caspase-3 analysis of each sample, a total of five fields
were assessed at magnification, ×40.
Western blot analysis
Frozen liver tissues were homogenized in RIPA buffer
(Thermo Fisher Scientific, Inc.) containing 1% protease inhibitor
cocktail (Sigma-Aldrich; Merck KGaA) and 1% phosphatase inhibitor
cocktail (ready-made; Thermo Fisher Scientific, Inc.). The protein
concentration was determined using a detergent compatible protein
assay kit (Bio-Rad Laboratories, Inc.). Equal amounts of 20
µg protein per lane were loaded onto 10% SDS-PAGE,
electrophoresed, and electrotransferred to a FluoroTrans membrane
(Pall Life Sciences). Following protein transfer, the membranes
were blocked with 2% skimmed milk (Cell Signaling Technology, Inc.)
for 1 h at room temperature and incubated overnight at 4°C with
primary antibody [antibodies against phosphorylated (p-)Smad2 (cat.
no. 3101; 1:1,000), total (t-)Smad2 (cat. no. 5339; 1:1,000),
p-Smad3 (cat. no. 9520; 1:1,000), t-Smad3 (cat. no. 9523; 1:1,000),
Smad4 (cat. no. 9515; 1:1,000), p-p38 mitogen-activated protein
kinase (MAPK; cat. no. 9216; 1:2,000), t-p38 MAPK (cat. no. 9212;
1:1,000), p-extracellular signal-regulated kinase (ERK)1/2 (cat.
no. 4377; 1:1,000) and t-ERK1/2 (cat. no. 4695; 1:1,000) were from
Cell Signaling Technology, Inc.; antibodies for matrix
metalloproteinase (MMP)-1 (cat. no. ab137332; 1:1,000) and MMP-13
(cat. no. ab39012; 1:3,000) were from Abcam; antibodies for PCNA
(cat. no. sc71858; 1:1,000) and GAPDH (cat. no. sc25778; 1:1,000)
were from Santa Cruz Biotechnology]. Visualization of protein
signals was achieved with a HRP-conjugated secondary antibodies
[anti-rabbit IgG, HRP-conjugated whole ab donkey (cat. no. NA934;
1:10,000; GE Healthcare Life Sciences) or anti-mouse IgG,
HRP-linked F (ab')2 fragment sheep (cat. no. NA9310;
1:10,000; GE Healthcare)] for 1 h at room temperature, and an
enhanced chemiluminescence western blot analysis system (Amersham;
GE Healthcare) using a lumino image analyzer (LAS 4000 mini;
Fujifilm). For re-blotting the membranes, the membranes were
striped with WB Stripping Solution Strong (Nacalai Tesque, Inc.)
for 1 h at 37°C. Relative amounts of protein were calculated based
on luminescence signals in each sample using Multigauge software
(version 3.11; Fujifilm).
Hydroxyproline measurement
Hydroxyproline measurement was performed using 10 mg
liver tissue that was homogenized in 100 µl distilled water
prior to the addition of 100 µl 12 N hydrochloride acid. The
samples were then hydrolyzed by incubation for 3 h at 120°C.
Following the addition of 5 mg activated charcoal and
centrifugation at 10,000 × g for 5 min at 4°C, the amount of
hydroxyproline in the supernatants was measured using
Hydroxyproline Assay kit (cat. no. STA675; Cell Biolabs Inc.).
Gelatin zymography. Frozen liver tissues were
homogenized at 4°C in buffer [50 mmol/l Tris-HCl (pH 7.6), 150
mmol/l NaCl, 0.1% SDS, 1% sodium deoxycholate, 1% NP-40 and 1%
Triton-X 100], and the protein concentration was determined using a
DC protein assay kit (Bio-Rad Laboratories, Inc.). Samples (50
µg protein) were loaded and resolved on a gelatin zymogram
gel (10% SDS-PAGE gel with 0.1% gelatin). Gels were placed in 2.5%
Triton X-100 solution for 30 min at 37°C and then incubated for 18
h at 37°C in enzyme activation buffer [50 mmol/l Tris-HCl (pH 7.5)
with 5 mmol/l CaCl2]. Gels were then stained at room
temperature with 0.2% Coomassie blue R-250 (Nacalai Tesque, Inc.)
for 30 min and de-stained at room temperature in 10% acetic acid
and 20% methanol for 15 min, as described previously (15). Relative amounts of protein were
calculated based on luminescence signals in each sample using
Multigauge software (version 3.11; Fujifilm).
Serum analysis
The biochemical parameters (AST, ALT, total-protein,
albumin and total-bilirubin) of serum samples were measured by
Oriental Yeast Co., Ltd.
Statistical analysis
Data analyses were conducted using JMP Pro software
v.13 (SAS Institute, Inc.). Differences between two groups were
examined for statistical significance using an unpaired Student's
t-test, and significant differences between multiple groups were
determined using one-way ANOVA followed by a Tukey's post hoc test.
All data are expressed as the mean ± SEM, and P<0.05 was
considered to indicate a statistically significant difference.
Results
TGF-β signaling via TGF-β receptor type I
kinase is inhibited by galunisertib in LX-2 cells
Galunisertib treatment of the human HSC line LX-2
dose-dependently decreased the levels of p-Smad2 and p-Smad3
independently of TGF-β1-induced signal activation, whereas
galunisertib treatment did not affect Smad4 expression (Figs. 1A and S1). In the human hepatoblastoma HepG2
cell line, the level of p-Smad2 was similarly decreased by
galunisertib treatment (Fig.
S2A-D). Conversely, galunisertib did not inhibit
phosphorylation of p38 MAPK or ERK1/2 in LX-2 cells (Figs. 1A and S1).
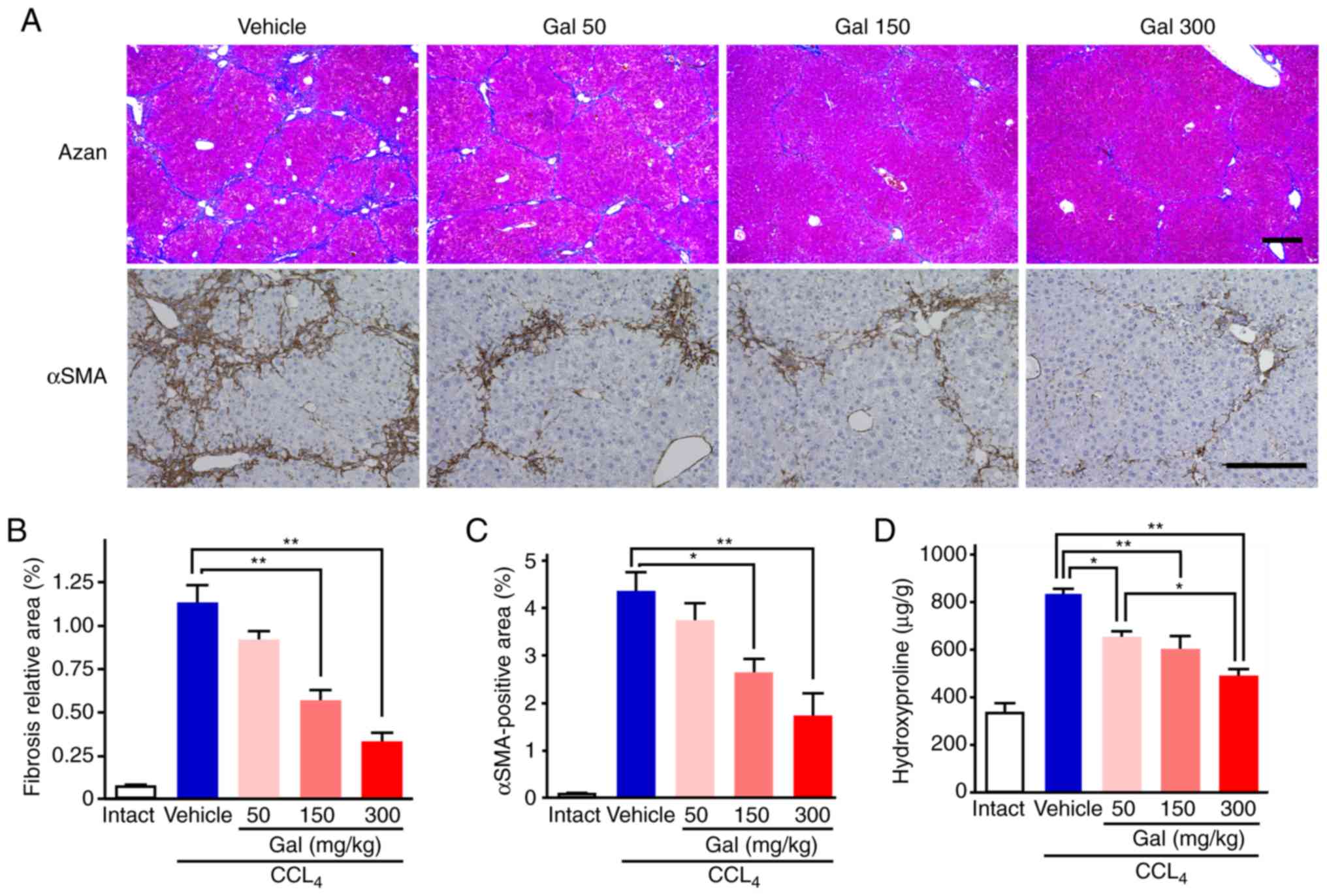
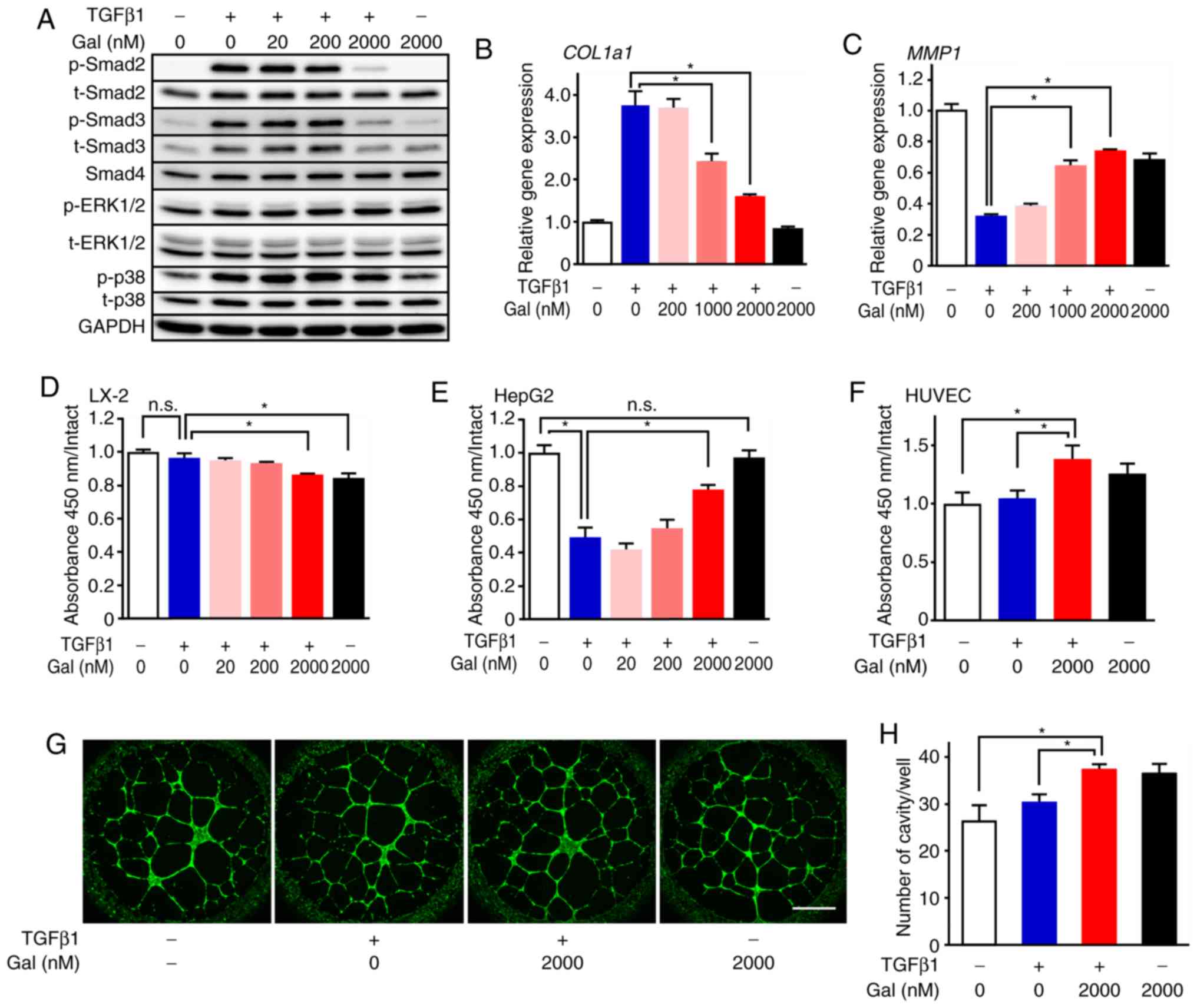 | Figure 1Anti-fibrotic and proliferative
effects of galunisertib. (A) Effects of galunisertib on TGF-β
signaling in LX-2 cells (n=4). p-Smad2, Smad3, p-Smad3, ERK1/2,
p-ERK1/2, p38 MAPK, p-p38 and Smad4 protein expression levels were
evaluated by western blot analysis. Galunisertib treatment
dose-dependently decreased p-Smad2 and p-Smad3 expression levels,
but not those of p-ERK1/2 and p-p38; (B and C) Effects of
galunisertib on collagen and MMP production in LX-2 cells (n=4).
(B) COL1a1 and (C) MMP1 mRNA expression levels were
evaluated by reverse transcription-quantitative PCR. Galunisertib
treatment dose-dependently inhibited and promoted Col1a1 and
MMP1 mRNA expression, respectively. The effects of
galunisertib on (D) LX-2, (E) HepG2 and (F) HUVEC cell
proliferation (n=4-6). Galunisertib treatment inhibited LX-2 cell
proliferation and promoted HepG2 and HUVEC proliferation even in
the presence of TGF-β1 stimulation. (G and H) Effects of
galunisertib in a tube formation assay with HUVEC (n=4). The number
of the cavities significantly increased following galunisertib
treatment. *P<0.05. Error bars represent the means ±
SEM. Scale bar=1 mm. n.s., not significant; Gal, galunisertib; ERK,
extracellular signal-regulated kinase; p-, phosphorylated; MMP,
matrix metalloproteinase; COL1a1, collagen type I alpha 1 chain;
TGF-β, transforming growth factor-β. |
Effect of galunisertib on collagen and
MMP synthesis, cell proliferation and angiogenesis
Galunisertib inhibited mRNA expression of collagen
type I alpha 1 chain (COL1a1) in LX-2 cells in a dose-dependent
manner (Fig. 1B). Conversely,
galunisertib treatment dose-dependently restored MMP1 expression
that was suppressed by TGF-β1 (Fig.
1C). Galunisertib treatment also dose-dependently inhibited
LX-2 cell proliferation even in the presence of TGF-β1 stimulation
(Fig. 1D). In addition,
galunisertib treatment dose-dependently promoted HepG2 and HUVEC
proliferation even in the presence of TGF-β1 stimulation (Fig. 1E and F). Furthermore, tube
formation by HUVECs was promoted by galunisertib, as evidenced by
the significant increase in the number of cavities observed
following 2,000 nM galunisertib treatment (Fig. 1G and H).
Galunisertib treatment inhibits liver
fibrosis progression in CCl4-induced fibrotic liver
In gross appearance, the livers of mice in the
vehicle-treated group had a dark red color, an irregular surface,
and were more rigid compared with the intact group. Concomitantly,
livers from the galunisertib-treated mice, particularly those in
the high-dose group, were bright pink and had only a slightly
irregular surface and little rigidity (Figs. 2A and S3). Galunisertib treatment increased
the liver/ body weight (BW) ratio in a dose-dependent manner
compared with the vehicle group [4.6±0.23% (vehicle) vs. 5.14±0.18%
(low), 5.25±.33% (middle) and 6.11±0.27% (high); Fig. 2B]. In particular, the ratio was
increased 1.7-fold in the high-dose group compared with the intact
group (6.11±0.27 vs. 3.63±0.15%; Fig.
2B).
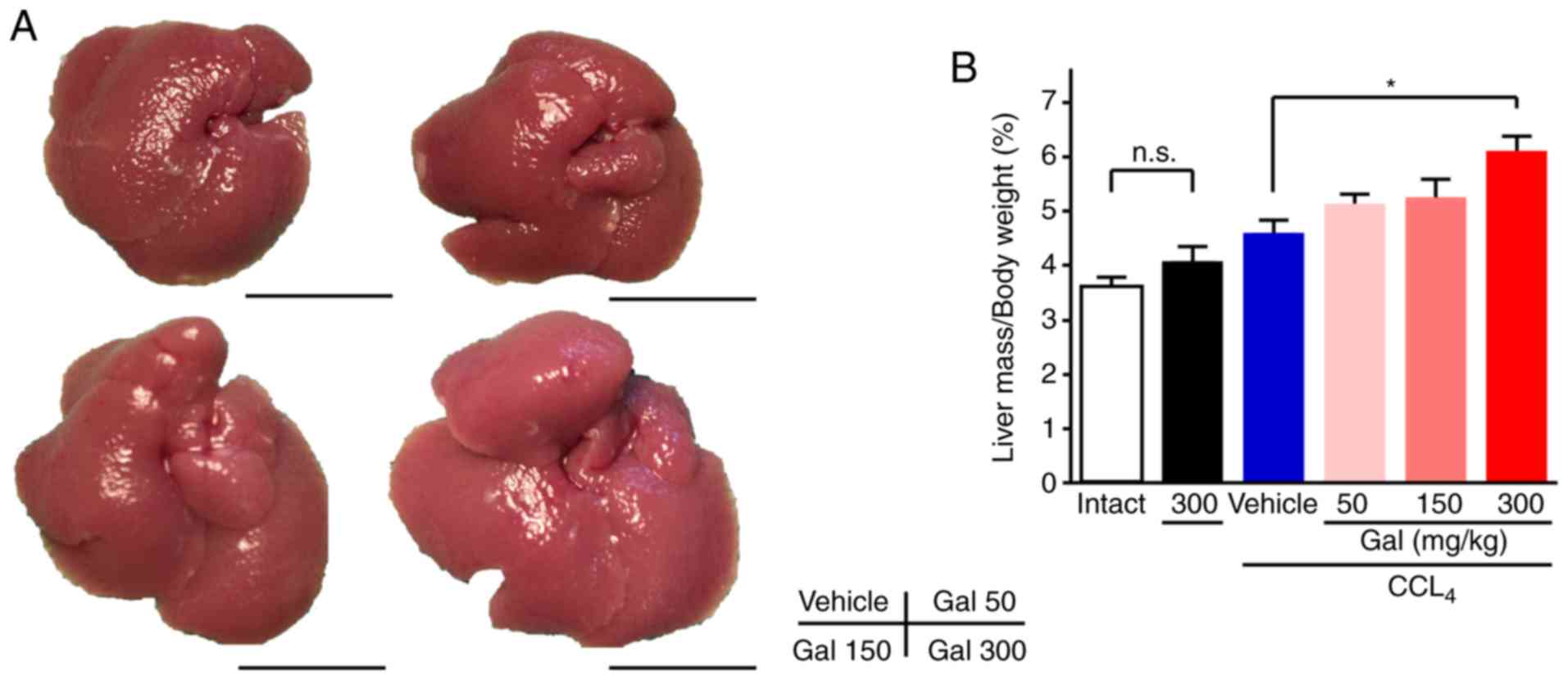 | Figure 2Gross appearance and liver/body
weight ratio for mice treated with galunisertib. (A) Livers from
CCl4-treated mice exhibited a dark red color, an
irregular surface and increased rigidity compared with livers from
the intact group. Livers from the galunisertib-treated mice
exhibited a bright pink color, a slightly irregular surface and
minimal rigidity. (B) The ratio of liver/body weight for mice
treated with galunisertib was dose-dependently and significantly
increased compared with that for mice treated with vehicle.
*P<0.05. Error bars represent the means ± SEM. Scale
bar=1 mm. Gal, galunisertib; CCl4, carbon tetrachloride;
Intact, no CCl4 or galunisertib treatment; Vehicle,
CCl4-treated with no galunisertib treatment; Gal 50,
livers from mice treated with low-dose galunisertib; Gal 150,
treatment with middle-dose galunisertib; Gal 300, treatment with
high-dose galunisertib; n.s., not significant. |
As demonstrated by the Mallory's Azan staining
results, the extent of the fibrosis was attenuated remarkably in
liver sections from mice treated with galunisertib compared with
those treated with the vehicle (Fig.
3A). Semi-quantitative analysis indicated that the size of the
fibrotic area was dose-dependently decreased with galunisertib
treatment compared with vehicle group [1.13±0.11% (vehicle) vs.
0.91±0.06% (low), 0.55±0.08% (middle) and 0.33±.06% (high);
Fig. 3B]. In addition, the
galunisertib-treated livers exhibited fewer α-SMA-positive cells
compared with the vehicle-treated livers (Fig. 3A). Semi-quantitative analysis of
immunohistochemical staining intensity using anti-α-SMA antibody
indicated that the sizes of the α-SMA-positive areas in mice
treated with galunisertib were decreased in a dose-dependent manner
compared with those in mice in the vehicle group [4.32±0.43%
(vehicle) vs. 3.72±0.38% (low), 2.62±0.31% (middle) and 1.71±0.49%
(high); Fig. 3C].
Hydroxyproline content, which is associated with the
amount of collagen, in liver samples was increased ~2.1-fold by
CCl4 administration during the 8-week treatment period
compared with the intact liver samples 849.7±52.1 (vehicle) vs.
408.9±19.4 (intact) (Fig. 3D).
All galunisertib doses produced statistically significant decreases
in the hydroxyproline content compared with that in the vehicle
group, and the high-dose galunisertib treatment group exhibited a
~40% decrease in hydroxyproline content compared with the vehicle
group [489.0±32.2 (high) vs. 849.7±52.1 µg/g (vehicle);
Fig. 3D].
The results from the RT-qPCR demonstrated that there
were no decreases in the expression levels of Col1a1,
fibronectin 1 (Fn1) and actin alpha 2, smooth muscle
(Acta2) mRNA after 8 weeks of CCl4 treatment. The
mRNA levels of these genes were also similar between mice treated
with vehicle and all dosages of galunisertib (Fig. S4A-C).
Galunisertib treatment activated MMP in
CCl4-induced fibrotic liver
The MMP-1 expression level in the vehicle-treated
group was significantly decreased compared with the level in the
intact group, as shown by the data from the western blot analysis
(Figs. 4A and S5) and RT-qPCR assays (Fig. S6A). In contrast to the in
vitro results, the MMP-1 expression levels after 8 weeks of
CCl4 treatment were similar in samples from the
galunisertib- and vehicle-treated livers. Semi-quantitative western
blot analysis also revealed that, after 8 weeks of CCl4
treatment, the expression levels of activated MMP-13 were similar
in liver samples from galunisertib- and vehicle-treated mice
(Figs. 4A and S5). Quantification of MMP-2 and MMP-9
expression by gelatin zymography indicated that the expression
levels of pro- and active MMP-9, but not MMP-2, were significantly
upregulated in galunisertib-treated livers compared with those in
the vehicle-treated livers (Figs.
4B-F and S5D). Furthermore,
RT-qPCR analysis indicated that the level of Mmp9 mRNA
expression after 8 weeks of CCl4 treatment was
significantly upregulated in livers from mice in the high-dose
galunisertib group compared with those from the vehicle-treated
group; however, there was no significant upregulation in expression
levels of Mmp1b, Mmp2, Mmp13 or TIMP
metalloproteinase inhibitor 1 mRNA (Figs. 4G-J and S6C).
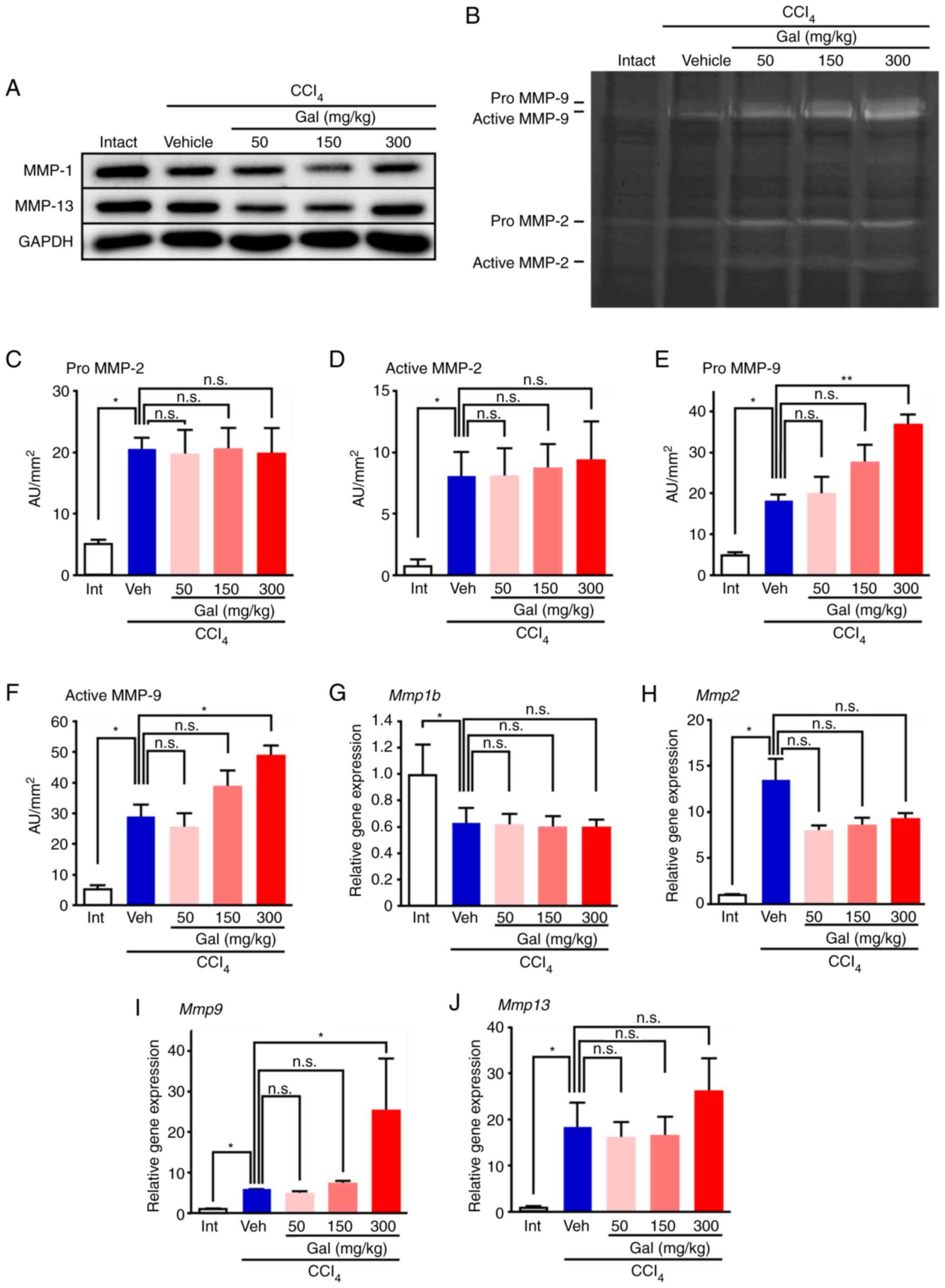 | Figure 4MMP expression and activity in
cirrhotic livers from mice treated with galunisertib. (A) Western
blot analysis data show that MMP-1 and MMP-13 expression levels
were not increased in the livers from galunisertib-treated mice
compared with vehicle-treated mice. (B) Gelatin zymography showing
the active- and pro-forms of MMP-2 and MMP-9 after 8 weeks of
CCl4 treatment. The levels of the active and pro-forms
of MMP-9 were significantly upregulated in livers from
galunisertib-treated mice compared with those from vehicle-treated
mice. (C-F) For semi-quantitative analyses, gelatin zymography
bands of (C) pro-form of MMP-2, (D) active form of MMP-2, (E)
pro-form of MMP-9 and (F) active form of MMP-9 were analyzed using
image analysis software. (G-J) qRT-PCR analysis of Mmp mRNA
expression. The levels of (G) Mmp1b and (H) Mmp2 were
similar between vehicle-treated mice and galunisertib-treated mice.
(I) Mmp9 mRNA expression was significantly upregulated in
livers from mice treated with high-dose galunisertib compared with
those from vehicle-treated livers. (J) Mmp13 mRNA expression
was similar between vehicle-treated mice and galunisertib-treated
mice. For semi-quantitative analysis of gelatin zymography, 2 gels
were assessed (n=4 in each group). The samples were derived from
the same experiment and the gels were processed in parallel.
*P<0.05 and **P<0.01. Error bars
represent the means ± SEM. n.s., not significant; Gal,
galunisertib; CCl4, carbon tetrachloride; Intact or Int,
no CCl4 or galunisertib treatment; Vehicle or Veh,
CCl4 treatment without galunisertib; AU, arbitrary unit;
MMP, matrix metalloproteinase. |
Galunisertib treatment promoted liver
regeneration in CCl4-induced fibrotic liver
In the galunisertib treatment groups, the percentage
of PCNA-positive hepatocytes observed in the high-power fields
increased in a dose-dependent manner (Fig. 5A). The number of PCNA-positive
hepatocytes increased around the portal vein, but not the central
vein. In the high-dose galunisertib treatment group, the percentage
of PCNA-positive hepatocytes increased by ~3-fold compared with
that in the vehicle group [29.1±4.47 (high) vs. 10.2±2.71
(vehicle); Fig. 5B]. Western blot
analysis of the levels of the PCNA protein in galunisertib-treated
livers also demonstrated dose-dependent increases compared with the
vehicle group (Figs. 5E and
S7). By contrast, in the liver
samples from the galunisertib-treated groups, the percentage of
TUNEL-positive hepatocytes and the number of cleaved
caspase-3-positive hepatocytes observed in high power fields was
similar to that for samples from vehicle-treated mice (Figs. 5C and S8). The proliferation/apoptosis (PA)
index for hepatocytes from mice treated with high-dose galunisertib
was significantly increased compared with that from mice treated
with the vehicle [4.70±1.36 (high) vs. 1.37±0.48 (vehicle);
Fig. 5D]. To further evaluate the
levels of cell proliferation and apoptosis, immunohistochemistry
using an anti-p-AKT antibody was performed, but no signals were
detected (data not shown). In the mice treated with high-dose
galunisertib, the number of PCNA+/isolectin
B4+ double-positive liver endothelial cells was
significantly increased after 8 weeks of CCl4 treatment
compared with those from the vehicle-treated mice [22.7±4.65 (high)
vs. 14.4±2.61 (vehicle); Fig. 5F and
G].
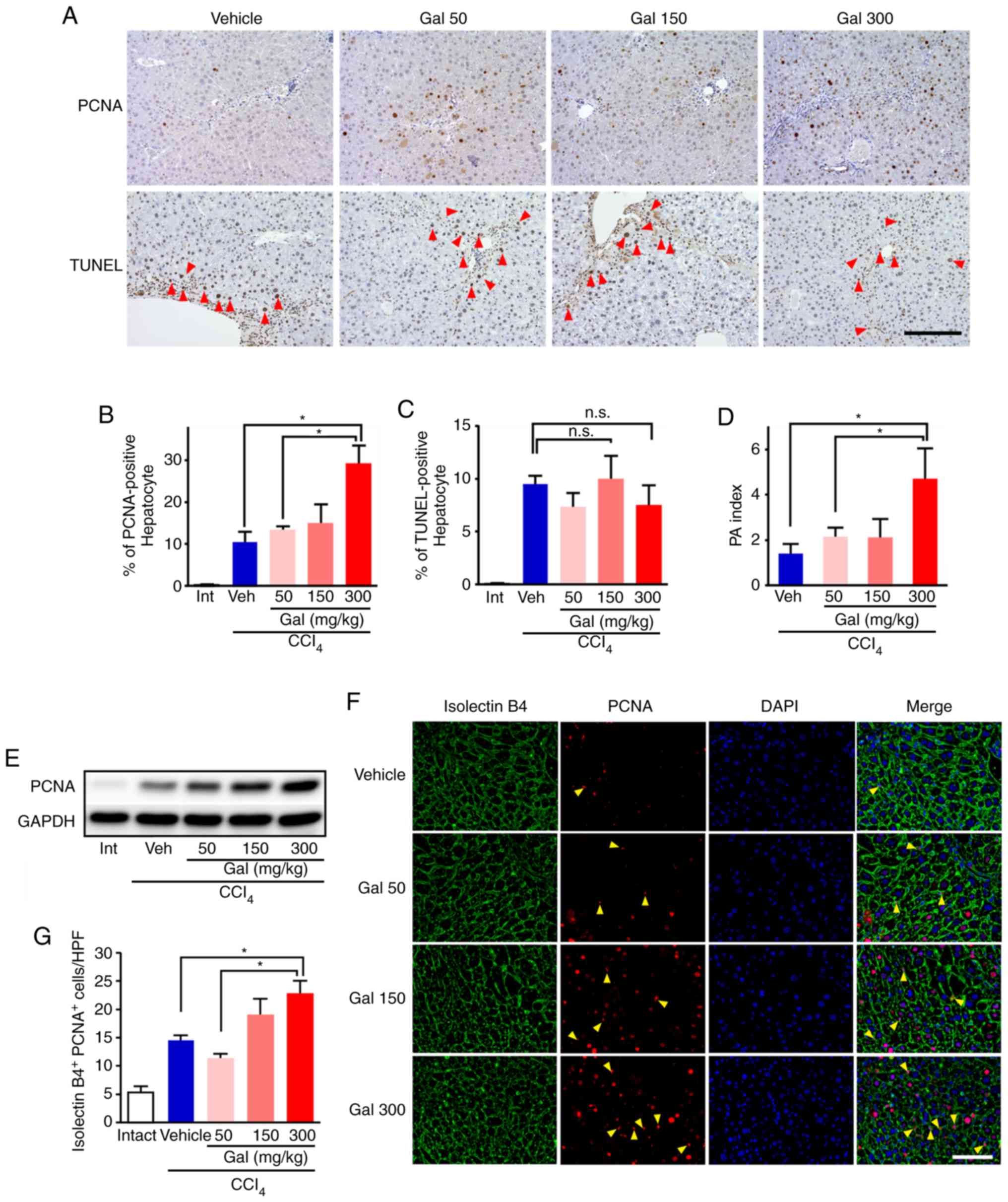 | Figure 5Galunisertib treatment accelerated
proliferation of hepatocytes and endothelial cells. (A) In the
galunisertib-treated mice, the numbers of PCNA-positive hepatocytes
increased dose-dependently (upper panels). Concomitantly, the
number of TUNEL-positive hepatocytes (arrowhead) was similar in
galunisertib- and vehicle-treated mice (lower panels). Scale
bar=200 µm; (B and C) For semi-quantitative analyses, (B)
PCNA- and (C) TUNEL-positive hepatocytes were assessed using image
analysis software. The percentage of PCNA-positive hepatocytes
dose-dependently increased in livers from mice treated with
galunisertib compared with those treated with vehicle, but the
percentage of TUNEL-positive hepatocytes was similar in both
groups. (D) The proliferation/apoptosis (PA) index was calculated
by % PCNA-positive hepatocytes/% TUNEL-positive hepatocytes. In
mice treated with high-dose galu-nisertib, the PA index was
significantly increased compared with vehicle-treated mice. (E)
PCNA protein expression levels in livers from galunisertib-treated
mice exhibited dose-dependent increases. (F) In
galunisertib-treated mice the number of PCNA+ (red) and
isolectin B4+ (green) cells (arrowhead) increased in a
dose-dependent manner. Scale bar=400 µm. (G) For
semi-quantitative analyses, PCNA+ isolectin
B4+ cells were assessed using image analysis software.
In livers from galunisertib-treated mice, the number of
PCNA+ isolectin B4+ cells was significantly
increased compared with that for the vehicle-treated mice.
*P<0.05. Error bars represent the means ± SEM. Gal,
galunisertib; CCl4, carbon tetrachloride; Intact or Int,
no CCl4- or galunisertib treatment; Vehicle or Veh,
CCl4-treated without galunisertib treatment; Gal 50,
low-dose galunisertib; Gal 150, middle-dose galunisertib; Gal 300,
high-dose galunisertib; PCNA, proliferating cell nuclear antigen;
TUNEL, terminal deoxynucleotidyl transferase mediated d-UTP nick
end labeling; n.s., not significant. |
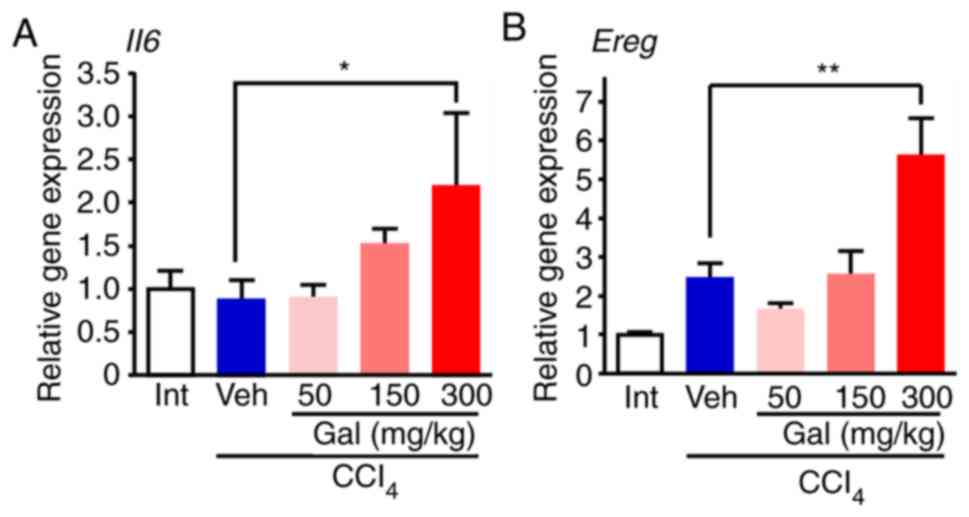
For the growth factors that are relevant to
hepatocytes, the RT2 profiler PCR
array indicated that the expression levels of interleukin-6
(Il6), epiregulin (Ereg), fibroblast growth factor-7
(Fgf7), epithelial growth factor (Egf) and hepatocyte
growth factor (Hgf) were upregulated in the livers from the
high-dose galunisertib-treated group compared with the livers from
the vehicle-treated group (Table
SII). RT-qPCR analysis verified that Il6 and Ereg
expression levels were significantly increased in the livers from
the high-dose galunisertib-treated group compared with the
vehicle-treated group (Fig. 6A and
B). Concomitantly, the RT-qPCR analysis of the mRNA expression
levels of other growth factors, including Fgf7, Egf, Hgf,
demonstrated that there were no significant differences between the
experimental groups (Fig.
S9A-C).
Toxic effects of galunisertib on liver
function in mice with and without CCl4 treatment
To evaluate the potential toxic effects of
galunisertib, mice were treated with 300 mg/kg/day galunisertib
(high-dose) for 4 weeks without CCl4 administration and
compared with the intact group. Although the serum levels of AST,
ALT and total bilirubin between the two groups did not
significantly change, those for total protein and albumin were
significantly decreased in mice treated with high-dose galunisertib
(Table I). Histologically, no
significant differences were observed between the two groups (data
not shown).
 | Table ISerum hepatobiliary parameters. |
Table I
Serum hepatobiliary parameters.
| Groups | Total protein,
g/dl | Albumin, g/dl | AST, IU/ml | ALT, IU/ml | Total bilirubin,
mg/dl |
|---|
| Intact | 5.38±0.05 | 3.75±0.03 | 45.00±2.36 | 21.25±0.85 | 0.07±0.00 |
| High-dose without
CCl4 | 4.90±0.08a | 3.28±0.03a | 67.75±16.15 | 61.50±30.30 | 0.10±0.02 |
| Vehicle | 5.40±0.04 | 3.54±0.05 | 130.43±41.55 | 124.71±8.63 | 0.16±0.02 |
| Low-dose | 5.54±0.04 | 3.59±0.05 | 120.00±40.71 | 112.71±12.22 | 0.16±0.04 |
| Middle-dose | 5.34±0.06 | 3.47±0.08 | 220.43±59.18 | 160.14±32.19 | 0.19±0.02 |
| High-dose | 5.28±0.09 | 3.28±0.09b | 255.75±62.71 | 203.25±66.17 | 0.17±0.00 |
Next, the effects of galunisertib in fibrotic liver
induced by CCl4 treatment were investigated. No loss of
BW was observed for any of the galunisertib treatment groups (data
not shown). There were also no significant differences in the
levels of serum total protein, AST, ALT and total bilirubin between
the galunisertib-treated and vehicle groups, although the serum
albumin level was significantly decreased in the high-dose group
compared with the vehicle group (Table I).
Discussion
The present study showed that TGF-β1 treatment in
LX-2 cells induced the phosphorylation of Smad2 and Smad3, and
increased the expression of fibrosis genes. These changes could be
inhibited by treatment with galunisertib at dosages that were in
the range of plasma concentrations in human clinical trials
(16,17). Similarly, it was shown in
proliferating HepG2 cells that TGF-β1-induced Smad2 phosphorylation
could be reversed by galunisertib. In addition, histology and
measurement of liver hydroxyproline content were used to
demonstrate that galunisertib dose-dependently suppressed liver
fibrosis in mice with liver cirrhosis induced by CCl4
treatment. These data strongly suggested that the TGF-β receptor
type I kinase inhibitor galunisertib would be a feasible and
effective anti-TGF-β therapy.
Galunisertib has been reported to affect the
activity of TGF-β type-I receptors ALK4 and ALK5 and markedly
inhibit the phosphorylation of Smad2 and Smad3 (10), and ERK1/2 phosphorylation
(18). Conversely, TGF-β
stimulation promotes phosphorylation of p38 MAPK independently of
TGF-β receptor type-1 kinases (19). The present study examined the
effects of galunisertib on intracellular signaling in HSC line
LX-2. The results showed that galunisertib treatment of LX-2 cells
markedly and dose-dependently decreased p-Smad2 and p-Smad3
expression levels, but did not affect p-p38 MAPK and p-ERK1/2
levels, even under activated conditions of TGF-β signaling. ERK1/2
was phosphorylated in the absence of TGF-β1 stimulation, which also
did not promote ERK1/2 phosphorylation. These results suggested
that phosphorylation of ERK1/2 is not regulated by TGF-β1 signaling
in LX-2 cells. HSCs are fibrogenic cells that make major
contributions to collagen accumulation during chronic liver
disease. TGF-β/Smad3 signaling in LX-2 cells not only enhances the
production of ECM, but also inhibits expression of MMP-1, which is
the primary interstitial collagenase during fibrolysis (20). Indeed, the present study
identified that galunisertib treatment inhibited the mRNA
expression levels of COL1a1 and increased those of
MMP1 in LX-2 cells in a dose-dependent manner. It was also
shown that galunisertib treatment promoted cell proliferation of
the human hepatoblastoma HepG2 cell line, even in the presence of
TGF-β stimulation, which typically inhibits hepatocyte
proliferation (5). In HCC
tumorigenesis, TGF-β serves a dual role through tumor suppressive
properties during the early disease stage and promotion of tumor
progression at later stages of disease (21). Among HCC cell lines, HepG2, HuH7
and Hep3B cells belong to the early TGF-β signature subgroup of HCC
models (22). As such, as an
early TGF-β signature HCC cell line, HepG2 cells exhibit
antiproliferative effects of TGF-β. Indeed, Serova et al
(18) indicated that HepG2 cells
exhibited the most marked TGF-β-mediated growth inhibition among 7
HCC cell lines. Together, these results suggested that galunisertib
inhibits the canonical TGF-β signaling pathway in vitro.
Fibrolysis mechanisms involve the inhibition of ECM
production and/or increase of collagenolytic activity (4). HSCs serve a central role in liver
fibrogenesis (4). In mice with
CCl4-induced cirrhosis, the present study showed that
galunisertib treatment dose-dependently decreased the percentage of
liver fibrotic areas and α-SMA-positive areas, as well as the
hydroxyproline content, compared with the vehicle-treated mice.
α-SMA is a well-known marker of activated HSCs and increased ECM
production is mediated by activated HSCs (23). However, the present study
identified that none of the administered doses of galunisertib
significantly decreased Col1a1, Fn1 and Acta2 mRNA
expression levels compared with the vehicle group. This result
could be attributed to the short (0.3 h) half-life of galunisertib
in mice (10) and could be
affected by the sample collection time. Furthermore, it was
identified that the activity of MMP-9 was increased in a
dose-dependent fashion following galunisertib administration.
Following activation by MMP-2, MMP-9 degrades fibronectin and
type-IV collagen, which are both major structural components of
basement membranes (24). The
results of the present study are consistent with those of a study
by Hammad et al (25),
which demonstrated that the amount of ECM/stromal components
fibronectin and laminin-332, as well as the activity of the
carcinogenic β-catenin pathway, is remarkably decreased in
galunisertib-treated Abcb4Ko mice, another mouse model of chronic
liver disease. Together, these data indicate that galunisertib
administration can promote fibrolysis. Meanwhile, MMP-1 in humans
and MMP-13 in mice have been shown to serve as the major MMPs that
degrade interstitial collagens (26-28). The present study showed that
galunisertib treatment dose-dependently increased MMP-1 expression
in LX-2 cells under TGF-β1 stimulation, but that MMP-13 levels were
similar in livers from galunisertib-treated and vehicle-treated
mice. Therefore, in mice, the fibrolytic action of galunisertib
appears to be mediated through the degradation of collagen, which
is a primary component of basement membranes.
Notably, enhanced regeneration of hepatocytes was
detected by immunohistochemistry and western blot analysis using an
anti-PCNA antibody. This result is consistent with that of
Karkampouna et al (29),
who reported that the co-administration of LY364947, an inhibitor
that targets the TGF-β type-I receptor kinase ALK5, with
CCl4 resulted in enhanced hepatic regeneration, as
measured by changes in PCNA levels. In addition, TGF-β regulates
the G1 to S phase transition of hepatocytes, but intact
TGF-β-mediated signaling is not required to inhibit liver
regeneration, as compensatory increases in activin A signaling can
occur (30).
Galunisertib is in Phase II trials for treatment of
liver cancer (NCT01246986 and NCT02178358). Therefore, the present
study explored the effect of clinical stage on the activity of
galunisertib as a TGF-β pathway inhibitor. The in vivo
antifibrotic and regenerative efficacy of galunisertib using a dose
of 75 mg/kg administered twice daily by oral gavage was evaluated.
This dosing schedule was defined by the PK/PD profile, as described
previously (11). Ongoing
clinical trials involving galunisertib are testing doses of 160 or
300 mg/day (BID) (NCT01246986 and NCT02178358). Therefore, the low-
and moderate-doses of galunisertib used in the present study
correspond to clinically relevant doses undergoing testing in human
clinical trials. However, 300 mg/kg/day galunisertib is considered
to be a toxic dose, and it has been already reported that
continuous administration of galunisertib at 300 mg/kg/day for 3
months causes side effects (31).
The present study identified that, in comparison
with the untreated mice, 300 mg/kg/day galunisertib was associated
with a significant increase in liver weight as well as a
significant decrease in albumin levels; the ALT and AST levels were
also increased, although not significantly, when compared with the
untreated mice. Concomitantly, albumin levels were also
significantly decreased in mice treated with 300 mg/kg/day
galunisertib that were not exposed to CCl4 as compared
with the intact group. To demonstrate that the observed
hepatomegaly was not hepatotoxic, the association between cell
proliferation and apoptosis during progression of
CCl4-induced liver fibrosis was examined. In liver
samples from mice treated with 300 mg/kg/day galunisertib, the
percentage of TUNEL-positive hepatocytes did not increase, but the
PA index was significantly increased compared with the hepatocytes
from the vehicle-treated mice. These results suggest that the
increase of liver weight may be attributed to an promotion of the
hepatocyte proliferation. Previous studies indicated that liver
weight was either maintained or increased following inhibition of
TGF-β and Activin A in a partial hepatectomy model and in normal
livers (32,33). Galunisertib inhibits the activity
of both ALK4 and ALK5, but may also inhibit the activin A signaling
pathway (11,34). Inhibition of both TGF-β/ALK5 and
Activin A/ALK4 signaling pathways may be associated with the
increased liver weight observed in mice treated with
galunisertib.
The effects of ALK5 inhibitors on angiogenesis have
recently been reported (35). In
the present study, the effects of galunisertib on liver fibrosis
associated with angiogenesis were evaluated using a cell
proliferation and tube formation assay with HUVEC and a mouse model
of angiogenesis. It was identified that galunisertib treatment
promoted angiogenesis by HUVEC regardless of TGF-β stimulation.
This result is consistent with that of Liu et al (35), who showed that a combination of
galunisertib and VEGF more potently induced angiogenesis compared
with VEGF alone. In the in vitro study, both the
proliferation and tube formation assays using HUVEC were performed
in the presence of several growth factors including VEGF. In
addition, our previous studies suggested that the VEGF levels in
liver tissue are elevated in the CCl4-induced cirrhosis
model (15,36). Taken together, these results
indicate that galunisertib treatment promoted angiogenesis in the
CCl4-induced cirrhosis model.
Another explanation for galunisertib-mediated
resolution of fibrosis and promotion of hepatic regeneration is an
increase in growth factor expression following galunisertib
administration. The results of the present study showed that
galunisertib treatment significantly upregulated the mRNA
expression levels of both Il6 and Ereg in liver
tissues. IL-6 is a cytokine that is primarily produced by T cells
and macrophages, and is also involved in inflammation and the
development of certain immune system diseases, in addition to its
ability to promote hepatocyte proliferation (37). TGF-β also has an immunosuppressive
effect. Therefore, inhibition of the TGF-β pathway by galunisertib
may affect IL-6 secretion by immune cells. Epiregulin is expressed
by the Ereg gene in Thy-1-positive cells, for example
mesenchymal cells or liver progenitor cells, and has been reported
to contribute to liver regeneration (38). Increases in epiregulin expression
may in turn increase the number of hepatic progenitor cells,
although the results of promoting hepatocyte proliferation support
that galunisertib-mediated inhibition of the TGF-β pathway may be
the main driver of increased proliferation of mature hepatocytes.
These possibilities require further investigation.
In terms of side effects, CCl4-treated
mice that were administered galunisertib exhibited an increased
liver/BW ratio, whereas mice that received only galunisertib and no
CCl4 showed no significant change in this ratio (data
not shown). These results appear reflect hepatic regeneration;
however, atypical increases in liver weight is known to be a
transient effect that can arise due to the inhibition of the TGF-β
and Activin A signaling pathways (33). In the biochemical analysis of
serum samples, the levels of total protein and albumin were
significantly decreased by high-dose galunisertib treatment without
CCl4 administration, in comparison with the intact
group. The cause of this effect is not clear, but no obvious
histological differences were observed between the two groups.
In conclusion, the present study demonstrated that
anti-TGF-β therapy using galunisertib ameliorated liver fibrosis by
decreasing the number of activated HSCs and increasing the activity
of MMP-9, as well as enhancing hepatic regeneration by elevating
growth factor levels, including Il6 and Ereg. Phase
II clinical trials of galunisertib for treatment of HCC are
currently ongoing. The majority of patients with HCC also have
liver cirrhosis, and thus determination of both the tolerability
and safety of galunisertib, as well as its ability to induce
fibrolysis and liver regeneration, is important. The results of the
present study may provide insights into the development of novel
therapies for liver cirrhosis.
Supplementary Data
Funding
The present study was supported in part by a grant
from the Kurume University Millennium Box Foundation for the
Promotion of Science and by the Medical Care Education Research
Foundation.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TN, HK and TTo made substantial contributions to the
conception and design of the study. AM and TN conceived and
designed the study, performed the experiments and wrote the
manuscript. MA and HI performed the experiments. TS, TTa and HS
performed the data analysis. All authors edited and reviewed the
final manuscript.
Ethics approval and consent to
participate
All experiments were approved by the Ethics Review
Committee for Animal Experimentation of the Kurume University
School of Medicine and were performed in accordance with Eli Lilly
and company animal care and use requirements for animal researchers
and suppliers.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors thank Ms. Mari Hagihara (Division of
Gastroenterology, Department of Medicine, School of Medicine,
Kurume University, Kurume, Japan) for technical assistance in the
preparation of tissue sections and Ms. Masako Hayakawa (Liver
Cancer Research Division, Research Center for Innovative Cancer
Therapy, Kurume University, Kurume, Japan) for excellent technical
assistance.
References
|
1
|
Schuppan D and Afdhal NH: Liver cirrhosis.
Lancet. 371:838–851. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fattovich G, Stroffolini T, Zagni I and
Donato F: Hepatocellular carcinoma in cirrhosis: Incidence and risk
factors. Gastroenterology. 127(5 Suppl 1): S35–S50. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tarao K, Nozaki A, Ikeda T, Sato A,
Komatsu H, Komatsu T, Taguri M and Tanaka K: Real impact of liver
cirrhosis on the development of hepatocellular carcinoma in various
liver diseases-meta-analytic assessment. Cancer Med. 8:1054–1065.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tsuchida T and Friedman SL: Mechanisms of
hepatic stellate cell activation. Nat Rev Gastroenterol Hepatol.
14:397–411. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nguyen LN, Furuya MH, Wolfraim LA, Nguyen
AP, Holdren MS, Campbell JS, Knight B, Yeoh GC, Fausto N and Parks
WT: Transforming growth factor-beta differentially regulates oval
cell and hepatocyte proliferation. Hepatology. 45:31–41. 2007.
View Article : Google Scholar
|
|
6
|
Nakamura T, Sakata R, Ueno T, Sata M and
Ueno H: Inhibition of transforming growth factor beta prevents
progression of liver fibrosis and enhances hepatocyte regeneration
in dimethylnitro-samine-treated rats. Hepatology. 32:247–255. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ueno H, Sakamoto T, Nakamura T, Qi Z,
Astuchi N, Takeshita A, Shimizu K and Ohashi H: A soluble
transforming growth factor beta receptor expressed in muscle
prevents liver fibrogenesis and dysfunction in rats. Hum Gene Ther.
11:33–42. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nakamura T, Ueno T, Sakamoto M, Sakata R,
Torimura T, Hashimoto O, Ueno H and Sata M: Suppression of
transforming growth factor-beta results in upregulation of
transcription of regeneration factors after chronic liver injury. J
Hepatol. 41:974–982. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yingling JM, Blanchard KL and Sawyer JS:
Development of TGF-beta signalling inhibitors for cancer therapy.
Nat Rev Drug Discov. 3:1011–1022. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Herbertz S, Sawyer JS, Stauber AJ,
Gueorguieva I, Driscoll KE, Estrem ST, Cleverly AL, Desaiah D, Guba
SC, Benhadji KA, et al: Clinical development of galunisertib
(LY2157299 monohydrate), a small molecule inhibitor of transforming
growth factor-beta signaling pathway. Drug Des Devel Ther.
9:4479–4499. 2015.PubMed/NCBI
|
|
11
|
Yingling JM, McMillen WT, Yan L, Huang H,
Sawyer JS, Graff J, Clawson DK, Britt KS, Anderson BD, Beight DW,
et al: Preclinical assessment of galunisertib (LY2157299
monohydrate), a first-in-class transforming growth factor-β
receptor type I inhibitor. Oncotarget. 9:6659–6677. 2017.
View Article : Google Scholar
|
|
12
|
Ikeda M, Morimoto M, Tajimi M, Inoue K,
Benhadji KA, Lahn MMF and Sakai D: A phase Ib study of transforming
growth factor-beta receptor I inhibitor galunisertib in combination
with sorafenib in Japanese patients with unresectable
hepatocellular carcinoma. Invest New Drugs. 37:118–126. 2019.
View Article : Google Scholar
|
|
13
|
Luangmonkong T, Suriguga S, Bigaeva E,
Boersema M, Oosterhuis D, de Jong KP, Schuppan D, Mutsaers HAM and
Olinga P: Evaluating the antifibrotic potency of galunisertib in a
human ex vivo model of liver fibrosis. Br J Pharmacol.
174:3107–3117. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
15
|
Nakamura T, Torimura T, Sakamoto M,
Hashimoto O, Taniguchi E, Inoue K, Sakata R, Kumashiro R, Murohara
T, Ueno T and Sata M: Significance and therapeutic potential of
endothelial progenitor cell transplantation in a cirrhotic liver
rat model. Gastroenterology. 133:91–107.e101. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gueorguieva I, Cleverly AL, Stauber A,
Sada Pillay N, Rodon JA, Miles CP, Yingling JM and Lahn MM:
Defining a therapeutic window for the novel TGF-β inhibitor
LY2157299 monohydrate based on a pharmacokinetic/pharmacodynamic
model. Br J Clin Pharmacol. 77:796–807. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Faivre S, Santoro A, Kelley RK, Gane E,
Costentin CE, Gueorguieva I, Smith C, Cleverly A, Lahn MM, Raymond
E, et al: Novel transforming growth factor beta receptor I kinase
inhibitor galunisertib (LY2157299) in advanced hepatocellular
carcinoma. Liver Int. 39:1468–1477. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Serova M, Tijeras-Raballand A, Dos Santos
C, Albuquerque M, Paradis V, Neuzillet C, Benhadji KA, Raymond E,
Faivre S and de Gramont A: Effects of TGF beta signalling
inhibition with galunisertib (LY2157299) in hepatocellular
carcinoma models and in ex vivo whole tumor tissue samples from
patients. Oncotarget. 6:21614–21627. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Heldin CH and Moustakas A: Signaling
receptors for TGF-β family members. Cold Spring Harb Perspect Biol.
8:a0220532016. View Article : Google Scholar
|
|
20
|
Yuan W and Varga J: Transforming growth
factor-beta repression of matrix metalloproteinase-1 in dermal
fibroblasts involves Smad3. J Biol Chem. 276:38502–38510. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Breuhahn K, Longerich T and Schirmacher P:
Dysregulation of growth factor signaling in human hepatocellular
carcinoma. Oncogene. 25:3787–3800. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Coulouarn C, Factor VM and Thorgeirsson
SS: Transforming growth factor-beta gene expression signature in
mouse hepatocytes predicts clinical outcome in human cancer.
Hepatology. 47:2059–2067. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bataller R and Brenner DA: Liver fibrosis.
J Clin Invest. 115:209–218. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ram M, Sherer Y and Shoenfeld Y: Matrix
metalloproteinase-9 and autoimmune diseases. J Clin Immunol.
26:299–307. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hammad S, Cavalcanti E, Werle J, Caruso
ML, Dropmann A, Ignazzi A, Ebert MP, Dooley S and Giannelli G:
Galunisertib modifies the liver fibrotic composition in the Abcb4Ko
mouse model. Arch Toxicol. 92:2297–2309. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Iimuro Y, Nishio T, Morimoto T, Nitta T,
Stefanovic B, Choi SK, Brenner DA and Yamaoka Y: Delivery of matrix
metal-loproteinase-1 attenuates established liver fibrosis in the
rat. Gastroenterology. 124:445–458. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Endo H, Niioka M, Sugioka Y, Itoh J,
Kameyama K, Okazaki I, Ala-Aho R, Kähäri VM and Watanabe T: Matrix
metallopro-teinase-13 promotes recovery from experimental liver
cirrhosis in rats. Pathobiology. 78:239–252. 2011. View Article : Google Scholar
|
|
28
|
Fallowfield JA, Mizuno M, Kendall TJ,
Constandinou CM, Benyon RC, Duffield JS and Iredale JP:
Scar-associated macrophages are a major source of hepatic matrix
metallopro-teinase-13 and facilitate the resolution of murine
hepatic fibrosis. J Immunol. 178:5288–5295. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Karkampouna S, Goumans MJ, Ten Dijke P,
Dooley S and Kruithof-de Julio M: Inhibition of TGFβ type I
receptor activity facilitates liver regeneration upon acute
CCl4 intoxication in mice. Arch Toxicol. 90:347–357.
2016. View Article : Google Scholar
|
|
30
|
Oe S, Lemmer ER, Conner EA, Factor VM,
Levéen P, Larsson J, Karlsson S and Thorgeirsson SS: Intact
signaling by transforming growth factor beta is not required for
termination of liver regeneration in mice. Hepatology.
40:1098–1105. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stauber AJ, Credille KM, Truex LL,
Ehlhardt WJ and Young JK: Nonclinical safety evaluation of a
transforming growth factor β receptor I kinase inhibitor in fischer
344 rats and beagle dogs. J Clin Pract. 4:1962014.
|
|
32
|
Kogure K, Omata W, Kanzaki M, Zhang YQ,
Yasuda H, Mine T and Kojima I: A single intraportal administration
of follistatin accelerates liver regeneration in partially
hepatectomized rats. Gastroenterology. 108:1136–1142. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ichikawa T, Zhang YQ, Kogure K, Hasegawa
Y, Takagi H, Mori M and Kojima I: Transforming growth factor beta
and activin tonically inhibit DNA synthesis in the rat liver.
Hepatology. 34:918–925. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fields SZ, Parshad S, Anne M, Raftopoulos
H, Alexander MJ, Sherman ML, Laadem A, Sung V and Terpos E: Activin
receptor antagonists for cancer-related anemia and bone disease.
Expert Opin Investig Drugs. 22:87–101. 2013. View Article : Google Scholar
|
|
35
|
Liu Z, Kobayashi K, van Dinther M, van
Heiningen SH, Valdimarsdottir G, van Laar T, Scharpfenecker M,
Löwik CW, Goumans MJ, Ten Dijke P and Pardali E: VEGF and
inhibitors of TGFbeta type-I receptor kinase synergistically
promote blood-vessel formation by inducing alpha5-integrin
expression. J Cell Sci. 122:3294–3302. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nakamura T, Tsutsumi V, Torimura T, Naitou
M, Iwamoto H, Masuda H, Hashimoto O, Koga H, Abe M, Ii M, et al:
Human peripheral blood CD34-positive cells enhance therapeutic
regeneration of chronically injured liver in nude rats. J Cell
Physiol. 227:1538–1552. 2012. View Article : Google Scholar
|
|
37
|
Tao Y, Wang M, Chen E and Tang H: Liver
regeneration: Analysis of the main relevant signaling molecules.
Mediators Inflamm. 2017:42563522017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tomita K, Haga H, Mizuno K, Katsumi T,
Sato C, Okumoto K, Nishise Y, Watanabe H, Saito T and Ueno Y:
Epiregulin promotes the emergence and proliferation of adult liver
progenitor cells. Am J Physiol Gastrointest Liver Physiol.
307:G50–G57. 2014. View Article : Google Scholar : PubMed/NCBI
|