1. Introduction
The severe acute respiratory syndrome coronavirus 2
(SARS-CoV-2), provisionally named as 2019-nCoV and colloquially
known as the coronavirus, is the pandemic-causing pathogen behind
coronavirus disease 2019 (COVID-19), which originated in Wuhan,
Hubei Province, China in the 4th quarter of 2019 (1). Despite the rapid delivery of the
SARS-CoV-2 genome by the group originally recognizing this novel
viral pneumonia and the subsequent production of a number of highly
specific and sensitive molecular diagnostics for SARS-CoV-2,
progress in fighting this disease is balanced by the continued lack
of ′quick win′ treatments that can suppress transmission or
ameliorate disease. Of the more than 100 clinical trials evaluating
a cadre of approved, or clinical stage antivirals and related
compounds, none have yet demonstrated unequivocal clinical utility
against COVID-19.
The eventual containment and mitigation of this
emerging disease will depend on our exponentially increasing
knowledge around SARS-CoV-2 virology, pathology, and cross-species
transmission. With viral genome organization resolved in
unprecedented speed, molecular pathogenesis and immunopathology
related to protein products, modes of transmission, host cell entry
mechanism and kinetics, vesicular trafficking, protein and small
molecule interaction, biological and replication machinery
mechanisms of action, are only beginning to be understood.
Likewise, the propensity for deleterious inflammatory responses is
episodically studied against SARS-CoV-2 antigen presentation,
antigen-specific antibody and cell mediated responses contributing
to protective adaptive immunity and the attenuation of disease
severity, as well as pulmonary and systemic tissue destruction.
Comprehensive interrogation of the viral-human interactome, and
associated chemoinformatic analyses (2,3)
nevertheless offer hope for the identification of nodal
intervention points across the continuum of COVID-19 disease
(4,5). As elucidation of these phenomena
will undoubtedly incur delays in developing appropriate
pharmacological interventions, clinical attention is understandably
focused on US Food and Drug Administration (FDA) approved drugs
which present putative treatments aligned to our present
under-standing of SARS-CoV (the agent of 2002-2003 Guangdong
Corinavirus epidemic) and Middle East respiratory syndrome virus
(MERS-CoV) infections, followed by compounds in clinical, or even
late pre-clinical investigation (6,7).
2. Virological, genomic, and host aspects
related to COVID-19 treatment
SARS-CoV-2 is a pleomorphic, enveloped,
positive-sense single-stranded RNA virus [(+)ssRNA] which encodes
15 or 16 replicate-related proteins, 4 or 5 structural proteins and
1-8 group-specific or accessory proteins from the
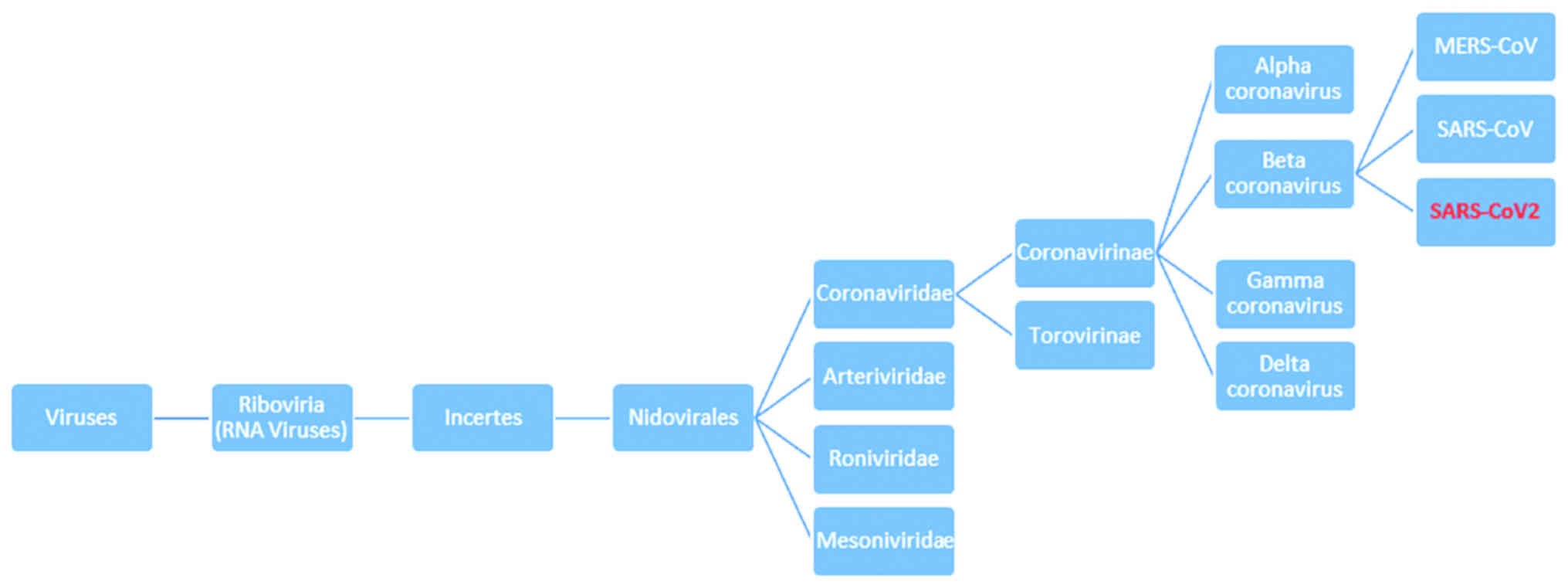
Orthocoronavirinae subfamily, Coronaviridae family,
order Nidovirales (8).
Coronaviridae are a large family that includes at least four
viruses responsible for common colds: four human coronaviruses
(HCoV 229E, NL63, OC43, and HKU1) account for up to a third of
adult upper respiratory tract infections (9). A phylogenetic tree of alpha-beta
coronavirus is presented in Fig.
1.
SARS-CoV-2 shows great phylogenetic similarity to
several strains of coronavirus isolated from bats in China, sharing
88% identity with bat-SL-CoVZC45 and bat-SL-CoVZXC21 which were
isolated in Zhoushan, eastern China (10), more than 96% genomic similarity
with BatCoV RaTG13 collected in Yunnan Province (11), and to a lesser extent to SARS-CoV
(approximately 79% homology). By contrast, the camelid-originating
MERS-CoV shares approximately 50% homology to SARS-CoV-2 (10).
Measuring a sizeable 29.8 kb, the SARS-CoV-2 genome
is flanked by short 5′ and 3′ highly structured untranslated
regions (UTRs) commonplace in RNA viruses and contains at least 14
open reading frames (ORFs) encoding 27 proteins organized in a
manner typical of other coronaviruses (CoVs) (12) (Fig.
2). The first ORFs are translated into two large polyproteins
(pp1a and pp1ab) cytosolically processed into the non-structural
proteins (nsp) 1-16. These contain the polyprotein-processing
protease nsp5, a host RNA restriction factor (nsp1) arresting
canonical translation, and collectively function as the viral
replicase-transcriptase complex (2) by rearranging rough endoplasmic
reticulum (RER) membranes into double-membrane vesicles (DMVs)
wherein replication, transcription and virion assembly take place
(13,14). The immunomodulatory protein nsp16
is 2′-O-ribose methyl transferase that renders the virus a
toll-like receptor (TLR) 7/8 antagonist (15) and is believed to enhance the
purported capacity of the virus to evade antiviral
interferon-generating cytosolic pathogen associated molecular
pattern recognition receptors, such as the double stranded RNA
(dsRNA) sensing protein kinase R (PKR) and TLR3, on account of the
DMV replication organelle structure (16). An additional protein of key
pharmacological interest the RNA-dependent RNA polymerase (RdRp)
nsp12, which is the primary target of the so-called directly acting
antiviral (DAA) drug family. The remaining ORFs encode the
so-called spike (S or SARS-2-S) protein responsible for virus
tropism, as well as envelope I, nucleocapsid (N) and membrane (M)
structural proteins. The mechanistic role of many of the remaining
accessory proteins that are redundant to viral replication remains
undetermined (7).
The human angiotensin-converting enzyme 2 (ACE2)
cell surface recaptor has been confirmed as the primary cell
attachment factor for SARS-CoV-2 (18). This single transmembrane domain
protein is expressed at appreciable protein levels on terminal
bronchiole as well as type I and II lung alveolar epithelium cells,
which aligns with the primary pathology of COVID-19: loss of
O2 saturation due to poor pulmonary gas exchange.
Likewise, noteworthy ACE2 levels have been reported among
enterocytes of the small intestine, arterial, and venous
endothelial cells, and arterial smooth muscle cells in most organs,
as well as the vagus nerve innervating the lung, heart and
digestive system (19,20), which mirror the diversity of
symptoms and pathologies associated with COVID-19: diarrhea,
myocarditis, fatigue, encephalopathy (headaches, confusion,
anosmia, stroke-like symptoms, and seizures). Following cellular
attachment, pivotal to cellular entry is the type II transmembrane
protease serine 2 (TMPRSS2) which process the SARS-CoV-2-S to
expose a cell-membrane fusion peptide (21). Once membrane fusion is achieved,
viral RNA is released into the infected host cell to initiate
pathogenic responses and viral replication. In a first step, the
open reading frame 1a/b (ORF1a/b) of the viral genome is translated
to produce the replicate proteins, after which the
replicase-transcriptase complex DMVs are assembled. In a second
step, the replication complex reverse-transcribes the positive RNA
genome into full-length negative-sense RNAs, which in turn template
the production of daughter full-length genomes, and a subset of
translation-focused mRNAs. These translation-dedicated transcripts
contain a common 5′ leader sequence cytosolically spliced to
downstream genes, which are added by discontinuous synthesis of
minus sense subgenomic RNAs templating the positive RNA genome
(22).
The genetic variance of SARS-CoV-2 is relatively
small compared to other RNA viruses (23), since CoVs possess mechanisms of
transcription error correction through the proofreading function of
nsp14 (24), a protein common to
MERS-CoV and SARS-CoV, which renders them collectively immune to
ribavirin (25), but is also
reported to drive recombinatory promiscuity (24). Consequently, ciral quasi-species
are relatively suppressed within patients, and mutation primarily
leads to single nucleotide polymorphisms (SNPs). Nearly 200 such
mutations have been reported as of mid-April 2020 (26). Based on the analysis of more than
100 virus isolates, two main genetic variants of SARS-CoV-2 were
initially identified, which are referred to by some members of the
community as L-type and S-type, differing only by 2 SNPs. The
L-type probably evolved from the S-type and is the more widespread
and ′aggressive′ variant (27).
Another study with 160 virus isolates from different countries
classified the virus into A, B and C type, the type A being the
oldest virus variant. Types A and C are most common in Europe and
America, while type B predominates in Asia (26). More recently, controversy over the
transmission propensity of the so-called L-type variant, which
harbours the noteworthy D614G mutation on SARS-CoV-2-S (28), has been ascribed to mutation
recurrence and multiple geographical introductions as elucidated
through the analysis of 7,666 published SARS-CoV-2 genomes
(29). However, the impact of
these mutations, particularly on protein core to the virus
replication cycle such as the cell tropism- and attachment
affinity-defining SARS-CoV-2-S remains experimentally
undetermined.
The current consensus among the scientific community
remains that the main sites of SARS-CoV-2 infection and replication
are both the upper and lower respiratory tracts, despite viral
loads being consistenly higher, and more reliable to detect in
lower respiratory tract speciments. However, this position, largely
driven by RT-PCR detection of SARS-CoV-2 and therefore, by
extrapolation replication, does not align with the near dearth of
ACE2 expression in the upper respiratory tract and oral mucosa
(20). The only exceptions to
these data are recent reports of olfactory epithelium ACE2
expression in mice (30) which is
in line with anosmic symptomatology in man, and early evidence of
interferon-induced increase in ACE2 expression in upper respiratory
tract sites, which would be secondary to infection and replication
in either distal, or physiologically non-epithelial local cell
types (31). Curiously,
endothelial ACE2 expression appears to be ubiquitous even in the
upper respiratory tract (20).
These findings postulate three hypotheses on the infection and
replication cascade of SARS-CoV-2. The simplest of these assumes
direct infection of upper respiratory tract epithelia from fomites
or large droplets (cough), followed by progressing infection to the
lower lung by week 2, which would explain deteriorating clinical
image in cases requiring hospitalization. However, this hypothesis
would rely on transcytotic epithelial mechanisms delivering
SARS-CoV-2 to ACE2-expressing basal upper respiratory/oral
epithelial cells or endothelia (20). Though transcytotic mechanisms are
not implausible (32,33), these cells would in turn propagate
the virus locally, causing interferon-induced ACE2 epithelial
overexpression, epithelial cell infection, and luminal virus
shedding, ultimately leading to the observations of low-level
detection in nasopharyngeal and oropharyngeal swabs by RT-PCR.
However, the immunomodulatory capacity of SARS-CoV-2 manifest
through DMVs and nsp16 (13,14,16), in addition to the consistently
higher viral load in the lower respiratory tract as opposed to the
relatively virus-free upper respiratory tract strongly disfavour
this hypothesis. The second hypothesis would require direct
infection of the olfactory epithelium again by fomites or droplets,
local replication and retrograde transport to the lower lung,
whereupon the virus would establish disease-causing replication.
However, the infrequent incidence of anosmia coupled to the
opposite direction of travel for tracheobronchial mucocilliary
clearance, and the need for sub-3 micron aerosol particle
dimensions to access the alveoli that would have to be generated in
the nose, present a formidable set of physiological barriers to the
virus for accessing the lower lung. The last hypothesis would
require direct infection of the lower respiratory tract through
inhalation of SARS-CoV-2 containing aerosols, directly infecting
the ACE2-overexpressing alveoli, sequentially driving upper
respiratory tract infection through mucocilliary clearance and
aerosol generation in exhaled breath. Poignantly, this putative
infection progression sequence aligns with ACE2 protein expression
data, SARS-CoV-2 levels reported by RT-PCR across the respiratory
tracts of patients, and offers a mechanistic explanation for the
rarity of anosmia and low nasopharyngeal carriage of the virus, as
diminishing quantities of aerosolized virus would need to diffuse
into the mucous found in these tissues upon exhalation. Critically,
data documenting viral particle production in upper respiratory
tract (or oral) epithelial or indeed endothelial cells remain
elusive from both human and animal studies.
3. Pathogenesis
Even though the replication site of the virus is
admittedly the lower respiratory system the virus can affect the
function of secondary organs that also express ACE2, such as the
kidney, GI tract, liver, and gall bladder. Typically, this results
in symptom severity that drives patients to seek medical attention,
usually by week 2 after symptom onset. However, to date no single
clincal report has documented measurable, or at least appreaciable
amounts of virus in the peripheral blood of patients admitted with
COVID-19. One possible explanation of the practically undetectable
levels of SARS-CoV-2 in blood might be the arrival time of patients
to hospitals (week 2), by which time seroconversion and
neutralizing antibodies are being generated and thus viraemia
suppressed. Alternatively, endothelial ACE2 expression reduces
blood viral load below the limit of detection of RT-PCR (~15,000
copies/ml whole blood) by sequestering free viral particles from
suspension. However, such extent of ACE2 expression systemically
across endothelia is not corroborated in public protein expression
data repositories (e.g., https://www.proteinatlas.org/ENSG00000130234-ACE2/tissue),
nor is it physiologically evidenced by the loss of endothelial
barrier function as is common in haemorrhagic fever viral
infections. Taken together, these findings cast doubts on
hypotheses regarding the mechanism of transfer of SARS-CoV-2 from
the respira-tory tract to the intestines (34), which is manifested through faecal
detection of SARS-CoV-2 RNA (35)
and protein in lower GI tract (36). By stark contrast, the main sites
of ACE2 expression, i.e., the lung, GI tract, kidneys, liver and
gall-bladder, and indeed the testis in men, wherein SARS-CoV-2
replication has also been documented (37), are all innervated by the vagus
nerve.
The median time from symptom onset to acute
respiratory distress syndrome, the principal reason for COVID-19
patients seeking medical attention, is approximately 8 days; around
this timepoint COVID-19 appears to morph from an acute viral
infection to what might appear to be a second attack, causing the
patient′s condition to worsen over the second week after symptom
onset. Critically, continued deterioration beyond day 14 is
associated with high mortality. Thus the disease could be divided
into three major stages after symptom onset: in the first stage
symptomatic patients report initially mild-to-moderate, but quality
of life impacting non-specific symptoms (days 0-7) associated with
a progressive drop in viral load; stage II, wherein severe dyspnoea
and associated pneumonia drive patients to seek medical attention;
and, finally stage III, a severe respiratory symptomatic stage
potentially with high viral load, but most commonly characterised
by runaway inflammatory responses associated with multiple organ
failure (38,39).
Thus, the ensuing innate immune response might be
two-phased; instead of the early innate immunity containing the
virus towards the developement of adaptive immunity, in some
patients there would appear to be a loss of key checkpoint
functions driving a secondary, damaging inflammatory phase. The
second inflammatory phase is driven by the cytokine release
syndrome (the so-called ′cytokine storm′) and affects patients with
severe conditions. Although genetic differences may underpin innate
response amplitude between individuals, viral amplification appears
to ensue (40) causing direct or,
more likely, indirect destruction of tissues affected by the
so-called ′happy hypoxia′ associated with COVID-19, the otherwise
asymptomatic critical reduction of blood oxygen saturation below
95%, sometimes reportedly as low as 50%, i.e., well below the level
normally associated with respiratory arrest.
The early antibody response against SARS-CoV-2
including IgA, IgM, and IgG response was examined by using an
ELISA-based assay on the recombinant SARS-CoV-2 protein against the
recombinant proteins for the other four common cold-causing human
coronaviruses. The median time to IgM and IgA antibody detection
was 5 days with interquartile range (IQR) 3-6, while IgG was
detected on 14 days (IQR 10-18) after symptom onset, with a
positive rate of 85.4, 92.7 and 77.9%, respectively (41). In a study of 23 patients with
COVID-19, viral load was reported to be declining across the first
week of illness, dropping below detection limits amongst most
patients by week 2. With analytical sensitivity lacking between
days 7-10, IgG and IgM antibodies were detectable more reliably at
approximately 10 days after symptom onset, with most patients
seroconverting within the first 3 weeks. Importantly, both IgG and
IgM were observed to neutralize SARS-CoV-2-N and the recaptor
binding domain of SARS-CoV-2-S (42).
Research data on late humoral response and
persistence of neutralizing antibodies in SARS-CoV-2 infection is
still limited. A major cause for concern are reports in China,
Japan and South Korea of patients testing still positive by RT-PCR
days, or even weeks after clinical discharge presumably on the
basis of two consecutive negative RT-PCR results. Some patientsa
are even reported to relapse, indicating that a sustained immune
response may be difficult to obtain, or that at least humoral
immunity is inadequate for infection elimination (40). Although the World Health
Organization (WHO) has recently pronounced these observations as a
consequence of RNA shedding from infected apoptotic epithelia in
the respiratory tract, corroborating data in the literature are
lacking. On the other hand, patients who have recovered from
SARS-CoV have been reported to harbour potent antibody responses to
SARS-S protein and its receptor binding domain (RBD) for at least
three years, indicating that RBD-targeting vaccines under
development might be adequately protective against COVID-19
(43,44).
Another aspect meriting attention is the progressive
lymphopenia documented during COVID-19, which is principally
manifest and most pronounced in severe cases, affecting
CD4+ and CD8+ T cell levels, as well as B
lymphocyte concentration, potentially adversely affecting antibody
production. Thus, T cell lymphopenia and associated cytokine
concentrations have been proposed as biomarkers of poor prognosis,
with interleukin (IL)-6, IL-10, and D-dimer continued elevation
marking transition to a severe disease state between days 7-14
after symptom onset, followed by further elevation of D-dimer,
neutrophilia, blood urea nitrogen, and creatinine marking in the
later, irreversible stages of disease. In contrast, IL-8 and tumor
necrosis factor alpha (TNF-α) elevation appears to be associated
and indeed peak early during the recovery phase (45,46).
4. Treatment strategies
In the absence of a clinically proven treatment for
COVID-19, frontline clinicians are restricted to offering
supportive symptomatic treatment such as oxygen therapy, steroids
on a per case basis, fluid management, and broad-spectrum
antibiotics against co- or secondary bacterial infections (45). Beyond these, as with any viral
disease, therapeutic intervention may extend to either direct
acting antivirals (DAAs) or immunomodulatory adjuvants and
substances (47). Considering the
drug discovery process, research efforts can be grouped in three
major classifications. The first relies on the repurposing of
approved, broad-spectrum antiviral drugs, through clinical trial
and error. The major advantage of this approach consists of the
clear knowledge about the drugs′ metabolic profile, dosages needed
and supported, potential side effects, and risks. The disadvantage
of these types of drugs is represented by their low selectivity
and, often, poor utility between virus families. The second
strategy is based on high-throughput screening, in silico,
in cellulo or, more recently, by means of artificial
intelligence drug discovery, to identify approved, or, at best,
late stage drugs with promising therapeutical effects either
through their intended mode of action (e.g., RNA dependent RNA
polymerase inhibition), or through their well-described off-target
effects (similar to kinase inhibitor promiscuity). The third
strategy involves identifying the molecular and pathological
characteristics of the disease in question, to develop entirely
novel drugs that may target them. The advantage here would be the
high selectivity and potentially high efficacy (48). Thankfully, the 18 years of head
start knowledge gained against SARS-CoV-2 on the back of the
SARS-CoV and MERS-CoV outbreaks of the early 21st century, have
resulted in an unprecedented speed by which the molecular
mechanisms of SARS-CoV-2 infection are being elucidated, not least
because of the rapid elucidation of the genomic organization of
SARS-CoV-2. These datasets have informed several rational, high
confidence potential therapeutic targets against SARS-CoV-2
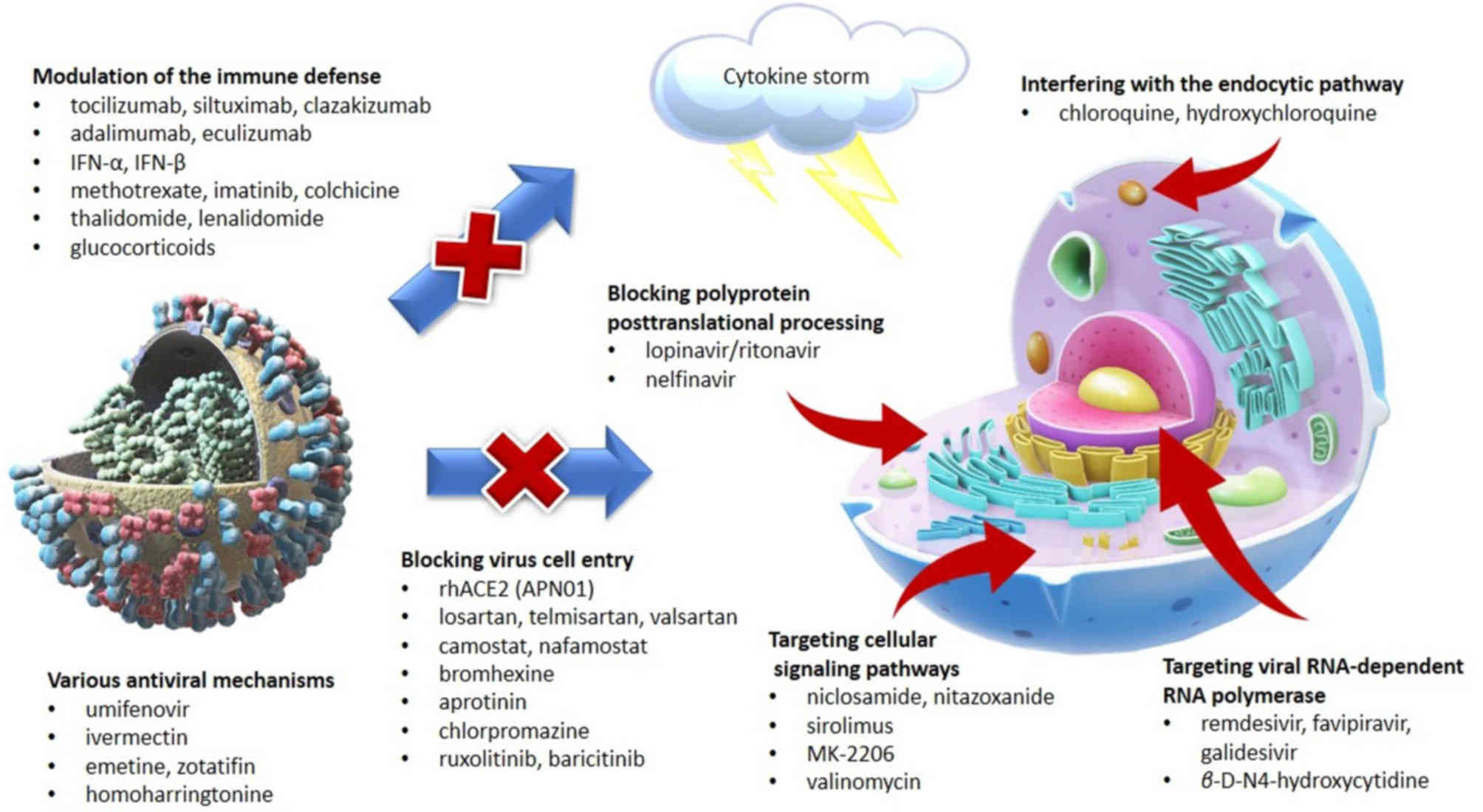
(10,49) (Fig.
3).
As the development of new drugs is a long and costly
process, the urgency of the pandemic targeted early efforts on
identifying candidate treatments among marketed drugs or approved,
but later abandoned compounds. Drug repurposing, also known as drug
repositioning, is one of the best solutions to identify therapeutic
solutions for COVID-19 in a time-critical fashion (50).
Blocking virus cell entry. Penetration of
host cells is an obligatory step in the life of any virus rendering
it a powerful target for drug development (51). The discovery of the intrinsic
mechanism of SARS-CoV-2-S to mediate cell entry focused efforts on
ACE2 and the whole renin-angiotensin-aldosterone system (RAAS)
(18,51). ACE2 is monocarboxypeptidase that
hydrolyzes the bond between the carboxyl-terminal phenylalanine and
proline in angiotensin II (Ang1-8) generating Ang1-7, a
physiological antagonist of Ang1-8 with vasodilator and
anti-inflammatory properties (52). Considering these effects, a
soluble recombinant human ACE2 (rhACE2) was developed under the
name APN01 (known also as GSK2586881) and evaluated in patients
with pulmonary arterial hypertension (53) and acute pulmonary injury (54). In healthy volunteers rhACE2 was
administered intravenously in doses from 100 to 1,200 µg/kg
and was generally well-tolerated, with no significant effect on
blood pressure and heart rate, and with only mild or moderate
adverse events reported (52).
SARS-CoV-2-S engages ACE2 as the entry receptor in a
similar way as the homologous SARS-CoV spike protein. The
expression and distribution of the ACE2 on alveolar epithelial
cells is consistent with the dry cough and dyspnea experienced by
the patients. Consequently, it is anticipated that rhACE2 may act
as a competitive interceptor of SARS-CoV-2 thereby reducing cell
penetration and offering protection against infection (55). A randomized placebo-controlled
study (NCT04335136) is scheduled to test rhACE2 in SARS-CoV-2
patients (56). Furthermore,
several peptide derivatives of ACE2 were designed using molecular
dynamics simulations as potential decoys that could be used as
future inhaled therapeutics (57). However, drug development history
(inhaled insulin, Exubera) denotes the brief residence time of
soluble proteins in the respiratory lumen, necessitating
modifications (58) that would
sustain rhACE2 decoy potency at the primary infection site of
interest for SARS-CoV-2, the respiratory epithelium.
Another strategy focused on ACE2 is based on the
hypothesis that ACE2 activity is reduced by SARS-CoV-2-S, exposing
the organism to higher levels of Ang1-8 which increases alveolar
permeability and causes lung injury (59). Two compounds,
resorcinolnaphthalein and 1-[(2-dimethylamino)ethylamino]-4-(hydro
xymethyl)-7-[(4-methylphenyl)sulfonyloxy]-9H-xanthene-9-one,
were identified in a large docking study as ACE2 activators and
were demonstrated to enhance ACE2 activity in a dose-depen-dent
manner, albeit with half-maximal effective concentration
(EC50) values of 19.5±0.4 and 20.1±0.8 µM,
respectively (60). Diminazene
aceturate is a veterinary-approved antitrypanosomal drug that
similarly activates ACE2 at comparable in vitro
EC50, rendering protective effects in cardiovascular and
type 1 diabetes experimental models (61,62). A recent review study presents the
toxic effect of diminazene aceturate on the nervous system of some
animals, but provides evidence that it was safe in thousands of
trypanosomiasis patients from Uganda (62). These compounds could be a
potential therapy in patients with SARS-CoV-2, provided topical
dosing and pharmacology afford adequate activation of respiratory
ACE2.
Likewise, use of Ang1-8 receptor 1 (AT1) antagonists
to reduce the aggressiveness and mortality of SARS-CoV-2 infection
based on two mechanisms, the blockade of the effects of excess
Ang1-8, and the upregulation of ACE2 (63). Several clinical studies are
underway to asses various AT1 antagonists: losartan 25 mg once
daily (NCT04335123, NCT04328012, and NCT04311177) or 50 mg daily
(NCT04312009), losartan 100 mg once daily alone or in combination
with aspirin 150 mg once daily or simvastatin 80 mg once daily in
an eight arms factorial trial (NCT04343001), telmisartan 80 mg
twice daily (NCT04355936), valsartan 80 mg up to 160 mg
(NCT04335786) (56).
Camostat is an orally active serine protease
inhibitor of trypsin and various inflammatory proteinases including
plasmin, kallikrein, thrombin and urokinase (64). It is approved clinically as
mesylate salt in Japan, but not Europe or the USA, for the
treatment of chronic pancreatitis and postoperative reflux
esophagitis due to its ability to inactivate trypsin and prevent
autodigestion (65). Camostat
mesylate has demonstrated inhibitory activity against TMPRSS2 and
other cell-surface proteases involved in SARS-CoV activation
(66). Indeed TMPRSS2 emerged as
a promising antiviral drug target after it was identified to have a
pivotal role for viral pathogenesis of monobasic H1N1, H3N2 and
H7N9 influenza A viruses (67).
Exposure of human bronchial epithelial Calu-3 cells
to 10 µM camostat mesylate was reported to cause a 10-fold
decrease in SARS-CoV viral entry and a 13-fold decrease in SARS-CoV
replication (66). Although from
a virology perspective such inhibitory effects are considered
mediocre, animal experiments in BALB/c mice at 30 mg/k administered
orally twice daily for 9 days starting 10 h before SARS-CoV
inoculation increased survival rate to 60 vs. 0% for untreated
animals (65). Similarly, Calu-3
cells pre-incubated with camostat mesylate at very high
concentrations (100 µM) before infection with SARS-CoV-2
significantly reduced, but failed to abrogate Calu-3 infection with
SARS-CoV-2 as measured by SARS-CoV-2-specific RT-PCR (21). Thus, a randomized,
placebo-controlled, phase IIa clinical trial (NCT04321096) is
evaluating camostat mesylate at 200 mg x 3/day for 5 days as
treatment for SARS-CoV-2. However, it is noteworthy that the study
design of this trial fails to meet the pre-treatment or, at least,
early treatment requirement common to most efficacious antivirals
and indeed the in vitro and in vivo performance
requirements exhibited previously. Elsewhere, the CLOCC-2020
clinical study (NCT04338906) will evaluate the efficacy and safety
of camostat mesylate in combination with the controversial agent
hydroxychloroquine in hospitalized patients with moderate COVID-19
infection (56).
Nafamostat, a chemical analog of camostat, has
potent inhibitory effects on several types of serine proteases
including trypsin, thrombin and plasmin (68) and was identified to effectively
reduce the MERS-CoV viral entry in Calu-3 cells at the
physiologically relevant concentration of 1 nM (69). Nafamostat at 100 µM
demonstrated promising inhibitory effects against SARS-CoV-2
infection of Vero E6 cells (70),
but it is anticipated that its very short half-life (71) significantly hinders clinical
utility in preventing SARS-CoV-2 infection.
Bromhexine, a widely used mucolytic cough medicine,
was identified as a TMPRSS2 inhibitor in a screening on various
chemical libraries, including over 1,200 FDA approved drugs. With
an IC50 of 0.75 µM, inhibition was reported as
selective, with low effects on the related hepsin and matriptase
proteases (72). However,
preliminary data on influenza-infected Calu-3 cells treated with
bromhexine showed no inhibitory effect (73). Despite a lack of in vitro
or in vivo data on SARS-CoV-2, it has been proposed as a
solution for COVID-19 patients based on its effect on TMPRSS2 and
favorable pharmacokinetic and safety profile (74). Several clinical trials are
registered to evaluate bromhexine as prophylaxis or therapy in
COVID-19 patients using various drug combinations (NCT04355026,
NCT04340349, NCT04273763) (56).
Aprotinin is a natural 58 amino acid peptide and a
serine protease inhibitor that blocks fibrinolysis and reduces
bleeding. It demonstrated significant effects against influenza
viruses (75). Preliminary
research reports have demonstrated that in Caco2 cells aprotinin is
more potent against SARS-CoV-2 (EC50 = 22.9 KIU/ml) than
against SARS-CoV (EC50 = 118 KIU/ml) cytopathogenic
effects (76) as measured in
kallikrein inactivator units (KIU). To this date, no clinical test
has been registered in Clinical Trials databases (56).
SARS-CoV nucleocapsid (protein N) binds to
cyclophilin A, which subsequently interacts with a member of the
immunoglobulin family of receptors, HAb18G/CD147, on the cell
membrane leading to viral cell invasion (77), in a mechanism similar with that of
HIV-1 virus (78). Based on this
model, a humanized anti-CD147 antibody was clinically tested in 17
patients with SARS-CoV-2 (NCT04275245) and published in a
preliminary fashion (79). Whilst
the results are interesting, interpretation should be approach with
caution as the source and characteristics of the antibody,
'meplazumab', are missing, and the number of tested patients is
very low.
Clathrin-mediated endocytosis is another mechanism
described for SARS-CoV cell invasion and possibly utilized by
SARS-CoV-2. Chlorpromazine inhibits the relocation of clathrin and
the adaptor protein 2 (AP2) from the cell surface, significantly
inhibiting SARS-CoV entry into HepG2 cells at the relatively high
concentration of 20 µM (80). Unfortunately, it showed no
significant effect on the SARS-CoV-2 cytopathic effects in Vero E6
cells when tested at 100 µM (81), yet a clinical trial is scheduled
to test the effect of 25 mg chlorpromazine administrated
intravenously every 6 h for 1 week (NCT04354805) (56).
An artificial intelligence method identified members
of the numb-associated kinases (NAK) family as potential
therapeutic targets against SARS-CoV-2 (82). The AP2-associated protein kinase 1
(AAK1) is an important member of NAK that binds to clathrin and
phosphorylates the medium subunit of AP2, playing an important role
in regulating clathrin-mediated endocytosis (83,84) and its inhibition was demonstrated
to reduce the infectivity of a wide range of viruses (82). The chemically related
pyrazole-pyrolopyrimidine derivatives baricitinib and ruxolitinib
are clinically approved Janus kinase (JAK) inhibitors with
important inhibitory effects on AAK1 (85), likewise predicted to reduce
SARS-CoV-2 infection (82). A
clinical trial (NCT04348071) is programmed to evaluate ruxolitinib
in patients receiving 10 mg twice daily for 14 days and a similar
trial (NCT04340232) will test baricitinib administered 2 mg once
daily for 14 days (56).
The role of cathepsins in coronavirus entry has been
established for SARS-CoV and MERS-CoV. In order to investigate
their role in SARS-CoV-2 cell entry a pseudovirus model was tested
in ACE2-expressing 293 cells. While cell treatment with the
cysteine protease cathepsin B inhibitor CA-074 (30 µM) had
no significant effect, cathepsin L inhibitor (SID 26681509, 2
µM) treatment reduced viral entry up to 76%. An inhibitory
effect close to 93% was reached after treatment with E64d (30
µM), an inhibitor of calpain and cathepsin B, H and L
(86). Cathepsin L is an
attractive target for drug development in several diseases, but
despite all the efforts, no cathepsin L inhibitor has advanced to
clinical trials (87) making
future cathepsin L-based SARS-CoV-2 therapies improbable.
Interfering with the endocytic pathway. Most
viruses use endocytic entry mechanisms and for the majority of
them, a reduction of pH serves as a cell penetration trigger
(88). Earlier studies have
described that endocytic pH modification suppresses viral
replication and may also have a virucidal effect. Such a minimal pH
value change, with no negative impact on the patient′s health
status, could be achieved by administering a simple small molecule
with hydrophobic weak base properties, like ammonium chloride,
which would lead to endocellular alkalosis by accumulating in
lysosomes following diffusion through plasma and lysosomal
membranes. Weak bases and ionophores, like ammonium chloride,
chloroquine, or monensin are frequently used in experimental
studies of viral cell penetration mechanisms (89). Ammonium chloride treatment
strongly inhibited the entry of pseudotypes bearing SARS-2-S in
293T cells lacking TMPRSS2, but was less effi-cient in Caco-2 cells
(TMPRSS2+) (21).
Chloroquine, chemically known as
7-chloro-4-((4-(diethy lamino)-1-methyl butyl)amino)quinoline, is a
9-aminoquinoline widely used for the treatment of malaria,
amebiasis and several auto-immune diseases (90) recently proposed to be efficient in
inhibiting SARS-CoV-2 in infected cells (70,91). Chloroquine has already
demonstrated in the past in vitro anti-viral activity
against several types of both RNA and DNA viruses, such as rabies
virus, poliovirus, several influ-enza virus types, hepatitis A, B
and C viruses, Dengue virus, Zika virus, and many others (92), as well as SARS-CoV (93) and MERS-CoV (94). The general anti-viral mechanism of
action relates to the ability of chloroquine, as a weak base, to
increase the endosomal pH required for viral/host cell fusion,
preventing endocytosis, and to interfere with viral particles bound
to the cell surface membrane (95). Additionally, in the case of
SARS-CoV and SARS-CoV-2, chloroquine is proposed to inhibit the
glycosylation of ACE2 receptor chains, thus limiting ligand
recognition of these receptors, rendering the viral spike protein
unable to mediate cell entry (92).
In light of the new pandemic, several clinical
trials have been conducted in different countries in order to test
the safety and efficacy of chloroquine phosphate in treating
SARS-CoV-2 patients (96), with
results confirming a significant improvement when compared to
control groups, as in the case of clinical trial ChiCTR2000029609
(97). When confronted with the
recent COVID-19 crisis in China, chloroquine has even been included
in the national treatment guidelines, with Chinese clinicians
recommending 500 mg chloroquine twice daily for 10 days for the
treatment of mild, moderate and severe cases of SARS-CoV-2-related
pneumonia with no contraindications to the drug (98).
Hydroxychloroquine is a less-toxic derivative of
chloroquine also used as an anti-malarial and in the management of
auto-immune disease, proposed through in vitro studies to
act also on SARS-CoV-2 infected cells (91). In Vero E6 models
hydroxychloroquine presented a better antiviral effect with EC50
values of 6.14 µM (after 24 h) and 0.72 µM (after 48
h), compared with chloroquine values of 23.90 µM (24 h) and
5.47 µM (48 h) (91).
Similar to chloroquine in many aspects with the
exception of a better pharmacotoxicological profile,
hydroxychloroquine is more frequently used instead of chloroquine
in COVID-19 clinical trials (99). Several studies confirmed
hydroxychloroquine sulfate administration to be significantly
associated with a reduction of the viral load in SARS-CoV-2
infected patients, as in the case of the Chinese randomized
clinical trial reported by Chen et al (ChiCTR2000029559)
(100) and Gautret et al,
who also reported after 6 days of treatment apparently significant
benefits compared to negative controls when administering daily 600
mg hydroxychloroquine in association with azithromycin, where
necessary (101). The drug is
currently perceived in certain quarters to be more promising than
chloroquine as a COVID-19 treatment due to the reduced probability
of causing retinopathy, psychiatric disorders, or severe
cardiovascular adverse effects such as arrhythmia (99); however, self-medication using both
drugs is unadvisable, as 9-aminoquinolines possess a narrow
therapeutic window (102) and
are associated with QT prologation. Currently, larger studies with
a more flexible design investigate the effect of
hydroxychloroquine, with the Solidarity study (EudraCT no.
2020-000982-18) and the Discovery study (NCT04315948) randomizing
treatment of SARS-CoV-2-infected patients using different antiviral
drugs, including hydroxychloroquine (103). However, preliminary reports are
highlighting a 2.61 adjusted hazard ratio of death among COVID-19
patients treated with hydroxychloroquine alone in the absence of
azithromycin, and no reduction in the risk of mechanical
ventilation (104).
Blocking polyprotein posttranslational
processing. Another promising strategy of treatment appears to
be interfering with viral replication, especially by acting at key
points following translation. After synthesis, the two newly-formed
viral polyproteins pp1a and pp1b require for posttranslational
processing two important proteinases encoded in these chains of
amino acids: papain-like proteinase, PLpro, and chymotrypsin-like
proteinase, 3CLpro, often referred to as the main protease or
Mpro (3). The activity
of these two cysteine proteases is essential for generating the 16
non-structural proteins, critical to the formation of the replicase
complex, and it is known that inhibiting the activity of either of
these two proteases affects the replication of SARS-CoV-2 (3,105).
Several inhibitors acting on SARS-CoV main protease,
which displays 96% similarity to the SARS-CoV-2 Mpro
(106), have recently been
synthesized and tested both in enzymatic assays and in cell lines,
showing promise for developing future drugs. Recently, Zhang et
al, after structure-based drug design and optimization of
different dipeptide α-ketoamides, derived several potential
Mpro inhibitors, of which
(S)-N-Benzyl-3-((S)-2-cinnamamido-3-cyclohexylpropanamido)-2-oxo-4-((S)-2-oxopyrrolidin-3-yl)
butanamide (referred to as 11r) displayed physiologically rele-vant
(nM) antiviral activity against MERS-CoV and SARS-CoV in hepatic
(Huh7) but not routine virology (Vero E6) cell lines, as well as
good inhibition of the SARS-CoV protease in enzymatic assays
(107). Based on previous
research, the same study group obtained a pyridone-containing
α-ketoamide deriva-tive of 11r, a compound (referred to as 13b)
with even lower sub-micromolar concentrations against SARS-CoV-2
Mpro in enzymatic assays, with half-maximal inhibitory
concentration (IC50) 0.67±0.18 µM, and
low-micromolar EC50 values (4-5 µM) in Calu-3
cells, further obtaining favorable results in mouse studies in
terms of pharmacokinetic parameters, reflecting lung tropism after
either subcutaneous or topical administration (108).
Drug repurposing appears to be another interesting
method of identification of Mpro inhibitors. Most
notable drugs showing inhibition of the main protease of SARS-COV-2
were found to be several anti-retroviral agents approved for HIV-1
treatment.
Lopinavir is a selective HIV-1 protease inhibitor
orally administered in combination with ritonavir, a CYP3A4
inhibitor used to increase its plasma concentration and clinical
efficacy (109). Lopinavir was
tested against SARS-CoV and showed a moderate activity in Vero-E6
cells (110). In a preliminary,
not yet peer-reviewed, study in Vero E6 cells, lopinavir showed a
mediocre EC50 = 5.73 µM against SARS-CoV-2
(111). A recent clinical trial
investigating the safety and efficacy of lopinavir (400 mg) and
ritonavir (100 mg) effect in 99 patients infected with SARS-CoV-2
has already ended, unfortunately concluding that the administration
of these drugs showed no significant benefit comparing to current
standard care (112). The
ineffectiveness of this combination could be explained by the
delayed treatment initiation, the median time from symptoms
identification to therapy onset being 13 days (51). Whether the ineffectiveness of the
treatment was due to the late therapy start or because of low
efficient concentrations at the site of action remains to be
explored.
The clinical use of lopinavir/ritonavir is
additionally linked to nausea and diarrhea (up to 28%) as well as
hepatotoxic effects (2-10%). Despite the limited data to support
use in the treatment of COVID-19, there is an important number of
clinical studies registered to test the lopinavir (400 mg) and
ritonavir (100 mg) coformulation alone or in combination with
hydroxychloroquine or interferon (51,56).
Other HIV-1 protease inhibitors, such as amprenavir,
tipranavir, saquinavir, ritonavir, nelfinavir, indinavir,
darunavir, and atazanavir that have been predicted to inhibit
SARS-CoV-2 Mpro (113) were also tested on Vero E6 cells
infected to SARS-CoV-2. Among these, nelfinavir was observed to be
the most efficient, with a mediocre EC50 of 1.13
µM suppressing viral replication in the post-entry step
(111). In a similar study on
SARS-CoV, nelfinavir inhibited the viral replication at non-toxic
doses with a selectivity index (SI) close to 300, while ritonavir,
lopinavir, saquinavir, and indinavir, did not have a significant
effect (114). Saquinavir and
tipranavir showed mediocre replication inhibition against
SARS-CoV-2 in cell lines (EC50 = 8.83 µM,
respectively 13.34), although with a lower selectivity index, while
amprenavir, darunavir and indinavir had low effects on viral
replication with EC50 values over 30 µM (111). Another preprint publication
reports the SARS-CoV-2 viral replication inhibition by atazanavir
with EC50 values of 2.0±0.12 µM (115).
No clinical study is registered in the Clinical
Trials database, nor in the Chinese Clinical Trial Registry to test
atazanavir, nelfinavir, tipranavir, or saquinavir as COVID-19
therapies. The clinical trials NCT04252274, NTC04303299 and
NCT04366089 will test darunavir in association with other
antivirals (56).
Targeting viral RNA-dependent RNA
polymerase. The RNA-dependent RNA polymerase (RdRp) is the
central subunit of the RNA synthesizing process for all
positive-strand RNA viruses and therefore an attractive target for
antiviral inhibitors. In SARS-CoV-2 nsp12 functions as RdRp
(116) and plays a central role
in the viral replication and transcription by catalyzing the
synthesis of viral RNA, assisted by nsp7 and nsp8 that serve as
co-factors (117). The
polymerase of RNA viruses is prone to replication mistakes, lacking
the proofreading capacity of its analog from DNA viruses. This high
mutation rate (10-3 to 10-5
mutations/nucleotide/round of replication) enables RNA viruses to
better adapt to environ-mental changes, but also introduces
damaging mutations that corrupt essential functions, a process
called lethal mutagenesis (118,119).
In CoVs, nsp14 has both 3′-5′ exoribonuclease
(ExoN) and guanine-N7-methyltransferase (N7-MTase) functions. ExoN
can hydrolyze RNA acting as a proofreading enzyme that is capable
of removing mismatched nucleotides (120). Targeting viral RNA-dependent RNA
polymerase has proven a successful strategy in treating various
viral infections (121), but is
difficult in the case of CoVs because of the ExoN activity
(116,119). Importantly, the catalytic site
of RdRp shares structural similarities with the hepatitis C virus
(HCV) ns5b RdRp (117),
introducing the prospect of repurposing ns5b HCV antivirals as
COVID-19 therapies. The results of a docking study indicate that
tenofovir and sofosbuvir, two approved HCV antivirals, can bind
tightly to SARS-CoV-2 RdRp (122). Still, these results should be
view with caution considering their effect could be theoretically
easily reversed by nsp14 activity. It is thus hypothesized that
finding inhibitors of ExoN activity could be a future direction for
COVID-19 therapeutics (119).
A number of compounds with proven inhibitory
effects on other virus RdRps were tested against SARS-CoV and
SARS-CoV-2, triaging some of them to clinical evaluation. Ribavirin
has the advantage of being clinically approved, but is associated
with important side effects and low efficacy. By contrast,
remdesivir, and to a lesser extent, favipiravir and galidesivir,
look like the most promising RdRp inhibitors for a future COVID-19
solution.
Remdesivir, formally known as GS-5734, is a
2-ethylbutyl-L-alaninate phosphoramidate prodrug that generates the
active compound, GS-441524, a 4-amino-5-cyano-pyrrolotriazine
analog of adenosine (123).
Remdesivir is metabolized more efficiently than its metabolite
GS-441524 into the active nucleoside triphosphate that function as
a chain-terminating nucleotide analog blocking RdRp (116). Remdesivir is not approved by the
FDA and not by the European Medicines Agency (EMA), but recently
received FDA emergency use authorization for the treatment of
COVID-19 on the basis of reduced hospitalisation time (124). However, this outcome is not
without controversy as the trial primary endpoint was altered days
before outcome announcement, raising important concerns with
respect to utility under the purported mechanism of action. Given
the structural similarity of remdesivir to naturally occurring
2′-O-methyl nucleoside analogues, antagonists of toll like receptor
7, and the initiation of treatment with the pre-symptomatic/early
symptomatic optimal treatment window, it is postulated remdesivir
may actually affect immunosuppression of cytokine storm syndrome,
as opposed to directly acting antiviral effects.
Remdesivir was developed to treat Ebola virus
infection, but demonstrated a broad antiviral spectrum against
SARS-CoV, MERS-CoV, and various other RNA viruses (116). In Vero E6 cells infected with
SARS-CoV-2, remdesivir potently blocked virus infection at high
nanomolar concentrations with an EC50 value of 0.77
µM demonstrating a low toxicity as the half-cytotoxic
concentration was reported to be above 100 µM (70). A separate study, however,
estimated EC50 at 26.90 µM (81). On the other hand, in Caco2 cells
remdesivir inhibited SARS-CoV-2 cytopathogenic effects with an
EC50 value of 0.23 µM (70). The protective effect was increased
10-fold by the addition of 8 µM of omeprazole. Omeprazole
alone has also demonstrated anti-SARS-CoV-2 protective effects, but
at physiologically irrelevant concentrations (EC50 = 34
µM), substantially above those used in clinical practice.
These results are presented in a yet to be peer-reviewed preprint
(76).
Remdesivir was clinically tested in 175 patients in
2018 during the Ebola outbreak in the Democratic Republic of Congo
by intravenous administration of 200 mg on day 1 followed by daily
maintenance doses of 100 mg from day 2 and continuing for 9 to 13
days. In pediatric patients, the doses were adjusted based on body
weight. The study revealed a favorable safety profile with higher
mean levels of creatinine and aspartate aminotransferase but failed
to achieve its primary endpoint (125).
A case report of the first COVID-19 patient
identified in the U.S. presents their treatment with intravenous
remdesivir on the seventh day of hospitalization and the
improvement of their condition afterward (126). A similar case report described
the remdesivir treatment of three hospitalized patients under
compassionate use protocol. The patient described as number 6 was
the same as the first case report. All three patients tolerated the
remdesivir treatment, with transient gastrointestinal symptoms and
a serum aminotransferase elevation (127). The administered doses were not
mentioned in the article.
A randomized, double-blind, placebo-controlled
clinical study (NCT04257656) on confirmed SARS-CoV-2 patients found
that remdesivir (n=158, vs. 79 placebo) intravenous administration
of a 200 mg loading dose on day 1 followed by 100 mg daily as
maintenance doses, up to a 10 days, produced no significant
difference in time to clinical improvement. The 28-day mortality
was similar between the remdesivir group (14%) compared to placebo
(13%). However, remdesivir reduced the time to clinical improvement
compared to placebo with a median of 18 vs. 23 days in the placebo
group. The study reported no significant impact of remdesivir use
on viral load measured on nasopharyngeal and oropharyngeal swabs,
implying either no direct acting antiviral effect, or an
unexpectedly long SARS-CoV-2 genome half-life (128). This result should be analyzed
considering the long average time (10-11 days) from symptom onset
to treatment start. We identified a number of other clinical trials
that are trying to evaluate the antiviral activity of remdesivir in
COVID-19 patients (NCT04252664, NCT04292899, NCT04292730,
NCT04302766, NCT04323761, NCT04280705, NCT04321616, NCT04315948,
NCT04314817, NCT04349410) using the same doses as NCT04257656
(56).
Favipiravir
(6-fluoro-3-hydroxy-2-pyrazinecarboxamide), also known as T-705,
was developed as a structural analog of
3-hydroxy-2-pyrazinecarboxamide (T-1105) compound identified in an
in vitro anti-influenza virus activity drug screening
(129). Favipiravir is a prodrug
that is activated by cellular enzyme phosphoribosylation to
favipiravir-ribofuranosyl-5′-triphosphate (favipiravir-RTP)
(130). It is reported to
selectively and potently inhibit the RNA-dependent RNA polymerase
of a broad range of RNA viruses: influenza viruses, arenaviruses,
bunyaviruses, noroviruses, flaviviruses and filoviruses (131). Favipiravir-RTP functions as a
guanosine and adenosine analog, its antiviral activity being
significantly inhibited by purine nucleotides and nucleosides, but
not by pyrimidine nucleotides in an MDCK cell model. Yet
favipiravir and its ribofuranosyl-5′-monophosphate showed no
significant inhibitory effect on influenza virus RNA polymerase
(100 µM). Favipiravir has a similar structure and also a
related antiviral spectrum as ribavirin increasing the lethal
mutations in the viral genome; this activity raises concerns about
teratogen risk potential (131).
The pharmacokinetic profile was characterized in Japanese healthy
volunteers to indicate a short half-life of 2 to 5.5 h and urinary
excretion in the form of the inactive 5-hydroxy-metabolite
(T-705M1) (132). Because of its
teratogenicity and embryotoxicity risks, favipiravir was approved
in Japan with strict regulations for use only in epidemics
(133).
Similar to remdesivir, favipiravir blocked
SARS-CoV-2 viral infection in Vero E6 cells, but at almost 80-fold
higher concentrations, with a calculated EC50 value of
61.88 µM (70). The
results of an open-label comparative controlled study of patients
with COVID-19 (ChiCTR2000029600) indicated that patients (n=35)
receiving favipiravir (1,600 mg x 2, day 1 and 600 mg x 2, days
2-14) had faster viral clearance and better chest imaging changes
than those (n=45) treated for 14 days with lopinavir (400 mg x 2)
and ritonavir (100 mg x 2). Both groups also received aerosol
inhalation of 5 million U IFN-α twice daily (134). Several clinical trials are
registered to test favipiravir in COVID-19 patients (NCT04358549,
NCT04346628, NCT04310228, NCT04349241, NCT04336904, NCT04319900,
NCT04359615, NCT04333589, NCT04303299, NCT04351295, NCT04356495,
NCT04345419) administered orally at a loading dose of 3,200 up to
3,600 mg (1,600-1,800 mg at 12 h) on day-1, followed by 1,200 to
1,800 mg maintenance dose (600-800 mg at 12 h) on days 2-10
(56).
Galidesivir, known also as BCX4430, is an adenosine
C-nucleoside analog similar to remdesivir, but has a nitrogen
replacing the oxygen in the ribose ring. It is converted in the
cell to the corresponding triphosphate nucleotide and after the
pyrophosphate′s cleavage to galidesivir monophosphate is
incorporated into nascent viral RNA, blocking viral RNA-dependent
RNA polymerase chain extension (135,136). It was reported to be active
against a wide range of viruses in cell and animal models (137). However, in a HeLa cell model of
SARS-CoV infection, galidesivir had an EC50 value of
57.7 µM (135).
Considering the high similarity of nsp12 in SARS-CoV-2 and SARS-CoV
(12), it was evaluated in Vero
E6 cells against SARS-CoV-2, but the inhibition of viral
replication was low at 100 µM (81). Nevertheless, a clinical
placebo-controlled, randomized study (NCT03891420) will test
galidesivir administered intravenously at 12 h intervals for 7 days
in COVID-19 patients (56).
A ribonucleoside derivative, β-D-N4-hydroxycytidine
(known also as NHC or EIDD-1931), was reported to demonstrate
potent inhibitory effects on MERS-CoV, SARS-CoV, and SARS-CoV-2 in
Calu-3 and human airway epithelial (HAE) cells. Encouragingly, NHC
inhibited SARS-CoV-2 infection in Vero E6 cells with
EC50 values of 0.3 µM and a value of 0.08
µM in Calu-3 cells. The isopropyl ester of the prodrug NHC,
EIDD-2801, was evaluated in C57BL/6 mice infected with a murine
adapted SARS-CoV to reduce significantly the viral titer and
associated infection symptoms when administered in doses of 500
mg/kg. The antiviral mechanism of action proposes the inhibition of
RdRp, with the effect of NHC being only slightly affected by ExoN
proofreading activity (138,139). Despite the potent antiviral
profile of EIDD-2801 and its advantage of oral administration
development for COVID, use is hindered by the lack of any clinical
data.
Ribavirin
(1-β-D-ribofuranosyl-1,2,4-triazole-3-carbox amide) is a
broad-spectrum antiviral agent clinically used primarily for
treating hepatitis C. The antiviral mechanism is complex, the most
important effects being the inhibition of human cells inosine
monophosphate dehydrogenase (IMPDH) by the monophosphate derivative
of the drug, and the incorporation of ribavirin 5′-triphosphate in
the nascent viral genome by RdRp (25). Both ribavirin and favipiravir act
as guanine analogs due to their carboxamide group that attaches to
cytosine by hydrogen bonds (137).
In cell models of SARS-CoV infection ribavirin had
a limited effect on viral replication (110). Similarly high concentrations are
needed to block SARS-CoV-2 viral infection in Vero E6 cells
(EC50 = 109.5 µM) (70) limiting clinical potential against
COVID-19. The resistance of SARS-CoV and SARS-CoV-2 to ribavirin is
produced by ExoN′s proofreading activity during virus replication
(119).
A systematic review of the therapies tested in the
SARS-CoV outbreak of 2002-2003 found that among the 20 studies on
ribavirin, results were inconclusive, with no discernible clinical
differences to other treatments. The review highlights various
toxic effects presented by several studies, the most frequent being
hemolytic anemia and liver dysfunction (140). The high doses needed to reduce
the viral replication highlight important toxic effects limiting
the therapeutic potential of ribavirin treatments in COVID-19
cases. Despite these reservations, an open-label, non-randomized,
clinical trial (NCT04356677) is programmed to test aerosolised
administration of ribavirin twice daily for up to 6 days at 50
mg/ml or 100 mg/ml concentrations (56). A combination of lopinavir (400
mg), ritonavir (100 mg), ribavirin (400 mg), and IFN-β 1b (0.25 mg)
is additionally under evaluation (NCT04276688) (56).
Targeting cellular signaling pathways.
Viruses highjack host cell-signaling pathways, redirecting cellular
processes in favor of their survival and replication, interfering
with normal cell metabolism, survival, by interrupting
transcription and replication (141). There are complex and not
completely understood mechanisms of viral interference with cell
signaling pathways. In the case of SARS-CoV-2, precise information
is scarce and amplified by the large number of proteins of unknown
function encoded in the virus genome, but research on new potential
therapies can successfully use knowledge gained on the related
SARS-CoV and MERS-CoV outbreak pathogens.
In cells infected with MERS-CoV perturbation of
several regulatory kinases was detected, such as the increased
phosphorylation of Akt1 and mTOR, modifications that have been
linked to a decrease in autophagy (142). Thus, the virus is believed to
block autophagy through a complex mechanism dependent on viral nsp6
that involves phosphorylation of S-phase kinase-associated protein
2 (SPK2) by a complex involving FK506-binding protein 51 (FKBP51),
PH domain leucine-rich repeat-containing protein phosphatases
(PHLPP), Akt, and Beclin1 (BECN1). The activated SPK2 promotes the
ubiquitination of BECN1 leading to its proteasomal degradation and
inhibition of autophagy (103).
A yet to be peer-reviewed report has proposed that SARS-CoV-2
infection may strongly reduce autophagic flux in experimental
models of human bronchial epithelial cells NCI-H1299 and VeroFM
cells (143). The analysis of
Vero E6 cells infected with SARS-CoV revealed higher levels of Akt
phosphorylated at serine 473 (S473) residue, but not at threonine
308 (T308) (144). The
anti-apoptotic effect of Akt depends on both S473 and T308
phosphorylation (145)
explaining low anti-apoptotic response to SARS-CoV infection
(144). These observations
highlight the potential of targeting SKP2 or the PI3K/AKT/mTOR
pathway.
MK-2206 is a selective and highly potent inhibitor
of all three Akt isoforms, currently under clinical evaluation
against various oncologic diseases (145). It reduced SARS-CoV-2
proliferation in VeroFM cells with a noteworthy EC50
value of 0.09 µM measured after 24 h in plaque-forming units
(143). This antiviral potential
of MK-2206 expands upon the previously reported activity on
influenza pH1N1 at EC50 values in the micromolar range
(146).
Several protein kinase inhibitors targeting the
ERK/MAPK and PI3K/AKT/mTOR signaling pathways were tested in Huh7
cells infected with MERS-CoV. Pretreatment with 10 µM
sirolimus (also known as rapamycin), an inhibitor of mTOR, reduced
viral infection by a mediocre 61%, while postinfection treatment
effected a comparable 57% reduction. Everolimus, a 2-hydroxyethyl
derivative of sirolimus, had similar inhibitory effects at 10
µM (56% reduction in pre-treated cells and 59% in
postinfection treatment) (147).
Surprisingly, the treatment of VeroFM cells with 0.3 µM
sirolimus increased the SARS-CoV-2 infection (143). Presently, a clinical trial
(NCT04341675) is scheduled to assess sirolimus administered as 6 mg
on day 1, followed by 2 mg daily for the next 13 days (56). Metformin could be another
repurposed drug for COVID-19 based on its autophagy induction by
activation of AMP-activated protein kinase (AMPK) and inhibition of
mTOR (148,149).
Niclosamide is an oral antihelminthic drug used to
treat a large array of parasitic infections being included on the
WHO list of essential medicines (150). In a screening of marketed drugs
it was identified to protect Vero E6 cells from the cytopathic
effects of SARS-CoV (151).
Niclosamide inhibited SARS-CoV replication in Vero E6 cells with a
corresponding EC50 value under 0.1 µM, as
measured by spike protein quantification (152). A yet not peer-reviewed preprint
presents the SARS-CoV-2 growth inhibition of niclosamide with an
EC50 value of 0.17 µM. The authors proposed a
mechanism of action that implicates the inhibition of SPK2
(143). SPK2 inhibitors reverse
the reduction of BECN1 levels produced by SARS-COV-2 in host cells
and enhance autophagy, inhibiting the viral replication (142,143). Another possible mechanism could
be niclosamide′s protonophore effect resulting in endosomal pH
neutralization preventing acid-dependent virus replication
(153). The maximum plasma
concentration after 500 mg x 3/day oral administration of
niclosamide ranged from 35.7 to 182 ng/ml, equivalent to 0.1-0.55
µM (154). The results
showed poor oral bioavailability that could hinder immediate
clinical utility against COVID-19. The clinical trial NCT04372082
will evaluate niclosamide 500 mg x 4/day (day 1) followed by 500 mg
x 2/day (days 2-10) in combination with diltiazem 60 mg x 3/day
(days 1-10) (56). The use of
diltiazem is supported by host-targeted antiviral effects
demonstrated in influenza mouse models (155).
Nitazoxanide, known as
2-(acetyloxy)-N-(5-nitro-2-thiazolyl)benzamide, is a structural
analog of niclosamide. Nitazoxanide is an antiprotozoal agent with
proven broad-spectrum antiviral properties. It is a prodrug
activated to its desacetyl derivative, tizoxanide, by hydrolysis.
Both nitazoxanide and tizoxanide inhibited MERS-CoV replication in
LLC-MK2 cells with mediocre EC50 values of 2.99
µM (0.92 µg/ml) and 3.13 µM (0.83
µg/ml), respectively (156). Nitazoxanide displayed similar
effects on SARS-CoV-2 cultured in Vero E6 cells with an
EC50 = 2.12 µM (70). The antiviral mechanism is possibly
the same as niclosamide, as observed in RAW264.7 cells treated with
tizoxanide. Tizoxanide increased BECN1 expression, inhibited Akt
and mTOR, and promoted the formation of autophagy vacuoles
(157). A placebo-controlled
study (NCT04343248) is scheduled to evaluate nitazoxanide (300 mg x
2/day) as post-exposure prophylaxis for COVID-19 elderly patients,
and a related study (NCT04359680) is focused on healthcare workers.
Other clinical studies will test nitazoxanide in combination with
hydroxychloroquine or with ivermectin (56).
Valinomycin is a toxic natural dodecadepsipeptide
that also inhibits SPK2 and blocked SARS-CoV replication in Vero E6
cells (EC50 = 1.63 µM) (152).
Antiviral mechanisms. Umifenovir (also known
as arbidol) is a broad-spectrum antiviral agent targeting both DNA
and RNA, and both enveloped and non-enveloped viruses, currently
used clinically for the prophylaxis and treatment of influenza in
Russia and China. The antiviral mechanism is not known and is
hypothesized to be directly virucidal coupled with host-targeting
effects (158,159), i.e., potentially
polypharmacological in nature. In a retrospective study on COVID-19
patients, 16 people received umifenovir 200 mg x 3/day, and 34
patients were treated with the lopinavir (400 mg)/ritonavir (100
mg) combination. The umifenovir group had a shorter duration of
positive SARS-CoV-2 RT-PCR tests, suggesting that umifenovir
monotherapy was more effective than lopinavir/ritonavir in reducing
viral load in COVID-19 patients (160). Separately, a retrospective
cohort study analyzed 16 patients who received umifenovir and
lopinavir/ritonavir versus 17 patients that received
lopinavir/ritonavir and found an apparent favorable clinical
response in the former group (161). In a retrospective study on 45
patients treated with 200 mg umifenovir three times a day for 5
days in 8 patients and 7 up to 10 mg in the other 37 patients, no
severe side effects were observed. The median time from admission
to the first negative test and hospitalization time was longer
compared to the control group, indicating that umifenovir alone may
not accelerate SARS-CoV-2 clearance (162).
The antiviral effect of umifenovir was only
recently investigated on Vero E6 infected with SARS-CoV-2.
Umifenovir treatment reduced the viral cytopathic effects with an
EC50 value of 4.11 µM, but with a relatively low
selectivity (SI=7.73). After a single oral dose of 800 mg, the
maximal plasma concentration in man was ascertained at ~4.1
µM. These data suggest that the doses of 200 mg three times
a day are too low for a physiological relevant antiviral effect
(163).
Ivermectin represents a mixture of two
semisynthetic avermectins and belongs to the chemical family of
16-membered macrocyclic lactones. It is used for over three decades
to treat various parasitic infections being included on the WHO
list of essential medicines (164). The antiviral effects of
ivermectin on various RNA viruses like influenza, Dengue virus, or
West Nile Virus prompted its evaluation against SARS-CoV-2.
Vero/hSLAM cells were infected with SARS-CoV-2 and treated with 5
µM ivermectin. After 24 h the viral RNA was reduced by 93%
in the supernatant and by 99.8% in the cells. After 48 h of
treatment, the viral RNA decrease was close to 5,000x (165). The EC50 value was
determined to be ~2 µM and the hypothesized mechanism of
action implicated blockade of viral protein transport in the
nucleus by dissociation of the host importin α/β1 heterodimer
(165,166) - mechanism not in line with the
exclusively cytosolic life cycle of SARS-CoV-2. Ivermectin is most
frequently used for onchocerciasis, strongyloidiasis, and
enterobiasis in doses of 150 to 200 µg/kg, and up to 400
µg/kg in patients with lymphatic filariasis (167). A clinical study (NCT04343092) is
programmed to test iver-mectin in COVID-19 patients as a single
dose of 200 µg/kg in combination with daily doses of
hydroxychloroquine (400 mg) and azithromycin (500 mg). Two other
clinical trials are registered to test ivermectin, one in
combination with chloroquine (NCT04351347), and another
(NCT04360356) in association with nitazoxanide (56).
Emetine is a natural tetrahydroisoquinoline
alkaloid from ipecac species with potent inhibitory effects on the
replication of a broad range of DNA and RNA viruses. Emetine
displayed potent anti-MERS-CoV activity in Vero E6 cells, with an
EC50 = 0.34 µM (after 72 h) and SI=9.06 (168). The EC50 value against
SARS-CoV-2 was evaluated as 0.5 µM after 48 h, a
concentration almost 3-fold higher than emetine′s therapeutic
plasma concentration (81),
hindering its clinical use in COVID-19 patients. Lycorine is a
structurally related alkaloid found in Amaryllidaceae
species, with potent anti-MERS-CoV effects (EC50 = 1.63
µM), but with therapeutic doses that exceed the toxic ones
(168). Homoharringtonine is an
alkaloid approved as a prodrug derivative, omacetaxine
mepesuccinate, for chronic myeloid leukemia treatment. It yields
potent and broad anti-RNA virus effects by reducing the
phosphorylation level of the eukaryotic initiation factor 4E
(eIF4E) (169).
Homoharringtonine reduced SARS-CoV-2 viral load in Vero E6 cells
with an EC50 value of 2.10 µM (81). Zotatifin (eFT226) is an inhibitor
of eukaryotic initiation factor 4A (eIF4A) under evaluation for
advanced solid tumor malignancies and produced strong antiviral
effects in Vero E6 cell models of SARS-CoV-2 viral infection
(170).
A number of antiviral compounds were evaluated
against SARS-CoV-2 with disappointing results. Oseltamivir, a
neuraminidase inhibitor frequently used for influenza patients
showed no significant antiviral effects at concentrations under 100
µM in Vero E6 cell models. The same result was obtained for
baloxavir acid, the active form of the baloxavir marboxil prodrug.
In influenza viruses it has been reported to block viral RNA
synthesis by inhibiting endonuclease activity (81). Other influenza neuraminidase
inhibitors such as laninamivir, peramivir, and zanamivir had no
result on the cytopathic effects of SARS-CoV-2 activity even at
high concentrations (163).
Modulation of the immune defense. The
response induced by viruses in the human host is a combination of
innate and acquired immunity that unfolds in a sequence, well known
in many aspects, but considerably nuanced between distinct insults.
Thus, foreign matter recognition initiates lympocyte recruitment,
to progresses into a resolution phase that normally seeks to return
to homeostasis (171). The
inflammatory response to SARS-CoV-2 generally begins when
pathogen-associated molecular patterns (PAMP), such as viral RNA,
and damage-associated molecular patterns (DAMPs), including ATP,
DNA, and ASC oligomers, are recognized by cell surface and
cytosolic PAMP recognition receptors (PRRs). The ensuing secretion
of pro-inflammatory cytokines and chemokines, such as IL-6, IFN-γ,
monocyte chemoattractant protein 1 (MCP1), and CXCL10, attracts
monocytes and T lymphocytes, but, unusually, not neutrophils, to
the infected site (172). The
impact of cytokines is generally increased in obesity, which is
associated with higher baseline inflammatory status, explaining the
higher risk of complications for obese patients infected with
SARS-CoV-2 (173).
The cytokine IL-6 is one of the most important
acute inflammatory mediators secreted early during an infection,
along with IL-1 and TNF-α (174). Immunosuppressants targeting
these molecules are needed to reverse the rampant immune activation
observed in late stage, severe forms of COVID-19 (175). A comprehensive list of
immunosuppressive therapies aimed at limiting immune-mediated
damage produced by SARS-CoV-2 infection are presented in Table I.
 | Table IImmunosuppressive therapies in
COVID-19. |
Table I
Immunosuppressive therapies in
COVID-19.
| Class | Drug | Mechanisms of
action | Doses | Clinical
studies |
|---|
| IL-6 | Tocilizumab | Humanized mAb that
binds selectively and competitively to soluble and
membrane-expressed IL-6 receptors, blocking IL-6 signal
transduction (176) | Several posologies,
e.g., in dose of 8 mg/kg (up to a maximum of 800 mg per dose)
within no less than 60 min, with an interval of 12 h (56) | NCT04317092,
NCT04345445, NCT04331795, |
| NCT04332094,
NCT04346355, NCT04359667, |
| NCT04372186,
NCT04320615, NCT04332913, |
| NCT04335071,
NCT04356937, NCT04363853, |
| NCT04363736,
NCT04361032, NCT04310228, |
| NCT04306705,
NCT04335305, NCT04315480, |
| NCT04370834,
NCT04339712, NCT04333914, |
| NCT04361552,
NCT04330638, NCT04331808, |
| NCT04322773,
NCT04347031, NCT02735707 (56) |
| Sarilumab | Human mAb, IgG1,
directed against IL-6R (177) | Doses of 400 mg, in
2 subcutaneous injections or by intravenous route (56) | NCT04357808,
NCT04315298, NCT04341870, |
| NCT04357860,
NCT04327388, NCT04359901, |
| NCT04324073,
NCT04322773, NCT04345289 (56) |
| Siltuximab | Chimeric anti-IL-6
mAb. | 11 mg/kg
administered by intravenous infusion | NCT04329650,
NCT04322188, NCT04330638 (56) |
| Clazakizumab | Humanized rabbit
mAb, anti-IL-6 | 25 mg intravenous
single dose, possibly repeated | NCT04351724,
NCT04363502, NCT04343989, |
| | | | NCT04348500
(56) |
| IL-1, | Anakinra | IL-1R natural
antagonist, reproduced industrially | Administered 400 mg
from day 1 to day 3 (two injections of 100 mg each 12 h) and 200 mg
the remaining 7 days | NCT04366232,
NCT04364009, NCT04357366, |
| IL-1β | NCT04324021,
NCT04339712, NCT04362111, |
| NCT04341584,
NCT04330638, NCT02735707 (56) |
| Canakinumab | mAb, IL-1β
antagonist | 300 or 600 mg (4 or
8 mg/kg) | NCT04365153,
NCT04348448, NCT04362813) (56) |
| TNF-α | XPro1595 | A protein biologic
that targets soluble TNF (sTNF) | 1 mg/kg for 2
weeks | NCT 04370236
(56) |
| Adalimumab | Human mAb against
TNF-α. | Not available | ChiCTR2000030089
(178) |
| IFN-α | IFN-α 2a, IFN-α 2b
nasal, aerosols | Type I IFN | Recombinant human
IFN-α 1b nasal drops, 2-3 drops for each nostril per time, 4
times/day IFN-α 1b in aerosols 10 million units twice daily | NCT04320238,
NCT04275388, NCT04293887, |
| NCT04251871,
NCT04344600, NCT04349410, |
| NCT04295551
(56) |
| IFN-β | IFN-β | Type I IFN, which
has the same receptors as INFα | IFN-β 1b 8 million
units per day, day 1-3 | NCT04350671,
NCT04343768, NCT04350684, |
| NCT04350281,
NCT04366245, NCT04324463, |
| NCT02735707,
NCT04276688, NCT04315948 (56) |
| IFN-λ | Pegylated
IFN-λ | Type III IFN | 180 µg
subcutaneous injection of pegylated IFN-λ | NCT04343976,
NCT04354259, NCT04344600 (56) |
IL-6 is produced in response to acute inflammation,
primarily in response to stimulation by TNF-α, lipopolysaccharides,
and IL-1 (179). The cellular
sensor for IL-5 is the surface-expressed IL-6 receptor (IL-6R;
classical signaling pathway) or through soluble IL-6R
(trans-signaling pathway) (180). The first pathway could be
anti-inflammatory as IL-6 plays a pivotal role in the polarization
of immune responses toward Th1, inhibiting the production of IFN-γ
by CD4 T cells, and by consequence CD8 T cell activation (174). The signaling through soluble
IL-6R has different consequences. The complex of IL-6 and soluble
IL-6R can bind to a membrane structure named gp130 on cells that do
not express IL-6R, initiating what appear to be pro-infammatory
cascades in cells otherwise non-responsive to IL-6 (181).
Several studies report elevations in IL-6 levels in
patients with COVID-19 (45,182,183). A retrospective study on 191
hospitalized COVID-19 patients from the Wuhan region found higher
IL-6 levels in the non-survivor group, compared to survivors
(184). A meta-analysis study
reported a 2.9-fold higher level of IL-6 in difficult COVID-19
cases in contrast to non-complicated ones (185). IL-6 has also an essential role
in the pathophysiology of rheumatoid arthritis being a validated
target for rheumatoid arthritis therapies (186), driving drug repurposing studies
for the IL-6 mediated-management of COVID-19.
Tocilizumab is a humanized monoclonal antibody
(mAb) directed against the IL-6 receptor used as an effective
therapeutic agent for rheumatoid arthritis patients who do not
respond to methotrexate (176).
A retrospective analysis of published studies involving the use of
tocilizumab in patients with COVID-19 highlighted the importance of
close patient monitoring for neutropenia, thrombocytopenia, and
elevated liver enzymes (187). A
pilot prospective open, single-arm study evaluated tocilizumab in
63 hospitalized patients with severe COVID-19 to report improved
respiratory and laboratory parameters (188). Sarilumab is a fully humanised
monoclonal antibody that specifically binds to both soluble and
membrane-bound IL-6 receptors (189). Siltuximab and clazakizumab are
antibodies targeting IL-6 that are under clinical evaluation for
severe lung disease associated with SARS-CoV-2 infection (190).
IL-1 receptor signaling is implicated early in
infection, preceeding IL-6 expression. However, it is also
responsible for acute tissue inflammatory damage. In mice, the
cytokine storm is associated with elevated levels of IL-1 (174), and a potentiation of TNF-α
(191). Using anakinra, the
synthetic form of natural IL-1 receptor antagonist, it has been
shown in mice that IL-1 signalling blockade could be a potential
treatment of influenza infections, arresting airway neutrophilia
early in the infection cycle, but also during the peak of viral
replication (192).
TNF-α is a key component of the cytokine response
and is implicated in COVID-19 severity, as it is found at elevated
levels among severe cases (45).
There is considerable clinical experience globally with several
antibodies targeting TNF-α in clinic against severe cases of
autoimmune inflammatory disease, such as rheumatoid arthritis,
Crohn's disease, ankylosing spondylitis, or psoriasis (193). The idea of repurposing
anti-TNF-α immunotherapies for the management of COVID-19 is
supported by preclinical studies on influenza mouse models
(193). Adalimumab, a fully
humanised monoclonal antibody, is under evaluation in COVID-19
patients (185).
Activation of the nicotinic receptor α7 on immune
cells also reduces the levels of IL-6 and TNF-α (171). Based on this effect, it has been
hypothesized that nicotine, a cholinergic agonist, could have
protective effects in COVID-19 patients (194). However, it is unlikely that
nicotine can be used clinically due to its addictive effects and
lack of specificity (171).
Rather, a review based on the available literature on the impact of
smoking on COVID-19 identified a number of studies highlighting a
history of smoking as a prognostic factor for severe outcomes,
while a meta-analysis study found no significant correlation
between smoking and the severity of COVID-19 disease (195).
The interferon family, whose expression is induced
directly and indirectly following viral PAMP recognition, is
critical in directing innate and adaptive immunity direction. Thus,
the interferon cascade induces multiple host antiviral factors or
immunomodulators. Activation of either of the two members of the
IFN-α receptor (IFNAR) family initiates cytosolic signal-ling
cascades culminating in the transcriptional activation of numerous
IFN-stimulated genes (ISGs). The protein products of these
transcripts exhibit antiviral principally through inhibition of
viral replication. Furthermore, type I IFNs enhance natural killer
cell activity and antiviral T cell responses, which are central to
antiviral defence (196).
A comprehensive evaluation of the therapies tested
in the SARS-CoV outbreak found two studies reporting IFN-α
therapies in combination with steroids and/or ribavirin, without a
significant difference in the clinical outcome when compared to
other therapies used (140). A
preprint report suggests a higher susceptibility of SARS-CoV-2 to
type I IFN treatment compared to SARS-CoV (197). These results were reaffirmed on
Vero cells treated with IFN-α or IFN-β, returning EC50
values for SARS-CoV-2 at 1.35 IU/ml for IFN-α, and 0.76 IU/ml for
IFN-β. In contrast, SARS-CoV EC50 values were reported
to range between 95 to 105 IU/ml depending on the viral strains
used (198).
Several immunosuppressants are also being examined
as potential solutions for severe COVID-19, with the highest
interest being in drugs such as methotrexate, JAK inhibitors
(ruxolitinib, baricitinib, tofacitinib, imatinib), colchicine,
thalidomide, and lenalidomide. Methotrexate-loaded nanoparticles
administered intravenously at 20 mg, once per week (total of 4
doses), is under evaluation in one trial (NCT04352465) (56). Imatinib, an Abl kinase inhibitor
and to a lesser extent, a JAK/STAT inhibitor, is used as an
experimental drug in elderly COVID-19 patients in out-of-hospital
settings at doses of 400 mg/day, 7 days (NCT04346147, NCT04357613,
NCT04356495) (56). Colchicine
presents long-established anti-inflammatory properties and
neutrophil inhibitory effects and is currently under evaluation in
COVID-19 patients (NCT04322682, NCT04322565, NCT04328480,
NCT04326790, NCT04350320) (184,199). Thalidomide plus low doses of
glucocorticoids, umifenovir, and IFN-α aerosols have been used in
NCT04273581 and NCT04273529. Lenalidomide, a structural derivative
of thalidomide, is currently under evaluation as 4 mg/day on days
1, 3 and 5, in less severe cases of COVID-19 associated with
low-molecular weight heparin (NCT04361643) (56). Eculizumab is a recombinant
humanized monoclonal antibody that targets complement component 5
and is under clinical study in 900-1200 mg doses administered
intravenously (NCT04355494) (56).
Dietary supplements. Ascorbic acid or
vitamin C is an established antioxidant and reactive oxygen species
scavenger widely considered to alleviate or prevent infections
(200). Vitamin C was proposed
as a supportive treatment for COVID-19 and several clinical trials
are registered to assess its effects. The clinical study
NCT04323514 uses intravenous administration of 10 g of vitamin C,
while study NCT04342728 uses 8 g of ascorbic acid divided into 2-3
doses/day alone or combined with 50 mg of zinc gluconate, and the
NCT04344184 study uses 100 mg/kg vitamin C intravenously
administered every 8 h for up to 72 h (56). An article details a Chinese expert
report on clinical improvement of 50 moderate to severe COVID-19
patients treated intravenously with 10-20 g/day (201). Vitamin D influences the immune
response by modulating several immune pathways and its deficiency
was correlated with increased risks of respiratory infections
(202). The clinical study
NCT04334005 will test administration of a single dose of 25,000 IU
of vitamin D as supplement in addition to standard therapies, while
in NCT04334005 vitamin D will be given orally as 50,000 IU once
weekly for 2 weeks in combination with 81 mg of acetylsalycilic
acid (56). Zinc has
anti-inflammatory effects moderating T cell functions that could
limit excess cytokine production in COVID-19 patients (203). Selenium supplementation was
shown to increase immunity and could be used in SARS-CoV-2
infections (204). The clinical
study NCT04323228 is scheduled to test an oral supplement enriched
in eicosapentaenoic acid, gamma-linolenic acid containing 18
µg selenium and 5.7 mg zinc (56).
Oligonucleotide and gene therapy-based
approaches. With the precise mechanism of SARS-CoV-2 migration
from respiratory epithelia onto distal ACE2-expressing organs
remaining unclear, direct intervention at the RNA genome level of
SARS-CoV-2 is also attracting research attention. However, efforts
in this space, compared to the advanced nature of vaccine and small
molecule intereventions, can only be characterized as speculative
and preliminary.
The most clinically advanced drug classes in this
space are short interfering RNA (siRNA) and ribonuclease H
(RNaseH)-active oligonucleotide antisense therapeutics, modalities
that have enjoyed serial regulatory approval successes and become
commercially viable since 2016. Thus, first pass metabolism affords
adequate passive liver uptake after systemic administration to
tackle hereditary hepatic diseases [e.g., mipomersen (205)]. Similarly, intrathecal
administration of antisense has proven an effective intervention
against spinal muscular atrophy [nusinersen (206)] and ushered true n-of-1 medicine
[milasen (207)]. Yet all
clinical candidates evaluated between 2000-2011 for lung diseases
of inflammatory or viral nature achieved ambiguous and inadequate
outcomes, despite the apparent facility of topical dosing to the
airways (208). This has been
largely attributed to misconceptions around systemic
bioavailability and topical cytosolic delivery potential, as rodent
studies of oligonucleotide bioavailability following respiratory
dosing (32) reflect
allometrically scaled human pharmacokinetics even onto
demonstrating effective hepatic and kidney targeting (32,209). Does this preclude topical
intervention to prevent ACE2 receptor docking of SARS-CoV-2?
Unfortunately, even the single clinical stage antisense candidate
AZD1419 targeting the cell surface TLR9 to improve time-to-loss of
eosinophilic, moderate-to-severe asthma control, also reportedly
failed to meet primary endpoints in the phase 2a INCONTRO trial
(NCT02898662) (56). Although
clarity over respiratory TLR9 coverage by the adopted dosing regime
remains undetermined, direct comparisons are ill-advised, thus
focusing future intervention efforts in four areas.
At first instance, aptamer-mediated inhibition of
ACE2 binding and TMPRSS2 activity, either solely or as a
combination therapy, presents a rational means of intervention to
prevent viral adhesion and cellular entry, similar to that proposed
by small molecule inhibitors. Although cost of goods, chemistry,
manufacturing and control requirements are significantly higher for
this chemotherapeutic class vs small molecule drugs, rapid hit
generation by modern aptamer development methods (210,211) and established orthogonal aptamer
optimization strategies can deliver clinical candidates compatible
with scalable manufacturing pertinent to pandemic outbreaks, in
timelines comparable or better than antibody biotherapeutics. Thus,
in the event of complete failure across the vast number of
repurposed medication trials described in this review, a second
wave of aptamer-based treatments could be ready for subsequent
clinical testing. Significantly, systemic access achieved through
the lung (32) may afford distal
tissue protection from SARS-CoV-2 infection.
The following approaches would involve the
generation of antisense, siRNA chemo- and biotherapeutics (DNA
Directed RNA interference mediators in the form of short hairpin
RNA (shRNA) precursors), or CRISPR-CAS13 systems (212) that would seek to directly cleave
the RNA genome of SARS-CoV-2. Cytosolic delivery challenges
notwithstanding, the paucity of reports indicating multi-log
suppression of viral replication by these chemotherapeutic classes
irrespective of target RNA virus, and the diverse evidence of rapid
viral evolutionary escape, combination therapy is likely to be
necessary (213) to afford
either protection, or treatment against SARS-CoV-2 by any such
agents. In this respect, RNA-cleaving CRISPR-CAS13 systems suffer
from macromolecular ribonucloprotein complex requirements that
compound drug delivery challenges vs the relatively smaller
molecular weight of oligonucleotides, that rely on host proteins to
exert their mechanism of action.
Poignantly, the only modality proven to achieve
cytosolic delivery in the lung are adeno-associated virus 6
biotherapeutics (214,215) for the minigene treatment of
cystic fibrosis (216). Although
this drug class has yet to be attempted against viral respiratory
diseases, the safety of AAV-mediated triple anti-viral shRNA
expression has been demonstrated in man even onto phase IIb against
HCV (NCT01899092). Consequently, a brief anti-SARS-CoV-2 shRNA
discovery and optimization campaign coupled to promoter
optimization experiments (217)
in physiologically relevant organoid models could yield a high
confidence-in-safety and confidence-in-rationale candidate drug
with the potential to suppress viral escape (218) if adequately tuned to express
shRNAs in vivo without interrupting microRNA homeostasis.
Significantly, the approach benefits from a ′fire and forget′,
single, inhaled administration that would mediate transgene
expression within 3 days of dosing and for up to 6 months (217), or potentially longer. It is,
therefore, proposed that such an intervention may compensate for
broad-spectrum, short-term protection from SARS-CoV-2 infection in
high risk populations in the event of failure to deliver protective
vaccines, or vaccination ineligibility due underlying
immunosuppression.
5. Discussion
Assembling this review was an exciting and
laborious daunting task, considering the astonishing volume of
research published and the urgency of finding effective, yet safe,
therapeutical solutions for COVID-19. The high pressure on the
research community leads both to very good studies and
publications, but also hasty and poorly designed studies, biased
conclusions, or unproven hypotheses, all mixed together. The
research directions were heavily influenced by the evolution of the
COVID-19 epidemic. At the early stages of the outbreak in China,
patients received oseltamivir based on the erroneous diagnosis of
influenza. Even if neuraminidase, the target of oseltamivir, was
invalidated for COVID-19 treatments, several ongoing clinical
trials still include oseltamivir as a comparison group (51).
Repurposing was also a rapid response solution
based on the experience with SARS-CoV, MERS-CoV, or with other
viruses. Many of the repurposed drugs undergoing clinical trials
were not tested in cell models against SARS-CoV-2, raising
questions about their real efficacy. Even so, the cell models can
be misleading because most compounds have not been tested on human
cells and because only EC50 values are reported without
clear information on the dose-response curve. Several conditions
such as different multiplicities of infection or time of exposure
can dramatically influence the EC50 values. There are
several other concerns that are worth mentioning: the potential of
a drug to reach the target tissue and how it would perform under
complex, disease-altered conditions (Fig. 4).
Future research should focus on how the virus
evades the immune response and try to better understand its
interaction with the T and B cell responses, both in the humans and
experimentally infected animals. The second goal should be to
reveal and assess details of viral replication strategies and
pathways, determining the relationship between sites of viral RNA
replication and virus assembly, as well as the extent to which
virus-host interactions are SARS-CoV-2 specific and organ-specific,
using genomics and proteomics, as well as new reagents. A third
future goal should be to translate new information about the
structure and function of SARS-CoV-2 proteins into specific
anti-virus therapies, particularly with respect to the SARS-CoV-2
proteins of unknown function. The next goal will be the
understanding of viral pathogenesis, to design and develop
effective live or attenuated safe vaccines. It is anticipated that,
like other RNA viruses, subgenomic SARS-CoV-2 replicon systems
adapted to lung, kidney, and even neuronal cell types will catalyse
the understanding of SARS-CoV-2 biology.
Funding
No funding was received.
Availability of data and materials
Data sharing is not applicable to this article, as
no datasets were generated or analyzed during the current
study.
Authors′ contributions
Conception: GMN, AT. Acquisition of data, design
and writing: GM, HP, SAM, DP, GN, GNDI, DAS, TKN, ND, AT. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
SAM is the CSO and Founder of PulmoBioMed Ltd., a
Northumbria University spin out company but had no personal
interest in product promotion, or any influence in terms of the
content of this article. DAS is the Editor-in-Chief for the
journal, but had no personal involvement in the reviewing process,
or any influence in terms of adjudicating on the final decision,
for this article. The other authors declare that they have no
competing interests.
Abbreviations:
|
AP2
|
adaptor protein 2
|
|
AMPK
|
AMP-activated protein kinase
|
|
AT1
|
Ang1-8 receptor 1
|
|
ACE2
|
angiotensin-converting enzyme 2
|
|
AAK1
|
AP2-associated protein kinase 1
|
|
BECN1
|
Beclin1
|
|
COVID-19
|
coronavirus disease 2019
|
|
EMA
|
European Medicines Agency
|
|
ExoN
|
exoribonuclease
|
|
EC50
|
half-maximal effective
concentration
|
|
IC50
|
half-maximal inhibitory
concentration
|
|
IFN
|
interferon
|
|
IL
|
interleukin
|
|
JAK
|
Janus kinases
|
|
MERS-CoV
|
Middle East respiratory syndrome
virus
|
|
mAb
|
monoclonal antibody
|
|
MCP1
|
monocyte chemoattractant protein
1
|
|
nsp
|
non-structural proteins
|
|
NAK
|
numb-associated kinases
|
|
ORFs
|
open reading frames
|
|
RBD
|
receptor binding domain
|
|
rhACE2
|
recombinant human ACE2
|
|
RdRp
|
RNA-dependent RNA polymerase
|
|
SNPs
|
single nucleotide polymorphisms
|
|
SPK2
|
S-phase kinase-associated protein
2
|
|
TLR
|
Toll-like receptor
|
|
TNF-α
|
tumour necrosis factor alpha
|
|
TMPRSS2
|
type II transmembrane protease serine
2
|
|
FDA
|
US Food and Drug Administration
|
Acknowledgements
Not applicable.
References
|
1
|
Lai CC, Shih TP, Ko WC, Tang HJ and Hsueh
PR: Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2)
and coronavirus disease-2019 (COVID-19): The epidemic and the
challenges. Int J Antimicrob Agents. 55:1059242020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fehr AR and Perlman S: Coronaviruses: An
overview of their replication and pathogenesis. Methods Mol Biol.
1282:1–23. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen Y, Liu Q and Guo D: Emerging
coronaviruses: Genome structure, replication, and pathogenesis. J
Med Virol. 92:418–423. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tsatsakis A, Petrakis D, Nikolouzakis TK,
Docea AO, Calina D, Vinceti M, Goumenou M, Kostoff RN, Mamoulakis
C, Aschner M and Hernández AF: COVID-19 an opportunity to
reevaluate the correlation between long-term effects of
anthropogenic pollutants on viral epidemic/pandemic events and
prevalence. Food Chem Toxicol. In Press.
|
|
5
|
Goumenou M, Sarigiannis D, Tsatsakis A,
Anesti O, Docea AO, Petrakis D, Tsoukalas D, Kostoff R, Rakitskii
V, Spandidos DA, et al: COVID-19 in Northern Italy: An integrative
overview of factors possibly influencing the sharp increase of the
outbreak (Review). Mol Med Rep. 22:20–32. 2020.PubMed/NCBI
|
|
6
|
Xu Z, Shi L, Wang Y, Zhang J, Huang L,
Zhang C, Liu S, Zhao P, Liu H, Zhu L, et al: Pathological findings
of COVID-19 associated with acute respiratory distress syndrome.
Lancet Respir Med. 8:420–422. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li X, Geng M, Peng Y, Meng L and Lu S:
Molecular immune pathogenesis and diagnosis of COVID-19. J Pharm
Anal. 10:102–108. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Docea AO, Tsatsakis A, Albulescu D,
Cristea O, Zlatian O, Vinceti M, Moschos SA, Tsoukalas D, Goumenou
M, Drakoulis N, et al: A new threat from an old enemy: Re-emergence
of coronavirus (Review). Int J Mol Med. 45:1631–1643.
2020.PubMed/NCBI
|
|
9
|
Paules CI, Marston HD and Fauci AS:
Coronavirus infections - more than just the common cold. JAMA.
323:7072020. View Article : Google Scholar
|
|
10
|
Lu R, Zhao X, Li J, Niu P, Yang B, Wu H,
Wang W, Song H, Huang B, Zhu N, et al: Genomic characterisation and
epidemiology of 2019 novel coronavirus: Implications for virus
origins and receptor binding. Lancet. 395:565–574. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou P, Yang XL, Wang XG, Hu B, Zhang L,
Zhang W, Si HR, Zhu Y, Li B, Huang CL, et al: A pneumonia outbreak
associated with a new coronavirus of probable bat origin. Nature.
579:270–273. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu A, Peng Y, Huang B, Ding X, Wang X, Niu
P, Meng J, Zhu Z, Zhang Z, Wang J, et al: Genome composition and
divergence of the Novel Coronavirus (2019-nCoV) originating in
China. Cell Host Microbe. 27:325–328. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Knoops K, Kikkert M, Worm SH,
Zevenhoven-Dobbe JC, van der Meer Y, Koster AJ, Mommaas AM and
Snijder EJ: SARS-coronavirus replication is supported by a
reticulovesicular network of modified endoplasmic reticulum. PLoS
Biol. 6:e2262008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Alsaadi EA and Jones IM: Membrane binding
proteins of coronaviruses. Future Virol. 14:275–286. 2019.
View Article : Google Scholar
|
|
15
|
Robbins M, Judge A, Liang L, McClintock K,
Yaworski E and MacLachlan I: 2′-O-methyl-modified RNAs act as TLR7
antagonists. Mol Ther. 15:1663–1669. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kikkert M: Innate immune evasion by human
respiratory RNA viruses. J Innate Immun. 12:4–20. 2020. View Article : Google Scholar :
|
|
17
|
Gordon DE, Jang GM, Bouhaddou M, Xu J,
Obernier K, White KM, O′Meara MJ, Rezelj VV, Guo JZ, Swaney DL, et
al: A SARS-CoV-2 protein interaction map reveals targets for drug
repurposing. Nature. https://doi.org/10.1038/s41586-020-2286-9.
|
|
18
|
Wrapp D, Wang N, Corbett KS, Goldsmith JA,
Hsieh CL, Abiona O, Graham BS and McLellan JS: Cryo-EM structure of
the 2019-nCoV spike in the prefusion conformation. Science.
367:1260–1263. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
To KF and Lo AWI: Exploring the
pathogenesis of severe acute respiratory syndrome (SARS): The
tissue distribution of the coronavirus (SARS-CoV) and its putative
receptor, angiotensin-converting enzyme 2 (ACE2). J Pathol.
203:740–743. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hamming I, Timens W, Bulthuis MLC, Lely
AT, Navis G and van Goor H: Tissue distribution of ACE2 protein,
the functional receptor for SARS coronavirus. A first step in
understanding SARS pathogenesis. J Pathol. 203:631–637. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hoffmann M, Kleine-Weber H, Schroeder S,
Krüger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu NH,
Nitsche A, et al: SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2
and is blocked by a clinically proven protease inhibitor. Cell.
181:271–280.e8. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sawicki SG, Sawicki DL and Siddell SG: A
contemporary view of coronavirus transcription. J Virol. 81:20–29.
2007. View Article : Google Scholar :
|
|
23
|
Rabi FA, Al Zoubi MS, Kasasbeh GA, Salameh
DM and Al-Nasser AD: Sars-cov-2 and coronavirus disease 2019: What
we know so far. Pathogens. 9:E2312020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gribble J, Pruijssers AJ, Agostini ML,
Anderson-Daniels J, Chappell JD, Lu X, Stevens LJ, Routh AL and
Denison MR: The coronavirus proofreading exoribonuclease mediates
extensive viral recombination. bioRxiv: https://doi.org/10.1101/2020.04.23057786.
|
|
25
|
Ferron F, Subissi L, Silveira De Morais
AT, Le NTT, Sevajol M, Gluais L, Decroly E, Vonrhein C, Bricogne G,
Canard B, et al: Structural and molecular basis of mismatch
correction and ribavirin excision from coronavirus RNA. Proc Natl
Acad Sci USA. 115:E162–E171. 2018. View Article : Google Scholar
|
|
26
|
Forster P, Forster L, Renfrew C and
Forster M: Phylogenetic network analysis of SARS-CoV-2 genomes.
Proc Natl Acad Sci USA. 117:9241–9243. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tang X, Wu C, Li X, Song Y, Yao X, Wu X,
Duan Y, Zhang H, Wang Y, Qian Z, et al: On the origin and
continuing evolution of SARS-CoV-2. Natl Sci Rev. nwaa0362020.
|
|
28
|
Korber B, Fischer W, Gnanakaran SG, Yoon
H, Theiler J, Abfalterer W, Foley B, Giorgi EE, Bhattacharya T,
Parker MD, et al: Spike mutation pipeline reveals the emergence of
a more transmissible form of SARS-CoV-2. bioRxiv: https://.org/10.1101/2020.04.29069054.
|
|
29
|
van Dorp L, Acman M, Richard D, Shaw LP,
Ford CE, Ormond L, Owen CJ, Pang J, Tan CCS, Boshier FAT, et al:
Emergence of genomic diversity and recurrent mutations in
SARS-CoV-2. Infect Genet Evol. May 5–2020.Epub ahead of print.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bilinska K, Jakubowska P, Von Bartheld CS
and Butowt R: Expression of the SARS-CoV-2 entry proteins, ACE2 and
TMPRSS2, in cells of the olfactory epithelium: identification of
cell types and trends with age. ACS Chem Neurosci. May 7–2020.Epub
ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ziegler CGK, Allon SJ, Nyquist SK, Mbano
IM, Miao VN, Tzouanas CN, Cao Y, Yousif AS, Bals J and Hauser BM:
SARS-CoV-2 receptor ACE2 is an interferon-stimulated gene in human
airway epithelial cells and is detected in specific cell subsets
across tissues. Cell. https://doi.org/10.1016/j.cell.2020.04.035.
|
|
32
|
Moschos SA, Frick M, Taylor B, Turnpenny
P, Graves H, Spink KG, Brady K, Lamb D, Collins D, Rockel TD, et
al: Uptake, efficacy, and systemic distribution of naked, inhaled
short interfering RNA (siRNA) and locked nucleic acid (LNA)
antisense. Mol Ther. 19:2163–2168. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Di Pasquale G and Chiorini JA: AAV
transcytosis through barrier epithelia and endothelium. Mol Ther.
13:506–516. 2006. View Article : Google Scholar
|
|
34
|
Lin L, Lu L, Cao W and Li T: Hypothesis
for potential pathogenesis of SARS-CoV-2 infection - a review of
immune changes in patients with viral pneumonia. Emerg Microbes
Infect. 9:727–732. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wu Y, Guo C, Tang L, Hong Z, Zhou J, Dong
X, Yin H, Xiao Q, Tang Y, Qu X, et al: Prolonged presence of
SARS-CoV-2 viral RNA in faecal samples. Lancet Gastroenterol
Hepatol. 5:434–435. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xiao F, Tang M, Zheng X, Liu Y, Li X and
Shan H: Evidence for gastrointestinal infection of SARS-CoV-2.
Gastroenterology. 158:1831–1833.e3. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Song C, Wang Y, Li W, Hu B, Chen G, Xia P,
Wang W, Li C, Hu Z, Yang X, et al: Detection of 2019 novel
coronavirus in semen and testicular biopsy specimen of COVID-19
patients. medRxiv: :https://doi.org/10.1101/2020.03.31.20042333.
|
|
38
|
Chen Y and Li L: SARS-CoV-2: Virus
dynamics and host response. Lancet Infect Dis. 20:515–516. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yi Y, Lagniton PNP, Ye S, Li E and Xu RH:
COVID-19: What has been learned and to be learned about the novel
coronavirus disease. Int J Biol Sci. 16:1753–1766. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shi Y, Wang Y, Shao C, Huang J, Gan J,
Huang X, Bucci E, Piacentini M, Ippolito G and Melino G: COVID-19
infection: The perspectives on immune responses. Cell Death Differ.
27:1451–1454. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Guo L, Ren L, Yang S, Xiao M, Chang D,
Yang F, Dela Cruz CS, Wang Y, Wu C, Xiao Y, et al: Profiling early
humoral response to diagnose novel Coronavirus Disease (COVID-19).
Clin Infect Dis ciaa. 310:2020.
|
|
42
|
To KKW, Tsang OTY, Leung WS, Tam AR, Wu
TC, Lung DC, Yip CC, Cai JP, Chan JM, Chik TS, et al: Temporal
profiles of viral load in posterior oropharyngeal saliva samples
and serum antibody responses during infection by SARS-CoV-2: An
observational cohort study. Lancet Infect Dis. 20:565–574. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cao Z, Liu L, Du L, Zhang C, Jiang S, Li T
and He Y: Potent and persistent antibody responses against the
receptor-binding domain of SARS-CoV spike protein in recovered
patients. Virol J. 7:2992010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Calina D, Docea AO, Petrakis D, Egorov A
M, Ishmukhametov AA, Gabibov AG, Shtilman MI, Kostoff R, Carvalho
F, Vinceti M, et al: Towards effective COVID-19 vaccines: Updates,
perspectives and challenges (Review). Int J Mol Med. 46:3–16.
2020.
|
|
45
|
Huang C, Wang Y, Li X, Ren L, Zhao J, Hu
Y, Zhang L, Fan G, Xu J, Gu X, et al: Clinical features of patients
infected with 2019 novel coronavirus in Wuhan, China. Lancet.
395:497–506. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang F, Nie J, Wang H, Zhao Q, Xiong Y,
Deng L, Song S, Ma Z, Mo P and Zhang Y: Characteristics of
peripheral lymphocyte subset alteration in COVID-19 pneumonia. J
Infect Dis. 221:1762–1769. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tu YF, Chien CS, Yarmishyn AA, Lin YY, Luo
YH, Lin YT, Lai WY, Yang DM, Chou SJ, Yang YP, et al: A Review of
SARS-CoV-2 and the Ongoing Clinical Trials. Int J Mol Sci.
21:212020. View Article : Google Scholar
|
|
48
|
Wu C, Liu Y, Yang Y, Zhang P, Zhong W,
Wang Y, Wang Q, Xu Y, Li M, Li X, et al: Analysis of therapeutic
targets for SARS-CoV-2 and discovery of potential drugs by
computational methods. Acta Pharm Sin B. Feb 27–2020.Epub ahead of
print. View Article : Google Scholar
|
|
49
|
Shojaei A and Salari P: COVID-19 and off
label use of drugs: An ethical viewpoint. Daru. May 8–2020.Epub
ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Senanayake SL: Drug repurposing strategies
for COVID-19. Future Drug Discov: fdd-2020-0010. 2020. View Article : Google Scholar
|
|
51
|
Sanders JM, Monogue ML, Jodlowski TZ and
Cutrell JB: Pharmacologic treatments for coronavirus disease 2019
(COVID-19): A Review. JAMA. 323:1824–1836. 2020.
|
|
52
|
Haschke M, Schuster M, Poglitsch M,
Loibner H, Salzberg M, Bruggisser M, Penninger J and Krähenbühl S:
Pharmacokinetics and pharmacodynamics of recombinant human
angio-tensin-converting enzyme 2 in healthy human subjects. Clin
Pharmacokinet. 52:783–792. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wilkins MR, Aman J, Harbaum L, Ulrich A,
Wharton J and Rhodes CJ: Recent advances in pulmonary arterial
hypertension. F1000Research. 7:F1000 Faculty Rev- 1128. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Khan A, Benthin C, Zeno B, Albertson TE,
Boyd J, Christie JD, Hall R, Poirier G, Ronco JJ, Tidswell M, et
al: A pilot clinical trial of recombinant human
angiotensin-converting enzyme 2 in acute respiratory distress
syndrome. Crit Care. 21:2342017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Batlle D, Wysocki J and Satchell K:
Soluble angiotensin-converting enzyme 2: A potential approach for
coronavirus infection therapy? Clin Sci (Lond). 134:543–545. 2020.
View Article : Google Scholar
|
|
56
|
US National Library of Medicine:
Recombinant Human Angiotensin-converting Enzyme 2 (rhACE2) as a
Treatment for Patients With COVID-19 (APN01-COVID-19). http://ClinicalTrials.gov
Identifier: NCT04335136. https://clinicaltrials.gov/ct2/show/NCT04335136?term=NCT04335136&draw=2amp;rank=1.
Accessed April 6, 2020.
|
|
57
|
Han Y and Král P: Computational design of
ACE2-based peptide inhibitors of SARS-CoV-2. ACS Nano.
14:5143–5147. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Heinemann L, Baughman R, Boss A and
Hompesch M: Pharmacokinetic and pharmacodynamic properties of a
novel inhaled insulin. J Diabetes Sci Technol. 11:148–156. 2017.
View Article : Google Scholar :
|
|
59
|
Deng J, Wang D-X, Deng W, Li C-Y and Tong
J: The effect of endogenous angiotensin II on alveolar fluid
clearance in rats with acute lung injury. Can Respir J. 19:311–318.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Hernández Prada JA, Ferreira AJ, Katovich
MJ, Shenoy V, Qi Y, Santos RA, Castellano RK, Lampkins AJ, Gubala
V, Ostrov DA, et al: Structure-based identification of
small-molecule angiotensin-converting enzyme 2 activators as novel
antihypertensive agents. Hypertension. 51:1312–1317. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Qi Y, Zhang J, Cole-Jeffrey CT, Shenoy V,
Espejo A, Hanna M, Song C, Pepine CJ, Katovich MJ and Raizada MK:
Diminazene aceturate enhances angiotensin-converting enzyme 2
activity and attenuates ischemia-induced cardiac pathophysiology.
Hypertension. 62:746–752. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Qaradakhi T, Gadanec LK, McSweeney KR,
Tacey A, Apostolopoulos V, Levinger I, Rimarova K, Egom EE, Rodrigo
L, Kruzliak P, et al: The potential actions of
angiotensin-converting enzyme II (ACE2) activator diminazene
aceturate (DIZE) in various diseases. Clin Exp Pharmacol Physiol.
47:751–758. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Gurwitz D: Angiotensin receptor blockers
as tentative SARS-CoV-2 therapeutics. Drug Dev Res. Mar 4–2020.Epub
ahead of print. View Article : Google Scholar
|
|
64
|
Ishikura H, Nishimura S, Matsunami M,
Tsujiuchi T, Ishiki T, Sekiguchi F, Naruse M, Nakatani T, Kamanaka
Y and Kawabata A: The proteinase inhibitor camostat mesilate
suppresses pancreatic pain in rodents. Life Sci. 80:1999–2004.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zhou Y, Vedantham P, Lu K, Agudelo J,
Carrion R Jr, Nunneley JW, Barnard D, Pöhlmann S, McKerrow JH,
Renslo AR, et al: Protease inhibitors targeting coronavirus and
filovirus entry. Antiviral Res. 116:76–84. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kawase M, Shirato K, van der Hoek L,
Taguchi F and Matsuyama S: Simultaneous treatment of human
bronchial epithelial cells with serine and cysteine protease
inhibitors prevents severe acute respiratory syndrome coronavirus
entry. J Virol. 86:6537–6545. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Tarnow C, Engels G, Arendt A, Schwalm F,
Sediri H, Preuss A, Nelson PS, Garten W, Klenk HD, Gabriel G, et
al: TMPRSS2 is a host factor that is essential for pneumotropism
and pathogenicity of H7N9 influenza A virus in mice. J Virol.
88:4744–4751. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Fuwa M, Kageyama M, Ohashi K, Sasaoka M,
Sato R, Tanaka M and Tashiro K: Nafamostat and sepimostat
identified as novel neuroprotective agents via NR2B
N-methyl-D-aspartate receptor antagonism using a rat retinal
excitotoxicity model. Sci Rep. 9:204092019. View Article : Google Scholar
|
|
69
|
Yamamoto M, Matsuyama S, Li X, Takeda M,
Kawaguchi Y, Inoue JI and Matsuda Z: Identification of Nafamostat
as a potent inhibitor of Middle East Respiratory Syndrome
Coronavirus S Protein-mediated membrane fusion using the
Split-Protein-Based Cell-Cell Fusion Assay. Antimicrob Agents
Chemother. 60:6532–6539. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wang M, Cao R, Zhang L, Yang X, Liu J, Xu
M, Shi Z, Hu Z, Zhong W and Xiao G: Remdesivir and chloroquine
effectively inhibit the recently emerged novel coronavirus
(2019-nCoV) in vitro. Cell Res. 30:269–271. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Lee YK, Lee HW, Choi KH and Kim BS:
Ability of nafamostat mesilate to prolong filter patency during
continuous renal replacement therapy in patients at high risk of
bleeding: A randomized controlled study. PLoS One. 9:e1087372014.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Lucas JM, Heinlein C, Kim T, Hernandez SA,
Malik MS, True LD, Morrissey C, Corey E, Montgomery B, Mostaghel E,
et al: The androgen-regulated protease TMPRSS2 activates a
proteolytic cascade involving components of the tumor
microenvironment and promotes prostate cancer metastasis. Cancer
Discov. 4:1310–1325. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Laporte M and Naesens L: Airway proteases:
An emerging drug target for influenza and other respiratory virus
infections. Curr Opin Virol. 24:16–24. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Maggio R and Corsini GU: Repurposing the
mucolytic cough suppressant and TMPRSS2 protease inhibitor
bromhexine for the prevention and management of SARS-CoV-2
infection. Pharmacol Res. 157:1048372020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Zhirnov OP, Matrosovich TY, Matrosovich MN
and Klenk HD: Aprotinin, a protease inhibitor, suppresses
proteolytic activation of pandemic H1N1v influenza virus. Antivir
Chem Chemother. 21:169–174. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Bojkova D, McGreig JE, McLaughlin KM,
Masterson SG, Widera M, Krähling V, Ciesek S, Wass MN, Michaelis M
and Cinatl J Jr: SARS-CoV-2 and SARS-CoV differ in their cell
tropism and drug sensitivity profiles. bioRxiv: :https://doi.org/10.1101/2020.04.03024257.
|
|
77
|
Chen Z, Mi L, Xu J, Yu J, Wang X, Jiang J,
Xing J, Shang P, Qian A, Li Y, et al: Function of HAb18G/CD147 in
invasion of host cells by severe acute respiratory syndrome
coronavirus. J Infect Dis. 191:755–760. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Watanabe A, Yoneda M, Ikeda F, Terao-Muto
Y, Sato H and Kai C: CD147/EMMPRIN acts as a functional entry
receptor for measles virus on epithelial cells. J Virol.
84:4183–4193. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Lythgoe MP and Middleton P: Ongoing
Clinical Trials for the Management of the COVID-19 Pandemic. Trends
Pharmacol Sci. Apr 9–2020.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Inoue Y, Tanaka N, Tanaka Y, Inoue S,
Morita K, Zhuang M, Hattori T and Sugamura K: Clathrin-dependent
entry of severe acute respiratory syndrome coronavirus into target
cells expressing ACE2 with the cytoplasmic tail deleted. J Virol.
81:8722–8729. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Choy KT, Wong AYL, Kaewpreedee P, Sia SF,
Chen D, Hui KPY, Chu DKW, Chan MCW, Cheung PP, Huang X, et al:
Remdesivir, lopinavir, emetine, and homoharringtonine inhibit
SARS-CoV-2 replication in vitro. Antiviral Res. 178:1047862020.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Stebbing J, Phelan A, Griffin I, Tucker C,
Oechsle O, Smith D and Richardson P: COVID-19: Combining antiviral
and anti-inflammatory treatments. Lancet Infect Dis. 20:400–402.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Conner SD and Schmid SL: Identification of
an adaptor-associated kinase, AAK1, as a regulator of
clathrin-mediated endocytosis. J Cell Biol. 156:921–929. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Sorrell FJ, Szklarz M, Abdul Azeez KR,
Elkins JM and Knapp S: Family-wide structural analysis of human
Numb-associated protein kinases. Structure. 24:401–411. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Eberl HC, Werner T, Reinhard FB, Lehmann
S, Thomson D, Chen P, Zhang C, Rau C, Muelbaier M, Drewes G, et al:
Chemical proteomics reveals target selectivity of clinical Jak
inhibitors in human primary cells. Sci Rep. 9:141592019. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Ou X, Liu Y, Lei X, Li P, Mi D, Ren L, Guo
L, Guo R, Chen T, Hu J, et al: Characterization of spike
glycoprotein of SARS-CoV-2 on virus entry and its immune
cross-reactivity with SARS-CoV. Nat Commun. 11:16202020. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Dana D and Pathak SK: A review of small
molecule inhibitors and functional probes of human Cathepsin L.
Molecules. 25:6982020. View Article : Google Scholar :
|
|
88
|
Cossart P and Helenius A: Endocytosis of
viruses and bacteria. Cold Spring Harb Perspect Biol.
6:a0169722014. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Nicola AV, McEvoy AM and Straus SE: Roles
for endocytosis and low pH in herpes simplex virus entry into HeLa
and Chinese hamster ovary cells. J Virol. 77:5324–5332. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Touret F and de Lamballerie X: Of
chloroquine and COVID-19. Antiviral Res. 177:1047622020. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Yao X, Ye F, Zhang M, Cui C, Huang B, Niu
P, Liu X, Zhao L, Dong E, Song C, et al: In vitro antiviral
activity and projection of optimized dosing design of
hydroxychloroquine for the treatment of severe acute respiratory
syndrome coronavirus 2 (SARS-CoV-2). Clin Infect Dis. Mar
9–2020.Epub ahead of print. View Article : Google Scholar :
|
|
92
|
Devaux CA, Rolain JM, Colson P and Raoult
D: New insights on the antiviral effects of chloroquine against
coronavirus: What to expect for COVID-19? Int J Antimicrob Agents.
Mar 12–2020.Epub ahead of print. View Article : Google Scholar
|
|
93
|
Biot C, Daher W, Chavain N, Fandeur T,
Khalife J, Dive D and De Clercq E: Design and synthesis of
hydroxyferroquine derivatives with antimalarial and antiviral
activities. J Med Chem. 49:2845–2849. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
de Wilde AH, Jochmans D, Posthuma CC,
Zevenhoven-Dobbe JC, van Nieuwkoop S, Bestebroer TM, van den Hoogen
BG, Neyts J and Snijder EJ: Screening of an FDA-approved compound
library identifies four small-molecule inhibitors of Middle East
respiratory syndrome coronavirus replication in cell culture.
Antimicrob Agents Chemother. 58:4875–4884. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Vincent MJ, Bergeron E, Benjannet S,
Erickson BR, Rollin PE, Ksiazek TG, Seidah NG and Nichol ST:
Chloroquine is a potent inhibitor of SARS coronavirus infection and
spread. Virol J. 2:692005. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Cortegiani A, Ingoglia G, Ippolito M,
Giarratano A and Einav S: A systematic review on the efficacy and
safety of chloroquine for the treatment of COVID-19. J Crit Care.
Mar 10–2020.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Gao J, Tian Z and Yang X: Breakthrough:
Chloroquine phosphate has shown apparent efficacy in treatment of
COVID-19 associated pneumonia in clinical studies. Biosci Trends.
14:72–73. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Guangdong Provincial Department of Science
and Technology and Guangdong Provincial Health Commission
Multi-center Collaborative Group of chloroquine phosphate for the
treatment of New Coronavirus Pneumonia: Expert consensus on the
treatment of new coronavirus pneumonia with chloroquine phosphate.
Zhonghua Jie He He Hu Xi Za Zhi. 43:185–188. 2020.
|
|
99
|
Liu J, Cao R, Xu M, Wang X, Zhang H, Hu H,
Li Y, Hu Z, Zhong W and Wang M: Hydroxychloroquine, a less toxic
derivative of chloroquine, is effective in inhibiting SARS-CoV-2
infection in vitro. Cell Discov. 6:162020. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Chen Z, Hu J and Zhang Z, Jiang S, Han S,
Yan D, Zhuang R, Hu B and Zhang Z: Efficacy of hydroxychloroquine
in patients with COVID-19: results of a randomized clinical trial.
medRxiv: :https://doi.org/10.1101/2020.03.22.20040758.
|
|
101
|
Gautret P, Lagier JC, Parola P, Hoang VT,
Meddeb L, Mailhe M, Doudier B, Courjon J, Giordanengo V, Vieira VE,
et al: Hydroxychloroquine and azithromycin as a treatment of
COVID-19: Results of an open-label non-randomized clinical trial.
Int J Antimicrob Agents. Mar 20–2020.Epub ahead of print.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Hickley NM, Al-Maskari A and McKibbin M:
Chloroquine and hydroxychloroquine toxicity. Arch Ophthalmol.
129:1506–1507. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Taccone FS, Gorham J and Vincent JL:
Hydroxychloroquine in the management of critically ill patients
with COVID-19: The need for an evidence base. Lancet Respir Med.
Apr 15–2020.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Magagnoli J, Narendran S, Pereira F,
Cummings T, Hardin JW, Sutton SS and Ambati J: Outcomes of
hydroxychloroquine usage in United States veterans hospitalized
with Covid-19. medRxiv: :https://doi.org/10.1101/2020.04.16.20065920.
|
|
105
|
Xue X, Yu H, Yang H, Xue F, Wu Z, Shen W,
Li J, Zhou Z, Ding Y, Zhao Q, et al: Structures of two coronavirus
main proteases: implications for substrate binding and antiviral
drug design. J Virol. 82:2515–2527. 2008. View Article : Google Scholar :
|
|
106
|
Hall DC Jr and Ji HF: A search for
medications to treat COVID-19 via in silico molecular docking
models of the SARS-CoV-2 spike glycoprotein and 3CL protease.
Travel Med Infect Dis. Apr 12–2020.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Zhang L, Lin D, Kusov Y, Nian Y, Ma Q,
Wang J, von Brunn A, Leyssen P, Lanko K, Neyts J, et al:
α-Ketoamides as broad-spectrum inhibitors of coronavirus and
enterovirus replication: structure-based design, synthesis, and
activity assessment. J Med Chem. 63:4562–4578. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Zhang L, Lin D, Sun X, Curth U, Drosten C,
Sauerhering L, Becker S, Rox K and Hilgenfeld R: Crystal structure
of SARS-CoV-2 main protease provides a basis for design of improved
α-ketoamide inhibitors. Science. 368:409–412. 2020.PubMed/NCBI
|
|
109
|
Croxtall JD and Perry CM:
Lopinavir/Ritonavir: A review of its use in the management of HIV-1
infection. Drugs. 70:1885–1915. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Chen F, Chan KH, Jiang Y, Kao RY, Lu HT,
Fan KW, Cheng VC, Tsui WH, Hung IF and Lee TS: In vitro
susceptibility of 10 clinical isolates of SARS coronavirus to
selected antiviral compounds. J Clin Virol. 31:69–75. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Yamamoto N, Matsuyama S, Hoshino T and
Yamamoto N: Nelfinavir inhibits replication of severe acute
respiratory syndrome coronavirus 2 in vitro. bioRxiv: :https://doi.org/10.1101/2020.04.06026476.
|
|
112
|
Cao B, Wang Y, Wen D, Liu W, Wang J, Fan
G, Ruan L, Song B, Cai Y, Wei M, et al: A Trial of
Lopinavir-Ritonavir in Adults Hospitalized with Severe Covid-19. N
Engl J Med. 382:1787–1799. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Boopathi S, Poma AB and Kolandaivel P:
Novel 2019 coronavirus structure, mechanism of action, antiviral
drug promises and rule out against its treatment. J Biomol Struct
Dyn. Apr 30–2020.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Yamamoto N, Yang R, Yoshinaka Y, Amari S,
Nakano T, Cinatl J, Rabenau H, Doerr HW, Hunsmann G, Otaka A, et
al: HIV protease inhibitor nelfinavir inhibits replication of
SARS-associated coronavirus. Biochem Biophys Res Commun.
318:719–725. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Fintelman-Rodrigues N, Sacramento CQ, Lima
CR, da Silva FS, Ferreira AC, Mattos M, de Freitas CS, Soares VC,
da Silva Gomes Dias S, Temerozo JR, et al: Atazanavir inhibits
SARS-CoV-2 replication and pro-inflammatory cytokine production.
bioRxiv: :https://doi.org/10.1101/2020.04.04020925.
|
|
116
|
Agostini ML, Andres EL, Sims AC, Graham
RL, Sheahan TP, Lu X, Smith EC, Case JB, Feng JY, Jordan R, et al:
Coronavirus susceptibility to the antiviral remdesivir (GS-5734) is
mediated by the viral polymerase and the proofreading
exoribonuclease. MBio. 9:e00221–e18. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Gao Y, Yan L, Huang Y, Liu F, Zhao Y, Cao
L, Wang T, Sun Q, Ming Z, Zhang L, et al: Structure of the
RNA-dependent RNA polymerase from COVID-19 virus. Science. Apr
10–2020.Epub ahead of print. View Article : Google Scholar
|
|
118
|
Smith EC: The not-so-infinite malleability
of RNA viruses: Viral and cellular determinants of RNA virus
mutation rates. PLoS Pathog. 13:e1006254. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Smith EC, Blanc H, Surdel MC, Vignuzzi M
and Denison MR: Coronaviruses lacking exoribonuclease activity are
susceptible to lethal mutagenesis: Evidence for proofreading and
potential therapeutics. PLoS Pathog. 9:e1003565. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Becares M, Pascual-Iglesias A, Nogales A,
Sola I, Enjuanes L and Zuñiga S: Mutagenesis of Coronavirus nsp14
reveals its potential role in modulation of the innate immune
response. J Virol. 90:5399–5414. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
De Clercq E and Li G: DC approved
antiviral drugs over the past 50 years. Clin Microbiol Rev.
29:695–747. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Elfiky AA: Ribavirin, Remdesivir,
Sofosbuvir, Galidesivir, and Tenofovir against SARS-CoV-2 RNA
dependent RNA polymerase (RdRp): A molecular docking study. Life
Sci. 253:1175922020. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Amirian ES and Levy JK: Current knowledge
about the antivirals remdesivir (GS-5734) and GS-441524 as
therapeutic options for coronaviruses. One Health. 9:1001282020.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
US Food and Drug Administration (FDA):
Recommendations for Investigational COVID-19 Convalescent Plasma.
https://www.fda.gov/vaccines-blood-biologics/investigational-new-drug-ind-or-device-exemption-ide-process-cber/recommendations-investigational-covid-19-convalescent-plasma.
Accessed May 1, 2020.
|
|
125
|
Mulangu S, Dodd LE, Davey RT Jr, Tshiani
Mbaya O, Proschan M, Mukadi D, Lusakibanza Manzo M, Nzolo D,
Tshomba Oloma A, Ibanda A, et al; PALM Writing Group. PALM
Consortium Study Team: A Randomized, Controlled Trial of Ebola
Virus Disease Therapeutics. N Engl J Med. 381:2293–2303. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Holshue ML, DeBolt C, Lindquist S, Lofy
KH, Wiesman J, Bruce H, Spitters C, Ericson K, Wilkerson S, Tural
A, et al: Washington State 2019-nCoV Case Investigation Team: First
Case of 2019 Novel Coronavirus in the United States. N Engl J Med.
382:929–936. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
COVID-19 Investigation Team: Clinical and
virologic characteristics of the first 12 patients with coronavirus
disease 2019 (COVID-19) in the United States. Nat Med. Apr
23–2020.Epub ahead of print.
|
|
128
|
Wang Y, Zhang D, Du G, Du R, Zhao J, Jin
Y, Fu S, Gao L, Cheng Z, Lu Q, et al: Remdesivir in adults with
severe COVID-19: A randomised, double-blind, placebo-controlled,
multicentre trial. Lancet. 395:P1569–P1578. 2020. View Article : Google Scholar
|
|
129
|
Furuta Y, Takahashi K, Kuno-Maekawa M,
Sangawa H, Uehara S, Kozaki K, Nomura N, Egawa H and Shiraki K:
Mechanism of action of T-705 against influenza virus. Antimicrob
Agents Chemother. 49:981–986. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Furuta Y, Gowen BB, Takahashi K, Shiraki
K, Smee DF and Barnard DL: Favipiravir (T-705), a novel viral RNA
polymerase inhibitor. Antiviral Res. 100:446–454. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Jordan PC, Stevens SK and Deval J:
Nucleosides for the treatment of respiratory RNA virus infections.
Antivir Chem Chemother. 26:20402066187644832018. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Madelain V, Nguyen THT, Olivo A, de
Lamballerie X, Guedj J, Taburet A-M and Mentré F: Ebola virus
infection: Review of the pharmacokinetic and pharmacodynamic
properties of drugs considered for testing in human efficacy
trials. Clin Pharmacokinet. 55:907–923. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Nagata T, Lefor AK, Hasegawa M and Ishii
M: Favipiravir: A new medication for the Ebola virus disease
pandemic. Disaster Med Public Health Prep. 9:79–81. 2015.
View Article : Google Scholar
|
|
134
|
Cai Q, Yang M, Liu D, Chen J, Shu D, Xia
J, Liao X, Gu Y, Cai Q, Yang Y, et al: Experimental Treatment with
Favipiravir for COVID-19: An Open-Label Control Study. Engineering
(Beijing). Mar 18–2020.Epub ahead of print.
|
|
135
|
Warren TK, Wells J, Panchal RG, Stuthman
KS, Garza NL, Van Tongeren SA, Dong L, Retterer CJ, Eaton BP,
Pegoraro G, et al: Protection against filovirus diseases by a novel
broad-spectrum nucleoside analogue BCX4430. Nature. 508:402–405.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Taylor R, Kotian P, Warren T, Panchal R,
Bavari S, Julander J, Dobo S, Rose A, El-Kattan Y, Taubenheim B, et
al: BCX4430 - A broad-spectrum antiviral adenosine nucleoside
analog under development for the treatment of Ebola virus disease.
J Infect Public Health. 9:220–226. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
De Clercq E: New nucleoside analogues for
the treatment of hemorrhagic fever virus infections. Chem Asian J.
14:3962–3968. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Sheahan TP, Sims AC, Zhou S, Graham RL,
Pruijssers AJ, Agostini ML, Leist SR, Schäfer A, Dinnon KH III,
Stevens LJ, et al: An orally bioavailable broad-spectrum antiviral
inhibits SARS-CoV-2 in human airway epithelial cell cultures and
multiple coronaviruses in mice. Sci Transl Med. 12:eabb58832020.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Agostini ML, Pruijssers AJ, Chappell JD,
Gribble J, Lu X, Andres EL, Bluemling GR, Lockwood MA, Sheahan TP,
Sims AC, et al: Small-molecule antiviral β-d-N4-Hydroxycytidine
inhibits a proofreading-intact Coronavirus with a high genetic
barrier to resistance. J Virol. 93:e01348–e19. 2019. View Article : Google Scholar :
|
|
140
|
Stockman LJ, Bellamy R and Garner P: SARS:
Systematic review of treatment effects. PLoS Med. 3:e343. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Moens U and Macdonald A: Effect of the
large and small T-antigens of human polyomaviruses on signaling
pathways. Int J Mol Sci. 20:202019. View Article : Google Scholar
|
|
142
|
Gassen NC, Niemeyer D, Muth D, Corman VM,
Martinelli S, Gassen A, Hafner K, Papies J, Mösbauer K, Zellner A,
et al: SKP2 attenuates autophagy through Beclin1-ubiquitination and
its inhibition reduces MERS-Coronavirus infection. Nat Commun.
10:57702019. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Gassen NC, Papies J, Bajaj T, Dethloff F,
Emanuel J, Weckmann K, Heinz DE, Heinemann N, Lennarz M, Richter A,
et al: Analysis of SARS-CoV-2-controlled autophagy reveals
spermidine, MK-2206, and niclosamide as putative antiviral
therapeutics. bioRxiv: :https://doi.org/10.1101/2020.04.15.997254.
|
|
144
|
Mizutani T, Fukushi S, Saijo M, Kurane I
and Morikawa S: Importance of Akt signaling pathway for apoptosis
in SARS-CoV-infected Vero E6 cells. Virology. 327:169–174. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Nitulescu GM, Margina D, Juzenas P, Peng
Q, Olaru OT, Saloustros E, Fenga C, Spandidos DA, Libra M and
Tsatsakis AM: Akt inhibitors in cancer treatment: The long journey
from drug discovery to clinical use (Review). Int J Oncol.
48:869–885. 2016. View Article : Google Scholar :
|
|
146
|
Denisova OV, Söderholm S, Virtanen S, Von
Schantz C, Bychkov D, Vashchinkina E, Desloovere J, Tynell J,
Ikonen N, Theisen LL, et al: Akt inhibitor MK2206 prevents
influenza pH1N1 virus infection in vitro. Antimicrob Agents
Chemother. 58:3689–3696. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Kindrachuk J, Ork B, Hart BJ, Mazur S,
Holbrook MR, Frieman MB, Traynor D, Johnson RF, Dyall J, Kuhn JH,
et al: Antiviral potential of ERK/MAPK and PI3K/AKT/mTOR signaling
modulation for Middle East respiratory syndrome coronavirus
infection as identified by temporal kinome analysis. Antimicrob
Agents Chemother. 59:1088–1099. 2015. View Article : Google Scholar :
|
|
148
|
De Santi M, Baldelli G, Diotallevi A,
Galluzzi L, Schiavano GF and Brandi G: Metformin prevents cell
tumorigenesis through autophagy-related cell death. Sci Rep.
9:662019. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Lehrer S: Inhaled biguanides and mTOR
inhibition for influenza and coronavirus (Review). World Acad Sci
J. 2:12020.PubMed/NCBI
|
|
150
|
Chen W, Mook RA Jr, Premont RT and Wang J:
Niclosamide: Beyond an antihelminthic drug. Cell Signal. 41:89–96.
2018. View Article : Google Scholar
|
|
151
|
Wu CJ, Jan JT, Chen CM, Hsieh HP, Hwang
DR, Liu HW, Liu CY, Huang HW, Chen SC, Hong CF, et al: Inhibition
of severe acute respiratory syndrome coronavirus replication by
niclosamide. Antimicrob Agents Chemother. 48:2693–2696. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Wen CC, Kuo YH, Jan JT, Liang PH, Wang SY,
Liu HG, Lee CK, Chang ST, Kuo CJ, Lee SS, et al: Specific plant
terpenoids and lignoids possess potent antiviral activities against
severe acute respi-ratory syndrome coronavirus. J Med Chem.
50:4087–4095. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Jurgeit A, McDowell R, Moese S, Meldrum E,
Schwendener R and Greber UF: Niclosamide is a proton carrier and
targets acidic endosomes with broad antiviral effects. PLoS Pathog.
8:e1002976. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Schweizer MT, Haugk K, McKiernan JS,
Gulati R, Cheng HH, Maes JL, Dumpit RF, Nelson PS, Montgomery B,
McCune JS, et al: A phase I study of niclosamide in combination
with enzalutamide in men with castration-resistant prostate cancer.
PLoS One. 13:e01983892018. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Pizzorno A, Terrier O, Nicolas de
Lamballerie C, Julien T, Padey B, Traversier A, Roche M, Hamelin
ME, Rhéaume C, Croze S, et al: Repurposing of drugs as novel
influenza inhibitors from clinical gene expression infection
signatures. Front Immunol. 10:602019. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Rossignol J-F: Nitazoxanide, a new drug
candidate for the treatment of Middle East respiratory syndrome
coronavirus. J Infect Public Health. 9:227–230. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Shou J, Wang M, Cheng X, Wang X, Zhang L,
Liu Y, Fei C, Wang C, Gu F, Xue F, et al: Tizoxanide induces
autophagy by inhibiting PI3K/Akt/mTOR pathway in RAW264.7
macrophage cells. Arch Pharm Res. 43:257–270. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Blaising J, Polyak SJ and Pécheur E-I:
Arbidol as a broad-spectrum antiviral: An update. Antiviral Res.
107:84–94. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Haviernik J, Štefánik M, Fojtíková M, Kali
S, Tordo N, Rudolf I, Hubálek Z, Eyer L and Ruzek D: Arbidol
(Umifenovir): A broad-spectrum antiviral drug that inhibits
medically important arthropod-borne flaviviruses. Viruses.
10:1842018. View Article : Google Scholar :
|
|
160
|
Zhu Z, Lu Z, Xu T, Chen C, Yang G, Zha T,
Lu J and Xue Y: Arbidol monotherapy is superior to
lopinavir/ritonavir in treating COVID-19. J Infect. Apr
10–2020.Epub ahead of print. View Article : Google Scholar
|
|
161
|
Deng L, Li C, Zeng Q, Liu X, Li X, Zhang
H, Hong Z and Xia J: Arbidol combined with LPV/r versus LPV/r alone
against Corona Virus Disease 2019: A retrospective cohort study. J
Infect. Mar 11–2020.Epub ahead of print. View Article : Google Scholar
|
|
162
|
Lian N, Xie H, Lin S, Huang J, Zhao J and
Lin Q: Umifenovir treatment is not associated with improved
outcomes in patients with coronavirus disease 2019: A retrospective
study. Clin Microbiol Infect. Apr 25–2020.Epub ahead of print.
View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Wang X, Cao R, Zhang H, Liu J, Xu M, Hu H,
Li Y, Zhao L, Li W, Sun X, et al: The anti-influenza virus drug,
arbidol is an efficient inhibitor of SARS-CoV-2 in vitro. Cell
Discov. 6:282020. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Crump A: Ivermectin: Enigmatic
multifaceted ′wonder′ drug continues to surprise and exceed
expectations. J Antibiot (Tokyo). 70:495–505. 2017. View Article : Google Scholar
|
|
165
|
Caly L, Druce JD, Catton MG, Jans DA and
Wagstaff KM: The FDA-approved drug ivermectin inhibits the
replication of SARS-CoV-2 in vitro. Antiviral Res. 178:1047872020.
View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Yang SNY, Atkinson SC, Wang C, Lee A,
Bogoyevitch MA, Borg NA and Jans DA: The broad spectrum antiviral
ivermectin targets the host nuclear transport importin α/β1
heterodimer. Antiviral Res. 177:1047602020. View Article : Google Scholar
|
|
167
|
Juarez M, Schcolnik-Cabrera A and
Dueñas-Gonzalez A: The multitargeted drug ivermectin: From an
antiparasitic agent to a repositioned cancer drug. Am J Cancer Res.
8:317–331. 2018.PubMed/NCBI
|
|
168
|
Shen L, Niu J, Wang C, Huang B, Wang W,
Zhu N, Deng Y, Wang H, Ye F, Cen S, et al: High-throughput
screening and identification of potent broad-spectrum inhibitors of
Coronaviruses. J Virol. 93:e00023–e19. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Dong HJ, Wang ZH, Meng W, Li CC, Hu YX,
Zhou L and Wang XJ: The natural compound Homoharringtonine presents
broad antiviral activity in Vitro and in Vivo. Viruses. 10:6012018.
View Article : Google Scholar :
|
|
170
|
Gordon DE, Jang GM, Bouhaddou M, Xu J,
Obernier K, White KM, O′Meara MJ, Rezelj VV, Guo JZ, Swaney DL, et
al: A SARS-CoV-2 protein interaction map reveals targets for drug
repurposing. Nature. Apr 30–2020.Epub ahead of print. View Article : Google Scholar
|
|
171
|
D′Elia RV, Harrison K, Oyston PC,
Lukaszewski RA and Clark GC: Targeting the 'cytokine storm' for
therapeutic benefit. Clin Vaccine Immunol. 20:319–327. 2013.
View Article : Google Scholar
|
|
172
|
Tay MZ, Poh CM, Rénia L, MacAry PA and Ng
LFP: The trinity of COVID-19: Immunity, inflammation and
intervention. Nat Rev Immunol. Apr 28–2020.Epub ahead of print.
View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Petrakis D, Margină D, Tsarouhas K, Tekos
F, Stan M, Nikitovic D, Kouretas D, Spandidos DA and Tsatsakis A:
Obesity a risk factor for increased COVID-19 prevalence, severity
and lethality (Review). Mol Med Rep. 22:9–19. 2020.PubMed/NCBI
|
|
174
|
Velazquez-Salinas L, Verdugo-Rodriguez A,
Rodriguez LL and Borca MV: The role of interleukin 6 during viral
infections. Front Microbiol. 10:10572019. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Mehta P, McAuley DF, Brown M, Sanchez E,
Tattersall RS and Manson JJ; HLH Across Speciality Collaboration,
UK: COVID-19: Consider cytokine storm syndromes and
immunosuppression. Lancet. 395:1033–1034. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Betts BC, St Angelo ET, Kennedy M and
Young JW: Anti-IL6-receptor-alpha (tocilizumab) does not inhibit
human monocyte-derived dendritic cell maturation or alloreactive
T-cell responses. Blood. 118:5340–5343. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Reichert JM: Which are the antibodies to
watch in 2013? MAbs. 5:1–4. 2013. View Article : Google Scholar :
|
|
178
|
Chinese Clinical Trial Registry (ChiCTR):
A clinical study for the efficacy and safety of Adalimumab
Injection in the treatment of patients with severe novel
coronavirus pneumonia (COVID-19). Registration number:
ChiCTR2000030089. http://www.chictr.org.cn/showprojen.aspx?proj=49889.
Accessed February 22, 2020.
|
|
179
|
Kamimura D, Ishihara K and Hirano T: IL-6
signal transduction and its physiological roles: The signal
orchestration model. Rev Physiol Biochem Pharmacol. 149:1–38.
2003.PubMed/NCBI
|
|
180
|
Kaur S, Bansal Y, Kumar R and Bansal G: A
panoramic review of IL-6: Structure, pathophysiological roles and
inhibitors. Bioorganic Med Chem. 28:1153272020. View Article : Google Scholar
|
|
181
|
Rose-John S: IL-6 trans-signaling via the
soluble IL-6 receptor: Importance for the pro-inflammatory
activities of IL-6. Int J Biol Sci. 8:1237–1247. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Chen N, Zhou M, Dong X, Qu J, Gong F, Han
Y, Qiu Y, Wang J, Liu Y, Wei Y, et al: Epidemiological and clinical
characteristics of 99 cases of 2019 novel coronavirus pneumonia in
Wuhan, China: A descriptive study. Lancet. 395:507–513. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Qin C, Zhou L, Hu Z, Zhang S, Yang S, Tao
Y, Xie C, Ma K, Shang K, Wang W and Tian DS: Dysregulation of
immune response in patients with COVID-19 in Wuhan, China. Clin
Infect Dis. Mar 12–2020.Epub ahead of print.
|
|
184
|
Zhou F, Yu T, Du R, Fan G, Liu Y, Liu Z,
Xiang J, Wang Y, Song B, Gu X, et al: Clinical course and risk
factors for mortality of adult inpatients with COVID-19 in Wuhan,
China: A retrospective cohort study. Lancet. 395:1054–1062. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Jamilloux Y, Henry T, Belot A, Viel S,
Fauter M, El Jammal T, Walzer T, François B and Sève P: Should we
stimulate or suppress immune responses in COVID-19? Cytokine and
anti-cytokine interventions. Autoimmun Rev. May 4–2020.Epub ahead
of print. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Srirangan S and Choy EH: The role of
interleukin 6 in the pathophysiology of rheumatoid arthritis. Ther
Adv Musculoskelet Dis. 2:247–256. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Alzghari SK and Acuña VS: Supportive
treatment with Tocilizumab for COVID-19: A systematic review. J
Clin Virol. 127:1043802020. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Sciascia S, Aprà F, Baffa A, Baldovino S,
Boaro D, Boero R, Bonora S, Calcagno A, Cecchi I, Cinnirella G, et
al: Pilot prospective open, single-arm multicentre study on
off-label use of tocilizumab in patients with severe COVID-19. Clin
Exp Rheumatol. May 1–2020.Epub ahead of print.
|
|
189
|
Bae SC and Lee YH: Comparison of the
efficacy and tolerability of tocilizumab, sarilumab, and sirukumab
in patients with active rheumatoid arthritis: A Bayesian network
meta-analysis of randomized controlled trials. Clin Rheumatol.
37:1471–1479. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Russell B, Moss C, George G, Santaolalla
A, Cope A, Papa S and Van Hemelrijck M: Associations between
immune-suppressive and stimulating drugs and novel COVID-19 - a
systematic review of current evidence. Ecancermedicalscience.
14:10222020. View Article : Google Scholar
|
|
191
|
Dinarello CA: The IL-1 family and
inflammatory diseases. Clin Exp Rheumatol. 20(Suppl 27): S1–S13.
2002.
|
|
192
|
Sichelstiel A, Yadava K, Trompette A,
Salami O, Iwakura Y, Nicod LP and Marsland BJ: Targeting IL-1β and
IL-17A driven inflammation during influenza-induced exacerbations
of chronic lung inflammation. PLoS One. 9:e984402014. View Article : Google Scholar
|
|
193
|
Feldmann M, Maini RN, Woody JN, Holgate
ST, Winter G, Rowland M, Richards D and Hussell T: Trials of
anti-tumour necrosis factor therapy for COVID-19 are urgently
needed. Lancet. 395:1407–1409. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Farsalinos K, Niaura R, Le Houezec J,
Barbouni A, Tsatsakis A, Kouretas D, Vantarakis A and Poulas K:
Editorial: Nicotine and SARS-CoV-2: COVID-19 may be a disease of
the nicotinic cholinergic system. Toxicol Rep. Apr 30–2020.Epub
ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Berlin I, Thomas D, Le Faou AL and Cornuz
J: COVID-19 and smoking. Nicotine Tob Res. Apr 3–2020.Epub ahead of
print. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Arimori Y, Nakamura R, Yamada H, Shibata
K, Maeda N, Kase T and Yoshikai Y: Type I interferon limits
influenza virus-induced acute lung injury by regulation of
excessive inflammation in mice. Antiviral Res. 99:230–237. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Lokugamage KG, Hage A, Schindewolf C,
Rajsbaum R and Menachery VD: SARS-CoV-2 sensitive to type I
interferon pretreatment. bioRxiv: :https://doi.org/10.1101/2020.03.07.982264.
|
|
198
|
Mantlo E, Bukreyeva N, Maruyama J,
Paessler S and Huang C: Antiviral activities of type I interferons
to SARS-CoV-2 infection. Antiviral Res. 179:1048112020. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Deftereos S, Giannopoulos G, Vrachatis DA,
Siasos G, Giotaki SG, Cleman M, Dangas G and Stefanadis C:
Colchicine as a potent anti-inflammatory treatment in COVID-19: can
we teach an old dog new tricks. Eur Hear J Cardiovasc Pharmacother.
Apr 27–2020.Epub ahead of print. View Article : Google Scholar
|
|
200
|
Hemilä H: Vitamin C and Infections.
Nutrients. 9:3392017. View Article : Google Scholar :
|
|
201
|
Cheng RZ: Can early and high intravenous
dose of vitamin C prevent and treat coronavirus disease 2019
(COVID-19)? Med drug Discov. 5:1000282020. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Marik PE, Kory P and Varon J: Does vitamin
D status impact mortality from SARS-CoV-2 infection? Med drug
Discov. Apr 29–2020.Epub ahead of print. View Article : Google Scholar :
|
|
203
|
Skalny AV, Rink L, Ajsuvakova OP, Aschner
M, Gritsenko VA, Alekseenko SI, Svistunov AA, Petrakis D, Spandidos
DA, Aaseth J, et al: Zinc and respiratory tract infections:
Perspectives for COVID 19 (Review). Int J Mol Med. 46:17–26.
2020.
|
|
204
|
Jayawardena R, Sooriyaarachchi P,
Chourdakis M, Jeewandara C and Ranasinghe P: Enhancing immunity in
viral infections, with special emphasis on COVID-19: A review.
Diabetes Metab Syndr. 14:367–382. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Langsted A and Nordestgaard BG: Antisense
oligonucleotides targeting lipoprotein(a). Curr Atheroscler Rep.
21:302019. View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Li Q: Nusinersen as a therapeutic agent
for spinal muscular atrophy. Yonsei Med J. 61:273–283. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
207
|
Kim J, Hu C, Moufawad El Achkar C, Black
LE, Douville J, Larson A, Pendergast MK, Goldkind SF, Lee EA,
Kuniholm A, et al: Patient-customized oligonucleotide therapy for a
rare genetic disease. N Engl J Med. 381:1644–1652. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
208
|
Moschos SA, Usher L and Lindsay MA:
Clinical potential of oligonucleotide-based therapeutics in the
respiratory system. Pharmacol Ther. 169:83–103. 2017. View Article : Google Scholar
|
|
209
|
Ng B, Cash-Mason T, Wang Y, Seitzer J,
Burchard J, Brown D, Dudkin V, Davide J, Jadhav V, Sepp-Lorenzino
L, et al: Intratracheal administration of siRNA triggers mRNA
silencing in the lung to modulate T cell immune response and lung
inflammation. Mol Ther Nucleic Acids. 16:194–205. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Le ATH, Krylova SM, Kanoatov M, Desai S
and Krylov SN: Ideal-filter capillary electrophoresis (IFCE)
facilitates the one-step selection of aptamers. Angew Chem Int Ed
Engl. 58:2739–2743. 2019. View Article : Google Scholar
|
|
211
|
Yufa R, Krylova SM, Bruce C, Bagg EA,
Schofield CJ and Krylov SN: Emulsion PCR significantly improves
nonequilibrium capillary electrophoresis of equilibrium
mixtures-based aptamer selection: Allowing for efficient and rapid
selection of aptamer to unmodified ABH2 protein. Anal Chem.
87:1411–1419. 2015. View Article : Google Scholar
|
|
212
|
Abbott TR, Dhamdhere G, Liu Y, Lin X,
Goudy L, Zeng L, Chemparathy A, Chmura S, Heaton NS, Debs R, et al:
Development of CRISPR as a prophylactic strategy to combat novel
coronavirus and influenza. bioRxiv: :https://doi.org/10.1101/2020.03.13991307.
|
|
213
|
Schopman NCT, ter Brake O and Berkhout B:
Anticipating and blocking HIV-1 escape by second generation
antiviral shRNAs. Retrovirology. 7:522010. View Article : Google Scholar : PubMed/NCBI
|
|
214
|
Duncan GA, Kim N, Colon-Cortes Y,
Rodriguez J, Mazur M, Birket SE, Rowe SM, West NE, Livraghi-Butrico
A, Boucher RC, et al: An Adeno-Associated Viral Vector Capable of
Penetrating the Mucus Barrier to Inhaled Gene Therapy. Mol Ther
Methods Clin Dev. 9:296–304. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Kurosaki F, Uchibori R, Mato N, Sehara Y,
Saga Y, Urabe M, Mizukami H, Sugiyama Y and Kume A: Optimization of
adeno-associated virus vector-mediated gene transfer to the
respiratory tract. Gene Ther. 24:290–297. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Steines B, Dickey DD, Bergen J, Excoffon
KJ, Weinstein JR, Li X, Yan Z, Abou Alaiwa MH, Shah VS, Bouzek DC,
et al: CFTR gene transfer with AAV improves early cystic fibrosis
pig phenotypes. JCI Insight. 1:e887282016. View Article : Google Scholar :
|
|
217
|
Suhy DA, Kao SC, Mao T, Whiteley L, Denise
H, Souberbielle B, Burdick AD, Hayes K, Wright JF, Lavender H, et
al: Safe, long-term hepatic expression of anti-HCV shRNA in a
nonhuman primate model. Mol Ther. 20:1737–1749. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Denise H, Moschos SA, Sidders B, Burden F,
Perkins H, Carter N, Stroud T, Kennedy M, Fancy SA, Lapthorn C, et
al: Deep sequencing insights in therapeutic shRNA processing and
siRNA target cleavage precision. Mol Ther Nucleic Acids.
3:e1452014. View Article : Google Scholar : PubMed/NCBI
|