Introduction
Gestational diabetes mellitus (GDM) is defined as
any degree of glucose intolerance identified for the first time
during pregnancy. This condition begins when the function of
pancreatic β cells is insufficient to overcome the insulin
resistance that occurs during pregnancy (1). GDM is associated with
pro-inflammatory processes, oxidative stress and endothelial cell
dysfunction in the microvasculature of the placenta (2).
Although several studies have been conducted to
identify biomarkers and to make a timely diagnosis of GDM (3,4),
the oral glucose tolerance test (OGTT) is still the gold standard
for the diagnosis of this condition. However, the cut-off values
established by the World Health Organization (WHO) have been
modified by several international consortiums, generating multiple
criteria for the diagnosis and management of GDM (5). Moreover, the OGTT is considered by
several authors to be invasive and to provide a late diagnosis
during pregnancy (4,6).
Maternal-fetal cellular communication depends on
factors secreted by the placenta. These factors can be markers of
altered placental functions that are detectable in the maternal
circulation from the early stages of pregnancy and, therefore,
could have a predictive value in various pregnancy-related diseases
(7). The human placenta expresses
>500 microRNAs (miRNAs or miRs), almost exclusively expressed in
this organ. For this reason, the study of placental miRNAs is
essential for the understanding of the regulatory mechanisms of
normal and complicated pregnancies (8).
Placental exosomes are released by exocytosis as
part of mechanisms for communication and molecular regulation in
normal pregnancy (9); however,
their composition and concen-tration is a reflection of the
cellular physiology, and important differences in these parameters
have been identified in association with pathological states
(10).
The chromosome 19 miRNA cluster (C19MC) located on
chromosome 19q13.41, is the largest group of miRNAs identified to
date. Some of the most important C19MC miRNAs are miR-516b-5p,
miR-517-5p and miR-518a-3p (11),
whose expression levels are modulated in response to stress,
alterations in circulating glucose levels and other stimuli
characteristic of pregnancy and are also contained in placental
exosomes. The cellular targets of C19MC miRNAs carried as exosomal
cargo have not been totally identified and may include
non-trophoblastic placental cells, maternal organs, maternal blood
system or fetal cells (12). The
downregulation of several C19MC miRNAs has been observed in
preeclampsia, gestational hypertension and fetal growth restriction
(13). Furthermore, some miRNAs
that are not coded by the C19CM, such as miR-16-5p and miR-222-3p,
have also been associated with pregnancy complications, such as
preeclampsia and GDM (14).
The expression of miRNAs contained in placental
exosomes purified from serum and plasma of patients with GDM has
been previously evaluated (15,16). However, to the best of our
knowledge, there are no studies available to date on the
characterization of miRNAs from urine exosomes of women with GDM.
Urine is a biological fluid very useful for clinical applications
due to its availability in large quantities, non-invasive
collection and simple sample treatment. In the present study, the
expression of miR-516-5p, miR-517-3p, miR-518-5p, miR-222-3p and
miR-16-5p was evaluated in placental exosomes purified from urine
in the 1st, 2nd and 3rd trimester of pregnancy with GDM and healthy
controls, since the participation of these microRNAs in the
pathophysiology of GDM is not yet fully understood.
Materials and methods
Study design and selection of
participants
The present longitudinal study included 61 pregnant
women who attended prenatal control between May, 2018 and April,
2019 at the Obstetrics Service of the Hospital Central 'Dr. Ignacio
Morones Prieto', San Luis Potosí, Mexico. Patients with gestational
hypertension, urinary infections, pre-existing diabetes mellitus,
and chronic kidney disease were excluded.
The criteria used for the diagnosis of GDM were
established by the Obstetrics Service of the Hospital Central 'Dr.
Ignacio Morones Prieto' (Table
SI) and following the WHO criteria (17). All participants were tested for
oral glucose tolerance between weeks 24 and 28 of gestation
(17).
Clinical and demographic data were obtained from the
medical records of each participant in the first prenatal visit.
The GDM group (n=27) was formed by patients who were diagnosed with
GDM during the second trimester of pregnancy. The control group
(n=34) was constituted by euglycemic women. The present study was
approved by the Ethics Committee of the Hospital Central 'Dr.
Ignacio Morones Prieto', with registration number 84-17 (Folio
CONBIOET ICA-24-CEI-001-201604279). The study was conducted in
accordance with the Declaration of Helsinki. The informed consent
signature was obtained from each participant prior to the
collection of the urine samples.
Sample collection and treatment
Three urine samples were collected per participant
in a sterile vessel: The first during the 1st trimester (8-20
weeks), the second during the 2nd trimester (24-28 weeks) and
finally in the 3rd trimester (32-39 weeks), as summarized in
Table SII.
After collection, the urine samples were centrifuged
at 300 × g for 15 min at room temperature to remove cells and cell
debris. The samples were then centrifuged again at 1,200 × g for 10
min at 4°C and divided into 10 ml aliquots. A cocktail of protease
inhibitors (PMSF, NaN3 and leupeptin) (18) was added to each aliquot and they
were stored at -80°C until analysis.
Isolation of urinary exosomes
Composite samples (pools) were established prior to
the isolation of exosomes and the amplification of miRNAs, mixing
equivalent amounts of individual samples. A total of 5 pools
containing 34 individual samples and 5 pools containing 27
individual samples were established from the healthy controls and
GDM groups, respectively. Each pool was matched by age and body
mass index (BMI).
Exosomes were purified from 10 ml of pooled urine
samples using the Urine Exosome Purification and RNA Isolation Midi
kit (Norgen Biotek Corp, Product # 58700) following the
manufacturer's instructions. The Urine Exosome Purification kit
isolates exosomes based on the isoelectric point of the proteins
present in the microvesicles that are filtered through silicon
carbide columns, which makes the method sensitive and specific
(19). Purified exosomes were
stored at -80°C for subsequent analysis.
Cryogenic transmission electron
microscopy (cryo-TEM)
The morphology and size of the exosomes were
examined by cryo-TEM. A volume of 3 to 5 µl of exosome
samples suspended in phosphate-buffered saline was transferred to a
Lacey carbon-coated grid. The grid was maintained at 85% humidity
for 10 sec and then was immersed in liquid ethanol prepared without
ice crystals at liquid nitrogen temperature (-196°C). Subsequenlty,
the grid was mounted on a cryo-transfer sample holder at -175°C.
The preparation was observed using a cryo-TEM Microscope JEM-2100
containing a LaB6 filament operated at different
acceleration voltages (80,100,120,160 and 200 kV).
Identification of exosomal surface
protein markers by western blot analysis
The identity of exosomes was assessed by western
blot analysis, evaluating the presence of the surface proteins,
CD9, CD63 and CD81. Additionally, the placental origin was
evaluated by the presence of placental alkaline phosphatase (PLAP).
Electrophoresis was performed under reducing conditions (12.5%
SDS-PAGE), from 20 µg of protein, extracted using the Urine
Exosome Purification and RNA Isolation Midi kit (Norgen Biotek
Corp., product #58700). The protein concentration was determined
using the bicinchoninic acid (BCA) method using the Pierce BCA
Protein Assay kit (Pierce; Thermo Fisher Scientific, Inc.) and 20
µg of protein was loaded by lane. Western blot analysis was
performed according to standard procedures using polyvinylidene
difluoride membranes and an enhanced chemiluminescence system
Optiblot ECL Substrate kit (ab133406, Abcam United Kingdom). The
following antibodies were used: Anti-CD9 mouse monoclonal antibody
(isotype IgG1; host species, mouse; this antibody reacts with human
CD9 antigen, ab2215, Abcam United Kingdom, 1:1,000), anti-CD81
mouse monoclonal antibody (isotype IgG1; host species, mouse; this
antibody reacts with human CD81 antigen, ab79559, Abcam United
Kingdom, 1:1,000), anti-CD63 mouse monoclonal antibody, (isotype
IgG1; host species, mouse; this antibody reacts with human CD63
antigen, ab8219, Abcam United Kingdom, 1:1,000) and anti-PLAP
rabbit monoclonal antibody (isotype IgG; host species, rabbit;
ab133602, Abcam United Kingdom, 1:500). The primary antibodies were
incubated at room temperature for 2 h. The secondary antibodies
used were as follows: Goat polyclonal secondary antibody to mouse
IgG-H&L (HRP) (ab6789) and goat anti-rabbit IgG H&L(HRP)
(ab6789), incubated overnight at 4°C. As a loading control,
vesicles isolated from HepG2 [HEPG2] cells (ATCC®
HB-8065™) were used. The stability of urinary exosomes was
evaluated after 12 months of storage at -80°C by western blot
analysis and cryo-TEM.
miRNA isolation and quantification
Total RNA was isolated from 400 µl of
purified exosomes using the Urine Exosome Purification and RNA
Isolation Midi kit (Norgen Biotek Corp.; cat. no. 58700) following
the manufacturer's instructions. The RNA concentration was assessed
using a NanoDrop 1000 spectrophotometer (NanoDrop Technologies
Inc.). The cDNAs for the mature miRNAs (U6, miR-16-5p, miR-222-3p,
miR-516b-5p, miR-517-5p and miR-518a-3p) were synthesized from 150
ng of total RNA using the Taq Man® microRNA test,
(Applied Biosystems); TaqMan probes were used for each of the
miRNAs to avoid unspecific amplification. The 5'-3' sequences of
the primers are presented in Table
SIII. The CFX96 touch thermal cycler was used (Bio-Rad
Laboratories, Inc.).
All RT-qPCR reactions were performed in duplicate;
Cq values were averaged and the 2−ΔΔCq method (20) was used to obtain the relative
expression, where ΔΔCq was calculated by subtracting the
ΔCq value from the mean of the control group (healthy
pregnant women) with the ΔCq value of GMD women. miRNA
levels were normalized with the use of the average of U6 snRNA Cq
value as the housekeeping gene. The experimental conditions for
reverse transcription and PCR are detailed in Tables SIV and SV, respectively.
miRNA target prediction
The evaluation for the interaction and target genes
networks was performed using the miRTar-getLink program, which
contains experimentally validated interactions in the latest
version of miRTarBase 1 (version 6.0: September 15, 2015). The
functional evaluation of the miRNAs was performed using
DIANA-miRPath v3.0 (http://www.microrna.gr/miRPathv3). Additionally, using
The Kyoto Encyclopedia of Genes and Genomes (KEGG, https://www.genome.jp/kegg/) bioinformatics tool,
signaling pathways related to the study of miRNAs were evaluated,
considering a value of P<0.05 as statistically significant.
Statistical analysis
Statistical analysis was performed with GraphPad
Prism 5.0 software (GraphPad Software, Inc.). The data are
expressed as the means ± SD or median ± IQR according data
distribution, respectively. For the relative expression of the
miRNAs, a Kruskal-Wallis ANOVA test and a non-parametric Dunn
post-hoc test were performed. A value of P<0.05 was considered
to indicate a statistically significant difference. The GDM
predictive capacity was evaluated by means of receiver operating
characteristic (ROC) curve analysis.
Results
Clinical characteristics of the
population studied
The anthropometric, clinical and biochemical
parameters of the pregnant women included in the present study are
summarized in Table I. The mean
age of the patients with GDM was 29.9±6.03 and 26.06±5.28 years for
the healthy controls. The BMI was significantly higher in women
that developed GDM (28.66±4.02 vs. 25.69±4.03, P=0.006; Table I).
 | Table IGeneral characteristics of pregnant
women included in the study. |
Table I
General characteristics of pregnant
women included in the study.
| Clinical data | GDM (n=27) | Healthy (n=34) | P-value |
|---|
| Age (years) | 29.93±6.03 | 26.06±5.28 | 0.01 |
| BMI
(kg/m2) | 28.66±4.02 | 25.69±4.03 | 0.006 |
| Glucose
(mg/dl) | 89.60±14.15 | 78.00±8.48 | 0.0002 |
| Systolic blood
pressure (mmHg) | 110.4±9.29 | 105.7±9.80 | 0.06 |
| Diastolic blood
pressure (mmHg) | 69.44±8.59 | 68.74±9.96 | 0.77 |
| Urea (mg/dl) | 14.18±3.62 | 17.84±5.99 | 0.007 |
| Creatinine
(mg/dl) | 0.55±0.08 | 0.57±0.12 | 0.35 |
| Hemoglobin
(g/dl) | 13.45±1.23 | 12.68±1.28 | 0.021 |
| Leukocytes
(×103/µl) | 8.64±2.06 | 8.40±1.77 | 0.64 |
Isolation and characterization of
placental exosomes from urine
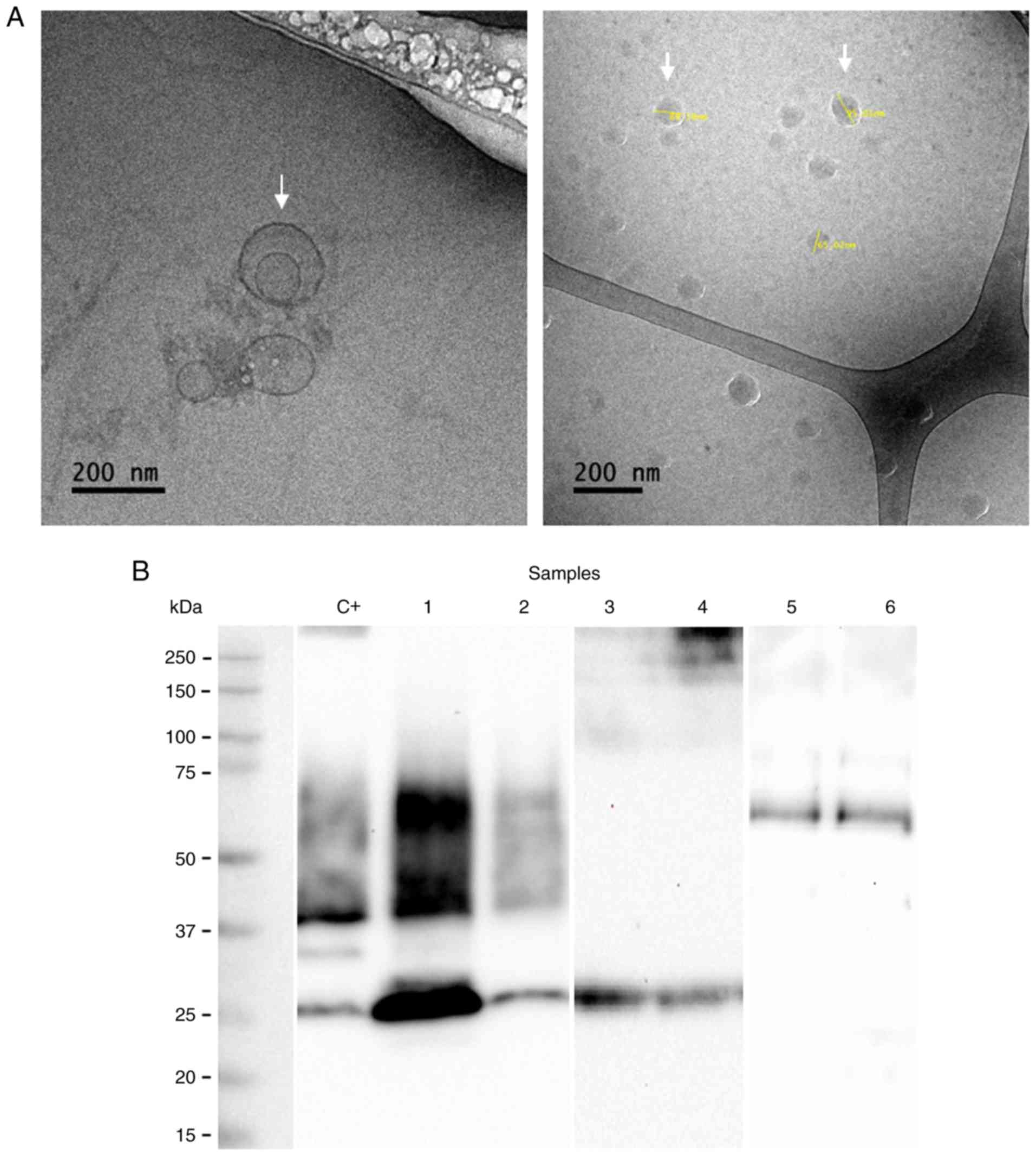
The presence of exosomes was demonstrated by
morphological (cryo-TEM) and molecular characterization (western
blot analysis) after 4 and 12 months of storage. The isolated
exosomes exhibited a spherical shape with a size between 80 and 140
nm (Fig. 1A). The arrows in
Fig. 1A indicate the presence of
exosomes. The presence of CD63, CD9 and CD81 proteins associated
with the exosome identity was corroborated by western blot analysis
(Fig. 1B). In addition, the
presence of PLAP indicates the placental origin of these vesicles
(Fig. 1B).
Differential expression of placental
exosomal miRNAs in the urine of patients with GDM and healthy
pregnant women
The relative expression of each miRNA was determined
by RT-qPCR, using the 2−ΔΔCq method and U6 as a
constitutive expression gene for normalization. In the 1st
trimester of pregnancy, non-significant differences were observed
in the expression of the five analyzed miRNAs. However,
statistically significant differences were found between the study
groups in the 2nd and 3rd trimester of pregnancy, for the miRNAs
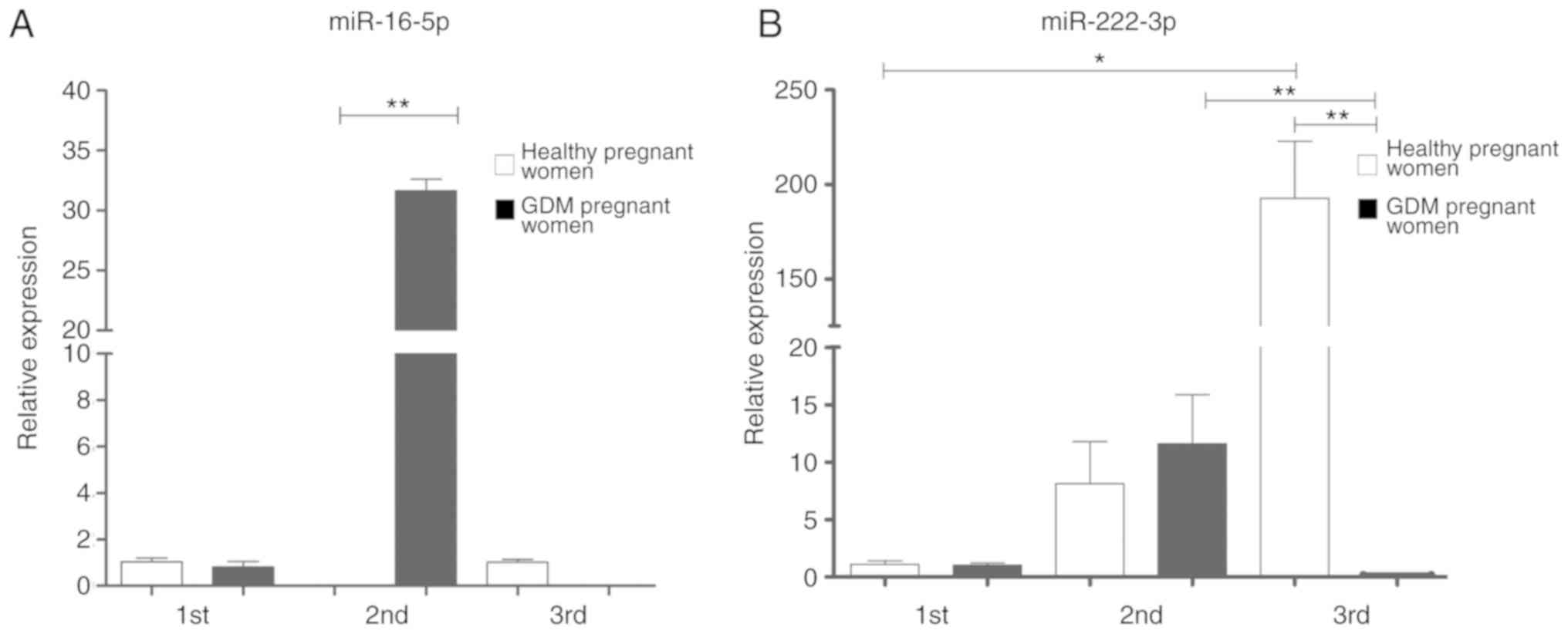
coded by C19MC, as well as for miR-222-3p (Figs. 2 and 3).
The expression of miR-222-3p in the placental
exosomes from healthy women increased significantly as the
pregnancy progressed, as shown in Fig. 2; however, the expression of this
miRNA in the pregnant women with GDM significantly decreased during
the 3rd trimester in comparison to the healthy controls (Fig. 2).
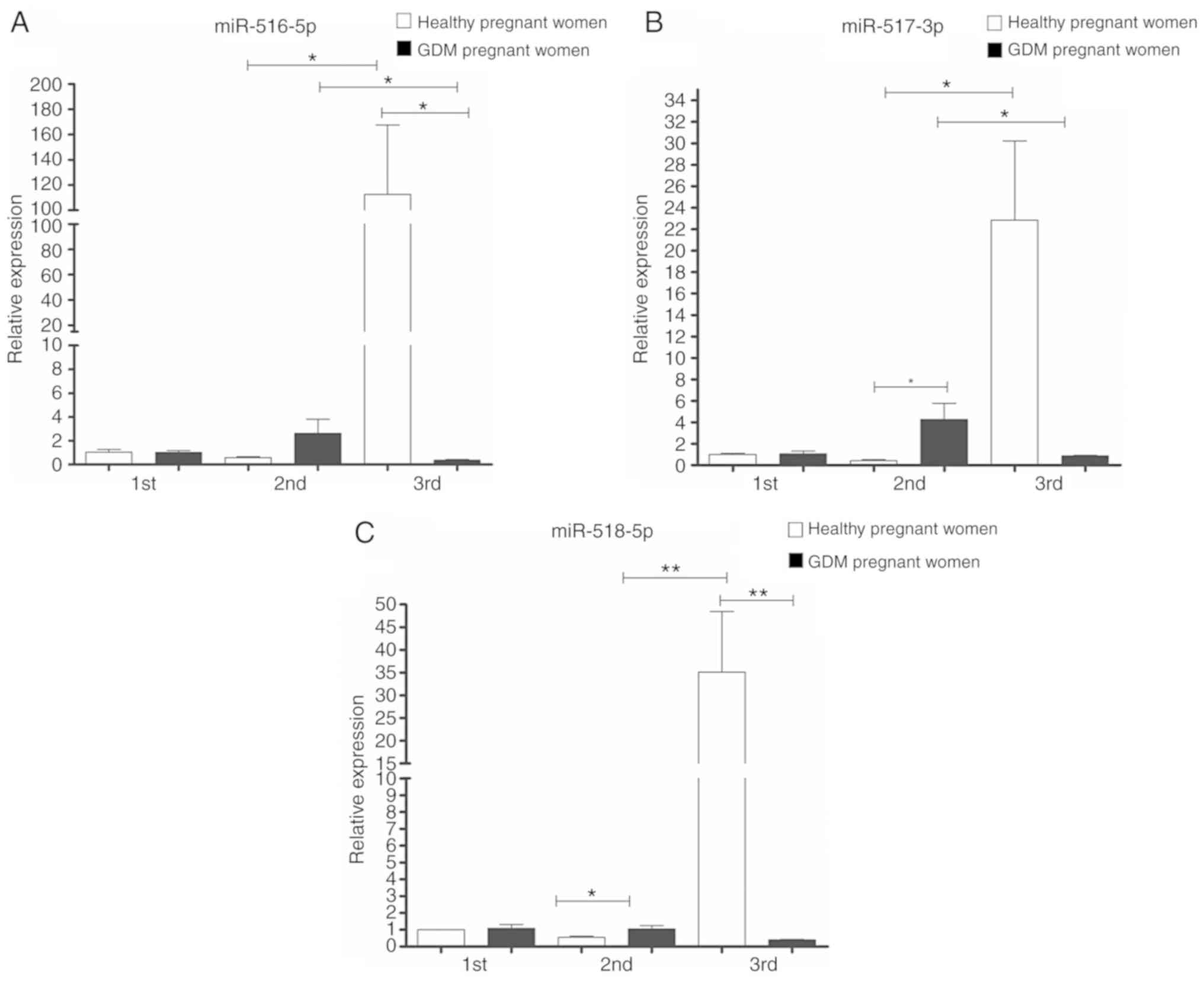
Regarding the expression of 'trophomiRs'
(miR-516-5p, miR-517-3p and miR-518-5p), the expression profile
observed in samples from healthy pregnant women exhibited an
increase in their expression as the pregnancy progressed, from the
1st to the 3rd trimester. In the case of the GDM group, the
expression of these trophomiRs increased from the 1st to the 2nd
trimester, with miR-517-3p and miR-518-5p exhibiting statistically
significant differences (upregulated) compared to the healthy
pregnant women. In the GDM group, a marked decrease was observed in
the 3rd trimester and the expression levels of the three miRNAs
were significantly downregulated compared to the healthy pregnant
group (Fig. 3).
Since the expression levels of miR-16-5p, miR-517-3p
and miR-518-5p in patients with GDM were differed markedly from
those of the healthy controls in the 2nd trimester, the diagnostic
potential of these miRNAs was further investigated. A ROC curve
constructed based on exosomal miRNAs levels in the early 2nd
trimester, revealed that the area under the curve (AUC) was 1 (CI
1-1) for miR-16-5p, miR-517-3p and miR-518-5p, indicating the high
diagnostic accuracy of these miRNAs for the differentiation between
patients with GDM and healthy women. The performance of these
miRNAs to predict GDM is shown in Table II.
| Table IIROC analysis of the microRNAs. |
Table II
ROC analysis of the microRNAs.
| MicroRNA | AUC | IC 95% | Standard error | P-value |
|---|
| miR-16-5p | 1.00 | 1.00-1.00 | 0 | 0.009 |
| miR-222-3p | 0.688 | 0.2893-1.086 | 0.203 | 0.387 |
| miR-516b-5p | 0.938 | 0.7615-1.113 | 0.090 | 0.043 |
| miR-517-5p | 1.00 | 1.00-1.00 | 0 | 0.034 |
| miR-518-3p | 1.00 | 1.00-1.00 | 0 | 0.021 |
Evaluation of interaction networks of
common target genes between the studied miRNAs
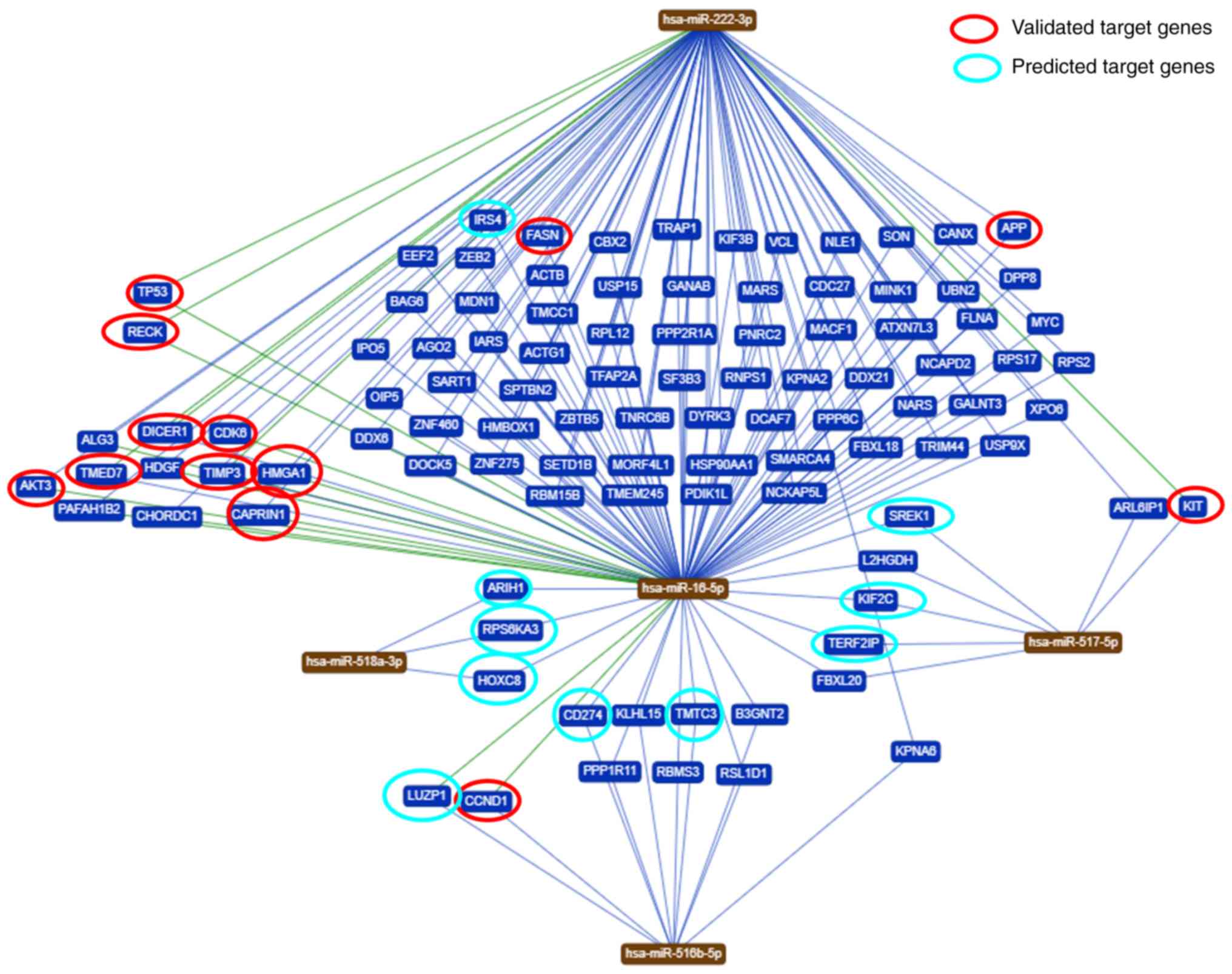
To identify genes regulated by the five miRNAs
analyzed in the present study, a bioinformatics analysis was
performed. Based on the bioinformatic analysis, it was demonstrated
that these miRNAs are predicted to regulate different target genes.
In total, 102 genes were shared among the five miRNAs evaluated in
the present study (Fig. 4). An
in-depth analysis of the potential functions of the five miRNAs
under study (https://ccb-web.cs.unisaarland.de/mirtargetlink/),
revealed that some of the target genes predicted by the
bioinformatics analysis have been previously experimentally
validated or predicted (Table
III).
| Table IIIMicroRNAs validated and/or predicted
target genes. |
Table III
MicroRNAs validated and/or predicted
target genes.
| Target Official
symbol | Official full
name | Sequence accession
ID (Gene) | MicroRNA associated
with regulation |
Predicted/validated | Gene function |
|---|
| FASN | Fatty acid
synthase | NM_004104.5 | miR-16-5p | Validated
(RT-qPCR) | Regulate components
of the insulin/PI3K-AKT (21). |
| AKT3
(miRTarBase ID MIRT003329) | AKT
serine/threonine kinase 3 | NM_005465.7 | miR-16-5p | Validated
(luciferase reporter assay) | Involved in a wide
variety of biological processes including cell proliferation,
differentiation, apoptosis, tumorigenesis, as well as glycogen
synthesis and glucose uptake. Key regulators of endothelial cell
functions and angiogenesis (22). |
| CAPRIN1
(miRTarBase ID MIRT005086) | Cell cycle
associated protein 1 | NM_005898.5 | miR-16-5p | Validated
(luciferase reporter assay/RT-qPCR/western blot analysis) | Involved in cell
proliferation (23). |
| HMGA1
(miRTarBase ID MIRT000270) | High mobility group
AT-hook 1 | NM_002131.4 | miR-16-5p | Validated
(luciferase reporter assay/RT-qPCR/western blot analysis) | Involved in cell
proliferation and in the development of human malignancies
(23,24). |
| CDK6
(miRTarBase ID MIRT000939) | Cyclin-dependent
kinase 6 | NM_001259.8 | miR-16-5p | Validated
(RT-qPCR/flow cytometry/luciferase reporter assay/western blot
analysis) | Gene related to the
cell cycle. Control of the G1/S transition (25). |
| TP53
(miRTarBase ID MIRT005764) | Tumor protein
p53 | NM_000546.6 | miR-16-5p and
miR-222-3p | Validated (western
blot anal ysis/luciferase reporter assay) | Encodes a tumor
suppressor protein that contains transcriptional activation, DNA
binding, and oligomerization domains. The encoded protein responds
to various cellular stresses to regulate the expression of target
genes, leading to cell cycle arrest, apoptosis, senescence, DNA
repair, or changes in metabolisms (26,27). |
| RECK
(miRTarBase ID MIRT031373) | Reversion inducing
cysteine rich protein with kazal motifs | NM_021111.3 | miR-16-5p and
miR-222-3p | Validated
(luciferase reporter assay/RT-qPCR/western blot analysis) | Associated with
cellular apoptosis. Suppresses tumor invasion and metastasis by
downregulating the activity of matrix metalloproteinase 9 (MMP9)
(28). |
| APP
(miRTarBase ID MIRT031838) | Amyloid-beta
precursor protein | NM_000484.4 | miR-16-5p | Validated
(ELISA/immunopre cipitation/luciferase reporter
assay/RT-qPCR/western blot analysis) | Regulator Aβ, Tau,
inflammation and oxidative stress in Alzheimer's disease (29). |
| CCND1
(miRTarBase ID MIRT001225) | Cyclin D1 | NM_053056.3 | miR-16-5p | Validated
(luciferase reporter assay/RT-qPCR/western blot analysis/flow
cytometry) | Well-known
regulator of cell cycle progression (30-32). |
| IRS4 | Insulin receptor
substrate 4 | NM_003604.2 | miR-16-5p | Predicted | Involved in cell
growth and glucose homeostasis (33). |
| DICER1
(miRTarBase ID MIRT006377) | Dicer 1,
ribonuclease III | NM_030621.4 | miR-222-3p | Validated
(luciferase reporter assay) | It plays a crucial
role in the biogenesis of small regulatory RNAs, such as microRNAs
(miRNAs) and small interfering RNAs (siRNAs). Dicer has also been
found to participate in chromosomal fragmentation during apoptosis
or in inflammatory processes (34). |
| TMED7
(miRTarBase ID MIRT002334) | Transmembrane p24
trafficking protein 7 | NM_181836.6 | miR-222-3p | Validated (western
blot analysis; RT-qPCR) | Involved in
vesicular protein trafficking between endosomal structures in
resting cells and the Golgi apparatus, therefore, it is
functionally similar to the Rab family (35,36). |
| TIMP3
(miRTarBase ID MIRT003451) | TIMP
metallopeptidase inhibitor 3 | NM_000362.5 | miR-222-3p | Validated (flow
cytometry/immunohistochemistry/luciferase reporter
assay/RT-qPCR/western blot analysis/immunohistochemistry/in
situ hybridization/ELISA) | The proteins
encoded by this gene family are inhibitors of the matrix
metalloproteinases, a group of peptidases involved in degradation
of the extracellular matrix (ECM). Expression of this gene is
induced in response to mito genic stimulation (37,38). |
| KIT
(miRTarBase ID MIRT001779) | KIT proto-oncogene,
receptor tyrosine kinase | NM_000222.3 | miR-222-3p | Validated
(luciferase reporter assay/RT-qPCR/microarray/western blot
analysis) | Related to cell
proliferation and melanogenesis (39,40). Involved in erythropoiesis,
erythroleukemic cell growth, and TRAIL apoptotic signal in lung
cancer cells (41). Oncogene in
many types of cancer, involved in the pathogenesis of PTC (42). Promotes expansion of the first
erythroblasts, linked to cancer therapy (43). Regulation of gastrointestinal
stromal tumors (GIST) (44). |
| ARIH1
(miRTarBase ID MIRT549302) | Ariadne RBR E3
ubiquitin-protein ligase 1 | NM_005744.5 | miR-518a-3p | Predicted [PAR-CLIP
(45)] | Associated with
cellular proliferation and modification of proteins in nuclear
bodies (46). Potent mediator of
DNA damage-induced translation arrest that protects stem and cancer
cells against genotoxic stress (47). |
| RPS6KA3
(miRTarBase ID MIRT554630) | Ribosomal protein
S6 kinase A3 | NM_004586.3 | miR-518a-3p | Predicted [PAR-CLIP
(48)] | Associated with
cellular processes such as cell growth, motility, survival, and
proliferation (49). Related to
Coffin-Lowry syndrome (50). |
| HOXC8
(miRTarBase ID MIRT562161) | Homeobox C8 | NM_022658.4 | miR-518a-3p | Predicted [PAR-CLIP
(45)] | Associated to
cellular processes such as cell growth, motility, survival, and
proliferation in cancer cells (51-53). |
| LUZP1
(miRTarBase ID MIRT444756) | Leucine zipper
protein 1 | NM_033631.4 | miR-516b-5p | Predicted [PAR-CLIP
(54)] | Involved in
exosomal processes, ventricular septum development (55,56). |
| CD274
(miRTarBase ID MIRT445289) | CD274 molecule | NM_001267706.1 | miR-516b-5p | Predicted [PAR-CLIP
(54)] | Expressed by T
cells, B cells, and monocytes is a potent regulator of immune
responses (57). Programmed death
ligand 1 (PD-L1, CD274) has been reported to be expressed
abnormally in many cancers (58). |
| TMTC3
(miRTarBase ID MIRT445289) | Transmembrane
O-mannosyltransferase targeting cadherins 3 | NM_181783.4 | miR-516b-5p | Predicted [PAR-CLIP
(59)] | Role in proteolysis
and stress response in the endoplasmic reticulum. It can play a
direct role in the physiology of immune cells in long-term graft
function (60). |
| KIF2C
(miRTarBase ID MIRT455884) | Kinesin family
member 2C | NM_001297655.2 | miR-517-5p | Predicted [PAR-CLIP
(59)] | Modulator in
microtubule depolymerization, bipolar spindle formation, and
chromosome segregation, has been reported to take roles in cancer
biology (61,62). |
| TERF2
(miRTarBase ID MIRT490157) | TERF2 interacting
protein | NM_018975.4 | miR-517-5p | Predicted [PAR-CLIP
(48)] | Folding human
telomeres into loops to prevent unwanted DNA repair and final
binding of chromosomes (63). |
| SREK1
(miRTarBase ID MIRT611178) | splicing regulatory
glutamic acid and lysine rich protein 1 | NM_001323533.2 | miR-517-5p | Predicted [PAR-CLIP
(64)] | Nucleic acid
binding and nucleotide binding (55,56). |
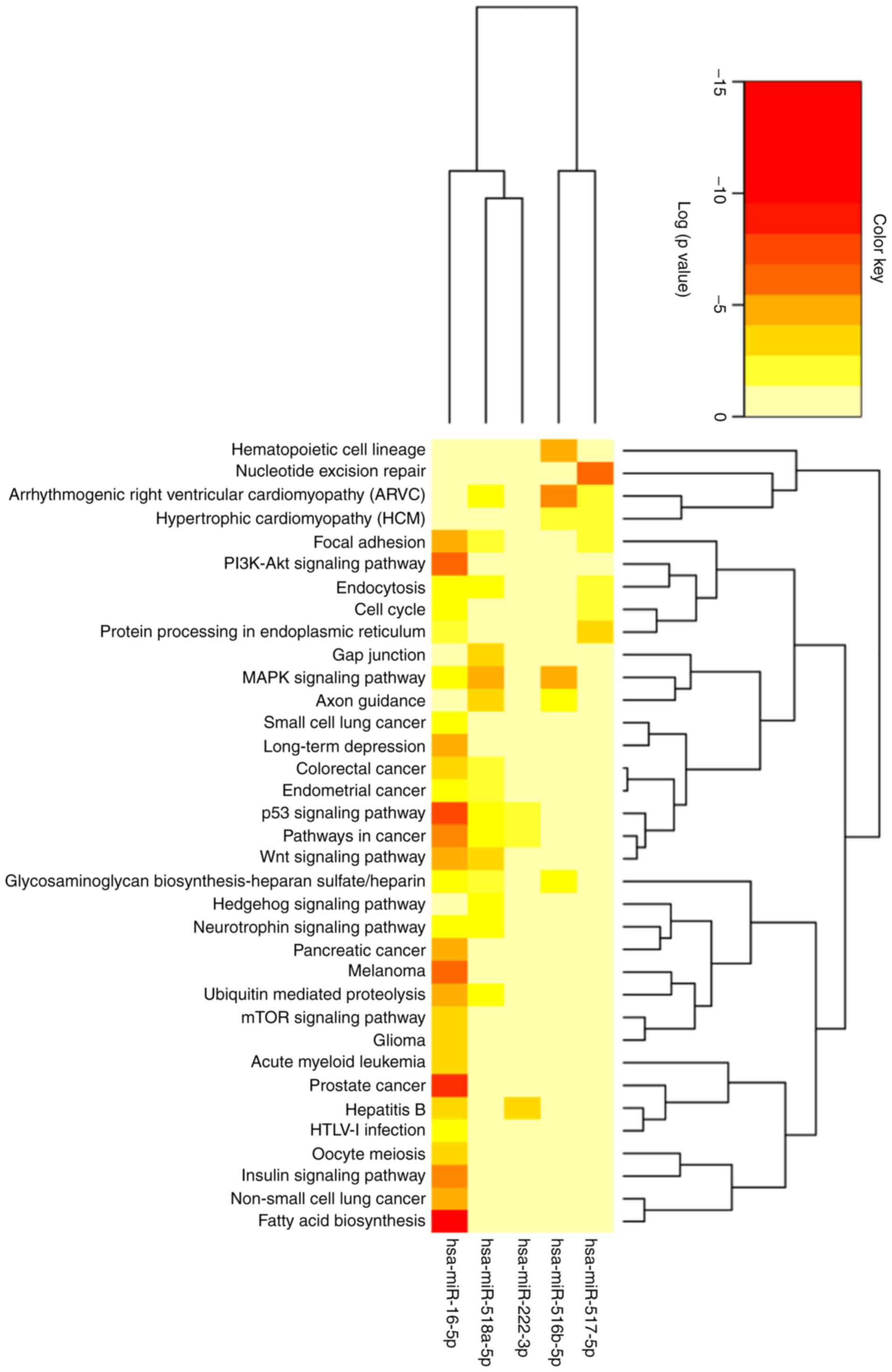
The 102 genes identified were related to 62 pathways
predicted to be involved in different metabolic, molecular and cell
regulation processes. In total, 32 routes presented a value of
P<0.05, indicating that these particular pathways are regulated
by at least one of the studied miRNAs. Some of the most
significative pathways are the biosynthesis of fatty acids, the
signaling pathway of PI3K-Akt and the signaling of insulin, as
shown in Fig. 5 and in further
detail in Table SVI.
Discussion
To date, there are several methods wich can be used
to examine miRNA profiling, such as RT-qPCR, microarrays and direct
sequencing. Each method has its advantages and limitations;
however, RT-qPCR seems to have better sensitivity and is the only
platform capable of generating absolute quantification. In the
present study, as well as in several studies (65), RT-qPCR techniques were used to
validate candidate biomarkers identified through a literature
search or based on previous findings (66-69).
The relative expression of miR-516-5p, miR-517-3p,
miR-518-5p, miR-222-3p and miR-16-5p in urinary placental exosomes
from patients with GDM were evaluated in the present study, at
different stages of pregnancy. To the best of our knowledge, there
is no previous study available in the literature to date
characterizing these miRNAs expressed in urinary placental
exosomes.
It was found that the miRNAs expression levels
differed across pregnancy. Accordingly, it has been reported that
in the maternal plasma, a substantial amount of
placental-associated miRNAs are differently expressed during the
1st, 2nd and 3rd trimester of gestation, probably to meet the
different regulatory demands of pregnancy (70).
In the present study, women with GDM exhibited
additional risk factors, such as being older and overweight,
compared to healthy pregnant women; both conditions are related to
high adiposity, which favors a pro-inflammatory response by
producing adipokines. Both the production of adipokines and the
high levels of glucose in circulation are considered factors that
induce the overexpression of miR-16-5p (14). In the case of miR-16-5p, it was
found that in the 2nd trimester, its expression was only detectable
in exosomes purified from patients with GDM, while in healthy
controls, it was undetectable. miR-16-5p is a modulator of the
PI3K/Akt signaling pathway, through the regulation of genes, such
as Pi3Kr1 and Pi3kr3, mTOR and Mapk3, among
others. In the present study, bioinformatics analysis revealed that
the signaling pathways of PI3K/Akt (P<0.0001), Wnt
(P<0.0001), insulin (P=0.004) and mTOR (P=0.016) were
significantly regulated by miR-16-5p (Table SVI). The overexpression of these
cell signaling pathways has been associated with diabetes mellitus
and GDM (21). In addition, genes
encoding for proteins 1 and 2 of the insulin receptor substrate
(IRS1/IRS2) are targets of miR-16-5p (21,71). Therefore, the upregulation of
miR-16-5p in patients with GDM at the 2nd trimester, will result in
a negative regulation of IRS1 and IRS2, which could lead to
abnormal Wnt/β-catenin signaling and, eventually, in diabetes
(72). However, following the
diagnosis of GDM, patients were subjected to pharmacological
treatment (metformin, insulin, or a combination of both), diet and
exercise. It is suggested that treatment with metformin may
influence the pattern of expression observed in the 3rd trimester
of pregnancy. Ortega et al identified that the circulating
expression of miR-140-5p (-16%, P=0.004) and miR-222 (-47%, P=0.03)
decreased significantly following metformin treatment in patients
with type 2 diabetes (T2D) (73).
Furthermore, Demirsoy et al demonstrated that a total of 13
miRNAs were found significantly downregulated following the
treatment of patients with T2D with metformin (P<0.05) (74). The effect of the treatment,
together with the complex and dynamic expression of miRNAs, which
vary according to the requirements in the regulation of specific
stages of development, may explain the differential expression of
miRNAs across gestation.
A similar profile was observed for miR-222-3p, with
the only difference that this microRNA was also expressed in the
second trimester of healthy pregnancy. On the other hand, the role
of miR-222-3p in the development of T2D has been previously
reported and has been associated with the pathogenesis of GDM
(75). Zhao et al reported
that miR-222-3p expression was significantly decreased in women
with GDM compared to controls at 17 weeks of gestation in serum
samples (76). Similarly,
Pheiffer et al reported a decrease in the expression of this
miRNA in the serum of women with GDM compared to the controls at 26
weeks of gestation (77).
Furthermore, the expresion of this miRNA increases in the adipose
tissue of women with GDM, and negatively correlates with the levels
of glucose transporter 4 (GLUT4) and estrogen receptor (ER)
(78). It is important to
consider that, since expression miR-222-3p is dependent on tissue
and insulin sensibility, the expression between adipose tissue and
placenta could be differential. Recently, the authors reported that
in the urine metabolome profile of women with GDM in the 3rd
trimester of pregnancy, 14 metabolites related with the steroid
hormone biosynthesis and tryptophan metabolism were significantly
elevated [i.e., L-tryptophan, urobilinogen, ceramide (d18:0/23:0),
21-deoxycortisol]. The upregulation of these pathways could trigger
insulin resistance and may respond to oxidative stress and
inflammation during GDM (79).
The majority of the genes involved in the insulin resistance
pathway are targets of miR-16-5p and miR-222-3p (8,80).
These miRNAs, contained in exosomes, can reach different tissues
and organs, thereby affecting metabolic processes in patients with
GDM.
In the present study, the amplification of miRNAs
coded by the C19MC (miR-516b-5p, miR-517-5p and miR-518a-3p) is an
indirect evidence of the placental origin of the exosomes that were
purified, which agrees with the detection of the exosomal marker,
PLAP, by western blot analysis. The expression profile of this
particular set of miRNAs was characterized by an increase across
gestation in healthy pregnant and no statistically significant
differences were found between both study groups in the 1st
trimester. However, in women with GDM the expression was
differential, and it was characterized by an increase in the 2nd
trimester followed by a reduction in the 3rd trimester of
pregnancy.
To date, there are only a few studies available on
the altered expression of C19MC miRNAs in pregnancy disorders
(13,81,82). None of these studies however, have
evaluated the expression of these miRNAs in urinary exosomes
derived from the placenta and, for this reason, the results of the
expression levels obtained in the present study cannot bed compared
directly with those of other reports in the literature.
Hromadnikova et al reported that 1st trimester circulating
plasma exosomes possess the identical C19MC miRNA expression
profile as placental tissues derived from patients with gestational
hypertension, preeclampsia and fetal growth restriction following
labor (83). Further studies are
required to elucidate whether the downregulation of C19MC miRNAs in
placental exosomes is associated with the pathophysiology of GDM or
if it is a compensatory mechanism.
The bioinformatics prediction carried out in the
present study through KEGG revealed that miR-16-5p, miR-222-3p,
miR-516b-5p, miR-517-5p and miR-518a-3p shared common target genes.
The fact that redundant regulation through several miRNAs is
required to modulate a physiological process constitutes a security
molecular process. Some of the target genes shared by the 5 miRNAs
selected in the present study (irs4, galnt, reck, alg3, akt3,
timp3, kit, l2hgdh, ki2fc, rap1, fb12, hoxc8 and pd-l1)
are expressed in a variety of tissues, including placenta and
adipose tissue. Exosomes potentially reach these tissues,
reflecting placental regulation as part of maternal-fetal
communication. Future functional studies are needed to elucidate
the effects of microRNAs expression on maternal and fetal tissues
and also to follow offspring during extrauterine life.
The present study supports the use of
placenta-derived exosomes as part of biomarker screening strategies
to identify women at risk of later development of pregnancy-related
complications. The placental exosomes released to the systemic
circulation and collected in the urine represent a non-invasive
source of signaling molecules, including miRNAs, whose abnormal
expression profile due to defective placental functions, reflects
the expression profile of the trophoblast cells and, consequently,
provides more precise information about pregnancy disorders. In the
1st trimester of gestation, at least for the selected set of miRNAs
under study, we were not able to find significant differences in
the levels of expression in GDM or healthy patients. At the 2nd
trimester, 3 miRNAs (mir-16-5p, miR-517-3p and miR-518-5p,
exhibited a good diagnostic value. However, since the sample size
in the present study was small, these results need to be
interpreted with caution and further studies, including an external
validation cohort with a larger size number are required.
The scenery for validating exosomal miRNAs for their
intended use in clinical applications is still complex. To date,
the most promising markers of gestational diabetes are: miR-29a,
miR-222, miR-16-5p, miR-17-5p and miR-20a-5p (14,71,76,78,84). In future miRNAs expression studies
during pregnancy, several aspects should be taken into account:
Gestational age (since the expression of miRNAs changes through
pregnancy), the type of sample studied, the sex of the offspring
and the route of delivery. Therefore, these variables must be
established between study groups. Further analyses need to be
conducted to explore a larger population to validate these findings
and to evaluate their diagnostic predictive value at the 1st
trimester of gestation. Despite this limitation, the results
revealed herein demonstrate alterations in the expression of miRNAs
across gestation and a differential expression profile within
urinary exosomes as a result of pathophysiological alterations in
women with GDM. In general, miRNAs that regulate the innate and
specific immune system are expressed throughout pregnancy. By
contrast, miRNAs that promote cell differentiation and control cell
cycle progression are mainly expressed in advanced gestation.
In conclusion, the expression profile of miRNAs
carried as the molecular cargo of placental urinary exosomes is
different between healthy and women with GDM; the differences are
evident and quantifiable from the 2nd trimester, being even more
evident towards the 3rd trimester. The alteration of miRNA
expression modulates pathways related to the development of insulin
resistance, pro-inflammatory response and metabolic homeostasis,
which are implicated in the physiopathology of GDM. The expression
of miR-16-5p in the 2nd trimester may be an important molecular
mechanism involved in insulin resistance in women who develop GDM,
although validation studies are required. C19MC miRNAs also exhibit
a differential profile in patients with GDM in comparison to
healthy controls, being upregulated in the second trimester in the
former group followed by a reduction in the third trimester of
pregnancy. The differential expression of miRNAs in various
development steps is probably to meet the different regulatory
demands of pregnancy. To the best of our knowledge, the present
study is the first to report the expression of miRNAs in placental
urinary exosomes, suggesting that urine is a potential biological
fluid for the research of pathological conditions during
pregnancy.
Supplementary Data
Funding
The present study was funded by CONACYT-Fondo
Sectorial de Salud (grant no. 290239 to YLH).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YLH and MSB were involved in the study
conceptualization. ASHVO, NVDF, EMM, DPPP and DAGL were involved in
the study methodology. MSB was involved in software analysis. ASHVO
and JAL were involved in data validation. YLH, MSB, EMM, DPPP and
ASHVO were involved in formal analysis. DEN was involved in the
analysis and interpretation of data. JCTO was involved in the
investigative aspects of the study. YLH was responsible for the
study resources. ASHVO was involved in the writing and preparation
of the original draft of the manuscript. YLH, EMM and JAL were
involved in the writing, reviewing and editing of the manuscript.
DEN was involved in the critical revision of the manuscript. MSB
and YLH supervised the study. YLH was involved in project
administration and in funding acquisition. All authors have read
and agreed to the published version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Hospital Central 'Dr. Ignacio Morones Prieto',
with registration number 84-17 (Folio CONBIOETICA-24-
CEI-001-201604279). The study was conducted in accordance with the
Declaration of Helsinki. The informed consent signature was
obtained from each participant before the collection of urine
samples.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank the participation of
Medical students of the specialty of Gynecology at the Hospital
Central 'Dr.Ignacio Morones Prieto' for their support in patient
recruitment.
References
|
1
|
Basina M: Gestational diabetogenesis. J
Women's Health Care. 01:e1062012. View Article : Google Scholar
|
|
2
|
Sáez T, de Vos P, Sobrevia L and Faas MM:
Is there a role for exosomes in foetoplacental endothelial
dysfunction in gestational diabetes mellitus? Placenta. 61:48–54.
2018. View Article : Google Scholar
|
|
3
|
Powe CE: Early pregnancy biochemical
predictors of gestational diabetes mellitus. Curr Diab Rep.
17:122017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rodrigo N and Glastras SJ: The emerging
role of biomarkers in the diagnosis of gestational diabetes
mellitus. J Clin Med. 7:1202018. View Article : Google Scholar :
|
|
5
|
Akgöl E, Abuşoğlu S, Gün FD and Ünlü A:
Prevalence of gestational diabetes mellitus according to the
different criterias. Turk J Obstet Gynecol. 14:18–22. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Karcaaltincaba D, Calis P, Ocal N, Ozek A,
Altug Inan M and Bayram M: Prevalence of gestational diabetes
mellitus evaluated by universal screening with a 75-g,2-h oral
glucose tolerance test and IADPSG criteria. Int J Gynecol Obstet.
138:148–151. 2017. View Article : Google Scholar
|
|
7
|
Ilekis JV, Tsilou E, Fisher S, Abrahams
VM, Soares MJ, Cross JC, Zamudio S, Illsley NP, Myatt L, Colvis C,
et al: Placental origins of adverse pregnancy outcomes: Potential
molecular targets: An executive workshop summary of the eunice
kennedy shriver national institute of child health and human
development. Am J Obstet Gynecol. 215(1 Suppl): S1–S46. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guarino E, Poggi CD, Grieco GE, Cenci V,
Ceccarelli E, Crisci I, Sebastiani G and Dotta F: Circulating
MicroRNAs as biomarkers of gestational diabetes mellitus: Updates
and perspectives. Int J Endocrinol. 2018:63804632018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kowal J, Tkach M and Théry C: Biogenesis
and secretion of exosomes. Curr Opin Cell Biol. 29:116–125. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cuffe JSM, Holland O, Salomon C, Rice GE
and Perkins AV: Review: Placental derived biomarkers of pregnancy
disorders. Placenta. 54:104–110. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fu G, Brkić J, Hayder H and Peng C:
MicroRNAs in human placental development and pregnancy
complications. Int J Mol Sci. 14:5519–5544. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ouyang Y, Mouillet JF, Coyne CB and
Sadovsky Y: Review: Placenta-specific microRNAs in exosomes-good
things come in nano-packages. Placenta. 35(Suppl): S69–S73. 2014.
View Article : Google Scholar
|
|
13
|
Hromadnikova I, Kotlabova K, Ondrackova M,
Pirkova P, Kestlerova A, Novotna V, Hympanova L and Krofta L:
Expression profile of C19MC microRNAs in placental tissue in
pregnancy-related complications. DNA Cell Biol. 34:437–457. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu Y, Tian F, Li H, Zhou Y, Lu J and Ge
Q: Profiling maternal plasma microRNA expression in early pregnancy
to predict gestational diabetes mellitus. Int J Gynecol Obstet.
130:49–53. 2015. View Article : Google Scholar
|
|
15
|
Salomon C, Torres MJ, Kobayashi M,
Scholz-Romero K, Sobrevia L, Dobierzewska A, Illanes SE, Mitchell
MD and Rice GE: A gestational profile of placental exosomes in
maternal plasma and their effects on endothelial cell migration.
PLoS One. 9:e986672014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Salomon C, Scholz-Romero K, Sarker S,
Sweeney E, Kobayashi M, Correa P, Longo S, Duncombe G, Mitchell MD,
Rice GE and Illanes SE: Gestational diabetes mellitus is associated
with changes in the concentration and bioactivity of
placenta-derived exosomes in maternal circulation across gestation.
Diabetes. 65:598–609. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Diagnostic Criteria and Classification of
Hyperglycaemia First Detected in Pregnancy: A world health
organization guideline. Diabetes Res Clin Pract. 103:341–363. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gonzales PA, Zhou H, Pisitkun T, Wang NS,
Star RA, Knepper MA and Yuen PS: Isolation and purification of
exosomes in urine. Methods Mol Biol. 641:89–99. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Abdalla M: Comprehensive coverage of
exosomes purification and exosomal RNA isolation from different
types of liquid biopsies. https://www.exosome-rna.com/upcoming-webinar-comprehensive-coverage-of-exosome-purification-and-exosomal-rna-isolation-from-different-types-of-liquid-biopsies/.
Accessed April 15, 2019.
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
21
|
Kwon DN, Chang BS and Kim JH: MicroRNA
dysregulation in liver and pancreas of CMP-Neu5Ac hydroxylase null
mice disrupts insulin/PI3K-AKT signaling. Biomed Res Int.
2014:2363852014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Spinetti G, Fortunato O, Caporali A,
Shantikumar S, Marchetti M, Meloni M, Descamps B, Floris I,
Sangalli E, Vono R, et al: MicroRNA-15a and MicroRNA-16 Impair
human circulating proangiogenic cell functions and are increased in
the proangiogenic cells and serum of patients with critical limb
ischemia. Circ Res. 112:335–346. 2013. View Article : Google Scholar :
|
|
23
|
Kaddar T, Rouault JP, Chien WW, Chebel A,
Gadoux M, Salles G, Ffrench M and Magaud JP: Two new miR-16
targets: Caprin-1 and HMGA1, proteins implicated in cell
proliferation. Biol Cell. 101:511–524. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Palmieri D, D'Angelo D, Valentino T, De
Martino I, Ferraro A, Wierinckx A, Fedele M, Trouillas J and Fusco
A: Downregulation of HMGA-targeting microRNAs has a critical role
in human pituitary tumorigenesis. Oncogene. 31:3857–3865. 2012.
View Article : Google Scholar
|
|
25
|
Liu Q, Fu H, Sun F, Zhang H, Tie Y, Zhu J,
Xing R, Sun Z and Zheng X: MiR-16 family induces cell cycle arrest
by regulating multiple cell cycle genes. Nucleic Acids Res.
36:5391–5404. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yin Y, Stephen CW, Luciani MG and Fåhraeus
R: p53 stability and activity is regulated by Mdm2-mediated
induction of alter-native p53 translation products. Nat Cell Biol.
4:462–467. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Marcel V, Perrier S, Aoubala M, Ageorges
S, Groves MJ, Diot A, Fernandes K, Tauro S and Bourdon JC: Δ160p53
is a novel N-terminal p53 isoform encoded by Δ133p53 transcript.
FEBS Lett. 584:4463–4468. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu Y, Xia Y, Niu H and Chen Y: MiR-16
induced the suppression of cell apoptosis while promote
proliferation in esophageal squamous cell carcinoma. Cell Physiol
Biochem. 33:1340–1348. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Parsi S, Smith PY, Goupil C, Dorval V and
Hébert SS: Preclinical evaluation of miR-15/107 family members as
multifactorial drug targets for Alzheimer's disease. Mol Ther
Nucleic Acids. 4:e2562015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bandi N, Zbinden S, Gugger M, Arnold M,
Kocher V, Hasan L, Kappeler A, Brunner T and Vassella E: MiR-15a
and miR-16 are implicated in cell cycle regulation in a
Rb-Dependent manner and are frequently deleted or down-regulated in
non-small cell lung cancer. Cancer Res. 69:5553–5559. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bonci D, Coppola V, Musumeci M, Addario A,
Giuffrida R, Memeo L, D'Urso L, Pagliuca A, Biffoni M, Labbaye C,
et al: The miR-15a-miR-16-1 cluster controls prostate cancer by
targeting multiple oncogenic activities. Nat Med. 14:1271–1277.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lerner M, Harada M, Lovén J, Castro J,
Davis Z, Oscier D, Henriksson M, Sangfelt O, Grandér D and Corcoran
MM: DLEU2, frequently deleted in malignancy, functions as a
critical host gene of the cell cycle inhibitory microRNAs miR-15a
and miR-16-1. Exp Cell Res. 315:2941–2952. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sebastiani G, Guarino E, Grieco GE,
Formichi C, Poggi CD, Ceccarelli E and Dotta F: Circulating
microRNA (miRNA) expression profiling in plasma of patients with
gestational diabetes mellitus reveals upregulation of miRNA
miR-330-3p. Front Endocrinol (Lausanne). 8:3452017. View Article : Google Scholar
|
|
34
|
Koralewska N, Ciechanowska K, Pokornowska
M, Figlerowicz M and Kurzyńska-Kokorniak A: Human ribonuclease
Dicer-structure and functions. Postepy Biochem. 65:173–182. 2019.In
Polish. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Doyle SL, Husebye H, Connolly DJ, Espevik
T, O'Neill LA and McGettrick AF: The GOLD domain-containing protein
TMED7 inhibits TLR4 signalling from the endosome upon LPS
stimu-lation. Nat Commun. 3:7072012. View Article : Google Scholar
|
|
36
|
Füllekrug J, Suganuma T, Tang BL, Hong W,
Storrie B and Nilsson T: Localization and recycling of gp27
(hp24gamma3): Complex formation with other p24 family members. Mol
Biol Cell. 10:1939–1955. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wisniewska M, Goettig P, Maskos K,
Belouski E, Winters D, Hecht R, Black R and Bode W: Structural
determinants of the ADAM inhibition by TIMP-3: Crystal structure of
the TACE-N-TIMP-3 complex. J Mol Biol. 381:1307–1319. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kotronis K, Zafrakas M, Papasozomenou P,
Timologou A, Miliaras D, Tarlatzis BC and Grimbizis G: Protein
expression pattern of tissue inhibitor of metalloproteinase-3
(TIMP3) in endometriosis and normal endometrium. Gynecol
Endocrinol. 35:1103–1106. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Felicetti F, Errico MC, Bottero L,
Segnalini P, Stoppacciaro A, Biffoni M, Felli N, Mattia G, Petrini
M, Colombo MP, et al: The Promyelocytic leukemia zinc
Finger-MicroRNA-221/-222 pathway controls melanoma progression
through multiple oncogenic mechanisms. Cancer Res. 68:2745–2754.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Felicetti F, Errico MC, Segnalini P,
Mattia G and Carè A: MicroRNA-221 and -222 pathway controls
melanoma progression. Expert Rev Anticancer Ther. 8:1759–1765.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Garofalo M, Quintavalle C, Di Leva G,
Zanca C, Romano G, Taccioli C, Liu CG, Croce CM and Condorelli G:
MicroRNA signatures of TRAIL resistance in human non-small cell
lung cancer. Oncogene. 27:3845–3855. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
He H, Jazdzewski K, Li W, Liyanarachchi S,
Nagy R, Volinia S, Calin GA, Liu C G, Franssila K, Suster S, et al:
The role of microRNA genes in papillary thyroid carcinoma. Proc
Natl Acad Sci USA. 102:19075–19080. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Felli N, Fontana L, Pelosi E, Botta R,
Bonci D, Facchiano F, Liuzzi F, Lulli V, Morsilli O, Santoro S, et
al: MicroRNAs 221 and 222 inhibit normal erythropoiesis and
erythroleukemic cell growth via kit receptor down-modulation. Proc
Natl Acad Sci USA. 102:18081–18086. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gits CM, van Kuijk PF, Jonkers MB, Boersma
AW, van Ijcken WF, Wozniak A, Sciot R, Rutkowski P, Schöffski P,
Taguchi T, et al: MiR-17-92 and miR-221/222 cluster members target
KIT and ETV1 in human gastrointestinal stromal tumours. Br J
Cancer. 109:1625–1635. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hafner M, Landthaler M, Burger L, Khorshid
M, Hausser J, Berninger P, Rothballer A, Ascano M Jr, Jungkamp AC,
Munschauer M, et al: Transcriptome-wide identification of
RNA-Binding protein and MicroRNA target sites by PAR-CLIP. Cell.
141:129–141. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Elmehdawi F, Wheway G, Szymanska K, Adams
M, High AS, Johnson CA and Robinson PA: Human homolog of drosophila
ariadne (HHARI) is a marker of cellular proliferation associated
with nuclear bodies. Exp Cell Res. 319:161–172. 2013. View Article : Google Scholar
|
|
47
|
von Stechow L, Typas D, Carreras Puigvert
J, Oort L, Siddappa R, Pines A, Vrieling H, van de Water B,
Mullenders LH and Danen EH: The E3 Ubiquitin ligase ARIH1 protects
against genotoxic stress by initiating a 4EHP-Mediated mRNA
translation arrest. Mol Cell Biol. 35:1254–1268. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kishore S, Jaskiewicz L, Burger L, Hausser
J, Khorshid M and Zavolan M: A quantitative analysis of CLIP
methods for identifying binding sites of RNA-binding proteins. Nat
Methods. 8:559–564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kennedy SA, Jarboui MA, Srihari S, Raso C,
Bryan K, Dernayka L, Charitou T, Bernal-Llinares M,
Herrera-Montavez C, Krstic A, et al: Extensive rewiring of the EGFR
network in colorectal cancer cells expressing transforming levels
of KRASG13D. Nat Commun. 11:4992020. View Article : Google Scholar :
|
|
50
|
Shen N, Liu Y, Zhang K, Lyu Y, Gao M, Ma
J, Xu L and Gai Z: Analysis of RPS6KA3 gene mutation in a Chinese
pedigree affected with coffinlowry syndrome. Zhonghua Yi Xue Yi
Chuan Xue Za Zhi. 36:798–800. 2019.In Chinese. PubMed/NCBI
|
|
51
|
Zhang J, Yang M, Li D, Zhu S, Zou J, Xu S,
Wang Y, Shi J and Li Y: Homeobox C8 is a transcriptional repressor
of E-cadherin gene expression in non-small cell lung cancer. Int J
Biochem Cell Biol. 114:1055572019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Gong C, Zou J, Zhang M, Zhang J, Xu S, Zhu
S, Yang M, Li D, Wang Y, Shi J and Li Y: Upregulation of MGP by
HOXC8 promotes the proliferation, migration, and EMT processes of
triple-negative breast cancer. Mol Carcinog. 58:1863–1875. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Fang X and Yan R: MiR-152 inhibits the
proliferation and invasion of chordoma cells by targeting HOXC8. J
Int Med Res. 47:5185–5193. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Gottwein E, Corcoran DL, Mukherjee N,
Skalsky RL, Hafner M, Nusbaum JD, Shamulailatpam P, Love CL, Dave
SS, Tuschl T, et al: Viral MicroRNA targetome of KSHV-Infected
primary effusion lymphoma cell lines. Cell Host Microbe.
10:515–526. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene Ontology: Tool for the unification of biology. Nat
Genet. 25:25–29. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
56
|
The Gene Ontology Consortium: The gene
ontology resource: 20 years and still GOing strong. Nucleic Acids
Res. 47:D330–D338. 2019. View Article : Google Scholar :
|
|
57
|
Cheng X, Veverka V, Radhakrishnan A,
Waters LC, Muskett FW, Morgan SH, Huo J, Yu C, Evans EJ, Leslie AJ,
et al: Structure and interactions of the human programmed cell
death 1 receptor. J Biol Chem. 288:11771–11785. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
He PX, Ma ZL, Han H, Zhang XY, Niu SH, Du
LN, Zheng YC and Liu HM: Expression of programmed death ligand 1
(PD-L1) is associated with metastasis and differentiation in
gastric cancer. Life Sci. 242:1172472020. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Whisnant AW, Bogerd HP, Flores O, Ho P,
Powers JG, Sharova N, Stevenson M, Chen CH and Cullen BR: In-Depth
analysis of the interaction of HIV-1 with cellular microRNA
biogenesis and effector mechanisms. MBio. 4:e0001932013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Racapé M, Duong Van Huyen JP, Danger R,
Giral M, Bleicher F, Foucher Y, Pallier A, Pilet P, Tafelmeyer P,
Ashton-Chess J, et al: The involvement of SMILE/TMTC3 in
endoplasmic reticulum stress response. PLoS One. 6:e193212011.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Gan H, Lin L, Hu N, Yang Y, Gao Y, Pei Y,
Chen K and Sun B: KIF2C exerts an oncogenic role in nonsmall cell
lung cancer and is negatively regulated by miR-325-3p. Cell Biochem
Funct. 37:424–431. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
McHugh T, Zou J, Volkov VA, Aurélie Bertin
A, Talapatra SK, Rappsilber J, Dogterom M and Welburn JPI: The
depolymerase activity of MCAK shows a graded response to Aurora B
kinase phosphorylation through allosteric regulation. J Cell Sci.
132:jcs2283532019. View Article : Google Scholar :
|
|
63
|
Nečasová I, Janoušková E, Klumpler T and
Hofr C: Basic domain of telomere guardian TRF2 reduces D-loop
unwinding whereas Rap1 restores it. Nucleic Acids Res.
45:12170–12180. 2017. View Article : Google Scholar
|
|
64
|
Karginov FV and Hannon GJ: Remodeling of
Ago2-mRNA interactions upon cellular stress reflects miRNA
complementarity and correlates with altered translation rates.
Genes Dev. 27:1624–1632. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Moldovan L, Batte KE, Trgovcich J, Wisler
J, Marsh CB and Piper M: Methodological challenges in utilizing
miRNAs as circulating biomarkers. Version 2 J Cell Mol Med.
18:371–390. 2014. View Article : Google Scholar
|
|
66
|
Munaut C, Tebache L, Blacher S, Noël A,
Nisolle M and Chantraine F: Dysregulated circulating miRNAs in
preeclampsia. Biomed Rep. 5:686–692. 2016. View Article : Google Scholar
|
|
67
|
Li D and Li J: Association of
miR-34a3p/5p, miR-1413p/5p, and miR-24 in decidual natural killer
cells with unexplained recurrent spontaneous abortion. Med Sci
Monit. 22:922–929. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Song GY, Song WW, Han Y, Wang D and Na Q:
Characterization of the role of microRNA-517a expression in low
birth weight infants. J Dev Orig Health Dis. 4:522–526. 2013.
View Article : Google Scholar
|
|
69
|
Li J, Chen L, Tang Q, Wu W, Gu H, Liu L,
Wu J, Jiang H, Ding H, Xia Y, et al: The role, mechanism and
potentially novel biomarker of microRNA-17-92 cluster in
macrosomia. Sci Rep. 5:172122015. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Cai M, Kolluru GK and Ahmed A: Small
molecule, big prospects: MicroRNA in pregnancy and its
complications. J Pregnancy. 2017:69727322017. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Cao YL, Jia YJ, Xing BH, Shi DD and Dong
XJ: Plasma microRNA-16-5p-17-5p and -20a-5p: Novel diagnostic
biomarkers for gestational diabetes mellitus. J Obstet Gynaecol
Res. 43:974–981. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Geng Y, Ju Y, Ren F, Qiu Y, Tomita Y,
Tomoeda M, Kishida M, Wang Y, Jin L, Su F, et al: Insulin receptor
substrate 1/2 (IRS1/2) regulates Wnt/β-catenin signaling through
blocking autophagic degradation of dishevelled2. J Biol Chem.
289:11230–11241. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Ortega FJ, Mercader JM, Moreno-Navarrete
JM, Rovira O, Guerra E, Esteve E, Xifra G, Martínez C, Ricart W,
Rieusset J, et al: Profiling of circulating MicroRNAs reveals
common micrornas linked to type 2 diabetes that change with insulin
sensitization. Diabetes Care. 37:1375–1383. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Demirsoy İH, Ertural DY, Balci Ş, Çınkır
Ü, Sezer K, Tamer L and Aras N: Profiles of circulating MiRNAs
following metformin treatment in patients with type 2 diabetes. J
Med Biochem. 37:499–506. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Collares CV, Evangelista AF, Xavier DJ,
Rassi DM, Arns T, Foss-Freitas MC, Foss MC, Puthier D,
Sakamoto-Hojo ET, Passos GA, et al: Identifying common and specific
microRNAs expressed in peripheral blood mononuclear cell of type 1,
type 2, and gestational diabetes mellitus patients. BMC Res Notes.
6:4912013. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Zhao C, Dong J, Jiang T, Shi Z, Yu B, Zhu
Y, Chen D, Xu J, Huo R, Dai J, et al: Early second-trimester serum
MiRNA profiling predicts gestational diabetes mellitus. PLoS One.
6:e239252011. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Pheiffer C, Dias S, Rheeder P and Adam S:
Decreased expression of circulating miR-20a-5p in South African
women with gestational diabetes mellitus. Mol Diagn Ther.
22:345–352. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Shi Z, Zhao C, Guo X, Ding H, Cui Y, Shen
R and Liu J: Differential expression of microRNAs in omental
adipose tissue from gestational diabetes mellitus subjects reveals
miR-222 as a regulator of Erα expression in estrogen-induced
insulin resistance. Endocrinology. 155:1982–1990. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
López-Hernández Y, Herrera-Van Oostdam A,
Toro-Ortiz JC, López JA, Salgado-Bustamante M, Murgu M and
Torres-Torres LM: Urinary metabolites altered during the third
trimester in pregnancies complicated by gestational diabetes
mellitus: Relationship with potential upcoming metabolic disorders.
Int J Mol Sci. 20:11862019. View Article : Google Scholar :
|
|
80
|
Ibarra A, Vega-Guedes B, Brito-Casillas Y
and Wägner AM: Diabetes in pregnancy and MicroRNAs: Promises and
limitations in their clinical application. Noncoding RNA.
4:322018.
|
|
81
|
Sadovsky Y, Mouillet JF, Ouyang Y, Bayer A
and Coyne CB: The function of trophomirs and other micrornas in the
human placenta. Cold Spring Harb Perspect Med. 5:a0230362015.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Hromadnikova I, Kotlabova K, Ivankova K
and Krofta L: First trimester screening of circulating C19MC
microRNAs and the evaluation of their potential to predict the
onset of preeclampsia and IUGR. PLoS One. 12:e01717562017.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Hromadnikova I, Dvorakova L, Kotlabova K
and Krofta L: The prediction of gestational hypertension,
preeclampsia and fetal growth restriction via the first trimester
screening of plasma exosomal C19MC microRNAs. Int J Mol Sci.
20:29722019. View Article : Google Scholar :
|
|
84
|
Esteves JV, Enguita FJ and Machado UF:
MicroRNAs-Mediated regulation of skeletal muscle GLUT4 expression
and translocation in insulin resistance. J Diabetes Res.
2017:72679102017. View Article : Google Scholar : PubMed/NCBI
|