Introduction
Human adipose-derived mesenchymal stem cells
(hADSCs) are a type of adult mesenchymal stem cells (MSCs) that
have the plasticity to differentiate into osteoblasts, chondrocytes
and adipocytes in response to the appropriate conditions (1-3).
In recent studies, hADSCs have been successfully induced into an
osteogenic lineage in vitro and have been used as seed cells
to effectively repair bone defects (4,5).
However, the molecular mechanisms governing the osteogenic
differentiation of hADSCs are not fully elucidated, and thus
investigating the potential mechanisms is of great importance.
MicroRNAs (miRNAs) are small (18-25 nucleotides in
length), single-stranded noncoding RNAs that mediate gene
suppression by binding to the 3′-untranslated region (3′UTR) of
target mRNAs by promoting degradation or inhibiting the translation
of target mRNAs (6,7). Recently, several studies have
revealed that miRNAs serve critical roles in the regulation of MSC
osteogenic differentiation. For instance, miR-145 was decreased
during osteogenic differentiation of C2C12 and MC3T3-E1 cells, and
may suppress their osteogenic differentiation potential by
targeting Sp7 (8). Wang et
al (9) further revealed that
miR-193a served a suppressive role in the osteogenic
differentiation of human bone marrow-derived stromal cells (hBMSCs)
via targeting high mobility group box 1 (HMGB1). In addition, Li
et al have indicated that miR-23a suppressed the osteogenic
differentiation of hBMSCs by possibly targeting low-density
lipoprotein receptor-related protein 5 (10). However, the roles of miRNAs in
regulating the osteogenic differentiation of hADSCs remain largely
unknown.
Osteogenic differentiation is a complex process
governed by the interplay of multiple signaling pathways, such as
bone morphogenetic protein (11),
Wnt (12) and mitogen-activated
protein kinase (MAPK) signaling pathways (13,14). These pathways are often
constitutively activated during osteogenic differentiation of MSCs.
It has been reported that MAPK signaling components, including
extracellular-signal regulated kinase 1/2 (ERK1/2), strongly
increased the expression of Runt-related transcription factor-2
(Runx2) protein, which is one of several key transcriptional
factors in osteogenesis (15).
Furthermore, several studies revealed that the ERK signaling
pathway is closely associated with the osteogenic differentiation
of rat and human MSCs (16,17). For example, Ye et al
(18) demonstrated that knockdown
of forkhead box protein A2 enhanced the osteogenic differentiation
of BMSCs partly via activation of the ERK signaling pathway. Wang
et al (19) further
reported that naringin, a traditional Chinese medicine, enhanced
the BMSC osteogenic differentiation through the activation of ERK
signaling. Each of these studies has led us to speculate that ERK
signaling may serve an important role in the differentiation of
hADSCs into an osteogenic lineage.
In the present study, the expression profiles of
miRNAs during osteogenic differentiation of hADSCs were analyzed
using miRNA microarray, and revealed that miR-143 was significantly
downregulated in this process. The study then investigated the
underlying mechanisms involved in the regulatory role of miR-143 on
hADSC osteogenic differentiation in order to identify a potential
molecular therapeutic strategy for bone regeneration.
Materials and methods
Cell culture
All protocols involving human subjects were approved
by the Ethics Committee of Minhang Hospital, Fudan University
(Shanghai, China). Adipose tissue specimens were obtained from five
healthy donors undergoing tumescence liposuction (age range, 32-53
years; median age, 41 years; 2 males and 3 females) who underwent
surgery at Minhang Hospital, Fudan University between April 2017
and April 2018. Clinical and biochemical examinations confirmed
that these subjects did not have acute inflammation, cancer,
endocrine diseases or infectious diseases. The inclusion criteria
were as follows: i) Patients who were willing to participate in the
study; and ii) clinical and biochemical examinations confirmed that
these subjects did not have acute inflammation, cancer, endocrine
diseases or infectious diseases. Patients who received chemotherapy
prior to the study were excluded from the present study. Written
informed consent for participation in the study was obtained from
all patients. The hADSCs were isolated from the adipose tissues
according to a previously described method (20).
Following isolation, hADSCs were cultured in basal
MSC culture medium (bM), containing Dulbecco's modified Eagle
medium (DMEM), 10% fetal calf serum, 1% antibiotics (100 U/ml
penicillin and 100 mg/ml streptomycin; Thermo Fisher Scientific,
Inc.) and 1% L-glutamine (200 mM; Lonza) at 37°C with 5%
CO2.
Adipogenic, osteogenic and chondrogenic
differentiation
For adipogenic differentiation, hADSCs
(3×105 cells/well) were seeded into 6-well culture
plates and cultured for 21 days with adipogenic differentiation
medium (Cyagen Bioscience, Inc.), and the medium was replaced every
3 days. Following the culture, the cells were washed with PBS and
then fixed for 30 min at room temperature with 4% paraformaldehyde
solution. Next, 0.6% Oil Red O (Sigma-Aldrich; Merck KGaA) solution
was used to stain the fixed cells for 1 h at room temperature, and
the cells were then observed using a phase contrast microscope
(Motic).
For osteogenic differentiation, hADSCs
(3×105 cells/well) were seeded into 6-well culture
plates containing 3 ml bM. On the following day, the osteogenic
differentiation was initiated by replacing the bM with osteogenic
differentiation medium (odM), which consisted of DMEM, 100 nM
dexamethasone, L-glutamine, 50 mM ascorbate, 1% antibiotics,
mesangial cell growth supplement and 10 mM β-glycerophosphate
(Lonza). The medium was replaced with fresh odM every 2-3 days
thereafter. After the cells were cultured with odM for a maximum of
21 days, Alizarin Red S staining was performed to assess the
osteogenic differentiation potential of hADSCs. For this, the cells
were fixed in 4% paraformaldehyde solution for 15 min, followed by
staining with 2% Alizarin Red S (Sigma-Aldrich; Merck KGaA)
solution for 40 min at room temperature. Images were captured under
a light microscope (DM-1000 Microscope; Leica Microsystems).
Subsequently, the stained cells were incubated with 10% acetic acid
for 30 min and collected with a cell scraper and vortexed. Samples
were then heated at 85°C for 10 min, cooled down, and centrifuged
at 20,000 × g for 15 min. The supernatant was neutralized with 10%
ammonium hydroxide. The absorbance was measured at 405 nm as
previously described (21).
For the differentiation of hADSCs into chondrocytes,
an MSC Chondrogenic Differentiation kit (Cyagen Bioscience, Inc.)
was used according to the manufacturer's protocol. Next, Alcian
Blue staining was performed to assess the chondrogenic
differentiation potential of hADSCs. Briefly, the cells were fixed
with 4% paraformaldehyde solution for 30 min, washed three times
with PBS, stained with 1% Alcian Blue (Solarbio Science &
Technology Co., Ltd.) solution, and finally observed under a
microscope (Olympus Corp.).
Alkaline phosphatase (ALP) activity
ALP activity was determined using an Alkaline
Phosphatase Diethanolamine Activity Detection kit (cat. no.
AP0100-1KT; Sigma-Aldrich; Merck KGaA) according to the
manufacturer's protocol.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total cellular RNAs were isolated 7, 14 and 21 days
after osteogenic differentiation using TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. For miR-143 detection, RT was performed using the
miScript II RT kit (Qiagen), while for the detection of Runx2, bone
sialoprotein (BSP), osteopontin (OPN), osteocalcin (OCN) and k-Ras
mRNA levels, RT was performed using the High Capacity cDNA reverse
transcription kit (Invitrogen; Thermo Fisher Scientific, Inc.).
miR-143 were measured using the miScript SYBR® Green PCR
kit (Exiqon; Qiagen), while Runx2, OCN, BSP, OPN and k-Ras
expression levels were measured using a Power SYBR Green PCR Master
mix (Roche Diagnostics), on a Light Cycler instrument (Bio-Rad
Laboratories, Inc.). The primers for qPCR analysis were as follows:
miR-143 forward, 5′-ACA CTC CAG CTG GGG GTG CAG TGC TGC ATC -3′,
and reverse, 5′-CTC AAC TGG TGT CGT GGA GTC GGC AAT TCA GTT GAG ACC
AGA -3′; U6 forward, 5′-GCT TCG GCA GCA CAT ATA CTA AAA T-3′, and
reverse, 5′-CGC TTC ACG AAT TTG CGT GTC AT-3′; Runx2 forward,
5′-GTC TCA CTG CCT CTC ACT TG-3′, and reverse, 5′-CAC ACA TCT CCT
CCC TTC TG-3′; OCN forward, 5′-ACA GAC AAG TCC CAC ACA GCA GC-3′,
and reverse, 5′-TGA AGG CTT TGT CAG ACT CAG GGC-3′; BSP forward,
5′-GCC AGA GGA GCA ATC ACC AA-3′, and reverse, 5′-CAG GCT GGA GGT
TCA CTG GT-3′; OPN forward, 5′-TTG GCT TTG CAG TCT CCT GCG G-3′,
and reverse, 5′-AGG CAA GGC CGA ACA GGC AAA-3′; k-Ras forward,
5′-ACT GAA TAT AAA CCT TGT GGT AG-3′, and reverse, 5′-TCA AAG AAT
GGT CCT GGA CC-3′; GAPDH forward, 5′-CGA GCC ACA TCG CTC AGA CA-3′,
and reverse, 5′-GTC TCA CTG CCT CTC ACT TG-3′. The thermocycling
were as follows: 95°C for 1 min; and 40 cycles of 95°C for 30 sec,
58°C for 30 sec and 68°C for 3 min/kb; followed by 68°C for 10 min.
The relative expression of each gene was calculated using the
2−∆∆Cq method (22).
Microarray assay
Total cellular RNA were extracted 14 days after
osteoinduction using miRNeasy mini kit (Qiagen). Next, the samples
were assessed using the miRCURY LNA™ Array kit, version 18.0
(Exiqon; Qiagen). The procedure and imaging processes were
performed as previously described (23).
Cell transfection
When hADSCs in a 6-well plate were grown to ~80%
confluence, miR-143 mimics, mimics negative control (NC), miR-143
inhibitor and inhibitor NC (Shanghai GenePharma Co., Ltd.) were
transfected into the cells using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol.
In addition, transfection of cultured hADSCs with
agomir-143, agomir NC, antagomir-143 and antagomir-NC (Shanghai
GenePharma Co., Ltd.) was performed at a final concentration of 100
nM in bM. The medium was replaced with osteogenic medium after 16
h, and hADSCs were continuously cultured in odM for 14 days and
then cells were harvested for RT-qPCR and western blot
analyses.
MEK/ERK inhibition
To block the activity of the ERK1/2 signaling
pathway, hADSCs were treated with the pathway inhibitors U0126 (10
µmol/l; cat no. 662009; Sigma-Aldrich; Merck KGaA) or
PD98059 (10 µmol/l; cat no. P215; Sigma-Aldrich; Merck KGaA)
at 37°C for 15 min.
Dual-luciferase reporter assay
miRNA target prediction tools, including Miranda
(http://miranda.org.uk) and TargetScan v7.0
(http://targetscan.org/) were used to search for
the putative targets of miR-143. Dual-Luciferase Reporter Assay
system (Promega Corporation) was used to analyze the double
luciferase activities according to the manufacturer's protocol. In
brief, a sequence containing miR-143-predicted target within the
k-Ras 3′ untranslated region (UTR) or a mutant sequence lacking any
complementarity with miR-143 seed sequence were cloned in the 3′
UTR of the luciferase gene, generating the reporter vector
wild-type-pGL3-k-Ras or mut-type-pGL3-k-Ras, respectively. When
hADSCs grew to 60-70% confluence in 12-well plates, the cells were
co-transfected with 50 nM miR-143 mimics, miR-143 inhibitor and 2
µg luciferase reporter plasmids using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). Luciferase activity
was determined 48 h after transfection. Normalization of firefly
luciferase activity to Renilla luciferase activity was
performed.
Western blot analysis
Western blot analysis was performed as previously
described (19). Briefly, cells
were lysed with RIPA buffer (cat. no. P0013C; Beyotime Institute of
Biotechnology), and the total protein concentration was determined
with using a bicinchoninic acid assay (cat. no. P0012; Beyotime
Institute of Biotechnology). Subsequently, 40 µg protein
samples were separated by 12% SDS-PAGE, and then transferred onto a
polyvinylidene difluoride (Millipore) membrane, and this membrane
was then blocked with 5% skim milk for 2 h at 4°C overnight. Next,
the samples were probed with primary antibodies at 37°C for 2 h as
follows: k-Ras (1:1,000; ab180772), p-p38 (1:1,000; ab31828) and
c-Raf/1 (1:1,000; cat. no. ab137435), purchased from Abcam;
p-c-Raf/1 (1:1,000; cat. no. sc-81513), obtained from Santa Cruz
Biotechnology, Inc., mitogen-activated protein kinase 1/2 (MEK1/2;
1:1,000; cat. no. 9122), p-MEK1/2 (1:1,000; cat. no. 9121), ERK1/2
(1:2,000; cat. no. 4695), p-ERK1/2 (1:2,000; cat. no. 4370), c-Jun
N-terminal kinase (JNK; 1:1,000; cat. no. 9258), p-JNK (1:1,000;
cat. no. 4671) and p38 (1:2,000; cat. no. 9212), purchased from
Cell Signaling Technology, Inc.; and β-actin (1:1,000; bs-0061R)
from Beijing Biosynthesis Biotechnology Co., Ltd. Membranes were
subsequently incubated with horseradish peroxidase-conjugated
anti-rabbit IgG secondary antibody (1:2,000; cat. no. 7074; Cell
Signaling Technology, Inc.) for 1 h at room temperature. Protein
bands were visual-ized using an enhanced chemiluminescence
detection system (GE Healthcare Life Sciences), and the blot bands
were then quantified with ImageJ software (version 1.46; National
Institutes of Health).
Statistical analysis
The difference between means was analyzed using
Student's t-test and analysis of variance using SPSS software
package, version 13.0 (SPSS, Inc.). All data are showed as the mean
± standard deviation (SD). A P-value of <0.05 was considered to
denote a statistically significant difference.
Results
Aberrant expression of miRNAs during
osteogenic differentiation
After hADSCs were isolated, the cells were cultured
to induce adipogenic, osteogenic and chondrogenic differentiation
to assess their multilineage differentiation potential. As shown in
Fig. 1A, hADSCs were able to
differentiate into adipocytes (Oil Red O staining), osteogenic
cells (Alizarin Red S staining) and chondrocytes (Alcian Blue
staining), which suggest that the isolated hADSCs exhibit the
typical characteristics of MSCs. Next, Alizarin Red S staining was
conducted to observe the changes of the extracellular matrix
mineralization and the results showed that extracellular matrix
mineralization was significantly increased under osteogenic
differentiation conditions in a time-dependent manner (Fig. 1B). In addition, ALP activity was
further measured by ALP assay. As shown in Fig. 1C, ALP activity increased with a
peak at 14 days and then decreased, typical of the well-established
program of osteoblast differentiation.
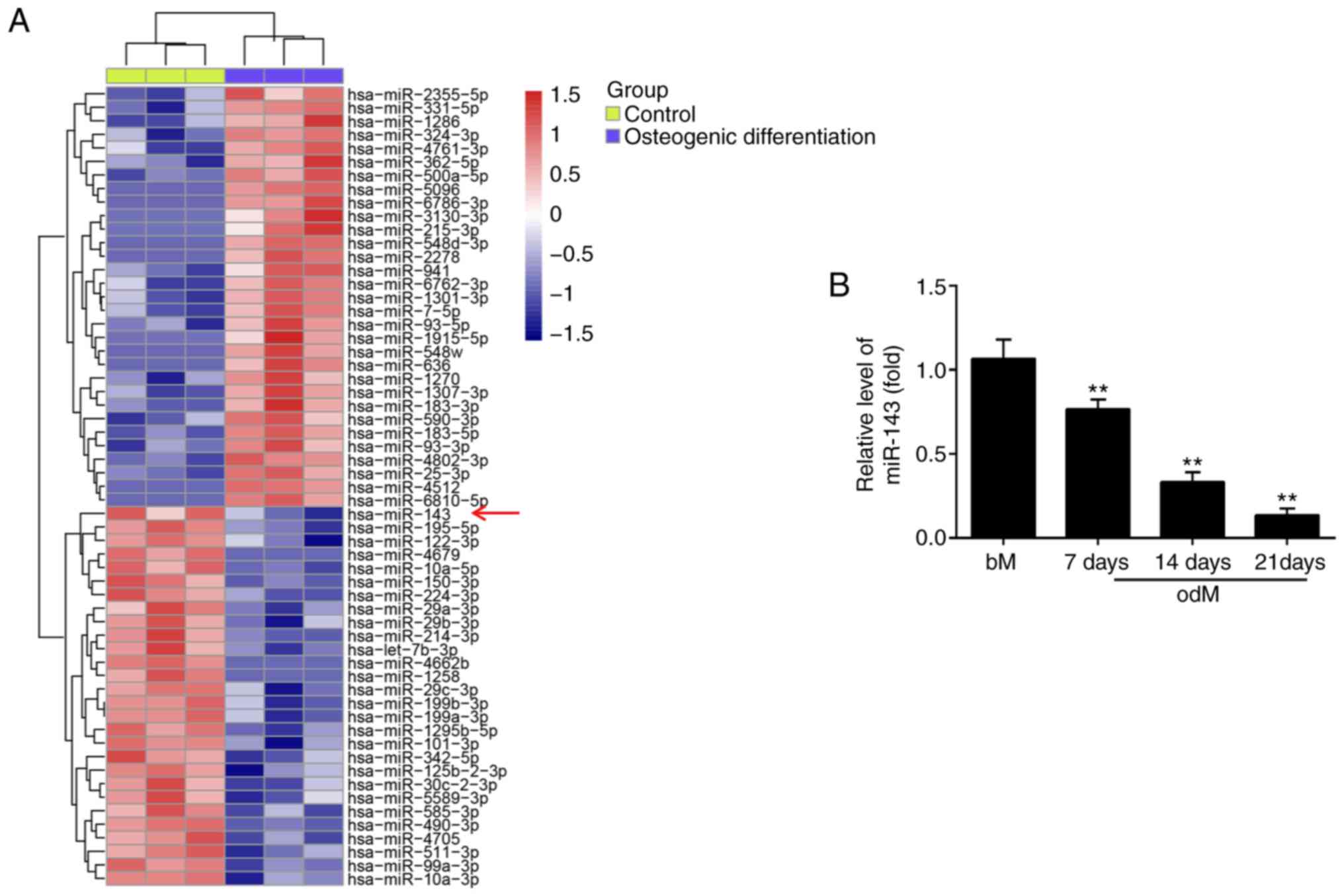
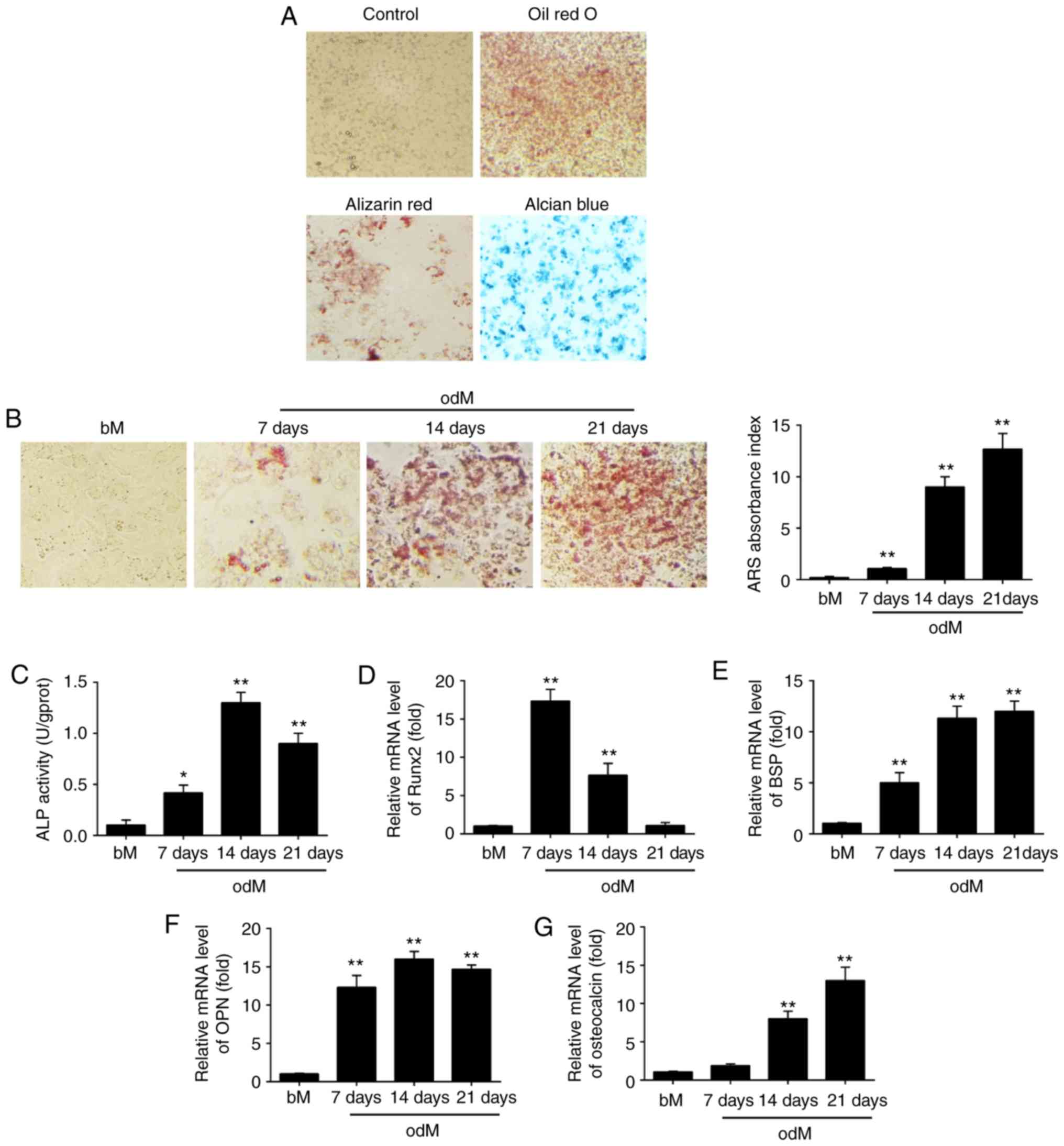 | Figure 1miR-143 was downregulated during
hADSC differentiation. (A) hADSCs were seeded in 6-well plates,
incubated in normal, adipogenic, osteogenic or chondrogenic
differentiation medium for 21 days, and then stained with Oil Red
O, Alizarin Red S or Alcian Blue, respectively. Representative
images are shown. The control group was stained by Alizarin Red S.
Magnification, ×200. (B) Alizarin Red S staining to examine the
mineralization of hADSCs cultured in odM or bM at 7, 14 and 21
days. A representative example of three independent experiments is
shown. Magnification, ×200. (C) ALP activity was assessed by
colorimetric assay. (D) Runx2, (E) BSP, (F) OPN and (G) OCN mRNA
levels were assessed by RT-qPCR analysis on days 7, 14 and 21
post-induction of osteogenic differentiation. Data are represented
as the mean ± standard deviation of three independent experiments.
*P<0.05, **P<0.01 vs. bM group. miR,
microRNA; hADSCs, human adipose-derived mesenchymal stem cells;
odM, osteogenic differentiation medium; bM, basal medium; ALP,
alkaline phosphatase; Runx2, Runt-related transcription factor-2;
BSP, bone sialoprotein; OPN, osteopontin; OCN, osteocalcin;
RT-qPCR, reverse transcription-quantitative polymerase chain
reaction. |
Multiple transcription factors drive the control of
osteogenesis, such as Runx2, BSP, OPN and OCN. Runx2 is a key
transcriptional regulator that promotes differentiation by inducing
the expression of osteo-specific genes during the early stages of
differentiation (24), whereas
OCN is one of the late-onset genes associated with osteogenesis
(25). BSP and OPN are
non-collagenous phosphoproteins proven to be involved in the
mineralization of bone (26). In
the present study, RT-qPCR was subsequently used to analyze the
expression levels of these osteogenic specific genes. The results
demonstrated that the expression of Runx2 was time-dependently
decreased under osteogenic differentiation conditions, while the
expression levels of BSP, OPN and OCN were significantly increased
(Fig. 1D-G). These data suggested
that osteogenic differentiation of hADSCs was successfully induced
using an osteogenic induction medium.
miR-143 expression was downregulated
during the osteogenic differentiation of hADSCs
To screen the expression patterns of miRNAs involved
in the osteogenic differentiation of hADSCs, a microarray analysis
was performed. Compared with the control group, 31 miRNAs were
upregulated and 28 miRNAs were downregulated following osteogenic
differentiation (Fig. 2A). Among
the aberrantly expressed miRNAs, miR-143 was selected for further
investigation in the present study, since its expression level was
the most significantly downregulated in the osteogenic
differentiation group. Previous studies have reported the
suppressive functions of miR-143 in different MSCs (18,27); however, whether miR-143 has a
similar role in the osteogenic differentiation of hADSCs remains
unknown. To further verify the microarray analysis results on
miR-143 expression, the miR-143 levels were detected at different
time points during the osteogenesis process. As shown in Fig. 2B, miR-143 expression was
significantly decreased during this process in a time-dependent
manner, compared with the control group. These results implied that
miR-143 may be involved in the osteogenic differentiation of
hADSCs.
miR-143 negatively regulates the
osteogenic differentiation of hADSCs
To investigate the role of miR-143 in the osteogenic
differentiation of hADSCs, the cells were transfected with
agomir-143 and antagomir-143, and the efficiency of transfection
was determined by qPCR on day 21 of osteogenic differentiation of
hADSCs. The results of RT-qPCR (Fig.
3A) revealed that miR-143 was notably upregulated and
downregulated following agomir-143 and antagomir-143 transfection,
respectively. Subsequently, Alizarin Red S and ALP staining were
performed to evaluate the effects of miR-143 on calcium deposits
and ALP activity during the osteogenic differentiation of hADSCs.
The results demonstrated that agomir-143 treatment led to a
significant decrease in both calcium deposits and ALP activity
compared with those in the agomir-NC group, while antagomir-143
transfection markedly enhanced these processes (Fig. 3B and C). RT-qPCR was also
conducted to assess the mRNA expression levels of the bone markers
Runx2, OPN, BSP and OCN. The results demonstrated that agomir-143
significantly reduced the expression levels of these markers,
whereas antagomir-143 significantly promoted these levels compared
with those in the corresponding NC groups (Fig. 3D-G). Collectively, these data
suggested that miR-143 was a negative regulator of hADSC
osteogenesis.
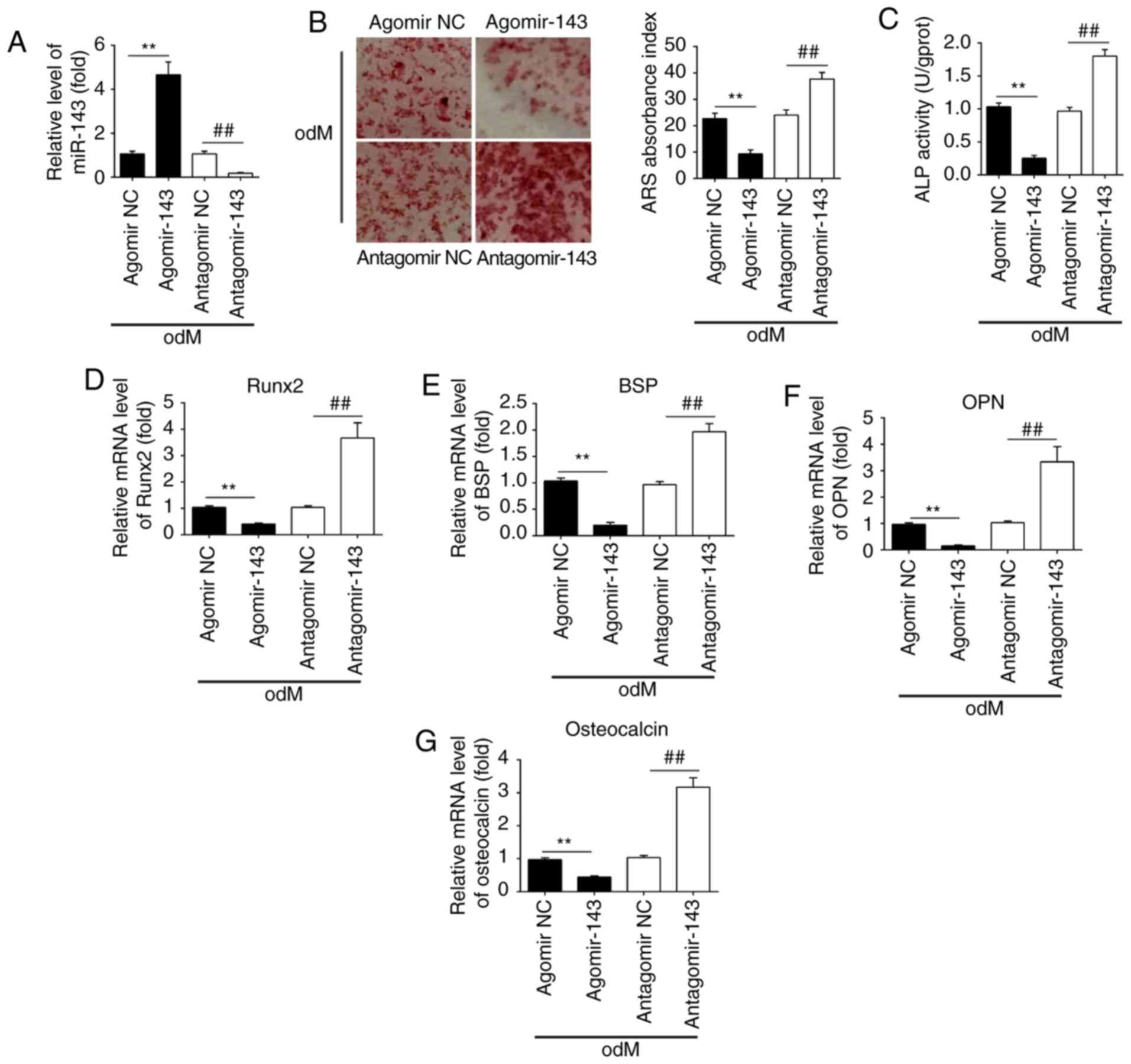 | Figure 3miR-143 negatively regulated the
osteogenic differentiation of hADSCs. hADSCs were transfected with
agomir-143, antagomir-143 or the corresponding NC (100 nM) for 16
h. The medium was then replaced with odM, and hADSCs were
continuously cultured for 14 days. (A) miR-143 expression was
assessed by RT-qPCR. (B) Osteogenic differentiation was determined
by Alizarin Red S staining and (C) ALP activity was assessed by
colorimetric assay on day 14 post-induction. Magnification, ×200.
(D) Runx2, (E) BSP, (F) OPN and (G) osteocalcin mRNA levels were
assessed by RT-qPCR analysis on day 14 post-induction. Data
represent the mean ± standard deviation of three independent
experiments. **P<0.01 and ##P<0.01.
miR, microRNA; hADSCs, human adipose-derived mesenchymal stem
cells; NC, negative control; odM, osteogenic differentiation
medium; ALP, alkaline phosphatase; Runx2, Runt-related
transcription factor-2; BSP, bone sialoprotein; OPN, osteopontin;
RT-qPCR, reverse transcription-quantitative polymerase chain
reaction. |
miR-143 negatively regulates the ERK1/2
signaling pathway during the osteogenic differentiation of
hADSCs
Previous studies have reported that MAPKs, including
ERK, JNK and p38-MAPK, are important signal transducers during
osteogenic differentiation of MSCs (28-30). Therefore, the present study
attempted to investigate whether miR-143 can influence the MAPK
(ERK1/2, JNK, and p38) signaling pathways during hADSC osteogenic
differentiation. Consistent with the findings of previous studies
(31,32), the levels of p-ERK1/2, p-JNK and
p-p38 were significantly increased during osteogenic
differentiation compared with undifferentiated control groups
(Fig. 4A and B). Notably,
agomir-143 and antagomir-143 transfection resulted in a decrease
and increase, respectively, only in the expression of p-ERK1/2,
whereas the expression levels of p-JNK or p-p38 were not markedly
affected (Fig. 4A and B). Taken
together, these results suggested that miR-143 was a negative
regulator of the ERK1/2 signaling pathway during the osteogenic
differentiation of hADSCs.
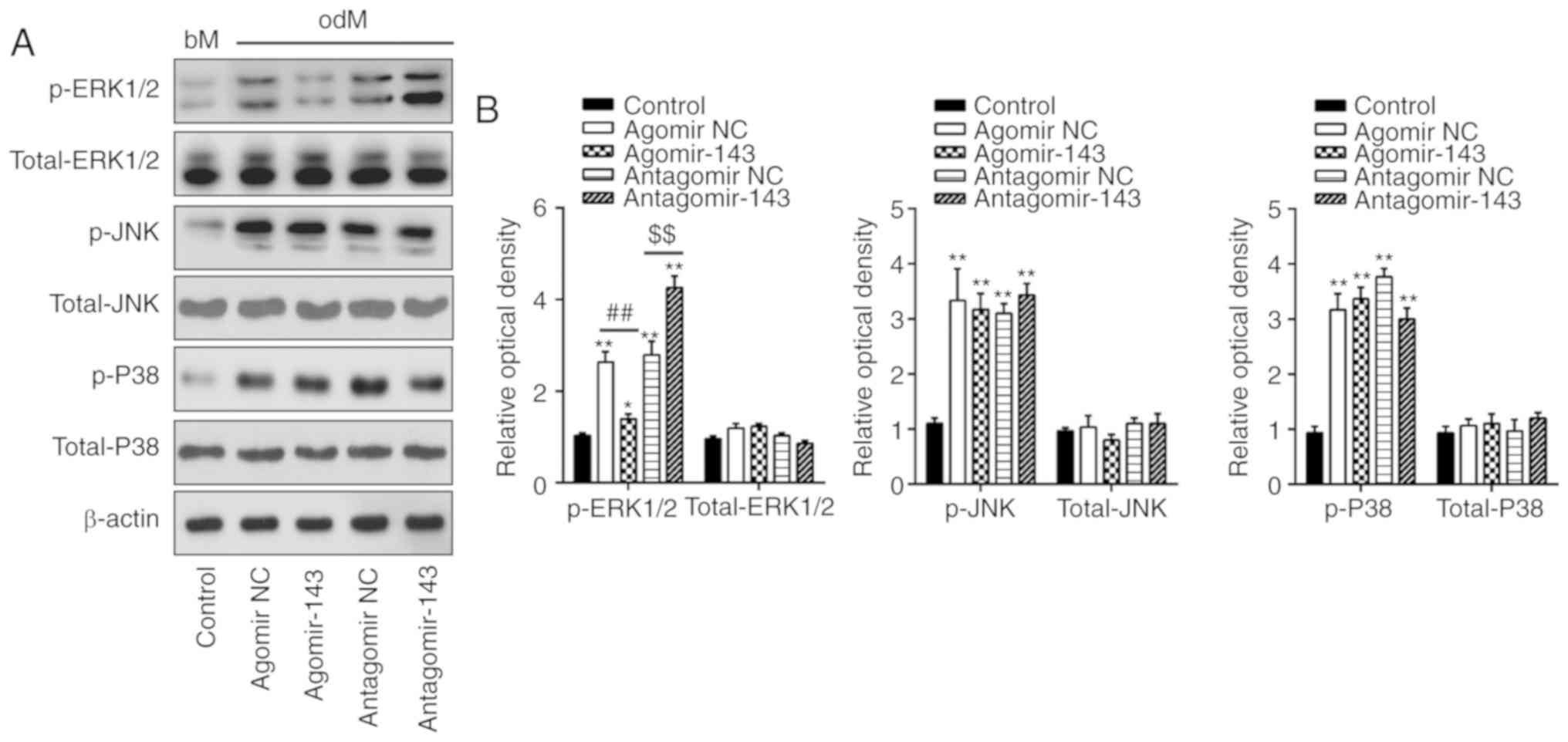 | Figure 4miR-143 negatively regulated the
ERK1/2 signaling pathway. hADSCs were transfected with agomir-143,
antagomir-143 or the corresponding NC (100 nM) for 16 h. The medium
was replaced with osteogenic medium, and hADSCs were continuously
cultured for 14 days. (A) Western blot analysis was used to assess
the protein expression levels of p-ERK1/2, ERK1/2, p-JNK, JNK,
p-P38 and P38, with β-actin used as an internal control. (B) The
bands were semi-quantitatively analyzed using ImageJ software and
normalized to β-actin density. Data represent the mean ± standard
deviation of three independent experiments. *P<0.05
and **P<0.01, vs. control group;
##P<0.01 and $$P<0.01. miR, microRNA;
hADSCs, human adipose-derived mesenchymal stem cells; NC, negative
control; ERK1/2, extracellular-signal regulated kinase 1/2; JNK,
c-Jun N-terminal kinase. |
miR-143 downregulation enhances
osteogenic differentiation of hADSCs through activation of the
ERK1/2 signaling pathway
To investigate whether the ERK1/2 signaling pathway
is critically involved in the positive effect of miR-143 on the
osteogenic differentiation of hADSCs, the activity of the ERK1/2
signaling pathway was blocked using the pathway inhibitors U0126
and PD98059 (33). The results
revealed that the level of p-ERK1/2 was markedly increased in the
odM + antagomir-143 group compared with that in odM group; however,
the protein levels of p-ERK1/2 were significantly decreased upon
U0126 or PD98059 treatment (Fig.
5A). Next, Alizarin Red S and ALP staining were conducted to
measure the calcium deposits and ALP activity, respectively. The
results indicated that both U0126 and PD98059 markedly reversed the
antagomir-143-induced increase in calcium deposits and ALP activity
(Fig. 5B and C). In addition, the
upregulation of Runx2, OPN, BSP and OCN levels induced by
antagomir-143 was also abrogated by U0126 and PD98059 treatment
(Fig. 5D-G). These results
indicated the significance of the ERK1/2 signaling pathway in the
osteogenic differentiation of hADSCs mediated by antagomir-143.
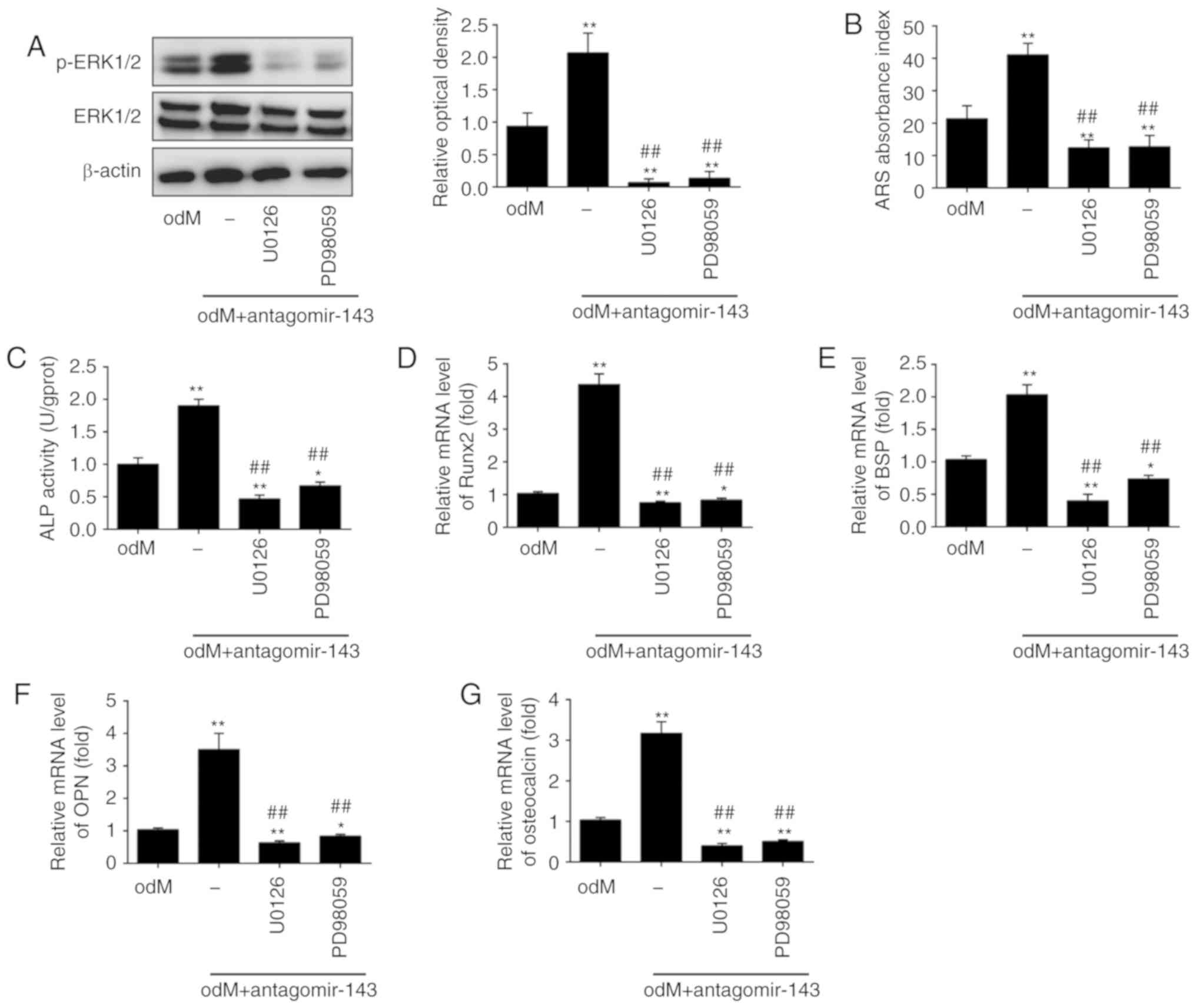 | Figure 5miR-143 downregulation enhanced the
osteogenic differentiation of human adipose-derived mesenchymal
stem cells through activation of the ERK1/2 signaling pathway.
Cells were treated with antagomir-143 or antagomir NC, with or
without 10 µmol/l U0126 or PD98059 for 15 min. (A) Protein
expression levels of p-ERK1/2 and ERK1/2, detected by western blot
analysis and quantified using ImageJ software. (B) Graphical
representation of the optical density following Alizarin Red S
staining at day 21 of culture. (C) ALP activity assessed by
colorimetric assay. (D) Runx2, (E) BSP, (F) OPN and (G)
osteo-calcin mRNA levels were assessed by reverse
transcription-quantitative polymerase chain reaction analysis. Data
represent the mean ± standard deviation of three independent
experiments. *P<0.05 and **P<0.01, vs.
odM group; ##P<0.01 vs. antagomir-143 + odM group.
miR, microRNA; ERK1/2, extracellular-signal regulated kinase 1/2;
NC, negative control; ALP, alkaline phosphatase; Runx2,
Runt-related transcription factor-2; BSP, bone sialoprotein; OPN,
osteopontin; odM, osteogenic differentiation medium. |
k-Ras is a direct target of miR-143
Through bioinformatics prediction using TargetScan
7.0 and miRanda, a putative target site of miR-143 was identified
in the 3′-UTR of k-Ras mRNA (Fig. 6A
and B). To further validate that miR-143 targeted k-Ras, a
dual-luciferase reporter assay was conducted. The results
demonstrated that miR-143 mimics markedly inhibited the luciferase
activity of the k-Ras-3′UTR WT reporter, whereas co-transfection
with the miR-143 inhibitor and WT reporter resulted in increased
luciferase activity; however, no evident changes were observed
following co-transfection of k-Ras 3′-UTR-Mut with miR-143 mimics
or inhibitor (Fig. 6C).
Furthermore, to determine whether miR-143 regulates k-Ras, the mRNA
and protein levels of k-Ras were measured by RT-qPCR and western
blot analysis, respectively. As shown in Fig. 6D and E, k-Ras expression was
markedly decreased following miR-143 mimics transfection, while its
expression was increased by miR-143 inhibitor in hADSCs at the mRNA
and protein levels. It was also observed that k-Ras levels were
markedly downregulated in hADSCs transfected with agomir-143 during
osteogenic differentiation, but upregulated in cells transfected
with antagomir-143 (Fig. 6F).
These data indicated that miR-143 directly targeted k-Ras during
the osteogenic differentiation of hADSCs.
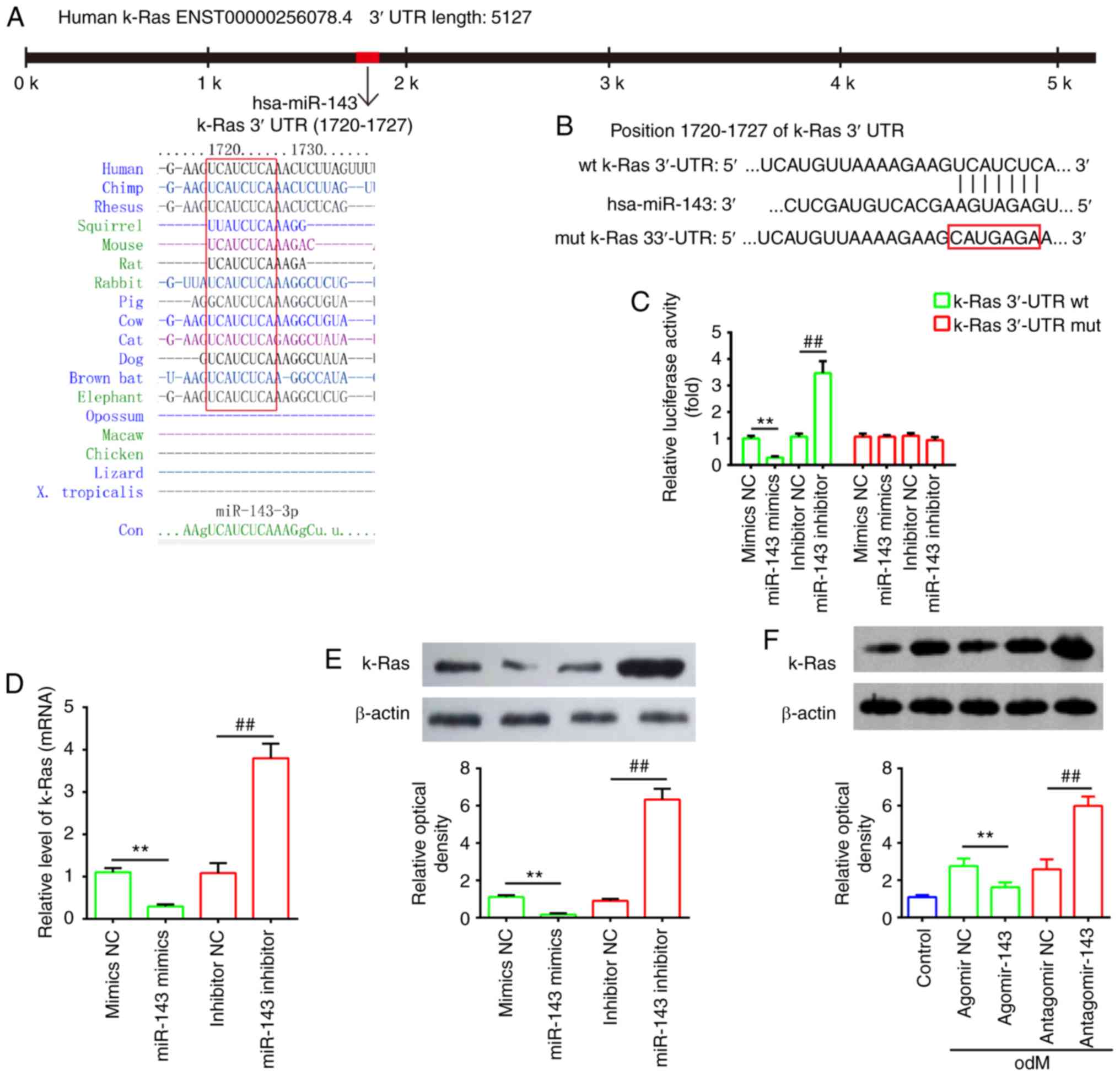 | Figure 6k-Ras was a direct target of miR-143.
(A) Sequences of miR-143 binding sites are highly conserved among
different species. (B) Putative binding site of miR-143 and k-Ras.
(C) Luciferase activity of hADSCs co-transfected with luciferase
reporter constructs containing WT or Mut k-Ras 3′-UTR, and with
miR-143 mimics, miR-143 inhibitor or NC (n=3). (D) Reverse
transcription-quantitative polymerase chain reaction and (E)
western blot analysis of mRNA and protein expression levels of
k-Ras in hADSCs transfected with miR-143 mimics, miR-143 inhibitor
or NC miRNA (n=3). (F) Western blot analysis of k-Ras protein
expression in hADSCs transfected with agomir-143, antagomir-143 and
the corresponding NC. The medium was replaced with odM after 16 h,
and hADSCs were continuously cultured for 14 days prior to protein
level assessment. Data represent the mean ± standard deviation of
three independent experiments. **P<0.01 and
##P<0.01. miR, microRNA; hADSCs, human
adipose-derived mesenchymal stem cells; WT, wild-type; Mut, mutant;
NC, negative control; odM, osteogenic differentiation medium. |
miR-143 negatively regulates the
k-Ras/MEK/ERK signaling pathway during osteogenic differentiation
of hADSCs
It has been reported that k-Ras functions as an
important activator of the ERK signaling pathway, which has been
implicated in the regulation of the osteogenic differentiation of
MSCs (34). Given this, in the
present study, it was speculated that miR-143 may modulate the ERK
signaling pathway via targeting k-Ras during osteogenic
differentiation of hADSCs. As shown in Fig. 7A and B, miR-143 overexpression
significantly decreased the expression levels of k-Ras, p-c-Raf/1,
p-MEK1/2 and p-ERK1/2, as compared with those in the agomir NC
group. By contrast, inhibition of miR-143 enhanced the expression
levels of these proteins when compared with those in antagomir-NC
group. These findings indicated that inhibition of miR-143 may
promote the osteogenic differentiation of hADSCs via activating the
k-Ras/MEK/ERK signaling pathway.
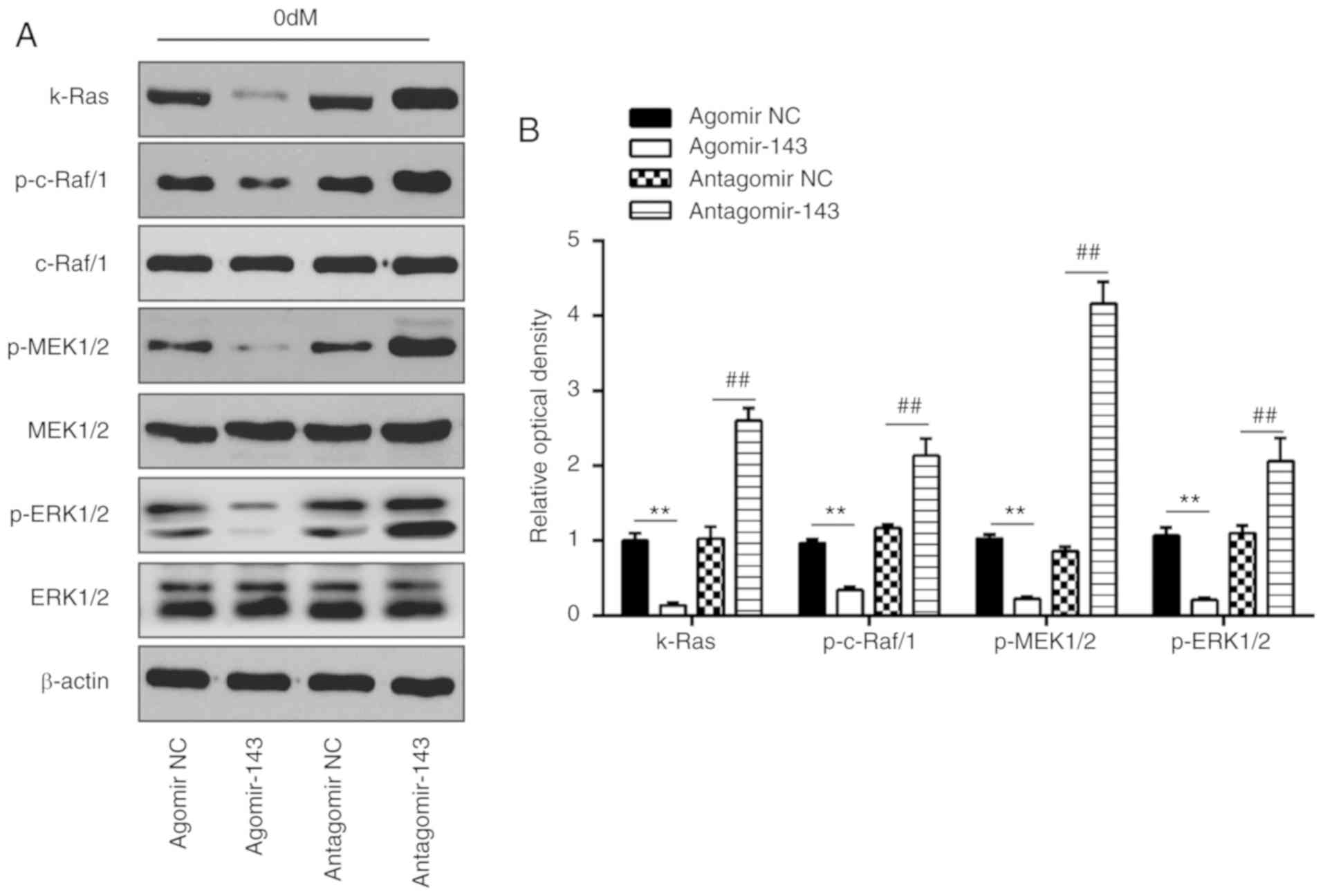 | Figure 7miR-143 negatively regulated
k-Ras/MEK/ERK signaling pathway during osteogenic differentiation
of hADSCs. hADSCs were transfected with agomir-143, antagomir-143
or the corresponding NC, and osteogenic differentiation medium was
added. (A) Western blot analysis was used to assess the protein
expression levels of k-Ras, p-Raf/1, Raf/1, p-MEK1/2, MEK1/2,
p-ERK1/2 and ERK1/2, with β-actin used as an internal control. (B)
The bands were semi-quantitatively analyzed using ImageJ software
and normalized to β-actin density. Data represent the mean ±
standard deviation of three independent experiments.
**P<0.01 and ##P<0.01. miR, microRNA;
hADSCs, human adipose-derived mesenchymal stem cells; MEK,
mitogen-activated protein kinase; ERK, extracellular-signal
regulated kinase; NC, negative control. |
Discussion
In the present study, the data revealed that miR-143
was down-regulated during the osteogenic differentiation of hADSCs.
Functional analyses indicated that miR-143 overexpression
suppressed the osteogenic potential of hADSCs, whereas miR-143
inhibition enhanced this potential. Notably, the current study data
revealed that miR-143 knockdown promoted the osteogenic
differentiation of these cells by activating the k-Ras/MEK/ERK
signaling pathway. These findings suggested that miR-143 serves a
potentially crucial role in the osteogenic differentiation of
hADSCs.
Mounting evidence has indicated the vital roles of
several other miRNAs in the osteogenic differentiation of hADSCs.
For instance, Kim et al (35) reported that miR-196a is a key
modulator of this process in hADSCs, and lentiviral overexpression
of this miRNA enhanced osteogenic differentiation by targeting
HOXC8. Luzi et al (36)
observed that miR-26a expression increased during hADSC
differentiation, while inhibition of miR-26a modulated late
osteogenic differentiation by targeting the SMAD1 transcription
factor. Zeng et al (37)
also identified miR-100 as a negative regulator of the osteogenic
differentiation of hADSCs, and overexpression of miR-100 inhibited
this differentiation. In the present study, using an miRNA
microarray, several miRNAs were found to be differently expressed
during hADSC osteogenic differentiation. In particular, the study
focused on miR-143, since its expression was decreased the most
during this process. It has previously been reported that miR-143
was downregulated in MSC osteogenic differentiation and negatively
regulated this process by targeting different genes (27,38). However, little is known regarding
the effect of miR-143 on the differentiation of hADSCs. To the best
of our knowledge, the present study investigated is the first to
investigate the role of miR-143 in the osteogenic differentiation
of hADSCs. The data revealed that overexpression of miR-143
significantly inhibited osteogenic differentiation, while
inhibition of miR-143 enhanced this process, suggesting the
important role of this miRNA in this cellular change.
Activation of the MAPK signaling pathway,
particularly of ERK1/2, has been implicated in the process of
osteogenic differentiation of hADSCs (18) and human periodontal ligament stem
cells (hPDLSCs) (33). For
instance, activation of the ERK1/2 signaling pathway has been
demonstrated to mediate the promoting effect of fibroblast growth
factor-7 on the osteogenic differentiation of embryonic stem cells
(39) and the naringin-induced
osteogenic differentiation of immortalized hPDLSCs (33). Notably, previous studies have
reported that miRNAs are important in the regulation of osteogenic
differentiation of MSCs through activating the ERK1/2 signaling
pathway. For example, Mei et al (40) revealed that miR-21 modulated
ERK-MAPK signaling activity by repressing SPRY2 expression to
affect the human MSC differentiation. Of note, Dong and Hu
(41) demonstrated that
overexpression of miR-143 suppressed the phosphorylation of ERK1/2
in non-small-cell lung cancer PC9/GR cells. To further elucidate
the mechanisms by which miR-143 promotes the osteogenic
differentiation of hADSCs, the effects of miR-143 downregulation on
key kinases in the MAPK signaling pathway were examined in the
present study. The data revealed that only the ERK1/2 signaling
pathway was regulated by miR-143, whereas the p38 and JNK pathways
did not appear to be involved. Further assessments indicated that
the promoting effect of antagomir-143 on the osteogenic
differentiation of hADSCs was markedly reversed by treatment with
MEK-specific inhibitors, U0126 and PD98059, which suggested that
the promoting effects of miR-143 on the osteogenic differentiation
of hADSCs may be mediated by the ERK1/2 signaling pathway.
It is well-known that k-Ras is an upstream regulator
of the ERK1/2 signaling pathway (42). Under physiological conditions,
k-Ras stimulates the transformation of Ras-GTP to Ras-GDP and
promotes the activation of downstream of MEK, which in turn
activates the ERK1/2 pathway (43,44). The Ras/Raf/MEK/ERK signaling
pathway has been reported to be closely associated with osteogenic
differentiation of MSCs. A previous study by Feng et al
(45) indicated that the HMGB1
promoted the secretion of multiple cytokines and potentiated the
osteogenic differentiation of MSCs through promoting the activation
of the Ras/MAPK signaling pathway. Another study also revealed that
strontium enhanced the MSC osteo-genic differentiation by
activating the Ras/MAPK signaling pathway (46). It has previously been reported
that miR-143 directly targets k-Ras in human cancer (47,48); however, whether k-Ras is a
functional target of miR-143 during hADSC osteogenic
differentiation was unclear. In the present study, k-Ras was
confirmed as a direct target of miR-143 in hADSCs. Furthermore, it
was observed that miR-143 blocked the activation of
k-Ras/c-Raf/MEK/ERK signaling, whereas inhibition of miR-143
enhanced this signaling. Collectively, these findings indicated
that miR-143 may regulate the osteogenic differentiation of hADSCs
through the k-Ras/MEK/ERK signaling pathway.
To date, the majority of cell-based therapies are
conducted using the well-characterized BMSCs. The osteogenic
process of BMSCs is a critical step for bone formation, and it has
been already used in clinical trials for bone damage treatment
(49-51). Compared with the BMSCs, hADSCs
displayed a significantly higher proliferating ability and
differentiation potential, thus confirming that hADSCs are a better
candidate for clinical application (52,53). In addition, hADSC harvesting
involves less pain and ethical issues (54). Since BMSCs have been involved for
some time in clinical protocols of cell therapy, we will further
verify the results of the present study on BMSCs in the future.
In conclusion, the data demonstrated that miR-143
was the most significantly downregulated miRNA during osteogenesis
in hADSC. Furthermore, miR-143 inhibition promoted the osteogenic
differentiation of hADSCs by activating k-Ras/c-Raf/MEK/ERK
pathway. The present study may provide a theoretical basis for the
development of stem cell-based therapy of bone fractures in the
future.
Acknowledgments
Not applicable.
Funding
This study was supported by the Shanghai Municipal
Planning Commission of Science and Research Fund (grant no.
201640219).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YZ, KZ, LW, HG and ZH performed the experiments,
analysed the data and wrote the paper. JX conceptualized the study
design, and contributed to data analysis and experimental
materials. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All individuals provided informed consent for the
use of human specimens for clinical research. The present study was
approved by the Ethics Committee of Minhang Hospital, Fudan
University (Shanghai, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Locke M, Feisst V and Dunbar PR: Concise
review: Human adipose-derived stem cells: Separating promise from
clinical need. Stem Cells. 29:404–411. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang S, Qu X and Zhao RC: Mesenchymal stem
cells hold promise for regenerative medicine. Front Med. 5:372–378.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gimble JM and Guilak F: Differentiation
potential of adipose derived adult stem (ADAS) cells. Curr Top Dev
Biol. 58:137–160. 2003. View Article : Google Scholar
|
|
4
|
Alvira-González J, Sánchez-Garcés MÀ,
Cairó JR, Del Pozo MR, Sánchez CM and Gay-Escoda C: Assessment of
bone regeneration using adipose-derived stem cells in critical-size
alveolar ridge defects: An experimental study in a dog model. Int J
Oral Maxillofac Implants. 31:196–203. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sánchez-Garcés MÀ, Alvira-González J,
Sánchez CM, Barbany Cairó JR, Del Pozo MR and Gay-Escoda C: Bone
regeneration using adipose-derived stem cells with fibronectin in
dehiscence-type defects associated with dental implants: An
experimental study in a dog model. Int J Oral Maxillofac Implants.
32:e97–e106. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jia J, Tian Q, Ling S, Liu Y, Yang S and
Shao Z: miR-145 suppresses osteogenic differentiation by targeting
Sp7. FEBS Lett. 587:3027–3031. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang SN, Zhao XQ, Yu B and Wang BW:
miR-193a inhibits osteogenic differentiation of bone marrow-derived
stroma cell via targeting HMGB1. Biochem Biophys Res Commun.
503:536–543. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li C, Yang D, Cao X, Wang F, Jiang H, Guo
H, Du L, Guo Q and Yin X: LFG-500, a newly synthesized flavonoid,
attenuates lipopolysaccharide-induced acute lung injury and
inflammation in mice. Biochem Pharmacol. 113:57–69. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Afzal F, Pratap J, Ito K, Ito Y, Stein JL,
van Wijnen AJ, Stein GS, Lian JB and Javed A: Smad function and
intranuclear targeting share a Runx2 motif required for osteogenic
lineage induction and BMP2 responsive transcription. J Cell
Physiol. 204:63–72. 2005. View Article : Google Scholar
|
|
12
|
Da Forno PD, Pringle JH, Hutchinson P,
Osborn J, Huang Q, Potter L, Hancox RA, Fletcher A and Saldanha GS:
WNT5A expression increases during melanoma progression and
correlates with outcome. Clin Cancer Res. 14:5825–5832. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Corrêa SA and Eales KL: The Role of p38
MAPK and its substrates in neuronal plasticity and
neurodegenerative disease. J Signal Transduct. 2012:6490792012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Thouverey C and Caverzasio J: The p38alpha
MAPK positively regulates osteoblast function and postnatal bone
acquisition. Cell Mol Life Sci. 69:3115–3125. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ying X, Cheng S, Wang W, Lin Z, Chen Q,
Zhang W, Kou D, Shen Y, Cheng X, Peng L, Zi Xu H and Zhu Lu C:
Effect of lactoferrin on osteogenic differentiation of human
adipose stem cells. Int Orthop. 36:647–653. 2012. View Article : Google Scholar :
|
|
16
|
Li Y, Wang J, Chen G, Feng S, Wang P, Zhu
X and Zhang R: Quercetin promotes the osteogenic differentiation of
rat mesen-chymal stem cells via mitogen-activated protein kinase
signaling. Exp Ther Med. 9:2072–2080. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou C, Zhang X, Xu L, Wu T, Cui L and Xu
D: Taurine promotes human mesenchymal stem cells to differentiate
into osteoblast through the ERK pathway. Amino Acids. 46:1673–1680.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ye C, Chen M, Chen E, Li W, Wang S, Ding
Q, Wang C, Zhou C, Tang L, Hou W, et al: Knockdown of FOXA2
enhances the osteogenic differentiation of bone marrow-derived
mesenchymal stem cells partly via activation of the ERK signalling
pathway. Cell Death Dis. 9:8362018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang H, Li C, Li J, Zhu Y, Jia Y, Zhang Y,
Zhang X, Li W, Cui L, Li W and Liu Y: Naringin enhances osteogenic
differentiation through the activation of ERK signaling in human
bone marrow mesenchymal stem cells. Iran J Basic Med Sci.
20:408–414. 2017.PubMed/NCBI
|
|
20
|
Boquest AC, Shahdadfar A, Brinchmann JE
and Collas P: Isolation of stromal stem cells from human adipose
tissue. Methods Mol Biol. 325:35–46. 2006.PubMed/NCBI
|
|
21
|
Li B, Qu C, Chen C, Liu Y, Akiyama K, Yang
R, Chen F, Zhao Y and Shi S: Basic fibroblast growth factor
inhibits osteogenic differentiation of stem cells from human
exfoliated deciduous teeth through ERK signaling. Oral Dis.
18:285–292. 2012. View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
23
|
Mei LL, Wang WJ, Qiu YT, Xie XF, Bai J and
Shi ZZ: miR-125b-5p functions as a tumor suppressor gene partially
by regulating HMGA2 in esophageal squamous cell carcinoma. PLoS
One. 12:e01856362017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu TM and Lee EH: Transcriptional
regulatory cascades in Runx2-dependent bone development. Tissue Eng
Part B Rev. 19:254–263. 2013. View Article : Google Scholar :
|
|
25
|
Lee MJ, Chen HT, Ho ML, Chen CH, Chuang
SC, Huang SC, Fu YC, Wang GJ, Kang L and Chang JK: PPARγ silencing
enhances osteogenic differentiation of human adipose-derived
mesenchymal stem cells. J Cell Mol Med. 17:1188–1193.
2013.PubMed/NCBI
|
|
26
|
Ying X, Chen X, Cheng S, Guo X, Chen H and
Xu HZ: Phosphoserine promotes osteogenic differentiation of human
adipose stromal cells through bone morphogenetic protein
signalling. Cell Biol Int. 38:309–317. 2014. View Article : Google Scholar
|
|
27
|
Li E, Zhang J, Yuan T and Ma B: MiR-143
suppresses osteogenic differentiation by targeting Osterix. Mol
Cell Biochem. 390:69–74. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ge C, Xiao G, Jiang D and Franceschi RT:
Critical role of the extracellular signal-regulated kinase-MAPK
pathway in osteoblast differentiation and skeletal development. J
Cell Biol. 176:709–718. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Greenblatt MB, Shim JH and Glimcher LH:
Mitogen-activated protein kinase pathways in osteoblasts. Annu Rev
Cell Dev Biol. 29:63–79. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Artigas N, Ureña C, Rodriguez-Carballo E,
Rosa JL and Ventura F: Mitogen-activated protein kinase
(MAPK)-regulated interactions between Osterix and Runx2 are
critical for the transcriptional osteogenic program. J Biol Chem.
289:27105–27117. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Park S, Arai Y, Kim BJ, Bello A, Ashraf S,
Park H, Park KS and Lee SH: SPRY4 suppression facilitates
osteogenic differentiation in adipose-derived mesenchymal stem
cells. Tissue Eng Part A. 25:1645–1657. 2019. View Article : Google Scholar
|
|
32
|
Yang X, Yang Y, Zhou S, Gong X, Dai Q,
Zhang P and Jiang L: Puerarin stimulates osteogenic differentiation
and bone formation through the ERK1/2 and p38-MAPK signaling
pathways. Curr Mol Med. 17:488–496. 2018. View Article : Google Scholar
|
|
33
|
Wei K, Xie Y, Chen T, Fu B, Cui S, Wang Y,
Cai G and Chen X: ERK1/2 signaling mediated naringin-induced
osteogenic differentiation of immortalized human periodontal
ligament stem cells. Biochem Biophys Res Commun. 489:319–325. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wei F, Liu Y, Bellail AC, Olson JJ, Sun
SY, Lu G, Ding L, Yuan C, Wang G and Hao C: K-Ras mutation-mediated
IGF-1-induced feedback ERK activation contributes to the rapalog
resistance in pancreatic ductal adenocarcinomas. Cancer Lett.
322:58–69. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim YJ, Bae SW, Yu SS, Bae YC and Jung JS:
miR-196a regulates proliferation and osteogenic differentiation in
mesenchymal stem cells derived from human adipose tissue. J Bone
Miner Res. 24:816–825. 2009. View Article : Google Scholar
|
|
36
|
Luzi E, Marini F, Sala SC, Tognarini I,
Galli G and Brandi ML: Osteogenic differentiation of human adipose
tissue-derived stem cells is modulated by the miR-26a targeting of
the SMAD1 transcription factor. J Bone Miner Res. 23:287–295. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zeng Y, Qu X, Li H, Huang S, Wang S, Xu Q,
Lin R, Han Q, Li J and Zhao RC: MicroRNA-100 regulates osteogenic
differentiation of human adipose-derived mesenchymal stem cells by
targeting BMPR2. FEBS Lett. 586:2375–2381. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang P, Yang W, Wang G and Li Y: miR-143
suppresses the osteogenic differentiation of dental pulp stem cells
by inactivation of NF-κB signaling pathway via targeting TNF-α.
Arch Oral Biol. 87:172–179. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jeon YM, Kook SH, Rho SJ, Lim SS, Choi KC,
Kim HS, Kim JG and Lee JC: Fibroblast growth factor-7 facilitates
osteogenic differentiation of embryonic stem cells through the
activation of ERK/Runx2 signaling. Mol Cell Biochem. 382:37–45.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mei Y, Bian C, Li J, Du Z, Zhou H, Yang Z
and Zhao RC: miR-21 modulates the ERK-MAPK signaling pathway by
regulating SPRY2 expression during human mesenchymal stem cell
differentiation. J Cell Biochem. 114:1374–1384. 2013. View Article : Google Scholar
|
|
41
|
Dong YZ and Hu T: Effects of miR-143
overexpression on proliferation, apoptosis, EGFR and downstream
signaling pathways in PC9/GR cell line. Eur Rev Med Pharmacol Sci.
22:1709–1716. 2018.PubMed/NCBI
|
|
42
|
Castellano E and Santos E: Functional
specificity of ras isoforms: So similar but so different. Genes
Cancer. 2:216–231. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cheng D, Zhao L, Xu Y, Ou R, Li G, Yang H
and Li W: K-Ras promotes the non-small lung cancer cells survival
by cooperating with sirtuin 1 and p27 under ROS stimulation. Tumour
Biol. 36:7221–7232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Georgieva M, Krasteva M, Angelova E,
Ralchev K, Dimitrov V, Bozhimirov S, Georgieva E and Berger MR:
Analysis of the K-ras/B-raf/Erk signal cascade, p53 and CMAP as
markers for tumor progression in colorectal cancer patients. Oncol
Rep. 20:3–11. 2008.PubMed/NCBI
|
|
45
|
Feng L, Xue D, Chen E, Zhang W, Gao X, Yu
J, Feng Y and Pan Z: HMGB1 promotes the secretion of multiple
cytokines and potentiates the osteogenic differentiation of
mesenchymal stem cells through the Ras/MAPK signaling pathway. Exp
Ther Med. 12:3941–3947. 2016. View Article : Google Scholar
|
|
46
|
Peng S, Zhou G, Luk KD, Cheung KM, Li Z,
Lam WM, Zhou Z and Lu WW: Strontium promotes osteogenic
differentiation of mesenchymal stem cells through the Ras/MAPK
signaling pathway. Cell Physiol Biochem. 23:165–174. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Qin HX, Cui HK, Pan Y, Hu RL, Zhu LH and
Wang SJ: miR-143 inhibits cell proliferation through targeted
regulating the expression of K-ras gene in HeLa cells. Zhonghua
Zhong Liu Za Zhi. 38:893–897. 2016.In Chinese. PubMed/NCBI
|
|
48
|
Takai T, Tsujino T, Yoshikawa Y, Inamoto
T, Sugito N, Kuranaga Y, Heishima K, Soga T, Hayashi K, Miyata K,
et al: Synthetic miR-143 exhibited an anti-cancer effect via the
down-regulation of K-RAS networks of renal cell cancer cells in
vitro and in vivo. Mol Ther. 27:1017–1027. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Caplan AI: Review: Mesenchymal stem cells:
Cell-based reconstructive therapy in orthopedics. Tissue Eng.
11:1198–1211. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Arinzeh TL: Mesenchymal stem cells for
bone repair: Preclinical studies and potential orthopedic
applications. Foot Ankle Clin. 10:651–665. viii2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Bhansali S, Dutta P, Kumar V, Yadav MK,
Jain A, Mudaliar S, Bhansali S, Sharma RR, Jha V, Marwaha N, et al:
Efficacy of autologous bone marrow-derived mesenchymal stem cell
and mononuclear cell transplantation in type 2 diabetes mellitus: A
randomized, placebo-controlled comparative study. Stem Cells Dev.
26:471–481. 2017. View Article : Google Scholar
|
|
52
|
Zuk PA, Zhu M, Mizuno H, Huang J, Futrell
JW, Katz AJ, Benhaim P, Lorenz HP and Hedrick MH: Multilineage
cells from human adipose tissue: Implications for cell-based
therapies. Tissue Eng. 7:211–228. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tobita M, Orbay H and Mizuno H:
Adipose-derived stem cells: Current findings and future
perspectives. Discov Med. 11:160–170. 2011.PubMed/NCBI
|
|
54
|
Sempere JM, Martinez-Peinado P, Arribas
MI, Reig JA, De La Sen ML, Zubcoff JJ, Fraga MF, Fernández AF,
Santana A and Roche E: Single cell-derived clones from human
adipose stem cells present different immunomodulatory properties.
Clin Exp Immunol. 176:255–265. 2014. View Article : Google Scholar : PubMed/NCBI
|